

 Unveiling the histology and anatomy of the lungfish Chaoceratodus portezuelensis (Sarcopterygii: Dipnoi) from the Portezuelo and Cerro Lisandro formations (Upper Cretaceous) of Argentine Patagonia
Unveiling the histology and anatomy of the lungfish Chaoceratodus portezuelensis (Sarcopterygii: Dipnoi) from the Portezuelo and Cerro Lisandro formations (Upper Cretaceous) of Argentine Patagonia
Article number: 27.2.a42
https://doi.org/10.26879/1408
Copyright Palaeontological Association, August 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 22 May 2024. Acceptance: 16 August 2024.
ABSTRACT
Chaoceratodus portezuelensis from the Cretaceous of Argentine Patagonia is undergoing re-examination based on both previously known and newly referred tooth plates and associated bones. This study expands upon the original diagnosis by incorporating characteristics specific to pterygopalatine and prearticular tooth plates, and dental histology features. Additionally, it broadens the information based on Mesozoic dipnoans from Patagonia for potential future systematic and phylogenetic studies. The pterygopalatine tooth plates are characterized by slightly curved mediolingual edges, with the inner angle positioned just behind the second denticulation and short, wide-based first denticulations. The prearticular tooth plates feature mediolingual edges tending towards straight, with the inner angle positioned at the level of the second denticulation, short first denticulations, and a bilobed spur. Histological cross-sections reveal rows of solid interdenteonal dentine corresponding to ridges that anastomose over the rest of the plateau. This pattern is visible on the external surface of the tooth plates, creating a characteristic wear pattern for the species. Both the morphology and histology of the dentition bear resemblance, albeit with differences, to other Patagonian species, such as Metaceratodus baibianorum and Atlantoceratodus iheringi.
Karen M. Panzeri. Museo Argentino de Ciencias Naturales Bernardino Rivadavia, División de Ictiología, Av. Ángel Gallardo 470, Buenos Aires, Argentina, C1405DJR (Corresponding author)
k.panzeri@fcnym.unlp.edu.ar
Federico A. Guzmán. Universidad Nacional de Río Negro, Estados Unidos 750, Río Negro, Argentina, R8332. federico.a.guzman@gmail.com
Key words: tooth plates; interdenteonal dentine; circumdenteonal dentine; dental histology; paleohistology; dipnoi; Patagonia
Final citation: Panzeri, Karen M. and Guzmán, Federico A. 2024. Unveiling the histology and anatomy of the lungfish Chaoceratodus portezuelensis (Sarcopterygii: Dipnoi) from the Portezuelo and Cerro Lisandro formations (Upper Cretaceous) of Argentine Patagonia. Palaeontologia Electronica, 27(2):a42.
https://doi.org/10.26879/1408
palaeo-electronica.org/content/2024/5317-cretaceous-lungfish-analysis
Copyright: August 2024 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Dipnoans are a group of sarcopterygians that emerged in the Devonian, initially displaying high taxonomic diversity (Clack et al., 2011; Clack and Ahlberg, 2016; Yanchao et al., 2021). These organisms were initially marine forms and had a global distribution (Clement, 2019). The skulls, often found, were characterized by many dermal and chondral ossifications, elongated snouts, and small orbits. Regarding dentition in these early forms, numerous dental types, such as isolated pseudo teeth, dentine plates, or tooth plates, have been identified (Ahlberg et al., 2006; Clack et al., 2011). As an evolutionary trend towards the present day, there is an observed reduction in the number of dermal and chondral ossifications. Additionally, since the Mesozoic, the dentition stabilizes in the form of tooth plates (Cloutier, 1996; Clack et al., 2011; Panzeri et al., 2022a). Simultaneously, they became restricted to continental freshwater environments, with a distribution limited to the Southern Hemisphere (Cavin, 2017). Currently, the only six living species of dipnoans are found in Africa, South America, and Australia.
Although rare, articulated Mesozoic skulls have been found in different periods and parts of the world (Teller, 1891; Martin 1981a; Antunes et al., 1990; Kemp, 1994; Cavin et al., 2007, 2020; Pardo et al., 2010; Panzeri et al., 2022a). Conversely, the fossil record of Mesozoic dipnoans is primarily represented by tooth plates due to their high resistance (Churcher and De Iullis, 2001; Apesteguía et al., 2007; Skrzycki, 2015; Frederickson and Cifelli, 2017; Panzeri et al., 2020; Pawlak et al., 2020). The latter occurs in the Cretaceous strata of Patagonia, and due to the nature of the deposits, these tooth plates are often found rounded and fragmented. Numerous species have been named based on these findings: Metaceratodus wichmanni (Apesteguía et al., 2007); M. kaopen (Apesteguía et al., 2007); M. baibianorum Panzeri et al., 2020; Atlantoceratodus iheringi Cione et al., 2007; A. patagonicus Agnolin, 2010; Ceratodus argentinus Apesteguía et al., 2007; and Chaoceratodus portezuelensis Apesteguía et al., 2007. Although mostly tooth plates are found, two three-dimensionally preserved articulated skulls along with part of a postcranial skeleton were recently described by Panzeri et al. (2022a), assigning them to the species Rinconodus salvadori.
Some of the dipnoan fossils from Argentina were previously reviewed by Apesteguía et al. (2007). In their work, these authors named the species Chaoceratodus portezuelensis from the Portezuelo Formation based on a single tooth plate. From the same layers, they also mentioned the presence of a bone with a certain pathology and a tooth plate assigned to the species “Ameghinoceratodus” iheringi. Additionally, materials from the Cerro Lisandro Formation were studied and referred to the family Ptychoceratodontidae and the genus Ceratodus. This contribution aims to make a review of these tooth plates, providing a detailed anatomical and histological description. This study provides a foundation of information on the anatomy of tooth plates for future analyses of dipnoans around the world and adds to recent contributions with similar methodologies conducted on other species of Argentine dipnoans.
GEOLOGICAL SETTING
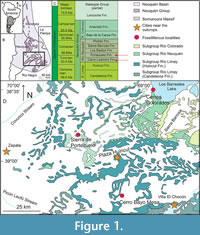 The Neuquén Basin is a retroarc basin developed during Mesozoic times on the Pacific margin of South America (Legarreta and Uliana, 1996; Figure 1A-B). It hosts an almost continuous marine and continental sedimentary sequence from the Late Triassic to the Early Cenozoic (Howell et al., 2005). Specifically, the Neuquén Group was deposited in the Neuquén Basin during the Late Cretaceous (Cenomanian-Campanian) (Garrido, 2010; Figure 1C). The sedimentary sequences that constitute the Neuquén Group have been known since early studies as “Dinosaur Beds” (“Estratos con dinosaurios” in Spanish, Wichmann, 1927) due to the numerous remains of these organisms they contain. During the Turonian and Coniacian stages, sedimentary deposits of the Cerro Lisandro and Portezuelo formations are present within the Neuquén Group.
The Neuquén Basin is a retroarc basin developed during Mesozoic times on the Pacific margin of South America (Legarreta and Uliana, 1996; Figure 1A-B). It hosts an almost continuous marine and continental sedimentary sequence from the Late Triassic to the Early Cenozoic (Howell et al., 2005). Specifically, the Neuquén Group was deposited in the Neuquén Basin during the Late Cretaceous (Cenomanian-Campanian) (Garrido, 2010; Figure 1C). The sedimentary sequences that constitute the Neuquén Group have been known since early studies as “Dinosaur Beds” (“Estratos con dinosaurios” in Spanish, Wichmann, 1927) due to the numerous remains of these organisms they contain. During the Turonian and Coniacian stages, sedimentary deposits of the Cerro Lisandro and Portezuelo formations are present within the Neuquén Group.
The Cerro Lisandro Formation (middle-upper Turonian) extends across the Neuquén and Rio Negro provinces. Lithologically, it comprises fine clastic deposits, including layers of red mudstones, interspersed with levels of siltstones and fine micaceous quartz sandstones, displaying yellowish and gray-green hues (Garrido, 2011). The formation is interpreted either as a sinuous fluvial system with a mixed or mud-dominant load (Salgado et al., 2009; Garrido, 2010) or as an alluvial and floodplain environment with seasonal climate (Sánchez and Asurmendi, 2014).
Most tooth plates from the Cerro Lisandro Formation originate from a 40 cm thick sandy channel at Cerro Bayo Mesa locality (Coria et al., 1996), situated 40 km south of Plaza Huincul, Neuquén province (Figure 1D). Explorations at the Cerro Bayo Mesa locality began in 1993 when Roberto Saldivia, a farmer from Plaza Huincul, made a notable discovery in the sediments: he found bones belonging to an ornithischian dinosaur. This finding prompted an expedition in 1996 led by a team from the MCF and the MUC (Coria and Calvo, 2002; Coria, 2022). Four specimens of the ornithopod Anabisetia saldiviai were discovered during this expedition alongside other findings, including numerous lungfish tooth plates (Coria et al., 1996). Other fossil remains found at this locality include a theropod (Coria et al., 2006), a crocodyliform (Barrios et al., 2016), and various isolated remains (Coria et al., 1996). Lastly, tooth plate discoveries in Cerros Colorados locality (near Los Barreales Lake) were made in 2003 by the team from the Museo Carmen Funes (Garrido, personal commun., 2024).
The Portezuelo Formation (upper Turonian-lower Coniacian) extends across Neuquén, Río Negro, and Mendoza provinces, with its type locality in Sierra del Portezuelo (Garrido, 2010), 20 km west of Plaza Huincul (Figure 1C-D). It consists of fine- to medium-grained sandstones alternating with reddish and yellowish mudstones (Sánchez et al., 2005; Garrido, 2010), indicative of a sinuous fluvial system with sand-dominant load (Salgado et al., 2009; Garrido, 2010). Tooth plates from this formation are primarily found at Sierra del Portezuelo (Figure 1D) alongside various fossils, including theropods, sauropods, turtles, birds, and remains of small-sized ornithopods, among others (Novas, 1996; de la Fuente, 2003; Agnolin et al., 2006; Apesteguía et al., 2007). The holotype of the species C. portezuelensis, along with other tooth plates, originated from this area and was obtained during two expeditions carried out in 1990 by a team from the MACN and in 2001 by a team from the MACN together with a team from the MCF, respectively. Additional tooth plates were discovered in 2003 at Cerros Colorados locality within Portezuelo Formation outcrops, in a fieldwork carried out by MCF and the NHMLA (Apesteguía et al., 2007; Garrido, personal commun., 2024). Few fossil remains have been reported from this area, including the tooth plates previously mentioned, as well as gymnosperm woods and other fossil remains from different formations and ages (see Martinez and Lutz, 2007; Calvo and Salgado, 2022).
MATERIALS AND METHODS
Materials
A total of 56 tooth plates were studied. From the Cerro Lisandro Formation, Cerro Bayo Mesa locality, 50 tooth plates (MCF-PVPH-425 to 440; MCF-PVPH-542, and some fragmentary tooth plates without numbering but belonging to the same batch), along with the three histological cross-sections MCF-PVPH-915A (one), 915B (two). From the same formation but at Cerros Colorados locality, two tooth plates (MCF-PVPH-500 and 506). From the Portezuelo Formation, Sierra de Portezuelo locality, only three tooth plates (MCF-PVPH-373, 374, 424), and from Cerros Colorados locality, one tooth plate (MCF-PVPH-572).
Methods
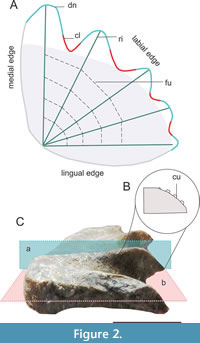 Morphological descriptions follow the terminology implemented in Panzeri et al. (2020, 2022a, 2022b), which follows a set of terms previously defined by various authors (Kemp, 1977; Smith and Campbell, 1987; Churcher and De Iullis, 2001). Tooth plates can be upper (pterygopalatine or vomerine) or lower (prearticular). In a prearticular or pterygopalatine tooth plate, a medial, a lingual, and a labial edge are identified (Figure 2A). Undulations are observed on the labial edge, where depressions correspond to clefts and elevations to denticulations. The crest, located at the maximum curvature of the denticulation, may exhibit cusps (Figure 2B). The crests align with the ridges on the occlusal surface, while the clefts correspond to the furrows. Lastly, the occlusal surface area which does not involve denticulations is the plateau, often exhibiting wear patterns.
Morphological descriptions follow the terminology implemented in Panzeri et al. (2020, 2022a, 2022b), which follows a set of terms previously defined by various authors (Kemp, 1977; Smith and Campbell, 1987; Churcher and De Iullis, 2001). Tooth plates can be upper (pterygopalatine or vomerine) or lower (prearticular). In a prearticular or pterygopalatine tooth plate, a medial, a lingual, and a labial edge are identified (Figure 2A). Undulations are observed on the labial edge, where depressions correspond to clefts and elevations to denticulations. The crest, located at the maximum curvature of the denticulation, may exhibit cusps (Figure 2B). The crests align with the ridges on the occlusal surface, while the clefts correspond to the furrows. Lastly, the occlusal surface area which does not involve denticulations is the plateau, often exhibiting wear patterns.
Tooth plates were photographed with a Canon digital camera and later measured using ImageJ software (Schneider et al., 2012). The inner angle was measured between the tip of the first and last denticulations, and the linear measurements of the medial and lingual edges were taken in centimeters.
The histological cross-sections were prepared by Dr. Maria Eugenia Pereyra (FCNyM, UNLP) following the methodology implemented in Cerda et al. (2020). Histological cross-sections were obtained following different cutting planes, here referred to as vertical (aligned along the ridge, Figure 2C) and horizontal (parallel to the occlusal surface, Figure 2C). Due to the scarcity and thinness of tooth plates from C. portezuelensis, conducting multiple cuts along a horizontal plane was not feasible. Only two pterygopalatine tooth plates were selected for analysis, one resulting in two vertical cross-sections, while the other provided a single horizontal cross-section. Histological cross-sections were analyzed at INREMI and at MACN (Ichnology laboratory), using a Nikon Optiphot-Pol 255884 petrographic microscope at various magnifications (2x, 5x, 10x, and 20x), utilizing transmitted light with a cross-polarizer (530 nm) and a 1/4 lambda filter. Glycerin was used to enhance the clarity of the sections. Microscope images were captured using a Xiaomi RedmiNote 10 digital camera.
Histological descriptions adhere to the terminology implemented by Panzeri et al. (2022b) and Panzeri (2024), which encompass terms previously defined by other authors (see cited references). Externally, a layer of enamel is briefly identified in a horizontal cross-section. Beneath the enamel lies a mantle dentine layer, and an interdenteonal dentine is observed internally. Denteons are defined as the combination of pulp canals and circumdenteonal dentine and are categorized into transitional (elongated and attached to the margins) and secondary denteons (embedded in interdenteonal dentine). In vertical cross-sections, denteons are typically observed in an occluso-pulpal direction, with interdenteonal dentine between them.
Institutional abbreviations. MACN, Museo Argentino de Ciencias Naturales Bernardino Rivadavia; MAU, Museo Argentino Urquiza, Rincón de los Sauces, Neuquén, Argentina; MCF, Museo Carmen Funes Plaza Huincul, Argentina; MML, Museo Paleontológico Municipal “Héctor Cabaza”; MPCA, Museo Provincial Carlos Ameghino, Cipolletti, Río Negro, Argentina; MPCN, Museo Patagónico de Ciencias Naturales, General Roca, Río Negro, Argentina; MPEF, Museo Paleontológico Egidio Feruglio, Trelew, Chubut, Argentina; MPM, Museo Provincial Padre Molina, Rio Gallegos, Santa Cruz, Argentina; MUC, Museo de la Universidad Nacional del Comahue; NHMLA, Natural History Museum of Los Ángeles County.
Anatomical abbreviations. af, anterior facet; ap, ascending process of the pterygopalatine bone; bid, birefringent interdenteonal dentine; bd, branched denteon; cd, circumdenteonal dentine; cl, cleft; cu, cusps; dn, denticulation; dt, dentinal tubules; en, enamel; fu, furrow; gr, granules; icb, inner circumdenteonal dentine; il, incremental line; ld, linear denteon; mid, monorefringent interdenteonal dentine; ocb, outer circumdenteonal dentine; pf, posterior facet; pun, punctuations; ri, ridge; ru, Ruge’s canal; spu, spur; st, step; trd, transitional denteons; tx, bone texture; wp, wear pattern; ld, linear denteon; mid, monorefringent interdenteonal dentine; pc, pulp canal.
SYSTEMATIC PALEONTOLOGY
Class SARCOPTERYGII Romer, 1955
Order DIPNOI Müller, 1845
Family CERATODONTIDAE Gill, 1872
Genus CHAOCERATODUS Apesteguía, Agnolin, and Claeson, 2007
Type species. Chaoceratodus portezuelensis Apesteguía, Agnolin and Claeson, 2007
Holotype. MCF-PVPH-373, prearticular tooth plate with the homonymous bone fused.
Type locality. Portezuelo Formation (locality Sierra de Portezuelo).
Referred material. MCF-PVPH-425 to 440: 425, pterygopalatine tooth plate; 426, pterygopalatine tooth plate; 427, pterygopalatine tooth plate; 428, pterygopalatine tooth plate; 429, prearticular tooth plate; 430, prearticular tooth plate; 431, pterygopalatine tooth plate; 432, fragmentary tooth plate; 433, pterygopalatine tooth plate (in Apesteguía et al., 2007, numbered as 453); 434, pterygopalatine tooth plate; 435, fragmentary tooth plate; 436, prearticular tooth plate; 437, pterygopalatine tooth plate; 438, pterygopalatine tooth plate; 439, pterygopalatine tooth plate; 440, prearticular tooth plate; and others without numbering; MCF-PVPH-542, three tooth plates; MCF-PVPH-500, prearticular tooth plate; MCF-PVPH-506, fragmented tooth plate; MCF-PVPH-374, indeterminate pterygopalatine bone; MCF-PVPH-424, prearticular tooth plate; MCF-PVPH-572, pterygopalatine tooth plate; MCF-PVPH-915A, B: sections of tooth plates, 915A: horizontal cross-section; 915B: two vertical cross-sections.
Diagnosis. Pterygopalatine tooth plates display five denticulations, with the inner angle positioned slightly behind the second denticulation. Prearticular tooth plates display four denticulations, and the inner angle is slightly ahead of the second denticulation. Superficial clefts, with the first being the deepest. Medial and lingual edges generally maintain straightness, with a curvature in the inner angle. In pterygopalatine tooth plates, the lingual edge forms a slightly obtuse angle with the sagittal plane. The first denticulation of pterygopalatine tooth plates is short and has a wide base, which is posteriorly curved at the end, with a small anterior facet. The first denticulation of prearticular tooth plates is straight, with a narrower base and a marked facet toward the inner angle. The second denticulation in prearticular tooth plates may exhibit a posterior wear facet. The fourth and fifth denticulations in pterygopalatine tooth plates are involved in the formation of the step. The fourth denticulation is involved in the formation of a bilobed spur in prearticular tooth plates. The ascending process is evident at the level of the second denticulation. The interdenteonal dentine forms bands with monorefringent centers coinciding with the ridges, externally evidenced as a reticulated wear pattern with pronounced ridges.
Morphological Description
The tooth plates of Chaoceratodus portezuelensis display varying sizes. The larger tooth plates measure 2.794 cm along the medial edge and 2.404 cm along the lingual edge, while the dimensions of the smaller fragments cannot be precisely determined. The clefts are subtly deep, and the denticulations feature sharp ridges with rounded or acute crests. The inner angle measures 85.91° in pterygopalatine tooth plates and 85.56° in prearticular tooth plates.
 Pterygopalatine tooth plates and associated bones. The pterygopalatine tooth plates are intricately fused to the pterygopalatine bone (Figure 3A-C). The external surface of the pterygopalatine bone displays a textured anastomosed pattern, creating small, delineated areas and featuring foramina (Figure 3A). Generally, these surfaces bear lichen marks, seamlessly blending with the bone texture. None of the specimens have a fully preserved pterygopalatine bone, and only the posterior portion of the symphysis is known. Dorsally, at the level of the second denticulation, the pterygopalatine process is evident, characterized by a large and circular base (Figure 3A-C).
Pterygopalatine tooth plates and associated bones. The pterygopalatine tooth plates are intricately fused to the pterygopalatine bone (Figure 3A-C). The external surface of the pterygopalatine bone displays a textured anastomosed pattern, creating small, delineated areas and featuring foramina (Figure 3A). Generally, these surfaces bear lichen marks, seamlessly blending with the bone texture. None of the specimens have a fully preserved pterygopalatine bone, and only the posterior portion of the symphysis is known. Dorsally, at the level of the second denticulation, the pterygopalatine process is evident, characterized by a large and circular base (Figure 3A-C).
The tooth plates exhibit five denticulations, and the punctuations are randomly distributed (Figure 3D). While the first denticulation is the longest, it does not differ significantly from the others. Its base is wide, and its tip abruptly directs posteriorly towards the end (Figure 3D). The remaining denticulations are of similar sizes, with the fifth being the shortest (Figure 3D). The ridges and crests tend to curve posteriorly, and the clefts have a comparable depth, although the first is slightly deeper (Figure 3E-H). The inner angle of the pterygopalatine tooth plates is rounded and positioned slightly posterior to the second denticulation (Figure 3D, E).
The medial edge surpasses the lingual edge in length, both displaying a subtle curvature but tending more towards straightness. Conversely, the area surrounding the inner angle is highly curved (Figure 3D, I-J). Toward their free portions, both the medial and lingual edges exhibit a projection that corresponds to the curvature of the first and the base of the fifth denticulations (Figure 3H-I). Enamel is arranged along the mediolingual and labial edges, forming distinct bands.
Wear resulting from the occlusion of the jaws is evident on the plateau area, the posterior facets of the denticulations, and the last denticulations (Figure 3G, K-M). The plateau area may either be flat or concave (Figure 3K-L). The posterior facets of the denticulations experience wear and enamel loss (Figure 3M). The furrows deepen, and the last denticulation, involving the fourth, forms a step (Figure 3G). On the occlusal surface, impacted by the action of the jaw, a wear pattern, characterized by clustered pits and pronounced acute ridges, emerges (Figure 3E, I-J). This pattern is masked in the larger tooth plate (Figure 3D). Only one specimen displays cusps on the ridges of the occlusal surface (Figure 3I), while others have cusps on the crests.
 Prearticular tooth plates and associated bones. The prearticular bone is not fully preserved in any of the specimens and exhibits a texture the same as that of the pterygopalatine bone (Figure 4A-C). On the ventral and labial side, the Ruge’s canal is divided by a ridge positioned at the level of the first cleft (Figure 4D). Unfortunately, the symphysis remains unpreserved in all specimens. The prearticular tooth plate is fused dorsally to the prearticular bone. It has four denticulations, and its surface is adorned with punctuations (Figure 4A, E). Both the denticulations and the clefts share similar lengths and depths, with the first denticulation being slightly longer and straight (Figure 4A, F-G). All the ridges tend to curve anteriorly (Figure 4A, F). The inner angle is positioned slightly anterior to the second denticulation (Figure 4A, G-H).
Prearticular tooth plates and associated bones. The prearticular bone is not fully preserved in any of the specimens and exhibits a texture the same as that of the pterygopalatine bone (Figure 4A-C). On the ventral and labial side, the Ruge’s canal is divided by a ridge positioned at the level of the first cleft (Figure 4D). Unfortunately, the symphysis remains unpreserved in all specimens. The prearticular tooth plate is fused dorsally to the prearticular bone. It has four denticulations, and its surface is adorned with punctuations (Figure 4A, E). Both the denticulations and the clefts share similar lengths and depths, with the first denticulation being slightly longer and straight (Figure 4A, F-G). All the ridges tend to curve anteriorly (Figure 4A, F). The inner angle is positioned slightly anterior to the second denticulation (Figure 4A, G-H).
Both the medial and lingual edges tend to be straight; however, they curve at the level of the inner angle. Abrupt changes in direction occur towards the free portions of the medial and lingual edges, one corresponding to the curvature of the first denticulation and the other to the base of the fourth denticulation (Figure 4A, F-H).
Similar to pterygopalatine tooth plates, wear occurs in the same regions due to jaw action. The plateau area becomes convex (occluding in the concavity of pterygopalatine tooth plates, Figure 4I), the furrows deepen, and the fourth denticulation forms a bilobed spur (Figure 4A, F, J). On the anterior side of the denticulations, a wear facet is generated due to jaw action (Figure 4F-G). In certain cases, a posterior wear facet is present (Figure 4K-M). The wear pattern is the same as in pterygopalatine tooth plates. Some prearticular tooth plates have cusps on the ridges and crests (Figure 4L-M).
Histological Description
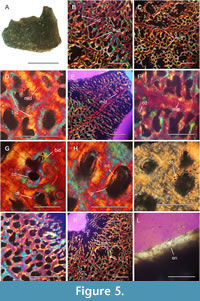 Horizontal cross-sections: section made in the middle part of the tooth plate (Figure 5A). A discernible disorganization characterizes the pattern of interdenteonal dentine (Figure 5B-D). The birefringent interdenteonal dentine surrounds the denteons, deviating from the formation of a cross-pattern. A row-like structure of monorefringent interdenteonal dentine is found within this disarray (Figure 5D-F). Originating from the center of each denticulation (Figure 5E-F), these rows of monorefringent interdenteonal dentine dendritically extend towards the plateau area, positioning themselves between the denteons and the surrounding birefringent interdenteonal dentine (Figure 5B-C).
Horizontal cross-sections: section made in the middle part of the tooth plate (Figure 5A). A discernible disorganization characterizes the pattern of interdenteonal dentine (Figure 5B-D). The birefringent interdenteonal dentine surrounds the denteons, deviating from the formation of a cross-pattern. A row-like structure of monorefringent interdenteonal dentine is found within this disarray (Figure 5D-F). Originating from the center of each denticulation (Figure 5E-F), these rows of monorefringent interdenteonal dentine dendritically extend towards the plateau area, positioning themselves between the denteons and the surrounding birefringent interdenteonal dentine (Figure 5B-C).
In the center of the denticulation, the monorefringent row of interdenteonal dentine is thicker, gradually thinning towards the sides and in the plateau area (Figure 5E). The denteons in the plateau area assume a non-rounded shape, exhibiting a kidney-shaped form. Typically, they are surrounded by dentinal tubules with a granular texture (Figure 5G-I) and are smaller in diameter in the interdenteonal dentine rows. Surrounding the denteons is the circumdenteonal dentine, appearing birefringent under the lambda filter at certain points (Figure 5G). In other regions, the circumdenteonal dentine lacks birefringence under the lambda filter and assumes a silver hue with a polarized light (Figure 5H-I). Despite this variation, the concentric arrangement of fibers remains evident.
Transitional denteons emerge towards the region close to the inner angle and in the clefts, forming dense clusters (Figure 5J-K). Externally, a row of birefringent enamel is observed, and there is no evidence of mantle dentine (Figure 5L). Towards the denticulations, the transitional denteons attached to the edge extend to the central area of the denticulation (Figure 5E). In this area, it is not possible to distinguish between mantle dentine and interdenteonal dentine.
 Vertical cross-sections. The denteons emerge in an occluso-pulp direction from a specific point. In one of the cross-sections, they exhibit some branching (Figure 5A-B). On the other, they are discontinuous and linear (Figure 6C). Between the denteons, birefringent interdenteonal dentine is arranged in two bands with diffuse boundaries (Figure 6D). A wider monorefringent interdenteonal dentine band is positioned between these bands. Towards the sides of the denteons, the circumdenteonal dentine is located (Figure 6D-F). The latter appears transparent in the vertical cross-sections and is crossed by numerous dentinal tubules. At their base, the dentinal tubules contain granules. Very little bone tissue is observed in all sections, with only one primary osteon observed. A total of approximately 9/10 incremental lines were counted (Figure 6A).
Vertical cross-sections. The denteons emerge in an occluso-pulp direction from a specific point. In one of the cross-sections, they exhibit some branching (Figure 5A-B). On the other, they are discontinuous and linear (Figure 6C). Between the denteons, birefringent interdenteonal dentine is arranged in two bands with diffuse boundaries (Figure 6D). A wider monorefringent interdenteonal dentine band is positioned between these bands. Towards the sides of the denteons, the circumdenteonal dentine is located (Figure 6D-F). The latter appears transparent in the vertical cross-sections and is crossed by numerous dentinal tubules. At their base, the dentinal tubules contain granules. Very little bone tissue is observed in all sections, with only one primary osteon observed. A total of approximately 9/10 incremental lines were counted (Figure 6A).
DISCUSSION
Remarks on the Species Chaoceratodus portezuelensis
Chaoceratodus portezuelensis was identified and described by Apesteguía et al. (2007) based on a single prearticular tooth plate from the Portezuelo Formation. Certain characteristics noted in the prior specific diagnosis warrant detailed discussion to elucidate their anatomical attributes. The holotype of C. portezuelensis, initially categorized as a pterygopalatine tooth plate, is revealed to be a right prearticular tooth plate with four denticulations. The presence of four denticulations aligns with other Mesozoic prearticular tooth plates from Argentina, all of which, up to this point, exhibit the same number of denticulations (Wichmann, 1924, 1927; Pascual and Bondesio, 1976; Cione, 1987; Apesteguía et al., 2007; Cione et al., 2007; Agnolin, 2010; Panzeri et al., 2020; 2022a; Agnolín et al., 2024; Panzeri, 2024). This tooth plate displays the second shortest denticulation in length, attributed to an anomaly of short denticulations (Kemp, 2003). It also shows pronounced wear with denticles along the crests. The original diagnosis describes the combination of these two features as “non-ornamented tubercles”. However, this significant wear may stem from malocclusion, attributed to anomalies within the tooth plate and potential irregularities in the opposing one. Such occurrences are a prevalent phenomenon in the dentition of these fishes (Kemp, 2003; Panzeri and Muñoz, 2022). The evidence of anomalies is evident in the holotype of Chaoceratodus portezuelensis, as healthy specimens with similar anatomical structures have been identified and attributed to the same species herein.
Conversely, the anomalous tooth plate reported by Apesteguía et al. (2007) from the same locality aligns with a pterygopalatine bone. The described bulge represents the base of the pterygopalatine ascending process, which is fragmented and positioned at the level of the second denticulation, consistent with other Argentine species. From the same formation and locality, a tooth plate is attributed to the family Ptychoceratodontidae (424), and from the Cerros Colorados locality, another tooth plate is referred to the species Atlantoceratodus iheringi (572). Both prearticular tooth plates are herein assigned to the species Chaoceratodus portezuelensis due to shared morphological characteristics, such as the presence of a bilobed spur.
The tooth plates from the Cerro Lisandro Formation are more numerous and come from the localities of Cerro Bayo Mesa and Cerros Colorados. Most of them have been named and illustrated in the work of Apesteguía et al. (2007) and assigned either to the families Ptychoceratodontidae (427, 430, 431, 433, 435, 436, 439, 440, 506) or Ceratodontidae (426, 432). Nowadays, the most supported idea is that classical Mesozoic dipnoan families do not represent monophyletic groups (Cavin et al., 2007; Pardo et al., 2010; Kemp et al., 2017). In addition to this, the tooth plates from the Cerro Lisandro Formation share certain morphological features belonging to the species Chaoceratodus portezuelensis. Even the fragmented tooth plates display morphological features and a wear pattern similar to the observed in this species. Therefore, the tooth plates from the Cerro Lisandro Formation are also referred to as C. portezuelensis.
 Salgado et al. (2009) reported a tooth plate (MPCA-AT 11) from the Anfiteatro area, Cerro Lisandro Formation, and provided a brief description of it, including an illustration under a clear camera. This plate is a pterygopalatine tooth plate, and the perforations referred to in their work correspond to the openings of pulp canals or punctuations (Figure 7A-B). The illustration and description provided show that the third denticulation is the largest, which is uncommon for specimens from Argentina (Salgado et al., 2009, fig. 3C-D). This denticulation is reinterpreted here as the second, with the first one being fractured. Additionally, the plateau area is concave in labial view, as commonly observed in this type of tooth plate (Figure 7E-F). In MPCA-AT 11, a pattern similar to that observed in Chaoceratodus portezuelensis is identified. Therefore, despite it being highly fragmented and based on its characteristic external pattern, this right pterygopalatine tooth plate is tentatively identified as cf. Chaoceratodus portezuelensis.
Salgado et al. (2009) reported a tooth plate (MPCA-AT 11) from the Anfiteatro area, Cerro Lisandro Formation, and provided a brief description of it, including an illustration under a clear camera. This plate is a pterygopalatine tooth plate, and the perforations referred to in their work correspond to the openings of pulp canals or punctuations (Figure 7A-B). The illustration and description provided show that the third denticulation is the largest, which is uncommon for specimens from Argentina (Salgado et al., 2009, fig. 3C-D). This denticulation is reinterpreted here as the second, with the first one being fractured. Additionally, the plateau area is concave in labial view, as commonly observed in this type of tooth plate (Figure 7E-F). In MPCA-AT 11, a pattern similar to that observed in Chaoceratodus portezuelensis is identified. Therefore, despite it being highly fragmented and based on its characteristic external pattern, this right pterygopalatine tooth plate is tentatively identified as cf. Chaoceratodus portezuelensis.
Morphological and Histological Comparison with Dipnoan Species from Patagonia
The species Chaoceratodus portezuelensis possesses unique characteristics that differentiate it from other Patagonian dipnoans. In the pterygopalatine tooth plates, the first denticulation is short with a wear facet near the apex, the remaining denticulations are of similar sizes, while the last one forms the step, and the edges are slightly curved at the inner angle, which is positioned behind the second denticulation. Whereas in the prearticular tooth plates, the first denticulation is short, the second denticulation may exhibit a posterior wear facet, a bilobed spur, and relatively straight edges with an inner angle positioned at the level of the first cleft or slightly ahead of it at the level of the second denticulation. The characteristic of being able to present a posterior wear facet in prearticular tooth plates is unique since, usually, in prearticular tooth plates from the Mesozoic of Argentina, wear facets are only present anteriorly on denticulations. One of the outstanding characteristics of the species is the arrangement of interdenteonal dentine in rows that reflect externally as a pattern on the occlusal surface. The rows of monorefringent interdenteonal dentine coincide with the denticulations and anastomose over the rest of the plateau, appearing as an externally reticulated pattern.
In comparison with the remaining Patagonian species, Chaoceratodus portezuelensis differs from Metaceratodus kaopen (Apesteguía et al., 2007; Cione and Gouiric Cavalli, 2012) in having a larger plateau area, with an acute inner angle between the first and last ridges. Additionally, the pterygopalatine tooth plates of M. kaopen have a wear facet anterior to the first denticulation that reaches the inner angle (Figure 7E). Metaceratodus kaopen also displays a more obtuse inner angle and a step formed by the last two denticulations, which, when viewed occlusally, adopts a square shape (Figure 7F). The prearticular tooth plates exhibit a first denticulation with a wide base, occasionally displaying a projection at the base level, which is absent in the species described here (Figure 7G). Chaoceratodus portezuelensis differs from M. wichmanni and M. baibianorum in several respects. It possesses a shorter first denticulation and a more pronounced curve along the mediolingual edge at the inner angle. Moreover, it has a smaller plateau area and a slightly deeper first cleft than the one found in the latter species. M. wichmanni has pterygopalatine tooth plates with a similar plateau area, a step formed by the last denticulation, and a second cleft deeper than the others (Apesteguía et al., 2007; Figure 7H-I); similar features are observed in the species Atlantoceratodus patagonicus (Agnolin, 2010). The prearticular tooth plates of M. wichmanni have a simple spur, and the first denticulation is longer with a narrow base (Figure 7J). The other species, M. baibianorum, has pterygopalatine tooth plates with wider plateau areas, slightly curved mediolingual edges, and prearticular tooth plates with a simple spur (Panzeri et al., 2020; Figure 7K-M).
Chaoceratodus portezuelensis also differs from Atlantoceratodus iheringi by featuring a shorter first denticulation on the pterygopalatine tooth plates along with an anterior wear facet. In contrast, A. iheringi showcases a longer first denticulation on the pterygopalatine and prearticular tooth plates, devoid of an anterior wear facet (Cione et al., 2007; Panzeri, 2024; Figure 7N-P). The species Ceratodus argentinus, is based by a single left pterygopalatine tooth plate, which shows signs of wear from transportation and lacks the first denticulation. This species is nominated in the same study by Apesteguía et al. (2007), and shares similarities with the pterygopalatine tooth plates of C. portezuelensis when they attain larger sizes (Figure 3D). However, the absence of material referred to C. argentinus complicates direct comparison.
Finally, Chaoceratodus portezuelensis differs from Rinconodus salvadori in that its first denticulation is short, relatively robust and with an anterior wear facet. In contrast, R. salvadori features a longer, posteriorly curved first denticulation with a raised medial edge forming a ridge and a markedly deep posterior wear facet (Panzeri et al., 2022a; Figure 7Q-S). The medial edge of R. salvadori curves smoothly without prominences and lacks anterior wear facets (Figure 7Q). Additionally, in R. salvadori, the prearticular tooth plates exhibit a pronounced anterior wear facet in the first denticulation (Figure 7S) without posterior wear facets in the remaining ones.
 Concerning the dental histology of lungfish from Patagonia, detailed descriptions have only been provided for histological cross-sections of tooth plates attributed to Metaceratodus baibianorum (Panzeri et al., 2022b) and Atlantoceratodus iheringi (Panzeri, 2024). The histology of Chaoceratodus portezuelensis differs in the arrangement of dentine. The interdenteonal dentine is arranged in rows of monorefringent dentine, coinciding with the ridges that anastomose towards the plateau area. Between these rows and the denteons, the interdenteonal dentine is arranged more erratically, forming patches of monorefringent and birefringent areas. The denteons have kidney-shaped forms, and the circumdenteonal dentine appears to be arranged in a single band. In M. baibianorum, the interdenteonal dentine is laid out in an ordered cross pattern surrounding small, circular denteons (Figure 8A-C; Panzeri et al., 2022b). The circumdenteonal dentine in M. baibianorum and C. portezuelensis is simple and arranged in a single band (Figure 8A-B). Finally, although in A. iheringi, the interdenteonal dentine forms disorganized patches, certain areas exhibit an ordered cross pattern similar to M. baibianorum (Figure 8D-E; Panzeri, 2024). Also, in A. iheringi, the circumdenteonal dentine is arranged in a double band, and the denteons are usually elongated and grouped (Figure 8D-F).
Concerning the dental histology of lungfish from Patagonia, detailed descriptions have only been provided for histological cross-sections of tooth plates attributed to Metaceratodus baibianorum (Panzeri et al., 2022b) and Atlantoceratodus iheringi (Panzeri, 2024). The histology of Chaoceratodus portezuelensis differs in the arrangement of dentine. The interdenteonal dentine is arranged in rows of monorefringent dentine, coinciding with the ridges that anastomose towards the plateau area. Between these rows and the denteons, the interdenteonal dentine is arranged more erratically, forming patches of monorefringent and birefringent areas. The denteons have kidney-shaped forms, and the circumdenteonal dentine appears to be arranged in a single band. In M. baibianorum, the interdenteonal dentine is laid out in an ordered cross pattern surrounding small, circular denteons (Figure 8A-C; Panzeri et al., 2022b). The circumdenteonal dentine in M. baibianorum and C. portezuelensis is simple and arranged in a single band (Figure 8A-B). Finally, although in A. iheringi, the interdenteonal dentine forms disorganized patches, certain areas exhibit an ordered cross pattern similar to M. baibianorum (Figure 8D-E; Panzeri, 2024). Also, in A. iheringi, the circumdenteonal dentine is arranged in a double band, and the denteons are usually elongated and grouped (Figure 8D-F).
Morphological and Histological Comparison with Dipnoan Species from Other Regions of the World
Regarding lungfish from other parts of the world, a relationship has been previously established between Patagonian lungfish and those from Madagascar, specifically with the species Ferganoceratodus madagascariensis (Priem, 1924; Martin, 1981b, 1982; Schultze, 1991; Martin et al., 1999). The similarity lies in this species and the Patagonian lungfish sharing the same number of denticulations, with medial edges longer than the lingual edges and similarly curved. Conversely, the remaining species of the genus Ferganoceratodus generally have a medial edge shorter than the lingual edge. Both edges are straight, unlike what is observed in Patagonian species. A possible close relationship has also been established between F. madagascariensis (from Madagascar), the Patagonian lungfish Chaoceratodus portezuelensis (Apesteguía et al., 2007), Rinconodus salvadori (Panzeri et al., 2022a), and Atlantoceratodus iheringi (Martin, 1981b, 1982; Martin et al., 1999; Panzeri 2024), and North American lungfish (Fernández et al., 1973; Martin, 1981b; 1982; Schultze, 1991; Martin et al., 1999). Particularly C. portezuelensis broadly resembles lungfish from Jurassic and Cretaceous formations in North America (e.g., Potamoceratodus guentheri; Apesteguía et al., 2007) in possessing pterygopalatine tooth plates with a wide plateau area relative to the area of the denticulations, medial edges longer than the lingual edges and equally curved, and prearticular tooth plates with the same number of denticulations and longer labial edges than lingual edges. However, they differ from C. portezuelensis in that the latter has the ascending pterygopalatine process at the level of the second denticulation with a circular base, a shorter first denticulation with an anterior wear facet, and prearticular tooth plates with a bilobed spur.
Recently, Challands et al. (2024) proposed a new taxonomic scheme in which all tooth plates with five denticulations on the upper tooth plate and four on the lower would be assigned to Ferganoceratodus. We opted to follow traditional classifications until a more solid taxonomic framework becomes available. This is because the diagnosis of Ferganoceratodus is primarily based on cranial characters, and according to Kemp (1997) and Kemp et al. (2017, suppl. data), the number of denticulations alone “is unreliable” and “unhelpful for most Mesozoic and Cenozoic tooth plates”. Additionally, in Challands et al. (2024) comparisons, many tooth plates with the 5/4 configuration are excluded from this genus due to other morphological and distinctive features of the dentition, which alters the criteria for reassignment to Ferganoceratodus and indicating that this proposal requires improvements (e.g., in species based on tooth plates from North America and Patagonia).
It is important to note that, to date, all pterygopalatine tooth plates from Patagonia have a medial edge that is longer than the lingual edge, placing the inner angle in a posteromedial position (Figure 7; Panzeri et al., 2020). This feature differs from all tooth plates assigned to Ferganoceratodus (except F. madagascariensis, see above). In Challands et al. (2024), this feature is mentioned only for Patagonian species assigned to Metaceratodus, raising doubts about their reassignment to Ferganoceratodus. Furthermore, among Patagonian species, Rinconodus salvadori is omitted from the discussion (Panzeri et al., 2022a), representing the first Mesozoic dipnoan with a preserved cranial roof from South America. The cranial bone configuration of R. salvadori is more similar to the North American species Potamoceratodus guentheri (Marsh, 1878) than to Ferganoceratodus and Ptychoceratodus. As mentioned earlier, this does not rule out a potential close phylogenetic relationship between the species from Africa (Madagascar) and Patagonia.
Detailed histological studies predominantly using polarized light with a lambda filter have been primarily conducted on the dentitions of Paleozoic dipnoans from other parts of the world. The arrangement of tissues in Paleozoic dipnoans varies, generating patterns not observed in Patagonian dipnoans (Barwick et al., 1997; den Blaauwen et al., 2006). For example, in Dipterus valenciennesi, the pulp canals are surrounded by thick monorefringent dentine, with birefringent dentine forming a square pattern between them. On the other hand, in Tarachomylax oepiki, although a lambda filter has not been used and the cross-sections are made in a different direction, the dentine surrounding the pulp canal is also thicker than the dentine between them. Similar observations are made in Chirodipterus australis, where the pulp canals often merge, forming a particular mesh (Smith and Campbell, 1987), as also seen in Sagenodus inequalis (Smith, 1979). In Adolopas moyasmithae, the dentition and tissue arrangement resemble those of the aforementioned species, with opaque dentine coinciding with the crests (interpreted as petrodentine; Campbell and Barwick, 1998), while this dense tissue is not observed towards the sides. Additionally, Mesozoic dipnoans from other parts of the world exhibit a tooth plate histology similar to that observed in Chaoceratodus portezuelensis. As mentioned earlier, C. portezuelensis shows a distinctive arrangement of tissues in the horizontal cross-section, externally manifested as rows coinciding with the ridges and anastomosing towards the sides. The arrangement of interdenteonal dentine in monorefringent rows coinciding with the ridges resembles the petrodentine found in lepidosirenids (Bemis, 1984) and certain species of Mioceratodus (Kemp, 2001). However, this dentine anastomoses forms monorefringent rows also towards the plateau area, including some small lumen denteons, suggesting that it would not form from the remnants of core dentine, as it occurs in petrodentine (Lison, 1941; Kemp, 2001).
Other Mesozoic species resembling histology present in Chaoceratodus portezuelensis include Neoceratodus forsteri, N. eyrensis, and Ferganoceratodus madagascariensis (Smith, 1984; Kemp, 2001). In the horizontal cross-sections of these species, interdenteonal dentine is organized in a cross pattern. On the other hand, in the vertical cross-sections, a clearer organization of monorefringent interdenteonal dentine is observed between the two bands of birefringent interdenteonal dentine (Smith, 1984), with the denteons being parallel to each other and not converging occlusally. Finally, in F. edwardsi, the vertical cross-sections resemble those of Ceratodus tuberculatus (Tabaste, 1963) where the denteons converge towards the occlusal surface and delineate areas of dentine between them (Challands et al., 2024). Although horizontal cross-sections have not been performed, the micro-CT reveals a tissue arrangement that reflects external wear patterns, which differ significantly from those observed in Patagonian species. that observed in Ferganoceratodus martini (Cavin et al., 2007) and in African species such as C. tuberculatus, C. parvus, C. africanus, and Lavocatodus humei (Foureau, 1907; Tabaste, 1963; Churcher and de Iuliis, 2001; Claeson et al., 2014).
CONCLUSIONS
In conclusion, Chaoceratodus portezuelensis displays distinctive features not found in lungfish from other regions, including Patagonia. These features, such as a medial edge longer than the lingual edge, a short first denticulation in pterygopalatine tooth plates with an anterior wear facet and prearticular tooth plates with a bilobed spur, along with a unique histological pattern, define the species. This contribution provides a diagnosis of the species that differentiates features present in pterygopalatine and prearticular tooth plates as they vary according to the type of tooth. Furthermore, our findings underscore the morphological similarities observed between Patagonian lungfish, the Madagascan species Ferganoceratodus madagascariensis, and North American lungfish. Lastly, we emphasize the importance of tissue arrangement as a valuable tool for future systematic and phylogenetic studies alongside detailed morphological descriptions.
ACKNOWLEDGEMENTS
We extend our heartfelt appreciation to CONICET for funding the present research, to M.E. Pereyra for preparing the histological sections, to the staff of the ichnology laboratory (MACN) for facilitating the use of the petrographic microscope, to the staff of the Museo Carmen Funes, particularly L. and R. Coria, and to the staff of the Museo Carlos Ameghino for their invaluable assistance. Additionally, we express our gratitude to A.C. Garrido, F. Agnolin and F. Novas for providing valuable information. Special thanks to the editor and the anonymous reviewers for their valuable comments.
REFERENCES
Agnolin, F. 2010. A new species of the genus Atlantoceratodus (Dipnoiformes: Ceratodontoidei) from the Uppermost Cretaceous of Patagonia and a brief overview of fossil dipnoans from the Cretaceous and Paleogene of South America. Brazilian Geographical Journal: Geosciences and Humanities Research Medium, 1(2):162–210.
Agnolin, F.L., Novas, F.E., and Lio, G. 2006. Neornithine bird coracoid from the Upper Cretaceous of Patagonia. Ameghiniana, 4(1):245–248.
Agnolín, F.L., Herrera, G.Á., Rolando, M.A., Motta, M., Rozadilla, S., Verdiquio, L., D´Angelo, J., Moyano-Paz, D., Varela, A., Sterli, J., Bogan, S., Miner, S., Moreno Rodriguez, A., Muñoz, G., Isasi, M., and Novas, F.E. 2024. Fossil vertebrates from the Cerro Fortaleza Formation (Upper Cretaceous), Santa Cruz Province, Argentina. Cretaceous Research, 154:105735.
https://doi.org/10.1016/j.cretres.2023.105735
Ahlberg, P.E., Smith, M.M., and Johanson, Z. 2006. Developmental plasticity and disparity in early dipnoan (lungfish) dentitions. Evolution & Development, 8(4):331–349.
https://doi.org/10.1111/j.1525-142X.2006.00106.x
Antunes, M.T., Maisey, J.G., Marques, M.M., Schaeffer, B., and Thomson, K.S. 1990. Triassic fishes from the Cassange Depression (RP de Angola). Ciências da Terra (UNL), Número Especial, 1–64.
Apesteguía, S., Agnolin, F., and Claeson, K. 2007. Review of Cretaceous dipnoans from Argentina (Sarcopterygii: Dipnoi) with descriptions of new species. Revista del Museo Argentino de Ciencias Naturales nueva serie 9(1):27–40.
Barrios, F., Paulina-Carabajal, A., and Bona, P. 2016. A new peirosaurid (Crocodyliformes, Mesoeucrocodylia) from the Upper Cretaceous of Patagonia, Argentina. Ameghiniana, 53(1): 14–25.
https://doi.org/10.5710/AMGH.03.09.2015.2903
Barwick, R.E., Campbell, K.S., and Mark-Kurik, E. 1997. Tarachomylax: A new Early Devonian dipnoan from Severnaya Zemlya, and its place in the evolution of the Dipnoi. Geobios, 30(1):45–73.
https://doi.org/10.1016/S0016-6995(97)80257-4
Bemis, W.E. 1984. Morphology and growth of lepidosirenid lungfish tooth plates (Pisces: Dipnoi). Journal of Morphology, 179:73–93.
https://doi.org/10.1002/jmor.1051790108
Calvo, J.O. and Salgado, L. 2022. El Museo de Geología y Paleontología de la Universidad Nacional del Comahue. Publicación Electrónica de la Asociación Paleontológica Argentina, 22(1):309–317.
Campbell, K.S.W. and Barwick, R.E. 1998. A new tooth-plated dipnoan from the Upper Devonian Gogo Formation and its relationships. Memoirs Queensland Museum, 42:403–437.
Cavin, L. 2017. Freshwater fishes: 250 million years of evolutionary history. ISTE Press-Elsevier, London and Oxford.
Cavin, L., Suteethorn, V., Buffetaut, E., and Tong, H. 2007. A new Thai Mesozoic lungfish (Sarcopterygii, Dipnoi) with an insight into post-Palaeozoic dipnoan evolution. Zoological Journal of the Linnean Society, 149(2):141–177.
https://doi.org/10.1111/j.1096-3642.2007.00238.x
Cavin, L., Deesri, U., and Chanthasit, P. 2020. A new lungfish from the Jurassic of Thailand. Journal of Vertebrate Paleontology 40:e1791895.
https://doi.org/10.1080/02724634.2020.1791895
Cerda, I.A., Pereyra, M.E., Garrone, M.C., Ponce, D., Navarro, T.G., González, R., Militello, M., Luna, C.A., and Jannello, J.M. 2020. A basic guide for sampling and preparation of extant and fossil bones for histological studies. Publicación Electrónica de la Asociación Paleontológica Argentina, 20.
Challands, T.J., Cavin, L., Zondo, M., Munyikwa, D., Choiniere, J.N., and Barrett, P.M. 2024. A new lungfish from the Upper Triassic of the Mid-Zambezi Basin, Zimbabwe. Journal of Vertebrate Paleontology, e2365391.
https://doi.org/10.1080/02724634.2024.2365391
Churcher, C.S. and De Iuliis, G. 2001. A new species of Protopterus and a revision of Ceratodus humei (Dipnoi: Ceratodontiformes) from the Late Cretaceous Mut Formation of eastern Dakhleh Oasis, Western desert of Egypt. Palaeontology, 44(4):305–323.
https://doi.org/10.1111/1475-4983.00181
Cione, A.L. 1987. The Late Cretaceous fauna of los Alamitos, Patagonia, Argentina. II: The fishes. Revista del Museo Argentino de Ciencias Naturales Bernardino Rivadavia e Instituto Nacional de Investigación de las Ciencias Naturales. Paleontología, 3:111–120.
Cione, A.L., Gouiric Cavalli, S., Goin, F J., and Poiré, D.G. 2007. Atlantoceratodus a new genus of lungfish from the upper Cretaceous of South America and Africa. Revista del Museo de La Plata, 10.
Cione, A.L. and Gouiric-Cavalli, S. 2012. Metaceratodus kaopen comb. nov. and M. wichmanni comb. nov., two Late Cretaceous South American species of an austral lungfish genus (Dipnoi). Alcheringa: An Australasian Journal of Palaeontology, 36(2):203–216.
Clack, J.A., Sharp, E.L., Long, J.A., Jørgensen, J.M., and Joss, J. 2011. The fossil record of lungfishes, p.1–42. In Jørgensen, J.M. and Joss, J. (eds.), The Biology of Lungfishes. Science Publishers, Enfield, USA.
https://doi.org/10.1080/03115518.2012.621804
Clack, J.A. and Ahlberg, P.E. 2016. Sarcopterygians: from lobe-finned fishes to the tetrapod stem group, p. 51–70. In Clack, J.A., Fay, R., and Popper, A.N. (eds.), Evolution of the Vertebrate Ear. Springer Handbook of Auditory Research, vol. 59. Springer, Cham.
https://doi.org/10.1007/978-3-319-46661-3_3
Claeson, K.M., Sallam, H.M., O’Connor, P.M., and Sertich, J.J. 2014. A revision of the Upper Cretaceous lepidosirenid lungfishes from the Quseir Formation, Western Desert, central Egypt. Journal of Vertebrate Paleontology, 34:760–766.
https://doi.org/10.1080/02724634.2014.838574
Clement, A.M. 2019. Sarcopterygian Fishes, the “Lobe-Fins”, p. 119–142. In Ziermann, J., Diaz Jr., R.E., and Diogo, R. (eds.), Heads, Jaws, and Muscles. Fascinating Life Sciences. Springer, Cham.
https://doi.org/10.1007/978-3-319-93560-7_6
Cloutier, R. 1996. Dipnoi (Akinetia: Sarcopterygii), p. 19–226. In Schultze, H.P. and Cloutier, R. (eds.), Devonian Fishes and Plants of Miguasha, Quebec. Verlag Dr. Freidrich Pfeil, München.
Coria, R.A. 2022. Pequeña historia de la paleoherpetología en el museo Carmen Funes de Plaza Huincul (Neuquén, Argentina): Hechos y protagonistas. Publicación Electrónica de la Asociación Paleontológica Argentina, 22(1):326–334.
https://doi.org/10.5710/PEAPA.26.05.2021.347
Coria, R.A., Cladera, G., and Salgado, L. 1996. Sobre una nueva localidad fosilífera en la Formación Río Limay? Cretácico Superior temprano, en la localidad de Cerro Bayo Mesa, Provincia de Neuquén. Ameghiniana, 33(4):463.
Coria, R.A. and Calvo, J.O. 2002. A new iguanodontian ornithopod from Neuquén Basin, Patagonia, Argentina. Journal of Vertebrate Paleontology, 22(3):503–509.
https://doi.org/10.1671/0272-4634(2002)022[0503:ANIOFN]2.0.CO;2
Coria, R.A., Currie, P.J., and Carabajal, A.P. 2006. A new abelisauroid theropod from northwestern Patagonia. Canadian Journal of Earth Sciences, 43(9):1283–1289.
https://doi.org/10.1139/e06-025
de La Fuente, M.S. 2003. Two new pleurodiran turtles from the Portezuelo Formation (Upper Cretaceous) of northern Patagonia, Argentina. Journal of Paleontology, 77(3):559–575.
https://doi.org/10.1666/0022-3360(2003)0772.0.CO;2
den Blaauwen, J.L., Barwick, R.E., and Campbell, K.S. 2006. Structure and function of the tooth plates of the Devonian lungfish Dipterus valenciennesi from Caithness and the Orkney Islands. Records of the Western Australian Museum, 23(1):91.
https://doi.org/10.18195/issn.0312-3162.23(1).2006.091-113
Fernández, J., Bondesio, P., and Pascual, R. 1973. Restos de Lepidosiren paradoxa (Osteichthyes, Dipnoi) de la Formación Lumbrera (Eogeno, ¿Eoceno?) de Jujuy. Ameghiniana, 10(2):152–172.
Foureau, F. 1907. Documents scientifiques de la Mission Saharienne Mission Foureau-Lamy `D'Alger au Congo par le Tchad'. Paris, Masson et Cie, 3:551–1210.
Frederickson, J.A. and Cifelli, R.L. 2017. New Cretaceous lungfishes (Dipnoi, Ceratodontidae) from western North America. Journal of Paleontology, 91:146–161.
https://doi.org/10.1017/jpa.2016.131
Garrido, A.C. 2010. Estratigrafía del Grupo Neuquén, Cretácico Superior de la Cuenca Neuquina (Argentina): nueva propuesta de ordenamiento litoestratigráfico. Revista del Museo Argentino de Ciencias Naturales, 12(2):121–177.
Garrido, A.C. 2011. El Grupo Neuquén (Cretácico Tardío) en la Cuenca Neuquina, p. 231–244. In Leanza, H.A., Arregui, C., Carbone, O., Danieli, J.C., and Vallés, J.M. (eds.), Geología y Recursos Naturales de la provincia del Neuquén. 18º Congreso Geológico Argentino, Relatorio.
Gill, T. 1872. Arrangement of the families of fishes, or classes Pisces, Marsipobranchii and Leptocardii. Smithsonian Miscellaneous Collections, 247.
Howell, J.A., Schwarz, E., Spalletti, L.A., and Veiga, G.D. 2005. The Neuquén basin: an overview, p. 1–14. In Veiga, G.D., Spalletti, L.A., Howell, J.A., and Schwarz, E. (eds.), The Neuquén Basin, Argentina: A Case Study in Sequence Stratigraphy and Basin Dynamics. Geological Society Special Publications No. 252. The Geological Society, London.
Kemp, A. 1977. The pattern of tooth plate formation in the Australian lungfish, Neoceratodus forsteri Krefft. Zoological Journal of the Linnean Society, 60(3):223–258.
https://doi.org/10.1111/j.1096-3642.1977.tb01028.x
Kemp, A. 1994. Australian Triassic lungfish skulls. Journal of Paleontology, 68: 647–654.
https://doi.org/10.1017/S0022336000025968
Kemp, A. 1997. A revision of Australian Mesozoic and Cenozoic lungfish of the family Neoceratodontidae (Osteichthyes: Dipnoi), with a description of four new species. Journal of Paleontology, 71(4):713–733.
https://doi.org/10.1017/S0022336000040166
Kemp, A. 2001. Petrodentine in derived dipnoan tooth plates. Journal of Vertebrate Paleontology, 21:422–437.
https://doi.org/10.1671/0272-4634(2001)021[0422:PIDDTP]2.0.CO;2
Kemp, A. 2003. Developmental anomalies in the tooth plates and jaw bones of lungfish. Journal of Vertebrate Paleontology, 23(3):517–531.
https://doi.org/10.1671/1868
Kemp, A., Cavin, L., and Guinot, G. 2017. Evolutionary history of lungfishes with a new phylogeny of post-Devonian genera. Palaeogeography, Palaeoclimatology, Palaeoecology, 471:209–219.
https://doi.org/10.1016/j.palaeo.2016.12.051
Legarreta, L. and Uliana, M.A. 1996. The Jurassic succession in west-central Argentina: stratal patterns, sequences and paleogeographic evolution. Palaeogeography, Palaeoclimatology, Palaeoecology, 120(4):303–330.
https://doi.org/10.1016/0031-0182(95)00042-9
Lison, L. 1941. The structure of teeth of lungfish. Petrodentine. Comptes Rendus des Seances de la Societe de Biologie et de Ses Filiales, 135:431.
Marsh O.C. 1878. A new species of Ceratodus from the Jurassic. American Journal of Science, 1:76.
Martin, M. 1981a. Les dipneustes et Actinistiens du Trias supérieur continental marocain. Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie), 69:1–29.
Martin, M. 1981b. Les Dipneustes mesozoiques malgaches, leurs affinites et leur interet paléobiogeographique. Bulletin de la Société géologique de France, 7:579–585.
https://doi.org/10.2113/gssgfbull.S7-XXIII.6.579
Martin, M. 1982. Nouvelles données sur la phylogénie et la systémique des dipneustes postpaléozoïques, conséquences stratigraphiques et paléogéographiques. Geobios, Mémoire Spécial, 6:53–64.
https://doi.org/10.1016/S0016-6995(82)80102-2
Martin, M., Barbieri, L., and Cuny, G. 1999. The Madagascan Mesozoic ptychoceratodontids (Dipnoi) systematic relationships and paleobiogeographical significance. Oryctos, 2:3–16.
Martínez, L.C. and Lutz, A.I. 2007. Especies nuevas de Baieroxylon Greguss y Circoporoxylon Kräusel en las formaciones Rayoso y Huincul (Cretácico), provincia del Neuquén, Argentina. Ameghiniana, 44(3):537–546.
Müller, J. 1845. Über den Bau und die Grenzen der Ganoiden und liber das natürliche System der Fische. Abhandlungen der Akademie der Wissenschaften zu Berlin, 1844:117–216.
Novas, F.E. 1996. Alvarezsauridae, Cretaceous basal birds from Patagonia and Mongolia. Memoirs-Queensland Museum, 39:675–702.
Panzeri, K.M. 2024. Revisiting the Cretaceous lungfish Atlantoceratodus iheringi (Ameghino, 1898) from the Mata Amarilla Formation (Argentina) with comments on tooth plates histology. Rivista Italiana di Paleontologia e Stratigrafia, 130(2):191–209.
https://doi.org/10.54103/2039-4942/21646
Panzeri, K.M., Gouiric Cavalli, S., Muñoz, N.A., and Cione, A.L. 2020. Metaceratodus baibianorum, a new dipnoan species from the Upper Cretaceous of southern South America supported by traditional and geometric morphometric analyses. Journal of Vertebrate Paleontology, 40(2):e1769640.
https://doi.org/10.1080/02724634.2020.1769640
Panzeri, K.M. and Muñoz, N.A. 2022. Metaceratodus baibianorum from the La Colonia Formation: tooth plate anomalies and the possible presence of tertiary dentine. Alcheringa: An Australasian Journal of Palaeontology, 46(2):198–207.
https://doi.org/10.1080/03115518.2022.2078882
Panzeri, K.M., Gouiric Cavalli, S., Cione, A.L., and Fillippi, L. 2022a. Description of the first Cretaceous (Santonian) articulated skeletal lungfish remains from South America, Argentina. Comptes Rendus Palevol, 2(37):815–835.
https://doi.org/10.5852/cr-palevol2022v21a37
Panzeri, K.M., Pereyra, M.E., and Cione, A.L. 2022b. The South American dipnoan Metaceratodus baibianorum (Dipnoi, Ceratodontidae) from the Upper Cretaceous La Colonia Formation, Patagonia, Argentina: an approach from the histology of the tooth plates. Cretaceous Research, 133:105144.
https://doi.org/10.1016/j.cretres.2022.105144
Pardo, J.D., Huttenlocker, A.K., Small, B.J., and Gorman, M.A. 2010. The cranial morphology of a new genus of lungfish (Osteichthyes: Dipnoi) from the Upper Jurassic Morrison Formation of North America. Journal of Vertebrate Paleontology, 30(5):1352–1359.
https://doi.org/10.1080/02724634.2010.501430
Pascual, R. and Bondesio, P. 1976. Notas sobre vertebrados de la Frontera Cretácica-Terciaria. III. Ceratodontidae (peces Osteichthyes, Dipnoi) de la Formación Coli-Toro y de otras unidades del Cretácico tardío de Patagonia y sur de Mendoza. Sus implicancias palaeobiogeográficas. Actas del Sexto Congreso Geológico Argentino, 1:565–577.
Pawlak, W., Talanda, M., Sulej, T., and Niedzwiedzki, G. 2020. Dipnoan from the Upper Triassic of East Greenland and remarks about palaeobiogeography of Ptychoceratodus. Acta Palaeontologica Polonica, 65(3):561–574
https://doi.org/10.4202/app.00679.2019
Priem, F. 1924. Paléontologie de Madagascar XII: les poissons fossiles. Annales de Paléontologie, 23:83–178.
Romer, A.S. 1955. Herpetichthyes, Amphibioidei, Choanichthyes or Sarcopterygii?. Nature, 176:126–127.
https://doi.org/10.1038/176126b0
Salgado, L., Canudo, J.I., Garrido, A.C., Ruiz-Omenaca, J.I., García, R.A., de la Fuente, M.S., Barco, J.L., and Bollati, R. 2009. Upper Cretaceous vertebrates from El Anfiteatro area, Río Negro, Patagonia, Argentina. Cretaceous Research, 30(3):767–784.
https://doi.org/10.1016/j.cretres.2009.01.001
Sánchez, M.L. and Asurmendi, E. 2014. Modelo de depósito de la Formación Cerro Lisandro: lóbulos de desembocadura y deltas de tipo Gilbert. Cretácico superior, región central de cuenca Neuquina, Argentina. Revista mexicana de ciencias geológicas, 31(2):141–162.
Sánchez, M.L., Calvo, J.O., and Heredia, S. 2005. Paleoambientes de sedimentación del tramo superior de la Formación Portezuelo, Grupo Neuquén (Cretácico Superior), Los Barreales, provincia del Neuquén. Revista de la Asociación Geológica Argentina, 60(1):142–158.
Schneider, C.A., Rasband, W.S., and Eliceiri, K.W. 2012. NIH Image to ImageJ: 25 years of image analysis. Nature methods, 9(7):671–675.
https://doi.org/10.1038/nmeth.2089
Schultze, H.P. 1991. Lungfish from the El Molino (Late Cretaceous) and Santa Lucia formations in South-central Bolivia. In Súarez-Soruco, R. (ed.), Fósiles y Facies de Bolivia, Vertebrados, Revista Técnica de YPFB, 12:441–448.
Skrzycki, P. 2015. New species of lungfish (Sarcopterygii, Dipnoi) from the Late Triassic Krasiejów site in Poland, with remarks on the ontogeny of Triassic dipnoan tooth plates. Journal of Vertebrate Paleontology, 35:e964357.
https://doi.org/10.1080/02724634.2015.964357
Smith, M.M. 1979. Structure and histogenesis of tooth plates in Sagenodus inaequalis Owen considered in relation to the phylogeny of post-Devonian dipnoans. Proceedings of the Royal Society of London. Series B. Biological Sciences, 204:15–39.
https://doi.org/10.1098/rspb.1979.0010
Smith, M.M. 1984. Petrodentine in extant and fossil dipnoan dentitions: microstructure, histogenesis and growth. Proceedings of the Linnean Society of New South Wales, 107:367–407.
Smith, M. and Campbell, K.S.W. 1987. Comparative morphology, histology and growth of the dental plates of the Devonian dipnoan Chirodipterus. Philosophical Transactions of the Royal Society of London. B, Biological Sciences, 317:329–363.
https://doi.org/10.1098/rstb.1987.0066
Tabaste, N. 1963 Etude de restes de poissons du Crétacé saharien. Mémoires de l'Institut Français d'Afrique Noire, Mélanges Ichthyol., 68:436–499.
Teller F.J. 1891. Über den Schädel eines Fossilen dipnoërs Ceratodus sturii nov. spec. aus den Schichten der oberen Trias der Nordalpen. Abhandlungen der Kaiserlich-königlichen Geologischen Reichsanstalt, 15(3):1–39.
Wichmann, R. 1924. Nuevas observaciones geológicas en la parte oriental del Neuquén y en el Territorio del Río Negro. Ministerio de Agricultura. Dirección General de Minas, Geología e Hidrología.
Wichmann, R. 1927. Sobre las facies lacustres Senoniana de los estratos con dinosaurios y su fauna. Boletín de la Academia Nacional de Ciencias de Córdoba, 30:383–405.
Yanchao, L., Xindong, C., Tuo Q., and Min, Z. 2021. A new dipnoan genus from the Middle Devonian of Huize, Yunnan, China. Journal of Systematic Palaeontology, 19:1303–1315.
https://doi.org/10.1080/14772019.2022.2042409
