

 Qualitative and quantitative analysis of the graptolite assemblage in the linnaei Biozone (Silurian, lowermost Telychian) at Želkovice, Prague Synform (Czech Republic)
Qualitative and quantitative analysis of the graptolite assemblage in the linnaei Biozone (Silurian, lowermost Telychian) at Želkovice, Prague Synform (Czech Republic)
Article number: 27.3.a58
https://doi.org/10.26879/1425
Copyright Palaeontological Association, December 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 28 June 2024. Acceptance: 27 November 2024.
ABSTRACT
This case study represents a detailed analysis of the graptolite assemblage from the middle part of the linnaei Biozone at the Želkovice locality (Litohlavy Formation, Prague Synform). From a hand-dug test pit which exposed one square meter of bedding plane, 7180 graptolite specimens were determined from three successive bulk samples (Z0, Z1 and Z2), each representing a 10-cm thick layer. Throughout the samples, changes in the graptolite assemblage are evident, both in terms of species composition and in total and relative abundance. Although overall the studied material may represent a climax stage of the graptolite community before the utilis extinction event, a detailed analysis revealed short-term fluctuations in the assemblage composition and population structure. In total, 25 species of graptolites were found, of which only 15 are present in all samples and subsamples. The most diverse and abundant graptolite fauna was found in sample Z1, in which 23 species were found. Metaclimacograptus asejradi, Streptograptus pericoi, and Glyptograptus fastigatus are identified for the first time from both Želkovice and the Prague Synform in general. Additionally, the first appearance datum of Parapetalolithus hispanicus has been identified, allowing for a detailed analysis of the boundary interval between the palmeus and hispanicus subzones of the linnaei Biozone. Graptolites of the linnaei Biozone have already been described and studied at Želkovice since the mid-nineteenth century. However, this work is the first to deal with small-scale variations in the percentage representation of taxa linked to observations on changing taphonomy and a comprehensive quantitative analysis of the graptolite community.
Zuzana Strossová. Institute of Geology and Palaeontology, Faculty of Science, Charles University, Albertov 6, Prague, 128 43, Czech Republic and Czech Geological Survey, Klárov 3, 118 21 Prague 1, Czech Republic strossova@natur.cuni.cz
Vojtěch Kovář. Czech Geological Survey, Klárov 3, 118 21 Prague 1, Czech Republic; vojtech.kovar@geology.cz
Petr Štorch. Department of Palaeobiology and Palaeoecology, Institute of Geology, Academy of Sciences of the Czech Republic, Rozvojová 269, 165 00 Praha 6, Czech Republic; storch@gli.cas.cz
Keywords: upper Llandovery; graptolites; Barrandian area; richness; species diversity; community structure
Final citation: Strossová, Zuzana, Kovář, Vojtěch, and Štorch, Petr. 2024. Qualitative and quantitative analysis of the graptolite assemblage in the linnaei Biozone (Silurian, lowermost Telychian) at Želkovice, Prague Synform (Czech Republic). Palaeontologia Electronica, 27(3:a58.
https://doi.org/10.26879/1425
palaeo-electronica.org/content/2024/5392-zelkovice-graptolite-analysis
Copyright: December 2024 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
The planktonic graptolites, extinct representatives of Pterobranchia, belong to the most utilized groups for biostratigraphy in the Paleozoic, allowing for highly precise correlations of strata both regionally and globally. The duration of individual biozones in the Ordovician and Silurian can vary from a little over 1.5 Myr to only hundreds of thousands of years (Maletz, 2017; Goldmann et al., 2020; Štorch, 2023; Štorch et al., 2024). Due to their planktonic mode of life, they can also be excellent indicators of Palaeozoic sea currents (Maletz, 2017). Finally, graptolite evolution provides numerous textbook examples of the effects of morphogenetic plasticity and adaptive radiation, thanks to which they survived several extinction events and, as “rebirths”, occupied empty niches with new species (Štorch, 1995; 2023). Nevertheless, after each event, the evolutionary plasticity was gradually exhausted and the number of newly evolved species decreased (Štorch, 1995, 2023), until their ultimate extinction in the late Early Devonian (Pragian/Emsian Boundary; Maletz, 2017). The response of graptolite communities to extinction events has been studied by numerous authors (e.g., Koren’, 1971; Lenz, 1993; Štorch, 1995; Manda et al., 2019). However, there has been less focus on population structure during climax periods, the population dynamics of which are consequently less well understood.
A small-scale, but detailed case study provides information on diversity and abundance as well as methodological insights that can support potential large-scale studies. Similar questions and topics were dealt with in the Ordovician, for example, by quantitative studies of Cisne and Chandlee (1982), Underwood (1998), Fan et al. (2013) and in the Silurian by Mazur (2014) or most recently by Manda et al. (2019). However, except for Manda et al. (2019), the listed studies are based primarily on specimens isolated from limestones and thus involve a substantially different methodology, in which dissolution of the samples and analysis of isolated specimens play an important role. Underwood (1998) outlined the methodological aspects of work with dissolved material with a high degree of precision and aptness. The advantage of the acid-extracted material is the acquisition of tubaria (according to Maletz et al., 2014) preserved in 3D, on which the development of the colony can be studied and important morphological features, poorly visible or not preserved in compression fossils, can be observed. A disadvantage is the fragility of tubaria, which can lead to the loss or disintegration of colonies into multiple fragments, making the accuracy of quantitative analysis questionable.
The Silurian sedimentary succession of the Prague Synform in the Barrandian area of central Bohemia, Czech Republic, has been continuously studied for more than 150 years; however, the first modern biostratigraphic division and the effort focused on regional correlation using graptolites was carried out by Bedřich Bouček in the 1930s (Bouček, 1932). The first documented finds of graptolites from the Želkovice locality date back to the time of Joachim Barrande's collections in the mid-nineteenth century (Barrande, 1850). The locality yields uniquely well-preserved and abundant graptolites. At present, unfortunately, it is no longer well-exposed and accessible. For this reason, limited rescue research took place in the summer of 2021, during which a hand-dug test pit was excavated in the black shale facies (linnaei Biozone). It was possible to detect short-term changes in graptolite assemblage (qualitatively and quantitatively) in the order of 10 cm and 5 cm. During the counting and determination, previously unpublished taxa from the given locality were also found and newly described. Finally, the lowest occurrence of P. hispanicus was also precisely captured, according to which the linnaei Biozone is divided into two subzones: palmeus and hispanicus. This work is aimed to i) find out species diversity and its short-term changes across the interval studied (S), total graptolite abundance; ii) detect short-term fluctuations and changes in the graptolite assemblage; iii) reveal the dominant species, community structure; and iv) application of various indices of dominance and alpha diversity.
GEOLOGICAL SETTING
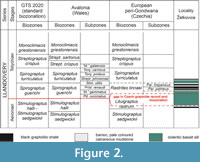
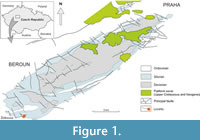 Prague Synform is located in the central part of the Teplá-Barrandian Unit of the Variscan basement of the Bohemian Massif. The inner part of the synform consists of Silurian and Devonian deposits, outer parts are formed by sedimentary and volcanic rocks of Ordovician age (see Figure 1). Its structural-tectonic development was described and summarized in detail by Havlíček (1981), Havlíček et al. (1994), Chlupáč et al. (1998), Melichar (2004), Röhlich (2007), and Vacek and Žák (2019). The Silurian volcano-sedimentary succession of the Prague Synform is formally divided into five successive formations: Želkovice, Litohlavy, Motol, Kopanina, and Požáry (Kříž, 1992); the units exhibit a progressive shift from shale-dominated to carbonate-dominated sedimentation. The characteristics of the individual formations and their relations to the graptolite biozones were summarized by Štorch (2023). At the studied locality, the upper part of the Želkovice and the lower part of the Litohlavy formations have been recorded. These formations consist of black silty shales, siliceous shales, clayey shales, and black silicites. Litohlavy Formation is characterized by irregular alternation of black shales and pale-coloured mudstones, the latter of which are barren of graptolites. Currently, the sedgwickii, rastrum, linnaei, and turriculatus biozones are known from the southern outskirts of Želkovice village. The boundary between the Aeronian Lituigraptus rastrum Biozone and the early Telychian Rastrites linnaei Biozone is hidden within a pale-coloured mudstone lacking graptolites and an overlying basalt sill, separating the linnaei Biozone (see Figure 2) (Štorch and Frýda, 2012; Štorch, 2023).
Prague Synform is located in the central part of the Teplá-Barrandian Unit of the Variscan basement of the Bohemian Massif. The inner part of the synform consists of Silurian and Devonian deposits, outer parts are formed by sedimentary and volcanic rocks of Ordovician age (see Figure 1). Its structural-tectonic development was described and summarized in detail by Havlíček (1981), Havlíček et al. (1994), Chlupáč et al. (1998), Melichar (2004), Röhlich (2007), and Vacek and Žák (2019). The Silurian volcano-sedimentary succession of the Prague Synform is formally divided into five successive formations: Želkovice, Litohlavy, Motol, Kopanina, and Požáry (Kříž, 1992); the units exhibit a progressive shift from shale-dominated to carbonate-dominated sedimentation. The characteristics of the individual formations and their relations to the graptolite biozones were summarized by Štorch (2023). At the studied locality, the upper part of the Želkovice and the lower part of the Litohlavy formations have been recorded. These formations consist of black silty shales, siliceous shales, clayey shales, and black silicites. Litohlavy Formation is characterized by irregular alternation of black shales and pale-coloured mudstones, the latter of which are barren of graptolites. Currently, the sedgwickii, rastrum, linnaei, and turriculatus biozones are known from the southern outskirts of Želkovice village. The boundary between the Aeronian Lituigraptus rastrum Biozone and the early Telychian Rastrites linnaei Biozone is hidden within a pale-coloured mudstone lacking graptolites and an overlying basalt sill, separating the linnaei Biozone (see Figure 2) (Štorch and Frýda, 2012; Štorch, 2023).
 The locality Želkovice (named after the village Želkovice near Libomyšl) is historically known for and represents an important site for graptolite research with unusually favourable preservation. Most species from the Rastrites linnaei Biozone of Želkovice have been described by Barrande (1850), Perner (1897) or Bouček and Přibyl (1941). Additional species from the locality were also documented by Bouček and Přibyl (1943, 1951). Bouček (1953) summarized initial findings about stratigraphic assignment and division of the locality and also mentioned the occurrence of the newly recognized sedgwickii Biozone (upper part of the biozone is currently referred to Lituigraptus rastrum Biozone (Štorch and Frýda, 2012; Štorch, 2023) and correlatable with Stimulograptus halli Biozone of Loydell (1991a, 1992). Bouček divided the linnaei Biozone into the lower Petalograptus palmeus and upper Petalograptus hispanicus subzones. Recently, Parapetalolithus species, including their juvenile stages from Želkovice, have been analyzed in detail by Strossová (2024).
The locality Želkovice (named after the village Želkovice near Libomyšl) is historically known for and represents an important site for graptolite research with unusually favourable preservation. Most species from the Rastrites linnaei Biozone of Želkovice have been described by Barrande (1850), Perner (1897) or Bouček and Přibyl (1941). Additional species from the locality were also documented by Bouček and Přibyl (1943, 1951). Bouček (1953) summarized initial findings about stratigraphic assignment and division of the locality and also mentioned the occurrence of the newly recognized sedgwickii Biozone (upper part of the biozone is currently referred to Lituigraptus rastrum Biozone (Štorch and Frýda, 2012; Štorch, 2023) and correlatable with Stimulograptus halli Biozone of Loydell (1991a, 1992). Bouček divided the linnaei Biozone into the lower Petalograptus palmeus and upper Petalograptus hispanicus subzones. Recently, Parapetalolithus species, including their juvenile stages from Želkovice, have been analyzed in detail by Strossová (2024).
MATERIAL AND METHODS
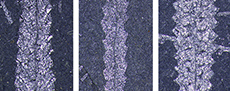
 A meter-by-meter-sized test pit was hand-excavated at a field tract exposing outcrops of the linnaei Biozone south of the farm at Želkovice (Figure 1). From the test pit, three consecutive bulk samples were collected. Each sample, represented by graptolite-bearing black shale, comprised a rock volume of 1 m x 1 m x 0.1 m (i.e., 105 cm3; see Figure 3). The lowest sample was divided into two subsamples of equal thickness. In total, 7180 determinable specimens of graptolites were counted through three bulk samples (Z0 - Z2). For exact numbers of species and specimens see Table 1 and Table 2.
A meter-by-meter-sized test pit was hand-excavated at a field tract exposing outcrops of the linnaei Biozone south of the farm at Želkovice (Figure 1). From the test pit, three consecutive bulk samples were collected. Each sample, represented by graptolite-bearing black shale, comprised a rock volume of 1 m x 1 m x 0.1 m (i.e., 105 cm3; see Figure 3). The lowest sample was divided into two subsamples of equal thickness. In total, 7180 determinable specimens of graptolites were counted through three bulk samples (Z0 - Z2). For exact numbers of species and specimens see Table 1 and Table 2.
Tubaria are usually preserved on black shale surface as flattened silvery impressions and preserved in a very low relief. Graptolites were photographed either dry or immersed in ethanol, using an Olympus SZX 16 stereomicroscope and an Olympus SZX 10 stereomicroscope fitted with a Canon EOS 2000D camera at the Institute of Geology of the Academy of Sciences. Figures with photos and line drawings were made using the Adobe Photoshop software and the CorelDraw software.
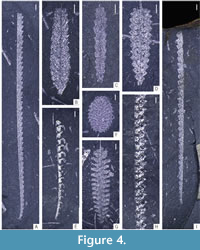
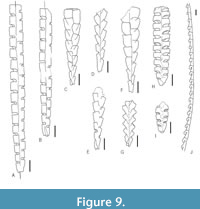
The obtained data was analysed using the PAST 4.11 software (Hammer et al., 2001). Statistical analyses focused on community structure (dominance/evenness), abundance (absolute) and species richness (across all samples). Rarefaction curves were computed for all three samples (and two subsamples) to evaluate the effect of sample size on species counts. For the evaluation of species richness across samples with substantially different sizes, two commonly utilized indices - Margalef’s index ((S-1)/ln(n)) and Menhinick’s index (S/√n) were calculated in each bulk sample where S is the number of species and n is the sample size. To characterize the evenness of graptolite assemblages preserved in the three respective samples and two subsamples, Dominance and Berger-Parker indices were calculated. Dominance was calculated using ![]() where ni is the number of individuals of taxon i and n is the sample size (total number of specimens in the sample). The Berger-Parker index, being more straightforward, is the number of individuals of the most common taxon divided by sample size. Given a large proportion of specimens belonging to genera Glyptograptus, Metaclimacograptus, and Streptograptus could not be determined to species level with certainty, the dominance index (D) has been calculated on genus level for each sample. The graptolite material is deposited in the collection of Zuzana Strossová (prefix ZAZ) in the Czech Geological Survey, Prague. Nearly all recognized taxa are depicted in Figure 4-Figure 5, examples of juvenile stages in Figure 6 and ranges of all species are shown in Figure 7. Newly documented taxa of graptolites from the Želkovice locality are depicted in separate Figure 8-Figure 9.
where ni is the number of individuals of taxon i and n is the sample size (total number of specimens in the sample). The Berger-Parker index, being more straightforward, is the number of individuals of the most common taxon divided by sample size. Given a large proportion of specimens belonging to genera Glyptograptus, Metaclimacograptus, and Streptograptus could not be determined to species level with certainty, the dominance index (D) has been calculated on genus level for each sample. The graptolite material is deposited in the collection of Zuzana Strossová (prefix ZAZ) in the Czech Geological Survey, Prague. Nearly all recognized taxa are depicted in Figure 4-Figure 5, examples of juvenile stages in Figure 6 and ranges of all species are shown in Figure 7. Newly documented taxa of graptolites from the Želkovice locality are depicted in separate Figure 8-Figure 9.
RESULTS
Sample Z0
 Sample Z0, 10 cm in thickness, represents the uppermost layer. Its topmost part is influenced by surface weathering. Secondary limonite formed through oxidation of small pyrite crystals, concretions or less common coatings. The shales are iron-red and cavities after Fe-rich concretions are occasionally found. In less weathered shales, graptolite tubaria are still pyritized, and pyrite was observed in the form of coatings.
Sample Z0, 10 cm in thickness, represents the uppermost layer. Its topmost part is influenced by surface weathering. Secondary limonite formed through oxidation of small pyrite crystals, concretions or less common coatings. The shales are iron-red and cavities after Fe-rich concretions are occasionally found. In less weathered shales, graptolite tubaria are still pyritized, and pyrite was observed in the form of coatings.
The graptolite assemblage consists mostly of mature specimens; siculae and juvenile stages are in the minority. Tubaria occur in clusters; isolated specimens are the exception. Rare conodonts have been observed. The graptolite assemblage comprises 23 species in 12 genera (see Table 1). The most represented species are Spirograptus guerichi (315 specimens), Parapetalolithus hispanicus, and Torquigraptus planus (150 specimens each). Conversely, the least represented were Pseudoretiolites dentatus (1 specimen), Metaclimacograptus asejradi, Parapetalolithus ovatus, and Rastrites fugax (2 specimens each). Representatives of genera Glyptograptus, Metaclimacograptus and Rastrites are not common; e.g., Rastrites schaueri present in lower samples disappears completely. On the contrary, some representatives of the genus Streptograptus (Str. pericoi and Str. plumosus) appear, which do not occur in older samples. In terms of morphological disparity, uniserial tubaria prevail in the assemblage. 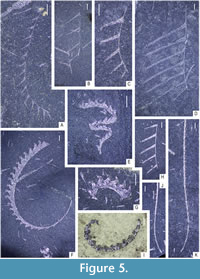 Their ratio is approximately 3:1 (i.e., 773 uniserial tubaria to 242 biserial tubaria). Although specimens with a straight type of tubarium predominate by genus and species (5 genera, 11 species - 304 specimens), spiralliform or coiled specimens of Spirograptus guerichi, Torquigraptus planus, and T. contortus dominate the assemblage (two genera, three species - 469 specimens). The abundance of the respective species is shown in Table 2 and in Figure 7. The identification of species and counting of specimens was hampered by surface weathering in the uppermost 2-3 cm of the sampled layer. However, it is unlikely to have affected the species diversity, population structure, or the total number of preserved specimens substantially. Compared to the other samples and subsamples (Z1, Z2, Z2 - A, Z2 - B), sample Z0 is characterized by a marked changed in specimen counts for all taxa. There is an exchange of species. Genera Glyptograptus and Rastrites retreat or disappear and, on the contrary, the first representatives of Streptograptus appear.
Their ratio is approximately 3:1 (i.e., 773 uniserial tubaria to 242 biserial tubaria). Although specimens with a straight type of tubarium predominate by genus and species (5 genera, 11 species - 304 specimens), spiralliform or coiled specimens of Spirograptus guerichi, Torquigraptus planus, and T. contortus dominate the assemblage (two genera, three species - 469 specimens). The abundance of the respective species is shown in Table 2 and in Figure 7. The identification of species and counting of specimens was hampered by surface weathering in the uppermost 2-3 cm of the sampled layer. However, it is unlikely to have affected the species diversity, population structure, or the total number of preserved specimens substantially. Compared to the other samples and subsamples (Z1, Z2, Z2 - A, Z2 - B), sample Z0 is characterized by a marked changed in specimen counts for all taxa. There is an exchange of species. Genera Glyptograptus and Rastrites retreat or disappear and, on the contrary, the first representatives of Streptograptus appear.
Sample Z1
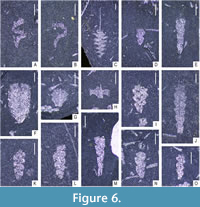 The 10 cm thick Z1 sample directly underlies sample Z0. When observing the excavated area (1 m2), approximately 70% of its upper bedding plane was filled with clustered graptolites. The remaining 30% of the surface was largely devoid of graptolites. Locally, tubaria are aligned, but the direction is not uniform for the entire layer surface. Lighter and darker laminae were visible in the cross-section. The shale was darker and more ferruginous than in the Z0 layer, with occasional cavities being left after pyrite concretions. Limonite occurred in the form of undispersed pseudomorphs. Pyrite was present in the form of coatings. Tubaria are preserved in accumulations as well as isolated. Rarely, conodont elements have been preserved. The graptolite assemblage is dominated by mature tubaria; however, in comparison with Z0 and Z2, there is also a high proportion of siculae and juvenile stages in Glyptograptus, Metaclimacograptus, Parapetalolithus, Spirograptus and others - see Figure 6; siculae and juveniles represent approximately one-third of the assemblage, which is substantially more than in the other samples. Graptolite assemblage is represented by 23 species and 12 genera. This bulk sample contained the largest number of specimens - a total of 3565. The most abundant species was Stimulograptus becki with 1009 specimens, which reached its peak dominance here. The species was also common in the other samples but with far fewer individual specimens (i.e., 29 specimens in Z0 and 241 specimens in Z2). No other species has reached such a high number of specimens even in the other layers. Other species abundant in sample Z1 include Spirograptus guerichi (688 specimens) and Torquigraptus planus (349 specimens). Conversely, the least abundant species include Rastrites cf. carnicus (4 specimens), Glyptograptus aff. latus (8 specimens), and Torquigraptus contortus (13 specimens). The number of species is comparable to layer Z0, because it contains representatives of the genera Rastrites and Glyptograptus, which do not occur in Z0, and on the contrary, Streptograptus pericoi and Streptograptus plumosus have not been recorded in Z1. In the assemblage, graptolites with uniserial tubaria (2824 specimens) predominate over biserial species (741 specimens). Their ratio is approximately 4:1. Compared to layer Z0, from the uniserial species, forms with a straight tubarium (1708 specimens) are more common than forms with a spiral tubarium (1050 specimens) represented by Spirograptus guerichi, Torquigraptus planus, and Torquigraptus contortus (1050 specimens). The exact species representation of individual taxa can be seen in Table 2 and in Figure 7. The sample represents a well-balanced climax assemblage of the linnaei Biozone. The high number of species, the high number of specimens across many species and the fact that in the assemblage the siculae, juvenile, and mature tubaria are evenly spread across many genera further support this assumption. Small-scale fluctuation, perhaps seasonality, is also evident from the laminae observed in the cross-section of the shale. However, these laminae are also visible in the lower part of the underlying Z2, where there is also a high abundance of species. During the fieldwork, an alignment of the tubaria, probably driven by a current, was noticeable. We assume that the Z1 layer could have been deposited under dysoxic conditions.
The 10 cm thick Z1 sample directly underlies sample Z0. When observing the excavated area (1 m2), approximately 70% of its upper bedding plane was filled with clustered graptolites. The remaining 30% of the surface was largely devoid of graptolites. Locally, tubaria are aligned, but the direction is not uniform for the entire layer surface. Lighter and darker laminae were visible in the cross-section. The shale was darker and more ferruginous than in the Z0 layer, with occasional cavities being left after pyrite concretions. Limonite occurred in the form of undispersed pseudomorphs. Pyrite was present in the form of coatings. Tubaria are preserved in accumulations as well as isolated. Rarely, conodont elements have been preserved. The graptolite assemblage is dominated by mature tubaria; however, in comparison with Z0 and Z2, there is also a high proportion of siculae and juvenile stages in Glyptograptus, Metaclimacograptus, Parapetalolithus, Spirograptus and others - see Figure 6; siculae and juveniles represent approximately one-third of the assemblage, which is substantially more than in the other samples. Graptolite assemblage is represented by 23 species and 12 genera. This bulk sample contained the largest number of specimens - a total of 3565. The most abundant species was Stimulograptus becki with 1009 specimens, which reached its peak dominance here. The species was also common in the other samples but with far fewer individual specimens (i.e., 29 specimens in Z0 and 241 specimens in Z2). No other species has reached such a high number of specimens even in the other layers. Other species abundant in sample Z1 include Spirograptus guerichi (688 specimens) and Torquigraptus planus (349 specimens). Conversely, the least abundant species include Rastrites cf. carnicus (4 specimens), Glyptograptus aff. latus (8 specimens), and Torquigraptus contortus (13 specimens). The number of species is comparable to layer Z0, because it contains representatives of the genera Rastrites and Glyptograptus, which do not occur in Z0, and on the contrary, Streptograptus pericoi and Streptograptus plumosus have not been recorded in Z1. In the assemblage, graptolites with uniserial tubaria (2824 specimens) predominate over biserial species (741 specimens). Their ratio is approximately 4:1. Compared to layer Z0, from the uniserial species, forms with a straight tubarium (1708 specimens) are more common than forms with a spiral tubarium (1050 specimens) represented by Spirograptus guerichi, Torquigraptus planus, and Torquigraptus contortus (1050 specimens). The exact species representation of individual taxa can be seen in Table 2 and in Figure 7. The sample represents a well-balanced climax assemblage of the linnaei Biozone. The high number of species, the high number of specimens across many species and the fact that in the assemblage the siculae, juvenile, and mature tubaria are evenly spread across many genera further support this assumption. Small-scale fluctuation, perhaps seasonality, is also evident from the laminae observed in the cross-section of the shale. However, these laminae are also visible in the lower part of the underlying Z2, where there is also a high abundance of species. During the fieldwork, an alignment of the tubaria, probably driven by a current, was noticeable. We assume that the Z1 layer could have been deposited under dysoxic conditions.
Sample Z2
Within the lowermost 10 cm thick layer changes in the graptolite assemblage were apparent already in the field. Therefore, the layer marked as Z2 was divided into two subsamples - A (upper 5 cm) and B (lower 5 cm). In the sample Z2, the graptolite assemblage is composed of 22 species and 12 genera. The species Spirograptus guerichi (684 specimens), Torquigraptus planus (428 specimens), and Parapetalolithus palmeus (331 specimens) are the most abundant in the sample. Among the least represented species are Monograptus marri (two specimens), Pseudoretiolites dentatus (three specimens), and Pristiograptus variabilis (five specimens). The number of species is comparable to samples Z0 and Z1; however, compared to Z0, Streptograptus is rare. There were more specimens counted in sample Z2 than in Z0, but less than in Z1. Uniserial tubaria (1843 specimens) predominate over biserial tubaria (757 specimens). Their ratio is approximately 2:1. Uniserial graptolites with a spiralliform tubarium are the most abundant. In Subsample A, 30-40% of the bedding plane surface was irregularly occupied by tubaria and fragments of Rastrites. Mature and juvenile specimens predominate here in general; however, in some taxa (e.g., genus Parapetalolithus) siculae are more abundant. Compared to layer Z1; however, the proportion of juvenile specimens in Z2 is notably lower across all genera and species. Tubaria were most often clustered in accumulations. Rare conodonts were also found. No current alignment was noted. Alternating lighter and darker laminae are visible in split shale. Limonite occurs in the form of rusty coatings, rarely as pseudomorphs after dispersed pyrite crystals. In this subsample, the graptolite assemblage yielded 22 species and 12 genera. The most common and least common species are identical to those listed in the overall characteristics of the sample Z2. However, the high abundance of some species found in the upper subsample (A) is pronounced. Spirograptus guerichi is represented by 591 specimens out of a total of 684 identified in the whole sample Z2 and Parapetalolithus palmeus comprises 277 specimens out of the total of 331. In the upper subsample, the occurrence of Parapetalolithus hispanicus, represented by 120 specimens, was recorded for the first time in the stratigraphic succession. Among uniserial species, the spirally enrolled tubaria predominate (a total of 898 specimens) over considerably less common straight forms (489 specimens).
 Subsample B represents the lowest studied stratigraphic level. In comparison with the overlying layers, graptolites are less common and generally less well-preserved. The assemblage consists almost exclusively of mature tubaria; there is an absolute minimum of juvenile growth stages, including siculae. Tubaria are scattered evenly on the surface; bedding-plane accumulations are rare. Rare conodonts have been observed. The shale is slightly ferruginous. Cavities filled with secondary Fe-minerals are rare, but limonite coatings are common. No evidence of current-driven alignment of tubaria was noted. The shale has a lighter shade than in subsample A, sometimes with a greenish tinge, which may be explained by a lower content of dispersed organic matter. Overall, the shale is more compact. In the subsample B, the graptolite assemblage contained 17 species assigned to nine genera. Compared to (sub)samples Z0, Z1, and Z2A, the subsample Z2B contained the lowest number of species, each represented by relatively low numbers of specimens. The most abundant species here include Torquigraptus planus (126 specimens), Spirograptus guerichi (93 specimens), and Stimulograptus halli (76 specimens). The least abundant species include Pristiograptus variabilis and Pseudoretiolites dentatus (both 1 specimen), Parapetalolithus ovatus (three specimens), and Glyptograptus fastigatus with Rastrites abbreviatus (four specimens each). Parapetalolithus palmeus was represented by 54 specimens while P. hispanicus is completely absent, the same as Glyptograptus aff. latus, Metaclimacograptus asejradi, Clinoclimacograptus aff. retroversus, Monograptus marri, or Rastrites cf. carnicus. The ratio of principal uniserial morphotypes is relatively even - 233 specimens with a straight tubarium and 223 specimens with a spiraliform or coiled tubarium. Compared to samples Z0 and Z1, the graptolite assemblage of subsamples Z2 (A and B) showed the highest fluctuation in the number of specimens. The lowest part of the excavated succession, which seemed substantially different from other layers lithologically and taphonomically already during the fieldwork, as it contains numerous fragments of Rastrites tubaria. This initial impression was subsequently confirmed by the detailed identification of the species and counting of their specimens. Probably the most interesting; however, is subsample B, i.e., the lowest part of the test pit where a sharp break in the number of species and the number of specimens was recorded. A lower number of species with relatively small populations may indicate less favourable environmental conditions, compared to layers Z0 or Z1. Due to the absence of crystallized pyrite and current-aligned graptolites, a distinct change in the sea-bottom ventilation and oxygenation of the water column and associated transition from dysoxic to anoxic conditions can be assumed.
Subsample B represents the lowest studied stratigraphic level. In comparison with the overlying layers, graptolites are less common and generally less well-preserved. The assemblage consists almost exclusively of mature tubaria; there is an absolute minimum of juvenile growth stages, including siculae. Tubaria are scattered evenly on the surface; bedding-plane accumulations are rare. Rare conodonts have been observed. The shale is slightly ferruginous. Cavities filled with secondary Fe-minerals are rare, but limonite coatings are common. No evidence of current-driven alignment of tubaria was noted. The shale has a lighter shade than in subsample A, sometimes with a greenish tinge, which may be explained by a lower content of dispersed organic matter. Overall, the shale is more compact. In the subsample B, the graptolite assemblage contained 17 species assigned to nine genera. Compared to (sub)samples Z0, Z1, and Z2A, the subsample Z2B contained the lowest number of species, each represented by relatively low numbers of specimens. The most abundant species here include Torquigraptus planus (126 specimens), Spirograptus guerichi (93 specimens), and Stimulograptus halli (76 specimens). The least abundant species include Pristiograptus variabilis and Pseudoretiolites dentatus (both 1 specimen), Parapetalolithus ovatus (three specimens), and Glyptograptus fastigatus with Rastrites abbreviatus (four specimens each). Parapetalolithus palmeus was represented by 54 specimens while P. hispanicus is completely absent, the same as Glyptograptus aff. latus, Metaclimacograptus asejradi, Clinoclimacograptus aff. retroversus, Monograptus marri, or Rastrites cf. carnicus. The ratio of principal uniserial morphotypes is relatively even - 233 specimens with a straight tubarium and 223 specimens with a spiraliform or coiled tubarium. Compared to samples Z0 and Z1, the graptolite assemblage of subsamples Z2 (A and B) showed the highest fluctuation in the number of specimens. The lowest part of the excavated succession, which seemed substantially different from other layers lithologically and taphonomically already during the fieldwork, as it contains numerous fragments of Rastrites tubaria. This initial impression was subsequently confirmed by the detailed identification of the species and counting of their specimens. Probably the most interesting; however, is subsample B, i.e., the lowest part of the test pit where a sharp break in the number of species and the number of specimens was recorded. A lower number of species with relatively small populations may indicate less favourable environmental conditions, compared to layers Z0 or Z1. Due to the absence of crystallized pyrite and current-aligned graptolites, a distinct change in the sea-bottom ventilation and oxygenation of the water column and associated transition from dysoxic to anoxic conditions can be assumed.
Species Richness and Dominance
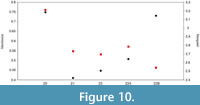
Species richness of the graptolite assemblage ranges from 22 to 23 species per sample. Margalef’s and Menhinick’s indices are 2.7-3.21 and 0.41-0.75, respectively (see Figure 10). When applying a diversity permutation test, there are no statistically significant differences (p < 0.05) in Margalef’s and Menhinick’s indices between the samples, except when comparing the Margalef’s index in subsamples Z2-A and Z2-B with sample Z0 and Menhinick’s index between Z2-A and Z0. Rarefaction curves for species richness only flatten out after approx. 500 specimens (see Figure 11).
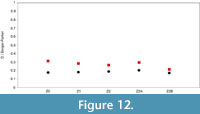
The dominance index is low and very similar for the individual samples and subsamples (D = 0.17-0.2; see Figure 12). The upper subsample of Z2 has a significantly increased dominance compared to the other (sub)samples (p<0.05 for each of the pairings); however, the difference is very small. Berger-Parker index is generally comparable among the samples, ranging from 0.26 to 0.31 (Figure 12). The subsample Z2B deviates slightly with a Berger-Parker index value of 0.21. Dominance is low in comparison with considerably higher values recorded in environmentally stressed graptolite assemblages affected by the late Aeronian sedgwickii mass extinction event (Štorch and Frýda, 2012) or upper Homerian assemblages that survived the devastating lundgreni mass extinction event (Manda et al., 2019).
SYSTEMATIC PALAEONTOLOGY
Phylum HEMICHORDATA Bateson, 1885, p. 111
Class PTEROBRANCHIA Lankester, 1877, p. 448
Subclass GRAPTOLITHINA Bronn, 1849, p. 149 (emended by Maletz, 2014b).
Family METACLIMACOGRAPTIDAE Koren’ and Rickards, 1996
Genus METACLIMACOGRAPTUS Bulman and Rickards, 1968, emend. Melchin et al., 2011
Type species. Diplograptus hughesi Nicholson, 1869; by original designation; from the Llandovery of the Lake District, England.
Metaclimacograptus asejradi Legrand, 1993
Figure 8B; Figure 9H, I
1989 Paraclimacograptus sp.; Rodríguez Núñez et al., pl. 2, fig. 5.
1993 Metaclimacograptus (?) asejradi nov. sp.; Legrand, p. 423, fig. 5e, f.
1998a Metaclimacograptus asejradi, Legrand 1993; Štorch, pl. I, fig. 1; text-fig.3, fig. 9.
1998 Metaclimacograptus cf. asejradi Legrand; Gutiérrez-Marco and Štorch, pp. 87-88, fig. 10e, f.
2003a Metaclimacograptus cf. M. asejradi Legrand, 1993; Lenz, Cuerda, and Peralta, p. 1200, figs. 3.7-3.9.
2003b Metaclimacograptus cf. asejradi Legrand, 1993; Lenz, Cuerda, and Peralta, figs. 3.1-3.3; 4.6-4.8
2012 Metaclimacograptus cf. asejradi Legrand, 1993; Tortello, Uriz, Alfaro, Cingolani, Bidone and Inchausti, p. 293-294, figs. 8.6, 8.7, 9.1.
2015 Metaclimacograptus asejradi Legrand; Loydell, Frýda, and Gutiérrez-Marco, p. 766, fig. 16AB.
2023 Metaclimacograptus asejradi Legrand, 1993; López and Kaufmann, fig. 3C-3D.
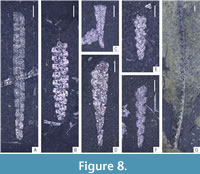 Holotype. specimen no. 1815 al, figured by Legrand (1993, fig. 5e); Tioukeline Claystones, north-west of Ouallene (Asejrad, Algeria); turriculatus s. l. Biozone (early Telychian)
Holotype. specimen no. 1815 al, figured by Legrand (1993, fig. 5e); Tioukeline Claystones, north-west of Ouallene (Asejrad, Algeria); turriculatus s. l. Biozone (early Telychian)
Material. 23 specimens, mostly complete, both mature and juvenile tubaria, preserved as flattened impressions, rarely in very low relief.
Diagnosis. The tubarium with an asymmetrical proximal end of normalograptid appearance, markedly widening from 0.7-0.9 mm at the first thecal pair to 1.2-1.4 mm at about th4. Median septum undulating. Thecae are densely spaced, sharply geniculated, with short (0.4-0.5 mm), parallel-sided supragenicular wall and deep and relatively broad apertural excavation perpendicular to the tubarium. Two thecae repeat distance of 1.3-1.4 mm proximally, and up to 1.7 mm distally.
Description. The tubarium is relatively robust. The length commonly varies between 7-8 mm. One exception was a specimen that was 13 mm long. The sicula is almost completely covered, with an aperture width of 0.35 mm. The proximal end is blunt, rounded and slightly asymmetrical. The virgella is 0.3-0.78 mm long, usually preserved. Dorso-ventral width (DVW) varies between 0.7 and 0.9 mm at the level of the 1st thecal pair and gradually increases through 1.2-1.3 mm at the third thecal pair to the maximum 1.3-1.4 mm attained at the fifth thecal pair. Thecae are sharply geniculated, and their double sigmoidal curvature is rarely visible. Short and slightly convex supragenicular ventral walls are 0.4-0.5 mm long at th5 and parallel to the tubarium axis. Each of the relatively narrow and deep apertural excavations without apparent hoods occupies approximately one-quarter to one-third of the tubarium width at th5. The 2TRD varies between 1.3 and 1.4 mm at th2 and th5. The 2TRD10 is about 1.7 mm. The median septum undulates, but details are not visible. The nema is occasionally preserved.
Remarks. In comparison to other species of Metaclimacograptus from the linnaei and lower turriculatus biozones, Metaclimacograptus asejradi is a well-recognizable species even in flattened specimens. It is characterized by its wide tubarium, short ventral supragenicular walls, regularly pronounced undulation of the median septum and slightly but distinctly asymmetric proximal end. Metaclimacograptus undulatus, described by Kurck (1882), is much smaller and narrower, with more closely spaced thecae and a rounded, symmetrical proximal end of the parallel-sided tubarium. The specimens studied here correspond to the description of Štorch (1998a) and Gutiérrez-Marco and Štorch (1998). However, the strongly undulating medium septum is poorly visible in most, flattened specimens.
Several authors recorded M. asejradi from the region of South America (see synonymy). López and Kaufmann (2023) recently described M. asejradi from Argentina from levels assigned to the Stimulograptus sedgwickii Biozone of the Aeronian age. Thus, its stratigraphic range may be longer than has been recorded so far.
Family NORMALOGRAPTIDAE Štorch and Serpagli, 1993, p. 14
Genus CLINOCLIMACOGRAPTUS Bulman and Rickards 1968, emend. Bulman and Rickards 1968 and Maletz, 2019
Type species. Clinoclimacograptus retroversus Bulman and Rickards 1968; by original designation; from the Llandovery of Tommarp, Sweden.
Clinoclimacograptus aff. retroversus (Bulman and Rickards, 1968)
Figure 8A; Figure 9A-B
aff. 1968 P. (Clinoclimacograptus) retroversus subgen. et. sp. nov.; Bulman and Rickards 1968, pp. 8-12, text- figs. 3a-c, 4a-c, 5.
aff. 1970 Pseudoclimacograptus (Clinoclimacograptus) retroversus Bulman and Rickards, 1968; Rickards, pp. 34-35, text-fig. 14, figs. 1-4.
aff. 1975 Pseudoclimacograptus retroversus (Bulman and Rickards, 1968); Bjerreskov, pp. 25-26, figs. 9D-E.
aff. 1991b Clinoclimacograptus retroversus Bulman and Rickards, 1968; Loydell, p. 674, pl. 1, figs. 5, 14.
 Holotype. Specimen SM A52951 from Tommarp, Sweden, probably sedgwickii Biozone, figured by Bulman and Rickards (1968, text-fig. 4b).
Holotype. Specimen SM A52951 from Tommarp, Sweden, probably sedgwickii Biozone, figured by Bulman and Rickards (1968, text-fig. 4b).
Material. 52 flattened specimens, including a number of juvenile tubaria.
Description. The tubarium is 8-10 mm long, exceptionally up to 14.65 mm. The sicula is largely covered, only in one specimen pressed through and showing length of ca. 1.2 mm and apertural width 0.26 mm. Virgella up to 0.44 mm long. Dorso-ventral width varies between 0.6 and 0.74 mm at the first thecal pair, increases to 0.8-0.9 mm at the third thecal pair, and 0.9-1.0 mm at the fifth thecal pair. A maximum width of 1.0-1.2 mm is attained at the tenth thecal pair. The proximal end is asymmetric but still somewhat rounded, length of th11 varies within 0.54-0.8 mm. Thecae are strongly geniculated with markedly long (0.65-0.77 mm), parallel-sided supragenicular walls. Deep, broadly slit-like excavations, generally perpendicular to the tubarium axis, occupy approximately one-third of the tubarium width at the level of th5. Strongly everted apertures claimed in pyritized specimens assigned to Clinoclimacograptus are not observed in flattened material. Also, the median septum is not visible in this mode of preservation. The 2TRD increases from 1.48-1.7 mm at th2, through 1.7-1.85 mm at th5 to 1.8-1.9 mm at th10. The nema is visible, but thread-like.
Remarks. None of the specimens studied can be assigned to C. retroversus with certainty. They differ from the genus Metaclimacograptus by their asymmetric and elongated proximal end and apertural excavations that are partially asymmetric (but not introverted) and slight incisions-like narrow apertural excavations, but deep. In addition, present specimens differ from Metaclimacograptus asejradi by having a significantly narrower flattened tubarium.
Most characters and parameters of the studied specimens correspond to those reported for Clinoclimacograptus retroversus by Bulman and Rickards (1968). However, the tubaria studied here show less densely spaced thecae (higher 2TRD values) than those reported by Bulman and Rickards (1968) and Rickards (1970). Present specimens possess 11-12 thecae in 10 mm, in the original description of C. retroversus this value ranges between 12-13 thecae in the proximal part of the tubarium and decreases to 10 thecae distally. Furthermore, the median septum is not visible in flattened imprints, making it impossible to say with certainty that this is truly a C. retroversus species. On the other hand, the other parameters correspond to the original descriptions of the holotype. Nevertheless, the work of Loydell (1991b), who worked with the extracted 3D tubarium should also be considered. The specimens assigned by them to the species Clinoclimacograptus retroversus show significantly higher dorso-ventral width than the specimens studied in this work. It cannot be overlooked that the width of these specimens preserved in full relief differs from the original descriptions of equally preserved specimens of C. retroversus by Bulman and Rickards (1968). Bjerreskov (1981) described Clinoclimacograptus washingtoni, from North Greenland which can be readily differentiated by its regularly widening tubarium and more concave supragenicular thecal walls. Finally, Bjerreskov (1975) mentions the occurrence of C. retroversus already from the convolutus Biozone, and the holotype of the species C. retroversus is described from the upper sedgwickii Biozone. Its occurrence in the linnaei Biozone in the Prague Synform would therefore continuously extend its stratigraphic range into the younger biozone.
Family RETIOLITIDAE Lapworth, 1873, emend. Melchin et al., 2011
Genus GLYPTOGRAPTUS Lapworth, 1873, emend. Melchin et al., 2011
Type species. Diplograptus tamariscus Nicholson, 1868; by original designation; from the Llandovery of Duffkinnel, Scotland.
Glyptograptus sp. A
Figure 8E; Figure 9G
Material. 15 specimens
Description. The tubarium is relatively narrow, usually only 4-5 mm long, with one exception reaching 11 mm. The proximal end is pointed. Most of the sicula is covered, the ventral wall is exposed by approximately 0.3 mm. The sicular aperture is approx. 0.2 mm in width. No virgella is preserved. The tubarium width increases slightly, but constantly from 0.7-0.8 mm at first thecal pair, through 1 mm at th3, to 1.1-1.2 mm at th10. Thecae inclined at 25-32° to the tubarium axis exhibit gently convex supragenicular walls. Concave thecal apertures are 0.27-0.3 mm wide, slightly everted and nearly perpendicular to the thecal axis. The length of th11 is 0.6 mm, the length of th12 varies between 0.7-0.8 mm. Thecal excavations are shallow in the proximal thecae. Thecae overlap for about one-half of their length. The 2TRD is 1.2-1.3 mm at th2 and about 1.3 mm at th5. The medium septum and nema were not recorded.
Remarks. All specimens of Glyptograptus sp. A were found in samples Z1 and Z2. Tubaria always were relatively narrow and short. Only one specimen attained the length of 11 mm. From the overall morphology and measured parameters, it appears that all specimens belong to the rich Glyptograptus tamariscus group. According to detailed descriptions and illustrations provided by Loydell (1992), the studied specimens resemble Glyptograptus elegans by Packham (1962) in tubarium width and 2TRD values. However, due to the large proportion of immature specimens in the present material, definite species assignment is not possible, and the species is left in open nomenclature. Commonly used 2TRD values have a limited value in juvenile specimens (e.g., in a specimen having 5-6 thecal pairs - the last pair does not reach full maturity and, therefore, does not allow for 2TRD5 measurement.
Glyptograptus fastigatus (Haberfelner, 1931)
Figure 8C-D; Figure 9C, F
1931 Glyptograptus tamariscus mut. fastigatus nov. mut.; Haberfelner, pp. 105-106, pl. 3, Figs. 17a-e.
1962 Glyptograptus sp. cf. G. tamariscus fastigans Haberfelner; Packham, pp. 512-513, pl. 71, fig. 5, text-fig. 3f.
1970 Glyptograptus aff. G. tamariscus fastigans Haberfelner, 1931; Hutt, Rickards, and Skevington, p. 6, pl. 1, fig. 14.
1979 Diplograptus (Orthograptus) ultimus Manck; Jaeger and Robardet, pl. 2, figs. 1, 20.
1989 Glyptograptus aff. G. fastigans Haberfelner; Melchin, p. 1740, fig. 11D (non B).
1991a G. fastigatus Haberfelner 1931; Loydell, p. 242, fig. 8c.
1992 Glyptograptus fastigatus Haberfelner, 1931a; Loydell, p. 30-31, pl. 1, figs. 4-5, text-fig. 11, figs. 2, 4, 22.
2015 Glyptograptus fastigatus Haberfelner; Loydell, Frýda, and Gutiérrez-Marco, figs. 16V, AA, 18B-D, 20K.
Holotype. Not assigned yet. Type collection of Gl. fastigatus described by Haberfelner (1931, pp. 105-106, pl. 3, Figs 17a-e) came from the turriculatus Biozone sensu lato of Hochwipfel in the Carnic Alps, Austria.
Material. 34 flattened, mostly complete tubaria
Description. The present, mostly immature tubaria have a pronounced wedge shape, usually 6.7-7.5 mm long, but one specimen attained a length of 28 mm. There is a sicula visible only in reverse view without an apex, and its length is 0.8-0.9 mm. The apex reaches the level of the middle of the second thecae, and the aperture width varies between 0.22-0.25 mm. A virgella has not been observed. The tubarium is only 0.5 mm wide at the first thecal pair, but then it widens significantly to 0.8-0.9 mm at th3, and 1.2-1.4 mm at th5. In some specimens having more than 10 thecal pairs, the width at th10 varies between 1.9 and 2.0 mm. The length of th11 varies between 0.6-0.7 mm, and th12 crosses the ventral wall of the sicula approximately 0.5 mm above the sicular aperture. Subsequent thecae are relatively long, regularly alternating in the proximal part. Towards the distal part of the tubarium, thecae become wider and form only a gentle sigmoidal curve, with very shallow apertural excavations. Thecal overlap varies between one-third and one-quarter of the thecal length. Thecal apertures, 0.15-0.21 mm wide, are perpendicular to the axis of the tubarium. Thecae are loosely spaced relative to the tubarium width, with 2TRD 2 1.7-2.13 mm, and 2TRD 5 1.9-2.1 mm. The nema fine, observed in four specimens, is little projected distally beyond the thecate part. The tubarium of a presumably circular or nearly circular cross section is commonly flattened in scalariform or sub-scalariform orientation.
Remarks. Almost all measured parameters correspond with descriptions of Gl. fastigatus provided by Packham (1962) and Loydell (1992). The only exception is the tubarium width, where the measured values in the distal part of the tubarium correspond to Glyptograptus auritus (Bjereskov, 1975). The 2TRD is generally lower than that reported for Glyptograptus fastigatus (2TRD2 1.55-2.15 mm; 2TRD5 2.00-2.25 mm) but, on the contrary, it is higher than 2TRD reported for Glyptograptus auritus (1.65 mm for 2TRD 2 and 1.9 mm for 2TRD 5) by Loydell (1992). Glyptograptus fastigatus was first described by Haberfelner (1931), who documented its occurrence from the maximus Subzone, which corresponds (according to Loydell 1992) in about the lower-middle guerichi Biozone. Haberfelner also mentioned the possibility of very limited stratigraphic occurrence within one biozone. His assumption was confirmed by Packham (1962), Melchin (1989), Loydell (1992), and Loydell et al. (2015), all of whom found this species in similar stratigraphic levels. Its present record in Bohemia (Prague Synform) in the linnaei Biozone is in accordance with previous findings. When comparing the descriptions of Glyptograptus fastigatus (Haberfelner, 1931) and Glyptograptus auritus (Bjerreskov, 1975) in detail, a considerable similarity was found. It should be stressed, however, that a basal disc developed in mature specimens of G. auritus has not been recorded in our material. In addition, the species Glyptograptus auritus, defined by Bjerreskov (1975), was documented from the lower part of her turriculatus Biozone. However, from the detailed notes of the work and associated graptolite fauna, it follows that this lower part also corresponds to the guerichi Biozone. It would be appropriate to subject the two species to a thorough revision in the future regarding the possible role of intraspecific variability. For example, in the genus Parapetalolithus (defined by Koren’ and Rickards, 1996), P. palmeus, and P. elongatus were distinguished as separate subspecies (and later species) for several decades (variants of P. p. palmeus, P. p. clavatus, P. e. elongatus, and P. e. linearis). The reason was the significantly expanded (even bulging) proximal part of the tubarium and thus increased DVW (dorsoventral-width) values. However, according to the works of Loydell (1992), Štorch (1998a) and the most recent systematic revision by Strossová (2024), it is still two species, not four.
Glyptograptus aff. latus (Packham, 1962)
Figure 8F; Figure 9D-E
aff. 1962 Glyptograptus enodis latus subsp. nov.; Packham, p. 518, pl. 71, fig. 20, text-fig. 4e.
aff. 2015 Glyptograptus latus Packham; Loydell, Frýda, and Gutiérrez-Marco, figs. 16H, 18R.
Holotype. Specimen no. SM A24969 from the lower Aeronian gregarius Biozone [horizon O after Sudbury (1958)], Rheidol Gorge, Wales figured by Packham (1962, pl. 71, fig. 20; text-fig. 4e).
Material. 16 flattened tubaria
Description. The relatively narrow, though markedly gradually widening, tubarium is usually 6-8 mm long. The longest measured specimens attain a length of 10-11 mm. The sicula is 1.9-1.0 mm long, 0.21-0.27 mm wide at the aperture. The virgella is very thin and 0.27-0.29 mm long. The tubarium width increases from 0.5-0.6 mm at th1, to 0.7-0.85 mm at th3, and 0.95-1.0 mm at th5. Specimens with more than 10 thecal pairs attain a maximum width of 1.1 mm. The length of th11 varies between 0.8 and 0.95 mm. Thecae markedly alternating, overlap for one-quarter to one-third of their length. Supragenicular walls are slightly inclined to the tubarium axis in the proximal thecae. Towards the distal part of the tubarium, they become gently convex and inclined at about 15-25° (values measured at th5). Thecal apertures are 0.2-0.25 mm wide and slightly everted. Apertural excavations are relatively prominent in the proximal part of the tubarium and occupy approximately one-third of the tubarium width. In distal thecae, genicula become less pronounced and excavations become shallower. The 2TRD ranges from 1.5-1.7 mm at th2 and 1.7-1.9 mm at th5. A slender nema is rarely projected beyond the thecate part of the tubarium.
Remarks. Packham (1962) described the new species in detail, based on a single specimen. Present specimens have almost identical parameter values with the original description (tubarium width, th1 length, thecal overlap, etc.). Nevertheless, the species G. latus is based on a specimen preserved in relief, found in the gregarius Biozone of Rheidol Gorge, UK, which corresponds to Demirastrites triangulatus - Demirastrites simulans biozones of the Prague Synform. Loydell et al. (2015) recorded G. latus in the halli Biozone (Upper Aeronian) in the El Pintado section in Spain. If all specimens referred to this species in Rheidol Gorge, El Pintado and here (in Želkovice) belong to the same species, then Glyptograptus latus has a rather large stratigraphic range - from the lower Aeronian to the lower Telychian. The overall morphology, and the fact that other species of the genus Glyptograptus also have long stratigraphic ranges, (e.g., Glyptograptus elegans) allow for the tentative assignment of the Czech specimens to G. latus.
Family MONOGRAPTIDAE Lapworth, 1873
Genus STREPTOGRAPTUS Yin, 1937, emend. Loydell, 1990 and Maletz and Loydell 2021
Type species. Original specimen Graptolitus plumosus Baily (1871, pp. 22-23, fig. 1a-c.) is lost. Currently, neotype BELUM K12275a is figured by Loydell (1990) and came from the turriculatus Biozone of Tieveshilly, County Down, Northern Ireland.
Streptograptus pericoi Štorch, 1998a
Figure 8G; Figure 9J
1998 “Paradiversograptus” aff. runcinatus (Lapworth); Gutiérrez-Marco and Štorch, p. 90, figs. 10 a, b, d.
1998a Streptograptus pericoi sp. n. Štorch; Štorch, p. 127, pl. II, fig. 6, text-fig. 3, fig. 4.
2015 Streptograptus pericoi Štorch; Loydell, Frýda, and Gutiérrez-Marco, p. 771, figs. 17G, p. 779, 21AB, AL, AM.
2019 Streptograptus pericoi Štorch; Loydell, Udchachon and Burrett, p. 372, fig. 8A.
Holotype. specimen no. MGM 3480 S, figured by Štorch (1998a, pl. II, fig. 6 and text-fig. 3, fig. 4), from the upper part of the linnaei Biozone (hispanicus Subzone, early Telychian) of the Bádenas Formation at Checa section (Western Iberian Cordillera, Guadalajara Province, Spain).
Material. 8 incomplete specimens and fragments.
Description. The tubarium is gently ventrally curved and gently widening. Immature specimens are 19.7-20 mm long, exceptionally over 40 mm in one distal fragment. The most proximal part of the tubarium can be, in some cases, weakly dorsally recurved (here in three specimens out of eight). The sicula is 1.08 mm long, with an apertural width 0.17-0.22 mm. The sicular apex reaches to half level of th1. Dorso-ventral width of the tubarium gradually increases from 0.25-0.4 mm at th1, to 0.29-0.37 mm at th2, 0.3 mm at th3, 0.36-0.46 mm at th5, 0.54 mm at th10, and 0,72 at th15. Mature specimens with more than 15 thecae attain a maximum distal width of 0.74-0.87 mm. The prothecae possess an inward-sloping ventral wall. Coiled metathecae of streptograptid form occupy approximately two-fifths of the DVW in proximal thecae and one-half of the DVW in distal thecae. Accordingly, a coiled metatheca takes up approx. one-half of the total thecal length in the distal part of the tubarium. Extended, inward-facing thecal apertures point towards the dorsal side of the tubarium. The 2TRD values are 1.69-1.75 mm at th2, 1.71-1.81 at th3, and 1.65-1.78 at th5. The presumed interthecal septum is steeply inclined.
Remarks. Streptograptus pericoi was described by Štorch (1998a) from the Checa Section in Spain. Currently, its occurrence is also recorded from another locality in Spain (El Pintado) and from the Sepon Mine (central Laos), along with the present record in the Prague Synform (Czechia). Loydell et al. (2019) considered Str. pericoi to be one of the typical (peri) Gondwanan graptolite species. The sicular apex reaches half the length of th1 in some specimens described in this study. However, in the original description by Štorch (1998a), a sicula reaches the top of th1. The 2TRD values are almost identical to those in the original description throughout the tubarium - the differences in 2TRD values fluctuate in the order of 0.1-0.2 mm which may be ascribed to intraspecific variability. The overall trend of 2TRD values and its slight decrease towards the distal part of the tubarium remained the same. Specimens are preserved as flattened silvery impressions in low relief, and, unfortunately, in most cases pyritized. The species has been recorded only in the uppermost, Z0 layer of the test pit, which corresponds to the hispanicus Subzone and observations by Štorch (1998a) or Loydell et al. (2015). At the same or similar level in which Str. pericoi is found, the following species have been recorded in the peri-Gondwanan sections: Paradiversograptus runcinatus (Lapworth, 1876), Streptograptus plumosus (Baily, 1871), Streptograptus pseudoruncinatus (Bjerreskov, 1975), Monograptus pseudobecki (Bouček and Přibyl, 1943), and Streptograptus picarrai (Loydell et al. 2015). When compared to Paradiversograptus runcinatus (Lapworth, 1876), the Str. pericoi differs in shape of thecae and lower values of 2TRD. Paradiversograptus runcinatus possesses simply hooked thecae and 2TRD values 1.8-2.2 mm at th2 as noted by Štorch (1998a). Streptograptus plumosus (Baily, 1871) has a tightly coiled tubarium with markedly dorsally recurved proximal end comprising 2-4 thecae, broader metathecae that occupy more than one-half of the tubarium width distally, and prothecae with conspicuous folds at their bases as noted by Loydell (1992). Streptograptus pseudoruncinatus (Bjerreskov, 1975) differs from Str. pericoi by having more closely spaced thecae as noted by Loydell (1993). Monograptus pseudobecki (Bouček and Přibyl, 1943) differs in having narrower, rather hooked than coiled thecae (like P. runcinatus) and slightly narrower proximal tubarium part (1.88 mm compared to 1.75 mm), with higher 2TRD2.
Streptograptus picarrai Loydell et al., 2015 from El Pintado section, has a similar tubarium curvature to other species listed above, but it has a greater DVW: 0.45-0.6 mm at th2, 0.55-0.8 mm at th5, and 0.65-0.9 mm at th10.
DISCUSSION
 When interpreting the obtained data (qualitative and quantitative) and reconstructing the palaeoenvironmental and palaeoecological conditions, various factors need to be considered. The lithology of the given locality, the ecology of graptolites, their morphology, the intensity, and influence of taphonomy and their mutual interactions require thorough consideration. Finally, methodological procedures, such as sampling, taxonomic work, and statistical analyses all influence the results of any palaeobiological or stratigraphic analyses.
When interpreting the obtained data (qualitative and quantitative) and reconstructing the palaeoenvironmental and palaeoecological conditions, various factors need to be considered. The lithology of the given locality, the ecology of graptolites, their morphology, the intensity, and influence of taphonomy and their mutual interactions require thorough consideration. Finally, methodological procedures, such as sampling, taxonomic work, and statistical analyses all influence the results of any palaeobiological or stratigraphic analyses.
Environmental conditions
 Except for abundant graptolites and occasional occurrences of conodont elements, no other fauna was recorded in the test pit. Nor have other authors (Bouček and Přibyl, 1941; Bouček, 1953; Štorch, 1994) reported any further fauna from the linnaei Biozone of this locality. The faunal record combined with occasional fine parallel lamination of the black shale, the conspicuous absence of trace fossils, widespread presence of pyrite coatings, crystals and small concretions are suggestive of a sedimentation in a low energy, likely deep-water environment under dysoxic to anoxic bottom conditions. In such a case, there was little risk that graptolite tubaria (or other faunal remains) would be disturbed by scavengers, which could have otherwise significantly affected the taphocenosis. However, it is also important to realize the scope and applicability of this study, resulting from a limited number of samples, which can be applied to a locality. Another crucial factor is the planktonic way of life of graptolites. It is important to realize that the colony of zooids moved diurnal vertical migration in the water column for most of its life, but was subject to lateral transport by sea currents, including possibly upwelling and downwelling (Maletz, 2017). This could have resulted in a considerable postmortem transfer before the colony sank to the sea bottom. On the contrary, an absence of currents would be required for the colony to be buried directly below the place of its demise (Maletz, 2014a, 2017).
Except for abundant graptolites and occasional occurrences of conodont elements, no other fauna was recorded in the test pit. Nor have other authors (Bouček and Přibyl, 1941; Bouček, 1953; Štorch, 1994) reported any further fauna from the linnaei Biozone of this locality. The faunal record combined with occasional fine parallel lamination of the black shale, the conspicuous absence of trace fossils, widespread presence of pyrite coatings, crystals and small concretions are suggestive of a sedimentation in a low energy, likely deep-water environment under dysoxic to anoxic bottom conditions. In such a case, there was little risk that graptolite tubaria (or other faunal remains) would be disturbed by scavengers, which could have otherwise significantly affected the taphocenosis. However, it is also important to realize the scope and applicability of this study, resulting from a limited number of samples, which can be applied to a locality. Another crucial factor is the planktonic way of life of graptolites. It is important to realize that the colony of zooids moved diurnal vertical migration in the water column for most of its life, but was subject to lateral transport by sea currents, including possibly upwelling and downwelling (Maletz, 2017). This could have resulted in a considerable postmortem transfer before the colony sank to the sea bottom. On the contrary, an absence of currents would be required for the colony to be buried directly below the place of its demise (Maletz, 2014a, 2017).
 Despite the problems mentioned above, several studies have managed to deal with the given problem and with the help of systematic facies data from the field and the application of the principle of actualism (e.g.: comparison with modern zooplankton - see Finney and Berry 1997) have proposed models of the depth distribution of graptolites. Most of these works are based on Ordovician graptolites, namely azygograptids or pendent didymograptids (see Cisne and Chandlee, 1982; Cooper et al., 1991; Beckly and Maletz, 1991; Goldman et al., 2002; Egenhoff and Maletz, 2007; Mitchell et al., 2008). Their models differentiated shallow-water and deep-water graptolite communities. Relatively low diversity communities are typical for outer-shelf settings, while deep-water environments are characterized by species-rich communities and the possible presence of pandemic taxa (Maletz, 2017). Lower faunal provincialism is documented in the Silurian; however, the principles of biogeographic distribution or depth distribution of graptolites apparently functioned similarly (Berry and Boucot, 1972; Kaljo, 1978; Štorch, 1998b; Cooper et al., 2012). In Želkovice, a highly diversified community was observed in the samples. If we apply the information from the above-mentioned Ordovician and Silurian works to our study, then the studied taphocenosis indicates a deep-water environment or a shelf-water environment during a high sea level in the Prague Basin.
Despite the problems mentioned above, several studies have managed to deal with the given problem and with the help of systematic facies data from the field and the application of the principle of actualism (e.g.: comparison with modern zooplankton - see Finney and Berry 1997) have proposed models of the depth distribution of graptolites. Most of these works are based on Ordovician graptolites, namely azygograptids or pendent didymograptids (see Cisne and Chandlee, 1982; Cooper et al., 1991; Beckly and Maletz, 1991; Goldman et al., 2002; Egenhoff and Maletz, 2007; Mitchell et al., 2008). Their models differentiated shallow-water and deep-water graptolite communities. Relatively low diversity communities are typical for outer-shelf settings, while deep-water environments are characterized by species-rich communities and the possible presence of pandemic taxa (Maletz, 2017). Lower faunal provincialism is documented in the Silurian; however, the principles of biogeographic distribution or depth distribution of graptolites apparently functioned similarly (Berry and Boucot, 1972; Kaljo, 1978; Štorch, 1998b; Cooper et al., 2012). In Želkovice, a highly diversified community was observed in the samples. If we apply the information from the above-mentioned Ordovician and Silurian works to our study, then the studied taphocenosis indicates a deep-water environment or a shelf-water environment during a high sea level in the Prague Basin.
Methodological implications
Counting and determining species and their specimens was influenced by their preservation. Although the locality is exceptional in terms of graptolite imprints, which show a high level of detail (e.g., a preserved inner membrane of P. dentatus - see Figure 4G, Figure 6C), the analysis is still constrained due to the flattened nature of the specimens and its differential impact on distinct graptolite morphotypes. While the species of the genera Petalolithus and Parapetalolithus have tubaria of a rather flat and box-like cross-section, on the other hand, the representatives of the genera Glyptograptus and Metaclimacograptus exhibit tubaria rather oval to circular in cross-section, as shown by Loydell (1991b), Melchin (1998), Koren’ and Rickards (2004), Loydell and Maletz (2009), or Melchin et al. (2011). This difference in tubarium cross-sections has a notable bearing on species-level taxonomical analyses. In many cases, tubaria of Petalolithus and Parapetalolithus sank with their longer axis parallel to the sea bottom and were deposited accordingly (e.g., Štorch, 1998a, 2001; Strossová, 2024); we can even recognize whether the tubarium is deposited reversely or obversely. When the tubaria of some Glyptograptus and Metaclimacograptus with oval or circular cross-sections reached the seafloor and were subsequently entombed in the sediments, their orientation was notably more random. This commonly resulted in a scalariform or sub-scalariform mode of diagenetic flattening that makes their species-level determination difficult or even impossible. It may also lead to the erroneous establishment of new species based on artefacts of preservation. Therefore, up to 80% of specimens in some genera were assignable to genus level only in our study, unavoidably impacting any study on diversity. A similar although less substantial problem also occurs in some fragmentary specimens of Streptograptus; however, in this study, their determination was mainly hampered by bedding surface weathering and disintegration of tubaria into fragments lacking either the proximal or distal part. The orientation of Streptograptus with upward-facing metathecae (i.e., scalariform view) is very common. Therefore, all specimens preserved on the bedding plane in unfavourable, scalariform orientation, which made their definite identification impossible, were also included; however, only determined to the genus level. Noteworthy is the impact of sample size on observed species richness. Rarefaction curves (Figure 11) clearly show that species richness only stabilizes after approx. 500 specimens in every sample. This should be noted in the case of studies in comparable assemblages as specimens of rare species can be severely outnumbered by the more common ones. Had this study been conducted on smaller samples, some of the species could have been missed. Only a handful of studies have so far analysed a short interval in such detail, none in the Silurian. This should bring new insights into graptolite sampling elsewhere. Statistical analyses based on the comparison of diversity and dominance indices showed either no or only extremely small differences between samples (compare e.g., with Manda et al., 2019). Similarly, the species richness indices are mostly comparable across the samples. This is interesting, as it shows that while the individual taxa changed quite substantially between the samples, the general structure of the assemblage remains relatively stable.
Graptolite biostratigraphy and assemblages
The studied taphocenosis characterized by low dominance and high species richness, likely reflects a thriving climax community characterizing the period between the sedgwickii and utilis events. Based on species richness, abundance, population structure, occurrence of specialized and surviving older taxa, and morphological disparity, it can be assumed that the best living conditions prevailed in the interval represented by samples Z0 and Z1. The relationships between the climax stages of graptolite communities and biotic crises were discussed in detail by Melchin et al. (1991), Urbanek (1993), and Štorch (1995).
According to the papers by Urbanek (1993), Loydell (1994), Štorch (1995), Štorch and Frýda (2012), or Štorch (2023), the species Parapetalolithus elongatus, Parapetalolithus hispanicus or Spirograptus guerichi can be identified and assumed as progenitor taxa or their descendants. On the other hand, representatives of the genus Streptograptus may be described as immigrant survivors. The presence of a well-developed, diverse community living likely under suitable conditions prevents us from establishing, which taxa have the potential to behave opportunistically under harsher conditions. Opportunists fill the void left by extinct taxa, where they dominate a low-species, high-abundance community. Subsequently, when new species radiate from progenitors, species immigrate or the so-called Lazarus taxa reappear from refugia, numbers of opportunists usually decrease again. There are several species only represented by a few specimens (e.g., Pristiograptus variabilis, Torquigraptus contortus, or Pseudoretiolites dentatus). The interpretation of why these taxa are less dominant is difficult.
From the oldest (lowermost) bulk sample Z2 to the youngest bulk sample Z0, abundance and species richness of biserial tubaria belonging to the genera Parapetalolithus, Glyptograptus, Metaclimacograptus, and Pseudoretiolites decreases gradually from the stratigraphically lowest sample to the highest one. Predomination of the uniserial graptolites, of genera Monograptus, Stimulograptus, and Spirograptus, exhibits an opposite trend. Among uniserial taxa, straight forms slightly outnumber spiral forms in diversity as well as abundance. In sample Z1, straight forms prevail substantially. However, in samples Z0 and Z2, where the abundance of specimens is lower than in Z1, spiral forms represented mainly by Spirograptus guerichi and Torquigraptus planus are more abundant. These taxa were likely able to better withstand the changes in environmental conditions. Similar phenomena have been observed in other fossil groups (see e.g., Vodička and Manda, 2019).
The onset of streptograptids accompanied by an overall high diversity of graptolites is typical for a pre-extinction fauna before the onset of the Utilis event (Loydell, 1994; Gutiérrez-Marco and Štorch, 1998). Therefore, could the assemblage of sample Z0 be a harbinger of the upcoming Utilis Event? Probably not, because species diversity does not decrease in the upper part of the linnaei Biozone, and sample Z0 is in its second half. It is therefore possible that this is only a small fluctuation. However, this possibility cannot be confirmed or denied with certainty. Unfortunately, the mentioned event is not recorded in the Prague Synform or in the Želkovice locality, due to the barren interval between the linnaei and turriculatus biozones (Štorch, 2023).
In conclusion, it would also be appropriate to mention why the Rastrites linnaei Biozone is used in the Prague Basin instead of the Spirograptus guerichi Biozone, which is part of the standard biozonation for the lower Telychian, as can be seen in Figure 2.
There are two reasons for the current use of the regional Rastrites linnaei Biozone instead of the international standard Spirograptus guerichi Biozone. First, juvenile specimens of S. guerichi can be easily misidentified as juvenile S. turriculatus, as demonstrated by Loydell et al. (1993). Secondly, the former species is common in the upper Parapetalolithus hispanicus Subzone of the linnaei Biozone, as exposed by a hand-dug test pit, but rare in its lower, Parapetalolithus palmeus Subzone. In contrast, Rastrites linnaei occurs throughout both subzones, though it is more abundant in the lower, palmeus Subzone. However, it is necessary to add that the palmeus Biozone, occupied only 5 cm of thickness in the hand dug test-pit, out of a total of 30 cm. The black shales of the Rastrites linnaei Biozone are bounded by barren mudstone layers both below and above. The base of the overlying Spirograptus turriculatus Biozone is not currently accessible in the Prague Synform, and the graptolite record at this level is poor. Rastrites linnaei, being more easily recognizable, was chosen as the index species for a separate biozone by Perner and Kodym (1919, 1922), Bouček (1953), and Štorch (1994, 2006, 2023). The occurrence of R. linnaei remains a more effective indicator in the Prague Synform for reliably differentiating between the uppermost part of the linnaei Biozone and the lowermost turriculatus Biozone.
Finally, the new information on occurrences of P. palmeus and P. hispanicus needs to be discussed. These taxa have been utilized for the determination of their eponymous biozones since they were first determined by Bouček (1953). In our study, an overlap of their ranges in at least 25 cm of the succession. FAD of P. hispanicus was captured in a layer with abundant P. palmeus. Considering the limited thickness of the overlap, this information may seem unimportant. Moreover, in layer Z0, where the occurrence of P. hispanicus reaches the highest point (150 specimens) in the studied interval, P. palmeus nearly disappears (four specimens). However, it still means that if in Želkovice or anywhere else in the same stratigraphic level we find a specimen of P. palmeus, we cannot be sure whether we are in the palmeus Subzone or already in the hispanicus Subzone. In addition, according to Bouček (1953) and Štorch (1994), both subzones can be up to several meters thick not only in Želkovice but also in other currently inaccessible localities of the Prague Synform. In the test pit, the overlap occurred at about 25 cm (five-sixths of the total thickness). In a section deposited more rapidly, the overlap could occur in a substantially larger thickness. Neither Bouček (1953) nor Štorch (1994, 2006, 2023) mentioned the overlap of subzonal index species, nor did other studies describing the Western Iberian Cordillera or Corral de Calatrava (Gutiérrez-Marco and Štorch, 1998; Štorch, 1998a), which partially adopted biozonations from Bohemia and Wales, provided this information. However, the delineation of FADs and LADs of index taxa is crucial for the precise definition of established biozones.
CONCLUSIONS
1) 7180 graptolite specimens belonging to 25 species were obtained from a hand-dug test pit in the linnaei Biozone. Several species have been recognized that have not been previously reported from the Prague Synform, namely: Streptograptus pericoi, Metaclimacograptus asejradi, Clinoclimacograptus aff. retroversus, Glyptograptus fastigatus, and Glyptograptus aff. latus.
2) The Z1 sample proved to be the richest in graptolites both in terms of species richness and abundance - 3565 specimens were found; 23 species were identified, and populations were most evenly represented here (siculae, juvenile stages, mature tubaria) across all genera. Due to the highest number of specimens, the highest diversity of species and the nature of the population, it can be assumed that the graptolite community reached its optimum at this level.
3) A gradual decrease in relative abundance and species richness of biserial graptolites is accompanied by an increase in both metrics in uniserial graptolites. Straight forms mostly outnumber spiral ones; however, their ratio is not constant. These shifts are likely ecologically conditioned.
4) Except for the index taxa Spirograptus guerichi, Pa. palmeus and Pa. hispanicus, the most abundant species was Stimulograptus becki. In layer Z1, the latter is even the most abundant species being represented by 1009 specimens. On the contrary, Sp. guerichi has “only” 688 specimens in the same layer. Among the least represented species are Rastrites cf. carnicus, Streptograptus pericoi, Glyptograptus aff. latus, and Streptograptus plumosus. Each was represented by less than 20 specimens in total.
5) Diversity and dominance indices show only small differences between the bulk samples representing three successive intervals, each 10 cm in thickness. Rarefaction curves show that a sample size below 500 specimens would result in rather inadequate species richness data. The relatively high diversity alongside low dominance shows a well-balanced thriving graptolite fauna hinting at favourable environmental conditions in the studied interval.
6) The present test pit, which exposed the boundary interval of the Parapetalolithus palmeus and Parapetalolithus hispanicus sub-biozones, enabled the validity of current subzones to be confirmed. However, there exists a partial overlap in occurrences of the index species that has not been previously reported.
ACKNOWLEDGEMENTS
The authors greatly acknowledge research funding through a study scholarship at Charles University, Czech Science Foundation project GA 20-23363S, and research plan RVO 67985831 of the Institute of Geology of the Czech Academy of Sciences. This manuscript also contributes to the Strategic Research Plan of the Czech Geological Survey (DKRVO/ČGS 2023-2027) through internal project 311410. Many thanks also go to L. Muir and one anonymous reviewer for their insightful comments and suggestions on the earlier version of the paper. ZS would like to express a big thanks to colleague M. Kočová Veselská for her unceasing support, plethora of valuable advice and for believing and listening to her.
REFERENCES
Baily, W.H. 1871. Palaeontological remarks. In Traill, W.A. and Egan, F.W. (eds.), Explanatory memoir to accompany Sheets 49, 50 and part of 61 of the maps of the Geological Survey of Ireland including the country around Downpatrick, and the shores of Dundrum Bay and Strangford Lough, County of Down. Alexander Thom. Dublin & London. p. 22-23.
Barrande, J. 1850. Graptolites de Bohême. Extrait du Systême Silurien du centre de la Bohême. Chez l’auteur, Prague. vi +74 pp., 4 pl. Imprimerie et papier de Théophile Haase Fils.
Bateson, W. 1885. The later stages in the development of Balanoglossus kowalevskii, with a suggestion as to the affinities of the Enteropneusta. Quarterly Journal of Microscopical Science 25:81-122.
Beckly, A. and Maletz, J. 1991. The Ordovician graptolites Azygograptus and Jishougraptus in Scandinavia and Britain. Palaeontology 34 (4):887-925.
Berry, W.B.N. and Boucot, A.J. 1972. Silurian graptolite depth zonation. In Proceedings of the 24th International Geological Congress, Montreal. Volume 7: pp. 59-65.
Bjerreskov, M. 1975. Llandoverian and Wenlockian graptolites from Bornholm. Fossils and Strata 8, 1-94.
https://doi.org/10.18261/8200093921-1975-01
Bjerreskov, M. 1981. Silurian graptolites from Washington Land, western North Greenland. Bulletin Grønlands Geologiske Undersøgelse, 142: 1-58.
https://doi.org/10.34194/bullggu.v142.6684
Bouček, B. 1932. Předbežna zpráva o některých nových druzích graptolitů z českého gotlandienu. Věstnik Státního geologického ústavu Československé republiky, 7:150-155.
Bouček, B. 1953. Biostratigraphy, development and correlation of the Želkovice and Motol Beds of the Silurian of Bohemia. Sborník Ústředního ústavu geologického, Oddíl paleontologický, 20:421-484.
Bouček, B. and Münch, A. 1944. Die Retioliten des mittel-europaischen Llandovery und unteren Wenlock. Rozpravy České Akademie věd a umění, Třída II 53(41):1-54.
Bouček, B. and Přibyl, A. 1941. O rodu Petalolithus Suess z českého siluru. Rozpravy České akademie věd a umění, Třída 2, 51(11):1-22.
Bouček, B. and Přibyl, A. 1943. Über böhmische Monograpten aus der Untergattung Streptograptus Yin. Mitteilungen der Tschechischen Akademie der Wissenschaften, 52:1-23.
Bouček, B. and Přibyl, A. 1951. On some slender species of the genus Monograptus Geinitz, especially of the subgenera Mediograptus and Globosograptus. Bulletin international de l’Académie tchèque des Sciences, 52:185-216.
Bronn, H.G. 1849. Handbuch der Geschichte der Natur. Dritter Band, Zweite Abtheilung. II. Theil: Organisches Leben (Schluß). Index palaeontologicus oder Ueberblick der bis jetzt bekannten fossilen Organismen. 1106 pp. Schweizerbart, Stuttgart.
https://doi.org/10.5962/bhl.title.102095
Bulman, O.M.B. and Rickards, R.B. 1968. Some new diplograptids from the Llandovery of Britain and Scandinavia. Palaeontology, 11(1):1-15.
Chlupáč, I., Havlíček, V., Kříž, J., Kukal, Z., and Štorch, P. 1998. Palaeozoic of the Barrandian (Cambrian to Devonian). 183 pp. Czech Geological Survey, Prague.
Cisne, J.L. and Chandlee, G.O. 1982. Taconic foreland basin graptolites: age zonation, depth zonation, and use in ecostratigraphic correlation. Lethaia, 15:343-363.
https://doi.org/10.1111/j.1502-3931.1982.tb01699
Cooper, R.A., Fortey, R.A., and Lindholm, K. 1991. Latitudinal and depth zonation of early Ordovician graptolites. Lethaia 24: 199-218.
https://doi.org/10.1111/j.1502-3931.1991.tb01468.x
Cooper, R. A., Rigby, S., Loydell, D.K., and Bates, D.E. 2012. Palaeoecology of the Graptoloidea. Earth-Science Reviews, 112(1-2): 23-41.
https://doi.org/10.1016/j.earscirev.2012.01.001
Egenhoff, S. and Maletz, J. 2007. Graptolites as indicators of maximum flooding surfaces in monotonous deepwater shelf successions. Palaios, 22:374-384.
https://doi.org/10.2110/palo.2005.p05-096r
Fan, J., Chen, Q., Melchin, M.J., Sheets, H.D., Chen, Z., Zhang, L., and Hou, X. 2013. Quantitative stratigraphy of the Wufeng and Lungmachi black shales and graptolite evolution during and after the Late Ordovician mass extinction. Palaeogeography, Palaeoclimatology, Palaeoecology, 389: 96-114.
https://doi.org/10.1016/j.palaeo.2013.08.005
Finney, S.C. and Berry, W.B. 1997. New perspectives on graptolite distributions and their use as indicators of platform margin dynamics. Geology, 25(10):919-922.
https://doi.org/10.1130/0091-7613(1997)025<0919:NPOGDA>2.3.CO;2
Goldman, D., Campbell, S.M., and Rahl, J.M. 2002. Three‐dimensionally preserved specimens of Amplexograptus (Ordovician, Graptolithina) from the North American mid-continent: taxonomic and biostratigraphic significance. Journal of Paleontology 76 (5):921-927.
https://doi.org/10.1666/0022-3360(2002)076<0921:TDPSOA>2.0.CO;2
Goldman, D., Sadler, P.M., Leslie, S.A., Melchin, M.J., Agterberg, F.P., and Gradstein, F.M. 2020. The Ordovician Period. In Geologic time scale 2020 (pp. 631-694). Elsevier.
https://doi.org/10.1016/b978-0-12-824360-2.00020-6
Gutiérrez-Marco, J.C. and Štorch, P. 1998. Graptolite biostratigraphy of the lower Silurian (Llandovery) shelf deposits of the Western Iberian Cordillera, Spain. Geological Magazine, 135(1):71-92.
https://doi.org/10.1017/S0016756897007802
Haberfelner, E. 1931. Graptolithen aus dem Obersilur der Karnischen Alpen. I. Hochwipfel Nordseite. Sitzungsberichte, Österreichische Akademie der Wissenschaften, Wien, Mathematisch-Naturwissenschaftliche Klasse, Abteilung, 140(1):89-168.
Hammer, Ø., Harper, D.A., and Ryan, P.D. 2001. Past: paleontological statistics software package for education and data analysis. Palaeontologia Electronica, 4(1):1.
https://palaeo-electronica.org/2001_1/past/issue1_01.htm
Havlíček, V. 1981. Development of a linear sedimentary depression exemplified by the Prague Basin (Ordovician-Middle Devonian; Barrandian area-central Bohemia). Sborník geologických věd, Geologie, 35:7-48.
Havlíček, V., Vaněk, J., and Fatka, O. 1994. Perunica microcontinent in the Ordovician (its position within the Mediterranean Province, series division, benthic and pelagic communities). Sborník geologických věd, Geologie, 46:23-56.
Hutt, J.E., Rickards, R.B., and Skevington, D. 1970. Isolated Silurian graptolites from the Bollerup and Klubbudden stages of Dalarna, Sweden. Geologica et Palaeontologica, 4(1).
Jaeger, H. and Robardet, M. 1979. Le Silurien et le Dévonien basal dans le Nord de la Province de Séville (Espagne). Geobios, 12(5): 687-714.
https://doi.org/10.1016/S0016-6995(79)80097-2
Kaljo, D.L. 1978. On the bathymetric distribution of graptolites. Acta Palaeontologica Polonica, 23(4):pp. 523-531. Warszawa.
Koren’, T.J.N. 1971. The zones of Monograptus hercynicus and Monograptus falcarius in Pai-Khoi. Lethaia, 4(2): 235-248.
https://doi.org/10.1111/j.1502-3931.1971.tb01291
Koren’, T.N. and Rickards, R.B. 1996. Taxonomy and evolution of Llandovery biserial graptoloids from the southern Urals, western Kazakhstan. Special Papers in Palaeontology, 54:1-103.
Koren’ T.N. and Rickards, R.B. 2004. An unusually diverse Llandovery (Silurian) diplograptid fauna from the southern Urals of Russia and its evolutionary significance. Palaeontology, 47(4):859-918.
https://doi.org/10.1111/j.0031-0239.2004.00411.x
Kříž, J. 1992. Silurian Field Excursions: Prague Basin (Barrandian), Bohemia. National Museum Wales, Geological Series 13:1-111.
Kurck, C. 1882. Nägra nya graptolitarter frä Skäne. Geologiska Föreningen i Stockholm Förhandlingar, 6(7):294-304.
Lankester, E.R. 1877. Notes on the embryology and classification of the animal kingdom; comprising a revision of speculations relative to the origin and significance of the germlayers. Quarterly Journal of Microscopical Science, New Series, 17:339-454.
Lapworth, C. 1873. On an improved classification of the Rhabdophora. Geological Magazine, 10:500-504, 555-560.
Lapworth, C. 1876. On Scottish Monograptidæ. Geological Magazine, 3(7): 308-321.
Legrand, P. 1993. A propos d’un Niveau à Neodiplograptus dans le Silurien inférieur à l’est de Ouallene, Asejrad (Sahara Algérien). Implications stratigraphiques et paléogéographiques. In 118 Congrès national des Sociétés historiques et scientifiques, 4 éme colloque sur la Géologie africaine, Pau, 409-424.
Lenz, A.C. 1993. Late Wenlock-Ludlow (Silurian) graptolite extinction, evolution, and biostratigraphy: perspectives from Arctic Canada. Canadian Journal of Earth Sciences, 30(3):491-498.
https://doi.org/10.1139/e93-038
Lenz, A., Cuerda, A., and Peralta, S. 2003a. Graptolites from the upper Llandovery of the Talacasto area, central Precordillera, San Juan, Argentina. Journal of Paleontology, 77(6): 1198-1202.
https://doi.org/10.1666/0022-3360(2003)077<1198:GFTULO>2.0.CO;2
Lenz, A.C., Cuerda, A.J., and Peralta, S.H. 2003b. A retiolitid graptolite fauna from the Talacasto area, central Precordillera of San Juan, Argentina, and its age significance. Journal of Paleontology, 77(6):1198-1202.
López, F.E. and Kaufmann, C. 2023. New insights on the Silurian graptolite biostratigraphy of the La Chilca Formation, Poblete Norte section, Central Precordillera of San Juan, Argentina: faunal replacement and paleoenvironmental implications. Andean geology, 50(2):201-216.
https://doi.org/10.5027/andgeov50n2-3617
Loydell, D.K. 1990. On the graptolites described by Baily (1871) from the Silurian of Northern Ireland and the genus Streptograptus Yin. Palaeontology, 33:937-943.
Loydell, D.K. 1991a. The biostratigraphy and formational relationships of the upper Aeronian and lower Telychian (Llandovery, Silurian) formations of western mid-Wales. Geological Journal, 26(3):209-244.
https://doi.org/10.1002/gj.3350260304
Loydell, D.K. 1991b. Isolated graptolites from the Llandovery of Kallholen, Sweden. Palaeontology, 34(3):671-693.
Loydell, D.K. 1992. Upper Aeronian and lower Telychian (Llandovery) graptolites from western mid-Wales. Monographs of the Palaeontographical Society, 147(592):55-164.
https://doi.org/10.1080/25761900.2022.12131772
Loydell, D.K. 1993. Worldwide correlation of Telychian (Upper Llandovery) strata using graptolites. Geological Society, London, Special Publications, 70(1):323-340.
https://doi.org/10.1144/gsl.sp.1993.070.01.21
Loydell, D.K. 1994. Early Telychian changes in graptoloid diversity and sea level. Geological Journal, 29(4):355-368.
Loydell, D.K., Štorch, P., and Melchin, M.J. 1993. Taxonomy, evolution, and biostratigraphical importance of the Llandovery graptolite Spirograptus. Palaeontology, 36:909-926.
Loydell, D.K. and Maletz, J. 2009. Isolated graptolites from the Lituigraptus convolutus Biozone (Silurian, Llandovery) of Dalarna, Sweden. Palaeontology, 52(2):273-296.
https://doi.org/10.1111/j.1475-4983.2008.00843.x
Loydell, D.K., Frýda, J., and Gutiérrez-Marco, J.C. 2015. The Aeronian/Telychian (Llandovery, Silurian) boundary, with particular reference to sections around the El Pintado reservoir, Seville Province, Spain. Bulletin of Geosciences, 90(4):743-794.
https://doi.org/10.3140/bull.geosci.1564
Loydell, D.K., Udchachon, M., and Burrett, C. 2019. Llandovery (lower Silurian) graptolites from the Sepon Mine, Truong Son Terrane, central Laos and their palaeogeographical significance. Journal of Asian Earth Sciences, 170:360-374.
https://doi.org/10.1016/j.jseaes.2018.11.013
Maletz, J. 2014a. Graptolite reconstructions and interpretations. Paläontologische Zeitschrift, 89:271-286.
https://doi.org/10.1007/s12542-014-0234-4
Maletz, J. 2014b. The classification of the Graptolithina Bronn, 1849. Bulletin of Geosciences 89, (3):477-540.
https://doi.org/10.3140/bull.geosci.1465
Maletz, J., Bates, E.B.D., Brussa, E.D., Cooper, A.R., Lenz, C.A., Riva, F.J., Toro, A.B., and Zhang, Y. 2014. Part V, Revision 2, Chapter 12: Glossary of the Hemichordata. Treatise Online 62:1-23.
Maletz, J. 2017. Graptolite Paleobiology. 336 pp. WileyBlackwell, UK.
https://doi.org/10.1002/9781118515624
Maletz, J. 2019. Part V, Second Revision, Chapter 24: Infraorder Neograptina and family Normalograptidae: Introduction, morphology, and systematic descriptions. Treatise Online 116:1-15.
https://doi.org/10.17161/to.v0i0.9768
Maletz, J. and Loydell, D.K. 2021. Part V, Second Revision, Chapter 28: Family Monograptidae: Introduction, morphology, and systematic descriptions. Treatise Online 156:1-43, 19 fig.
Manda, Š., Štorch, P., Frýda, J., Slavík, L., and Tasáryová, Z. 2019. The mid-Homerian (Silurian) biotic crisis in offshore settings of the Prague Synform, Czech Republic: Integration of the graptolite fossil record with conodonts, shelly fauna and carbon isotope data. Palaeogeography, Palaeoclimatology, Palaeoecology, 528:14-34.
https://doi.org/10.1016/j.palaeo.2019.04.026
Mazur, E. M. 2014. Population Variation in Fossil Graptolites: a Quantitative Study Based on Single Species Assemblages. Honors Theses, papers 36, 1-121pp.
Melchin, M.J. 1989. Llandovery graptolite biostratigraphy and paleobiogeography, Cape Phillips Formation, Canadian Arctic Islands. Canadian Journal of Earth Sciences, 26(9): 1726-1746.
https://doi.org/10.1139/e89-147
Melchin, M.J. 1998. Morphology and phylogeny of some early Silurian 'diplograptid' genera from Cornwallis Island, Arctic Canada. Palaeontology, 41(2):263-315.
Melchin, MJ., Mitchell, C. E., Barnes, C.R., and Williams, S.H. 1991. Late Ordovician extinction in the Graptoloidea. Advances in Ordovician Geology, 90(9):143-156.
Melchin, M.J., Mitchell, C.E., Naczk-Cameron, A., Fan, J.X., and Loxton, J. 2011. Phylogeny and Adaptive Radiation of the Neograptina (Graptoloida) During the Hirnantian Mass Extinction and Silurian Recovery. Proceedings of the Yorkshire Geological Society, volume 58(4):281-309.
https://doi.org/10.1144/pygs.58.4.301
Melichar, R. 2004. Tectonics of the Prague Synform: a hundred years of scientific discussion. Krystalinikum, 30:167-187.
Mitchell, C., Brussa, E.D., and Maletz, J. 2008. A mixed isograptid-didymograptid graptolite assemblage from the Middle Ordovician of west Gondwana (NW Bolivia): implications for graptolite paleoecology. Journal of Paleontology, 82(6):1114-1126.
https://doi.org/10.1666/06-069.1
Nicholson, H.A. 1868. On the graptolites of the Coniston Flags; with notes on the British species of the genus Graptolites. Quarterly Journal of the Geological Society, 24(1-2):521-545.
https://doi.org/10.1144/GSL.JGS.1868.024.01-02.67
Nicholson, H.A. 1869. On some new species of graptolites. Annals and Magazine of Natural History, 4(4):231-242.
https://doi.org/10.1080/00222936908696041
Packham, G.H. 1962. Some diplograptids from the British lower Silurian. Palaeontology, 5(3):498-526.
Perner, J. 1897. Études sur les Graptolites de Bohême. Part 3. section a. 25 pp. Raimond Gerhard, Prague.
https://doi.org/10.5962/bhl.title.100697
Perner, J. and Kodym, O. 1919. O rozčlenění svrchního siluru v Čechách. Časopis Musea Království českého 93:6-24.
Perner, J. and Kodym, O. 1922. On the zonal division and correlation of the Silurian of Bohemia. American Journal of Science 4: 53-72.
https://doi.org/10.2475/ajs.s5-4.19.53
Přibyl, A. 1940. O českých zástupcích monograptidů ze skupiny https://doi.org/10.5962/bhl.title.100697. Rozpravy České akademie věd a umění, Třída 2, 1(16):1-16.
Rickards, R.B. 1970. The Llandovery (Silurian) graptolites of the Howgill Fells, northern England. Monographs of the Palaeontographical Society, 123(524): 2-104.
Rodríguez-Núñez, V.M., Gutiérrez-Marco, J.C., and Sarmiento, G. 1989. Rasgos bioestratigráficos de la sucesión silúrica del Sinclinal del Guadarranque (provincias de Cáceres, Badajoz y Ciudad Real). COL-PA, 42:83-106.
Röhlich, P. 2007. Structure of the Prague Basin: The deformation diversity and its causes (the Czech Republic). Bulletin of Geosciences, 82(2):175-182.
https://doi.org/10.3140/bull.geosci.2007.02.175
Seelmeier, H. 1936. Obersilurische Graptolithen von der Gugel: (Karnische Alpen). Sitzungsberichte der Akademie der Wissenschaften, Wien, 145:217-226.
Štorch, P. 1994. Graptolite biostratigraphy of the Lower Silurian (Llandovery and Wenlock) of Bohemia. Geological Journal, 29:137-165.
https://doi.org//10.1002/gj.3350290204
Štorch, P. 1995. Biotic crises and post-crisis recoveries recorded by Silurian planktonic graptolite faunas of the Barrandian area (Czech Republic). Geolines, 3:59-70.
Štorch, P. 1998a. New data on Telychian (Upper Llandovery, Silurian) graptolites from Spain. Journal of the Czech Geological Society, 43(3):113-142.
Štorch, P. 1998b. Biostratigraphy, palaeobiogeographical links and environmental interpretation of the Llandovery and Wenlock graptolite faunas of peri-Gondwanan Europe. In Sixth International Graptolite Conference, Madrid, Temas Geológicos-Mineros (Vol. 23, pp. 126-129).
Štorch, P. 2001. Graptolites, stratigraphy and depositional setting of the middle Llandovery (Silurian) volcanic-carbonate facies at Hýskov (Barrandian area, Czech Republic). Bulletin of the Czech Geological Survey, 76:55-76.
Štorch, P. 2006. Facies development, depositional settings and sequence stratigraphy across the Ordovician-Silurian boundary: a new perspective from Barrandian area of the Czech Republic. Geological Journal, 41:163-192.
https://doi.org/10.1002/gj.1038
Štorch, P. 2023. Graptolite biostratigraphy and biodiversity dynamics in the Silurian System of the Prague Synform (Barrandian area, Czech Republic). Bulletin of Geosciences, 98(1).
https://doi.org/10.3140/bull.geosci.1862
Štorch, P., and Loydell, D. 1992. Graptolites of the Rastrites linnaei Group from the European Llandovery (lower Silurian). Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen, 184(1): 63-86.
Štorch, P. and Serpagli, E. 1993. Lower Silurian graptolites from Southwestern Sardinia. Bolletino della Società Paleontologica Italiana, 32, 3-57.
Štorch, P. and Frýda, J. 2012. The late Aeronian graptolite https://doi.org/10.5962/bhl.title.100697 Event, associated positive carbon isotope excursion and facies changes in the Prague Synform (Barrandian area, Bohemia). Geological Magazine, 149(6):1089-1106.
https://doi.org/10.1017/S001675681200026X
Štorch, P., Loydell, D., Melchin, M.J., and Goldman, D. 2024. Graptolites in biostratigraphy: the primary tool for subdivision and correlation of Ordovician, Silurian, and Lower Devonian offshore marine successions. Newsletters on Stratigraphy, (1):1-37.
https://doi.org/10.1127/nos/2024/0810
Strossová, Z. 2024. Lower Telychian (Silurian) species of https://doi.org/10.5962/bhl.title.100697from the linnaei, turriculatus and crispus biozones in the Prague Synform: taxonomy in the light of astogeny and intraspecific variability. Bulletin of Geosciences, 99(1):43-71.
https://doi.org/10.3140/bull.geosci.1889
Sudbury, M. 1958. Triangulate monograptids from the Monograptus gregarius Zone (lower Llandovery) of the Rheidol Gorge (Cardiganshire). Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 241(685):485-554.
Tortello, M.F., Uriz, N.J., Alfaro, M.B., Cingolani, C.A., Bidone, A.R., and Inchausti, J.C.G. 2012. Trilobites and graptolites from the Vargas Peña Formation (early Silurian), Paraná Basin, eastern Paraguay. Revue de Paléobiologie, 10:279-298.
Underwood, C. J. 1998. Population structure of graptolite assemblages. Lethaia, 31(1):33-41.
https://doi.org/10.1111/j.1502-3931.1998.tb00487
Urbanek, A. 1993. Biotic crises in the history of Upper Silurian graptoloids: a palaeobiological model. Historical Biology, 7(1):29-50.
https://doi.org/10.1080/10292389309380442
Vacek, F. and Žák, J. 2019. A lifetime of the Variscan orogenic plateau from uplift to collapse as recorded by the Prague Basin, Bohemian Massif. Geological Magazine, 156(3): 485-509.
https://doi.org/10.1017/S0016756817000875
Vodička, J. and Manda, Š. 2019. A taxonomical and statistical study of chitinozoan distribution across the lundgreni Event (Wenlock, Silurian) from the Prague Basin, Czech Republic: A specific pattern driven by ecological changes. Marine Micropaleontology, 149: 44-63.
https://doi.org/10.1016/j.marmicro.2019.04.002
Yin, T.H. 1937. Brief description of the Ordovician and Silurian fossils from Shihtien. Bulletin of the Geological Society of China, 16:281-302.
https://doi.org/10.1111/j.1755-6724.1937.mp16001012.x
