


Late Triassic small and medium-sized vertebrates from the Fleming Fjord Group of the Jameson Land Basin, central East Greenland
Article number: 28.1.a18
https://doi.org/10.26879/1423
Copyright Palaeontological Association, April 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 19 June 2024. Acceptance: 1 April 2025.
ABSTRACT
The Late Triassic deposits in the Jameson Land Basin, central East Greenland, stand as a crucial fossil area, yielding a diverse Norian vertebrate fauna. This basin, situated at a palaeolatitude of 41° N on the northern rim of Pangea and bordered in the North by the Boreal Sea, was a hub of activity during the Late Triassic. A large ephemeral to perennial lake system developed in the central and eastern parts of the basin, with rivers transporting sediment from the uplands northwest of the basin. Our research focused on the microvertebrate remains recovered from an expedition to Jameson Land in 1991 which yielded significant findings. These specimens, meticulously photographed and listed in a catalogue, comprise 950 vertebrate remains. Notably, we have identified new taxa never described from the Late Triassic sediments of Greenland, including sharks (Lissodus, Rhomphaiodon), bony fish (Gyrolepis), and reptiles (Doswelliidae and Rhynchocephalia). The revision of two sphenodontians jaw fragments as Clevosauridae, in association with Lissodus lepagei, Lissodus cf. Lissodus minimus, Rhomphaiodon sp., Saurichthys sp., and Gyrolepis sp., which are taxa that are highly documented in Europe and Asia, confirms a relationship between faunas of Greenland and Eurasia during the Late Triassic, a statement previously based on macro vertebrate studies.
Valerian J.P. Jésus. LAPC, Centro de Ciências Naturais e Humanas, Universidade Federal do ABC, Rua São Paulo, s/n, Jardim Antares, São Bernardo do Campo/ SP, CEP 09606-070, Brazil. valerian.jesus@ufabc.edu.br
Octávio Mateus. GeoBioTec, Dept. Earth Sciences, Faculdade de Ciências e Tecnologia da Universidade Nova de Lisboa, 2829-516 Monte de Caparica, Portugal and Museu da Lourinhã, Rua João Luís de Moura, 95, 2530-158 Lourinhã, Portugal. omateus@fct.unl.pt
Jesper Milan. Geomuseum Faxe, Østsjællands Museum, Rådhusvej 2, DK-4640 Faxe, Denmark. jesperm@oesm.dk
Lars B. Clemmensen. Department of Geosciences and Natural Resource Management, University of Copenhagen, Oester Voldgade 10, DK-1350 Copenhagen K, Denmark. larsc@ign.ku.dk
Keywords: Chondrichthyes; Actinopterygii; Amphibia; Reptilia; Norian; East Greenland
Final citation: Jésus, Valerian J.P., Mateus, Octávio, Milàn, Jesper, and Clemmensen, Lars B. 2025. Late Triassic small and medium-sized vertebrates from the Fleming Fjord Group of the Jameson Land Basin, central East Greenland. Palaeontologia Electronica, 28(1):a18.
https://doi.org/10.26879/1423
palaeo-electronica.org/content/2025/5520-triassic-greenland-vertebrates
Copyright: April 2025 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
The Triassic follows one of the greatest extinction crises at the Permian-Triassic boundary. The Triassic then saw the emergence of several groups of vertebrates from which the main ones known today evolved, such as dinosaurs, mammals, amphibians, and lizards. For example, it was during the Triassic period that the archosaurs underwent a major phase of diversification, resulting in two major groups, the Pseudosuchia (crocodilians) and the Avemetatarsalia (pterosaurs and dinosaurs) (Nesbitt, 2011). The Triassic also saw the appearance of other archosauriforms, such as the crocodile-like phytosaurs, which occupied an ecological niche comparable to modern crocodiles. Although many groups appeared after the crisis, some were only slightly affected by the events. This is the case of the Hybodontiformes, cartilaginous fishes related to modern sharks and rays, whose diversity was not adversely affected by the Permian-Triassic crisis (Wen et al., 2023). On the contrary, from the Triassic to the Early Jurassic, hybodontiforms were the dominant elasmobranch group in most marine and freshwater environments (Rees and Underwood, 2008).
The Late Triassic lake and fluvial deposits from the Jameson Land Basin of central East Greenland have provided an abundant and diverse vertebrate fauna, comprising chondrichthyans and actinopterygian fishes, dipnoi, theropod and sauropodomorph dinosaurs, temnospondyls, turtles, aetosaurs, phytosaurs, pterosaurs, and mammals (Jenkins et al., 1994, 1997; Sulej et al., 2014, 2020; Clemmensen et al., 2016, 2020; Klein et al., 2016; Lallensack et al., 2017; Marzola et al., 2017a, 2018; Pawlak et al., 2020). Recently, the first occurrence of lissamphibian microvertebrate remains from the Late Triassic sediments was reported (Jésus et al., 2022). However, no extensive study on the microvertebrate assemblages, other than mammals (Jenkins et al., 1994), has been done so far. Evidence of chondrichthyans and actinopterygian remains from the Jameson Land Basin is highlighted by previous works (Clemmensen, 1980a, 1980b; Jenkins et al., 1994). This paper aims to review and describe these small- to medium-sized vertebrate remains collected during previous expeditions and to discuss their palaeobiological importance.
GEOLOGICAL SETTING
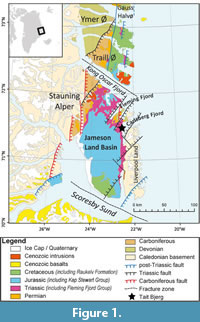 The Jameson Land Basin is located in Jameson Land and Scoresby Land on the central east coast of Greenland, around 71° N (Clemmensen et al., 2020) (Figure 1). Its boundaries are the Stauning Alper in the west, the Liverpool Land area in the east, the fracture zones in the Kong Oscar Fjord region in the north, and the fracture zone in Scoresby Sund in the south (Guarnieri et al., 2017; Clemmensen et al., 2022). During the Late Triassic, East Greenland was located at 41° N on the northern rim of the supercontinent Pangaea, bordered by the Boreal Sea to the North (Clemmensen, 1980a, 1980b; Clemmensen et al., 1998, 2020; Kent and Tauxe, 2005; Nøttvedt et al., 2008; Kent et al., 2014; Andrews and Decou, 2019). The Jameson Land Basin was situated at the southern end of the East Greenland rift system, which formed part of a larger rift complex separating Greenland from Norway before the opening of the Atlantic (Ziegler, 1988; Nøttvedt et al., 2008; Guarnieri et al., 2017). Since the Late Triassic, the basin has rotated 45° clockwise and translated 30° northward, relative to present-day meridians (Kent and Tauxe, 2005).
The Jameson Land Basin is located in Jameson Land and Scoresby Land on the central east coast of Greenland, around 71° N (Clemmensen et al., 2020) (Figure 1). Its boundaries are the Stauning Alper in the west, the Liverpool Land area in the east, the fracture zones in the Kong Oscar Fjord region in the north, and the fracture zone in Scoresby Sund in the south (Guarnieri et al., 2017; Clemmensen et al., 2022). During the Late Triassic, East Greenland was located at 41° N on the northern rim of the supercontinent Pangaea, bordered by the Boreal Sea to the North (Clemmensen, 1980a, 1980b; Clemmensen et al., 1998, 2020; Kent and Tauxe, 2005; Nøttvedt et al., 2008; Kent et al., 2014; Andrews and Decou, 2019). The Jameson Land Basin was situated at the southern end of the East Greenland rift system, which formed part of a larger rift complex separating Greenland from Norway before the opening of the Atlantic (Ziegler, 1988; Nøttvedt et al., 2008; Guarnieri et al., 2017). Since the Late Triassic, the basin has rotated 45° clockwise and translated 30° northward, relative to present-day meridians (Kent and Tauxe, 2005).
The Jameson Land Basin contains a relatively thick Late Triassic succession of continental sediments, including braided and meandering river, ephemeral lake, and perennial lake deposits (Clemmensen, 1980a; Clemmensen et al., 1998, 2020). The lake deposits in the uppermost part of the Triassic succession (the Fleming Fjord Group) are particularly well exposed on steep cliff sides facing the Carlsberg Fjord (Clemmensen et al., 2016, 2020). The Fleming Fjord Group, which has a thickness of about 350 m, consists of the basal Edderfugledal Formation, the middle Malmros Klint Formation, and the uppermost Ørsted Dal Formation, including the Carlsberg Fjord Member (Clemmensen et al., 2020). The basal Edderfugledal Formations may have had sporadic connections to the Boreal Sea, while marine influence was rare or absent in the Malmros Klint and Ørsted Dal Formations. Vertebrate fossils have primarily been found in lake and mudflat deposits in the two uppermost formations (Clemmensen et al., 2020). Most of these vertebrates are continental land animals, as testified by the occurrences of amphibians (Marzola et al., 2017b; Jésus et al., 2022), dinosaurs (Milàn et al., 2004; Lallensack et al., 2017; Beccari et al., 2021), and mammal-like synapsids (Sulej et al., 2014, 2020). Phytosaurs (Milàn et al., 2021; Lopez-Rojas et al., 2022) are usually associated with lake deposits. Hybodonditiforms are attributed to both marine and freshwater environments during the Triassic and the Jurassic (Cuny, 2012), indicating potential connection to the sea.
MATERIALS AND METHODS
The material presented here has been collected during five expeditions involving collaborations between the National History Museum of Denmark, the University of Copenhagen and Harvard University. Professor Farish A. Jenkins Jr. led the first four expeditions, which took place in 1991, 1992, 1995, and 2001. In 1991, the team collected fossils from 84 sites for 40 days (Marzola, 2019: Appendices A7-A11, p. 171-228). Specimens studied here have been recovered during the 1991 expedition and come from three sites on the west side of the Tait Bjerg Mountain (62/91/G, 64/91/G and “No field” #/92/G). Overall, 950 specimens have been observed and identified in this study (Table 1). They include taxa already reported in these sites by previous authors, like phytosaurs and temnospondyls, and newly described taxa that were briefly mentioned, such as chondrichthyans, actinopterygians, and various reptiles.
Sample NHMD 62/91/G is a thin bone bed in the lowermost part of the Carlsberg Fjord Member of the Ørsted Dal Formation, western side of Tait Bjerg. The bone bed is probably formed by a rapid transgression of dry mud flats during an episode of increased water levels in a large ephemeral lake. The bone bed contained two isolated mammalian premolariform teeth as well as isolated reptilian teeth (theropod, prosauropod, and maybe phytosaur ones), amphibian dermal amour, coprolites, and fish remains (Jenkins et al., 1994). Sample “No field” #/92/G is from an unspecified lake mudstone in the uppermost Carlsberg Fjord Member (Ørsted Dal Formation), western side of Tait Bjerg. The bed contained unidentified reptiles and fish remains, including a large jaw. Sample 64/91/G is from a lacustrine limestone/marlstone bed in the uppermost Tait Bjerg Member (Ørsted Dal Formation) on the western side of Tait Bjerg. The bed contained amphibian remains, including Gerrothorax (Jenkins et al., 1994). Here, a more detailed description of the rich vertebrate fauna preserved in these beds is given.
Each material has been individually photographed with a camera (Nikon model D3300 with the lens AF-P DX NIKKOR 18-55 mm 1:3.5-5.6G), a digital microscope (Dinolite pro AM4111T (R4)), or with an autonomous camera (FLEXACAM C1) fixed to a stereomicroscope (Leica M165 C) by stacking multiple pictures (~50) per specimen with Leica Application Suite X (LAS X) from the NOVA. In average, around two or three photographs have been taken for each specimen, among the 3390 pictures in total for 950 specimens. The smallest fossils, like hybodontiform teeth, required using a stereomicroscope (Nexius Zoom Evo, euromex NZ. 1702-S) for observation.
Systematic identifications were kept as conservative as possible, according to the preservation, conservation, and preparation of each specimen. Only material with suitable enough characteristics could be identified at the species level, thanks to the comparison with contemporaneous micro-vertebrate faunas.
Institutional Abbreviations
BGM, Bath Royal Literary & Scientific Institution, Bath, England; BM (NH), Fossil Fish Section, Department of Palaeontology, Natural History Museum, London, England; BRSUG, University of Bristol School of Earth Sciences, Bristol, England; MGUH - NHMD Geological Museum - Natural History Museum of Denmark, Copenhagen, Denmark; PEFO, Petrified Forest National Park, Arizona, United States SMNS, Staatliches Museum für Naturkunde Stuttgart, Stuttgart, Germany; UFRGS-PV-T, Universidade Federal do Rio Grande do Sul (PV, Paleontologia de Vertebrados (Vertebrate Palaeontology); T, Triassic Collection), Porto Alegre, Rio Grande do Sul, Brazil.
SYSTEMATIC PALAEONTOLOGY
Class CHONDRICHTHYES Huxley, 1880
Order HYBODONTIFORMES Patterson, 1966
Superfamily HYBODONTOIDEA, Owen, 1846
Genus LISSODUS Brough, 1935
Lissodus lepagei Duffin, 1993a
(Figure 2A-H)
 Specimens referred. NHMD-1811650 - tooth; NHMD-1811651 - tooth; NHMD-1811652 - tooth; and 270 additional teeth were identified and stored under collections number NHMD-1811705.
Specimens referred. NHMD-1811650 - tooth; NHMD-1811651 - tooth; NHMD-1811652 - tooth; and 270 additional teeth were identified and stored under collections number NHMD-1811705.
Locality. From site 62/91/G, western side of Tait Bjerg, Jameson Land, East Greenland (71°28´34´´ North 22°40´43´´ West).
Horizon and age. Thin bone bed in Carlsberg Fjord Member of the Ørsted Dal Formation in the Fleming Fjord Group. Late Triassic (Norian).
Emended diagnosis. Teeth of Lissodus, which range from very small (around 0.5 mm) to moderately small (up to 3 mm), showing moderate heterodonty. Coronal morphology is generally conservative with a low to high principal cusp, in posterior and anterior teeth, respectively, up to three pairs of lateral cusplets and a crenulate occlusal crest. The moderate to strong labial peg (around 0.5 mm) possesses a strong accessory cusplet and shows considerable variation in shape in occlusal view. Fine to strong vertical ridges are confined to upper lingual and upper labial faces of the crown. A horizontal ridge surrounds the crown at the top of the crown shoulder, without anastomosis.
Description. From the collection, 273 teeth have been assigned to Lissodus lepagei. Those teeth strongly vary in size, from less than 1 mm to almost 3 mm long mesiodistally, never exceedingly further. The crown profile is gracile and narrow, although it can be thicker. The height and width of the crown vary on the tooth, but it is usually 0.5 mm high, excluding the cusps, and 0.4 mm wide labiolingually. The tooth is weakly curved lingually. The crown possesses a high principal cusp and up to three pairs of lateral cusplets in some teeth. The horizontal ridge circles all around the crown at the crown shoulder, at the middle height of the crown. The occlusal crest, being particularly salient, is positioned at the mesiodistal length of the crown, extending through the apices of the principal cusp and lateral cusplets. Each cusp/cusplet is ornamented by at least one vertical ridge on both lingual and labial faces descending from the apices to the horizontal ridge. The principal cusp may display two other non-branching vertical ridges on both sides. Otherwise, the crown surface is smooth. The labial peg is moderately strong and rounded, surmounted by an additional cusplet, discernible depending on the tooth, while the lingual peg is less prominent (Figure 2A-B). The root is preserved on a dozen specimens. It is as high as the crown, approximately 0.5 mm. The crown/root junction is deeply incised around the tooth. It is lingually directed, with its lingual and labial sides showing aligned foramina. Two morphotypes have been identified (Figure 2A-B, E-H), though there is a continuous spectrum of shape variation, with teeth showing an intermediate state of characters suggesting a position between the two morphotypes (Figure 2C-D), representing only the extreme of this spectrum.
Morphotype “Lissodus lepagei 1” (LL1) (Figure 2A-B): It is usually the longest mesiodistally morphotype present. It has the highest cusp, with a height of 1 mm from the crown and lateral cusplets. Concerning the latter, they are longer than high; their height is never higher than half of the principal cusp. The labial peg is prominent and distinguishes itself from the rest of the tooth. The accessory cusplet ornamenting the labial peg is present, although, due to erosion, it can be difficult to discern it or even absent in strongly eroded specimens.
Morphotype “Lissodus lepagei 2” (LL2) (Figure 2E-H): This morphotype is stockier with a lower principal cusp. The lateral cusplets may be absent or less distinguishable. The base is usually concave. In some specimens, the labial vertical ridge descending from the apex of the principal cusp may end at the labial peg ornamented by an accessory cusplet, usually more prominent than in Morphotype LL1.
Remarks. The assignment to Lissodus is based on the presence of a labial protuberance gently sloping towards the crown base (the labial peg), a triangular crown shape in occlusal view, a root lingually inclined marked by a row of small circular foramina on both lingual and labial side (Rees and Underwood, 2002). The description of the numerous specimens from Jameson Land Basin matches the diagnosis of Lissodus lepagei from the middle Norian of Luxembourg, as described by Duffin (1993a). Both specimens from Greenland and Luxembourg have a crenulated occlusal crest, with up to three pairs of lateral cusplets and a variable-shaped labial peg surmounted by a cusplet in some cases (Duffin, 1993a). This species, unlike other Lissodus species, is barely known in the fossil record, reported only in middle Norian of Western Europe countries like France (Grozon), Luxembourg (Syren and Medernach), and probably the Middle Triassic of Spain (Duffin, 1993a; Cuny et al., 1998; Godefroit et al., 1998; Pla et al., 2013). The labial peg is moderately to strongly prominent, sometimes angled mesiodistally. As in the Medernach specimens, the accessory cusplet is present in most of the teeth, but it can be absent due to erosion. Contrary to the specimens from Medernach (Duffin, 1993a), there is a higher degree of variation in size, and morphologically, it could be interpreted as a monognatic heterodonty where Morphotype LL1 might be anterior or lateral teeth, and Morphotype LL2 represents posterior teeth. Morphotype LL2 shows strong morphological similarities with the specimens from Medernach (e.g., holotype BM(NH) P. 62633 in Duffin, 1993a), with a similar size (1 mm long mesiodistally), an occlusal crest with a cutting edge surmounted by a low principal cusp and up to three pairs of lateral cusplets, ornamented by vertical ridges restricted to the upper part of the crown, and a prominent labial peg with an accessory cusplet. Morphotype LL1 differs only by having a very high principal cusp, demarcating itself from the rest of the crown. Following these differences (higher degree of heterodonty, anterior teeth with more prominent principal cusp and a greater range of size variation), we emended the diagnosis as seen previously in the text.
We exclude the identification as to Lissodus nodosus (Seilacher, 1943), known from the Norian of Germany (Duffin, 2001), by the presence of labial and lingual nodes, the lack of prominent lateral cusplets and horizontal ridge on the crown and a labial peg devoid of any accessory cusplet as described in the holotype SMNS 50.214. For similar reasons, the possibility of Lissodus angulatus (Stensiö, 1921) or Lissodus africanus (Broom, 1909) is ruled out as both differ from our specimens by the absence of the accessory cusplet on the labial peg, the lack of lateral cusplets, and no horizontal ridge around the crown shoulder. Moreover, L. lepagei has more gracile teeth with entirely distinguishable cusps, as opposed to the wide labiolingually and low crown present in most Lissodus species (Duffin, 2001). For this study, L. lepagei is considered a valid species of Lissodus, but we recognise that more work on the systematics of Hybodontiformes is needed. For example, L. lepagei was thought to represent juvenile specimens of Lissodus minimus; however, due to the absence of adult L. minimus from the material of Medernach (Luxembourg) this hypothesis might be refuted (Duffin, 1993a). Interestingly, Morphotype LL1 teeth also share similarities with teeth of Parvodus rugianus (Ansorge, 1990), an Early Cretaceous species displaying moderately high principal cusp, two to three pairs of lateral cusplets and a labial peg rounded (Rees, 2002). Parvodus dentition is characterised by a low degree of monognatic heterodonty (Rees, 2002, Rees et al., 2013). The original author, Ansorge, assigned his specimens to Lissodus before they were reviewed as Parvodus (Rees and Underwood, 2002).
Lissodus cf. Lissodus minimus (Agassiz, 1836)
(Figure 2I-N)
Specimens referred. NHMD-1811662 - tooth; NHMD-1811663 - tooth; NHMD-1811664 - tooth; and 174 additional teeth were identified and stored under collections number NHMD-1811707.
Locality. From site 62/91/G, western side of Tait Bjerg, Jameson Land, East Greenland (71°28´34´´ North 22°40´43´´ West).
Horizon and age. Thin bone bed in Carlsberg Fjord Member of the Ørsted Dal Formation in the Fleming Fjord Group. Late Triassic (Norian).
Description. Among the 177 teeth of Lissodus cf. Lissodus minimus, the size varies between morphotypes but never exceeds 3 mm long, except for one incomplete specimen measuring 3.5 mm mesiodistally, with one cusp end missing. The height and width of the crown vary but are around 0.3 mm and 0.5 mm, respectively. The tooth is often curved lingually and has a concave base. The crown comprises a diamond-shaped principal cusp. One pair of low lateral cusplets may be present in some teeth. Different types of ridges ornament the surface of the crown. The horizontal ridge is at the level of the crown shoulder and runs all around the tooth. The occlusal crest runs along the mesiodistal length, passing through the apices of the principal cusp and lateral cusplets. Vertical ridges descend from the apices of the principal cusp and the occlusal crest and lateral cusplets when present. The labial side exhibits a low labial peg perpendicular to the occlusal crest, pointing toward the base of the crown. It can be prominent according to the morphotype. The lingual peg may be less discernible (Figure 2N). The distal ends of the crown are rounded. When preserved, the root is less deep than the crown. The crown/root junction is deeply incised around the tooth. The root follows the same concavity as the base. It is as high as the crown, approximately 0.5 mm. It is lingually directed and ridged horizontally on the labial side, with vascular foramina running parallel to the crown base. Variations have been observed between the numerous specimens, some with eroded cusps and crown surfaces, removing the ridges. At least three morphotypes have been observed in the collection.
Morphotype “ Lissodus minimus 1” (Lm1) (Figure 2I-J): It is usually the smallest morphotype, up to 1.5 mm long. Unlike the pyramidal principal cusp, the crown is lower distally and weakly ridged, almost smooth. Lateral cusplets can be present, their top being lingually curved. The peg is triangular in occlusal view.
Morphotype “Lissodus minimus 2” (Lm2) (Figure 2K-M): It is an intermediate morphology between Morphotype Lm1 and Morphotype Lm3 (see below). They are 2 mm long and 0.5--1 mm wide. They are slightly curved lingually and concave at the base. The teeth show a low crown; the horizontal ridge is closer to the base. The principal cusp is pyramidal and very wide, more bulbous than in the previous morphotype Lm1, while the mesiodistal ends are thinner (half the width of the principal cusp). Lateral cusplets are absent in this morphotype. The labial peg is lower and rounded, shaped like a ‘U.’ The surface is strongly vertically ridged, but the occlusal crest is less sharp than on the anterior teeth.
Morphotype “Lissodus minimus 3” (Lm3) (Figure 2N): This morphotype regroups the most robust teeth. The crown is longer mesiodistally and wider labiolingually. The principal cusp is domed-shaped and lower. The peg cannot be distinguished from the cusp. The base is curved but far less than in the other morphotypes. The tooth is also curved lingually at the distal part. The horizontal ridge is difficult to distinguish as the crown profile is very low. This is the morphotype with most vertical ridges descending from the apex, showing bifurcation between them. No root has been preserved.
Remarks. As for L. lepagei, the assignation to Lissodus is based on a labial peg, a triangular crown shape in occlusal view and a lingually inclined root ornamented by foramina on both the lingual and labial sides (Rees and Underwood, 2002). It is helped by the overall morphology similar to Lissodus minimus (Agassiz, 1836): domed teeth with low cusp, broad and triangular labial peg, and ornamentation with numerous ridges (Duffin, 1985). This species is well known in Northwestern and central Europe: Britain (Allard et al., 2015; Moreau et al., 2021), France (Duffin, 1993b [Saint-Nicolas-de-Port]; Cuny et al., 2000 [Lons-le-Saunier]), Belgium (Duffin and Delsate, 1993), Luxembourg (Syren and Medernach) (Godefroit et al., 1998), Germany (Konietzko-Meier et al., 2019), and Poland (Duffin and Gaździcki, 1977) (Fischer, 2008). Recently, it has been observed in the English Rhaetian on numerous occasions (Korneisel et al., 2015; Norden et al., 2015; Slater et al., 2016; Cross et al., 2018; Moreau et al., 2021), allowing comparison of recently discovered specimens showing multiple examples of the monognathic heterodonty, with three to five different morphotypes: anterior, anterolateral, lateral, postero-lateral, and posterior. Lm1 could be an anterolateral tooth morphotype of Lissodus cf. L. minimus, Lm2 a lateral tooth, and Lm3 a posterior tooth. Duffin (1985) observed two main tooth types that are matched by morphotypes Lm1 and Lm2 (e.g., BGM CD 55 and BGM CD 56 for the first type), and Lm3 (e.g., BGM CD 57 and BGM CD 58 for the second type). According to the author, BGM CD 55 is a mesial tooth, BGM CD 56 an anterolateral tooth, BGM CD 57 a lateral tooth, and BGM CD 58 an extreme lateral tooth (Duffin, 1985), corresponding to the attributed position of Greenland morphotypes. Also, most of the changes mentioned by Duffin (1985) are observed between morphotypes, taking place distally through the dentition starting from the anterior teeth like elongation mesiodistally, the crown becoming shallower, an increase of vertical ridges number and their bifurcation. Since lateral cusplets are absent in most teeth, and their dimensions do not match those of L. minimus described by Duffin (1985, 1993b, 2001), we prefer to use L. cf. L. minimus to refer to the specimens from Greenland.
Hybodontiformes indet.
(Figure 2O-S)
Specimens referred. NHMD-1811665 - dorsal spine; NHMD-1811666 - dermal denticle; NHMD-1811667 - cephalic spine; NHMD-1811726 - dorsal spine; NHMD-1811727 - dorsal spine; NHMD-1811728 - dorsal spine; NHMD-1811729 - dorsal spine; NHMD-1811730 - dorsal spine; NHMD-1811731 - dorsal spine; NHMD-1811732 - dermal denticle; NHMD-1811733 - dermal denticle; NHMD-1811734 - dermal denticle; NHMD-1811735 - dermal denticle; NHMD-1811736 - dermal denticle; NHMD-1811737 - dermal denticle; NHMD-1811738 - cephalic spine; NHMD-1811739 - cephalic spine; NHMD-1811740 - cephalic spine; NHMD-1811741 - cephalic spine; NHMD-1811742 - cephalic spine; NHMD-1811743 - cephalic spine; NHMD-1811744 - cephalic spine; NHMD-1811745 - cephalic spine; NHMD-1811746 - cephalic spine; NHMD-1811747 - cephalic spine; NHMD-1811748 - cephalic spine; NHMD-1811749 - cephalic spine; NHMD-1811750 - cephalic spine; NHMD-1811751 - cephalic spine.
Locality. From site 62/91/G, western side of Tait Bjerg, Jameson Land, East Greenland (71°28´34´´ North 22°40´43´´ West).
Horizon and age. Thin bone bed in Carlsberg Fjord Member of the Ørsted Dal Formation in the Fleming Fjord Group. Late Triassic (Norian).
Description. Seven dorsal spines, about 15 cephalic spines and seven dermal denticles of Hybodontiformes have been identified, although more precise identification was impossible.
Dorsal spines (Figure 2O): The largest dorsal spine is 1.7 cm long and 6 mm wide. They have very pronounced horizontal ridges on the lateral sides and cusplets along the posterior margin. They tend to curve posteriorly in their distal parts.
Cephalic spines (Figure 2P-Q): The cephalic spines have two parts: a base and a crown. The crown is not preserved on any specimen presented here. Most specimens are the T-shaped, tri-radiate base comprising three equally wide branches. The branches are rounded at the end and slightly bent. The surface is usually porous. When the specimen is complete, the crown is the enamelled fourth and most extended branch (compare with Leuzinger et al., 2017: figure 4A-D, p. 476).
Dermal denticles (Figure 2R-S): The dermal denticles have a circular base and a crown. The crown is a cone-like structure with vertical ridges originating at the apex, extending basally to the cusp neck (compare with Klug et al., 2010: figure 5, p.252). The ridges can be more or less pronounced.
Order SYNECHODONTIFORMES Duffin and Ward, 1993
Genus RHOMPHAIODON Duffin, 1993b
Rhomphaiodon sp.
(Figure 3)
 Specimens referred. NHMD-1811668 - tooth; NHMD-1811669 - tooth; NHMD-1811670 - tooth; and 69 additional teeth stored under the collections number NHMD-1811711.
Specimens referred. NHMD-1811668 - tooth; NHMD-1811669 - tooth; NHMD-1811670 - tooth; and 69 additional teeth stored under the collections number NHMD-1811711.
Locality. From site 62/91/G, western side of Tait Bjerg, Jameson Land, East Greenland (71°28´34´´ North 22°40´43´´ West).
Horizon and age. Thin bone bed in Carlsberg Fjord Member of the Ørsted Dal Formation in the Fleming Fjord Group. Late Triassic (Norian).
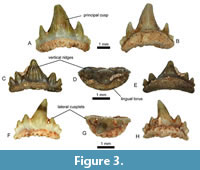
Description. The teeth of this species are the third most abundant remains from site 62/91/G, with 72 specimens. The size is around 1.5--2 mm, but it can reach 3 mm apicobasally and 2 mm mesiodistally. Teeth are symmetrical tricuspids (Figure 3A, D-E), pentacuspid (Figure 3B), or heptacuspid (Figure 3C), but they can be asymmetrical. They display one principal cusp and up to three pairs of lateral cusplets. In rare cases, the number of cusplets is unequal on each side. The connection between the cusp and cusplets has the shape of a V, with an acute angle, sometimes even a right angle in some specimens. Principal cusp is triangular in labial and lingual views. The edges of the triangles can be straight or slightly curved. The principal cusp can be conical or shaped as a grain of rice in occlusal view. It can be curved lingually and occasionally laterally curved, as for the lateral cusplets. The principal cusp is one to 3 mm, and the lateral cusplets decrease in size towards the distal end. On the teeth with a 1 mm high principal cusp, a pattern can be observed in the decrease in size of the lateral cusplets. The first pair of lateral cusplets are inferior to or equal to half of the principal cusp height. When there is a second pair of lateral cusplets, they are half the height of the first pair of lateral cusplets. If there is one, this pattern continues for the third pair of lateral cusplets. Vertical striations run from the apices of the cusp and cusplets to the base of the crown on the lingual and labial sides. The root is usually present. It is lower but wider than the crown, and its height is around one-fifth of the tooth total height or even less than half of the crown in the highest teeth in lingual view. The root is more elevated in labial view, around one-third of the tooth. The base can be curved or completely flat. The root is more extended in its lingual part and gives the shape of a wall to the front of the tooth, as the cusps are close to the labial part. The lingual part of the root is extended lingually forming a broad, bulbous structure called the lingual torus (Figure 3D). The root possesses multiple foramina aligned on both lingual and labial sides. The foramina are close to the base on the labial, and sometimes the pits are not closed. On the lingual side, some foramina are more elongated vertically.
Remarks. The identification is supported by the presence of a crown profile with very high principal cusp and lateral cusplets and the presence of lingual torus on the root. Two species belong to this genus, Rhomphaiodon minor (Agassiz, 1837) and Rhomphaiodon nicolensis Duffin, 1993b. The species R. minor was first described as Hybodus minor by Agassiz (1837) based on a dorsal fin spine. Similarities were noted between R. nicolensis and ‘ Hybodus’ minor by Duffin (1993b). Then, the isolated teeth of R. minor were assigned to the genus Rhomphaiodon by Cuny and Risnes (2005), based on the enameloid microstructure and the morphology, and identified as a member of the Synechodontiformes after a suggestion that the species shares vascularised root typical of Synechodontiformes (Cuny et al., 2000). The genus is common in Europe during the Late Triassic and Early Jurassic (Duffin, 1993b; Duffin and Delsate, 1993; Delsate and Godefroit, 1995; Cuny et al., 2000; Cuny and Risnes, 2005; Norden et al., 2015).
Superclass OSTEICHTHYES Huxley, 1880
Class ACTINOPTERYGII Cope, 1887
Order SAURICHTHYIFORMES Aldinger, 1937
Family SAURICHTHYIDAE Owen, 1860 (sensu Stensiö, 1925)
Genus SAURICHTHYS Agassiz, 1834
Saurichthys sp.
(Figure 4A-C)
 Specimens referred. NHMD-1811674 - tooth; NHMD-1811675 - tooth; NHMD-1811676 - tooth; and 32 additional teeth stored under the collections number NHMD-1811712.
Specimens referred. NHMD-1811674 - tooth; NHMD-1811675 - tooth; NHMD-1811676 - tooth; and 32 additional teeth stored under the collections number NHMD-1811712.
Locality. From site 62/91/G and “No field” # 92/G, western side of Tait Bjerg, Jameson Land, East Greenland (71°28´34´´ North 22°40´43´´ West).
Horizon and age. Thin bone bed in Carlsberg Fjord Member of the Ørsted Dal Formation in the Fleming Fjord Group. Late Triassic (Norian).
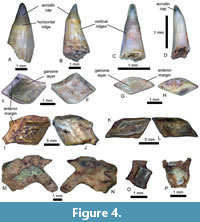
Description. The 35 teeth identified as Saurichthys sp. are elongated and conical, with a circular cross-section. The teeth can be slender or stocky, slightly curved, or even sigmoidal in general shape (Figure 4A-B). Their size varies considerably; the biggest is around 5.5 mm long (Figure 4A), while the smallest measures 1.75 mm (Figure 4C). The acrodin cap, the tip of the tooth, is smooth and can be translucent for the smallest specimens. The cap of the largest teeth can be pearl coloured at the base. There are strong vertical ridges all along the lower part of the teeth, named the shaft. The ridges are thicker closer to the base, giving a folded appearance to the flared base. The shaft and the cap are separated by a neck, a horizontal ridge of one to 2 mm in diameter. The height of the cap depends on the neck position, but usually, the cap corresponds to 10--30% of the total height of the tooth.
Remarks. The teeth are assigned to Saurichthys due to the relative height of the cap, of the total height of the tooth, and the vertically ridged shaft. The dentition of Saurichthys is composed of two distinct sizes: the largest, around 3 mm and the smallest, 1 mm long (see Kogan and Romano, 2016). The largest and longest caps are usually assigned to Saurichthys sp., while the smallest specimens remain doubtful. In that case, they are counted as actinopterygian indeterminate teeth. The teeth have been compared to the teeth of Saurichthys longidens Agassiz, 1844, described in a dozen articles on the microvertebrate fauna of the Rhaetian of England. The morphology of the teeth of Birgeria acuminata differs from that of Saurichthys longidens by being larger with ridges on the lingual side of the cap and both taxa have been described as two morphotypes of Severnichthys acuminatus (Agassiz, 1844) in a dozen of studies on the fauna of the English Rhaetian (Allard et al., 2015; Korneisel et al., 2015; Norden et al., 2015; Lakin et al., 2016; Mears et al., 2016; Slater et al., 2016; Landon et al., 2017; Cavicchini et al., 2018; Cross et al., 2018; Moreau et al., 2021). However, Diependaal and Reumer (2021) considered the genus Severnichthys as a nomen dubium by arguing that Birgeria and Saurichthys are two different animals. Considering the hypothesis of Diependaal and Reumer (2021), and the absence of any teeth sharing a similar morphology with Birgeria among our material, it seems that our material tends to support that said hypothesis. Regarding the material from Greenland, it was not possible to identify to the species-level.
The genus has a global distribution, with records in the Lower Triassic of Alberta, British Columbia, China, France, Great Britain, Greenland, Spitsbergen, Madagascar, Nepal, Tasmania, Australia, Russia, and South Africa (Rieppel, 1992; Bender and Hancox, 2004). The oldest Saurichthys ever found is from the Early Triassic Wordie Creek Formation of East Greenland (Kogan, 2011). Its presence was mainly restricted to the north-western Tethys in the Late Triassic, especially in Europe (Romano et al., 2012). The genus was also observed in the Fleming Fjord Formation, only reported as Saurichthys sp. by Jenkins et al. (1994) but first described here.
Order PALAEONISCIFORMES Hay, 1902
Family PALAEONISCIDAE Vogt, 1852
Genus GYROLEPIS Agassiz, 1835
Gyrolepis sp.
(Figure 4D-H)
Specimens referred. NHMD-1811677 - tooth; NHMD-1811678 - ganoid scale; NHMD-1811679 - ganoid scale; NHMD-1811752 - ganoid scale; NHMD-1811771 - ganoid scale; NHMD-1811772 - ganoid scale; NHMD-1811773 - ganoid scale; NHMD-1811774 - ganoid scale; NHMD-1811775 - ganoid scale; NHMD-1811777 - ganoid scale; NHMD-1811778 - ganoid scale; NHMD-1811779 - ganoid scale; NHMD-1811794 - ganoid scale; NHMD-1811795 - ganoid scale; NHMD-1811796 - ganoid scale; NHMD-1811800 - ganoid scale; NHMD-1811781 - ganoid scale; NHMD-1811782 - ganoid scale; NHMD-1811783 - ganoid scale; NHMD-1811784 - ganoid scale; NHMD-1811785 - ganoid scale; NHMD-1811786 - ganoid scale.
Locality. From site 62/91/G, western side of Tait Bjerg, Jameson Land, East Greenland (71°28´34´´ North 22°40´43´´ West).
Horizon and age. Thin bone bed in Carlsberg Fjord Member of the Ørsted Dal Formation in the Fleming Fjord Group. Late Triassic (Norian).
Description. 22 specimens have been assigned to Gyrolepis sp. including teeth and two different morphotypes of scales.
Teeth (Figure 4D): The teeth are conical, weakly curved, and circular in cross-section. They range in height from around 1 to 1.5 mm. The acrodin cap corresponds to 10% of the tooth total height and is separated from the shaft by a horizontal ridge. The cap is smooth and translucent. Its tip is rounded, but it can be slender and curved. The shaft can have fine vertical striations, but the best-preserved tooth has a smooth shaft and is curved. The base is star-like shaped in cross-section due to grooves at this level, giving the base a flared appearance.
Scale Morphotype “Gyrolepis 1” (Gyr1) (Figure 4E-F): The scale is a rhomboid-shaped piece. The external side is covered by a ganoine layer, ornamented anteroposteriorly by striations that can bifurcate (Figure 4E). The ganoine does not cover the anterior margin of the scale. A broad vertical ridge often marks the internal side (Figure 4F).
Scale Morphotype “Gyrolepis 2” (Gyr2) (Figure 4G-H): It is more elongated in the anteroposterior direction, showing the shape of a lozenge. The scale is thinner than the first morphotype. The external side has the same ganoine layer with a similar pattern of striations (Figure 4G). The internal side does not show any ridge (Figure 4H).
Remarks. The species Gyrolepis albertii Agassiz, 1835 was described from scales from the Muschelkalk (Middle Triassic) of Germany and the Rhaetian of Wickwar (England) (Agassiz, 1835; Mears et al., 2016). Mears et al. (2016) described five scale morphotypes, including one as G. albertii. In this study, the scales and the teeth described are assigned only to the genus Gyrolepis by following the same reasoning from previous studies: similar lozenge shape, striations that bifurcate, and presence of the anterior margin articulating with the other scales (Allard et al., 2015; Mears et al., 2016; Landon et al., 2017; Cavicchini et al., 2018; Cross et al., 2018; Diependaal and Reumer 2021). De Lange et al. (2023) and this paper noted the absence of ornamentation on some scale morphotypes, probably due to erosion.
Scales with similar morphology to the morphotype Gyr1 have been previously reported (Mears et al., 2016; Cavicchini et al., 2018; Cross et al., 2018), although some were not attributed to G. albertii (e.g., specimen BRSUG 29385 in Landon et al., 2017: figure 6k). Scales with similar morphotypes were later assigned to G. albertii by Diependaal and Reumer (2021). It is difficult to distinguish the smallest teeth of Saurichthys sp. from those of Gyrolepis sp., as they are not complete and are vertically striated. Only one tooth, specimen NHMD-1811677, correctly matches the morphology of Gyrolepis sp. dentition. Gyrolepis is assigned to the Palaeoniscidae, a wastebasket group typical of the Carboniferous and Permian likely to be paraphyletic (Korneisel et al., 2015). The taxon has a wide stratigraphic range and needs a taxonomic review (Mears et al., 2016). The sharpness of the teeth suggests a carnivorous diet (Moreau et al., 2021). Gyrolepis presence in Europe has been firmly documented (Allard et al., 2015; Mears et al., 2016; Diependaal and Reumer, 2021). It has already been mentioned in Greenland (Clemmensen, 1980b; Jenkins et al., 1994). Another Palaeoniscidae, Glaucolepis artica Stensiö, 1932, was previously described from the Early Triassic of Greenland, but no teeth have been reported, preventing any comparison with the new specimens.
Actinopterygii indet.
(Figure 4I-P)
Specimens referred. NHMD-1811680 - ganoid scale; NHMD-1811681 - ganoid scale; NHMD-1811682 - ganoid scale; NHMD-1811683 - fin ray element; NHMD-1811684 - vertebral centrum; NHMD-1811788 - ganoid scale; NHMD-1811789 - ganoid scale; NHMD-1811790 - ganoid scale; NHMD-1811791 - ganoid scale; NHMD-1811792 - ganoid scale; NHMD-1811793 - ganoid scale; NHMD-1811797 - ganoid scale; NHMD-1811798 - ganoid scale; NHMD-1811799 - ganoid scale; NHMD-1811801 - ganoid scale; NHMD-1811802 - ganoid scale; NHMD-1811803 - ganoid scale; NHMD-1811804 - ganoid scale; NHMD-1811805 - ganoid scale; NHMD-1811806 - ganoid scale; NHMD-1811807 - ganoid scale; NHMD-1811808 - ganoid scale; NHMD-1811809 - vertebral centrum; NHMD-1811810 - vertebral centrum; NHMD-1811811 - vertebral centrum; NHMD-1811812 - vertebral centrum; NHMD-1811813 - vertebral centrum; NHMD-1811814 - vertebral centrum; NHMD-1811815 - vertebral centrum; NHMD-1811816 - vertebral centrum; NHMD-1811817 - vertebral centrum; NHMD-1811818 - vertebral centrum; NHMD-1811819 - vertebral centrum; NHMD-1811820 - vertebral centrum. nd 18 additional ganoid scales, and 12 vertebral centrums stored under the collections number NHMD-1811714.
Locality. From site 62/91/G on western side of Tait Bjerg, Jameson Land, East Greenland (71°28´34´´ North 22°40´43´´ West).
Horizon and age. Thin bone bed in Carlsberg Fjord Member of the Ørsted Dal Formation in the Fleming Fjord Group. Late Triassic (Norian).
Description. Various actinopterygians remains, such as scales, fin ray elements, teeth acrodin caps, and vertebral centra.
Individual Scales (Figure 4I-N): Many fish scales of various shapes that could not be attributed to a specific actinopterygian clade have been gathered. NHMD-1811680 (Figure 4I-J) is vaguely rhomboid with a rounded angle, with the dorsal edge curved and a peg in the anterior extremity. The external side shows a ganoin layer ornamented by convex sinuous ridges, while the internal side has an oval-shaped convexity. NHMD-1811681 (Figure 4K-L) is a rhomboid that is longer than wider and has a thicker ganoin layer. This layer is smooth with two irregularities: one elongated convexity on the anterior margin and one elongated concavity posteriorly. NHMD-1811682 (Figure 4M-N) presents two pegs, one very short and spike-shaped and the second longer and rectangular. Two foramina are visible on the external side, one next to the second peg and the last foramen adjacent to the first peg. On the internal side, a concavity is present and is shaped like the first peg. This structure could have received the peg of the preceding scale.
Fin ray element (Figure 4O): It is short, 2 mm long, and composed of two parts. The first one is the bony core, which is circular in cross-section. The second part is the ganoine-covered scale, which is flat. The two parts are connected by bone.
Vertebrae (Figure 4P): The vertebral centra are amphicoelic and can be perforated. The base of the haemal arc or the neural arc can be present. Duffin and Gaździcki (1977) described a similar centrum in the Rhaetian of Poland.
Remarks. Site 62/91/G yields unidentified actinopterygian remains, including scales, fin ray elements, teeth acrodin caps, and vertebral centra. They lack diagnostic features to allow further identification. On top of the two-scale morphotypes previously described, seven other ganoid scale shapes are represented, although no species identification could be made. Based on multiple previous studies, it is approved that ganoid scales are assigned to non-teleost fish (Buscalioni et al., 2008), with some Teleosteomorpha taxa displaying ganoid scales (Arratia, 2015; Arratia and Schultze, 2024). Thus, every individual ganoid scale not assigned to Gyrolepis sp. is tentatively attributed to non-teleostean Actinopterygii without going further in the identification process.
Class AMPHIBIA Linnaeus, 1758
Order TEMNOSPONDYLI Zittel, 1888
Family PLAGIOSAURIDAE Abel, 1919
Genus GERROTHORAX Nilsson, 1934
Gerrothorax pulcherrimus (Fraas, 1913)
(Figure 5A)
 Specimens referred. NHMD-1811685 - ornamented skull fragment; NHMD-1811821 - ornamented skull fragment.
Specimens referred. NHMD-1811685 - ornamented skull fragment; NHMD-1811821 - ornamented skull fragment.
Locality. From site 62/91/G, western side of Tait Bjerg, Jameson Land, East Greenland (71°28´34´´ North 22°40´43´´ West).
Horizon and age. Thin bone bed in Carlsberg Fjord Member of the Ørsted Dal Formation in the Fleming Fjord Group. Late Triassic (Norian).
Description. Bone fragment with tubercular ornamentation. The tubercules are conical structures with smooth surfaces.
Remarks. The bone lacks any anatomical feature other than the tubercular ornamentation necessary to identify it. This type of ornamentation is typical of the dermal bone of the skull of Plagiosaurinae since similar ornamentation has been noted in Gerrothorax pulcherrimus, notably the skull, the mandible, and the pectoral girdle, with the other members of Plagiosaurinae Abel, 1919 (Jenkins et al., 2008). However, it displays more prominent tubercles (pustular), as can be seen in several specimens used to reconstruct the skull of G. pulcherrimus (e.g., MGUH 28923 and MGUH 28925 in Jenkins et al., 2008). Of all temnospondyl remains of Greenland, only Gerrothorax displays such ornamentation (Jenkins et al., 1994). In addition, the context of age and location of the specimen is consistent with previous discoveries of the species in three other localities in the basin: MacKnight Bjerg, Sydkronen, and Lepidopteriselv (Jenkins et al., 1994; Marzola et al., 2018). Considering the pustular aspect of the ornamentation, without any continuous ridges, NHMD-1811685 could belong to a large specimen (Schoch and Witzmann, 2011).
Temnospondyli indet.
(Figure 5B-G)
Specimens referred. NHMD-1811686 - tooth; NHMD-1811687 - dentary; NHMD-1811688 - tooth; NHMD-1811822 - tooth; NHMD-1811823 - tooth; NHMD-1811824 - tooth; NHMD-1811825 - tooth; NHMD-1811826 - tooth; NHMD-1811827 - tooth; NHMD-1811828 - tooth; NHMD-1811829 - tooth; NHMD-1811830 - tooth; NHMD-1811831 - tooth; NHMD-1811832 - tooth; NHMD-1811833 - tooth; NHMD-1811834 - tooth; NHMD-1811835 - tooth; NHMD-1811836 - tooth; NHMD-1811837 - tooth; NHMD-1811838 - tooth; NHMD-1811839 - tooth; NHMD-1811840 - maxilla.
Locality. From site 62/91/G, western side of Tait Bjerg, Jameson Land, East Greenland (71°28´34´´ North 22°40´43´´ West).
Horizon and age. Thin bone bed in Carlsberg Fjord Member of the Ørsted Dal Formation in the Fleming Fjord Group. Late Triassic (Norian).
Description. More than 20 temnospondyl remains are reported here. Most of these remains are teeth and two jaw fragments that are too poorly preserved for further identification.
Jaw fragments (Figure 5B-C): There are two jaw fragments. The first, NHMD-1811687 is a dentary of 2 cm long, 2--4 mm wide and 0.5--1 cm high (Figure 5B). The anterior part is broken ventrally. In occlusal view, 14 teeth emplacements are visible with the crown base. All the teeth show folds (Figure 5C). Teeth are circular in cross-section, 2 mm wide. However, only the base is preserved, preventing them from further description. The second jaw fragment is too damaged to identify what bone it is accurately. The circular structures with a similar pattern representing the dentary might suggest it was a tooth-bearing bone of a temnospondyl; however, we cannot determine whether it is a maxillary or a dentary.
Teeth (Figure 5D-G): The 20 isolated teeth are conical and display longitudinal grooves on the lower portion of the crown surface. These teeth are conical, massive, weakly lingually curved, and asymmetrical. The labial surface is more convex in lateral view and rounded in cross-section than the lingual surface. On both mesial and distal sides, the carina exhibits sharp edges. The folds run from the base to the middle of the crown surface but never to the upper part. They vary significantly; the largest tooth has a crown height of 1.7 cm long and a crown base of 75 mm long and 0.6 mm wide; the smallest has a crown height of 3 mm long and a crown base of 2 mm long and 1.5 mm wide.
Remarks. The longitudinal grooves on the isolated teeth and the folds in the teeth of the jaw are typical of temnospondyls, known to possess labyrinthodont teeth. The infolding of their enamel and dentine results in longitudinal grooves on the lower part of the teeth (Kowalski et al., 2009; Rinehart and Lucas, 2013). Many temnospondyls have been found in the Late Triassic of Jameson Land, such as Aquiloniferus kochi, Selenocara groenlandica, Stoschiosaurus nielseni, Tupilakosaurus heilmani, Cyclotosaurus naraserluki, and Gerrothorax (Marzola et al., 2018), which has already been mentioned earlier. The material is only referred to as Temnospondyli indet. since it does not have any characters, allowing a deeper identification.
Class REPTILIA Laurenti, 1768
Clade ARCHOSAURIFORMES Gauthier, 1986
Clade PHYTOSAURIA Jäger, 1828
Family PARASUCHIDAE Lydekker, 1885
Genus MYSTRIOSUCHUS Fraas, 1896
cf. Mystriosuchus alleroq López-Rojas et al. 2022
(Figure 6A-C)
 Specimens referred. NHMD-1811690 - tooth; NHMD-1811691 - tooth; NHMD-1811841 - tooth; NHMD-1811842 - tooth; NHMD-1811843 - tooth; NHMD-1811844 - tooth; NHMD-1811845 - tooth; NHMD-1811846 - tooth; NHMD-1811847 - tooth; NHMD-1811848 - tooth; NHMD-1811850 - tooth; NHMD-1811851 - tooth; NHMD-1811852 - tooth; NHMD-1811853 - tooth; NHMD-1811854 - tooth; NHMD-1811855 - tooth; NHMD-1811856 - tooth; NHMD-1811857 - tooth; NHMD-1811858 - tooth.
Specimens referred. NHMD-1811690 - tooth; NHMD-1811691 - tooth; NHMD-1811841 - tooth; NHMD-1811842 - tooth; NHMD-1811843 - tooth; NHMD-1811844 - tooth; NHMD-1811845 - tooth; NHMD-1811846 - tooth; NHMD-1811847 - tooth; NHMD-1811848 - tooth; NHMD-1811850 - tooth; NHMD-1811851 - tooth; NHMD-1811852 - tooth; NHMD-1811853 - tooth; NHMD-1811854 - tooth; NHMD-1811855 - tooth; NHMD-1811856 - tooth; NHMD-1811857 - tooth; NHMD-1811858 - tooth.
Locality. From site 62/91/G, western side of Tait Bjerg, Jameson Land, East Greenland (71°28´34´´ North 22°40´43´´ West).
Horizon and age. Thin bone bed in Carlsberg Fjord Member of the Ørsted Dal Formation in the Fleming Fjord Group. Late Triassic (Norian).
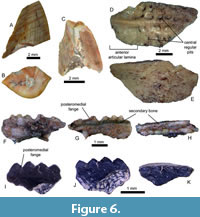
Description. At least 19 isolated teeth have been identified as phytosaur teeth, including NHMD-1811690 and NHMD-1811691. They are conical, slightly compressed labio-lingually on the carina, forming sharp edges on the mesial and distal margins. No strong curvature distally is visible for most teeth observed from this site because only the apical parts are present, except for NHMD-1811690 (Figure 6A). The cross-section varies from D-shaped to oval-flattened, with strong edges slightly closer to the lingual side (Figure 6B). Most preserved specimens consist only of the upper parts of teeth, shaped as an isosceles triangle in lingual or labial views with a long base. Due to bad preservation, apex is not always present, or only fragments of the crown are conserved. The size varies significantly between specimens; the largest tooth, NHMD-1811690, is 6 mm high and has a crown base of 8 mm long and 5 mm wide. NHMD-1811691 is a bit smaller, 5 mm high with a crown base of 4.5 mm long and 2 mm wide. Few specimens preserved their serrated carinae. NHMD-1811691 possesses serration on a 1.5 mm mesial margin towards the crown apex. Even though the denticles are complicated to distinguish as there is no clear separation, the preserved denticles are domed-shaped and wider than high (Figure 6C).
Remarks. The teeth of phytosaurs have already been mentioned in Tait Bjerg (Jenkins et al., 1994) but were barely described back then. Then, another specimen has been described but without teeth associated to the material (Mateus et al., 2014). Today, specimens observed here can be compared to Mystriosuchus alleroq, a phytosaur recently described in the Jameson Land Basin (López-Rojas et al., 2022), whose heterodont teeth have been studied. However, the specimens in this study are poorly preserved, preventing us from pushing further the description (e.g., serration density). Thus, NHMD-1811690 could be assigned to the posterior maxilla region due to its rather flattened and stocky appearance and oval cross-sections. It is more difficult to determine the anatomical tooth position of NHMD-1811691 since an important part from the middle crown to the base is missing. However, from its slightly D-shaped cross-section and straight lingual and labial faces, it could be assigned to the posterior premaxilla set. Still, based on their morphology and the recently described M. alleroq, these teeth are tentatively assigned to this species considering the anatomy and geographical and geological context. Because of the absence of diagnostic tooth characters, we can only assign them to cf. M. alleroq.
Clade PROTEROCHAMPSIA Bonaparte, 1971
Clade DOSWELLIIDAE Weems, 1980
Doswelliidae indet.
(Figure 6D-E)
Specimens referred. NHMD-1811719 - osteoderm.
Locality. From site 62/91/G, western side of Tait Bjerg, Jameson Land, East Greenland (71°28´34´´ North 22°40´43´´ West).
Horizon and age. Thin bone bed in Carlsberg Fjord Member of the Ørsted Dal Formation in the Fleming Fjord Group. Late Triassic (Norian).
Description. Missing its edges, the osteoderm is sub-rectangular. Its dorsal surface is ornamented by a prominent keel running all along the plate and more than 20 central regular pits of subequal size and contour on two-thirds of the total length of the osteoderm (Figure 6D). The remaining third, the anterior articular lamina, is a smooth surface devoid of pits with a less prominent keel, thinner than the rest of the plate. The internal surface is convex and crossed by longitudinal striations bifurcating to the centre, corresponding to the position of the keel (Figure 6E).
Remarks. Since phytosaurs are known with the recently described M. alleroq, it might seem logical to attribute this osteoderm to that species. López-Rojas et al. (2022) described four morphotypes of osteoderms in their supplementary data. We immediately refute the fourth morphotype hypothesis due to the obvious different ornamentation between them and NHMD-1811719, as it consists of small, interconnected ridges in the fourth morphotype. The second morphotype is characterised by a dorsal keel near the medial edge, while it is present in the middle of the dorsal surface in NHMD-1811719. Both the first and third morphotypes display a central mid-dorsal keel, however, the first morphotype keel is more sub-circular and not as prominent and marked as in NHMD-1811719. Regarding the third morphotype, it displays strong lateral projections giving a triangular shape to the osteoderm, which are not present in NHMD-1811719. Moreover, pits in osteoderms of M. alleroq are irregular in size contrary to the regular ones in NHMD-1811719.
Another group of vertebrates wearing osteoderm in the Triassic of Greenland is Aetosauria, represented by Aetosaurus ferratus Fraas, 1877 and Paratypothorax andressorum Long and Ballew, 1985 (Jenkins et al., 1994), both specimens preserved with some osteoderms. Osteoderms of A. ferratus are ornamented with “radiating, oblong, closely, appressed grooves on smooth-surfaced plates”, while in P. andressi they display “an eccentrically placed eminence and radiating groves” (see Jenkins et al., 1994). The difference in ornamentation between Aetosauria and NHMD-1811719 immediately refutes the attribution of the latter to the said group.
Another possibility could be Aetosauriformes such as Revueltosaurus callenderi Hunt and Lucas, 1989 with dorsal paramedian osteoderms (PEFO 34561 and PEFO 42442) described by Parker et al, (2021) as “ornamented with a random pattern or incised circular and oblong pits and having a distinct raised anterior bar along the antero-dorsal edge”. NHMD-1811719 differs by the regularity of its pits and the presence of a more prominent dorsal keel. In R. callenderi, there are also caudal osteoderms (PEFO 34561) which are remarkable for a sharp, raised keel, much more than in NHMD-1811719, but a weak pits ornamentation (Parker et al., 2021), contrary to NHMD-1811719 which has at least two rows of pits on both side of its keel.
The specimen is comparable to the osteoderm of Doswelliidae, a group of non-Archosauria archosauriform known from the Middle Triassic to Upper Triassic in North America, Germany, Poland, and South America (Sues et al., 2013; Czepiński et al., 2023). The clade has two unambiguous synapomorphies among non-Archosauria archosauriform on the osteoderm established by Desojo et al. (2011): (1) ornamentation coarse, incised, and composed of central regular pits of equal size and (2) the presence of an unornamented anterior articular lamina. The description seen before does match these two synapomorphies. The clade is represented in the Norian by one species, Doswellia kaltenbachi Weems, 1980 from the Chinle Formation (USA) (Parker and Barton, 2008; Parker et al., 2021). It is the closest to our specimen, both in space and time. The only species known in Europe is Jaxtasuchus salomoni Schoch and Sues, 2014, from the Ladinian of Erfurt Formation, Germany (Schoch and Sues, 2014) and in the Lower Keuper of Poland (Czepiński et al., 2023). The difference that can be noted between Jaxtasuchus and our material is that the dorsal keel of NHMD-1811719 seems to develop anteriorly, covering the lamina, while in Jaxtasuchus, the anterior articular lamina is flat and smooth. Despite this, and due to the small amount of material available, only one osteoderm, we keep the identification as Doswelliidae indet. since more material is required to go further in the identification.
Superorder LEPIDOSAURIA Haeckel, 1866
Order RHYNCHOCEPHALIA Günther, 1866
Suborder SPHENODONTIA Williston, 1925
Family CLEVOSAURIDAE Bonaparte and Sues, 2006
Clevosauridae indet.
(Figure 6F-K)
Specimens referred. NHMD-1811692 - left maxilla; NHMD-1811693 - right maxilla.
Locality. From site 62/91/G and 64/91/G, western side of Tait Bjerg, Jameson Land, East Greenland (71°28´34´´ North 22°40´43´´ West).
Horizon and age. Thin bone bed in Carlsberg Fjord Member of the Ørsted Dal Formation in the Fleming Fjord Group. Late Triassic (Norian).
Description. The first specimen, NHMD-1811692, is a section from the left maxilla, measuring 3.95 mm long, 0.85 mm wide, and 1.3 mm high without the teeth (Figure 6F-H). Its anterior and posterior margins are both missing. During previous preparation, the specimen was fixed on a nail by a glue bubble, preventing any description of the dorsal margin. To avoid damaging it, it is kept as is. In the lateral view, the maxilla is partially covered by sediments, (Figure 6F). The medial surface is smooth and displays a thick layer that forms a ledge at the bases of the teeth (Figure 6G-H). The ledge is 0.48 mm long labio-lingually. It bears five acrodont teeth, circular in cross-section and with rounded tips, attached to a smooth and thin secondary bone. Their base is elongated anteroposteriorly due to posteromedial flanges, visible in lateral and occlusal views (Figure 6G-H). Except for the rounded apex resulting from use, the lack of important wear patterns on the specimen indicate it must be a juvenile or a sub-adult individual. The average dimensions of the teeth are 0.47 mm long mesio-distally and 0.37 mm wide labiolingually at the crown base and 0.52 mm high for the crown height.
NHMD-1811693 is a tooth-bearing bone, a right maxilla. It measures 2.25 mm long, 1 mm wide and 1.5 mm high (Figure 6I-K). The shape of the anterior extremity suggests that it is the most anterior part of the bone, showing the first four maxillary teeth (Figure 6I, K). Its dorsal margin is broken, as is the posterior part. The lingual and labial edges deviate from each other as they approach the dorsal margin (which is absent). In the dorsal view, the internal structure of the bone is visible, characterised by spongiosis. The dorsal part is concave. Four acrodont conical additional teeth are present on the bone. They are circular in cross-section, slightly extended anteroposteriorly, with sharp triangular posteromedial flanges (Figure 6I-J). They are sharper and higher than the previous specimen but still rounded at the tip. Similarly, they have no sign of wear pattern, suggesting it is a sub-adult considering the additional teeth. The teeth measure, on average, 0.5 mm long and 0.25 mm wide at the base, and 0.5 mm high.
Remarks. The two specimens were previously identified and mentioned as Sphenodontia indet. by Jenkins et al. (1994). NHMD-1811692 was found in association with two premolariform teeth of Kuehneotherium sp. (Jenkins et al., 1994) of the Carlsberg Fjord Member, while NHMD-1811693 was with eight teeth of the same species in a dolomitic limestone at the top of Tait Bjerg, corresponding to the Tait Bjerg Member (Jenkins et al., 1994). Both specimens are considered as maxilla fragments, NHMD-1811692 being the middle part as the three most anterior teeth are hatchling teeth while the two last are additional ones, and NHMD-1811693 would be the part that articulate with the jugal, indicated by the concave dorsal surface (see Chambi-Trowell et al., 2021). Both specimens show multiple features that are typical of sphenodontians. First, the presence of an acrodont dentition is a typical trait of the group (Hsiou et al., 2019; Chambi-Trowell et al., 2021). Then, the secondary bone at the base of the marginal dentition of NHMD-1811692 is characteristic of sphenodontians (Williston, 1925; Hsiou et al., 2019). However, the secondary layer of the first specimen is very thick compared to other specimens. Still, a similar condition seems to be present in a juvenile specimen (UFRGS-PV-0613-T) of Microsphenodon bonapartei Chambi-Trowell et al., 2021, from the Late Triassic of Brazil (Romo-de-Vivar-Martínez et al., 2021: figure 4b-c, p. 5). NHMD-1811693 does not have a secondary bone, making it difficult to identify it accurately. The alternation in size of the hatchling teeth is also characteristic of sphenodontians (Harrison, 1901; Fraser, 1988). The posteromedial flanges are present in many sphenodontians, particularly the species of Clevosaurus Swinton, 1939 (Sues et al., 1994; Säilä, 2005; Jones, 2006a; Romo-de-Vivar-Martínez and Bento Soares, 2015; Hsiou et al., 2015, 2019; O’Brien et al., 2018). The shape of the teeth and the sharp posteromedial flanges are according to the omnivorous or carnivorous diet of clevosaurs (Jones, 2006b; Rauhut et al., 2012; Martínez et al., 2013). NHMD-1811692 and NHMD-1811693 are similar to juvenile Clevosaurus maxillae from the Triassic of Britain and Brazil, displaying the same type of dentition (Fraser, 1988, figure 40; Romo-de-Vivar-Martínez and Bento Soares, 2015, figure 4f-g; Romo-de-Vivar-Martínez et al., 2021, figure 4b-c). Based on these similarities with clevosaurs, we tentatively assign these two specimens to Clevosauridae.
DISCUSSION
Of the 950 specimens photographed, only 840 have been identified to the class. Throughout the collection, chondrichthyan is the most common (~68% of all material), primarily due to Hybodontiformes remains (~60%). Actinopterygians (~14%) are less represented, while tetrapods (~6%) are rare. The last 11% are the indeterminate material that could not be assigned to any taxa. Tait Bjerg shows great diversity, as most vertebrate groups are present, including mammals described by Jenkins et al. (1994) (Table 1). It also yields a dozen coprolite specimens present in the collection. Despite this taxonomic diversity, many specimens remain to be identified, and other areas need to be treated appropriately in relation to the study of microvertebrates.
Deposition of the Late Triassic Fleming Fjord Group in the Jameson Land Basin occurred in a large lake system that evolved from semi-perennial to ephemeral and finally back to perennial (Clemmensen et al., 2020). Rivers brought coarse-grained sediments from source areas northwest of the basin to the lake system. Lake deposition lasted from about 220 Ma to about 209 Ma, while deposition of the vertebrate-bearing Carlsberg Fjord Member took place between 214 and 211 Ma (Kent and Clemmensen, 2021; Mau et al., 2022). Fossil evidence, including a rich terrestrial vertebrate fauna including amphibians, mammals and dinosaurs (e.g., Jenkins et al., 1994), seemed to underline the strictly continental nature of the basin. The presence of hybodontiforms, such as Lissodus lepagei, known as freshwater sharks, reinforces the hypothesis of the lake system. However, Saurichthys is known to have a global distribution in both freshwater and marine environments (Romano et al., 2012; Kligman et al., 2017), and Gyrolepis might have had a lower tolerance to freshwater than the former (de Lange et al., 2023). Therefore, the last two taxa support the hypothesis of occasional connections to the sea during the initial and final phases of lake evolution, as suggested by Clemmensen (1980a).
Identifying Hybodontiformes teeth was the most difficult struggle in this study due to size, shape and character variations. Many specimens are too eroded to allow an accurate identification beyond hybodontiform. Others are preserved enough to display characters but lack the details to confirm their morphotype. Indeterminate teeth could not be precisely assigned, as they are similar to L. lepagei and L. cf. Lissodus minimus. On top of the morphological complexity of the teeth from Greenland, Lissodus phylogenetic position is unresolved among Hybodontiformes because of shared characters with both Lonchidiidae and Acrodontinae, the former showing a significant disparity, with delicate, thin teeth or wide and low crushing teeth (Rees, 2008; Stumpf and Kriwet, 2019). Hence, intermediate forms between each morphotype prevent accurate identification.
On another point concerning Lissodus, although it has been proven that L. minimus and L. lepagei are two different species and not an ontogenetic sequence, the differentiation between them often seems confusing. The confirmed presence of L. lepagei in France and Luxembourg (Duffin, 1993a, 1993b; Cuny et al., 1998) suggests a very restricted distribution in Western Europe, unlike L. minimus. However, this new occurrence in Greenland, in addition to that possible one in Spain (Pla et al. 2013), leads us to rethink a more extensive distribution and, if possible, to review the Norian and Rhaetian fossil record in other European countries. Another possibility to explain this absence could be an identification bias in the publications of recent years. For example, L. lepagei has never been observed in vertebrate assemblages from the Rhaetian described recently, despite several specimens sharing morphological similarities and characters in common with L. lepagei (see Korneisel et al., 2015; Norden et al., 2015; Slater et al., 2016; Cross et al., 2018; Moreau et al., 2021; de Lange et al., 2023). It is, therefore, potentially probable that these specimens from Rhaetian of England, identified as anterior teeth of L. minimus, could be teeth of L. lepagei, but further study needs to be done.
More importantly, Jenkins et al. (1994) already reported the presence of indeterminate hybodont sharks, Saurichthys and Gyrolepis, although no descriptions or pictures are available. Yet, the authors did not mention the presence of Lissodus lepagei or Rhomphaiodon. This is the first time Lonchidiidae, Synechodontiformes, and Doswelliidae have been reported and described in Greenland. The two first are well documented in the Triassic of Europe (Duffin and Gaździcki, 1977; Duffin and Delsate, 1993; Bernardi et al., 2010; Allard et al., 2015; Antczak et al., 2020; Moreau et al., 2021). The presence of temnospondyls is not a surprise since expeditions of Greenland have revealed multiple well-preserved specimens from the expeditions that have been previously reported and described (Marzola et al., 2017b, 2018). Phytosaur remains have already been registered in Jameson Land Basin, to which can be added Mystriosuchus alleroq, with a nearly complete mandible as a holotype recently published (López-Rojas et al., 2022).
The presence of rhynchocephalians has already been reported, although not studied yet. The possible affiliation to Diphydontosaurus or other basal rhynchocephalian might suggest that the two first specimens mentioned by Jenkins et al. (1994) could be referred to as similar taxa. Plus, those rhynchocephalian specimens are associated with specimens of Lissodus, Rhomphaiodon, Saurichthys, and Gyrolepis, comparable with Eurasian species (Wenz, 1967; Duffin and Gaździcki, 1977; Chen et al., 2007; Allard et al., 2015; Keeble et al., 2018). It is congruent with the hypothesis of a strong relationship between the Eurasian and Greenland faunas, at least for the Jameson Land Basin and its surroundings and adds new information on the palaeogeographical and palaeolatitudinal distribution of Late Triassic faunistic provinces (Jenkins et al., 1994; Marzola et al., 2017c, 2018; Clemmensen et al., 2015; Agnolin et al., 2018; Marzola, 2019; Pawlak et al 2020).
CONCLUSION
The microvertebrate assemblage in this study yielded more than 900 specimens mainly from site 62/91/G, in the Carlsberg Fjord Member of Tait Bjerg, Jameson Land, East Greenland, recording the substantial diversity of the Carlsberg Fjord Member vertebrates, with the main Triassic clades represented. Most specimens were identified but more than 120 specimens could not be identified to genus or family level. The result of this work strongly supports the hypothesis that the relationships between faunas from Greenland and Eurasia highlight the importance of detailed studies of microvertebrate assemblages of ancient bone beds. Overall, the Jameson Land Basin Norian aquatic fauna is comparable to coeval European and Asian faunas, predominantly composed of sharks (Hybodontiformes and Synechodontiformes) and bony fishes (Saurichthyiformes and Palaeonisciformes), represented mainly by teeth. This is the first time Synechodontiformes, Lonchidiidae, and Doswelliidae have been reported and described in Greenland. Until this work, there was no extensive study of Jameson Land Basin microvertebrate assemblage other than mammals undertaken by Jenkins et al. (1994). For this reason, this work aimed to point out the richness of Jameson Land Basin fauna, presenting the aquatic fauna and tetrapod remains ignored before such as hybodontiforms, doswelliids, and clevosaurids.
The study of the microvertebrate fauna of Jameson Land Basin has reinforced the previous hypothesis of European connection and faunal exchange, with a closer look into the smaller specimens of previously known and unknown taxa.
ACKNOWLEDGEMENTS
This research benefited from the GeoBioTec-GeoBioSciences, GeoTechnologies, and GeoEngineering NOVA [GeoBioCiências, GeoTecnologias e GeoEngenharias], grant UIDB/04035/2020 by the Fundação para a Ciência e Tecnologia. This study could not have been possible without the team that uncovered these specimens, F.A. Jenkins Jr., W.W. Amaral, W.R. Downs III, S.M. Gatesy, N.H. Shubin Niels Bonde, and L.B. Clemmensen. We thank Harvard University and L. Cotton and I. Bonnevier Wallstedt from the National History Museum of Denmark. Heartfelt thanks to A. Guillaume, V. López-Rojas and V. Beccari for reviewing and bringing improvements to the original manuscript and C. Tomás for laboratory support. Thanks to M.X. Mailho Maia for translation to Portuguese of the abstract. And finally, the anonymous reviewers are thanked for their comments on the manuscript, increasing its quality.
REFERENCES
Abel, O. 1919. Die Stämme der Wirbeltiere.Walter de Gruyter. Berlin.
Agassiz, L. 1833-1843. Recherches sur les poissons fossiles (5 volumes). Imprimerie de Petitpierre, Neuchâtel. Retrieved from
https://doi.org/10.5962/bhl.title.4275
Agnolin, F.L., Mateus, O., Milàn, J., Marzola, M., Wings, O., Adolfssen, J.S., and Clemmensen, L.B. 2018. Ceratodus tunuensis, sp. nov., a new lungfish (Sarcopterygii, Dipnoi) from the Upper Triassic of central East Greenland. Journal of Vertebrate Paleontology, 38:e1439834.
https://doi.org/10.1080/02724634.2018.1439834
Aldinger, H. 1937. Permische ganoidfische aus Ostgrönland. Reitzels Forlag.
Allard, H., Carpenter, S.C., Duffin, C.J., and Benton, M.J. 2015. Microvertebrates from the classic Rhaetian bone beds of Manor Farm Quarry, near Aust (Bristol, UK). Proceedings of the Geologists’ Association, 126:762–776. https://doi.org/10.1016/j.pgeola.2015.09.002
Andrews, S.D. and Decou, A. 2019. The Triassic of Traill Ø and Geographical Society Ø, East Greenland: Implications for North Atlantic palaeogeography. Geological Journal, 54:2124–2144.
https://doi.org/10.1002/gj.3287
Ansorge, J. 1990. Fischreste (Selachii, Actinopterygii) aus der Wealdentonscholle von Lobber Ort (Münchgut/Rügen/DDR). Fischreste (Selachii, Actinopterygii) aus der Wealdentonscholle von Lobber Ort (Münchgut/Rügen/DDR), 64:133–144.
Antczak, M., Ruciński, M.R., Stachacz, M., Matysik, M., and Król, J.J. 2020. Diversity of vertebrate remains from the Lower Gogolin Beds (Anisian) of southern Poland. Annales Societatis Geologorum Poloniae, 90:419–433.
https://doi.org/10.14241/asgp.2020.22
Arratia, G. 2015. Complexities of Early Teleostei and the Evolution of Particular Morphological Structures through Time. Copeia, 103:999–1025.
https://doi.org/10.1643/CG-14-184
Arratia, G. and Schultze, H.-P. 2024. The oldest teleosts (Teleosteomorpha): their early taxonomic, phenotypic, and ecological diversification during the Triassic. Fossil Record, 27:29–53.
https://doi.org/10.3897/fr.27.115970
Beccari, V., Mateus, O., Wings, O., Milan, J., and Clemmensen, L.B. 2021. Issi saaneq gen. et sp. nov.--A New Sauropodomorph Dinosaur from the Late Triassic (Norian) of Jameson Land, Central East Greenland.
https://doi.org/10.3390/d13110561
Bender, P.A. and Hancox, P.J. 2004. Newly Discovered Fish Faunas from the Early Triassic, Karoo Basin, South Africa, and their Correlative Implications. Gondwana Research, 7:185–192.
https://doi.org/10.1016/S1342-937X(05)70317-8
Bernardi, M., Petti, F.M., and Avanzini, M. 2010. A webbed archosaur footprint from the Upper Triassic (Carnian) of the Italian Southern Alps. New Mexico Museum of Natural History and Science, Bulletin, 51:65–68.
Bonaparte, J.F. 1971. Cerritosaurus binsfeldi Price, tipo de una nueva familia de tecodontes (Pseudosuchia-Prorterochampsia). Anais da Academia Brasileira de Ciencias, 43:417–422.
Bonaparte, J.F. and Sues, H.-D. 2006. A new species of Clevosaurus (Lepidosauria: Rhynchocephalia) from the Upper Triassic of Rio Grande do sul, Brazil. Palaeontology, 49:917–923.
https://doi.org/10.1111/j.1475-4983.2006.00568.x
Broom, R. 1909. The fossil fishes of the Upper Karoo beds of South Africa. Annals of the South Africa Museum, 7:251–269.
Brough, J. 1935. On the structure and relationships of the hybodont sharks. Memoirs and Proceeding of the Manchester Literary and Philosophical Society, 79:35–49.
Buscalioni, A.D., Fregenal, M.A., Bravo, A., Poyato-Ariza, F.J., Sanchíz, B., Báez, A.M., Cambra Moo, O., Martín Closas, C., Evans, S.E., and Marugán Lobón, J. 2008. The vertebrate assemblage of Buenache de la Sierra (Upper Barremian of Serrania de Cuenca, Spain) with insights into its taphonomy and palaeoecology. Cretaceous Research, 29:687–710.
https://doi.org/10.1016/j.cretres.2008.02.004
Cavicchini, I., Heyworth, H.C., Duffin, C.J., Hildebrandt, C., and Benton, M.J. 2018. April 1. A Rhaetian microvertebrate fauna from Stowey Quarry, Somerset, U.K. Proceedings of the Geologists’ Association, 129:144–158.
https://doi.org/10.1016/j.pgeola.2018.02.001
Chambi-Trowell, S.A.V., Martinelli, A.G., Whiteside, D.I., Vivar, P.R.R.D., Soares, M.B., Schultz, C.L., Gill, P.G., Benton, M.J., and Rayfield, E.J. 2021. The diversity of Triassic South American sphenodontians: a new basal form, clevosaurs, and a revision of rhynchocephalian phylogeny. Journal of Systematic Palaeontology, 19:787–820.
https://doi.org/10.1080/14772019.2021.1976292
Chen, L., Cuny, G., and Wang, X. 2007. The chondrichthyan fauna from the Middle-Late Triassic of Guanling (Guizhou province, SW China). Historical Biology, 19:291–300.
https://doi.org/10.1080/08912960701248234
Clemmensen, L., Milàn, J., Adolfssen, J., Estrup, E., Frobøse, N., Klein, N., Mateus, O., and Wings, O. 2016. The vertebrate-bearing Late Triassic Fleming Fjord Formation of central East Greenland revisited: Stratigraphy, palaeoclimate and new palaeontological data. Geological Society, London, Special Publications, 434.
https://doi.org/10.1144/SP434.3
Clemmensen, L.B. 1980a. Triassic rift sedimentation and palaeogeography of central East Greenland. Bulletin Grønlands Geologiske Undersøgelse, 136:1–72.
https://doi.org/10.34194/bullggu.v136.6678
Clemmensen, L.B. 1980b. Triassic lithostratigraphy of East Greenland between Scoresby Sund and Kejser Franz Josephs Fjord. Bulletin Grønlands Geologiske Undersøgelse, 139:1–56.
Clemmensen, L.B., Kent, D.V., and Jenkins, F.A. 1998. A Late Triassic lake system in East Greenland: facies, depositional cycles and palaeoclimate. Palaeogeography, Palaeoclimatology, Palaeoecology, 140:135–159.
https://doi.org/10.1016/S0031-0182(98)00043-1
Clemmensen, L.B., Kent, D.W., Mau, M., Mateus, O., and Milàn, J. 2020. Triassic lithostratigraphy of the Jameson Land Basin (central EastGreenland), with emphasis on the new Fleming Fjord Group. Bulletin of the Geological Society of Denmark, 68:95–132.
https://doi.org/10.37570/bgsd-2020-68-05-rev
Clemmensen, L.B., Lindström, S., Mateus, O., Mau, M., Milàn, J., and Kent, D.V. 2022. A new vertebrate fossil-bearing layer in the Rhætelv Formation (Kap Stewart Group) of central East Greenland: evidence of a Hettangian marine incursion into the continental Jameson Land Basin. Lethaia, 55:1–12.
https://doi.org/10.1111/let.12449
Cope, E.D. 1887. Zittel’s manual of palaeontology. American Naturalist, 21:1014–1019.
Cross, S.R.R., Ivanovski, N., Duffin, C.J., Hildebrandt, C., Parker, A., and Benton, M.J. 2018. Microvertebrates from the basal Rhaetian Bone Bed (latest Triassic) at Aust Cliff, S.W. England. Proceedings of the Geologists’ Association, 129:635–653.
https://doi.org/10.1016/j.pgeola.2018.06.002
Cuny, G., Martin, M., Rauscher, R., and Mazin, J.-M. 1998. A new neoselachian shark from the Upper Triassic of Grozon (Jura, France). Geological Magazine, 135:657–668.
https://doi.org/10.1017/S0016756898001435
Cuny, G., Hunt, A., Mazin, J.-M., and Rauscher, R. 2000. Teeth of enigmatic neoselachian sharks and an ornithischian dinosaur from the uppermost Triassic of Lons-le-Saunier (Jura, France). Paläontologische Zeitschrift, 74:171–185.
https://doi.org/10.1007/BF02987959
Cuny, G. and Risnes, S. 2005. The enameloid microstructure of the teeth of synechodontiform sharks (Chondrichthyes: Neoselachii). PalArch’s Journal of Vertebrate Palaeontology, 3:1–19.
De Lange, B., Chenal, E., Diependaal, H.J., and Reumer, J.W.F. 2023. Fish remains from the Rhaetian (Late Triassic) of Winterswijk, the Netherlands (Pisces: Chondrichthyes and Actinopterygii). Netherlands Journal of Geosciences, 102:e10.
https://doi.org/10.1017/njg.2023.10
Delsate, D. and Godefroit, P. 1995. Chondrichthyens du Toarcien inférieur d’Aubange (Lorraine belge). Belgian Geological Survey Professional Paper, 278:23–43.
Desojo, J.B., Ezcurra, M.D., and Schultz, C.L. 2011. An unusual new archosauriform from the Middle-Late Triassic of southern Brazil and the monophyly of Doswelliidae. Zoological Journal of the Linnean Society, 161:839–871.
https://doi.org/10.1111/j.1096-3642.2010.00655.x
Diependaal, H.J. and Reumer, J.W.F. 2021. Note on the Rhaetian fish fauna from a subrosion pipe in Winterswijk (the Netherlands), with a discussion on the validity of the genus Severnichthys Storrs, 1994. Netherlands Journal of Geosciences, 100:e9.
https://doi.org/10.1017/njg.2021.5
Duffin, C.J. 1985. Revision of the hybodont selachian genus Lissodus Brough (1935). Palaeontographica, A, 188:105–152.
Duffin, C.J. 2001. Synopsis of the selachian genus Lissodus Brough. Neues Jahrbuch fur Geologie und Palaontotoqie Abhandlungen, 221:145–218.
Duffin, C.J. 1993a. Mesozoic chondrichthyan faunas. 1. Middle Norian (Upper Triassic) of Luxembourg. Palaeontographica Abteilung A, 229:15–36.
Duffin, C.J. 1993b. Late Triassic shark teeth (Chondrichthyes, Elasmobranchii) from Saint-Nicolas-de-Port (north-east France). Professional Paper Geological Survey of Belgium, 7–32.
Duffin, C.J. and Gaździcki, A. 1977. Rhaetian fish remains from the Tatra Mountains. Acta Geologica Polonica, 27:333–348.
Duffin, C.J. and Delsate, D. 1993. The age of the Upper Triassic vertebrate fauna from Attert (Province of Luxembourg, Belgium). Professional Paper Geological Survey of Belgium, 33–44.
Duffin, C.J. and Ward, D.J. 1993. The Early Jurassic palaeospinacid sharks of Lyme Regis, southern England. Professional Paper Geological Survey of Belgium, 53–102.
Fischer, J. 2008. Brief synopsis of the hybodont form taxon Lissodus Brough, 1935, with remarks on the environment and associated fauna. Freiberger Forschungshefte, 528:1–23.
Fraas, E. 1896. Die schwäbischen Trias-Saurier nach dem Material der Kgl. Naturalien-Sammlung in Stuttgart zusammengestellt. E. Schweizerbart’sche Verlagshandlung.
Fraas, E. 1913. Neue Labyrinthodonten aus der schwabischen Trias. Palaeontographica, 60:275–294.
Fraas, O. 1877. Aetosaurus ferratus Fr. Schweizerbart’sche Verlagsbuchhandlung.
Fraser, N.C. 1988. The osteology and relationships of Clevosaurus (Reptilia: Sphenodontida). Philosophical Transactions of the Royal Society of London Series B, 321:125–178.
https://doi.org/10.1098/rstb.1988.0092
Gauthier, J. 1986. Saurischian monophyly and the origin of birds. Memoirs of the California Academy of sciences, 8:1–55.
Godefroit, P., Cuny, G., Delsate, D., and Roche, M. 1998. Late Triassic Vertebrates from Syren (Luxembourg). Neues Jahrbuch fur Geologie und Palaontologie - Abhandlungen, 210:305–343.
https://doi.org/10.1127/njgpa/210/1998/305
Guarnieri, P., Brethes, A., and Rasmussen, T.M. 2017. Geometry and kinematics of the Triassic rift basin in Jameson Land (East Greenland). Tectonics, 36:602–614.
https://doi.org/10.1002/2016TC004419
Günther, A. 1866. Contribution to the Anatomy of Hatteria (Rhynchocephalus, Owen). Philosophical Transactions of the Royal Society of London, 15:460–462.
Haeckel, E. 1866. Generelle morphologie der organismen. Allgemeine grundzüge der organischen formen-wissenschaft, mechanisch begründet durch die von Charles Darwin reformirte descendenztheorie. Verlag von Georg Reimer, Berlin.
https://doi.org/10.5962/bhl.title.3953
Harrison, H.S. 1901. The development and succession of teeth in Hatteria punctata. Journal of Cell Science, 2:161–213. https://doi.org/10.1242/jcs.s2-44.174.161
Hay, O.P. 1902. Bibliography and catalogue of the fossil vertebrata of North America. Bulletin of the United States Geological Survey, 179:1–868.
https://doi.org/10.5962/bhl.title.20094
Hsiou, A.S., De França, M.A.G., and Ferigolo, J. 2015. New data on the Clevosaurus (Sphenodontia: Clevosauridae) from the Upper Triassic of southern Brazil. PLoS One, 10:e0137523. Public Library of Science.
https://doi.org/10.1371/journal.pone.0137523
Hsiou, A.S., Nydam, R.L., Simões, T.R., Pretto, F.A., Onary, S., Martinelli, A.G., Liparini, A., Martínez, P.R.R. de V., Soares, M.B., Schultz, C.L., and Caldwell, M.W. 2019. A new clevosaurid from the Triassic (Carnian) of Brazil and the rise of sphenodontians in Gondwana. Scientific Reports, 9:11821.
https://doi.org/10.1038/s41598-019-48297-9
Hunt, A.P. and Lucas, S.G. 1989. A new? ornithischian dinosaur from the Bull Canyon Formation (Upper Triassic) of east-central New Mexico, p. 355–358. In Lucas, S.G. and Hunt, A.P. (eds), Dawn of the age of dinosaurs in the American Southwest: Albuquerque, New Mexico Museum of Natural History.
Huxley, T.H. 1880. On the application of the laws of evolution to the arrangement of the vertebrate and more particularly of the Mammalia, p. 43, 649–662. Proceedings of the Zoological Society, London.
Jäger, G.F. von. 1828. Über die fossile Reptilien, welche in Würtemberg aufgefunden worden sind. Metzler.
Jenkins Jr., F.A., Shubin, N.H., Amaral, W.W., Gatesy, S.M., Schaff, C.R., Clemmensen, L.B., Downs, W.R., Davidson, A.R., Bonde, N., and Osbæck, F. 1994. Late Triassic continental vertebrates and depositional environments of the Fleming Fjord Formation, Jameson Land, east Greenland. Meddelelser om Greenland Geoscience, 32:1–25.
https://doi.org/10.7146/moggeosci.v32i.140904
Jenkins Jr., F.A., Gatesy, S.M., Shubin, N.H., and Amaral, W.W. 1997. Haramiyids and Triassic mammalian evolution. Nature, 385:715–718.
https://doi.org/10.1038/385715a0
Jenkins Jr., F.A., Shubin, N.H., Gatesy, S.M., and Warren, A. 2008. Gerrothorax pulcherrimus from the Upper Triassic Fleming Fjord Formation of East Greenland and a reassessment of head lifting in temnospondyl feeding. Journal of Vertebrate Paleontology, 28:935–950.
https://doi.org/10.1671/0272-4634-28.4.935
Jésus, V.J.P., Mateus, O., Milan, J., and Clemmensen, L.B. 2022. First occurrence of a frog-like batrachian (Amphibia) in the Late Triassic Fleming Fjord Group, central. Bulletin of the Geological Society of Denmark, 70:117–130.
https://doi.org/10.37570/bgsd-2022-70-08
Jones, M.E.H. 2006a. The Early Jurassic clevosaurs from China (Diapsida: Lepidosauria), p. 37, 548–562. New Mexico Museum of Natural History and Science Bulletin.
Jones, M.E.H. 2006b. Tooth diversity and function in the Rhynchocephalia (Diapsida: Lepidosauria), p. 55–58. In Barrett, P.M. and Evans, S.E. (eds.), Ninth International Symposium on Mesozoic Terrestrial Ecosystems and Biota. Natural History Museum, London.
Keeble, E., Whiteside, D.I., and Benton, M.J. 2018. The terrestrial fauna of the Late Triassic Pant-y-ffynnon Quarry fissures, South Wales, UK and a new species of Clevosaurus (Lepidosauria: Rhynchocephalia). Proceedings of the Geologists’ Association, 129:99–119.
https://doi.org/10.1016/j.pgeola.2017.11.001
Kent, D.V. and Clemmensen, L.B. 2021. Northward dispersal of dinosaurs from Gondwana to Greenland at the mid-Norian (215-212 Ma, Late Triassic) dip in atmospheric p CO2. Proceedings of the National Academy of Sciences, 118:e2020778118.
https://doi.org/10.1073/pnas.2020778118
Kent, D.V. and Tauxe, L. 2005. Corrected Late Triassic latitudes for continents adjacent to the North Atlantic. Science, 307:240–244.
https://doi.org/10.1126/science.1105826
Kent, D.V., Santi Malnis, P., Colombi, C.E., Alcober, O.A., and Martínez, R.N. 2014. Age constraints on the dispersal of dinosaurs in the Late Triassic from magnetochronology of the Los Colorados Formation (Argentina). Proceedings of the National Academy of Sciences, 111:7958–7963.
https://doi.org/10.1073/pnas.1402369111
Klein, H., Milàn, J., Clemmensen, L.B., Frobøse, N., Mateus, O., Klein, N., Adolfssen, J.S., Estrup, E.J., and Wings, O. 2016. Archosaur footprints (cf. Brachychirotherium) with unusual morphology from the Upper Triassic Fleming Fjord Formation (Norian-Rhaetian) of East Greenland. Geological Society, London, Special Publications, 434:71–85.
https://doi.org/10.1144/SP434.1
Kligman, B., Parker, W., and Marsh, A. 2017. First record of Saurichthys (Actinopterygii) from the Upper Triassic (Chinle Formation, Norian) of western North America. Journal of Vertebrate Paleontology, 37:e1367304.
https://doi.org/10.1080/02724634.2017.1367304
Klug, S., Tütken, T., Wings, O., Pfretzschner, H.-U., and Martin, T. 2010. A Late Jurassic freshwater shark assemblage (Chondrichthyes, Hybodontiformes) from the southern Junggar Basin, Xinjiang, northwest China. Palaeobiodiversity and Palaeoenvironments, 90:241–257.
https://doi.org/10.1007/s12549-010-0032-2
Kogan, I. 2011. Remains of Saurichthys (Pisces, Actinopterygii) from the Early Triassic Wordie Creek Formation of East Greenland. Bulletin of the Geological Society of Denmark, 59:93–100.
https://doi.org/10.37570/bgsd-2011-59-09
Kogan, I. and Romano, C. 2016. Redescription of Saurichthys madagascariensis Piveteau, 1945 (Actinopterygii, Early Triassic), with implications for the early saurichthyid morphotype. Journal of Vertebrate Paleontology, 36:e1151886.
https://doi.org/10.1080/02724634.2016.1151886
Konietzko-Meier, D., Werner, J.D., Wintrich, T., and Martin Sander, P. 2019. A large temnospondyl humerus from the Rhaetian (Late Triassic) of Bonenburg (Westphalia, Germany) and its implications for temnospondyl extinction. Journal of Iberian Geology, 45:287–300.
https://doi.org/10.1007/s41513-018-0092-0
Korneisel, D., Gallois, R.W., Duffin, C.J., and Benton, M.J. 2015. Latest Triassic marine sharks and bony fishes from a bone bed preserved in a burrow system, from Devon, UK. Proceedings of the Geologists’ Association, 126:130–142.
https://doi.org/10.1016/j.pgeola.2014.11.004
Kowalski, J., Bodzioch, A., Janecki, P.A., Ruciński, M.R., and Antczak, M. 2019. Preliminary report on the microvertebrate faunal remains from the Late Triassic locality at Krasiejów, SW Poland. Annales Societatis Geologorum Poloniae, 89:291–305.
https://doi.org/10.14241/asgp.2019.10
Lakin, R.J., Duffin, C.J., Hildebrandt, C., and Benton, M.J. 2016. The Rhaetian vertebrates of Chipping Sodbury, South Gloucestershire, UK, a comparative study. Proceedings of the Geologists’ Association, 127:40–52.
https://doi.org/10.1016/j.pgeola.2016.02.010
Lallensack, J.N., Klein, H., Milàn, J., Wings, O., Mateus, O., and Clemmensen, L.B. 2017. Sauropodomorph dinosaur trackways from the Fleming Fjord Formation of East Greenland: evidence for Late Triassic sauropods. Acta Palaeontologica Polonica, 62:833–843.
https://doi.org/10.4202/app.00374.2017
Landon, E.N.U., Duffin, C.J., Hildebrandt, C., Davies, T.G., Simms, M.J., and Benton, M.J. 2017. The first discovery of crinoids and cephalopod hooklets in the British Triassic. Proceedings of the Geologists’ Association, 128:360–373.
https://doi.org/10.1016/j.pgeola.2017.03.005
Laurenti, J.N. 1768. Specimen medicum : exhibens synopsin reptilium emendatam cum experimentis circa venena et antidota reptilium austriacorum. Joan Thomae, Viennae.
https://doi.org/10.5962/bhl.title.5108
Leuzinger, L., Cuny, G., Popov, E., and Billon-Bruyat, J.-P. 2017. A new chondrichthyan fauna from the Late Jurassic of the Swiss Jura (Kimmeridgian) dominated by hybodonts, chimaeroids and guitarfishes. Papers in Palaeontology, 3:471–511.
https://doi.org/10.1002/spp2.1085
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species; cum characteribus, differentiis, synonymis, locis. Holmiae [Stockholm].
Long, R.A. and Ballew, K.L. 1985. Aetosaur dermal armor from the Late Triassic of southwestern North America, with special reference to material from the Chinle Formation of Petrified Forest National Park. Museum of Northern Arizona Bulletin 47:45–68.
López-Rojas, V., Clemmensen, L.B., Milàn, J., Wings, O., Klein, N., and Mateus, O. 2022. A new phytosaur species (Archosauriformes) from the Upper Triassic of Jameson Land, central East Greenland. Journal of Vertebrate Paleontology, 42:e2181086.
https://doi.org/10.1080/02724634.2023.2181086
Lydekker, R. 1885. The Reptilia & Amphibia of the Maleri & Denwa Groups. Geological Survey Office.
Martínez, R.N., Apaldetti, C., Colombi, C.E., Praderio, A., Fernandez, E., Malnis, P.S., Correa, G.A., Abelin, D., and Alcober, O. 2013. A new sphenodontian (Lepidosauria: Rhynchocephalia) from the Late Triassic of Argentina and the early origin of the herbivore opisthodontians. Proceedings of the Royal Society B: Biological Sciences, 280:20132057.
https://doi.org/10.1098/rspb.2013.2057
Marzola, M. 2019. The Late Triassic vertebrate fauna of the Jameson Land Basin, East Greenland: description, phylogeny, and paleoenvironmental implications. Faculdade de Ciências e Tecnologia, PhD Thesis, Universidade Nova de Lisboa, Portugal, Lisboa.
Marzola, M., Mateus, O., Milàn, J., and Clemmensen, L.B. 2017a. The 2016 Dinosaur Expedition to the Late Triassic of the Jameson Land Basin, East Greenland, 249-253. Abstract book of the XV Encuentro de Jóvenes Investigadores en Paleontología, Pombal.
Marzola, Marco, Mateus, O., Shubin, N.H., and Clemmensen, L.B. 2017b. Cyclotosaurus naraserluki, sp. nov., a new Late Triassic cyclotosaurid (Amphibia, Temnospondyli) from the Fleming Fjord Formation of the Jameson Land Basin (East Greenland). Journal of Vertebrate Paleontology, 37:e1303501.
https://doi.org/10.1080/02724634.2017.1303501
Marzola, M., Mateus, O., Milàn, J., and Clemmensen, L. 2017c. European affinities of the Late Triassic biota from Greenland are related to paleolatitude. Presented at the 15th Annual Meeting of the European Association of Vertebrate Palaeontologists, Munich, Germany, p. Zitteliana, 91.
Marzola, M., Mateus, O., Milàn, J., and Clemmensen, L.B. 2018. A review of Palaeozoic and Mesozoic tetrapods from Greenland. Bulletin of the Geological Society of Denmark, 66:21–46.
https://doi.org/10.37570/bgsd-2018-66-02
Mateus, O., Butler, R.J., Brusatte, S.L., Whiteside, J.H., and Steyer, J.S. 2014. The first phytosaur (Diapsida, Archosauriformes) from the Late Triassic of the Iberian Peninsula. Journal of Vertebrate Paleontology, 34:970–975.
https://doi.org/10.1080/02724634.2014.840310
Mau, M., Bjerrum, C.J., and Clemmensen, L.B. 2022. Late Triassic paleowinds from lacustrine wave ripple marks in the Fleming Fjord Group, central East Greenland. Palaeogeography, Palaeoclimatology, Palaeoecology, 586:110776.
Mears, E.M., Rossi, V., MacDonald, E., Coleman, G., Davies, T.G., Arias-Riesgo, C., Hildebrandt, C., Thiel, H., Duffin, C.J., and Whiteside, D.I. 2016. The Rhaetian (Late Triassic) vertebrates of Hampstead Farm Quarry, Gloucestershire, UK. Proceedings of the Geologists’ Association, 127:478–505.
Milàn, J., Clemmensen, L., and Bonde, N. 2004. Vertical sections through dinosaur tracks (Late Triassic lake deposits, East Greenland) - undertracks and other subsurface deformation structures revealed. Lethaia, 37:285–296.
https://doi.org/10.1080/00241160410002036
Milàn, J., Octávio, M., Mau, M., Rudra, A., Sanei, H., and Clemmensen, L.B. 2021. A possible phytosaurian (Archosauria, Pseudosuchia) coprolite from the Late Trias-sic Fleming Fjord Group of Jameson Land, central East Greenland. Bulletin of the Geological Society of Denmark, 69:71–80.
https://doi.org/10.37570/bgsd-2021-69-05
Moreau, M., Duffin, C.J., Hildebrandt, C., Hutchinson, D., Parker, A., Carpenter, S., and Benton, M.J. 2021. Microvertebrates from the Rhaetian basal bone bed of Saltford, near Bath, SW England. Proceedings of the Geologists’ Association, 132:174–187.
https://doi.org/10.1016/j.pgeola.2020.11.003
Nesbitt, S.J. 2011. The Early Evolution of Archosaurs: Relationships and the Origin of Major Clades. Bulletin of the American Museum of Natural History, 2011(352):1–292.
https://doi.org/10.1206/352.1
Nilsson, T. 1934. Vorläufige Mitteilung über einen Stegocephalenfund aus dem Rhät Schonens. Geologiska Föreningen i Stockholm Förhandlingar, 56:428–442.
https://doi.org/10.1080/11035893409444893
Nordén, K.K., Duffin, C.J., and Benton, M.J. 2015. A marine vertebrate fauna from the Late Triassic of Somerset, and a review of British placodonts. Proceedings of the Geologists’ Association, 126:564–581.
https://doi.org/10.1016/j.pgeola.2015.07.001
Nøttvedt, A., Johannessen, E.P., and Surlyk, F. 2008. The Mesozoic of Western Scandinavia and East Greenland. Episodes Journal of International Geoscience, 31:59–65. International Union of Geological Sciences.
https://doi.org/10.18814/epiiugs/2008/v31i1/009
O’Brien, A., Whiteside, D.I., and Marshall, J.E.A. 2018. Anatomical study of two previously undescribed specimens of Clevosaurus hudsoni (Lepidosauria: Rhynchocephalia) from Cromhall Quarry, UK, aided by computed tomography, yields additional information on the skeleton and hitherto undescribed bones. Zoological Journal of the Linnean Society, 183:163–195.
https://doi.org/10.1093/zoolinnean/zlx087
Owen, R. 1846. Lectures on the comparative anatomy and physiology of the vertebrate animals1. Longman, Brown, Green, and Longmans.
Owen, R. 1860. Palaeontology, or a systematic summary of extinct animals and their geological relations. Adam and Charles Black.
Parker, W.G. and Barton, B.J. 2008. New information on the Upper Triassic archosauriform Vancleavea campi based on new material from the Chinle Formation of Arizona. Palaeontologia Electronica, 11:14A.
https://palaeo-electronica.org/2008_3/158/index.html
Parker, W.G., Nesbitt, S.J., Marsh, A.D., Kligman, B.T., and Bader, K. 2021. First occurrence of Doswellia cf. D. kaltenbachi (Archosauriformes) from the Late Triassic (middle Norian) Chinle Formation of Arizona and its implications on proposed biostratigraphic correlations across North America during the Late Triassic. Journal of Vertebrate Paleontology, 41:e1976196.
https://doi.org/10.1080/02724634.2021.1976196
Patterson, C. 1966. British wealden sharks. Bulletin of the British Museum (Natural History) Geology, 11:281--350.
https://doi.org/10.5962/p.150189
Pawlak, W., Tałanda, M., Sulej, T., and Niedźwiedzki, G. 2020. Dipnoan from the Upper Triassic of East Greenland and remarks about palaeobiogeography of Ptychoceratodus. Acta Palaeontologica Polonica 65 (3): 561-574.
Pla, C., Márquez-Aliaga, A., and Botella, H. 2013. The chondrichthyan fauna from the Middle Triassic (Ladinian) of the Iberian Range (Spain). Journal of Vertebrate Paleontology, 33:770–785.
https://doi.org/10.1080/02724634.2013.748668
Rauhut, O.W., Heyng, A.M., López-Arbarello, A., and Hecker, A. 2012. A new rhynchocephalian from the Late Jurassic of Germany with a dentition that is unique amongst tetrapods. PLOS One, 7:e46839.
Rees, J. 2002. Shark fauna and depositional environment of the earliest Cretaceous Vitabäck Clays at Eriksdal, southern Sweden. Earth and Environmental Science Transactions of The Royal Society of Edinburgh, 93:59–71.
Rees, J. 2008. Interrelationships of Mesozoic hybodont sharks as indicated by dental morphology - preliminary results. Acta Geologica Polonica, 58:217–221.
Rees, J. and Underwood, C.J. 2002. The status of the shark genus Lissodus Brough, 1935, and the position of nominal Lissodus species within the Hybodontoidea (Selachii). Journal of Vertebrate Paleontology, 22:471–479.
https://doi.org/10.1671/0272-4634(2002)022[0471:TSOTSG]2.0.CO;2
Rees, J. and Underwood, C.J. 2008. Hybodont sharks of the English Bathonian and Callovian (Middle Jurassic). Palaeontology, 51:117–147.
https://doi.org/10.1111/j.1475-4983.2007.00737.x
Rees, J., Cuny, G., Pouech, J., and Mazin, J.-M. 2013. Non-marine selachians from the basal Cretaceous of Charente, SW France. Cretaceous Research, 44:122–131.
https://doi.org/10.1016/j.cretres.2013.04.002
Rieppel, O. 1992. A new species of the genus Saurichthys (Pisces: Actinopterygii) from the Middle Triassic of Monte San Giorgio (Switzerland), with comments on the phylogenetic interrelationships of the genus. Palaeontographica. Abteilung A, Paläozoologie, Stratigraphie, 221:63–94.
Rinehart, L.F. and Lucas, S.G. 2013. Tooth form and function in temnospondyl amphibians: relationship of shape to applied stress. New Mexico Museum of Natural History Bulletin, 61:533–542.
Romano, C., Kogan, I., Jenks, J., Jerjen, I., and Brinkmann, W. 2012. Saurichthys and other fossil fishes from the late Smithian (Early Triassic) of Bear Lake County (Idaho, USA), with a discussion of saurichthyid palaeogeography and evolution. Bulletin of Geosciences, 543–570.
https://doi.org/10.3140/bull.geosci.1337
Romo-de-Vivar-Martínez, P.R. and Bento Soares, M. 2015. Dentary morphological variation in Clevosaurus brasiliensis (Rhynchocephalia, Clevosauridae) from the Upper Triassic of Rio Grande do Sul, Brazil. PLoS One, 10:e0119307.
Romo-de-Vivar-Martínez, P.R., Martinelli, A.G., Paes Neto, V.D., Scartezini, C.A., Lacerda, M.B., Rodrigues, C.N., and Bento Soares, M. 2021. New rhynchocephalian specimen in the Late Triassic of southern Brazil and comments on the palatine bone of Brazilian rhynchocephalians. Historical Biology, 33:209–217.
https://doi.org/10.1080/08912963.2019.1602616
Säilä, L.K. 2005. A new species of the sphenodontian reptile Clevosaurus from the Lower Jurassic of South Wales. Palaeontology, 48:817–831.
https://doi.org/10.1111/j.1475-4983.2005.00486.x
Schoch, R.R. and Witzmann, F. 2011. Cranial morphology of the plagiosaurid Gerrothorax pulcherrimus as an extreme example of evolutionary stasis. Lethaia, 45: 371–385.
https://doi.org/10.1111/j.1502-3931.2011.00290.x
Schoch, R.R. and Sues, H.-D. 2014. A new archosauriform reptile from the Middle Triassic (Ladinian) of Germany. Journal of Systematic Palaeontology, 12:113–131.
https://doi.org/10.1080/14772019.2013.781066
Seilacher, A. 1943. Elasmobranchier-Reste aus dem oberen Muschelkalk und dem Keuper Württembergs. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Monatshefte B, 256–292.
Slater, T.S., Duffin, C.J., Hildebrandt, C., Davies, T.G., and Benton, M.J. 2016. Microvertebrates from multiple bone beds in the Rhaetian of the M4-M5 motorway junction, South Gloucestershire, U.K. Proceedings of the Geologists’ Association, 127:464–477.
https://doi.org/10.1016/j.pgeola.2016.07.001
Stensiö, E.A. 1921. Triassic Fishes from Spitsbergen. Part IA. Holzhausen, Vienna.
Stensiö, E.A. 1925. Triassic fishes from Spitzbergen 2. Kungliga Svenska Vetenskapsakademiens Handlingar 3(2), 1–261.
Stensiö, E.A. 1932. Triassic fishes from East Greenland: ollected by the Danish expeditions in 1929-1931. Meddelelser om Greenland, 83:1–305.
Storrs, G.W. 1994. Fossil vertebrate faunas of the British Rhaetian (latest Triassic). Zoological Journal of the Linnean Society, 112:217–259.
https://doi.org/10.1111/j.1096-3642.1994.tb00319.x
Stumpf, S. and Kriwet, J. 2019. A new Pliensbachian elasmobranch (Vertebrata, Chondrichthyes) assemblage from Europe, and its contribution to the understanding of late Early Jurassic elasmobranch diversity and distributional patterns. PalZ, 93:637–658.
https://doi.org/10.1007/s12542-019-00451-4
Sues, H.-D., Desojo, J.B., and Ezcurra, M.D. 2013. Doswelliidae: a clade of unusual armoured archosauriforms from the Middle and Late Triassic. Geological Society, London, Special Publications, 379:49–58.
https://doi.org/10.1144/SP379.13
Sues, H.-D., Shubin, N.H., and Olsen, P.E. 1994. A new sphenodontian (Lepidosauria: Rhynchocephalia) from the McCoy Brook Formation (Lower Jurassic) of Nova Scotia, Canada. Journal of Vertebrate Paleontology, 14:327–340.
https://doi.org/10.1080/02724634.1994.10011563
Sulej, T., Krzesiński, G., Tałanda, M., Wolniewicz, A.S., Błażejowski, B., Bonde, N., Gutowski, P., Sienkiewicz, M., and Niedźwiedzki, G. 2020. The earliest-known mammaliaform fossil from Greenland sheds light on origin of mammals. Proceedings of the National Academy of Sciences, 117:26861–26867.
https://doi.org/10.1073/pnas.2012437117
Sulej, T., Wolniewicz, A., Bonde, N., Błażejowski, B., Niedźwiedzki, G., and Tałanda, M. 2014. New perspectives on the Late Triassic vertebrates of East Greenland: preliminary results of a Polish−Danish palaeontological expedition. Polish Polar Research, 35:541–552.
https://doi.org/10.2478/popore-2014-0030
Swinton, W.E. 1939. A new Triassic rhynchocephalian from Gloucestershire. Annals and Magazine of Natural History, 4:591–594.
https://doi.org/10.1080/00222933908655407
Vogt, C. 1852. Ueber die Siphonophoren. Zeitschrift für wissenschaftliche Zoologie, 3:522–525.
Weems, R.E. 1980. An unusual newly discovered archosaur from the Upper Triassic of Virginia, USA. Transactions of the American Philosophical Society, 70:1–53.
Wen, W., Zhang, Q., Kriwet, J., Hu, S., Zhou, C., Huang, J., Cui, X., Min, X., and Benton, M.J. 2023. First occurrence of hybodontid teeth in the Luoping Biota (Middle Triassic, Anisian) and recovery of the marine ecosystem after the end-Permian mass extinction. Palaeogeography, Palaeoclimatology, Palaeoecology, 617:111471.
https://doi.org/10.1016/j.palaeo.2023.111471
Wenz, S. 1967. Compléments à l’étude des poissons actinoptérygiens du Jurassique français6. Éditions du Centre national de la recherche scientifique.
Williston, S.W., Williston, S.W., and Gregory, W.K. 1925. The osteology of the reptiles1–324. Harvard University Press, Cambridge, Massachussetts.
https://doi.org/10.5962/bhl.title.6573
Ziegler, P.A. 1988. Evolution of the Arctic-North Atlantic and the Western Tethys.
Zittel, K.A. von. 1880. Handbuch der palaeontologie. Abteilung Palaeozoologie. R. Oldenbourg, Munich and Leipzig.
