

 Theropod tooth morphotypes from the Andrés fossil site: Insights into a highly diverse fauna of European Late Jurassic carnivore dinosaurs
Theropod tooth morphotypes from the Andrés fossil site: Insights into a highly diverse fauna of European Late Jurassic carnivore dinosaurs
Article number: 28.3.a53
https://doi.org/10.26879/1504
Copyright Paleontological Society, November 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Appendices
Submission: 22 October 2024. Acceptance: 9 November 2025.
ABSTRACT
The Andrés fossil site (Pombal, Portugal) is well known by its abundant record of fossils attributed to the theropod Allosaurus. However, a diversity of other vertebrates has also been identified in this Upper Jurassic locality, including fishes, lepidosaurs, crocodylomorphs, pterosaurs and different dinosaur groups. The Late Jurassic vertebrate fossil record in other European areas is scarce and mostly represented by isolated teeth, highlighting the importance of detailed descriptions and comprehensive analyses of these fossils to approach the faunal diversity in these landmasses. Here, an assemblage of isolated theropod teeth collected from Tithonian levels at the Andrés locality in the Lusitanian Basin is described. A combination of different methods, including discriminant and cladistic analyses as well as machine learning tools, was used to assess their taxonomic identification. These analyses allowed to characterize ten different morphotypes attributed to Allosaurus and to several groups of small coelurosaurian theropods, including indeterminate Coelurosauria, early-branching Tyrannosauroidea, Neocoelurosauria, Velociraptorinae, and Dromaeosaurinae. This study revealed an unusually diverse theropod dinosaur fauna, showing some similarities to that described from the Kimmeridgian lignite levels of the famous Guimarota fossil site (Leiria, Portugal). Other tooth morphotypes from different Upper Jurassic European localities are comparable to those described at the Andrés fossil site, suggesting similarities in theropod faunal composition. The study of this large assemblage of isolated theropod teeth allow us to better understand the palaeobiodiversity, paleogeographic distribution, and evolutionary history of the Late Jurassic theropod fauna of the Iberian Peninsula and other correlative European areas.
Elisabete Malafaia. Instituto Dom Luiz, Faculdade de Ciências, Universidade de Lisboa. Campo Grande, 1749-016 Lisboa, Portugal and Grupo de Biología Evolutiva, Facultad de Ciencias, Universidad Nacional de Educación a Distancia (UNED), Avenida de Esparta S/N, 28232 Las Rozas de Madrid, Madrid, Spain (corresponding author). efmalafaia@ciencias.ulisboa.pt
Filipa Batista. Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, Campus da Caparica, 2829-516 Caparica, Portugal. filipa.lopes.batista@gmail.com
Bruno Maggia. Instituto Dom Luiz, Faculdade de Ciências, Universidade de Lisboa. Campo Grande, 1749-016 Lisboa, Portugal and Grupo de Biología Evolutiva, Facultad de Ciencias, Universidad Nacional de Educación a Distancia (UNED), Avenida de Esparta S/N, 28232 Las Rozas de Madrid, Madrid, Spain. BrunoMaggia@gmail.com
Carolina S. Marques. Centro de Estatística e Aplicações, Faculdade de Ciências, Universidade de Lisboa. Campo Grande, 1749-016 Lisboa, Portugal. csmarques@ciencias.ulisboa.pt
Fernando Escaso. Grupo de Biología Evolutiva, Facultad de Ciencias, Universidad Nacional de Educación a Distancia (UNED), Avenida de Esparta S/N, 28232 Las Rozas de Madrid, Madrid, Spain. fescaso@ccia.uned.es
Pedro Dantas. Museu Nacional de História Natural e da Ciência, Universidade de Lisboa, Rua da Escola Politécnica 56, 1250-102 Lisboa, Portugal and Sociedade de História Natural, Apartado 25, 2564-909 Torres Vedras, Portugal. pedromdantas@gmail.com
Francisco Ortega. Grupo de Biología Evolutiva, Facultad de Ciencias, Universidad Nacional de Educación a Distancia (UNED), Avenida de Esparta S/N, 28232 Las Rozas de Madrid, Madrid, Spain. fortega@ccia.uned.es
Keywords: Allosaurus; Coelurosauria; Lusitanian Basin; morphometric; cladistic; machine learning
Final citation: Malafaia, Elisabete, Batista, Filipa, Maggia, Bruno, Marques, Carolina S., Escaso, Fernando, Dantas, Pedro, and Ortega, Francisco. 2025. Theropod tooth morphotypes from the Andrés fossil site: insights into a highly diverse fauna of European Late Jurassic carnivore dinosaurs. Palaeontologia Electronica, 28(3):a53.
https://doi.org/10.26879/1504
https://palaeo-electronica.org/content/2025/5721-diversity-of-theropod-teeth-from-the-upper-jurassic-of-portugal
Copyright: November 2025 Paleontological Society
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
The Late Jurassic fossil record of theropod dinosaurs from the Iberian Peninsula is relatively abundant, particularly in the sedimentary units representing the Mesozoic infill of the Lusitanian Basin (central west of Portugal). Despite several more complete specimens having been found in these levels, isolated teeth are the most common theropod fossils in the Lusitanian Basin. In other European Upper Jurassic localities, body fossils of theropods are scarcer and are mostly represented by isolated teeth (see Appendix 1). Therefore, detailed descriptions and taxonomic discussions of tooth morphotypes are critical for accessing the diversity, evolutionary history, and palaeoecology of these dinosaurs. A large sample of isolated teeth, along with some postcranial material of theropod dinosaurs, has been described from different Upper Jurassic (Kimmeridgian) localities in northern Germany. This material represents a relatively diverse fauna composed of possible Ceratosauria, different Megalosauroidea, including cf. Marshosaurus and cf. Torvosaurus, Allosauroidea (some tentatively related to Allosaurus), indeterminate Coelurosauria, and early-branching Tyrannosauroidea (e.g., Gerke and Wings, 2016; Evers and Wings, 2020). Some of these specimens were first interpreted as belonging to velociraptorine dromaeosaurids (Lubbe et al., 2009). In the Bavaria region (southern Germany), an important record of avialan theropods has been described from Kimmeridgian-Tithonian levels of the “Solnhofen Limestones” (e.g., Owen, 1863; Wellnhofer, 1988; Mayr et al., 2005; Foth and Rauhut, 2017; Rauhut et al., 2018a). Some non-avian theropods have also been found in these levels, including the megalosauroid Sciurumimus albersdoerferi and the coelurosaurians Juravenator starki and Compsognathus longipes (Wagner, 1861; Göhlich and Chiappe, 2006; Rauhut et al., 2012).
From different Upper Jurassic (Oxfordian and Kimmeridgian) localities in England several osteological remains referred to Theropoda have been described, including the allosauroid Metriacanthosaurus parkeri and the early branching tyrannosauroid Stokesosaurus langhami (e.g., Huxley, 1869; Lydekker, 1888; Walker, 1964; Benson, 2008; Martill et al., 2006; Naish and Martill, 2007). The Late Jurassic of France has also yielded a relatively large sample of theropod remains, including an almost complete and well-preserved skeleton collected from a Tithonian lithographic limestone quarry at Canjuers, attributed to Compsognathus longipes (e.g., Bidar et al., 1972; Peyer, 2006). The Middle to Upper Jurassic Vaches Noires cliffs in Calvados (Normandy) have produced several theropod remains, including the holotypes of Streptospondylus altdorfensis and Piveteausaurus divesensis, as well as cranial and postcranial elements attributed to indeterminate tetanurans, megalosaurids, and allosauroids, (Piveteau, 1923; Taquet and Welles, 1977; Knoll et al., 1999; Allain, 2001; Buffetaut et al., 2010; Monvoisin et al., 2022). From the Tithonian of Chassiron (western France), a relatively large sample of isolated theropod teeth, representing morphotypes interpreted as belonging to spinosaurids, possible megalosaurids and allosaurids, dromaeosaurids, and troodontids has been described (Vullo et al., 2014).
Skeletal theropod remains are scarce in the Upper Jurassic of Spain and mostly consist of isolated teeth and fragmentary postcranial elements. Several isolated teeth and a few postcranial elements from Kimmeridgian localities in Asturias have been assigned to indeterminate theropods, probable ceratosaurians, indeterminate tetanurans, megalosaurids, possible carcharodontosaurids, and indeterminate maniraptorans (e.g., Canudo and Ruiz-Omeñaca, 2001; Ruiz-Omeñaca et al., 2008; Rauhut et al., 2018b). Also, from Tithonian-Berriasian localities in Teruel, isolated teeth and scarce postcranial material have been identified to several theropod clades, including megalosaurids, allosaurids, and dromaeosaurids (e.g., Canudo et al., 2005, 2006; Gascó et al., 2012; Cobos et al., 2014).
The Lusitanian Basin has yielded an abundant Late Jurassic theropod fossil record, mostly represented by large and medium size forms, including Ceratosaurus, Torvosaurus, Lourinhanosaurus, Allosaurus, and Lusovenator (e.g., Mateus, 1998; Pérez-Moreno et al., 1999; Rauhut and Fechner, 2005; Mateus et al., 2006; Hendrickx and Mateus, 2014a; Malafaia et al., 2015, 2017a, 2020). A diverse assemblage of small theropods attributed to different coelurosaurian clades has also been described, but these are mostly represented by isolated remains (e.g., Zinke, 1998; Rauhut, 2000, 2003; Hendrickx and Mateus, 2014b; Malafaia et al., 2017b). The Andrés fossil site (Pombal, Portugal) is one of the most relevant localities for studying Late Jurassic continental vertebrates from the Lusitanian Basin. This site has yielded an unusually abundant and diverse vertebrate assemblage, including ginglymodian fishes, abundant material of at least one lepidosaur taxon, neosuchian crocodylomorphs, indeterminate pterosaurs and different dinosaur groups (Pérez-Moreno et al., 1999; Ortega et al., 2006; Malafaia et al., 2010, 2024a). Theropod dinosaurs are well represented at Andrés, with abundant cranial and postcranial remains attributed to Allosaurus, alongside a relatively large collection of isolated teeth, some previously attributed to indeterminate dromaeosaurids (Pérez-Moreno et al., 1999; Malafaia et al., 2010, 2025). The diversity of vertebrate fossils at Andrés has been compared to that of the Upper Jurassic lignite levels of the Guimarota mine (Leiria, Portugal). Guimarota is primarily known for its rich and diverse record of early mammals (e.g., Henkel and Krebs, 1977; Martin, 2013; Martin and Schultz, 2023). However, a highly diverse fauna of other vertebrates has also been described (e.g., Bräm, 1973; Buscalioni et al., 1996; Krebs and Schwarz, 2000; Kriwet, 2000; Weichmann, 2000; Wiechmann and Gloy, 2000; Rauhut, 2001; Schwarz and Fechner, 2004; Caldwell et al., 2015; Mocho et al., 2017). Theropods are the most abundant dinosaurs in Guimarota, mostly represented by isolated teeth tentatively assigned to Compsognathus, Dromaeosaurus, indeterminate velociraptorines, troodontids, Richardoestesia, Paronychodon, tyrannosauroids, allosaurids, and Ceratosaurus (e.g., Zinke and Rauhut, 1994; Zinke, 1998; Rauhut, 2000). Some isolated teeth were firstly referred to cf. Archaeopteryx (Weigert, 1995), but were later considered to be distinct from this taxon (Rauhut et al., 2018a) and an attribution to a yet undescribed non-avian theropod was proposed (Louchart and Pouech, 2017). More complete theropod specimens are also known from Guimarota, such as a hatchling Allosaurus maxilla (Rauhut and Fechner, 2005), and pelvic elements, including the holotype of the tyrannosauroid Aviatyrannis jurassica (Rauhut, 2003).
The use of morphometric and cladistic analyses to support the identification of isolated theropod teeth has been extensively explored over the last two decades. Morphometric tools for studying large samples of theropod teeth were first approached by Farlow et al. (1991) and later developed by Smith (2005) and Smith et al. (2005). These authors compiled a large dataset of morphometric variables based on crown and denticle measurements across different theropod taxa. This dataset has since been expanded in subsequent studies, both in terms of the number of specimens and the range of dental-based variables (e.g., Smith, 2007; Larsson and Currie, 2013; Hendrickx et al., 2015a, 2020a; Gerke and Wings, 2016; Isasmendi et al., 2024). However, the effectiveness of these methods is sometimes limited by significant morphospace overlap among different taxa, which reduces their utility for identifying isolated theropod teeth (Hendrickx et al., 2020a, 2023). To improve accuracy, a combination of morphometric methods and cladistic analyses based on dentition-based character datasets (e.g., Hendrickx et al., 2015b) has been widely employed to support the taxonomic identification of isolated theropod teeth (e.g., Serrano-Martínez et al., 2016; Malafaia et al., 2017b; Alonso et al., 2018; Young et al., 2019; Delcourt et al., 2020; Hendrickx et al., 2020b; Isasmendi et al., 2024). More recently, machine learning tools applied to morphometric datasets have also been used, producing promising results that may significantly enhance the classification of these fossils (e.g., Wills et al., 2021, 2023; Hendrickx et al., 2023; Marques et al., 2025).
Here a sample of twenty-one isolated theropod teeth collected from the Andrés site is described to assess the diversity at this Upper Jurassic locality. Analyses of the identified tooth morphotypes, based on different methodologies (cladistic and discriminant analyses as well as machine learning tools), are presented to support their taxonomic assignment. Also, the assemblage of tooth morphotypes from Andrés is compared with other correlative European fossil records to assess the distribution of Late Jurassic theropod faunas in this region.
GEOLOGICAL SETTING
 The Andrés fossil site is located southeast of the town of Pombal, at the northern end of the Central Sector of the Lusitanian Basin (Figure 1). The sedimentary levels consist of massive, fine-grained, micaceous sandstones with parallel or cross-lamination, abundant plant debris, and some thin lenticular levels of red and grey claystone with abundant freshwater bivalves and some gastropods (Malafaia et al., 2010, 2025). This sedimentary sequence is interpreted as belonging to the Bombarral Formation (sensu Azerêdo et al., 2010), which is a diachronic unit but that has mostly been considered Tithonian in age (Manuppella et al., 1974, 1978; Fürsich et al., 2021).
The Andrés fossil site is located southeast of the town of Pombal, at the northern end of the Central Sector of the Lusitanian Basin (Figure 1). The sedimentary levels consist of massive, fine-grained, micaceous sandstones with parallel or cross-lamination, abundant plant debris, and some thin lenticular levels of red and grey claystone with abundant freshwater bivalves and some gastropods (Malafaia et al., 2010, 2025). This sedimentary sequence is interpreted as belonging to the Bombarral Formation (sensu Azerêdo et al., 2010), which is a diachronic unit but that has mostly been considered Tithonian in age (Manuppella et al., 1974, 1978; Fürsich et al., 2021).
MATERIAL AND METHODS
Material
The studied material consists of twenty-one isolated theropod teeth collected during different fieldwork campaigns conducted between 1988 and 2010 at the Andrés fossil site. All specimens are housed in the Museu Nacional de História Natural e da Ciência, Universidade de Lisboa (Lisboa, Portugal), and are inventoried as MNHN/UL.AND# (MNHN/UL, Museu Nacional de História Natural of the Universidade de Lisboa, AND, Andrés, and #, number of the element).
Methods
Comparative methodology and terminology. The teeth were described following anatomical, positional, directional, and morphometric nomenclatures proposed by Smith and Dodson (2003), Smith et al. (2005), and Hendrickx et al. (2015a). Each tooth was photographed in labial, lingual, mesial, distal, apical, and basal views using a Canon EOS 6D Mark II camera, with a macro-EF 100 mm lens. For smaller specimens, a Leica Z6 macroscope equipped with an integrated Leica DFC450 camera was used. The morphology of the denticles and enamel ornamentation were also observed and photographed using the Leica Z6 with the integrated Leica DFC450 camera.
Discriminant Function Analysis. The twenty-one theropod teeth from Andrés were added to the dataset of 1334 teeth compiled by Hendrickx et al. (2023). Twenty-eight variables were taken for each tooth (see Appendix 2). For the morphometric analyses, we used a final dataset comprising twelve variables (CBL, CBW, CH, AL, CMA, MCL, MCW, MSL, MDL, DDL, LAF, and LIF; morphometric abbreviations detailed in Appendix 3). Because both mesial and distal crown angles were measured, we used the abbreviation CMA (equivalent to CA in Hendrickx et al. 2020a) and CDA, respectively. To reduce potential noise in the results, we included only Late Jurassic and Cretaceous taxa from the original dataset. Additionally, in each analysis, we excluded all the groups with fewer cases than variables (i.e., fewer than 12 cases) to enhance the robustness of the results (e.g. Hair et al. 2010; Zavorka and Perrett, 2014). A series of discriminant analyses were conducted using Past v4 software (Hammer et al., 2001), following the methodology outlined in previous works (Hendrickx et al. 2020a). Following an initial set of analyses performed following this methodology, some taxa that were clearly unrelated to the studied specimens and well separated from all other morphospaces (Compsognathus and Buitreraptor in the genus level dataset, and Compsognathidae in the clade-level dataset) were also excluded to improve visualization. Due to a high number of missing values across several variables in some groups, an additional set of LDAs were performed using Python (version 3.8.16), and the package scikit-learn (Pedregosa et al., 2011). In this case, all variables with more than 20% missing data (i.e., AL, CMA, MCL, MCW, MSL, and MDL) were excluded, and all specimens with missing values, were removed, following the approach used by Marques et al. (2025).
To compare the assemblage of isolated teeth from Andrés with other tooth morphotypes described in correlative European levels, a dataset of morphometric variables was compiled from specimens found in different Upper Jurassic localities in Portugal, Spain, and Germany, based on previously published data (Ruiz-Omeñaca et al., 2008; Gascó et al., 2012; Malafaia et al., 2017b). Because several measurements used in the previous analyses are unknown for some of these specimens, a reduced principal component analysis (PCA) and t-distributed Stochastic Neighbor Embedding (t-SNE), following the methodology used by Wills et al. (2023) were performed in Python using the packages scikit-learn and open TSNE, respectively. Following Marques et al. (2025), variables with more than 20% of missing data and the specimens with missing values were removed. The final dataset for these analyses included the following variables: CBL, CBW, CH, and AL (see Appendix 1 and Appendix 4).
Cladistic analysis. The studied teeth were grouped in ten morphotypes, based on morphological and morphometric similarities and were scored as different taxonomic units. One morphotype (morphotype 2) was scored as mesial teeth, two morphotypes (morphotype 6 and 7) were scored as both mesial and lateral teeth due to uncertain preliminary identification, and the other seven morphotypes were scored as lateral teeth. The scores of each morphotype were included in the dentition-based database of Hendrickx et al. (2020a) (see Appendix 5). A cladistic analysis was then conducted for each morphotype using this database and following the search strategy outlined by Hendrickx et al. (2020a). See Appendix 3 for further details on the methodology used for the cladistic analyses.
Random Forest (RF) classifier. The isolated teeth from Andrés were analysed based on machine learning tools, using different RF models previously trained on the dataset of morphometric variables of theropod teeth compiled by Hendrickx et al. (2023). The trained models were made publicly available by Marques et al. (2025). A Quantile Transformer standardization was applied to approximate a normal distribution for the higher-level classification, while a standard scaler (mean = 0 and standard deviation =1) was used for the genus-level classification, both following the preprocessing methods used for the trained models. These analyses were conducted in Python (version 3.8.16) using the “scikit-learn” package (Pedregosa et al., 2011). Following the methodology described by Marques et al. (2025), seven morphometric variables were used (CBL, CBW, CH, CBR, CHR, DC, and DDL). The inclusion of both raw variables (CBW, CBL, CH) and their ratio-derived counterparts (CBR and CHR) in the analysis was chosen, as most machine learning methods, such as random forests, are less sensitive to multicollinearity, prioritizing variable splits over linear dependencies. By including both types of variables, we increase the likelihood of the models identifying subtle yet important patterns in the dataset that might be missed if either ratios or the raw variables were excluded. Specimens with missing data were excluded, resulting in a subset of fourteen specimens from the Andrés sample suitable for analysis. Two classification models were applied to predict the taxonomic interpretation of the studied teeth: one based on a dataset organized by genus, and another based on higher taxonomic groups. For further details on the methodology used in the RF analyses, see Appendix 3 and Marques et al. (2025).
SYSTEMATIC PALEONTOLOGY
DINOSAURIA Owen, 1842
THEROPODA Marsh, 1881
TETANURAE Gauthier, 1986
ALLOSAUROIDEA Currie and Zhao, 1994
ALLOSAURIDAE Marsh, 1878
ALLOSAURUS Marsh, 1877
Remarks. Three morphotypes (1, 2, and 3) are referred to Allosaurus.
 Morphotype 1. This morphotype is represented by five specimens (MNHN/UL.AND.23, MNHN/UL.AND.27, MNHN/UL.AND.30. MNHN/UL.AND.31, and MNHN/UL.AND.45; Figure 2) that consist of generally well-preserved tooth crowns, with three of them also preserving fragments of the root. The crowns are moderately large, slightly recurved distally, moderately elongated (CHR mean = 2.18), and weakly compressed labiolingually (CBR between 0.67 and 1.14; mean = 0.95; see Appendix 2). They have salinon-shaped basal cross-sections due to the presence of vertical concavities on the lingual surface adjacent to the mesial and distal carinae (Figure 2M). The lingual surface is slightly concave in distal or mesial view, while the labial surface is strongly convex. In lateral view, the crowns are triangular, with a mostly straight to slightly concave distal margin and a convex mesial margin. The apex is centrally positioned and does not extend beyond the level of the distal carina. The root is larger than the crown (at least 1.2 times the crown height), subcircular in cross-section, with mostly parallel mesial and distal margins but seem slightly tapered distally (Figure 2A-D). In MNHN/UL.AND.30 the root has a deep and well-delimited longitudinal depression centrally positioned that corresponds to a resorption pit (Figure 2A-D).
Morphotype 1. This morphotype is represented by five specimens (MNHN/UL.AND.23, MNHN/UL.AND.27, MNHN/UL.AND.30. MNHN/UL.AND.31, and MNHN/UL.AND.45; Figure 2) that consist of generally well-preserved tooth crowns, with three of them also preserving fragments of the root. The crowns are moderately large, slightly recurved distally, moderately elongated (CHR mean = 2.18), and weakly compressed labiolingually (CBR between 0.67 and 1.14; mean = 0.95; see Appendix 2). They have salinon-shaped basal cross-sections due to the presence of vertical concavities on the lingual surface adjacent to the mesial and distal carinae (Figure 2M). The lingual surface is slightly concave in distal or mesial view, while the labial surface is strongly convex. In lateral view, the crowns are triangular, with a mostly straight to slightly concave distal margin and a convex mesial margin. The apex is centrally positioned and does not extend beyond the level of the distal carina. The root is larger than the crown (at least 1.2 times the crown height), subcircular in cross-section, with mostly parallel mesial and distal margins but seem slightly tapered distally (Figure 2A-D). In MNHN/UL.AND.30 the root has a deep and well-delimited longitudinal depression centrally positioned that corresponds to a resorption pit (Figure 2A-D).
Both the mesial and distal carinae are serrated and extend to the cervix. The serrated carinae also extend to the apex of the crown, forming a continuous serrated margin. The distal carina is centrally positioned in the distal margin, whereas the mesial carina is located on the mesial surface in the apical end of the crown but strongly twists onto the lingual surface toward the base (Figure 2F). On average, there are 13 denticles per 5 mm in the mid-section of both carinae (DSDI mean = 0.99; see Appendix 2). The mesial denticles are subquadrangular, with slightly convex external margins, and become smaller toward the crown base (Figure 2O-Q). The distal denticles are mesiodistally rectangular, also with convex external margins, and become slightly smaller basally as well (Figure 2R-S). The denticles are separated by relatively wide interdenticular spaces and are oriented perpendicularly to the carina. Some specimens show subtle transverse undulations and poorly developed interdenticular sulci between the distal denticles, particularly in the middle and basal sections of the crown. The enamel has a braided texture that is well-visible with light, particularly on the lingual surface (Figure 2T).
 Morphotype 2. This morphotype is represented by seven specimens (MNHN/UL.AND.24, MNHN/UL.AND.25, MNHN/UL.AND.28, MNHN/UL.AND.29. MNHN/UL.AND.32a and MNHN/UL.AND.32b, and MNHN/UL.AND.33; Figure 3), consisting mostly of well-preserved tooth crowns, with some specimens also preserving root fragments. The crowns are medium sized, relatively short, strongly compressed labiolingually (CBR between 0.42 and 0.63; mean = 0.53), and slightly recurved distally (see Appendix 2). The distal margin is concave, whereas the mesial one is strongly convex, with the apex positioned distally to the most basal level of the distal carina. The lingual surface is mostly flat or slightly convex, while the labial surface is convex. Basal cross-sections are elliptical or lenticular in shape (Figure 3E and K). The mesial and distal carinae are serrated and extend to the basal section of the crown. The distal carina reaches the cervix, but the denticulated mesial carina ends well above it. The mesial carina is mostly centrally positioned in the mesial margin or slightly twists lingually toward the crown base, whereas the distal carina is markedly displaced labially to the base. The number of denticles in the central section ranges between 18 to 19 denticles and 11 to 17 denticles per 5 mm in the mesial and distal carinae, respectively (DSDI mean = 1.15; see Appendix 2). The interdenticular space is relatively wide on both carinae. The mesial denticles are apicobasally rectangular with rounded to mostly flat distal margins, whereas the distal denticles are mesiodistally rectangular with somewhat asymmetrically convex distal margins. Subtle transverse undulations are visible, particularly on the labial surface. Poorly developed interdenticular sulci are present on both mesial and distal carinae, though they are more pronounced adjacent to the denticles of the distal carina. The enamel shows a texture consisting of a series of thin and irregular crenulations, which are only visible with light.
Morphotype 2. This morphotype is represented by seven specimens (MNHN/UL.AND.24, MNHN/UL.AND.25, MNHN/UL.AND.28, MNHN/UL.AND.29. MNHN/UL.AND.32a and MNHN/UL.AND.32b, and MNHN/UL.AND.33; Figure 3), consisting mostly of well-preserved tooth crowns, with some specimens also preserving root fragments. The crowns are medium sized, relatively short, strongly compressed labiolingually (CBR between 0.42 and 0.63; mean = 0.53), and slightly recurved distally (see Appendix 2). The distal margin is concave, whereas the mesial one is strongly convex, with the apex positioned distally to the most basal level of the distal carina. The lingual surface is mostly flat or slightly convex, while the labial surface is convex. Basal cross-sections are elliptical or lenticular in shape (Figure 3E and K). The mesial and distal carinae are serrated and extend to the basal section of the crown. The distal carina reaches the cervix, but the denticulated mesial carina ends well above it. The mesial carina is mostly centrally positioned in the mesial margin or slightly twists lingually toward the crown base, whereas the distal carina is markedly displaced labially to the base. The number of denticles in the central section ranges between 18 to 19 denticles and 11 to 17 denticles per 5 mm in the mesial and distal carinae, respectively (DSDI mean = 1.15; see Appendix 2). The interdenticular space is relatively wide on both carinae. The mesial denticles are apicobasally rectangular with rounded to mostly flat distal margins, whereas the distal denticles are mesiodistally rectangular with somewhat asymmetrically convex distal margins. Subtle transverse undulations are visible, particularly on the labial surface. Poorly developed interdenticular sulci are present on both mesial and distal carinae, though they are more pronounced adjacent to the denticles of the distal carina. The enamel shows a texture consisting of a series of thin and irregular crenulations, which are only visible with light.
 Morphotype 3. This morphotype is represented by a single, almost complete and well-preserved tooth crown (MNHN/UL.AND.206; Figure 4A -J). It is relatively small, short, strongly compressed labiolingually (CBR = 0.47), and distally recurved (see Appendix 2). The distal margin is concave, while the mesial one is strongly convex, with the apex positioned distally to the level of the distal carina. The labial surface is mostly flat distally and slightly convex mesially, whereas the lingual surface is strongly convex. The basal cross-section of the crown is elliptical shaped (Figure 4F). Both mesial and distal carinae are serrated. The distal carina extends to the cervix, while the mesial one is restricted to the apical half of the crown. The mesial carina is mostly centrally positioned in the mesial margin but slightly twists onto the lingual surface toward the crown base. On the other hand, the distal carina is markedly displaced labially, resulting in an asymmetrical crown in distal view. There are 18 denticles per 5 mm in the central section of the distal carina. The interdenticular space is relatively wide on both carinae. The mesial denticles in the apical part of the crown are apicobasally rectangular with slightly convex to almost flat external margins, whereas the distal denticles are mesiodistally rectangular and have somewhat asymmetrically convex external margins with the apex pointing apically (Figure 4G-J). A few thin transverse undulations are visible in the basal section of the labial surface. Poorly developed interdenticular sulci are present on both mesial and distal carinae but they are more marked adjacent to the denticles in the central section of the distal carina. The enamel shows a texture consisting of a series of thin and irregular crenulations only visible with light.
Morphotype 3. This morphotype is represented by a single, almost complete and well-preserved tooth crown (MNHN/UL.AND.206; Figure 4A -J). It is relatively small, short, strongly compressed labiolingually (CBR = 0.47), and distally recurved (see Appendix 2). The distal margin is concave, while the mesial one is strongly convex, with the apex positioned distally to the level of the distal carina. The labial surface is mostly flat distally and slightly convex mesially, whereas the lingual surface is strongly convex. The basal cross-section of the crown is elliptical shaped (Figure 4F). Both mesial and distal carinae are serrated. The distal carina extends to the cervix, while the mesial one is restricted to the apical half of the crown. The mesial carina is mostly centrally positioned in the mesial margin but slightly twists onto the lingual surface toward the crown base. On the other hand, the distal carina is markedly displaced labially, resulting in an asymmetrical crown in distal view. There are 18 denticles per 5 mm in the central section of the distal carina. The interdenticular space is relatively wide on both carinae. The mesial denticles in the apical part of the crown are apicobasally rectangular with slightly convex to almost flat external margins, whereas the distal denticles are mesiodistally rectangular and have somewhat asymmetrically convex external margins with the apex pointing apically (Figure 4G-J). A few thin transverse undulations are visible in the basal section of the labial surface. Poorly developed interdenticular sulci are present on both mesial and distal carinae but they are more marked adjacent to the denticles in the central section of the distal carina. The enamel shows a texture consisting of a series of thin and irregular crenulations only visible with light.
COELUROSAURIA von Huene, 1914
COELUROSAURIA INDET.
Remarks. Two morphotypes (4 and 5) are attributed to indeterminate Coelurosauria.
Morphotype 4. This morphotype is represented by an almost complete tooth crown and a fragment of the root (MNHN/UL.AND.213; Figure 4K-Q). The apical part of the crown is worn due to a well-developed wear surface that also extends to the distal margin. The crown is relatively small and only weakly compressed labiolingually (CBR = 0.88), with a subcircular basal cross-section (Figure 4P). It is relatively elongated (CHR = 1.83) and mostly straight in lateral view, with a straight distal margin and a slightly convex mesial margin. Both labial and lingual surfaces are strongly convex. The mesial carina is absent in the preserved part of the crown, while the distal carina is serrated, slightly deflected lingually, and extends to the basal part of the crown, but ends well above the cervix (Figure 4M-N). The distal denticles are slightly longer mesiodistally than apicobasally, with symmetrically convex and rounded distal margins (Figure 4Q). There are 9.25 denticles per millimetre in the central section of the distal carina. The denticles are separated by relatively narrow spaces. Interdenticular sulci are not visible or are very short and poorly developed. Marginal and transverse undulations are also absent in the preserved part of the crown. The enamel has a braided and oriented texture that is barely visible with light.
Morphotype 5. This morphotype is represented by a poorly preserved fragment of a tooth crown (MNHN/UL.AND.209; Figure 4R-W). The crown is markedly recurved distally, with a strongly concave mesial margin and a convex distal margin. It is relatively high but mesiodistally short, resulting in an overall elongated profile (CHR = 1.81; see Appendix 2). The crown is broken in several parts, missing the apical part and the enamel in different sections of the lingual surface. The basal section is oval-shaped with low labiolingual compression (CBR = 0.62). Both the mesial and distal carinae are denticulated. The mesial carina is straight and centrally positioned on the mesial surface, ending around mid-height of the crown (Figure 4U). In contrast, the distal carina extends along the entire preserved length of the distal margin and is strongly deflected lingually, resulting in an asymmetrical appearance in distal view (Figure 4T). The lingual surface is mostly flat, while the labial surface is strongly convex. There are 11 denticles per one millimetre in the central section of the distal carina. These denticles are apicobasally rectangular in the apical part of the crown and subquadrangular in the central section, with symmetrically convex distal margins (Figure 4V). Denticle size decreases towards the base of the crown, with the most basal ones being almost imperceptible crenulations. The mesial denticles are very small and cannot be individually distinguished or measured in the preserved fragment. Denticles are separated by very narrow spaces and no interdenticular sulci are observed in the preserved fragment of the crown. The enamel has an oriented braided texture that is clearly visible with light, but lacks marginal or transverse undulations.
TYRANNOSAUROIDEA Osborn, 1906
TYRANNOSAUROIDEA INDET.
Remarks. One morphotype (6) is here interpreted as belonging to an early branching tyrannosauroid.
 Morphotype 6. This morphotype is represented by one specimen (MNHN/UL.AND.105; Figure 5A-G) that consists of a well-preserved crown and an almost complete root, missing only its distal end. The crown is relatively small, strongly compressed labiolingually (CBR = 0.41), elongated (CHR = 2.14), and distinctly recurved distally (see Appendix 2). The distal margin is strongly concave, while the mesial margin is convex, with the apex positioned well distal to the basal most level of the distal carina. The labial surface is mostly flat, whereas the lingual surface is slightly convex. The crown likely has a figure-eight-shaped basal cross-section due to the presence of shallow, centrally positioned longitudinal depression on both the lingual and labial surfaces of the root. However, these depressions do not significantly project into the basal section of the crown (Figure 5A-B). The preserved root fragment has parallel mesial and distal margins. The distal carina is denticulated, mostly straight in distal view, strongly deflected labially, and extends to the cervix (Figure 5C). The mesial carina is also serrated, centrally positioned, and straight in mesial view, but ends at about the mid-height of the crown (Figure 5D). Due to poor preservation, the morphology of the mesial denticles cannot be described. In the central section of the distal carina, there are 25.5 denticles per 5 mm. The denticles are separated by relatively wide interspaces, have symmetrically convex external margins, and project perpendicularly from the carina. They are mesiodistally rectangular in the basal and central section of the crown, becoming quadrangular toward the apex, and very reduce in size to the crown base (Figure 5F-G). Denticles on both the mesial and distal carinae extend to the cervix, forming an almost continuous serrated apical margin. Very faint transverse undulations are visible on both labial and lingual surfaces, which are also present in the preserved root fragment. Short, poorly developed interdenticular sulci are present adjacent to the denticles in the central and basal sections of the distal carina. The enamel shows a braided ornamentation that is barely visible with light.
Morphotype 6. This morphotype is represented by one specimen (MNHN/UL.AND.105; Figure 5A-G) that consists of a well-preserved crown and an almost complete root, missing only its distal end. The crown is relatively small, strongly compressed labiolingually (CBR = 0.41), elongated (CHR = 2.14), and distinctly recurved distally (see Appendix 2). The distal margin is strongly concave, while the mesial margin is convex, with the apex positioned well distal to the basal most level of the distal carina. The labial surface is mostly flat, whereas the lingual surface is slightly convex. The crown likely has a figure-eight-shaped basal cross-section due to the presence of shallow, centrally positioned longitudinal depression on both the lingual and labial surfaces of the root. However, these depressions do not significantly project into the basal section of the crown (Figure 5A-B). The preserved root fragment has parallel mesial and distal margins. The distal carina is denticulated, mostly straight in distal view, strongly deflected labially, and extends to the cervix (Figure 5C). The mesial carina is also serrated, centrally positioned, and straight in mesial view, but ends at about the mid-height of the crown (Figure 5D). Due to poor preservation, the morphology of the mesial denticles cannot be described. In the central section of the distal carina, there are 25.5 denticles per 5 mm. The denticles are separated by relatively wide interspaces, have symmetrically convex external margins, and project perpendicularly from the carina. They are mesiodistally rectangular in the basal and central section of the crown, becoming quadrangular toward the apex, and very reduce in size to the crown base (Figure 5F-G). Denticles on both the mesial and distal carinae extend to the cervix, forming an almost continuous serrated apical margin. Very faint transverse undulations are visible on both labial and lingual surfaces, which are also present in the preserved root fragment. Short, poorly developed interdenticular sulci are present adjacent to the denticles in the central and basal sections of the distal carina. The enamel shows a braided ornamentation that is barely visible with light.
MANIRAPTORA Gauthier, 1966
PARAVES Sereno, 1997
DROMAEOSAURIDAE Matthew and Brown, 1922
cf. DROMAEOSAURINAE Matthew and Brown, 1922
Remarks. Two morphotypes (7 and 8) are tentatively assigned to dromaeosaurine dromaeosaurids.
Morphotype 7. This morphotype is represented by a crown fragment lacking the base, with a large wear surface on the apical part that extends to the mesial margin and slightly onto the lingual surface (MNHN/UL.AND.26; Figure 5H-Q). The preserved fragment is relatively small and strongly compressed labiolingually (CBR = 0.57; see Appendix 2). The crown is slightly recurved, with a slightly concave distal margin, a convex mesial margin, and the apex positioned at the basal-most level of the preserved crown fragment. The basal section of the preserved crown is lanceolate-shaped, with a well-developed distal carina, a mostly flat lingual surface, and a slightly convex labial surface (Figure 5M). Both the mesial and distal carinae are denticulated. The distal denticles extend along the entire preserved height of the crown, while the mesial carina also extends along the preserved height of the crown but the denticles end slightly above the base. The distal carina is mostly straight but is somewhat displaced toward the labial side (Figure 5J), whereas the mesial carina is centrally positioned along the mesial surface (Figure 5K). A slightly concave surface is present adjacent to the distal carina on the labial side. The distal denticles are mostly subquadrangular, slightly larger apicobasally than mesiodistally, with rounded and symmetrically convex distal margins (Figure 5O-Q). The mesial denticles are also subquadrangular, with symmetrically convex margins (Figure 5N). The distal denticles are larger than the mesial denticles (DSDI = 1.57; see Appendix 2). Short interdenticular sulci are present between the distal denticles, particularly in the central and basal sections of the crown. Denticles on both mesial and distal carinae are separated by relatively narrow interspaces. The enamel has an irregular and non-oriented texture. No marginal or transverse undulations are visible on the preserved fragment of the crown.
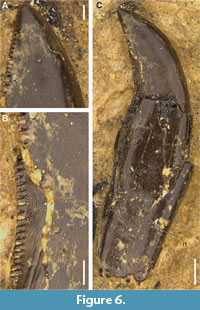 Morphotype 8. This morphotype is represented by an almost complete tooth with a well-preserved crown and root (MNHN/UL.AND.107; Figure 6). However, it still embedded in a sediment block, with only one surface (likely the labial one) exposed. Therefore, some measurements and morphological features cannot be confidently assessed at this moment. The crown is very small and seems to be strongly compressed labiolingually. It is markedly recurved distally, with a concave distal margin concave, a convex mesial margin, and the apex positioned beyond the level of the distal carina. Both the mesial and distal carinae are denticulated. The distal denticles extend along the entire height of the carina. Although the mesial carina is not fully visible, it appears to terminate around mid-crown height or extend slightly into the basal section, projecting onto the lingual surface near the crown base. The distal carina is also partially obscured by sediment but seems mostly straight, and either centrally positioned or slightly displaced labially. The distal denticles are mostly mesiodistally rectangular, with rounded and symmetrically convex distal margins (Figure 6B-C). The mesial denticles are very small and somewhat apicobasally rectangular, with slightly convex external margins. There are 9.5 denticles per millimetre in the visible apical part of the mesial carina, and between 9 and 7 denticles per millimetre in the apical and central/basal sections of the distal carina, respectively (DSDI = 1.36; see Appendix 2). Denticles on both carinae are separated by broad interspaces. Short, poorly developed interdenticular sulci are visible adjacent to the distal denticles, becoming slightly more pronounced in the basal section of the crown. Thin, barely visible transverse undulations are present along the preserved surface of the root, but not on the exposed surface of the crown (Figure 6A). The enamel has an irregular, non-oriented texture that is barely visible with light. The root is very elongated, with mostly parallel mesial and distal margins, and has a shallow, centrally positioned longitudinal concavity that extends along the entire height of the root to the crown base.
Morphotype 8. This morphotype is represented by an almost complete tooth with a well-preserved crown and root (MNHN/UL.AND.107; Figure 6). However, it still embedded in a sediment block, with only one surface (likely the labial one) exposed. Therefore, some measurements and morphological features cannot be confidently assessed at this moment. The crown is very small and seems to be strongly compressed labiolingually. It is markedly recurved distally, with a concave distal margin concave, a convex mesial margin, and the apex positioned beyond the level of the distal carina. Both the mesial and distal carinae are denticulated. The distal denticles extend along the entire height of the carina. Although the mesial carina is not fully visible, it appears to terminate around mid-crown height or extend slightly into the basal section, projecting onto the lingual surface near the crown base. The distal carina is also partially obscured by sediment but seems mostly straight, and either centrally positioned or slightly displaced labially. The distal denticles are mostly mesiodistally rectangular, with rounded and symmetrically convex distal margins (Figure 6B-C). The mesial denticles are very small and somewhat apicobasally rectangular, with slightly convex external margins. There are 9.5 denticles per millimetre in the visible apical part of the mesial carina, and between 9 and 7 denticles per millimetre in the apical and central/basal sections of the distal carina, respectively (DSDI = 1.36; see Appendix 2). Denticles on both carinae are separated by broad interspaces. Short, poorly developed interdenticular sulci are visible adjacent to the distal denticles, becoming slightly more pronounced in the basal section of the crown. Thin, barely visible transverse undulations are present along the preserved surface of the root, but not on the exposed surface of the crown (Figure 6A). The enamel has an irregular, non-oriented texture that is barely visible with light. The root is very elongated, with mostly parallel mesial and distal margins, and has a shallow, centrally positioned longitudinal concavity that extends along the entire height of the root to the crown base.
cf. VELOCIRAPTORINAE Barsbold, 1983
Remarks. Two morphotypes (9 and 10) are interpreted here as possibly belonging to velociraptorine dromaeosaurids.
 Morphotype 9. This morphotype is represented by two well preserved tooth crowns (MNHN/UL.AND.104 and MNHN/UL.AND.208; Figure 7A-N). The crowns are very small, relatively low, strongly compressed labiolingually (CBR mean = 0.51), and slightly recurved distally (see Appendix 2). The distal margin is concave, while the mesial margin is strongly convex, with the apex positioned distal to the most basal level of the distal carina. The lingual surface is mostly flat, whereas the labial surface is strongly convex. The crowns have an elliptical or lanceolate shaped basal cross-section (Figure 7E and J). The distal carinae are serrated and extend to the cervix. The mesial carina extends about half the height of the crown but lacks denticles, bearing only small crenulations at the apical end. The mesial carina is straight and centrally positioned along the mesial margin. In MNHN/UL.AND.208, the distal carina is mostly centrally positioned. In MNHN/UL.AND.104, it is centrally positioned in the apical part of the crown but becomes strongly displaced lingually toward the base, so the crown is asymmetrical in distal view (Figure 7C and H). There are between 32.5 and 35 denticles per 5 mm in the central section of the distal carina. The denticles are separated by relatively wide interspaces and have symmetrically convex external margins. In the basal and central sections of the crown, they are mesiodistally rectangular, becoming more quadrangular toward the apex (Figure 7K-N). Subtle and barely visible transverse undulations are present on the labial surface. Short and poorly developed interdenticular sulci occur in the central section of the distal carina. The enamel shows a faint braided texture that is only visible with light.
Morphotype 9. This morphotype is represented by two well preserved tooth crowns (MNHN/UL.AND.104 and MNHN/UL.AND.208; Figure 7A-N). The crowns are very small, relatively low, strongly compressed labiolingually (CBR mean = 0.51), and slightly recurved distally (see Appendix 2). The distal margin is concave, while the mesial margin is strongly convex, with the apex positioned distal to the most basal level of the distal carina. The lingual surface is mostly flat, whereas the labial surface is strongly convex. The crowns have an elliptical or lanceolate shaped basal cross-section (Figure 7E and J). The distal carinae are serrated and extend to the cervix. The mesial carina extends about half the height of the crown but lacks denticles, bearing only small crenulations at the apical end. The mesial carina is straight and centrally positioned along the mesial margin. In MNHN/UL.AND.208, the distal carina is mostly centrally positioned. In MNHN/UL.AND.104, it is centrally positioned in the apical part of the crown but becomes strongly displaced lingually toward the base, so the crown is asymmetrical in distal view (Figure 7C and H). There are between 32.5 and 35 denticles per 5 mm in the central section of the distal carina. The denticles are separated by relatively wide interspaces and have symmetrically convex external margins. In the basal and central sections of the crown, they are mesiodistally rectangular, becoming more quadrangular toward the apex (Figure 7K-N). Subtle and barely visible transverse undulations are present on the labial surface. Short and poorly developed interdenticular sulci occur in the central section of the distal carina. The enamel shows a faint braided texture that is only visible with light.
Morphotype 10. This morphotype is represented by an almost complete crown that has a well-developed wear surface affecting both the labial and lingual sides (MNHN/UL.AND.212; Figure 7O-S). The crown is very short apicobasally but relatively elongated mesiodistally (CHR = 1.09; see Appendix 2). It is slightly recurved distally and strongly compressed labiolingually (CBR = 0.53), with a lanceolate shaped basal section (Figure 7S). Both the labial and lingual surfaces are slightly convex, although the lingual surface is somewhat more flattened. A serrated distal carina is present, while the mesial denticles and carina are absent in the portion of the crown not affected by the wear surface. The distal carina extends to the cervix, is mostly straight in distal view, but slightly displaced lingually (Figure 7Q). There are 7 and 7.75 denticles per millimetre in the central and basal section of the distal carina, respectively. The distal denticles, especially the most basal ones, are mesiodistally rectangular with symmetrically convex external margins, and are oriented perpendicularly to the carina (Figure 7J). The denticles are separated by relatively narrow spaces at mid-crown, which become slightly wider in the apical and basal sections. The enamel has a braided texture that is barely visible with light. No marginal or transverse undulations are visible. Interdenticular sulci are not observable adjacent to the distal denticles.
RESULTS
Discriminant Function Analyses (DFA)
 The results obtained on the LDA performed in PAST, based on the genus- and clade-level datasets, correctly classified 74.59% and 62.01% of the cases, respectively. The eigenvalues for axis 1 and 2 were 77.51 and 83.82, respectively. In these analyses, two specimens of morphotype 1 (MNHN/UL.AND.23 and 30) were classified as Allosaurus and Allosauridae, while two others (MNHN/UL.AND.27 and 31) were classified as Australovenator and Neovenatoridae, and one (MNHN/UL.AND.45) as Gorgosaurus and Neovenatoridae (see Figure 8 and Table 1 for details on the results of these analyses). The classification for morphotype 2 was more ambiguous. Three specimens (MNHN/UL.AND.24, 28, and 29) were classified as Alioramus, two (MNHN/UL.AND.32a and 33) as Fukuiraptor, one (MNHN/UL.AND.25) as Majungasaurus, and another (MNHN/UL.AND.32b) as Australovenator, based on the genus-level dataset. Using the clade-level dataset, most specimens of this morphotype were classified as Neovenatoridae, except MNHN/UL.AND.25, which was classified as Abelisauridae, and MNHN/UL.AND.24, which was assigned to non-abelisaurid Ceratosauria (see Figure 8 and Table 1 for details on the results of these analyses). Morphotype 3 (MNHN/UL.AND.206) and morphotype 7 (MNHN/UL.AND.26) were classified as Deinonychus and as belonging to Abelisauridae and Noasauridae, respectively. Morphotype 4 (MNHN/UL.AND.213) was identified as Dromaeosaurus and
The results obtained on the LDA performed in PAST, based on the genus- and clade-level datasets, correctly classified 74.59% and 62.01% of the cases, respectively. The eigenvalues for axis 1 and 2 were 77.51 and 83.82, respectively. In these analyses, two specimens of morphotype 1 (MNHN/UL.AND.23 and 30) were classified as Allosaurus and Allosauridae, while two others (MNHN/UL.AND.27 and 31) were classified as Australovenator and Neovenatoridae, and one (MNHN/UL.AND.45) as Gorgosaurus and Neovenatoridae (see Figure 8 and Table 1 for details on the results of these analyses). The classification for morphotype 2 was more ambiguous. Three specimens (MNHN/UL.AND.24, 28, and 29) were classified as Alioramus, two (MNHN/UL.AND.32a and 33) as Fukuiraptor, one (MNHN/UL.AND.25) as Majungasaurus, and another (MNHN/UL.AND.32b) as Australovenator, based on the genus-level dataset. Using the clade-level dataset, most specimens of this morphotype were classified as Neovenatoridae, except MNHN/UL.AND.25, which was classified as Abelisauridae, and MNHN/UL.AND.24, which was assigned to non-abelisaurid Ceratosauria (see Figure 8 and Table 1 for details on the results of these analyses). Morphotype 3 (MNHN/UL.AND.206) and morphotype 7 (MNHN/UL.AND.26) were classified as Deinonychus and as belonging to Abelisauridae and Noasauridae, respectively. Morphotype 4 (MNHN/UL.AND.213) was identified as Dromaeosaurus and  Therizinosauria. Morphotypes 5 (MNHN/UL.AND.209), morphotype 9 (MNHN/UL.AND.104 and 208), and morphotype 10 (MNHN/UL.AND.212) were classified as Masiakasaurus and Therizinosauria, except morphotype 5, which was assigned to Noasauridae in the analysis based on the clade-level dataset. Morphotype 6 (MNHN/UL.AND.105) was identified as Raptorex and Dromaeosauridae, and morphotype 8 as Australovenator and Neovenatoridae. The LDA performed in Python recovered an overall RR of 73.7%, with eigenvalues for axes 1 and 2 of 74.7% and 19.2%, respectively, using the clade-level dataset and an RR of 64.9%, with eigenvalues for axes 1 and 2 of 81.5% and 13.9%, respectively, using the genus-level dataset. These analyses classified all morphotype 1 specimens with complete data (MNHN/UL.AND.23, 30, and 31) as Allosaurus and Allosauridae (see Figure 9 and Table 1 for details on the results of these analyses). In contrast, morphotype 2 specimens were classified as Alioramus (MNHN/UL.AND.24 and 29), Fukuiraptor (MNHN/UL.AND.33), and Majungasaurus (MNHN/UL.AND.25). Using the clade-level dataset, these specimens were assigned to Abelisauridae (MNHN/UL.AND.24, 25, and 29) and Metriacanthosauridae (MNHN/UL.AND.33). The specimen of morphotype 4 was classified as Zanabazar and Troodontidae, while morphotype 5 was identified as Falcarius and as an early-branching Coelurosauria. Specimens of morphotypes 6, 7, 9, and 10 were classified as Dromaeosauridae using the clade-level dataset and as Saurornitholestes (morphotypes 6 and 9), Deinonychus (morphotype 7), and Falcarius (morphotype 10) using the genus-level dataset.
Therizinosauria. Morphotypes 5 (MNHN/UL.AND.209), morphotype 9 (MNHN/UL.AND.104 and 208), and morphotype 10 (MNHN/UL.AND.212) were classified as Masiakasaurus and Therizinosauria, except morphotype 5, which was assigned to Noasauridae in the analysis based on the clade-level dataset. Morphotype 6 (MNHN/UL.AND.105) was identified as Raptorex and Dromaeosauridae, and morphotype 8 as Australovenator and Neovenatoridae. The LDA performed in Python recovered an overall RR of 73.7%, with eigenvalues for axes 1 and 2 of 74.7% and 19.2%, respectively, using the clade-level dataset and an RR of 64.9%, with eigenvalues for axes 1 and 2 of 81.5% and 13.9%, respectively, using the genus-level dataset. These analyses classified all morphotype 1 specimens with complete data (MNHN/UL.AND.23, 30, and 31) as Allosaurus and Allosauridae (see Figure 9 and Table 1 for details on the results of these analyses). In contrast, morphotype 2 specimens were classified as Alioramus (MNHN/UL.AND.24 and 29), Fukuiraptor (MNHN/UL.AND.33), and Majungasaurus (MNHN/UL.AND.25). Using the clade-level dataset, these specimens were assigned to Abelisauridae (MNHN/UL.AND.24, 25, and 29) and Metriacanthosauridae (MNHN/UL.AND.33). The specimen of morphotype 4 was classified as Zanabazar and Troodontidae, while morphotype 5 was identified as Falcarius and as an early-branching Coelurosauria. Specimens of morphotypes 6, 7, 9, and 10 were classified as Dromaeosauridae using the clade-level dataset and as Saurornitholestes (morphotypes 6 and 9), Deinonychus (morphotype 7), and Falcarius (morphotype 10) using the genus-level dataset.
Cladistic analysis
 The cladistic analysis, using a constrained topology with all terminal taxa scored in the original dataset of Hendrickx et al. (2020a,b) along with morphotypes 1 and 2, yielded one most parsimonious tree (MPT) and placed the specimens from Andrés within Allosauroidea (Figure 10A). Morphotype 1 is allied with the metriacanthosaurids Sinraptor and Erectopus, based on the straight distal margin of the crown in lateral view and the presence of short interdenticular sulci. On the other hand, morphotype 2 is recovered as the sister taxon of Allosaurus, based on the shared presence of a mesial carina strongly twisting onto the mesiolingual surface, a strongly labially deflected distal carina in distal view, and presence of transverse undulations below the cervix. The analysis preformed for morphotype 3 yielded five MPT’s, and the strict consensus tree places MNHN/UL.AND.206 within Tyrannosauroidea, in a large
The cladistic analysis, using a constrained topology with all terminal taxa scored in the original dataset of Hendrickx et al. (2020a,b) along with morphotypes 1 and 2, yielded one most parsimonious tree (MPT) and placed the specimens from Andrés within Allosauroidea (Figure 10A). Morphotype 1 is allied with the metriacanthosaurids Sinraptor and Erectopus, based on the straight distal margin of the crown in lateral view and the presence of short interdenticular sulci. On the other hand, morphotype 2 is recovered as the sister taxon of Allosaurus, based on the shared presence of a mesial carina strongly twisting onto the mesiolingual surface, a strongly labially deflected distal carina in distal view, and presence of transverse undulations below the cervix. The analysis preformed for morphotype 3 yielded five MPT’s, and the strict consensus tree places MNHN/UL.AND.206 within Tyrannosauroidea, in a large 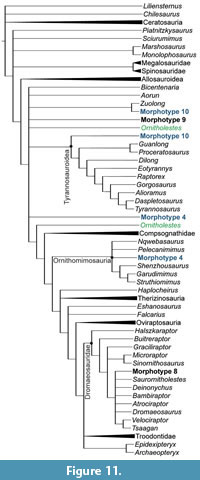 polytomy with Alioramus, Gorgosaurus, Raptorex, Eotyranus, Dilong, Guanlong, Proceratosaurus, and a group comprising Tyrannosaurus and Daspletosaurus (Figure 10B). For morphotype 4, analyses conducted with the specimen scored as a mesial tooth found three MPT’s. The consensus tree recovers this morphotype from Andrés within Coelurosauria, in a polytomy with Ornitholestes, Zuolong, Aorun, Bicentenaria, Tyrannosauroidea, and Neocoelurosauris (Figure 11). When MNHN/UL.AND.213 is scored as a lateral tooth, the analysis yields two MPT’s, and the consensus tree places it within Ornithomimosauria, in a polytomy with Pelecanimimus and Nqwebasaurus (Figure 11).
polytomy with Alioramus, Gorgosaurus, Raptorex, Eotyranus, Dilong, Guanlong, Proceratosaurus, and a group comprising Tyrannosaurus and Daspletosaurus (Figure 10B). For morphotype 4, analyses conducted with the specimen scored as a mesial tooth found three MPT’s. The consensus tree recovers this morphotype from Andrés within Coelurosauria, in a polytomy with Ornitholestes, Zuolong, Aorun, Bicentenaria, Tyrannosauroidea, and Neocoelurosauris (Figure 11). When MNHN/UL.AND.213 is scored as a lateral tooth, the analysis yields two MPT’s, and the consensus tree places it within Ornithomimosauria, in a polytomy with Pelecanimimus and Nqwebasaurus (Figure 11).
The analysis performed for morphotype 6 yielded two MPT’s. In one of these trees, MNHN/UL.AND.105 is recovered within Tyrannosauroidea, allied with Eotyrannus (Figure 10A). In the other tree, it is placed within Megalosauroidea as the sister taxon of Piatnitzkysaurus. The results for morphotype 5 are ambiguous. When the specimen is codified as a mesial tooth, the analysis yields two MPT’s, and the consensus tree places it at the base of Theropoda, in a polytomy with Dracovenator, Coelophysis, and Dilophosaurus (Figure 10C). When MNHN/UL.AND.209 is scored as a lateral tooth, the analysis finds eight MPT’s, and the consensus tree places it in a large polytomy with several early-branching theropods, including Dilophosaurus, Coelophysis, and Liliensternus, as well as more deeply nested forms such as Zuolong, Aorun, and Bicentenaria. However, in six of the MPT’s, this morphotype from Andrés is recovered at the base of Coelurosauria, and in the other two trees, as a member of Tyrannosauroidea allied with non-tyrannosauroid Coelurosauria.
For morphotype 7, the analysis yields five MPT’s, and the consensus tree recovers MNHN/UL.AND.26 as allied with abelisauroids, in a polytomy with the noasaurids Masiakasaurus, Noasaurus, and Limusaurus (Figure 10C). The analysis of morphotype 8 recovers four MPT’s, and the consensus tree places MNHN/UL.AND.107 within Dromaeosauridae, in a polytomy with Saurornitholestes, Deinonychus, Bambiraptor, Atrociraptor, Dromaeosaurus, and a group composed of Velociraptor and Tsaagan (Figure 11). The analysis performed for morphotype 9 finds five MPT’s, and the consensus tree places it within Coelurosauria, in a polytomy with Zuolong, Aorun, Bicentenaria, Tyrannosauroidea and Neocoelurosauria (Figure 11). Finally, for morphotype 10, the analysis yields two MPT’s. In one tree, the specimen from Andrés is placed at the base of Coelurosauria, as the sister taxon of Zuolong and in the other, it is recovered at the base of Tyrannosauroidea (Figure 11). See Appendix 3 for more details on the results of the cladistic analyses.
Random Forest (RF) classifier
The results of the RF classification for the studied specimens are summarized in Table 1 and Table 2. Specimens of morphotype 1 are consistently classified, with high probabilities, as Allosaurus based on the genus-level dataset. However, when using the clade-level dataset, only one specimen (MNHN/UL.AND.23) is assigned to Allosauridae, whereas the other two specimens (MNHN/UL.AND.30 and 31) are classified as Tyrannosauridae. In contrast, the predicted classification for morphotype 2 is more ambiguous, similar to the results from the LDA analysis. Two specimens (MNHN/UL.AND.24 and 29) are identified as Alioramus and Tyrannosauridae, one specimen (MNHN/UL.AND.33) as Fukuiraptor and Neovenatoridae, and another specimen (MNHN/UL.AND.25) as Majungasaurus and Abelisauridae. Morphotype 4 (MNHN/UL.AND.213) is classified as Dilong and Therizinosauria, while morphotype 5 (MNHN/UL.AND.209) is classified as Coelophysis and Therizinosauria. Morphotypes 6 (MNHN/UL.AND.105), 7 (MNHN/UL.AND.26), 9 (MNHN/UL.AND.104 and 208) are all classified as Dromaeosauridae based on the clade-level dataset, and at the genus level, they are identified as Saurornitholestes, Atrociraptor, Richardoestesia, and Eoraptor, respectively. Finally. Morphotype 10 (MNHN/UL.AND.212) is assigned to Richardoestesia and to a non-averostran Neotheropoda.
DISCUSSION
Taxonomic identification
Allosaurus (morphotype 1, 2, and 3)
Morphotype 1. These specimens are interpreted as mesial teeth based on the weak labiolingual compression of the crowns and the strong twist of the mesial carina onto the lingual surface. Both the discriminant and random forest analyses classify most of these specimens (except MNHN/UL.AND.27 and 45) as Allosaurus and Allosauridae. On the other hand, the cladistic analysis places this morphotype from Andrés within Metriacanthosauridae. These teeth share several features with mesial teeth of Allosaurus and metriacanthosaurids, including denticulated mesial and distal carinae extending to the root and well-beneath the cervix, a lingually positioned longitudinal groove adjacent to the mesial carina, a salinon-shaped basal cross-section of the crown, a strongly labially deflected distal carina, and a braided enamel texture (Hendrickx et al., 2019, 2020b). While some of these features are also convergently present in abelisaurids (e.g., Majungasaurus: Smith, 2007), the combination of dental characters described above is exclusive for Allosaurus and Sinraptor and these two taxa have almost indistinguishable mesial tooth morphology (Hendrickx et al., 2020b). Based on this combination of features, the results of the different performed analyses, and the currently known fossil record in Andrés, morphotype 1 can be confidently interpreted as mesial teeth of Allosaurus.
Morphotype 2. These specimens are interpreted as lateral teeth based on strong labiolingual compression of the crowns and the position of the distal and mesial carinae in the distal and mesial margins, respectively. The results of the discriminant and random forest analyses are ambiguous, with the teeth classified as belonging to different taxa. On the other hand, the cladistic analysis provides more robust results, placing this morphotype as the sister taxon of Allosaurus. The morphology of these teeth is similar to some isolated theropod teeth from the Upper Jurassic of Germany interpreted as belonging to Allosauroidea and Allosaurus (morphotype I and K of Gerke and Wings, 2016). The specimens from Andrés and the German morphotypes share similar crown size, compression ratios, and identical denticle densities in the mesial and distal carinae. They also share the strongly labially deflected distal carina in some teeth and a mesial carina that extends to the basal section of the crown (Gerke and Wings, 2006). Morphotype 2 also resembles an isolated tooth from the Upper Jurassic of Asturias (Ruiz-Omeñaca et al. 2008), which was tentatively assigned to a carcharodontosaurid based on the presence of well-developed wrinkles adjacent to the mesial carina that is a feature initially considered diagnostic of Carcharodontosaurus (Sereno et al. 1996). However, as this feature has been also described in teeth of several other theropods, including in Allosaurus (Brusatte et al., 2007; Hendrickx et al., 2019), the attribution of isolated teeth to carcharodontosaurids based on this trait should be reevaluated. Another similar tooth morphotype was described from the Upper Jurassic of Teruel and attributed to an indeterminate allosaurid (Gascó et al., 2012). These teeth share with morphotype 2 from Andrés the strongly labially deflected distal carina, comparable denticle density, and identical denticle morphology. Morphotype 2 shares several features with Allosaurus and Sinraptor, including a mesial carina that extends to the basal section of the crown and twists strongly onto the lingual surface, a strongly labially deflected distal carina, and the presence of relatively well-developed transverse undulations that, in some specimens, extend below the cervix (Hendrickx et al., 2019, 2020a,b). This combination of features supports the identification of morphotype 2 as belonging to Allosaurus, consistent with the cladistic analysis results. The ambiguous results of the discriminant analyses may be partially explained by the generally smaller size of the specimens from Andrés compared to Allosaurus teeth in the original dataset.
Morphotype 3. This specimen is interpreted as a lateral tooth based on the strongly labiolingually compressed crown and the position of the distal and mesial carinae mostly in the distal and mesial margins, respectively. The discriminant analyses performed with PAST classifies this tooth as Deinonychus and as a non-megalosauran Megalosauroidea. The cladistic analysis places this specimen within Tyrannosauridae but found three autapomorphies for MNHN/UL.AND.206: strong labiolingual compression of the crown, weak baso-apical elongation, and a subrectangular shape of the mesial margin of rounded denticles on mesial carina in lateral view. The morphology of this specimen is similar to teeth of morphotype 2, sharing several features such as a strongly labially deflected distal carina and presence of short, poorly developed interdenticular sulci adjacent to the distal carina. However, MNHN/UL.AND.206 has a much smaller crown and slightly higher number of denticles in the central section of the distal carina compared to morphotype 2. It also differs from the teeth of this morphotype in that the denticles of the mesial carina terminate around the mid-height of the crown. This specimen from Andrés has a morphology similar to other isolated teeth from the Upper Jurassic of the Lusitanian Basin interpreted as belonging to juvenile individuals of Allosaurus (morphotype 11 of Malafaia et al., 2017b). They share comparable crown heights and a strongly labially deflected distal carina. Both morphotypes have a higher denticle density on the mesial and distal carinae relative to other specimens attributed to Allosaurus. Despite some differences, the overall combination of features of morphotype 3 from Andrés is compatible with the morphology of lateral teeth of Allosaurus. Based on its similarities to morphotype 2, this specimen is here interpreted as possibly belonging to a juvenile individual of Allosaurus.
Coelurosauria indet. (morphotype 4)
Morphotype 4. MNHN/UL.AND.213 is interpreted as a mesial tooth crown based on its nearly circular basal cross-section. However, because some coelurosaurian theropods have weakly labiolingually compressed lateral teeth, it cannot be ruled out that this specimen may correspond to a lateral tooth. In the discriminant analyses based on the genus-level dataset, the specimen is classified as Dromaeosaurus and Zanabazar, while in the analysis using the clade-level dataset, it is identified as Therizinosauria and Troodontidae (Table 1). In the random forest analyses, it is assigned to Dilong and Therizinosauria. The cladistic analyses recovered it allied with non-tyrannosauroid Coelurosauria when scored as a mesial tooth, and within Ornithomimosauria when scored as a lateral tooth. This morphotype shares some similarities with a specimen from Guimarota tentatively identified as a mesial tooth of Dromaeosaurus (Zinke, 1998), including the rounded basal cross-section and mostly straight distal margin. However, the two differ in the extent of the denticulated distal carina, which reaches the cervix in the Guimarota tooth but ends well above it in MNHN/UL.AND.213. Also, the specimen from Andrés lacks a mesial carina in the preserved part of the crown (though it could have been present at the apical end, which was worn by a large wear facet) whereas the Guimarota specimen shows a mesial carina extending nearly to mid-crown height. A distal carina terminating well above the cervix is an unusual feature in theropod dentition, though it has been documented in the lateral teeth of some taxa such as Compsognathus, Juravenator, and Ornitholestes (Hendrickx et al., 2020a). Some dentary teeth of Dromaeosaurus also seem to have a similar short distal carina (Currie et al., 1990). This condition is also present in isolated teeth from the Lower Cretaceous of Siberia, interpreted as belonging to juvenile individuals of indeterminate dromaeosaurids or tyrannosauroids (Averianov et al., 2019). The specimen from Andrés shares several features with these Siberian teeth, including a similar crown height with rounded basal cross-section, the distal carina ending well above the cervix, absence of a mesial carina, and smooth enamel, but differs on the size of the denticles that are much smaller in MNHN/UL.AND.213. Morphotype 4 has an unusual combination of features not yet described in theropod dentition from the Upper Jurassic of the Lusitanian Basin. The results of both discriminant and cladistic analyses are also ambiguous, but all consistently relate the specimen to coelurosaurian theropods. Therefore, it is here assigned to an indeterminate Coelurosauria.
Neocoelurosauria indet. (morphotype 5)
Morphotype 5. MNHN/UL.AND.209 is interpreted as a mesial tooth crown based on its nearly circular basal cross-section. However, given that some coelurosaurian theropods have weakly labiolingually compressed lateral teeth, it cannot be ruled out that the specimen may instead represent a lateral tooth. In the genus-level dataset, the LDA classify this specimen as Masiakasaurus and Falcarius, whereas based on the clade-level dataset it is assigned to Noasauridae and early-branching Coelurosauria. The random forest analyses assign this specimen to Coelophysis and Therizinosauria as the most probable classification, and to Richardoestesia and Dromaeosauridae as the second most probable, with similar probabilities (Table 2), respectively.. The cladistic analysis yields uncertain results, placing the specimen in a polytomy at the base of Theropoda, possibly because the specimen is much incomplete. The morphology of MNHN/UL.AND.209 is somewhat similar to some teeth of Richardoestesia (or tentatively related to this taxon) from the Upper Cretaceous of North America (e.g., Currie et al., 1990; Frederickson et al., 2018; Larson and Currie, 2013) and Spain (e.g., Isasmendi et al., 2024), particularly in its strongly elongated crown and the high number of small denticles on the distal carina. However, teeth attributed to Richardoestesia generally have a nearly straight distal margin, whereas it is much concave in the specimen from Andrés. Morphotype 5 also shares several features with some neocoelurosaurian theropods (sensu Hendrickx et al., 2019), including a strongly labially deflected distal carina, a denticulated mesial carina terminating in the apical part of the crown, a subcircular basal cross-section, and a strongly distally recurved crown. Based on this combination of features and because it is poorly preserved, MNHN/UL.AND.209 is here tentatively assigned to an indeterminate neocoelurosaurian theropod.
Early-branching Tyrannosauroidea (morphotype 6)
Morphotype 6. This specimen is interpreted as a lateral tooth based on its strongly labiolingually compressed and markedly distally recurved crown. Both discriminant and random forest analyses primarily classify MNHN/UL.AND.105 as Saurornitholestes and Dromadeosauridae. The cladistic analysis places the specimen within Tyrannosauroidea in one of the MPTs and allied with the megalosauroid Piatnitzkysaurus in another. This specimen shares several features with teeth described for Marshosaurus and other piatnitzkysaurids, including a slender and elongated crown, a mesial carina terminating near mid-height of the crown, and a strongly labially deflected distal carina (Madsen, 1976; Hendrickx et al., 2019). The Andrés specimen is particularly similar to some isolated teeth tentatively related to Marshosaurus from other Late Jurassic localities in Portugal (morphotypes 7 and 8 of Malafaia et al., 2017b) and Germany (morphotype J of Gerke and Wings, 2016). However, MNHN/UL.AND.105 differs in having a smaller crown, a more pronounced distal curvature, and a higher denticle density on the distal carina compared to the specimens from Portugal and Germany, as well as to most known teeth of Marshosaurus and Piatnitzkysaurus (Gerke and Wings, 2016; Malafaia et al., 2017b; Hendrickx et al., 2020a).
The overall morphology of MNHN/UL.AND.105 also resembles the lateral teeth of the juvenile megalosauroid Sciurumimus, sharing the strongly recurved crowns, but in this taxon the mesial carina is unserrated (Rauhut et al., 2012). The combination of features seen in morphotype 6 is more similar to those of some early-branching tyrannosauroids, such as Proceratosaurus (Rauhut et al., 2010) and Eotyrannus (Hutt et al., 2001; Hendrickx et al., 2020a). The specimen from Andrés probably has an eight-shaped basal cross-section due to the presence of shallow, centrally positioned longitudinal depressions on both the labial and lingual surfaces of the root that extend into the base of the crown. This feature is present in most dromaeosaurids and troodontids and is considered as a synapomorphy for Pennaraptora (Hendrickx et al., 2019), but has also been described in some tyrannosauroids, including Proceratosaurus (Rauhut et al., 2010). Based on this combination of morphological features and the results of the cladistic analysis, morphotype 6 is here interpreted as possibly belonging to an early-branching tyrannosauroid.
Dromaeosauridae (cf. Dromaeosaurinae) (morphotypes 7 and 8)
Morphotype 7. MNHN/UL.AND.26 is interpreted as a fragment of a lateral tooth crown based on the strong labiolingual compression and the position of the mesial and distal carina mostly in the mesial and distal surfaces, respectively. Most discriminant and random forest analyses classify this specimen as belonging to a dromaeosaurid taxon (Deinonychus and Atrociraptor) or as Dromaeosauridae. The cladistic analysis recovers it within Noasauridae, in a polytomy with Limusaurus, Noasaurus, and Masiakasaurus. The specimen shares some morphological similarities with noasaurids, such as a crown height of less than 2 cm, a mesial carina extending to or just above the cervix, and a DSDI greater than 1.2. However, it lacks flutes and has a lanceolate basal cross-section, which contrasts with the salinon-shape present in the lateral teeth of some noasaurids, such as Masiakasaurus (Carrano et al., 2002; Hendrickx et al., 2019). Besides, the small crown size and relatively higher number of denticles on the mesial carina compared to the distal carina are features also shared with most dromaeosaurids (e.g., Hendrickx et al., 2019). Nevertheless, most dromaeosaurids, including microraptorines, some velociraptorines and dromaeosaurines have unserrated mesial carinae in some lateral teeth (e.g., Larson, 2008; Hendrickx et al., 2019; Wills et al., 2023). The extension of the mesial carina to the basal section of the crown in the specimen from Andrés is similar to the condition present in dromaeosaurines, whereas in velociraptorines, when present, the mesial carina ends around mid-height of the crown (e.g., Currie et al., 1990; Hendrickx et al., 2019). The shape of the denticles on the distal carina, which are mostly vertically rectangular with symmetrically convex external margins, also resembles those of dromaeosaurines more than to the apically hooked denticles typical of most velociraptorines (Currie et al., 1990; Currie and Varricchio, 2004; Larson, 2008; Hendrickx et al., 2019). Distal denticles slightly hooked apically have been also described in isolated teeth attributed to velociraptorine dromaeosaurids from Guimarota (Zinke, 1998). MNHN/UL.AND.26 also differs from the lateral teeth of Dromaeosaurus in the mostly centrally positioned mesial carina, whereas in this taxon it is characteristically strongly twisted onto the lingual surface (Currie et al., 1990; Fiorillo and Gangloff, 2000; Hendrickx et al., 2019). Besides, the distal carina on this specimen from Andrés seems almost straight and thus less distally recurved than is typical for the lateral dentition of dromaeosaurine dromaeosaurids (e.g., Currie et al., 1990; Hendrickx et al., 2019). However, this may be related to the incomplete preservation of the specimen, which lacks the basal section of the crown. Based on this combination of features, morphotype 7 is here assigned to an indeterminate dromaeosaurid, possibly related to dromaeosaurines, which is also supported by the results obtained on both the discriminant and randon forest analyses.
Morphotype 8. MNHN/UL.AND.107 is interpreted as a lateral tooth based on the strong labiolingual compression of the crown. Because it lacks information for some variables, only the discriminant analyses performed in PAST was able to classify this specimen, which assign it as Australovenator and Neovenatoridae. The cladistic analysis places it within a poorly resolved Dromaeosauridae group in a large polytomy with Saurornitholestes, Deinonychus, Bambiraptor, Atrociraptor, Dromaeosaurus, Velociraptor, and Tsaagan. This specimen has a combination of feature typical for lateral teeth of dromaeosaurids, including much smaller mesial denticle relative to the distal ones (DSDI > 1.2), a strongly concave distal margin, and a possible figure-of-eight (or bean-shaped) basal cross-section due to the presence of labial and lingual depressions on the tooth root extending to the crown base (Hendrickx et al., 2019). Within this clade, morphotype 8 differs from the lateral teeth of most Unenlagiinae on the absence of ridged or flutes, presence of denticulated carinae, and crown strongly labiolingual compressed (Novas et al., 2009; Hendrickx et al., 2019). It also differs from the teeth of most microraptorine dromaeosaurids in the presence of mesial carina and an unconstricted crown (Hendrickx et al., 2019). Distinguishing between the teeth of dromaeosaurines and velociraptorines is more challenging. One feature that has been used to distinguish isolated teeth of velociraptorines is the presence of apically hooked denticles, whereas dromaeosaurines typically have denticles oriented perpendicularly to the carina, with symmetrically convex or mostly flat external margins (Currie et al., 1990; Kirkland et al., 1993; Larson, 2008). However, this denticle morphology has also been reported in some dromaeosaurines (Hendrickx et al., 2019). Another common feature of dromaeosaurine lateral teeth, particularly in taxa such as Dromaeosaurus and Utahraptor, is a mesial carina that is strongly twisted toward the lingual surface (Currie et al., 1990; Kirkland et al., 1993).
MNHN/UL.AND.107 has a morphology similar to other specimens from Andrés (morphotype 9), differing mainly in the presence of a denticulated mesial carina and a slightly higher number of denticles in the apical section of the distal carina. Morphotype 8 has a combination of features compatible with Eudromaeosauria but the currently known distribution of some characters within this clade is somewhat ambiguous. The presence of a mesial carina and the symmetrically convex distal margin of the denticles are features more typical for dromaeosaurine lateral teeth, while the absence of a mesial carina and the presence of strongly hooked or slightly asymmetrical denticles are more widespread among velociraptorines. Based on this combination of features and supported by the results of the cladistic analysis, this specimen from Andrés is here attributed to a dromaeosaurid, and tentatively related to Dromaeosaurinae.
Dromaeosauridae (cf. Velociraptorinae) (morphotypes 9 and 10)
Morphotype 9. These specimens are interpreted as lateral tooth crowns based on their strong labiolingual compression. Both the discriminant analyses performed in Python and the random forest analysis classify these specimens as Saurornitholestes and Dromaeosauridae. In the cladistic analysis, this morphotype is recovered at the base of Coelurosauria as the sister taxon of Aorun. These specimens are characterized by the absence of denticulated mesial carina, instead displaying only small crenulations in the apical end of the crown. The absence of mesial denticles, together with the strongly labiolingual compressed and distally recurved crown, are features traditionally used to identify isolated dromaeosaurid teeth (Krumenacker et al., 2017). However, a denticulated mesial carina is present in most dromaeosaurids, including dromaeosaurines and most velociraptorines, such as Velociraptor, Saurornitholestes, and Deinonychus (e.g., Ostrom, 1969; Currie, 1995; Currie et al., 1990; Larson, 2008; Hendrickx et al., 2019). Lateral teeth lacking mesial denticles are common for microraptorines and unenlagiines (which have unserrated mesial and distal carinae), but have also been reported in some early-branching coelurosaurians such as Ornitholestes, in the velociraptorine Tsaagan, and in some teeth of Saurornitholestes (Hendrickx et al., 2019, 2020a). This feature is also present in some isolated teeth from the Upper Cretaceous of Canada attributed to dromaeosaurines (Larson, 2008), as well as in some specimens from the Middle Jurassic of United Kingdom attributed to indeterminate dromaeosaurids (Wills et al., 2023). Similarly, some isolated teeth from the Upper Jurassic of France and Spain, attributed to indeterminate dromaeosaurids, also lack mesial carinae (Gascó et al., 2012; Vullo et al., 2014). MNHN/UL.AND.104 has a strongly labially deflected distal carina, which is a feature described in lateral teeth of Ornitholestes and Dromaeosaurus among coelurosaurian theropods (Hendrickx et al., 2019). The basal cross-section of the specimens from Andrés is lenticular or lanceolate, differing from the figure-of-eight shape shared by most velociraptorines (Sweetman, 2004; Hendrickx et al., 2019). Some isolated teeth from Guimarota, attributed to velociraptorines, have denticulated mesial carinae (Zinke, 1998), while others show only small crenulations in the apical end, similar to that present in morphotype 9 (E.M. pers. obs.). The distal denticles of the specimens from Andrés are mostly oriented perpendicularly to the carina, as occur in dromaeosaurines, not apically hooked as is typical for most (but not all) velociraptorines (Currie et al., 1990; Hendrickx et al., 2019). However, there are some asymmetrically convex denticles, which is similar to the morphology found in specimens from Guimarota (Zinke, 1998). Although the teeth from Guimarota are considerably smaller (mean crown height around 2.3 mm), their overall morphology is otherwise comparable to the specimens from Andrés (Zinke, 1998). Despite the specimens of morphotype 9 show some features that have a not yet well understood distribution, their morphology is compatible with that of velociraptorine dromaeosaurids and they are here tentatively related to this clade.
Morphotype 10. MNHN/UL.AND.212 is interpreted as a lateral tooth crown based on its strong labiolingual compression and the nearly parallel position of the mesial and distal carinae. Despite somewhat ambiguous, the results of the LDAs performed in Python using the clade-level dataset and the random forest analysis based on the genus-level dataset assign this specimen as Dromaeosauridae and Richardoestesia, respectively. The cladistic analysis places this specimen within Coelurosauria, but in a large polytomy with Bicentenaria, Aorun, and Zuolong. This specimen shares several features with morphotype 9 from Andrés such as the absence of a mesial carina, at least in the preserved part of the crown (a large wear facet affects most of the apical end, so the possible presence of denticles apically cannot be exclude). The crown of MNHN/UL.AND.212 seems much less elongated than those of morphotype 9, although this may be due to the presence of the large wear facet. Both morphotypes also have a comparable number of denticles in the distal carina as well as similar shape of the denticles. Additionally, the centrally positioned distal carina in MNHN/UL.AND.212 is similar to the condition observed in MNHN/UL.AND.208. Although the specimen is somewhat incomplete apically and the results from the discriminant and cladistic analyses are inconclusive, it is here interpreted as belonging to a dromaeosaurid, possibly related to Velociraptorinae, based on the previously mentioned similarities with morphotype 9.
The results of the different analyses performed, along with the corresponding taxonomic interpretation for all theropod tooth morphotypes from Andrés, are summarized in Table 1 and Table 2.
Comments on the different methodologies used for the identification of isolated theropod teeth
The different methodological approaches applied to support the taxonomic identification of isolated theropod teeth from the Andrés fossil site yielded generally compatible results, particularly between the discriminant and random forest analyses. This consistency is expected, as both methodologies primarily rely on morphometric variables. The three methodologies (i.e., discriminant, random forest, and cladistics analyses) demonstrated relatively high accuracy in identifying the large specimens attributed to Allosaurus, particularly those of morphotype 1, which are interpreted as mesial teeth. The Reclassification Rates (RR) for the LDA performed in PAST exceeds 74%, with more than 68% of the specimens in the original dataset correctly classified to Allosaurus. The random forest analysis also assigns the specimens of morphotype 1 to this taxon, with probabilities exceeding 70%. However, the results for the Allosaurus lateral teeth, as well as for the smaller teeth, interpreted as belonging to more deeply nested theropods, were more ambiguous. For the identification of these smaller specimens, the discriminant and random forest analyses tended to provide more consistent results than the cladistics analyses, which may be in part due to the scarce Upper Jurassic fossil record currently known for these theropods. As was pointed out on previous works (e.g., Hendrickx et al., 2023; Wills et al., 2023), standard morphometric approaches, such as PCA and LDA, may produce unreliable results. This is often due to missing information for some variables and particularly to unequal sample sizes among taxonomic groups that may highly influence the predicted classification. In fact, the analyses performed in Python, considering only specimens with complete data, showed better results, especially for the classification of smaller teeth, compared to the analyses conducted in PAST. Machine learning techniques, such as those employed in random forest analyses, offer valuable contributions by applying algorithms capable of oversampling and standardization datasets. However, these methods usually require large samples to effectively train the models, which is not always possible when working with the fossil record.
Cladistic analyses often provide more robust results for classifying isolated theropod teeth than those obtained from the discriminant analyses (Hendrickx et al., 2020a, 2023). The use of this tool on the studied sample had relatively good results on classifying the specimens at higher taxonomic level, but recovered poor resolution on less inclusive ranks, particularly for clades with similar dentition or with higher variation on the dental morphology along the tooth row. Overall, the combined application of these different methods proves highly valuable to support the identification of isolated theropod teeth as emphasized in recent studies (e.g., Hendrickx et al., 2020a, 2023; Wills et al., 2023), but the results should be evaluated carefully.
Comparison on the diversity of theropod tooth morphotypes from the Upper Jurassic of the Lusitanian Basin and other European landmasses
A diversity of theropod tooth morphotypes has been described from the Upper Jurassic of the Lusitanian Basin, including both medium to large-sized forms as well as smaller teeth attributed to more deeply nested clades. The assemblage of isolated teeth of medium to large forms is compatible with the fossil record known based on more complete specimens. Non-abelisauroid Ceratosauria (Ceratosaurus), megalosaurids (Torvosaurus), possible metriacanthosaurids, and allosaurids (Allosaurus) have been identified (e.g., Hendrickx and Mateus, 2014b; Hendrickx et al., 2015b; Malafaia et al., 2017a,b). Despite scarcer, tooth morphotypes related to some of these theropods have also been described from other correlative European landmasses (e.g., Canudo et al. 2005; Ruiz-Omeñaca et al., 2008; Gascó et al., 2012; Cobos et al., 2014; Vullo et al., 2014; Gerke and Wings, 2016). Non-abelisauroid ceratosaurian theropods were not identified in the studied sample from Andrés but several mesial and lateral teeth collected from different Kimmeridgian and Tithonian localities of the coastal region of the Lusitanian Basin, as well as an isolated tooth from the Kimmeridgian of Guimarota, have been attributed to Ceratosaurus (Rauhut, 2000; Malafaia et al., 2017b, 2024a). In a few other Upper Jurassic European localities, isolated teeth have been tentatively assigned to this clade, including in Kimmeridgian levels of northern Germany (Gerke and Wings, 2016) and the Oxfordian of Switzerland (Meyer and Thüring, 2003).
The European fossil record of Late Jurassic megalosaurids is relatively abundant (see Appendix 1). These theropods are well represented in the Lusitanian Basin by cranial (including several isolated teeth) and postcranial material, as well as a nest with associated embryo remains collected in different Kimmerigdian and Tithonian localities, all attributed to Torvosaurus (e.g. Araújo et al., 2013; Henrickx and Mateus, 2014a; Malafaia et al., 2017a, b, 2024a). Some isolated teeth from Kimmeridgian-Tithonian levels of Porto das Barcas, assigned to indeterminate “Carnosauria” (sensu Molnar et al., 1990; Rauhut and Kriwet, 1994), may also belong to this taxon, based on the large crown size and the extension of the mesial carina, which ends around the mid-height of the crown. However, this taxon has not yet been identified in more northern regions (e.g., Pombal, Leiria, Batalha), possibly indicating palaeoenvironmental constraints on its geographic distribution (Malafaia et al., 2024b). In other European Late Jurassic areas, megalosaurid records are scarcer and mostly represented by isolated elements. Megalosaurids have been described from Kimmeridgian levels of Asturias (northern Spain) based on some isolated teeth, vertebrae, and footprints (Rauhut et al., 2018b). Other isolated teeth described from different Kimmeridgian localities of Asturias, assigned to possible “Carnosauria” and Carcharodontosauridae (Ruiz-Omeñaca et al., 2008), may also belong to this clade. The Upper Jurassic fossil record of these theropods in Spain also includes a large isolated tooth from the Kimmeridgian-Berriasian Villar del Arzobispo Formation (Cobos et al., 2013). Kimmeridgian levels of northern Germany and the Tithonian of western France have also yielded isolated teeth tentatively assigned to megalosaurids (Vullo et al., 2014). An isolated tooth crown from the Tithonian of western France was assigned to Spinosauridae based on the presence of well-marked ridges on the lingual surface, and comparisons with specimens from the Upper Jurassic Tendaguru Formation, first interpreted as belonging to an early-branching spinosaurid (Buffetaut, 2008, 2013; Vullo et al., 2014). However, the presence of ridges on the lingual surface is also known in the teeth of other theropods, such as Ceratosaurus (e.g., Madsen and Welles, 2000), including isolated teeth assigned to ?Ceratosaurus stechowi from the Tendaguru Formation, and an attribution to a closely related form has been suggested (e.g., Rauhut, 2011). Finally, some isolated teeth from the Upper Jurassic of Portugal and Germany have been assigned to non-megalosauran megalosauroids tentatively related to Marshosaurus (Gerke and Wings, 2016; Malafaia et al., 2017b). These teeth share a general morphology similar to morphotype 5 from Andrés, which is here interpreted as possibly belonging to an early-branching tyrannosauroid. However, MNHN/UL.AND.105 is much smaller, with a more strongly recurved crown and a higher denticles density on the distal carina, suggesting these teeth belong to different theropod groups.
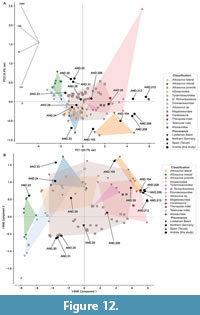 Isolated teeth attributed to allosaurids or indeterminate allosauroids are also relatively abundant in different Upper Jurassic European localities. These theropods are well represented by abundant cranial and postcranial remains attributed to Allosaurus from different Kimmeridgian and Tithonian levels of the Lusitanian Basin (e.g., Pérez-Moreno et al., 1999; Rauhut and Fechner, 2005; Mateus et al., 2006; Malafaia et al. 2017b, 2024a, 2025). Additionally, some isolated teeth from these levels have been tentatively attributed to metriacanthosaurids (Hendrickx et al., 2020a) and carcharodontosaurian allosauroids (Malafaia et al., 2017b). From the Kimmeridgian of northern Germany, an isolated tooth has been interpreted as possibly belonging to Allosaurus, alongside different morphotypes assigned to indeterminate allosauroids (Gerke and Wings, 2016). Tooth morphotypes from the Upper Jurassic of Teruel have also been interpreted as allosaurid (Gascó et al. 2012) and some isolated teeth from Chassiron (western France) were suggested to be related to indeterminate allosauroids (Vullo et al. 2014). Discriminant analyses incorporating theropod tooth morphotypes from different Upper Jurassic European localities show similarities among morphotype 2 from Andrés and other morphotypes from Portugal, northern Germany and Teruel (Figure 12), supporting previous hypotheses that some of these morphotypes may be related to Allosaurus (Gerke and Wings, 2016).
Isolated teeth attributed to allosaurids or indeterminate allosauroids are also relatively abundant in different Upper Jurassic European localities. These theropods are well represented by abundant cranial and postcranial remains attributed to Allosaurus from different Kimmeridgian and Tithonian levels of the Lusitanian Basin (e.g., Pérez-Moreno et al., 1999; Rauhut and Fechner, 2005; Mateus et al., 2006; Malafaia et al. 2017b, 2024a, 2025). Additionally, some isolated teeth from these levels have been tentatively attributed to metriacanthosaurids (Hendrickx et al., 2020a) and carcharodontosaurian allosauroids (Malafaia et al., 2017b). From the Kimmeridgian of northern Germany, an isolated tooth has been interpreted as possibly belonging to Allosaurus, alongside different morphotypes assigned to indeterminate allosauroids (Gerke and Wings, 2016). Tooth morphotypes from the Upper Jurassic of Teruel have also been interpreted as allosaurid (Gascó et al. 2012) and some isolated teeth from Chassiron (western France) were suggested to be related to indeterminate allosauroids (Vullo et al. 2014). Discriminant analyses incorporating theropod tooth morphotypes from different Upper Jurassic European localities show similarities among morphotype 2 from Andrés and other morphotypes from Portugal, northern Germany and Teruel (Figure 12), supporting previous hypotheses that some of these morphotypes may be related to Allosaurus (Gerke and Wings, 2016).
A highly diverse assemblage of small, mostly isolated remains attributed to coelurosaurian theropods has been identified in different Upper Jurassic European localities (see Appendix 1). However, their taxonomic affinities remain difficult to ascertain due to the fragmentary fossil record currently known for most of these clades. The tooth morphotypes from Andrés here interpreted as belonging to coelurosaurian theropods represent a diverse fauna that includes early-branching tyrannosauroids and dromaeosaurids, some tentatively related to dromaeosaurines and velociraptorines. The earliest record of tyrannosauroids in European landmasses is Proceratosaurus from the Middle Jurassic (Bathonian) of England (Rauhut et al., 2010). The Upper Jurassic record of these theropods is represented by Aviatyrannis and Stokesosaurus from Portugal and England, respectively (Rauhut, 2003; Benson, 2008). Isolated teeth attributed to early-branching tyrannosauroids have also been described from Portugal and Germany (Zinke, 1998; Rauhut, 2000; Gerke and Wings, 2016; Malafaia et al., 2017b). Isolated teeth mostly attributed to indeterminate dromaeosaurids have been described in different Upper Jurassic localities in Portugal, Spain, France, and Switzerland (e.g., Zinke, 1998; Rauhut, 2000; Gascó et al., 2012; Hendrickx and Mateus, 2014b). Some isolated teeth from northern Germany were first interpreted as velociraptorine dromaeosaurids (Lubbe et al., 2009) but were later reinterpreted as belonging to different early-branching theropods (Gerke and Wings, 2016). The absence of denticles on the mesial carina in morphotypes 9 and 10 from Andrés is a feature shared with some specimens from the Upper Jurassic of Spain and France, as well as with isolated teeth from the Middle Jurassic of England, all assigned to indeterminate dromaeosaurids (Gascó et al., 2012; Vullo et al., 2014; Wills et al., 2023). The specimens from Andrés have a combination of features compatible with the lateral dentition of velociraptorines, including the absence of mesial denticles, which is a feature more common (though not exclusive) to this clade among dromaeosaurids (e.g., Hendrickx et al., 2019). This shared absence of denticulated mesial carina and similar crown morphology and size suggest the presence of related dromaeosaurids with possible velociraptorine affinities in the Middle Jurassic of England and the Upper Jurassic of Portugal, Spain and France. However, morphotype C specimens from Gascó et al. (2012) show similarities to teeth from Guimarota, tentatively assigned to Compsognathus (Zinke, 1998), sharing the presence of a slight constriction at the crown base and absence of mesial denticles. The assemblage of small coelurosaurian tooth morphotypes identified in the Tithonian of Andrés represents an unusually high diversity. Although the isolated theropod teeth from Guimarota need revision using updated methodologies, some similarities can be identified between the two assemblages.. The fossil record of these two notable Portuguese fossil sites provide important insights for better understanding the diversity and paleogeographic distribution of Late Jurassic theropods.
CONCLUSION
The studied sample of isolated theropod teeth collected from Tithonian levels of Andrés allowed the identification of different morphotypes corresponding to mesial and lateral teeth of Allosaurus. Additionaly, a single tooth is tentatively assigned to a juvenile individual of Allosaurus, a taxon well represented at the site by abundant cranial and postcranial material. Besides, seven other morphotypes were described and interpreted as belonging to several coelurosaurian clades, including indeterminate Coelurosauria, early-branching Tyrannosauroidea, Neocoelurosauria, and dromaeosaurids tentatively related to Velociraptorinae and Dromaeosaurinae. A combination of methodologies (discriminant, machine learning, and cladistic analyses) proved effective in supporting the identification of these isolated teeth. Among these, morphometric-based analyses yielded better results for the identification of small coelurosaurian teeth than cladistic analyses, possibly due to the scarce Upper Jurassic fossil record known for these theropods. This study allowed the identification of the first specimens attributed to tyrannosauroids and possible dromaeosaurines and velociraptorines at the Andrés fossil site. The remarkable diversity and abundance of theropods found here is unusual for the Upper Jurassic European fossil record and provides valuable insights for a better understanding of these faunas, particularly small coelurosaurians. The assemblage of theropod tooth morphotypes from Andrés has several similarities with those described from the Kimmeridgian lignite levels of the Guimarota fossil site. Most theropod clades are represented in both localities, although some morphotypes that are relatively abundant at Guimarota (e.g., Compsognathus and troodontids) appear to be absent at Andrés.
ACKNOWLEDGMENTS
This work was supported by the Portuguese Fundação para a Ciência e a Tecnologia (FCT) I.P./MCTES through the contract [CEECIND/01770/2018] (https://doi.org/10.54499/CEECIND/01770/2018/CP1534/CT0004) and two doctoral scholarships [FCT/IDL (UI/BD/154498/2022)] and [FCT/CEAUL (UI/BD/154258/2022)] as well as national funds (PIDDAC) [UIDB/50019/2020] (https://doi.org/10.54499/UIDB/50019/2020), [UIDP/50019/2020] (https://doi.org/10.54499/UIDP/50019/2020), [LA/P/0068/2020] (https://doi.org/10.54499/LA/P/0068/2020) and [UIDB/00006/2020] (https://doi.org/10.54499/UIDB/00006/2020). We want to thank J. Amorim and Câmara Municipal de Pombal for assistance during fieldwork, J. Alves, R. Keller, and L. Póvoas (Museu Nacional de História Natural e da Ciência, Universidade de Lisboa) for access to the specimens and to the equipment and software for taking the pictures and measurements of the specimens, to L. Castro and P. Moita for discussion on some parts of the work.
REFERENCES
Allain, R. 2001. Redescription de Streptospondylus altdorfensis, le dinosaure théropode de Cuvier, du Jurassique de Normandie. Geodiversitas, 23:349-367.
Alonso, A., Gasca, J.M., Navarro-Lorbés, P., Rubio, C., and Canudo, J.I. 2018. A new contribution to our knowledge of the largebodied theropods from the Barremian of the Iberian Peninsula: The “Barranco del Hocino” site (Spain). Journal of Iberian Geology, 44(1):7-23.
https://doi.org/10.1007/s41513-018-0051-9
Araújo, R., Castanhinha, R., Martins, R.M.S., Mateus, O., Hendrickx, C., Beckmann, F., Schell, N., and Alves, L.C. 2013. Filling the gaps of dinosaur eggshell phylogeny: Late Jurassic theropod clutch with embryos from Portugal. Scientific Reports, 3:1924.
https://doi.org/10.1038/srep01924
Averianov, A.O., Ivantsov, S.V., and Skutschas, P.P. 2019. Theropod teeth from the Lower Cretaceous Ilek Formation of Western Siberia, Russia. Proceedings of the Zoological Institute of the Russian Academy of Sciences, 323:65-84.
https://doi.org/10.31610/trudyzin/2019.323.2.65
Azerêdo, A.C., Cabral, M.C., Martins, M.J., Loureiro, I.M., and Inês, N. 2010. Estudo estratigráfico dum novo afloramento da Formação de Cabaços (Oxfordiano) na região da Serra do Bouro (Caldas da Rainha). Comunicações Geológicas, 97:5-22.
Benson, R.B.J. 2008. New information on Stokesosaurus, a tyrannosauroid (Dinosauria: Theropoda) from North America and the United Kingdom. Journal of Vertebrate Paleontology, 28:732-750.
https://doi.org/10.1671/0272-4634(2008)28[732:NIOSAT]2.0.CO;2
Bidar, A., Demay, L., and Thomel, G. 1972. Compsognathus corallestris, une nouvelle espèce de dinosaurien théropode du Portlandien de Canjuers (Sud-Est de la France), Annales du Muséum of d’histoire naturelle de Nice, 1:9-40.
Bräm, H. 1973. Chelonia from the Upper Jurassic of Guimarota Mine (Portugal). Memórias dos Serviços Geológicos de Portugal, 22:135-141.
Brusatte, S.L., Benson, R.B., Carr, T.D., Williamson, T.E., and Sereno, P.C. 2007. The systematic utility of theropod enamel wrinkles. Journal of Vertebrate Paleontology, 27(4):1052-1056.
https://doi.org/10.1671/0272-4634(2007)27[1052:TSUOTE]2.0.CO;2
Buffetaut, E. 2008. Spinosaurid teeth from the Late Jurassic of Tendaguru, Tanzania, with remarks on the evolutionary and biogeographical history of the Spinosauridae. Travaux et Documents des Laboratoires de Géologie de la Faculté de Sciences de Lyon, 164:26-28.
Buffetaut, E. 2013. An early spinosaurid dinosaur from the Late Jurassic of Tendaguru (Tanzania) and the evolution of the spinosaurid dentition. Oryctos, 10:1-8.
Buffetaut, E., Hébert, F., and Rebours, T. 2010. Un métatarsien de dinosaure théropode dans le Jurassique des falaises des Vaches Noires (Calvados, Normandie, France). Bulletin des Sciences Géologique de Normandes, 1:49-53.
Buscalioni, A.D., Ortega, F., Pérez-Moreno, B.P., and Evans, S.E. 1996. The Upper Jurassic maniraptoran theropod Lisboasaurus estesi (Guimarota, Portugal) reinterpreted as a crocodylomorph. Journal of Vertebrate Paleontology, 16:358-362.
https://doi.org/10.1080/02724634.1996.10011322
Caldwell, M.W., Nydam, R.L., Palci, A., and Apesteguía, S. 2015. The oldest known snakes from the Middle Jurassic-Lower Cretaceous provide insights on snake evolution. Nature Communications., 6:5996.
https://doi.org/10.1038/ncomms6996
Canudo, J.I., and Ruiz-Omeñaca, J.I. 2001. Los restos directos de dinosaurios terópodos (excluyendo aves) en España. Ciencias de la Tierra, 26:347-373.
Canudo, J.I., Aurell, M., Barco, J.L., Cuenca-Bescós, G., and Ruiz-Omeñaca, J.I. 2005. Los dinosaurios de la Formación Villar del Arzobispo (Titónico medio - Berriasiense inferior) en Galve (Teruel). Geogaceta, 38:39-42.
Canudo, J.I., Ruiz-Omeñaca, J., Aurell, M., Barco, J.L., and Cuenca-Bescos, G. 2006. A megatheropod tooth from the late Tithonian - middle Berriasian (Jurassic-Cretaceous transition) of Galve (Aragon, NE Spain). Neues Jahrbuch für Geologie und Paläontologie. Abh., 239:77-99.
Carrano, M.T., Sampson, S.D., and Forster, C.A. 2002. The osteology of Masiakasaurus knopfleri, a small abelisauroid (Dinosauria: Theropoda) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology, 22:510-534.
https://doi.org/10.1671/0272-4634(2002)022[0510:TOOMKA]2.0.CO;2
Cobos, A., Lockley, M.G., Gascó, F., Royo-Torres, R., and Alcalá, L. 2014. Megatheropods as apex predators in the typically Jurassic ecosystems of the Villar del Arzobispo Formation (Iberian Range, Spain). Palaeogeography, Palaeoclimatology, Palaeoecology, 399:31-41.
https://doi.org/10.1016/j.palaeo.2014.02.008
Currie, P.J. 1995. New Information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda). Journal of Vertebrate Paleontology, 15:576-591.
https://doi.org/10.1080/02724634.1995.10011250
Currie, P.J., and Varricchio, D.J. 2004. A new dromaeosaurid from the Horseshoe Canyon Formation (upper Cretaceous) of Alberta, Canada, p. 112-132. In Currie, P.J., Koppelhus, E.B., Shugar, M.A., and Wright, J.L. (ed.), Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Indiana University Press, Bloomington.
Currie, P.J., Rigby Jr.,J.K., and Sloan, R.E. 1990. Theropod teeth from the Judith River Formation of southern Alberta, Canada. P. 107-125. In Carpenter, K., and Currie, P.J. (ed.), Dinosaur Systematics: Perspectives and Approaches 8. Cambridge University Press, Cambridge.
Delcourt, R., Brilhante, N.S., Grillo, O.N., Ghilardi, A.M., Augusta, B.G., and Ricardi-Branco, F. 2020. Carcharodontosauridae theropod tooth crowns from the Upper Cretaceous (Bauru Basin) of Brazil: A reassessment of isolated elements and its implications to palaeobiogeography of the group. Palaeogeography, Palaeoclimatology, Palaeoecology, 556:109870.
https://doi.org/10.1016/j.palaeo.2020.109870
Evers, S.W., and Wings, O. 2020. Late Jurassic theropod dinosaur bones from the Langenberg Quarry (Lower Saxony, Germany) provide evidence for several theropod lineages in the central European archipelago. PeerJ, 8:e8437.
http://doi.org/10.7717/peerj.8437
Farlow, J.O., Brinkman, D.L., Abler, W.L., and Currie, P.J. 1991. Size, shape, and serration density of theropod dinosaur lateral teeth. Modern Geology, 16(1-2):161-198.
Fiorillo, A.R. and Gangloff, R.A. 2000. Theropod teeth from the Prince Creek Formation (Cretaceous) of Northern Alaska, with speculations on Arctic dinosaur paleoecology. Journal of Vertebrate Paleontology, 20:675-682.
https://doi.org/10.1671/0272-4634(2000)020[0675:TTFTPC]2.0.CO;2
Foth, C., and Rauhut, O.W.M. 2017. Re-evaluation of the Haarlem Archaeopteryx and the radiation of maniraptoran theropod dinosaurs. BMC Evolutionary Biology, 17(236):1-16.
http://doi.org/10.1186/s12862-017-1076-y
Frederickson, J.A., Engel, M.H., and Cifelli, R.L. 2018. Niche Partitioning in theropod dinosaurs: diet and habitat preference in predators from the uppermost Cedar Mountain Formation (Utah, U.S.A.). Scientific Reports, 8:17872.
https://doi.org/10.1038/s41598-018-35689-6
Fürsich, F., Schneider, S., Werner, W., Lopez-Mir, B., and Pierce, C.S. 2021. Life at the continental-marine interface: palaeoenvironments and biota of the Alcobaça Formation (Late Jurassic, Central Portugal), with a formal definition of the unit appended. Palaeobiodiversity and Palaeoenvironments, 102:265-329.
https://doi.org/10.1007/s12549-021-00496-x
Gascó, F., Cobos, A., Royo-Torres, R., Mampel, L., and Alcalá, L. 2012. Theropod teeth diversity from the Villar del Arzobispo Formation (Tithonian-Berriasian) at Riodeva (Teruel, Spain). Palaeobiodiversity and Palaeoenvironments, 92:273-285.
https://doi.org/10.1007/s12549-012-0079-3
Gerke, O., and Wings, O. 2016. Multivariate and cladistic analyses of isolated teeth reveal sympatry of theropod dinosaurs in the Late Jurassic of Northern Germany. PLoS ONE, 11:e0158334.
https://doi.org/10.1371/journal.pone.0158334
Göhlich, U.B., and Chiappe, L.M. 2006. A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago. Nature, 440:329-332.
https://doi.org/10.1038/nature04579
Hair, J. F., Black, W. C., Babin, B. J., and Anderson, R. E. 2010. Multivariate Data Analysis (7th ed.). Pearson.
Hammer, Ø., Harper, D.A.T., and Ryan, P.D. 2001. Past: paleontological Statistics Software Package for education and data analysis. Palaeontologia Electronica, 4:1-9.
https://palaeo-electronica.org/2001_1/past/issue1_01.htm
Hendrickx, C., and Mateus, O. 2014a. Torvosaurus gurneyi n. sp., the largest terrestrial predator from Europe, and a proposed terminology of the maxilla anatomy in nonavian theropods. PLoS ONE, 9(3):e88905.
https://doi.org/10.1371/journal.pone.0088905
Hendrickx, C., and Mateus, O. 2014b. Abelisauridae (Dinosauria: Theropoda) from the Late Jurassic of Portugal and dentition-based phylogeny as a contribution for the identification of isolated theropod teeth. Zootaxa, 3759(1):1-74. http://doi.org/10.11646/zootaxa.3759.1.1
Hendrickx, C., Mateus, O., and Araújo, R. 2015a. A proposed terminology of theropod teeth (Dinosauria, Saurischia). Journal of Vertebrate Paleontology, 35:1-18.
https://doi.org/10.1080/02724634.2015.982797
Hendrickx, C., Mateus, O., and Araújo, R. 2015b. The dentition of megalosaurid theropods. Acta Palaeontologica Polonica, 60:627-642.
https://doi.org/10.4202/app.00056.2013
Hendrickx, C., Mateus, O., Araújo, R., and Choiniere, J. 2019. The distribution of dental features in non-avian theropod dinosaurs: taxonomic potential, degree of homoplasy, and major evolutionary trends. Palaeontologia Electronica, 22:1-110.
https://doi.org/10.26879/820
Hendrickx, C., Tschopp, E., and Ezcurra, M. 2020a. Taxonomic identification of isolated theropod teeth: the case of the shed tooth crown associated with Aerosteon (Theropoda: Megaraptora) and the dentition of Abelisauridae. Cretaceous Research, 108:104312.
https://doi.org/10.1016/j.cretres.2019.104312
Hendrickx, C., Stiegler, J., Currie, P.J., Han, F., Xu, X., Choiniere, J.N., and Wu, X.-C. 2020b. Dental anatomy of the apex predator Sinraptor dongi (Theropoda: Allosauroidea) from the Late Jurassic of China. Canadian Journal of Earth Sciences, 57:1127-1147.
https://doi.org/10.1139/cjes-2019-0231
Hendrickx, C., Trapman, T. H., Wills, S., Holwerda, F. M., Stein, K. H. W., Rauhut, O.W.M., Melzer, R.R., Woensel, J.V., and Reumer, J.W.F. 2023. A combined approach to identify isolated theropod teeth from the Cenomanian Kem Kem Group of Morocco: cladistic, discriminant, and machine learning analyses. Journal of Vertebrate Paleontology, 43:e2311791.
https://doi.org/10.1080/02724634.2024.2311791
Henkel, S., and Krebs, B. 1977. Der erste Fund eines Saugetier-Skelettes aus der Jura-Zeit. Umschau in Wissenschaft und Technik, 77:217 -218.
Hutt, S., Naish, D., Martill, D.M., Barker, M.J., and Newbery, P. 2001. A preliminary account of a new tyrannosauroid theropod from the Wessex Formation (Early Cretaceous) of southern England. Cretaceous Research, 22:227-242.
https://doi.org/10.1006/cres.2001.0252
Huxley, T.H. 1869. On the upper jaw of Megalosaurus. Quarterly Journal of the Geological Society of London, 25:311-314.
Isasmendi, E., Perez-Pueyo, M., Moreno-Azanza, M., Alonso, A., Puertolas-Pascual, E., Badenas, B., and Canudo, J.I. 2024. Theropod teeth palaeodiversity from the uppermost Cretaceous of the South Pyrenean Basin (NE Iberia) and the intra-Maastrichtian faunal turnover. Cretaceous Research, 162:105952.
https://doi.org/10.1016/j.cretres.2024.105952
Kirkland, J.I., Burge, D., and Gaston, R. 1993. A large dromasosaur (Theropoda) from the Lower Cretaceous of Eastern Utah. Hunteria, 2:1-16.
Knoll, F., Buffetaut, E., and Bülow, M. 1999. A theropod braincase from the Jurassic of the Vaches Noires cliffs (Normandy, France); osteology and palaeoneurology. Bulletin de la Societe Geologique de France, 170(1):103-109.
Krebs, B., and Schwarz, D. 2000. The crocodiles from the Guimarota mine, p. 69-75. In Martin, T., and Krebs, B. (ed.), Guimarota: a Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, München.
Kriwet, J. 2000. The fish fauna from the Guimarota mine, p. 41-50. In Martin, T., and Krebs, B. (ed.), Guimarota - A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, München.
Krumenacker, L.J., Simon, D.J., Scofield, G., and Varricchio, D.J. 2017. Theropod dinosaurs from the Albian-Cenomanian Wayan Formation of eastern Idaho. Historical Biology, 29:170-186.
https://doi.org/10.1080/08912963.2015.1137913
Kullberg, J.C., Rocha, R.B., Soares, A.F., Rey, J., Terrinha, P., Azerêdo, A.C., Callapez, P., Duarte, L.V., Kullberg, M.C., Martins, L., Miranda, R., Alvez, C., Mata, J., Madeira, J., Mateus, O., Moreira, M., and Nogueira, C.R. 2013. A Bacia Lusitaniana: Estratigrafia, Paleogeografia e Tectónica, p. 195-347. In Dias, R., Araújo, A., Terrinha, P., and Kullberg, J.C. (ed), Geologia de Portugal, Volume II - Geologia Meso-cenozóica de Portugal. Escolar Editora, Lisboa.
Larson, D.W. 2008. Diversity and variation of theropod dinosaur teeth from the uppermost Santonian Milk River Formation (Upper Cretaceous), Alberta: a quantitative method supporting identification of the oldest dinosaur tooth assemblage in Canada. Canadian Journal of Earth Sciences, 45:1455-1468.
https://doi.org/10.1139/E08-070
Larson, D.W., and Currie, P.J. 2013. Multivariate analyses of small theropod dinosaur teeth and implications for paleoecological turnover through time. PLoS One, 8(1):e54329.
https://doi.org/10.1371/journal.pone.0054329
Louchart, A., and Pouech, J. 2017. A tooth of Archaeopterygidae (Ayes) from the Lower Cretaceous of France extends the spatial and temporal occurrence of the earliest birds. Cretaceous Research, 73:40-46.
https://doi.org/10.1016/j.cretres.2017.01.004
Lubbe, T. van der, Richter, U., and Knötschke, N. 2009. Velociraptorine dromaeosaurid teeth from the Kimmeridgian (Late Jurassic) of Germany. Acta Palaeontologica Polonica, 54:401-408.
https://doi.org/10.4202/app.2008.0007
Lydekker, R. 1888. Catalogue of the Fossil Reptilia and Amphibia in the British Museum (Natural History). Part 1, Ornithosauria, Crocodylia, Dinosauria, Squamata, Rhynchocephalia, and Proterosauria. British Museum (Natural History), London.
Madsen, J.H. 1976. A second new theropod from the Late Jurassic of east central Utah. Utah Geology, 3(1):51-60.
Madsen, J.H., and Welles, S.P. 2000. Ceratosaurus (Dinosauria, Theropoda): a revised osteology. Utah Geological Survey, Miscellaneous Publication, 00-2:1-89.
Malafaia, E., Ortega, F., Escaso, F., Dantas, P., Pimentel, N., Gasulla, J.M., Ribeiro, B., Barriga, F., and Sanz, J.L. 2010. Vertebrate fauna at the Allosaurus fossil-site of Andrés (Upper Jurassic), Pombal, Portugal. Journal of Iberian Geology, 36:193-204. http://doi.org/10.5209/rev_JIGE.2010.v36.n2.7
Malafaia, E., Ortega, F., Escaso, F., and Silva, B. 2015. New evidence of Ceratosaurus (Dinosauria: Theropoda) from the Late Jurassic of the Lusitanian Basin, Portugal. Historical Biology, 27:938-946.
https://doi.org/10.1080/08912963.2014.915820
Malafaia, E., Mocho, P., Escaso, F., and Ortega, F. 2017a. New data on the anatomy of Torvosaurus and other remains of megalosauroid (Dinosauria, Theropoda) from the Upper Jurassic of Portugal. Journal of Iberian Geology, 43:33-59.
https://doi.org/10.1007/s41513-017-0003-9
Malafaia, E., Escaso, F., Mocho, P., Serrano-Martínez, A., Torices, A., Cachão. M., and Ortega, F. 2017b. Analysis of diversity, stratigraphic and geographical distribution of isolated theropod teeth from the Upper Jurassic of the Lusitanian Basin, Portugal. Journal of Iberian Geology, 43:257-291.
https://doi.org/10.1007/s41513-017-0021-7
Malafaia, E., Mocho, P., Escaso, F., and Ortega, F. 2020. A new carcharodontosaurian theropod from the Lusitanian Basin: evidence of allosauroid sympatry in the European Late Jurassic. Journal of Vertebrate Paleontology, 40(1):e1768106.
https://doi.org/10.1080/02724634.2020.1768106
Malafaia, E., Mocho, P., Escaso, F., Narvaéz, I., and Ortega, F. 2024a. Taxonomic and stratigraphic update of the material historically attributed to Megalosaurus from Portugal. Acta Palaeontologica Polonica, 69:127-171.
https://doi.org/10.4202/app.01113.2023
Malafaia, E., Mocho, P., Escaso, F., Narvaéz, I., Gasulla, J. M., Pérez-García, A., Marcos-Fernandez, F., and Ortega, F. 2024b. The continental vertebrate fossil record from Pombal: contribution to the knowledge of Upper Jurassic faunas of the Lusitanian Basin. Comunicações Geológicas (in press).
Malafaia, E., Dantas, P., Escaso, F., Mocho, P., and Ortega, F. 2025. Cranial osteology of a new specimen of Allosaurus Marsh, 1877 (Theropoda: Allosauridae) from the Upper Jurassic of Portugal and a specimen-level phylogenetic analysis of Allosaurus. Zoological Journal of the Linnean Society, 204(1):zlaf029.
https://doi.org/10.1093/zoolinnean/zlaf029
Manuppella, G., Rodrigues, A., Oliveira, J., and Carreira de Deus, P. 1974. Carta Geológica de Portugal, folha 23-A (Pombal). Escala 1:50 000 (1ª ed). Serviços Geológicos de Portugal.
Manuppella, G., Zbyszewski, G., and Veiga Ferreira, O. 1978. Notícia Explicativa da Folha 23-A (Pombal). Serviços Geológicos de Portugal.
Marques, C. S., Dufourq, E., Pereira, S., Santos, V.F., Malafaia, E. 2025. Enhancing the classification of isolated theropod teeth using machine learning: a comparative study. PeerJ, 13:e19116.
https://doi.org/10.7717/peerj.19116
Martill, D.M., Naish, D., and Earland, S. 2006. Dinosaurs in marine strata: evidence from the British Jurassic, including a review of the allochthonous vertebrate assemblage from the marine Kimmeridge Clay Formation (Upper Jurassic) of Great Britain. Actas de las III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno, Salas de los Infantes, Spain, p. 47-84.
Martin, T. 2013. Mammalian postcranial bones from the Late Jurassic of Portugal and their implications for forelimb evolution. Journal of Vertebrate Paleontology, 33:1432-1441.
https://doi.org/10.1080/02724634.2013.771780
Martin, T., and Schultz, J.A. 2023. Deciduous dentition, tooth replacement, and mandibular growth in the Late Jurassic docodontan Haldanodon exspectatus (Mammaliaformes). Journal of Mammalian Evolution, 30:507-531.
https://doi.org/10.1007/s10914-023-09668-2
Mateus, O. 1998. Lourinhanosaurus antunesi, a new Upper Jurassic allosauroid (Dinosauria: Theropoda) from Lourinhã, Portugal. Memórias da Academia de Ciências de Lisboa, 37:111-124.
Mateus, O., Walen, A., and Antunes, M.T. 2006. The large theropod fauna of the Lourinhã Formation (Portugal) and its similarity to the Morrison Formation, with a description of a new species of Allosaurus, p. 123-129. In Foster, J.R., and Lucas, S.G. (ed.), Paleontology and Geology of the Upper Jurassic Morrison Formation 36. Albuquerque: New Mexico Museum of Natural History and Science, Albuquerque.
Meyer, C.A., and Thüring, B. 2003. Dinosaurs of Switzerland. Comptes Rendus Paleovol, 2(1):103-117.
https://doi.org/10.1016/S1631-0683(03)00005-8
Mocho, P., Royo-Torres, R., Malafaia, E., Escaso, F., Narváez, I., and Ortega, F. 2017. New data on Late Jurassic sauropods of central and northern sectors of the Bombarral Sub-basin (Lusitanian Basin, Portugal). Historical Biology, 29:151-169.
https://doi.org/10.1080/08912963.2015.1137912
Molnar, R.E., Kurzanov, S.M., and Dong, Z. 1990. Carnosauria, p. 169-209. In Weishampel, D.B., Dodson, P., and Osmólska, H. (ed.), The Dinosauria. University of California Press, Berkeley.
Monvoisin, E., Allain R., Buffetaut E., and Picot L. 2022. New data on the theropod diversity from the Middle to Late Jurassic of the Vaches Noires cliffs (Normandy, France). Geodiversitas, 44:385-415.
https://doi.org/10.5252/geodiversi-tas2022v44a12
http://geodiversitas.com/44/12
Naish, D., and Martill, D.M. 2007. Dinosaurs of Great Britain and the role of the Geological Society of London in their discovery: basal Dinosauria and Saurischia. Journal of the Geological Society, London, 164:493-510.
https://doi.org/10.1144/0016-76492006-032
Novas, F.E., Pol, D., Canale, J.I., Porfiri, J.D., and Calvo, J.O. 2009. A bizarre Cretaceous theropod dinosaur from Patagonia and the evolution of Gondwanan dromaeosaurids. Proceedings of the Royal Society B, 276:1101-1107.
https://doi.org/10.1098/rspb.2008.1554
Ortega, F., Escaso, F., Gasulla, J.M., Dantas, P., and Sanz, J.L. 2006. Dinosaurios de la Península Ibérica. Estudios Geológicos, 62(1):219-240.
Ostrom, J.H. 1969. Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana. Bulletin of the Peabody Museum of Natural History, 30:1-165.
Owen, R. 1863. On the Archaeopteryx of von Meyer with a description of a long-tailed species, from the lithographic stone of Solenhofen. Philosophic Transactions, Royal Society, London, 153:3-47.
Pedregosa, F., Varoquaux, G., Gramfort, A., Michel, V., Thirion, B., Grisel, O., Blondel, M., Prettenhofer, P., Weiss, R., Dubourg, V., Vanderplas, J., Passos, A., Cournapeau, D., Brucher, M., Perrot, M., and Duchesnay, É. 2011. Scikit-learn: machine learning in Python. Journal of Machine Learning Research, 12:2825-2830.
Peyer, K. 2006. A reconsideration of Compsognathus from the upper Tithonian of Canjuers, Southeastern France. Journal of Vertebraye Paleontology, 26:879-896.
https://doi.org/10.1671/0272-4634(2006)26[879:AROCFT]2.0.CO;2
Pérez-Moreno, B.P., Chure, D.J., Pires, C., Marques da Silva, C., dos Santos, V., Dantas, P., Póvoas, L., Cachão, M., Sanz, J.L., and Galopim de Carvalho, A.M. 1999. On the presence of Allosaurus fragilis (Theropoda: Carnosauria) in the Upper Jurassic of Portugal: first evidence of an intercontinental dinosaur species. Journal of the Geological Society, London, 156, 449-452.
https://doi.org/10.1144/gsjgs.156.3.0449
Piveteau, J. 1923. L’arrière-crâne d’un dinosaurien carnivore de l’Oxfordien de Dives. Annales de Paléontology, 12:1-11.
Rauhut, O.W.M. 2000. The dinosaur fauna from the Guimarota mine, p. 75-82. In Martin, T., and Krebs, B. (ed.), Guimarota - A Jurassic Ecosystern. Verlag Dr. Friedrich Pfeil, München.
Rauhut, O.W.M. 2001. Herbivorous dinosaurs from the Late Jurassic (Kimmeridgian) of Guimarota, Portugal. Proceedings of the Geologists’ Association, 112:273-283.
https://doi.org/10.1016/S0016-7878(01)80007-9
Rauhut, O.W.M. 2003. A tyrannosauroid dinosaur from the Upper Jurassic of Portugal. Palaeontology, 46:903-910.
https://doi.org/10.1111/1475-4983.00325
Rauhut, O.W.M. 2011. Theropod dinosaurs from the Late Jurassic of Tendaguru (Tanzania). Special Papers in Palaeontology, 86:195-239.
Rauhut, O.W.M., and Fechner, R. 2005. Early development of the facial region in a non-avian theropod dinosaur. Proceedings of the Royal Society B, 272:1179-1183.
https://doi.org/10.1098/rspb.2005.3071
Rauhut, O.W.M., and Kriwet, J. 1994. Teeth of a big theropod dinosaur from Porto das Barcas (Portugal). Berliner geowiss. Abh. (E), 13:179-185.
Rauhut, O.W.M., Milner, A.C., and Moore-Fay, S. 2010. Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England. Zoological Journal of the Linnean Society, 158:155-195.
https://doi.org/10.1111/j.1096-3642.2009.00591.x
Rauhut, O.W.M., Foth, C., Tischlinger, H., and Norell, M. A. 2012. Exceptionally preserved juvenile megalosauroid theropod dinosaur with filamentous integument from the Late Jurassic of Germany. PNAS, 109:11746-11751.
https://doi.org/10.1073/pnas.1203238109
Rauhut, O.W.M., Foth, C., and Tischlinger, H. 2018a. The oldest Archaeopteryx (Theropoda: Avialiae): a new specimen from the Kimmeridgian/Tithonian boundary of Schamhaupten, Bavaria. PeerJ, 6:e4191.
https://doi.org/10.7717/peerj.4191
Rauhut, O.W.M., Piñuela, L., Castanera, D., García-Ramos, J.C., and Cela, I.S. 2018b. The largest European theropod dinosaurs: remains of a gigantic megalosaurid and giant theropod tracks from the Kimmeridgian of Asturias, Spain. PeerJ, 6:e4963.
https://doi.org/10.7717/peerj.4963
Ruiz-Omeñaca, J.I., Piñuela, L., García-Ramos, J.C., and Canudo J.I. 2008. Dientes de dinosaurios carnívoros (Saurischia: Theropoda) del Jurásico Superior de Asturias. Actas de las III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno, Salas de los Infantes, Spain, p. 273-291.
Ruiz-Omeñaca, J.I., Pereda Suberbiola, X., Julio Company, J., and Belinchón, M. 2010. Dientes de arcosaurios (Crocodyliformes, Theropoda) del Jurásico Superior de Buñol (Valencia). Geogaceta, 48:79-82.
Schwarz, D., and Fechner, R. 2004. Lusitanisuchus, a new generic name for Lisboasaurus mitracostatus (Crocodylomorpha: Mesoeucrocodylia), with a description of new remains from the Upper Jurassic (Kimmeridgian) and Lower Cretaceous (Berriasian) of Portugal. Canadian Journal of Earth Sciences, 41:1259-1271.
https://doi.org/10.1139/e04-059
Sereno, P.C., Dutheil, D.B., Hans, M.I., Larsson, C.E., Lyon, G.H., Magwene, P.M., Sidor, C.A., Varricchio, D.J., and Wilson, J.A. 1996. Predatory dinosaurs from the Sahara and Late Cretaceous faunal differentiation. Science, 272:986-991. http://doi.org/10.1126/science.272.5264.986
Serrano-Martínez, A., Vidal, D., Sciscio, L., Ortega, F., and Knoll, F. 2016. Isolated theropod teeth from the Middle Jurassic of Niger and the early dental evolution of Spinosauridae. Acta Palaeontologica Polonica, 61:403-415. https://doi.org/10.4202/app.00101.2014
Smith, J.B. 2005. Heterodonty in Tyrannosaurus rex: implications for the taxonomic and systematic utility of theropod dentitions. Journal of Vertebrate Paleontology, 25:865-887.
https://doi.org/10.1671/0272-4634(2005)025[0865:HITRIF]2.0.CO;2
Smith, J.B. 2007. Dental morphology and variation in Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology, 27:103-126.
https://doi.org/10.1671/0272-4634(2007)27[103:DMAVIM]2.0.CO;2
Smith, J.B, and Dodson, P. 2003. A proposal for a standard terminology of anatomical notation and orientation in fossil vertebrate dentitions. Journal of Vertebrate Paleontology, 23(1):1-12.
https://doi.org/10.1671/0272-4634(2003)23[1:APFAST]2.0.CO;2
Smith, J.B. Vann, D.R., and Dodson, P. 2005. Dental morphology and variation in theropod dinosaurs: implications for the taxonomic identification of isolated teeth. The Anatomic Record Part A, 285:699-736.
https://doi.org/10.1002/ar.a.20206
Sweetman, S.C. 2004. The first record of velociraptorine dinosaurs (Saurischia, Theropoda) from the Wealden (Early Cretaceous, Barremian) of southern England. Cretaceous Research, 25:353-364.
https://doi.org/10.1016/j.cretres.2004.01.004
Taquet, P., and Welles, S. 1977. Redescription du crâne de dinosaure théropode de Dives (Normandie). Annales de Paléontologie, 63:191-206.
Teixeira, C., Zbyszewski, G., Torre de Assunção, C., and Manuppella, G. 1968. Notícia Explicativa da Folha 23-C (Leiria). Serviços Geológicos de Portugal.
Vullo, R., Abit, D., Ballèvre, M., Billon-Bruyat, J.-P., Bourgeais, R., Buffetaut, É., Daviero-Gomez, V., Garcia, G., Gomez, B., Mazin, J.-M., Morel, S., Néraudeau, D., Pouech, J., Rage, J.-C., Schnyder, J., and Tong, H. 2014. Palaeontology of the Purbeck-type (Tithonian, Late Jurassic) bonebeds of Chassiron (Oléron Island, western France). Comptes Rendus Palevol, 13:421-441.
https://doi.org/10.1016/j.crpv.2014.03.003
Wagner, A. 1861. Neue Beiträge zur Kenntnis der urweltlichen Fauna des lithographischen Schiefers; V. Compsognathus longipes Wagner. Abhandlungen der Bayerischen Akademie der Wissenschaften, 9, 30-38.
Walker, A.D. 1964. Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs. Philosophical Transactions of the Royal Society of London, Series B, 248:53-135.
https://doi.org/10.1098/rstb.1964.0009
Weigert, A. 1995. Isolierte Zähne von cf. Archaeopteryx sp. aus dem Oberen Jura der Kohlengrube Guimarota (Portugal). Neues Jahrbuch für Geologie und Paläontologie, Monatshefte H, 9:562-576.
Wellnhofer, P. 1988. A new specimen of Archaeopteryx. Science, 240(4860):1790-1792.
https://doi.org/10.1126/science.240.4860.1790
Wiechmann, M.F. 2000. The albanerpetontids from the Guimarota mine, p. 51-54. In Martin, T., and Krebs, B. (ed.), Guimarota: a Jurassic ecosystem. Verlag Dr. Friedrich Pfeil, München.
Wiechmann, M.F., and Gloy, U. 2000. Pterosaurs and urvogels from the Guimarota mine, p. 83-86. In Martin, T., and Krebs, B. (ed.), Guimarota: a Jurassic ecosystem. Verlag Dr. Friedrich Pfeil, München.
Wills, S., Underwood, C.J., and Barrett, P.M. 2021. Learning to see the wood for the trees: Machine learning, decision trees, and the classification of isolated theropod teeth. Palaeontology, 64(1):75-99.
https://doi.org/10.1111/pala.12512
Wills, S., Underwood, C.J., and Barrett, P.M. 2023. Machine learning confirms new records of maniraptoran theropods in Middle Jurassic UK microvertebrate faunas. Papers in Palaeontology, 9:e1487.
https://doi.org/10.1002/spp2.1487
Young, C.M.E., Hendrickx, C., Challands, T.J., Foffa, D., Ross, D.A., Butler, I.B., and Brusatte, S.L. 2019. New theropod dinosaur teeth from the Middle Jurassic of the Isle of Skye, Scotland. Scottish Journal of Geology, 55(1):7-19.
https://doi.org/10.1144/sjg2018-020
Zavorka, M., and Perrett, G. 2014. Discriminant Function Analysis: An Application for Predicting Fish Habitat Use. Fisheries Management and Ecology, 21(5):392-400.
Zinke, J. 1998. Small theropod teeth from the Upper Jurassic coal mine of Guimarota (Portugal). Paläontologische Zeitschrift, 72(1/2):179-189.
Zinke, J., and Rauhut, O.W.M. 1994. Small theropods (Dinosauria, Saurischia) from the Upper Jurassic and Lower Cretaceous of the Iberian Peninsula. Berliner geowissenschaftliche Abhandlungen, E13:163-177.
