

 An intriguing new species of dabbling duck (Aves: Anseriformes) from the middle Miocene of Austria
An intriguing new species of dabbling duck (Aves: Anseriformes) from the middle Miocene of Austria
Article number: 26.3.a52
https://doi.org/10.26879/1334
Copyright Paleontological Society, December 2023
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 25 August 2023. Acceptance: 6 November 2023.
ABSTRACT
Several dozen species of Miocene anatids are known worldwide, but few are known from abundant finds and even fewer from nearly complete specimens. Most fossil ducks have been described from very scant fragmentary remains, making it very difficult or impossible to compare them with one another. In Austria, only a few heavily damaged remains of Miocene anatids have been found so far. Here, we describe a new genus and species of a nearly complete medium-sized duck preserved on four slabs from the middle Miocene of Lavanttal, southern Austria. A number of features on various parts of the skeleton including the coracoid, humerus and pelvis indicate that it was a dabbling duck in an ecological sense, and exclude the option of it being a diving duck. The new species shows a unique mosaic of features typical of different groups of ducks and some features specific to it. Its more precise systematic position is uncertain, pending future finding of more fossil specimens.
Zbigniew M. Bochenski. Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Slawkowska 17, 31-016 Krakow, Poland. (corresponding author). bochenski@isez.pan.krakow.pl
https://orcid.org/0000-0002-9405-3933
Johannes Happ. Reptilienzoo Happ, Villacherstrasse 237, 9020 Klagenfurt, Austria. johanneshapp@yahoo.de
https://orcid.org/0009-0007-3206-4315
Grzegorz Salwa. Regional Subcarpathian John Paul II Hospital in Krosno, Department of Neurology, Korczyńska 57, 38-400 Krosno, Poland. grzegorz.tomasz.salwa@gmail.com
https://orcid.org/0000-0003-0589-4675
Teresa Tomek. Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Slawkowska 17, 31-016 Krakow, Poland. toteresa45@gmail.com
https://orcid.org/0000-0002-8278-3031
Key words: new species; new genus; Miocene; fossil dabbling duck; Anseriformes; Lavanttal, Austria
Final citation: Bochenski, Zbigniew M., Happ, Johannes, Salwa, Grzegorz, and Tomek, Teresa. 2023. An intriguing new species of dabbling duck (Aves: Anseriformes) from the middle Miocene of Austria. Palaeontologia Electronica, 26(3):a52.
https://doi.org/10.26879/1334
palaeo-electronica.org/content/2023/5038-a-miocene-dabbling-duck
Copyright: December 2023 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
https://zoobank.org/82EEBDFB-7DE0-44F6-9FD3-BA4903721526
INTRODUCTION
The Lavanttal in Carinthia, southern Austria, is known for numerous and diverse middle Miocene fossils, which include plants, invertebrates and vertebrates representing terrestrial, marine and freshwater environments (e.g., Grímsson et al., 2015, 2020; Nolf and Brzobohatý, 2009; Prieto et al., 2019; Wank, 1991). Among these numerous finds are the remains of only two different passerine birds (Happ et al., 2022). Here, we describe a new genus and species of a medium-sized duck preserved on four slabs from this middle Miocene site.
The early evolution of the Anseriformes is poorly understood (Mayr, 2022); also the relationships and systematic division within the family Anatidae are constantly discussed and subject to change. The oldest definitely “duck-like” anseriforms are Romainvillia stehlini from the Late Eocene of France (Lebedinsky, 1927) and Romainvillia kazakhstanensis from Kazakhstan (Zelenkov, 2018). Romainvilliinae are also known from the Early Oligocene of Belgium (Mayr and Smith, 2001), England (Mayr 2008) and France (Mayr and De Pietri, 2013). The oldest representatives of the crown group Anatidae date back to the late Oligocene. These include the genera Mionetta and Pinpanetta from France and Australia, respectively (Mayr, 2022), and Aminornis and Teleornis from Argentina (Tambussi et al., 2023). In recent years, there have been a number of attempts to determine affinities among anatids (Worthy, 2009; Worthy and Lee, 2008; Worthy et al., 2022; Zelenkov et al., 2018), but this problem has still not been fully resolved (Mayr et al., 2022; Worthy et al., 2022; Zelenkov, 2020).
Worldwide, several dozen anatid species are known from the Miocene (Mlíkovský, 2002; Zelenkov, 2020, 2023b). Some Miocene anatids, such as Mionetta blanchardi from Europe or Manuherikia lacustrina from New Zealand, are known from abundant finds (Cheneval, 1987; Worthy et al., 2007). Sites rich in a diverse fauna of anseriforms are known from Europe (Mlíkovský, 2002), Asia (Gorobets, 2013; Zelenkov, 2011, 2012a, 2012b, 2020, 2023b; Zelenkov and Kurochkin, 2012), Australia and New Zealand (Worthy et al., 2007, 2022; Worthy, 2009). However, a number of fossil species have been described from very scant fragmentary remains, making it very difficult or impossible to compare them with one another (Mayr et al., 2022).
In Austria, only a few heavily damaged remains of Miocene anatids have been found so far. The oldest duck remains (Anatidae gen. and sp. indet.) come from the early Miocene of Oberdorf (Mlíkovský, 1998). Fragmentary remains of “Dendronessa” sp. from Götzendorf, and Chenoanas cf. sansaniensis and Anserinae indet. from Atzelsdorf are dated to the late Miocene (Mlíkovský, 1991 and Göhlich, 2009, respectively). Apart from the bone fragment of Anatidae indet. from the early Miocene, the new specimen described here is the oldest, most complete and best preserved duck from Austria, and one of the few near-complete ducks in the world.
MATERIAL AND METHODS
The nomenclature and classification of modern taxa follow Dickinson and Remsen (2013) and osteological terminology follows Baumel and Witmer (1993). Measurements are in millimeters.
The nearly complete specimen described here is preserved on four slabs (A-D) in the form of fossilized bones and their imprints. The thin layer of matrix covering some bone fragments was removed during the tedious, time-consuming preparation of the specimen by one of the co-authors (GS). Originally, the well-preserved right coracoid was visible on slab A in ventral view, which made it impossible to see the diagnostic features on its dorsal side. Taking advantage of the fact that this bone was not articulated with humerus, scapula or sternum, it was gently carved from the slab and the remnants of the surrounding rock were removed. Currently, it is a three-dimensional object and can be viewed from all sides.
The following extant species of the Anatidae were available for comparison in the osteological collection of the Institute of Systematics and Evolution of Animals, Polish Academy of Sciences (ISEA PAS): Dendrocygninae: Dendrocygna arborea, D. autumnalis, D. bicolor, D. eytoni, D. viduata. Oxyurinae: Oxyura jamaicensis. Anserinae: Anser albifrons, A. anser, A. caerulescens, A. canagicus, A. erythropus, A. fabalis, A. indicus, Branta bernicla, B. canadensis, B. leucopsis, B. ruficollis, B. sandvicensis, Cereopsis novaehollandiae, Coscoroba coscoroba, Cygnus atratus, C. columbianus, C. cygnus, C. melanocoryphus, C. olor. Anatinae: Mergini: Bucephala clangula, B. islandica, Clangula hyemalis, Lophodytes cucullatus, Melanitta fusca, M. nigra, Mergellus albellus, Mergus merganser, M. serrator, Somateria mollissima, S. spectabilis. Tadornini: Alopochen aegyptiaca, Chloephaga picta, Tadorna ferruginea, T. tadorna, T. variegata. Aythyni: Aythya ferina, A. fuligula, A. marila, A. nyroca, Netta rufina. Anatini: Amazonetta brasiliensis, Anas acuta, A. bahamensis, A. capensis, A. crecca, A. formosa, A. gibberifrons, A. platyrhynchos, A. poecilorhyncha, A. rubripes, A. superciliosa, Malacorhynchus membranaceus, Mareca strepera, M. falcata, M. penelope, M. sibilatrix, Spatula clypeata, S. cyanoptera, S. discors, S. querquedula, S. rhynchotis. Genera incertae sedis: Aix galericulata, A. sponsa, Biziura lobata, Cairina moschata, Callonetta leucophrys, Chenonetta jubata, Nettapus auritus, N. coromandelianus, N. pulchellus, Stictonetta naevosa.
In the case of Miocene ducks, in addition to original papers describing a given taxon, other publications describing features and/or containing informative illustrations were also used for comparisons. These additional publications, if any, are listed here in square brackets after the taxonomic authorities of the species: Dunstanetta Worthy, Tennyson, Jones, McNamara, and Douglas, 2007 [Worthy, 2009; Worthy and Lee, 2008; Worthy et al., 2008, 2022]; Manuherikia Worthy, Tennyson, Jones, McNamara, and Douglas, 2007 [Worthy, 2009; Worthy and Lee, 2008; Worthy et al., 2008, 2022]; Mionetta blanchardi (Milne-Edwards, 1863) [Cheneval, 1983; Howard, 1964; Livezey and Martin, 1988; Milne-Edwards, 1867-1868; Canterbury Museum’s account on www.sketchfab.com; Worthy and Lee, 2008]; Mionetta consobrina (Milne-Edwards, 1867-1868) [Lambrecht, 1933; Howard, 1964; Cheneval, 1983]; Mionetta natator (Milne-Edwards, 1867-1868) [Lambrecht, 1933; Švec, 1981; Cheneval, 1983; Göhlich, 2002]; Pinpanetta Worthy, 2009 [Worthy et al., 2022]; Aythya denesi (Kessler, 2013) [Zelenkov 2016]; Aythya molesta (Kurochkin, 1985) [Zelenkov, 2012a, 2016]; Aythya shihuibas Hou, 1985 [Stidham, 2015]; Miotadorna sanctibathansi Worthy, Tennyson, Jones, McNamara, and Douglas, 2007 [Worthy and Lee, 2008]; Anas kurochkini Zelenkov et Panteleyev, 2015 [Zelenkov, unpublished photo]; Matanas enrighti Worthy, Tennyson, Jones, McNamara and Douglas, 2007; Mioquerquedula minutissima Zelenkov and Kurochkin, 2012 [Zelenkov, 2023a]; Mioquerquedula soporata (Kurochkin, 1976) [Zelenkov, 2023a; Zelenkov and Kurochkin, 2012]; Bambolinietta lignitifila (Portis, 1884) [Mayr and Pavia, 2014]; Chenoanas asiatica Zelenkov, Stidham, Martynovich, Volkova, Li, Qui, 2018; Chenoanas deserta Zelenkov, 2012b [Zelenkov et al., 2018]; Chenoanas sansaniensis (Milne-Edwards, 1867-1868) [Göhlich, 2009; Zelenkov et al., 2018]; Aix praeclara Zelenkov and Kurochkin, 2012; Nogusunna conflictoides Zelenkov, 2011; Protomelanitta gracilis Zelenkov, 2011 [Zelenkov, 2023a]; Protomelanitta velox (Milne-Edwards, 1867-1868) [Švec, 1981; Zelenkov, 2023a; Zelenkov and Kurochkin, 2012]; Sharganetta mongolica Zelenkov, 2011.
Three ratios between skeletal elements were calculated: humerus length/ulna length (brachial index), humerus length/carpometacarpus length, and carpometacarpus length/coracoid medial length.
SYSTEMATIC PALAEONTOLOGY
Class AVES Linnaeus, 1758
Order ANSERIFORMES Wagler, 1831
Family ANATIDAE Leach, 1819
Genus LAVANTTALORNIS gen. nov.
zoobank.org/9592EB4E-37D8-457E-881B-0DBAA710D6EF
Type species. Lavanttalornis hassleri gen. et sp. nov.
Etymology. After the Lavanttal valley where the specimen was found, added to the Ancient Greek ὄρνις (órnis) meaning “bird”.
Taxonomic remarks. The new species resembles Anatidae in overall morphology and a unique combination of the following characters: the end of the beak bears foramina neurovascularia; the shape and proportions of the pelvis correspond to those of anatids; the coracoid bears a deeply concave cotyla scapularis of a rounded-triangular shape, short processus procoracoideus, and laterally protruding facies articularis humeralis; the humerus bears the facet of the tuberculum supracondylare ventrale that is cranially buttressed and tilted distally, and there is no prominent tuberculum supracondylare dorsale; in tibiotarsus, the condylus medialis projects much medially in relations to distal shaft, and lateral side of distal shaft forms a sharp edge that slants anteriorly towards condylus lateralis.
Differential diagnosis. Lavanttalornis hassleri gen. et sp. nov. is distinguished from:
* Mionetta blanchardi (Milne-Edwards, 1863), from the late Oligocene to middle Miocene of Europe, in: coracoid with medial margin of the omal section of the shaft strongly inclined medially and almost straight, processus acrocoracoideus hardly protruding beyond the medial margin of shaft, and facies articularis sternalis almost straight, humerus with roundish condylus ventralis, and distal end expanded little ventrally, and relatively wide phalanx proximalis digiti majoris with bulging caudal edge;
* Mionetta consobrina (Milne-Edwards, 1867-1868), from the early Miocene of Europe, in: coracoid with medial margin of the omal section of the shaft strongly inclined medially and almost straight, and processus acrocoracoideus hardly protruding beyond the medial margin of shaft;
* Mionetta natator (Milne-Edwards, 1867-1868), from the late Oligocene to early Miocene of Europe, in: coracoid with medial margin of the omal section of the shaft strongly inclined medially and almost straight, processus acrocoracoideus hardly protruding beyond the medial margin of shaft, and facies articularis sternalis almost straight;
* Miotadorna sanctibathansi Worthy, Tennyson, Jones, McNamara, and Douglas, 2007, from the early middle Miocene of New Zealand, in: coracoid with facies articularis sternalis almost straight, and humerus with distal end expanded little ventrally;
* Manuherikia Worthy, Tennyson, Jones, McNamara, and Douglas, 2007, and Dunstanetta Worthy, Tennyson, Jones, McNamara, and Douglas, 2007, from the early middle Miocene of New Zealand, in: coracoid with medial margin of the omal section of the shaft strongly inclined medially and almost straight, and humerus with relatively thick shaft that does not narrow distally;
* Pinpanetta Worthy, 2009, from the late Oligocene/early Miocene of Australia, in: coracoid with medial margin of the omal section of the shaft strongly inclined medially and almost straight, and processus acrocoracoideus hardly protruding beyond the medial margin of shaft;
* Sharganetta mongolica Zelenkov, 2011, Nogusunna conflictoides Zelenkov, 2011, and Protomelanitta gracilis Zelenkov, 2011, from the middle Miocene of Mongolia, in: humerus with shallow notch of incisura capitis in proximal outline of the bone, and distal end expanded little ventrally;
* Chenoanas deserta Zelenkov, 2012b, and Chenoanas asiatica Zelenkov, Stidham, Martynovich, Volkova, Li, Qui, 2018, both from the middle Miocene of eastern Russia and Mongolia, Chenoanas sansaniensis (Milne-Edwards, 1867-1868), from the early to middle Miocene of Europe, eastern Russia and Mongolia, Aix praeclara Zelenkov et Kurochkin, 2012, Mioquerquedula soporata (Kurochkin, 1976), and Mioquerquedula minutissima Zelenkov et Kurochkin, 2012, all from the middle Miocene of Mongolia, and Protomelanitta velox (Milne-Edwards, 1867), from the middle Miocene of France, in: coracoid with medial margin of the omal section of the shaft strongly inclined medially and almost straight, and processus acrocoracoideus hardly protruding beyond the medial margin of shaft;
* Aythya denesi (Kessler, 2013) from the late Miocene of Hungary, and Bambolinetta lignitifila (Portis, 1884) from the late Miocene of Italy, in: humerus with relatively thick shaft that does not narrow distally;
* Aythya shihuibas Hou, 1985, from the late Miocene of China, and Aythya molesta (Kurochkin, 1985), from the late Miocene of Mongolia, in: humerus with shallow notch of incisura capitis in proximal outline of the bone;
* Matanas enrighti Worthy, Tennyson, Jones, McNamara and Douglas, 2007, from the early-middle Miocene of New Zealand, in: coracoid with processus acrocoracoideus hardly protruding beyond the medial margin of shaft;
* Anas kurochkini Zelenkov et Panteleyev, 2015, from the late Miocene of northeastern Sea of Azov region, Russia, in: coracoid with medial margin of the omal section of the shaft strongly inclined medially and almost straight, thick crista acrocoracoidea, facies articularis humeralis with acute cranial angle and projecting little laterally, and facies articularis sternalis almost straight;
* all extant ducks by a unique combination of characters: coracoid with medial margin of the omal section of the shaft strongly inclined medially and almost straight, ventral plane of processus acrocoracoideus subparallel to the ventral plane of the extremitas sternalis, processus acrocoracoideus hardly protruding beyond the medial margin of shaft, and facies articularis sternalis almost straight, humerus with distal end expanded little ventrally, and relatively wide phalanx proximalis digiti majoris with bulging caudal edge.
Lavanttalornis hassleri sp. nov.
Figure 1, Figure 2, Figure 3A-E, Figure 4, Figure 5A, Figure 6A
zoobank.org/9E7230E3-FC50-4AE6-987B-E1F0DD6C7C95
Etymology. The species is named after the finder of the specimen, Dr. Andreas Hassler of Sankt Andrä, a veterinarian by profession but a paleontologist by passion.
Holotype. LMK-Pal 7453a+a’+b+c+d (Figure 1, Figure 2, Figure 3A-E, Figure 4, Figure 5A, Figure 6A), partly articulated nearly complete skeleton on four slabs from which the right coracoid has been removed, cleaned and is visible as a 3D object; housed at the paleontological collection of the Landesmuseum Kärnten (LMK-Pal), Klagenfurt, Austria.
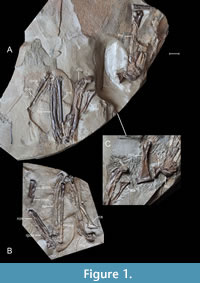 Type locality and horizon. A site near the village of Schaßbach, about 50 km northeast of Klagenfurt in the Lavanttal, Carinthia, southern Austria. GPS coordinates using WGS 84 datum: 46.79752°N 14.80676°E at an altitude of 471 m above sea level. The specimen is dated to the Upper Sarmatian s. s. (upper middle Miocene), ca 12 Ma. It was found in freshwater sediments that contain two coal seams lying above the Middle Sarmatian layers (Reischenbacher et al., 2007).
Type locality and horizon. A site near the village of Schaßbach, about 50 km northeast of Klagenfurt in the Lavanttal, Carinthia, southern Austria. GPS coordinates using WGS 84 datum: 46.79752°N 14.80676°E at an altitude of 471 m above sea level. The specimen is dated to the Upper Sarmatian s. s. (upper middle Miocene), ca 12 Ma. It was found in freshwater sediments that contain two coal seams lying above the Middle Sarmatian layers (Reischenbacher et al., 2007).
Diagnosis. As for the genus.
Measurements. Measurements (in mm) including total length (TL) and others, taken from slabs A, B or C: coracoid (A, dext) TL, 47.6, medial length, 42.4, sternal width, 17.7; scapula (A, sin) articular height, 11.2; humerus (A, sin) TL, 75.5, proximal width, 18.7, distal width, 10.6, least shaft width, 6.3; radius (B, dext) TL, 62.6; ulna (B, dext) TL, 69.5; carpometacarpus (B, dext) TL, 46.4, proximal width, 11.9; phalanx proximalis dig. majoris (B, dext) TL, 19.6; phalanx dig. minoris (B, dext) TL=9.6; pelvis (C) TL, 64.4, width between antitrochanters, 30.5.
Description and Comparison
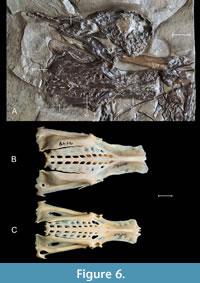
 Skull. The skull, visible in lateral view on slabs C and D, is partly an imprint and partly fossilized bone remains (Figure 2, Figure 6A). As in Mionetta blanchardi, the length of the beak is approximately the same as that of the skull. The braincase is too damaged for meaningful comparisons which also applies to the preserved small fragment of ramus mandibulae. As in extant anseriforms, the foramina neurovascularia are visible at the end of the beak. The caudal part of the nostril is elongated and tapered, while in the Miocene Mioquerquedula sp., Bambolinietta lignitifila and extant anseriforms, the caudal part of the nostril is rounded.
Skull. The skull, visible in lateral view on slabs C and D, is partly an imprint and partly fossilized bone remains (Figure 2, Figure 6A). As in Mionetta blanchardi, the length of the beak is approximately the same as that of the skull. The braincase is too damaged for meaningful comparisons which also applies to the preserved small fragment of ramus mandibulae. As in extant anseriforms, the foramina neurovascularia are visible at the end of the beak. The caudal part of the nostril is elongated and tapered, while in the Miocene Mioquerquedula sp., Bambolinietta lignitifila and extant anseriforms, the caudal part of the nostril is rounded.
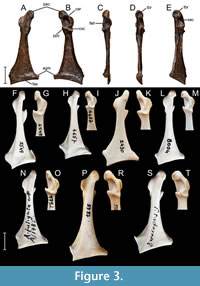 Coracoid. The well-preserved right coracoid removed from slab A (Figure 1, Figure 3A-E) shows a very thick, massive shaft that is somewhat similar to that of the Miocene Anas kurochkini, Mionetta blanchardi and extant Anas, Spatula, Mareca, and Melanitta; Mionetta natator and extant species of the genera Aythya, Bucephala, and Mergus have much slender shafts. A feature specific to Lavanttalornis hassleri is the medial margin of the omal section of the shaft, which is strongly inclined medially and almost straight. In Chenoanas, medial margin of shaft is also straight but it does not incline medially, whereas in Mionetta blanchardi, Mionetta consobrina, Mionetta natator, Aix praeclara, Mioquerquedula minutissima, Mioquerquedula soporata, Anas kurochkini, Pinpanetta, Manuherikia, Dunstanetta, Protomelanitta velox, and most extant taxa (e.g., Aix, Anas, Spatula, Mareca, Aythya, Clangula, Melanitta) the medial margin of shaft is much less medially inclined, and is usually slightly arched. As, for example, in the extant genus Anas s.l., the ventral plane of the processus acrocoracoideus is subparallel to the ventral plane of the extremitas sternalis (Worthy and Lee, 2008: character 45). The processus acrocoracoideus hardly protrudes beyond the medial margin of the shaft. This distinguishes it from the genera Chenoanas and Pinpanetta, Matanas enrighti, Aix praeclara, Mioquerquedula minutissima, Mioquerquedula soporata, Protomelanitta velox, Mionetta consobrina, Mionetta natator, and to a lesser extent from Mionetta blanchardi and many extant taxa (e.g., Aix, Anas, Mareca, Spatula, Clangula). The tuberculum brachiale (name by Zelenkov and Kurochkin, 2012; Zelenkov and Panteleyev, 2015) is concave and slightly wider dorsoventrally than craniocaudally high. As in extant species of the genus Anas but unlike the extinct Anas kurochkini, the crista acrocoracoidea (name by Elzanowski et al., 2012) is thick. As in all anatids, the margin of the facies articularis humeralis projects laterally, although not as far as in Anas kurochkini, and in lateral view it has an acute cranial angle. The sulcus m. acrocoracoidei is deeply excavated near the facies articularis humeralis, which is observed in taxa from various groups. As in many ducks, the processus proccoracoideus is relatively short, projecting craniomedially, and the cotyla scapularis is deeply concave of a rounded-triangular shape. The angulus medialis is short, blunt ended, and its short cranial edge is almost perpendicular to the shaft; in Mionetta blanchardi the angulus medialis merges with the shaft gradually, in Pinpanetta a distinct flange of various shape extends cranially along the medial shaft margin, and in Protomelanitta velox, Manuherikia, and Miotadorna sanctibathansi the angulus medialis is acute. In ventral view, the edge of the facies articularis sternalis is almost straight, whereas in Mionetta blanchardi, Mionetta natator, Anas kurochkini, Mioquerquedula minutissima, Manuherikia lacustrina, and Miotadorna sanctibathansi it arches sternally.
Coracoid. The well-preserved right coracoid removed from slab A (Figure 1, Figure 3A-E) shows a very thick, massive shaft that is somewhat similar to that of the Miocene Anas kurochkini, Mionetta blanchardi and extant Anas, Spatula, Mareca, and Melanitta; Mionetta natator and extant species of the genera Aythya, Bucephala, and Mergus have much slender shafts. A feature specific to Lavanttalornis hassleri is the medial margin of the omal section of the shaft, which is strongly inclined medially and almost straight. In Chenoanas, medial margin of shaft is also straight but it does not incline medially, whereas in Mionetta blanchardi, Mionetta consobrina, Mionetta natator, Aix praeclara, Mioquerquedula minutissima, Mioquerquedula soporata, Anas kurochkini, Pinpanetta, Manuherikia, Dunstanetta, Protomelanitta velox, and most extant taxa (e.g., Aix, Anas, Spatula, Mareca, Aythya, Clangula, Melanitta) the medial margin of shaft is much less medially inclined, and is usually slightly arched. As, for example, in the extant genus Anas s.l., the ventral plane of the processus acrocoracoideus is subparallel to the ventral plane of the extremitas sternalis (Worthy and Lee, 2008: character 45). The processus acrocoracoideus hardly protrudes beyond the medial margin of the shaft. This distinguishes it from the genera Chenoanas and Pinpanetta, Matanas enrighti, Aix praeclara, Mioquerquedula minutissima, Mioquerquedula soporata, Protomelanitta velox, Mionetta consobrina, Mionetta natator, and to a lesser extent from Mionetta blanchardi and many extant taxa (e.g., Aix, Anas, Mareca, Spatula, Clangula). The tuberculum brachiale (name by Zelenkov and Kurochkin, 2012; Zelenkov and Panteleyev, 2015) is concave and slightly wider dorsoventrally than craniocaudally high. As in extant species of the genus Anas but unlike the extinct Anas kurochkini, the crista acrocoracoidea (name by Elzanowski et al., 2012) is thick. As in all anatids, the margin of the facies articularis humeralis projects laterally, although not as far as in Anas kurochkini, and in lateral view it has an acute cranial angle. The sulcus m. acrocoracoidei is deeply excavated near the facies articularis humeralis, which is observed in taxa from various groups. As in many ducks, the processus proccoracoideus is relatively short, projecting craniomedially, and the cotyla scapularis is deeply concave of a rounded-triangular shape. The angulus medialis is short, blunt ended, and its short cranial edge is almost perpendicular to the shaft; in Mionetta blanchardi the angulus medialis merges with the shaft gradually, in Pinpanetta a distinct flange of various shape extends cranially along the medial shaft margin, and in Protomelanitta velox, Manuherikia, and Miotadorna sanctibathansi the angulus medialis is acute. In ventral view, the edge of the facies articularis sternalis is almost straight, whereas in Mionetta blanchardi, Mionetta natator, Anas kurochkini, Mioquerquedula minutissima, Manuherikia lacustrina, and Miotadorna sanctibathansi it arches sternally.
 Scapula. Articular parts with small fragments of the shaft of the left and right scapula are visible on slab A in medial (costal) view; slab B has an imprint of the right scapula (Figure 1, Figure 4A). As in Mionetta blanchardi and some extant taxa (e.g., Anas acuta, A.platyrhynchos, Spatula quercquedula, S. clypeata, Melanitta fusca), the acromion deviates very slightly dorsally; its dorsal edge forms an almost straight line with the margo dorsalis of the corpus scapulae. In many other species (e.g., Aix galericulata, Anas crecca, Mareca strepera, Aythya ferina, A.fuligula, Clangula hyemalis, Bucephala clangula, Mergellus albellus), the acromion is much more dorsally curved.
Scapula. Articular parts with small fragments of the shaft of the left and right scapula are visible on slab A in medial (costal) view; slab B has an imprint of the right scapula (Figure 1, Figure 4A). As in Mionetta blanchardi and some extant taxa (e.g., Anas acuta, A.platyrhynchos, Spatula quercquedula, S. clypeata, Melanitta fusca), the acromion deviates very slightly dorsally; its dorsal edge forms an almost straight line with the margo dorsalis of the corpus scapulae. In many other species (e.g., Aix galericulata, Anas crecca, Mareca strepera, Aythya ferina, A.fuligula, Clangula hyemalis, Bucephala clangula, Mergellus albellus), the acromion is much more dorsally curved.
Humerus. The left humerus is visible on slab A in cranial view and the right humerus in cranioventral view. Slab B shows an imprint of the dorsocaudal side of the right bone with a preserved tuberculum ventrale (Figure 1, Figure 4A-C, F). As in the Miocene Aythya denesi, the notch of incisura capitis in proximal outline of the humerus is shallow (Worthy and Lee, 2008: character 59) whereas in the late Oligocene Pinpanetta from Australia as well as in extant dendrocygnines, anserines and tadornines it is (almost) missing. The notch is more pronounced in Mionetta blanchardi, Sharganetta mongolica, Nogusunna conflictoides, Protomelanitta gracilis, Aythya molesta, A. shihuibas, Manuherikia, and Dunstanetta. Extant species of such genera as Aix, Anas, Spatula, Mareca, Aythya, Melanitta, or Mergellus also show a more pronounced notch. The tuberculum ventrale is directed caudo-cranially so its distal margin is approximately at right angles to the fossorial plane (Worthy and Lee, 2008: character 57). A similarly directed tuberculum ventrale is found in Pinpanetta, Aythya shihuibas and Aythya denesi, whereas in Miotadorna sanctibathansi, Mionetta, and the genus Chenoanas, the tuberculum ventrale is proximally oriented. As in extant Aix galericulata, Anas crecca, Spatula clypeata, Spatula querquedula, the tuberculum ventrale is narrow (forms a narrow ridge); the tuberculum is wide in many other extant taxa including Anas platyrhynchos, Anas acuta, Aythya ferina, Aythya fuligula, Bucephala clangula, Clangula hyemalis, Melanitta. The length of the crista deltopectoralis is comparable with the proximal width of the humerus. Other details of the crista deltopectoralis as well as details of the remaining structures of the proximal humerus, including tuberculum dorsale, crista bicipitalis, fossa pneumotricipitalis and capital shaft ridge, are not visible. As in Pinpanetta fromensis, Pinpanetta vickersrichae and Mionetta blanchardi the shaft is relatively thick, and it does not narrow distally but its sides are essentially parallel to each other (Worthy and Lee, 2008: character 61). The humerus of Bambolinetta lignitifila was definitely even more robust and stouter. In Aythya denesi, Manuherikia and Dunstanetta, Pinpanetta tedfordi and the similarly sized extant species, the shaft narrows distally. The shaft is s-shaped, in dorsal view. A similarly bent shaft is found in many extant taxa, including Aix galericulata, Anas crecca, Anas acuta, Anas platyrhynchos, Mareca strepera, Mareca penelope, Spatula clypeata, Spatula querquedula, Bucephala clangula, and Clangula hyemalis. In contrast, the shaft is more straight in such species as Aythya fuligula or Mergellus albellus. As in Mionetta, Manuherikia, Dunstanetta, Malacorhynchus, Stictonetta, Nomonyx, and Pinpanetta, the facet of the tuberculum supracondylare ventrale (attachment of the anterior articular ligament) is cranially buttressed (Worthy and Lee, 2008: character 65), whereas the unbuttressed facet is observed in Anseranas, Thalassornis, Biziura, and Oxyura. Moreover, in Lavanttalornis hassleri, the facet is tilted distally. As in Miocene Mionetta blanchardi and extant species such as Aix galericulata or Aythya fuligula, but unlike e.g., Spatula clypeata, fossa m. brachialis (brachial fossa) is large, well-defined and forms a marked depression (Worthy and Lee, 2008: character 68). It differs from that in Pinpanetta, where it is elongate and dorsal margin barely extends past midshaft width. As in Mionetta blanchardi, the processus flexorius extends roughly as far distally as condylus dorsalis (Worthy, 2009: character 63). As in Miocene Mionetta blanchardi and Sharganetta mongolica, and extant Anatinae, there is no prominent tuberculum supracondylare dorsale (ectepicondylar prominence) (Worthy and Lee 2008, character 64). A distinct ectepicondylar prominence is present in the Miocene Pinpanetta as well as in extant anserines. Contrary to Mionetta blanchardi, Pinpanetta, Miotadorna sanctibathansi, Sharganetta mongolica, Nogusunna conflictoides, and Protomelanitta gracilis, the distal end of the humerus is little expanded ventrally, and the condylus ventralis is roundish (more oblong in Mionetta, Pinpanetta, and Sharganetta but also roundish in Protomelanitta and Miotadorna).
Ulna. The right ulna is visible on plate A in ventral view, and its imprint is on plate B. The bone is clearly less stout and relatively longer than that of Bambolinetta lignitifila. Its length is in the range of Mionetta blanchardi; ulna of M. consorbina was longer, and that of M. natator was shorter. The proximal and distal parts are crushed, preventing meaningful comparisons.
Radius. The right proximal radius is visible on plate B in anterior view, and its distal posterior side is imprinted there. Plate A shows the distal right radius with a large portion of shaft in posterior view, and an imprint of the anterior proximal part (Figure 1, Figure 4F-G). The bone differs significantly from that of Bambolinetta lignitifila, from which it is clearly less stout and relatively longer. As in Mionetta blanchardi, the shaft is more straight than, for example, in extant Spatula clypeata. As in Spatula clypeata, the capital tuberosity (name used by Howard, 1929) is roundish but clearly larger than in Spatula, whereas in some other extant species, such as Aythya ferina or Melanitta nigra the tuberculum is more oval. As in Mionetta blanchardi and Spatula clypeata, the tuberculum aponeurosis ventralis is proximodistally relatively short, whereas in some extant species, such as Clangula hyemalis or Melanitta fusca the tuberculum is significantly longer.
 Carpometacarpus. The right carpometacarpus is visible on slab B in dorsal view, and its imprint is preserved on slab A. Moreover, the distal part of the left bone is visible on slab D in dorsal view (Figure 2, Figure 4D). As in Mionetta blanchardi and such modern taxa as Anas, Spatula, Mareca or Bucephala, the processus extensorius is broad proximo-distally and a distinct arcuate indentation separates it from the processus alularis; in Clangula hyemalis, the indentation is very small. Also as in Mionetta blanchardi and such extant taxa as Aix galericulata, Anas acuta, Anas crecca, A. platyrhynchos, Spatula clypeata, Spatula querquedula, Mareca penelope, Mareca strepera, Aythya fuligula, and A. ferrina, the symphysis metacarpalis distalis is long in relation to the carpometacarpus width measured just distad of spatium intermetacarpale (Worthy and Lee, 2008: character 84). The symphysis is relatively shorter in many other species (e.g., Bucephala clangula, Clangula hyemalis, Melanitta fusca, M.nigra, Mergellus albellus. There is a distinct nodule in the distal dorsal side of the os metacarpale majus. A similar nodule is in extant Anas platyrhynchos, Anas acuta, Anas crecca, Mareca penelope, Mareca strepera, Spatula querquedula, Spatula clypeata, Aythya fuligula, Aythya ferina, and Clangula hyemalis. In Bucephala clangula, Melanitta fusca, M.nigra, Mergus merganser, Mergus serrator the nodule is small or absent.
Carpometacarpus. The right carpometacarpus is visible on slab B in dorsal view, and its imprint is preserved on slab A. Moreover, the distal part of the left bone is visible on slab D in dorsal view (Figure 2, Figure 4D). As in Mionetta blanchardi and such modern taxa as Anas, Spatula, Mareca or Bucephala, the processus extensorius is broad proximo-distally and a distinct arcuate indentation separates it from the processus alularis; in Clangula hyemalis, the indentation is very small. Also as in Mionetta blanchardi and such extant taxa as Aix galericulata, Anas acuta, Anas crecca, A. platyrhynchos, Spatula clypeata, Spatula querquedula, Mareca penelope, Mareca strepera, Aythya fuligula, and A. ferrina, the symphysis metacarpalis distalis is long in relation to the carpometacarpus width measured just distad of spatium intermetacarpale (Worthy and Lee, 2008: character 84). The symphysis is relatively shorter in many other species (e.g., Bucephala clangula, Clangula hyemalis, Melanitta fusca, M.nigra, Mergellus albellus. There is a distinct nodule in the distal dorsal side of the os metacarpale majus. A similar nodule is in extant Anas platyrhynchos, Anas acuta, Anas crecca, Mareca penelope, Mareca strepera, Spatula querquedula, Spatula clypeata, Aythya fuligula, Aythya ferina, and Clangula hyemalis. In Bucephala clangula, Melanitta fusca, M.nigra, Mergus merganser, Mergus serrator the nodule is small or absent.
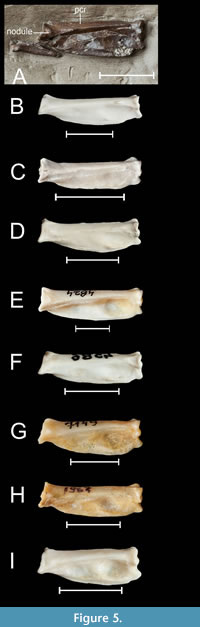
Phalanx proximalis digiti majoris. The right phalanx, detached from the carpometacarpus, is visible on slab B in dorsal view, and its imprint is preserved on slab A (Figure 1, Figure 5). For a duck, the phalanx is relatively wide in relation to its length, which is the result of a bulging of the caudal edge. In this respect, it differs from Mionetta blanchardi and most extant duck species whose phalanx is narrower and its caudal and cranial edges are often almost parallel to each other. One of the few taxa with a similarly wide phalanx is Anas platyrhynchos, but other members of the Anatini, such as Spatula clypeata, have narrower phalanges. A broad phalanx is also found e.g., in Aix galericulata of uncertain systematic affiliation. In the proximal part of the phalanx there is a nodule clearly raised both above the dorsal surface and the pila cranialis. A similar nodule is found in species of the genus Anas s.l. Other modern ducks either do not have such a nodule, or if they do, it is less pronounced, especially in relation to pila cranialis.
Pelvis. Pelvis is visible on slab C in dorsal view, and its imprint is preserved on slab D (Figure 2, Figure 6). The overall proportions of the pelvis, that is, length to width between antitrochanters and to width of the ala preacetabularis ilii, are similar to that of Anas platyrhynchos; some of the extant genera, e.g., Netta, Aythya, Oxyura or Biziura have a much narrower pelvis. Mionetta blanchardi also had a narrower pelvis, especially for ala preacetabularis ilii. Also as in Anas platyrhynchos, Spatula clypeata or Aix gallericulata, the pre-acetabular and post-acetabular parts are of similar length. In Aythya, Netta, Oxyura, Bucephala, or Biziura, the post-acetabular region is elongated to varying degrees. As in Mionetta blanchardi and many extant taxa including Aix, Anas, Aythya, Bucephala, and Melanitta, the caudal edges of the ala ischii and ala postacetabularis ilii extend roughly equally far caudally; in Dendrocygna, the ala ischii reaches further caudally. As in all Anatidae, foramina intertransversaria can be discerned.
 Femur. Portions of the left and right femur are visible on slab C, and their imprints are on slab D. The bones are too damaged to allow meaningful comparisons.
Femur. Portions of the left and right femur are visible on slab C, and their imprints are on slab D. The bones are too damaged to allow meaningful comparisons.
Tibiotarsus. The distal part of the right tibiotarsus is visible on slab D in anterolateral view and its imprint is on slab C. Portions of the left tibiotarsus and their imprints are on both slabs (Figure 2, Figure 4E). As in Mionetta blanchardi and extant Anatidae, the condylus medialis projects much medially in relations to distal shaft and the lateral side of distal shaft forms a sharp edge that slants anteriorly towards condylus lateralis (Bochenski and Tomek, 2009: characters 7a, 7b and 14’b). The pons supratendineus is located in the middle of the shaft and opens into the incisura intercondylaris, which is also typical of anseriforms. In contrast to the Miocene genus Chenoanas, the lateral margin of the condylus lateralis is medially inclined (in Chenoanas, it is positioned in line with the lateral margin of the shaft), and condyli lateralis and medialis are proximodistally long (in Chenoanas, the condyli are shorter than the width of the incisura intercondylaris).
Ratios between selected skeletal elements. For Lavanttalornis hassleri, the brachial index (humerus length/ulna length) is 1.1; similar values of this ratio are also observed in some extant species in all other groups of Anatinae (Appendix 1). Slightly lower values (approx. 1.0) are typical for all Dendrocygninae and parts of Tadornini, and slightly higher values (approx. 1.2) occur in various groups. The ratio humerus length/carpometacarpus length for Lavanttalornis hassleri is 1.6, and similar values were most often recorded in extant Anatini (in 15 species out of 20 examined) and in several species of Mergini, Tadornini and genera incertae sedis. It is true that higher values of this ratio (1.7–2.2) were recorded in some species among all taxonomic groups examined, but only in Dendrocygninae, Oxyurinae, and Aythyni, i.e., in the groups of good divers, all species showed these higher values. The ratio of carpometacarpus length/coracoid medial length for Lavanttalornis hassleri is 1.1, and similar values were recorded in most of the examined Anatini (12 species out of 18), and most of the examined species of uncertain taxonomic position (incertae sedis: 7 out of 10). Again, the most homogeneous groups with different values of this ratio than in Lavanttalornis were Dendrocygninae and Tadornini, in which all species showed higher values (1.2–1.4). In other species that dive for food, including Aythyni (4 out of 5), Mergini (6 out of 9) and Oxyura jamacensis, these values were lower (0.9–1.0).
DISCUSSION
A number of features on various parts of the skeleton can be used to infer about the duck’s way of life. In the examined specimen, such features are preserved mainly on three skeletal elements. In coracoid, the ventral plane of the processus acrocoracoideus is not much deviated from (lies subparallel to) the ventral plane of the extremitas sternalis, which is a usual condition in dabbling ducks; in Dendrocygninae, Oxyurinae, Aythyni, and Mergini, i.e., in ducks that dive while feeding, the ventral plane of the processus acrocoracoideus is more inclined (more perpendicular to) the ventral plane of the extremitas sternalis (Zelenkov, 2011; Zelenkov and Panteleyev, 2015; Worthy and Lee, 2008: character 45). In addition, the acute cranial angle of the facies articularis humeralis is also typical of dabbling ducks (Zelenkov and Panteleyev, 2015). Also, the distally tilted facet of the tuberculum supracondylare ventrale (distal humerus) is a usual condition in dabbling ducks whereas a medial rotation of the facet is common to all divers (McCracken et al., 1999; Worthy, 2009). Finally, the broad pelvis and its unextended post-acetabular part are also typical of dabbling ducks. Ducks that dive while foraging, such as Oxyura, Biziura, or Aythya, usually have a much narrower pelvis with extended post-acetabular section, which is interpreted as an adaptation to this type of activity (Raikow, 1970, 1971; Worthy and Lee, 2008). Although none of the three studied ratios gives a clear answer as to the ecology of Lavanttalornis hassleri, certain trends are clearly visible. It appears that the brachial index is relatively the least diverse among different groups of anatids, which is probably related to similar wing kinematics in all ducks (Nudds et al., 2007). The other two ratios (humerus length/carpometacarpus length and carpometacarpus length/coracoid medial length) seem to do a somewhat better job of dividing extant ducks into different groups. In both cases, Lavanttalornis is found in one group with most extant species of Anatini and other non-diving ducks. Species from good-diving groups (Dendrocygninae and Aythyni) have different proportions of bones examined. These preliminary results are promising but should be treated with caution because only one specimen per species was examined and not all species of anatids were included. More detailed analyses of the proportions of individual skeletal elements in all anatids are beyond the scope of the current study; they may be the subject of separate research. The above arguments taken together indicate unequivocally that Lavanttalornis hassleri was not a diving duck. Many osteological features are similar to those of dabbling ducks in an ecological sense.
Most of the new species of Miocene anatids have been described in new genera specially created for them (Mayr et al., 2022). Modern genera of ducks begin to dominate only in the late Miocene (Zelenkov, 2020). Given the unique mosaic of features typical of different ducks as well as features specific to Lavanttalornis hassleri, and the fact that it is of middle Miocene age, we decided to describe it in a new genus. Many features indicate that ducks that dive for food, including Dendrocygninae, Oxyurinae, and Aythyni can be excluded from the groups to which Lavanttalornis could belong, and Tadornini can probably be excluded from among non-diving ducks. It is more difficult to indicate the taxonomic group to which Lavanttalornis could belong. The similarity to Anatini is supported by many features but may be superficial, and considering the large number of ducks of uncertain systematic affiliation (genera incertae sedis) and the fact that the caudal side of humerus with significant phylogenetic features has not been preserved, it is difficult to draw more precise conclusions about the systematic affiliations of Lavanttalornis.
Most skeletal elements are preserved in articulation, but some bones, including both scapulae, the right coracoid, the left humerus, and the right phalanx proximalis digiti majoris, are no longer connected to the adjacent bones, but lie close to them. This indicates that the carcass was exposed to natural decomposition agents, including putrefaction gases, for some time before being buried in sediments, in a calm environment without much underwater turbulences that could stimulate its disarticulation and loss of skeletal elements (Schäfer, 1972; Bienkowska-Wasiluk, 2010). This was also the case with the passerine bird described earlier from the same site (Happ et al., 2022).
Many Miocene ducks are known from very few finds, sometimes even single incomplete bones (e.g., Boev, 2007, 2020; Gál et al., 2000; Göhlich, 2009; Kessler and Venczel, 2009). This makes it very difficult or even impossible to compare such specimens with those previously described. Only occasionally can they be identified as previously described taxa; just as rarely they may be references to fossil specimens to be described in the future. Lavanttalornis hassleri does not suffer from this defect. The unique feature of specimens preserved on slabs is the certainty that all skeletal elements come from the same individual (species). This is not only helpful in identifying fossil specimens, but can also be helpful in comparative, ecomorphological and phylogenetic analyses.
Typical habitats of modern dabbling ducks, in the ecological sense of the word, are shallow freshwater and estuaries. Compared to diving ducks, dabblers have relatively larger wings with a more elliptical shape, which enables them to rocket upwards out of the water and land even on small bodies of water that are spatially broken up by surrounding or emergent vegetation (Raikow, 1973). In the middle Miocene in Lavanttal, lowland wetland forests covered the shores of lakes, rivers, streams, marshes, and floodplains, with a warm and humid climate in the basin and dryer environmental conditions in the surrounding lowlands, highlands and mountains (Grímsson et al, 2015, 2020). The paleogeographic and ecological setting must have been a suitable environment for Lavanttalornis hassleri.
The remains of three birds were found at this site, all very well preserved. In addition to the duck described here, an almost complete passerine bird and a perfectly preserved sternum of another passerine were also found there (Happ et al., 2022). This may be a coincidence but it may also indicate a particular form of Lagerstaette. There is also the question of whether the birds died as a result of a single event and were buried at the same time, or whether they were deposited at different intervals and why, then, they were in the same place and in such a good state of preservation. The site was closed for environmental reasons shortly after these discoveries, so no more bird fossils have been found. But it will be reopened in the near future, which gives hope for further discoveries.
ACKNOWLEDGEMENTS
We are grateful to D. Sobieraj (Poland) for processing the photographs, and to N. Zelenkov (Russia) for providing us with unpublished photos of fossil specimens from his collection. We also appreciate the comments of N. Zelenkov and another anonymous referee whose reviews improved the manuscript.
REFERENCES
Baumel, J.J. and Witmer, L.M. 1993. Osteologia, p. 45–132. In Baumel, J.J., King, A.S., Breazile, J.E., Evans, H.E., and Vanden Berge, J.C. (eds.), Handbook of avian anatomy: Nomina anatomica avium. Publications of the Nuttall Ornithological Club, No 23. Nuttal Ornithological Club, Cambridge, Massachusetts.
Bienkowska-Wasiluk, M. 2010. Taphonomy of Oligocene teleost fishes from the Outer Carpathians of Poland. Acta Geologica Polonica, 60:479–533.
Bochenski, Z.M. and Tomek, T. 2009. A key for the identification of domestic birds bones in Europe: preliminary determination. Institute of Systematics and Evolution of Animals, Kraków, Poland.
Boev, Z. 2007. Neogene avifaunas of Bulgaria (a brief review), p. 26–35. In Bakardjieva, N., Chankova, St., Krastanov, B., and Gateva, Sv. (eds.), Evolution and Ecology-2007. Union of the Scientists of Bulgaria, Sophia.
Boev, Z. 2020. A little dabbling duck (Anatini Vigors, 1825 - Anseriformes Wagler, 1831) from the Late Miocene of Kremikovtsi (Bulgaria). Bulletin of the Natural History Museum - Plovdiv, 5:25–31.
Canterbury Museum. 2020. Mionetta blanchardi humerus, 3D model available from Sketchfab at https://skfb.ly/6VUYs. Accessed 5 December 2023.
Cheneval, J. 1983. Les Anatidae (Aves, Anseriformes) du Gisement aquitanien de Saint-Gerand-Le-Puy (Allier, France), p.525-538. In Buffetaut, E., Mazin, J.M., and Salmon, E. (eds.), Actes du symposium paléontologique G. Cuvier. Montbeliard.
Cheneval, J. 1987. Les Anatidae (Aves, Anseriformes) du Miocène de France. Révision systématique et évolution. Travaux et Documents des Laboratoires de Géologie de Lyon, 99:137–157.
Dickinson, E.C. and Remsen, Jr., J.V. 2013. The Howard and Moore complete checklist of the birds of the world, Volume 1, Aves Press. Eastbourne, UK.
Gál, E., Hír, J., Kessler, E., Kókay, J., and Márton, V. 2000. Middle Miocene fossils from the section of the road at the Rákóczi Chapel, Mátraszőlős. II. Locality Mátraszőlős 2. Folia Historico Naturalia Musei Matraensis, 24:39–75. [In Hungarian with English summary]
Göhlich, U.B. 2002. The Avifauna of the Miocene Fossil-Lagerstätte Sandelzhausen (Bavaria, Southern Germany). Zitteliana, 22:169–190.
Göhlich, U.B. 2009. The early Vallesian vertebrates of Atzelsdorf (late Miocene, Austria). Annalen des Naturhistorischen Museums in Wien, 111A:509–514.
Gorobets, L.V. 2013. Remains of Miocene birds from the Olkhon Island in the collections of the National Science and Natural History Museum of the National Academy of Sciences of Ukraine, p. 68–73. In Kalyakin, M.V., Zelenkov, N.V., and Gavrilov, V.M. (eds.), Problemy evolyutsii ptits: sistematika, morfologiya, ekologiya i povedenie (Avian evolution: systematics, morphology, ecology, and behavior). KMK Scientific Press Ltd., Moscow. [In Russian]
Grímsson, F., Meller, B., Bouchal, J. M., and Zetter, R. 2015. Combined LM and SEM study of the middle Miocene (Sarmatian) palynoflora from the Lavanttal Basin, Austria: part III. Magnoliophyta 1-Magnoliales to Fabales. Grana, 54(2):85–128.
https://doi.org/10.1080/00173134.2015.1007081
Grímsson, F., Bouchal, J.M., Xafis, A., and Zetter, R. 2020. Combined LM and SEM study of the middle Miocene (Sarmatian) palynoflora from the Lavanttal Basin, Austria: Part V. Magnoliophyta 3-Myrtales to Ericales. Grana, 59(2-3):127–193.
https://doi.org/10.1080/00173134.2019.1696400
Happ, J., Elsler, A., Kriwet, J., Pfaff, C., and Bochenski, Z.M. 2022. Two passeriform birds (Aves: Passeriformes) from the middle Miocene of Austria. PalZ, 96:313–321.
https://doi.org/10.1007/s12542-021-00579-2
Hou, L.H. 1985. Upper Miocene birds from Lufeng, Yunnan. Acta Anthropologica Sinica, 4:118–126.
Howard, H. 1929. The avifauna of Emeryville Shellmound. University of California Publications in Zoology, 32:301–394.
Howard, H. 1964. Fossil Anseriformes, p. 233–326. In Delacour J. The Waterfowl of the World, volume 4, Country Life Limited, London.
Kessler, E. 2013. A Kárpát medence madárvilágának őslénytani kézikönyv [Paleontological handbook of birdlife in the Carpathian Basin]. Könyvmühel, Miskolc.
Kessler, E. and Venczel, M. 2009. Bird remains from the middle Miocene of Subpiatră. Nymphaea. Folia naturae Bihariae, 36:27–36.
Kurochkin, E.N. 1976. New data on Pliocene birds of Western Mongolia. Transactions of the Joint Soviet-Mongolian Palaeontological Expedition, 3:51–67. (In Russian)
Kurochkin, E.N. 1985. Birds of Central Asia in the Pliocene. Transactions of the Joint Soviet-Mongolian Palaeontological Expedition, 26:1-119. (In Russian)
Lambrecht, K. 1933. Handbuch der Palaeornithologie. Borntraeger, Berlin.
Leach, W.E. 1819. Eleventh room, p. 63–68. In British Museum. Synopsis of the contents of the British Museum (15th ed.). Richard & Arthur Taylor.
Lebedinsky, N.G. 1927. Romainvillia stehlini n.g. n.sp., canard éocène provenant des marnes blanches du Bassin de Paris. Mémoires de la Société Paléontologique Suisse, 17:1–8.
Linnaeus, C. 1758. Systema Naturae per Regna Tria Naturae (10th ed.), volume 1: Regnum Animale. Salvii, L. Holmiae.
Livezey, B.C. and Martin, L.D. 1988. The Systematic Position of the Miocene Anatid Anas [?] blanchardi Milne-Edwards. Journal of Vertebrate Paleontology, 8:196–211.
https://doi.org/10.1080/02724634.1988.10011698
Mayr, G. 2008. Phylogenetic affinities and morphology of the late Eocene anseriform bird Romainvillia stehlini Lebedinsky, 1927. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, 248:365–380.
https://doi.org/10.1127/0077-7749/2008/0248-0365
Mayr, G. 2022. Paleogene fossil birds. Springer, Berlin.
Mayr, G. and De Pietri, V.L. 2013. A goose-sized anseriform bird from the late Oligocene of France: the youngest record and largest species of Romainvilliinae. Paläontologische Zeitschrift, 87:423–430.
https://doi.org/10.1007/s12542-013-0165-5
Mayr, G. and Pavia, M. 2014. On the true affinities of Chenornis graculoides Portis, 1884, and Anas lignitifila Portis, 1884–an albatross and an unusual duck from the Miocene of Italy. Journal of Vertebrate Paleontology, 34:914–923.
https://doi.org/10.1080/02724634.2013.821076
Mayr, G. and Smith, R. 2001. Ducks, rails, and limicoline waders (Aves: Anseriformes, Gruiformes, Charadriiformes) from the lowermost Oligocene of Belgium. Geobios, 34:547–561.
https://doi.org/10.1016/S0016-6995(01)80069-3
Mayr, G., Lechner, T., and Böhme, M. 2022. Nearly complete leg of an unusual, shelduck-sized anseriform bird from the earliest late Miocene hominid locality Hammerschmiede (Germany). Historical Biology, 4:465–474.
https://doi.org/10.1080/08912963.2022.2045285
McCracken, K.G., Harshman, J., McClellan, D.A., and Afton, A.D. 1999. Data set incongruence and correlated character evolution: an example of functional convergence in the hind-limbs of stifftail diving ducks. Systematic Biology, 48:683–714.
https://doi.org/10.1080/106351599259979
Milne-Edwards, A. 1863. Mémoire sur la distribution géologique des oiseaux fossiles et description de quelques espècies nouvelles. Annales des Sciences Naturelles, 4:133–176.
Milne-Edwards, A. 1867-1868. Recherches anatomiques et paléontologiques pour servir à l’histoire des oiseaux fossiles de la France. Éditions Masson, Paris.
Mlíkovský, J. 1991. Late Miocene birds of Götzendorf/Leitha, Austria. Annalen des Naturhistorischen Museums in Wien (A), 92:97–100.
Mlíkovský, J. 1998. Vertebrates from the Early Miocene lignite deposits of the Open-Cast Mine Oberdorf (Western Styrian basin, Austria). 4. Aves. Annalen des Naturhistorischen Museums in Wien (A), 99:39–42.
Mlíkovský, J. 2002. Cenozoic Birds of the World, volume 1. Europe: Ninox Press, Praha.
Nolf, D. and Brzobohatý, R. 2009. Lower Badenian fish otoliths of the Styrian and Lavanttal basins, with a revision of Weinfurter’s type material. Annalen des Naturhistorischen Museums in Wien (A), 111:323–356.
Nudds, R.L., Dyke, G.J., and Rayner, J.M.V. 2007. Avian brachial index and wing kinematics: putting movement back into bones. Journal of Zoology, 272(2):218–226.
https://doi.org/10.1111/j.1469-7998.2006.00261.x
Portis, A. 1884. Contribuzioni alla ornitolitologia italiana. Memorie della Reale Accademia delle Scienze di Torino, 36:361–384.
Prieto, J., Hassler, A., and Gross, M. 2019. Ein seltenes Fossil eines Stachelbilches aus Kärnten. Carinthia (II), 209/129:581–586.
Raikow, R.J. 1970. Evolution of diving adaptations in the stifftail ducks. University of California Press, Berkeley, Los Angeles, London.
Raikow, R.J. 1971. The osteology and taxonomic position of the White-backed Duck, Thalassornis leuconotus. The Wilson Bulletin, 83:270–277.
Raikow, R.J. 1973. Locomotor mechanisms in North American ducks. The Wilson Bulletin, 85:295–307.
Reischenbacher, D., Rifelj, H., Sachsenhofer, R.F., Jelen, B., Coric, S., Gross, M. and Reischenbacher, B. 2007. Early Badenian paleoenvironment in the Lavanttal Basin (Mühldorf Formation; Austria): Evidence from geochemistry and paleontology. Austrian Journal of Earth Sciences, 100:202–229.
Schäfer, W. 1972. Ecology and palaecology of marine environments. The University of Chicago Press, Chicago.
Stidham, T.A. 2015. Re-description and phylogenetic assessment of the Late Miocene ducks Aythya shihuibas and Anas sp. (Aves: Anseriformes) from Lufeng, Yunnan, China. Vertebrata PalAsiatica, 53:335–349.
http://www.vertpala.ac.cn/EN/Y2015/V53/I4/335
Švec, P. 1981. Lower Miocene Birds from Dolnice (Cheb Basin), Western Bohemia: Part 2, Časopis pro Mineralogii a Geologii, 26:45–56.
Tambussi, C.P., Degrange, F.J., and de Mendoza, R.S. 2023. “The present state of knowledge of the Cenozoic birds of Argentina" by Tonni 1980: four decades after. Publicación Electrónica de la Asociación Paleontológica Argentina, 23(1):255–295.
https://doi.org/10.5710/PEAPA.13.08.2022.418
Wagler, J.G. 1831. Einige Mitteilungen über Thiere Mexicos. Isis Von Oken 5:510–534.
Wank, M. 1991. Die Fossilien von Schönweg bei St. Andrä im Lavanttal (Kärnten). Carinthia (II) 181/101:301–313.
Worthy, T.H. 2009. Descriptions and phylogenetic relationships of two new genera and four new species of Oligo-Miocene waterfowl (Aves: Anatidae) from Australia. Zoological Journal of the Linnean Society, 156:411–454.
https://doi.org/10.1111/j.1096-3642.2008.00483.x
Worthy, T.H. and Lee, M.S. 2008. Affinities of Miocene waterfowl (Anatidae: Manuherikia, Dunstanetta and Miotadorna) from the St Bathans Fauna, New Zealand. Palaeontology, 51:677–708.
http://doi.org/10.1111/j.1475-4983.2008.00778.x
Worthy, T.H., Tennyson, A.J., Jones, C., McNamara, J.A., and Douglas, B.J. 2007. Miocene waterfowl and other birds from Central Otago, New Zealand. Journal of Systematic Palaeontology, 5:1–39.
https://doi.org/10.1017/S1477201906001957
Worthy, T.H., Tennyson, A.J., Hand, S.J., and Scofield, R.P. 2008. A new species of the diving duck Manuherikia and evidence for geese (Aves: Anatidae: Anserinae) in the St Bathans Fauna (Early Miocene), New Zealand. Journal of the Royal Society of New Zealand, 38:97–114.
https://doi.org/10.1080/03014220809510549
Worthy, T.H., Scofield, R.P., Salisbury, S.W., Hand, S.J., De Pietri, V.L., Blokland, J.C., and Archer, M. 2022. A new species of Manuherikia (Aves: Anatidae) provides evidence of faunal turnover in the St Bathans fauna, New Zealand. Geobios, 70:87–107.
https://doi.org/10.1016/j.geobios.2021.08.002
Zelenkov, N.V. 2011. Diving ducks from the middle Miocene of western Mongolia. Paleontological Journal, 45:191–199.
https://doi.org/10.1134/S0031030111020195
Zelenkov, N.V. 2012a. Neogene geese and ducks (Aves: Anatidae) from localities of the Great Lakes Depression, western Mongolia. Paleontological Journal, 46:607–619.
https://doi.org/10.1134/S0031030112060123
Zelenkov, N.V. 2012b. A new duck from the middle Miocene of Mongolia, with comments on Miocene evolution of ducks. Paleontological Journal, 46:520–530.
https://doi.org/10.1134/S0031030112050139
Zelenkov, N.V. 2016. Revision of non-Passeriform birds from Polgárdi (Hungary, Late Miocene): 1. Anseriformes. Paleontological Journal, 50:514–517.
https://doi.org/10.1134/S0031030116050142
Zelenkov, N.V. 2017. Finds of fragmentary bird skeletons in the Middle Miocene of the northern Caucasus. Doklady Biological Sciences, 477:223–226.
https://doi.org/10.1134/S0012496617060072
Zelenkov, N.V. 2018. The earliest Asian duck (Anseriformes: Romainvillia) and the origin of Anatidae. Doklady Biological Sciences, 483:225–227.
https://doi.org/10.1134/S0012496618060030
Zelenkov, N.V. 2020. Cenozoic evolution of Eurasian anatids (Aves: Anatidae s. l.). Biology Bulletin Reviews, 10:417–426.
https://doi.org/10.1134/S2079086420050096
Zelenkov, N.V. 2023a. Small ducks (Aves: Anatidae) from the Early-Middle Miocene of Eurasia. 1. A revision of Anas velox Milne-Edwards, 1868 and Anas soporata Kurochkin, 1976. Paleontological Journal, 57(4):452–462.
https://doi.org/10.1134/S0031030123040159
Zelenkov, N.V. 2023b. Small ducks (Aves: Anatidae) from the Early-Middle Miocene of Eurasia. Part 2. The fauna of Tagay locality (Baikal Region; eastern Siberia). Paleontological Journal, 57(5):560–572.
https://doi.org/10.1134/S003103012305012X
Zelenkov, N.V. and Kurochkin, E.N. 2012. Dabbling ducks (Aves: Anatidae) from the middle Miocene of Mongolia. Paleontological Journal, 46:421–429.
https://doi.org/10.1134/S0031030112040132
Zelenkov, N.V. and Panteleyev, A.V. 2015. Three bird taxa (Aves: Anatidae, Phasianidae, Scolopacidae) from the Late Miocene of the Sea of Azov (Southwestern Russia), Paläontologische Zeitschrift, 89:515–527.
https://doi.org/10.1007/s12542-014-0238-0
Zelenkov, N.V., Stidham, T.A., Martynovich, N.V., Volkova, N.V., Li, Q., and Qiu, Z. 2018. The middle Miocene duck Chenoanas (Aves, Anatidae): new species, phylogeny and geographical range. Papers in Palaeontology, 4:309–326.
https://doi.org/10.1002/spp2.1107
