

 A fragmentary leptonectid ichthyosaurian from the lower Pliensbachian of Luxembourg
A fragmentary leptonectid ichthyosaurian from the lower Pliensbachian of Luxembourg
Article number: 25.2.a24
https://doi.org/10.26879/1205
Copyright Palaeontological Association, August 2022
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Supplementary Information
Submission: 5 January 2022. Acceptance: 2 August 2022.
ABSTRACT
Despite abundant fossils, the quality of the fossil record of Early Jurassic marine reptiles strongly fluctuates with time and space. Pliensbachian strata have yielded very few marine reptile remains, especially outside of England, obscuring the evolution of marine reptiles during the middle part of the Early Jurassic. We report a new Pliensbachian locality from Luxembourg that contains abundant marine fauna and ichthyosaurian remains likely representing a single individual, composed of a partial snout, a possible surangular, two centra, and several ribs and gastralia. Ammonites and belemnites place this locality within the Valdani-Luridum Ammonite subzones of the Ibex Ammonite Zone, lower Pliensbachian. We assign the new ichthyosaur specimen to the clade Leptonectidae, using a combination of features from the snout and teeth. This specimen indicates that large neoichthyosaurians were present in multiple places of the European archipelago in all stages of the Early Jurassic and suggests that the ichthyosaurian faunae of western Europe remained essentially similar across the Sinemurian-Pliensbachian interval.
Valentin Fischer. Evolution & Diversity Dynamics Lab, UR Geology, Université de Liège, Belgium (corresponding author). v.fischer@uliege.be
Antoine Laboury. Evolution & Diversity Dynamics Lab, UR Geology, Université de Liège, Belgium. a.laboury@uliege.be
Kamil Bernacki. Department of Palaeontology, Natural History Museum Luxembourg, Luxembourg, Grand-Duchy of Luxembourg. kamil_bernacki@yahoo.com
Laurent Garbay. Department of Palaeontology, Natural History Museum Luxembourg, Luxembourg, Grand-Duchy of Luxembourg. garblaur@gmail.com
Yan Gillen. Department of Palaeontology, Natural History Museum Luxembourg, Luxembourg, Grand-Duchy of Luxembourg. gillen.yan@gmail.com
Charel Rollinger. Department of Palaeontology, Natural History Museum Luxembourg, Luxembourg, Grand-Duchy of Luxembourg. Charel.ROLLINGER@mnhn.lu
Anjin Thill. Department of Palaeontology, Natural History Museum Luxembourg, Luxembourg, Grand-Duchy of Luxembourg. anjin@caspersclimbingshop.com
Robert Weis. Department of Palaeontology, Natural History Museum Luxembourg, Luxembourg, Grand-Duchy of Luxembourg. RWEIS@mnhn.lu
Ben Thuy. Department of Palaeontology, Natural History Museum Luxembourg, Luxembourg, Grand-Duchy of Luxembourg. Ben.THUY@mnhn.lu
Keywords: Neoichthyosauria; Leptonectidae; Early Jurassic; Luxembourg-Trier
Final citation: Fischer, Valentin, Laboury, Antoine, Bernacki, Kamil, Garbay, Laurent, Gillen, Yan, Rollinger, Charel, Thill, Anjin, Weis, Robert, and Thuy, Ben. 2022. A fragmentary leptonectid ichthyosaurian from the lower Pliensbachian of Luxembourg. Palaeontologia Electronica, 25(2):a24. https://doi.org/10.26879/1205
palaeo-electronica.org/content/2022/3685-pliensbachian-leptonectid
Copyright: August 2022 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
Mesozoic marine reptiles have an abundant fossil record, documenting the beginning of their radiation during the earliest Triassic (Motani et al., 2015, 2017) to centimetres below the Cretaceous-Palaeogene boundary horizon (Jouve et al., 2008; Gallagher et al., 2012). However, the quality of their fossil record strongly fluctuates with time and space (Benson et al., 2010; Benson and Butler, 2011; Butler et al., 2011), and possibly less so with phylogeny (Tutin and Butler, 2017). This fluctuation appears fractal, being also present within Early Jurassic ichthyosaurians: the early Toarcian shales have provided thousands of fossils in western Europe, with most specimens from Germany and England (Hauff, 1953; Godefroit, 1994; Benson et al., 2010), while the Pliensbachian and late Toarcian assemblages are extremely poorly sampled (Bardet et al., 2008; Fernández et al., 2018; Lomax and Massare, 2018a; Fischer et al., 2021). Pliensbachian ichthyosaurian faunas generally resemble those from the Sinemurian, with the presence of Leptonectes (McGowan and Milner, 1999; Fernández et al., 2018; Lomax and Massare, 2018a), Ichthyosaurus (Lomax, 2010; Lomax and Massare, 2015; Massare and Lomax, 2016), and Temnodontosaurus (Huene, 1931a; Hungerbühler and Sachs, 1996; Maisch and Hungerbühler, 1997), but also Hauffiopteryx (Maisch and Reisdorf, 2006; Maxwell and Cortés, 2020). However, the vast majority of Pliensbachian ichthyosaurians come from England and Germany; one (Maisch and Reisdorf, 2006) is from Switzerland and another (Fernández et al., 2018) is from Spain. In this context, documenting new Pliensbachian occurrences of ichthyosaurians is important to better understand how marine predator faunas changed throughout the entire Early Jurassic. In this paper, we describe the likely associated remains of a large ichthyosaur showing leptonectid affinities from a new lower Pliensbachian locality in central Luxembourg.
MATERIAL AND METHODS
Institutional Abbreviations
BRSMG: Bristol City Museum and Art Gallery, Bristol, UK. MHNH: Muséum d’Histoire Naturelle du Havre, Le Havre, France. MNHN: Muséum National d’Histoire Naturelle, Paris, France. MNHNL: Muséum National d’Histoire Naturelle du Luxembourg, Luxembourg-ville, Luxembourg. NHMUK: Natural History Museum, London, UK. OUM: Oxford University Museum, Oxford, UK.
The Cloche d’Or Locality
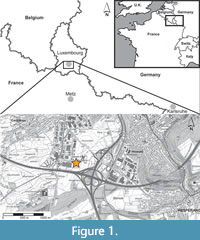 The remains described herein consist of a series of articulated and disarticulated remains of a large ichthyosaurian, MNHNL LM266 from the Cloche d’Or locality, in the Gasperich quarter in southern Luxembourg City, central Luxembourg (Figure 1). The outcrop at Cloche d’Or is situated along the banks of a recently re-natured Weierbaach creek. The first fragments were found during preliminary prospections in April 2021, and most of the remains were collected during controlled excavations and prospections lead by several of us (B.T., R.W., K.B., L.G., C.R., A.T., and Y.G.) in July 2021 (Figure 2), complemented by subsequent finds cropping out after the millennial floods of summer 2021.
The remains described herein consist of a series of articulated and disarticulated remains of a large ichthyosaurian, MNHNL LM266 from the Cloche d’Or locality, in the Gasperich quarter in southern Luxembourg City, central Luxembourg (Figure 1). The outcrop at Cloche d’Or is situated along the banks of a recently re-natured Weierbaach creek. The first fragments were found during preliminary prospections in April 2021, and most of the remains were collected during controlled excavations and prospections lead by several of us (B.T., R.W., K.B., L.G., C.R., A.T., and Y.G.) in July 2021 (Figure 2), complemented by subsequent finds cropping out after the millennial floods of summer 2021.
Lithostratigraphy of Cloche d’Or
Along the Weierbaach creek, two lithostratigraphic formations, as defined by the Geological map of Luxembourg, are exposed. The lower one, the Li4 (“Marnes pauvres en fossiles”) unit, consists of blue-greyish marls with occasional micritic nodules, and yields ammonites, bivalves (mostly Gryphaea macculocchi, as well as cardiniids and pectinids), brachiopods (notably Spiriferina, Zeilleria), crinoids (Pentacrinites sp.), and small passaloteuthid belemnites (Nannobelus oppeli). The Li4 succession corresponds to our units I and II (Figure 3). The lithological transition towards the overlying formation Lm1 is not clearly visible, and approximately corresponds to unit III or the basal part of unit IV. The upper part of unit IV as well as the overlying units V-VII belong to the Lm1 (“Calcaire ocreux”) formation, which consists here of sandy, yellow marls yielding numerous ammonites, echinoderm remains, and a more diversified belemnite fauna than the underlying Li4 formation. Usually, this formation consists of three members, a lower calcareous member (“Raricostaten-Schichten”, roughly corresponding to the uppermost Sinemurian), a middle marly member (“Numismalis-Mergel”, corresponding to the lower Pliensbachian) and an upper calcareous member (“Davoei-Kalk”, also lower Pliensbachian) (Lucius, 1948; Maubeuge, 1950). The yellow marls at Cloche d’Or are interpreted here as an equivalent of the middle marly member of the Calcaire ocreux formation. This means that the lower calcareous member is missing at Cloche d’Or. A similar hiatus at the base of the formation had previously been observed for some localities in Luxembourg (Lucius, 1948; Maubeuge, 1950; Maubeuge and Antun, 1967). A more complete succession of the Li4 and Lm1 formations has been recorded in other localities, for instance Capellen (Guérin-Franiatte, 2003), to the west of Luxembourg-City. The ichthyosaur remains described in the present paper were found in unit V, corresponding to the middle marly member of the Lm1 formation.
(Nannobelus oppeli). The Li4 succession corresponds to our units I and II (Figure 3). The lithological transition towards the overlying formation Lm1 is not clearly visible, and approximately corresponds to unit III or the basal part of unit IV. The upper part of unit IV as well as the overlying units V-VII belong to the Lm1 (“Calcaire ocreux”) formation, which consists here of sandy, yellow marls yielding numerous ammonites, echinoderm remains, and a more diversified belemnite fauna than the underlying Li4 formation. Usually, this formation consists of three members, a lower calcareous member (“Raricostaten-Schichten”, roughly corresponding to the uppermost Sinemurian), a middle marly member (“Numismalis-Mergel”, corresponding to the lower Pliensbachian) and an upper calcareous member (“Davoei-Kalk”, also lower Pliensbachian) (Lucius, 1948; Maubeuge, 1950). The yellow marls at Cloche d’Or are interpreted here as an equivalent of the middle marly member of the Calcaire ocreux formation. This means that the lower calcareous member is missing at Cloche d’Or. A similar hiatus at the base of the formation had previously been observed for some localities in Luxembourg (Lucius, 1948; Maubeuge, 1950; Maubeuge and Antun, 1967). A more complete succession of the Li4 and Lm1 formations has been recorded in other localities, for instance Capellen (Guérin-Franiatte, 2003), to the west of Luxembourg-City. The ichthyosaur remains described in the present paper were found in unit V, corresponding to the middle marly member of the Lm1 formation.
Biostratigraphy of Cloche d’Or
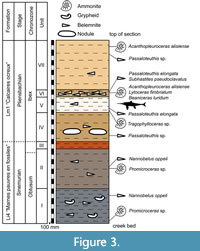 The biostratigraphic interpretation of the exposed succession is based on ammonites (Figure 3). Two biozones are recognised: the Obtusum Zone of the upper Sinemurian and the Ibex Zone of the lower Pliensbachian. A characteristic association has been identified in unit VI: Lytoceras fimbriatum (Sowerby), Acanthopleuroceras alisiense (Reynès in Haug), and Beaniceras luridum (Simpson), an association corresponding to the upper part of the Ibex Zone (Valdani-Luridum subzones) (Christian Meister, personal communication). Belemnites retrieved from the succession corroborate and, in part, complement the ammonite-based zonation, although the biostratigraphic value of the belemnites is less precise compared to the ammonites (Riegraf, 1980; Combémorel, 1997; Schlegelmilch, 1998). Belemnites of the genus Nannobelus, in particular Nannobelus oppeli, found in units I and II, indicate an upper Sinemurian age. Belemnites of the genus Passaloteuthis, known from the lower Plienbachian onwards, were found in units IV to VII. Together with the ammonite association from unit VI, the belemnite evidence thus suggests that the ichthyosaur-bearing bed V is of lower Pliensbachian age. Multiple fragmentary specimens of the ammonite Tragophylloceras further corroborates the lower Pliensbachian age (Ibex Zone) of the ichthyosaurian specimen MNHNL LM266.
The biostratigraphic interpretation of the exposed succession is based on ammonites (Figure 3). Two biozones are recognised: the Obtusum Zone of the upper Sinemurian and the Ibex Zone of the lower Pliensbachian. A characteristic association has been identified in unit VI: Lytoceras fimbriatum (Sowerby), Acanthopleuroceras alisiense (Reynès in Haug), and Beaniceras luridum (Simpson), an association corresponding to the upper part of the Ibex Zone (Valdani-Luridum subzones) (Christian Meister, personal communication). Belemnites retrieved from the succession corroborate and, in part, complement the ammonite-based zonation, although the biostratigraphic value of the belemnites is less precise compared to the ammonites (Riegraf, 1980; Combémorel, 1997; Schlegelmilch, 1998). Belemnites of the genus Nannobelus, in particular Nannobelus oppeli, found in units I and II, indicate an upper Sinemurian age. Belemnites of the genus Passaloteuthis, known from the lower Plienbachian onwards, were found in units IV to VII. Together with the ammonite association from unit VI, the belemnite evidence thus suggests that the ichthyosaur-bearing bed V is of lower Pliensbachian age. Multiple fragmentary specimens of the ammonite Tragophylloceras further corroborates the lower Pliensbachian age (Ibex Zone) of the ichthyosaurian specimen MNHNL LM266.
Taphonomy
The ichthyosaurian remains described in the present paper were either collected directly in bed V or picked from the weathered surface below and in the creek downstream. In some cases, fragments collected from the stream bed could be associated with fragments directly retrieved from bed V (in particular the two fragments of a possible surangular), corroborating that all the remains, in fact, originate from the same bed. Unit V consists of conspicuously bright-coloured clayey marl, readily distinguishable from the over- and underlying strata. Furthermore, unit V has yielded an extraordinary microfauna, including numerous minute spines of the echinoid Cuspidentechinus Smith, 2016 and cyrtocrinid remains that are absent from the under- and overlying strata. Thanks to the distinctive colour and microfauna of the adhering matrix, even bone fragments that were found downslope or in the creek could be unambiguously assigned to unit V.
The marls of unit V show no signs of re-sedimentation, reworking, or condensation, neither in macroscopic terms (e.g., reworked ammonites or belemnites, intraclasts, encrusted pebbles) nor in the microfaunal spectrum. Furthermore, the bones show no evidence of encrusting organisms or adhering matrix other than the marls from unit V. We therefore exclude the possibility of reworking or condensation to explain the co-occurrence of the bone fragments. While we cannot rule out that the bones belong to more than one individual, continuously controlled excavation allowed us to confirm the single origin of the bone fragments. All the remains can be traced down to a restricted area within bed V, approximately 5 m wide, on both sides of the creek.
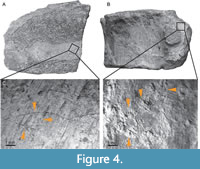 The bone tissue is well preserved in most cases. Interestingly, some fragments, in particular the vertebra and the possible surangular, show millimetric, radially oriented scratch marks (Figure 4). The size, depth, orientation, and arrangement of the scratch marks are typical features of Gnathichnus, an ichnogenus attributed to the grazing activity of regular echinoids (Bromley, 1975). Gnathichnus is usually found on invertebrate shells or skeletal parts (e.g., Breton et al., 1992; Carrasco, 2003) but also on vertebrate bones (Meyer, 2011; Reolid et al., 2015), including on ichthyosaur remains (Danise et al., 2014). The sediment surrounding the bone fragments yields abundant spines, test plates, and masticatory apparatus components of a small regular echinoid with teeth of trapezoidal cross-section, reminiscent of the common and widespread Early Jurassic echinoid genus Cunidentechinus, which therefore qualifies as a possible producer of the grazing traces on the bones.
The bone tissue is well preserved in most cases. Interestingly, some fragments, in particular the vertebra and the possible surangular, show millimetric, radially oriented scratch marks (Figure 4). The size, depth, orientation, and arrangement of the scratch marks are typical features of Gnathichnus, an ichnogenus attributed to the grazing activity of regular echinoids (Bromley, 1975). Gnathichnus is usually found on invertebrate shells or skeletal parts (e.g., Breton et al., 1992; Carrasco, 2003) but also on vertebrate bones (Meyer, 2011; Reolid et al., 2015), including on ichthyosaur remains (Danise et al., 2014). The sediment surrounding the bone fragments yields abundant spines, test plates, and masticatory apparatus components of a small regular echinoid with teeth of trapezoidal cross-section, reminiscent of the common and widespread Early Jurassic echinoid genus Cunidentechinus, which therefore qualifies as a possible producer of the grazing traces on the bones.
SYSTEMATIC PALAEONTOLOGY
Order ICHTHYOSAURIA De Blainville, 1835
Clade PARVIPELVIA Motani, 1999
Clade NEOICHTHYOSAURIA Sander, 2000
Family LEPTONECTIDAE Maisch, 1998
Leptonectidae indet.
Referred material. MNHNL LM266, from the lower Pliensbachian (Valdani to Luridum Ammonite subzones of the Ibex Ammonite Zone) of the Cloche d’Or locality, central Luxembourg.
Snout
 Two contiguous fragments of a rostrum are preserved over 51 mm, containing articulated premaxillae, dentaries, splenials, and teeth (Figure 5). The premaxilla and the dentary are slender, being as dorsoventrally high as the apicobasal height of the corresponding tooth crowns. The premaxillary and dentary fossae are present, in the form of a series of elongated foramina that are 13 mm long and 3 mm high for the premaxillary fossa and slightly longer for the dentary fossa. The anterior part of the splenial is preserved in articulation with the dentary and has a slit-like ventral exposure. The splenial thus possesses a long ventral exposure, which is similar to that of Leptonectes moorei (V.F. pers. obs. holotype specimen NHMUK PV R 14370) and differs from the condition seen in the rather short-snouted Hauffiopteryx spp., where the nasal rapidly protrude from the dorsal margin of the rostrum, extending anteriorly as far (or further) than the splenial (Marek et al., 2015; Maxwell and Cortés, 2020). In MNHNL LM266, the nasal is not even present internally, indicating its dorsal exposure is de facto set more posteriorly than that of the splenial.
Two contiguous fragments of a rostrum are preserved over 51 mm, containing articulated premaxillae, dentaries, splenials, and teeth (Figure 5). The premaxilla and the dentary are slender, being as dorsoventrally high as the apicobasal height of the corresponding tooth crowns. The premaxillary and dentary fossae are present, in the form of a series of elongated foramina that are 13 mm long and 3 mm high for the premaxillary fossa and slightly longer for the dentary fossa. The anterior part of the splenial is preserved in articulation with the dentary and has a slit-like ventral exposure. The splenial thus possesses a long ventral exposure, which is similar to that of Leptonectes moorei (V.F. pers. obs. holotype specimen NHMUK PV R 14370) and differs from the condition seen in the rather short-snouted Hauffiopteryx spp., where the nasal rapidly protrude from the dorsal margin of the rostrum, extending anteriorly as far (or further) than the splenial (Marek et al., 2015; Maxwell and Cortés, 2020). In MNHNL LM266, the nasal is not even present internally, indicating its dorsal exposure is de facto set more posteriorly than that of the splenial.
The tooth crowns are slightly recurved and markedly elongated: the apicobasal height / basal diameter ratio is 11 mm / 4 mm = 2.75 (Figure 5, Figure 6), very similar to some species of Stenopterygius (Maxwell, Fernández, et al., 2012), Hauffiopteryx (Maxwell and Cortés, 2020), and in leptonectids such as most species of Leptonectes (Fraas, 1892; Huene, 1922; McGowan, 1989, 1993) (with the exception of Leptonectes moorei [McGowan and Milner, 1999]), Eurhinosaurus longirostris (Reisdorf et al., 2011), and Wahlisaurus massarae (Lomax, 2016) (Figure 6). This condition is clearly distinct from the stouter teeth seen in Temnodontosaurus spp. (Fraas, 1891; McGowan, 1974; Godefroit, 1993), Suevoleviathan spp. (Maxwell, 2018), Protoichthyosaurus (Lomax and Massare, 2018b; Lomax et al., 2019), and some species of Ichthyosaurus (Fraas, 1891; Maisch, 1997; Maisch et al., 2008). Another similarity with Stenopterygius, Hauffiopteryx, and leptonectids is the reduction of longitudinal striations along the crown (Maisch, 1998; Maxwell, 2012; Lomax, 2016; Fernández et al., 2018) (Figure 5, Figure 6). Yet, sparse striations are present in MNHNL LM266 but do not reach the apical quarter of the crown (as in Leptonectes spp.; Figure 6), and many crowns exhibit a single basal ring, as in Wahlisaurus massarae (Lomax, 2016), Leptonectes spp. (Figure 6), and Eurhinosaurus longirostris (in which more than one basal ring is usually present (e.g., Godefroit, 1994; Fischer et al., 2011)). The acellular cementum ring is not ridged, unlike in Temnodontosaurus (Godefroit, 1993; Maxwell, Caldwell, et al., 2012), Ichthyosaurus (McGowan, 1973; Vincent et al., 2014), and Protoichthyosaurus (Lomax et al., 2019). The root bears fine striations, but only in the basal half, as in some leptonectids (Fraas, 1892; Godefroit, 1994). This differs from the condition of Leptonectes moorei (Figure 6), Leptonectes solei (Figure 6), Stenopterygius (Godefroit, 1994), and Hauffiopteryx (Maxwell and Cortés, 2020), where the root ridges reach the base of the acellular cementum ring. The root is slightly expanded mesiodistally and slightly compressed labiolingually, another usual feature that is often present in leptonectids (Fraas, 1891; Reisdorf et al., 2011; Lomax, 2016).
(Reisdorf et al., 2011), and Wahlisaurus massarae (Lomax, 2016) (Figure 6). This condition is clearly distinct from the stouter teeth seen in Temnodontosaurus spp. (Fraas, 1891; McGowan, 1974; Godefroit, 1993), Suevoleviathan spp. (Maxwell, 2018), Protoichthyosaurus (Lomax and Massare, 2018b; Lomax et al., 2019), and some species of Ichthyosaurus (Fraas, 1891; Maisch, 1997; Maisch et al., 2008). Another similarity with Stenopterygius, Hauffiopteryx, and leptonectids is the reduction of longitudinal striations along the crown (Maisch, 1998; Maxwell, 2012; Lomax, 2016; Fernández et al., 2018) (Figure 5, Figure 6). Yet, sparse striations are present in MNHNL LM266 but do not reach the apical quarter of the crown (as in Leptonectes spp.; Figure 6), and many crowns exhibit a single basal ring, as in Wahlisaurus massarae (Lomax, 2016), Leptonectes spp. (Figure 6), and Eurhinosaurus longirostris (in which more than one basal ring is usually present (e.g., Godefroit, 1994; Fischer et al., 2011)). The acellular cementum ring is not ridged, unlike in Temnodontosaurus (Godefroit, 1993; Maxwell, Caldwell, et al., 2012), Ichthyosaurus (McGowan, 1973; Vincent et al., 2014), and Protoichthyosaurus (Lomax et al., 2019). The root bears fine striations, but only in the basal half, as in some leptonectids (Fraas, 1892; Godefroit, 1994). This differs from the condition of Leptonectes moorei (Figure 6), Leptonectes solei (Figure 6), Stenopterygius (Godefroit, 1994), and Hauffiopteryx (Maxwell and Cortés, 2020), where the root ridges reach the base of the acellular cementum ring. The root is slightly expanded mesiodistally and slightly compressed labiolingually, another usual feature that is often present in leptonectids (Fraas, 1891; Reisdorf et al., 2011; Lomax, 2016).
The small size of these rostral fragments can be interpreted in two ways: (i) it belongs to a small individual, distinct from the other remains from the site or (ii) it represents the anterior extremity of a long snouted ichthyosaurian, where the tip of the rostrum can be extremely small compared to the size of the animal (McGowan, 1993, 2003). The presence of a discontinuous premaxillary fossa and, to a lesser extent, the absence of both a nasal and a maxilla (even internally) suggests that this fragment was located from the anterior quarter of the rostrum. The exposure of the splenial is less indicative of a position within the rostrum, because at least one leptonectid (Leptonectes moorei) possesses a very long ventral splenial exposure (V.F. pers. obs. holotype specimen NHMUK PV R 14370). In the absence of other evidence and given the taphonomy, stratigraphy, and the controlled excavations, we regard all the ichthyosaur fragments described here as likely belonging to a single individual. Even so, we detail the taxonomic information present in all fragments in isolation (Table 1), and the conclusions of the paper do not rely on this association.
Possible Surangular
 Two contiguous fragments are interpreted here as the posterior part of a large right surangular (Figure 4, Supplementary Information Figure 1). The lateral surface is convex and forms a shallow anteroposterior ridge at mid-height. The medial surface is concave and forms a wide prominent ridge located within the ventral half of the medial surface. The ventral surface is rounded while the bone tapers dorsally to a thin, saddle-shaped ridge, as might be seen close to the coronoid process in neoichthyosaurians (Sollas, 1916; McGowan, 1973). The perfectly straight and parallel bone fibres on the concave side (Figure 4, Supplementary Information Figure 1) are features of ichthyosaurian surangulars (V.F. pers. obs. on material from MNHNL, MHNH). We do not derive taxonomic information from these fragments, because we consider their identification too tentative.
Two contiguous fragments are interpreted here as the posterior part of a large right surangular (Figure 4, Supplementary Information Figure 1). The lateral surface is convex and forms a shallow anteroposterior ridge at mid-height. The medial surface is concave and forms a wide prominent ridge located within the ventral half of the medial surface. The ventral surface is rounded while the bone tapers dorsally to a thin, saddle-shaped ridge, as might be seen close to the coronoid process in neoichthyosaurians (Sollas, 1916; McGowan, 1973). The perfectly straight and parallel bone fibres on the concave side (Figure 4, Supplementary Information Figure 1) are features of ichthyosaurian surangulars (V.F. pers. obs. on material from MNHNL, MHNH). We do not derive taxonomic information from these fragments, because we consider their identification too tentative.
Centra
Two centra fragments are present. One (Figure 4, Figure 5) is a large dorsal centrum with an anteroposterior length of 52 mm and a diameter certainly above 100 mm (as the preserved portion is 100 mm wide), and probably between 120 and 130 mm. One apophysis is visible; the fact that the dorsal part of the centrum is missing suggests that this apophysis is the parapophysis. This apophysis is >20 mm long anteroposteriorly and 15 mm high dorsoventrally. It is teardrop-shaped, with anterior ridge connecting to the anterior margin of the centrum, as is typically of parvipelvians and clearly distinct from the dorsoventrally elongated diapophyses and often minuscule parapophyses seen in shastasaurids (Merriam, 1902; Sander, 1997; Fischer et al., 2014). Ventrally, the centrum forms a slightly concave surface. We interpret this as a parapophysis, present on the lateral surface of the centrum at mid-height. Accordingly, this centrum is regarded as an anterior dorsal centrum. The large size and the aspect ratio match with the morphology of Temnodontosaurus spp. (McGowan, 1974; Godefroit, 1993; Martin et al., 2012; Swaby and Lomax, 2021) and Leptonectes solei (McGowan, 1993). However, most other leptonectids have smaller centra, with diameters ranging from 25 to 65 mm (Huene, 1951; McGowan and Milner, 1999; McGowan, 2003; Lomax, 2016).
A second fragmentary bone solely consists of the margin of the articular surface of a small centrum. The lateral or ventral surface is concave and lacks chevron facets or apophyses. It is interpreted as a posterior caudal centrum, given the small size.
Ribs and Gastralia
Several tens of rib fragments are preserved (Figure 5, Supplementary Information Figure 1). Most have wide anterior and posterior grooves, giving the rib an “8” shape in cross-section, as is usually the case in neoichthyosaurians. However, the dorsal ribs of Leptonectes tenuirostris, Temnodontosaurus azerguensis, and some species of Ichthyosaurus seem to lack the grooves, giving their ribs a rounded cross-section (Martin et al., 2012; Lomax and Massare, 2016). In one fragment, one of the grooves can be observed vanishing medially; the largest fragments have a single groove, and the opposite side is gently convex (Figure 5). A series of other, small fragments are straight and exhibit a rugose texture; we interpret these as fragmentary gastralia.
DISCUSSION
The presence of long and slender teeth, with feint longitudinal striations, a ring/annulus at the base of the crown, and with a slightly rectangular root, combined with a slender rostrum suggest leptonectid affinities for the rostral fragments of MNHNL LM266 (Figure 6). Within leptonectids, the absence of an overbite and the slender teeth rule out Eurhinosaurus longirostris, Excalibosaurus costini, and Leptonectes moorei (Huene, 1931b; McGowan, 1986, 2003; McGowan and Milner, 1999). The large size of the associated centra - if they, as we think is the case, belong to the same individual - suggests an animal of large size, comparable to Leptonectes solei, and multiple species within Temnodontosaurus, notably Temnodontosaurus platyodon and Temnodontosaurus trigonodon (Godefroit, 1993; McGowan, 1993, 1996). Leptonectes is known to be present in the European Archipelago from the Hettangian to the Pliensbachian, including in nearby Belgium (Godefroit, 1992; McGowan and Milner, 1999; Lomax and Massare, 2018a) (if one regards the Pre-Planorbis beds as earliest Hettangian in age, as suggested by Hillebrandt and Kystyn (2009)) and is thus a solid candidate for the remains from the Cloche d’Or locality. However, we assign these remains to Leptonectidae indet., pending discovery of cranial and appendicular material that could clarify the taxonomic relationships of this specimen. It should be stressed here that the presence of leptonectids in the Luxembourg-Trier basin (see also Godefroit, 1992) during the early Pliensbachian does not require the skeletal elements we describe here to origin from a single individual; only the presence of a large leptonectid does (Table 1).
Even if leptonectid neoichthyosaurians have been abundantly described (e.g., Fraas, 1891; Huene, 1922; McGowan, 1989, 1993; Godefroit, 1994; McGowan and Milner, 1999; Maisch and Matzke, 2003; Lomax, 2016), the details of the dental morphology of leptonectids are rarely documented, notably the concentric rings found towards the base of the tooth crown (but see Godefroit, 1994; Reisdorf et al., 2011; Lomax, 2016). These slightly concave rings, which we term here “annuli”, clearly differ from the similarly positioned “enamel bands” of pliosaurids, which have a positive relief (Zverkov et al., 2018). Because it is not well distributed among neoichthyosaurians, this feature can be useful to segregate leptonectids from other taxa with smoothed and elongated tooth crowns, such as Hauffiopteryx and Stenopterygius (Figure 6). Indeed, even if Hauffiopteryx spp., Stenopterygius spp., and leptonectids all share a reduction of longitudinal ridges on tooth crown (Reisdorf et al., 2011; Maxwell, Fernández, et al., 2012; Fernández et al., 2018; Maxwell and Cortés, 2020), the presence of a such annuli has not been reported yet in Hauffiopteryx and Stenopterygius. A slightly labio-lingually compressed root whose apicobasal ridges do not reach the acellular cementum ring, combined with the occurrence of one of more annuli close to the base of the crown can, therefore, at the current state of our knowledge, identify a tooth belonging to a leptonectid (Figure 6). Several of these rings have been reported on the crown of Eurhinosaurus (Godefroit, 1994; Fischer et al., 2011; Reisdorf et al., 2011) and might represent a diagnostic feature of this taxon.
The continued absence of baracromian ichthyosaurians in Pliensbachian strata suggests a sense of continuity in the ichthyosaurian assemblages during the Hettangian-Pliensbachian interval, which essentially (but not exclusively) contains leptonectids, Ichthyosaurus, and Temnodontosaurus (McGowan and Motani, 2003). A novelty of the Pliensbachian is to record the slightly more derived taxon Hauffiopteryx (Maxwell and Cortés, 2020). Thanks to the Cloche d’Or locality, all stages of the Early Jurassic are now known to contain large early neoichthyosaurians (with centra diameter > 10 cm) in many places of the European Archipelago (Godefroit, 1993; McGowan, 1996; Martin et al., 2012; Maxwell and Vincent, 2015; Fischer et al., 2021). By contrast, baracromians dominate ichthyosaurian assemblages in the European Archipelago by the early Toarcian with the ubiquitous presence of multiple species of Stenopterygius (Huene, 1931b; Hauff, 1953; Godefroit, 1994; Maisch, 2010; Maxwell, 2012; Dick et al., 2016) and are the only ichthyosaurians found after the Bajocian, with the exception of Malawania anachronus (Fernández, 1994; Fischer et al., 2013, 2021). The Pliensbachian gap in the fossil record of ichthyosaurians might contain this change of dominance; in any case, these remains further highlight the importance of the Luxembourg-Trier basin deposits in understanding the diversity dynamics of Jurassic marine reptiles.
CONCLUSIONS
Marine reptile remains are rare in Pliensbachian strata, worldwide. We describe remains of large ichthyosaurians from a new lower Pliensbachian locality in central Luxembourg (Ibex Ammonite zone). At least some of these remains belong to Leptonectidae, confirming the continuous presence of early neoichthyosaurians during the entire Hettangian-Pliensbachian interval in Western Europe.
ACKNOWLEDGEMENTS
We warmly thank Handling Editor Dr. M. Pole and our two reviewers, D. Cortés and an anonymous colleague, for making our paper better with thoughtful and constructive suggestions. The work of A.L. is supported by a FRIA grant of the F.R.S.-FNRS (FC38761).
REFERENCES
Bardet, N., Fernandez, M., Garcia-ramos, J.C., Pereda-Suberbiola, X., Pinuela, L., Ruiz-omenaca, J.I., and Vincent, P. 2008. A juvenile plesiosaur from the Pliensbachian (Lower Jurassic) of Asturias, Spain. Journal of Vertebrate Paleontology, 28:258-263. https://doi.org/10.1671/0272-4634(2008)28[258:AJPFTP]2.0.CO;2
Benson, R.B.J. and Butler, R.J. 2011. Uncovering the diversification history of marine tetrapods: ecology influences the effect of geological sampling biases, p. 191-208. In McGowan, A.J. and Smith, A.B. (eds.), Comparing the geological and fossil records: implications for biodiversity studies. Geological Society, Special Publications, 358. London.
Benson, R.B.J., Butler, R.J., Lindgren, J., and Smith, A.S. 2010. Mesozoic marine tetrapod diversity: mass extinctions and temporal heterogeneity in geological megabiases affecting the vertebrates. Proceedings of the Royal Society B: Biological Sciences, 277:829-834. https://doi.org/10.1098/rspb.2009.1845
Breton, G., Neraudeau, D., and Cuenca-Boulat, C. 1992. Gnathichnus stellarum ichnosp. nov., trace de broutage d’un echinide du Campanien des Charentes (France). Revue de Paléobiologie, 11:219-229.
Bromley, R.G. 1975. Comparative analysis of fossil and recent echinoid bioerosion. Palaeontology, 18:725-739.
Butler, R.J., Brusatte, S.L., Andres, B., and Benson, R.B.J. 2011. How do geological sampling biases affect studies of morphological evolution in deep time? A case study of pterosaur (Reptilia: Archosauria) disparity. Evolution, 66:147-162. https://doi.org/10.1111/j.1558-5646.2011.01415.x
Carrasco, J.F. 2003. Presencia de la icnoespecie Gnathichnus pentax sobre Dimya richei (Bivalvia) en el Eoceno inferior de La Puebla de Roda (Huesca). Batalleria, 11:37-40.
Combémorel, R. 1997. Bélemnites, p. 157-168. In Cariou, E. and Hantzpergue, P. (eds.), Biostratigraphie du Jurassique Ouest-Européen et Méditerranéen. Zonations parallèles et distribution des invertébrés et microfossiles. Bulletin du Centre de Recherches Elf Productions, Mémoire 17.
Danise, S., Twitchett, R.J., and Matts, K. 2014. Ecological succession of a Jurassic shallow-water ichthyosaur fall. Nature communications, 5:1-8. https://doi.org/10.1038/ncomms5789
de Blainville, H.M.D. 1835. Description de quelques espèces de reptiles de la Californie, précédée de l’analyse d’un système général d’Erpetologie et d’Amphibiologie. Nouvelles annales du Muséum d’Histoire naturelle, Paris, 4:233-296.
Dick, D.G., Schweigert, G., and Maxwell, E.E. 2016. Trophic niche ontogeny and palaeoecology of early Toarcian Stenopterygius (Reptilia: Ichthyosauria). Palaeontology, 59:423-431. https://doi.org/10.1111/pala.12232
Fernández, M. 1994. A new long-snouted ichthyosaur from the early Bajocian of Neuquén basin (Argentina). Ameghiniana, 31:291-297.
Fernández, M.S., Piñuela, L., and García-Ramos, J.C. 2018. First report of Leptonectes (Ichthyosauria: Leptonectidae) from the Lower Jurassic (Pliensbachian) of Asturias, northern Spain. Palaeontologia Electronica, 21.2.29A:1-15. https://doi.org/10.26879/802
Fischer, V., Guiomar, M., and Godefroit, P. 2011. New data on the palaeobiogeography of Early Jurassic marine reptiles: the Toarcian ichthyosaur fauna of the Vocontian Basin (SE France). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 261:111-127. https://doi.org/10.1127/0077-7749/2011/0155
Fischer, V., Appleby, R.M., Naish, D., Liston, J., Riding, J.B., Brindley, S., and Godefroit, P. 2013. A basal thunnosaurian from Iraq reveals disparate phylogenetic origins for Cretaceous ichthyosaurs. Biology Letters, 9:1-6. https://doi.org/10.1098/rsbl.2013.0021
Fischer, V., Cappetta, H., Vincent, P., Garcia, G., Goolaerts, S., Martin, J.E., Roggero, D., and Valentin, X. 2014. Ichthyosaurs from the French Rhaetian indicate a severe turnover across the Triassic-Jurassic boundary. Naturwissenschaften, 101:1027-1040. https://doi.org/10.1007/s00114-014-1242-7
Fischer, V., Weis, R., and Thuy, B. 2021. Refining the marine reptile turnover at the Early-Middle Jurassic transition. PeerJ, 9:1-45. https://doi.org/10.7717/peerj.10647
Fraas, E.E. 1891. Ichthyosaurier der Süddeutschen Trias und Jura-Ablagerungen. H. Laupp, Tübingen.
Fraas, E.E. 1892. Ichthyosaurus numismalis. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg, 48:22-31.
Gallagher, W.B., Miller, K.G., Sherrell, R.M., Browning, J.V., Field, M.P., Olsson, R.K., Sugarman, P.J., Tuorto, S., and Wahyudi, H. 2012. On the last mosasaurs: Late Maastrichtian mosasaurs and the Cretaceous-Paleogene boundary in New Jersey. Bulletin de la Société géologique de France, 183:145-150. https://doi.org/10.2113/gssgfbull.183.2.145
Godefroit, P. 1992. Présence de Leptopterygius tenuirostris (Reptilia, Ichthyosauria) dans le Lias moyen de Lorraine belge. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique Sciences de la Terre, 62:163-170.
Godefroit, P. 1993. Les grands ichthyosaures sinémuriens d’Arlon. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique Sciences de la Terre, 63:25-71.
Godefroit, P. 1994. Les reptiles marins du Toarcien (Jurassique inférieur) belgo-luxembourgeois. Mémoires pour servir à l’Explication des Cartes Géologiques et Minières de la Belgique, 39:98.
Guérin-Franiatte, S. 2003. Biostratigraphie dans le Lias du Grand-Duché de Luxembourg : le sondage de Capellen, p. 65-78. Paléontologie au Luxembourg. Ferrantia - Travaux scientifiques du Musée national d’histoire naturelle, Luxembourg, 36.
Hauff, B. 1953. Das Holzmadenbuch. Verlag der Hohenlohe'schen Buchhandlung, Öhringen.
Hungerbühler, A. and Sachs, S. 1996. Ein großer Ichthyosaurier aus dem Pliensbachium von Bielefeld. Neue Einblicke in die Ichthyosaurier des Mittleren Lias und das Gebiß von Temnodontosaurus. Bericht des Naturwissenschaftlichen Vereins Bielefeld und Umgegend, Bielefeld, 37:15-52.
Jouve, S., Bardet, N., Jalil, N.E., Suberbiola, X.P., Bouya, B., and Amaghzaz, M. 2008. The oldest African crocodylian: phylogeny, paleobiogeography, and differential survivorship of marine reptiles through the Cretaceous-Tertiary boundary. Journal of Vertebrate Paleontology, 28:409-421. https://doi.org/10.1671/0272-4634(2008)28[409:TOACPP]2.0.CO;2
Lomax, D. and Massare, J. 2018a. A forefin of Leptonectes solei from the Lower Jurassic (Pliensbachian) of Dorset, UK. Proceedings of the Geologists’ Association, 129:770-773. https://doi.org/10.1016/j.pgeola.2018.07.005
Lomax, D. and Massare, J. 2018b. A second specimen of Protoichthyosaurus applebyi (Reptila: Ichthyosauria) and additional information on the genus and species. Paludicola, 11:164-178.
Lomax, D.R. 2010. An Ichthyosaurus (Reptilia, Ichthyosauria) with gastric contents from Charmouth, England: first report of the genus from the Pliensbachian. Paludicola, 8:22-36.
Lomax, D.R. 2016. A new leptonectid ichthyosaur from the Lower Jurassic (Hettangian) of Nottinghamshire, England, UK, and the taxonomic usefulness of the ichthyosaurian coracoid. Journal of Systematic Palaeontology, 15:387-401. https://doi.org/10.1080/14772019.2016.1183149
Lomax, D.R. and Massare, J.A. 2015. A new species of Ichthyosaurus from the Lower Jurassic of West Dorset, England, U.K. Journal of Vertebrate Paleontology, 35:e903260. https://doi.org/10.1080/02724634.2014.903260
Lomax, D.R. and Massare, J.A. 2016. Two new species of Ichthyosaurus from the lowermost Jurassic (Hettangian) of Somerset, England. Papers in Palaeontology, 3:1-20. https://doi.org/10.1002/SPP2.1065
Lomax, D.R., Porro, L.B., and Larkin, N.R. 2019. Descriptive anatomy of the largest known specimen of Protoichthyosaurus prostaxalis (Reptilia: Ichthyosauria) including computed tomography and digital reconstruction of a three-dimensional skull. PeerJ, 7:e6112. https://doi.org/10.7717/peerj.6112
Lucius, M. 1948. Geologie Luxemburgs. Das Gutland. Erläuterungen zu der geologischen Spezialkarte Luxemburgs, 5:1-397.
Maisch, M.W. 1997. The cranial osteology of Ichthyosaurus intermedius Conybeare, 1822 from the Lias of Great Britain. Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie), 258:1-27.
Maisch, M.W. 1998. A new ichthyosaur genus from the Posidonia Shale (Lower Toarcian, Jurassic) of Holzmaden, SW-Germany with comments on the phylogeny of post-Triassic ichthyosaurs. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 209:47-78. https://doi.org/10.1127/njgpa/209/1998/47
Maisch, M.W. 2010. Phylogeny, systematics, and origin of the Ichthyosauria - the state of the art. Palaeodiversity, 3:151-214.
Maisch, M.W. and Hungerbühler, A. 1997. Revision of Temnodontosaurus nuertingensis (v. Huene, 1931), a large ichthyosaur from the Lower Pliensbachian (Lower Jurassic) of Nürtingen, South Western Germany. Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie), 248:1-11.
Maisch, M.W. and Matzke, A.T. 2003. The cranial osteology of the ichthyosaur Leptonectes cf. tenuirostris from the Lower Jurassic of England. Journal of Vertebrate Paleontology, 23:116-127. https://doi.org/10.1671/0272-4634(2003)23[116:TCOOTI]2.0.CO;2
Maisch, M.W. and Reisdorf, A.G. 2006. Evidence for the longest stratigraphic range of a post-Triassic Ichthyosaur: a Leptonectes tenuirostris from the Pliensbachian (Lower Jurassic) of Switzerland. Geobios, 39:491-505. https://doi.org/10.1016/j.geobios.2005.04.005
Maisch, M.W., Reisdorf, A., Schlatter, R., and Wetzel, A. 2008. A large skull of Ichthyosaurus (Reptilia: Ichthyosauria) from the Lower Sinemurian (Lower Jurassic) of Frick (NW Switzerland). Swiss Journal of Geoscience, 101:617-627. https://doi.org/10.1007/s00015-008-1299-4
Marek, R.D., Moon, B.C., Williams, M., and Benton, M.J. 2015. The skull and endocranium of a Lower Jurassic ichthyosaur based on digital reconstructions. Palaeontology, 58:723-742. https://doi.org/10.1111/pala.12174
Martin, J.E., Fischer, V., Vincent, P., Suan, G., Martin, J.E., Fischer,V., Vincent, P., Suan, G., Martin, J.E., Fischer, V., Vincent, P., and Suan, G. 2012. A longirostrine Temnodontosaurus (Ichthyosauria) with comments on Early Jurassic ichthyosaur niche partitioning and disparity. Palaeontology, 55:995-1005. https://doi.org/10.1111/j.1475-4983.2012.01159.x
Massare, J.A. and Lomax, D.R. 2016. A new specimen of Ichthyosaurus conybeari (Reptilia, Ichthyosauria) from Watchet, Somerset, England, U.K., and a re-examination of the species. Journal of Vertebrate Paleontology, e1163264. https://doi.org/10.1080/02724634.2016.1163264
Maubeuge, P.L. 1950. Observations sur le Lotharingien et le Carixien du Grand-Duché de Luxembourg et comparaisons avec les régions voisines. Archives de l’Institut Grand-Ducal de Luxembourg, Section des Sciences naturelles, physiques et mathématiques, 19:357-364.
Maubeuge, P.L. and Antun, P. 1967. Observations sur le Lotharingien et le Carixien du Grand-Duché de Luxembourg. Bulletin de l’Académie royale de Belgique (Classe des Sciences) 5e série, 57:249-285.
Maxwell, E.E. 2012. New metrics to differentiate species of Stenopterygius (Reptilia: Ichthyosauria) from the Lower Jurassic of southwestern Germany. Journal of Paleontology, 86:105-115. https://doi.org/10.1666/11-038.1
Maxwell, E.E. 2018. Redescription of the lost holotype of Suevoleviathan integer (Bronn, 1844) (Reptilia: Ichthyosauria). Journal of Vertebrate Paleontology, 38:e1439833. https://doi.org/10.1080/02724634.2018.1439833
Maxwell, E.E. and Vincent, P. 2015. Effects of the early Toarcian Oceanic Anoxic Event on ichthyosaur body size and faunal composition in the Southwest German Basin. Paleobiology, 42:117-126. https://doi.org/10.1017/pab.2015.34
Maxwell, E.E. and Cortés, D. 2020. A revision of the Early Jurassic ichthyosaur Hauffiopteryx (Reptilia: Ichthyosauria), and description of a new species from Southwestern Germany. Palaeontologia Electronica, 23(2):a31. https://doi.org/10.26879/937
Maxwell, E.E., Caldwell, M.W., and Lamoureux, D.O. 2012. Tooth histology, attachment, and replacement in the Ichthyopterygia reviewed in an evolutionary context. Paläontologische Zeitschrift, 86:1-14. https://doi.org/10.1007/s12542-011-0115-z
Maxwell, E.E., Fernández, M.S., and Schoch, R.R. 2012. First diagnostic marine reptile remains from the Aalenian (Middle Jurassic): a new ichthyosaur from southwestern Germany. PLoS ONE, 7:e41692. https://doi.org/10.1371/journal.pone.0041692
McGowan, C. 1973. The cranial morphology of the Lower Liassic latipinnate ichthyosaurs of England. Bulletin of the British Museum (Natural History) Geology, 24:1-109.
McGowan, C. 1974. A revision of the longipinnate ichthyosaurs of the Lower Jurassic of England, with description of the new species (Reptilia, Ichthyosauria). Life Science Contributions, Royal Ontario Museum, 97:1-37.
McGowan, C. 1986. A putative ancestor for the swordfish-like ichthyosaur Eurhinosaurus. Nature, 322:454-456. https://doi.org/10.1038/322454a0
McGowan, C. 1989. Leptopterygius tenuirostris and other long-snouted ichthyosaurs from the English Lower Lias. Palaeontology, 32:409-427.
McGowan, C. 1993. A new species of a large, long-snouted ichthyosaur from the English lower Lias. Canadian Journal of Earth Sciences, 30:1197-1204.
McGowan, C. 1996. Giant ichthyosaurs of the Early Jurassic. Canadian Journal of Earth Sciences, 33:1011-1021.
McGowan, C. 2003. A new specimen of Excalibosaurus from the English Lower Jurassic. Journal of Vertebrate Paleontology, 23:950-956. https://www.jstor.org/stable/4524397
McGowan, C. and Milner, A.C. 1999. A new Pliensbachian ichthyosaur from Dorset, England. Palaeontology, 42:761-768. https://doi.org/10.1111/1475-4983.00096
McGowan, C. and Motani, R. 2003. Part 8. Ichthyopterygia. In Sues, H.-D. (ed.), Handbook of Paleoherpetology, 8. Verlag Dr. Friedrich Pfeil, München.
Merriam, J.C. 1902. Triassic Ichthyopterygia from California and Nevada. University of California Publications: Bulletin of the department of Geology, 3:63-108.
Meyer, C.A. 2011. Amazing graze - Grazing traces of sea urchins on turtles - An example from the Late Jurassic of Switzerland. Annalen des Naturhistorischen Museums in Wien, A113:555-565.
Motani, R. 1999. Phylogeny of the Ichthyopterygia. Journal of Vertebrate Paleontology, 19:473-496. https://www.jstor.org/stable/4524011
Motani, R., Jiang, D.-Y., Chen, G.-B., Tintori, A., Rieppel, O., Ji, C., and Huang, J.-D. 2015. A basal ichthyosauriform with a short snout from the Lower Triassic of China. Nature, 517:485-488. https://doi.org/10.1038/nature13866
Motani, R., Jiang, D.Y., Tintori, A., Ji, C., and Huang, J.D. 2017. Pre- versus post-mass extinction divergence of Mesozoic marine reptiles dictated by time-scale dependence of evolutionary rates. Proceedings of the Royal Society B: Biological Sciences, 284:e20170241. https://doi.org/10.1098/rspb.2017.0241
Reisdorf, A., Maisch, M.W., and Wetzel, A. 2011. First record of the leptonectid ichthyosaur Eurhinosaurus longirostris from the Early Jurassic of Switzerland and its stratigraphic framework. Swiss Journal of Geoscience, 104:211-224. https://doi.org/10.1007/s00015-011-0069-x
Reolid, M., Santos, A., and Mayoral, E. 2015. Grazing activity as taphonomic record of necrobiotic interaction: A case study of a sea turtle carapace from the Upper Jurassic of the Prebetic (south Spain). Revista mexicana de ciencias geológicas, 32:21-28.
Riegraf, W. 1980. Revision der Belemniten des Schwäbischen Jura. Palaeontographica Abteilung A, 169:128-206.
Sander, P.M. 1997. The paleobiogeography of Shastasaurus, p. 17-43. In Callaway, J.M. and Nicholls, E.L. (eds.), Ancient Marine Reptiles. Academic Press, San Diego, California.
Sander, P.M. 2000. Ichthyosauria: their diversity, distribution, and phylogeny. Paläontologische Zeitschrift, 74:1-35. https://doi.org/10.1007/BF02987949
Schlegelmilch, R. 1998. Die Belemniten des süddeutschen Jura: Ein Bestimmungsbuch für Geowissenschaftler und Fossiliensammler. Spektrum Akademischer Verlag, Heidelberg. https://doi.org/10.1007/978-3-8274-3083-0
Sollas, W.J. 1916. The skull of Ichthyosaurus, studied in serial sections. Philosophical transactions of the Royal Society of London, serie B, 208:63-126.
Swaby, E.J. and Lomax, D.R. 2021. A revision of Temnodontosaurus crassimanus (Reptilia: Ichthyosauria) from the Lower Jurassic (Toarcian) of Whitby, Yorkshire, UK. Historical Biology, 33:2715-2731. https://doi.org/10.1080/08912963.2020.1826469
Tutin, S.L. and Butler, R.J. 2017. The completeness of the fossil record of plesiosaurs, marine reptiles from the Mesozoic. Acta Palaeontologica Polonica, 62:563-573. https://doi.org/10.4202/APP.00355.2017
Vincent, P., Taquet, P., Fischer, V., Bardet, N., Falconnet, J., and Godefroit, P. 2014. Mary Anning’s legacy to French vertebrate palaeontology. Geological Magazine, 151:7-20. https://doi.org/10.1017/S0016756813000861
von Hillebrandt, A. and Krystyn, L. 2009. On the oldest Jurassic ammonites of Europe (Northern Calcareous Alps, Austria) and their global significance. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 253:163-195. https://doi.org/10.1127/0077-7749/2009/0253-0163
von Huene, F. 1922. Die Ichthyosaurier des Lias und ihre Zusammenhänge. Monographien zur Geologie und Paläontologie,1. Verlag von Gebrüder Borntraeger, Berlin.
von Huene, F. 1931a. Neue Ichthyosaurier aus Württemberg. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie. Beilage. Abteilung B, 65:305-320.
von Huene, F. 1931b. Neue Studien über Ichthyosaurier aus Holzmaden. Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, 42:345-382.
von Huene, F. 1951. Ein neuer Fund von Eurhinosaurus longirostris. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 93:277-283.
Zverkov, N.G., Fischer, V., Madzia, D., and Benson, R.B.J. 2018. Increased pliosaurid dental disparity across the Jurassic-Cretaceous transition. Palaeontology, 61:825-846. https://doi.org/10.1111/pala.12367
