

 A revision of the Early Jurassic ichthyosaur Hauffiopteryx (Reptilia: Ichthyosauria), and description of a new species from southwestern Germany
A revision of the Early Jurassic ichthyosaur Hauffiopteryx (Reptilia: Ichthyosauria), and description of a new species from southwestern Germany
Article number: 23(2):a31
https://doi.org/10.26879/937
Copyright Palaeontological Association, June 2020
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 2 November 2018. Acceptance: 12 June 2020.
ABSTRACT
Hauffiopteryx typicus is an Early Jurassic ichthyosaur species from Europe, for which geographically partitioned morphological variation between specimens from England and Germany has been described. We provide a complete anatomical description of the German material to address this taxonomical issue. We also identify and describe a new species of Hauffiopteryx from the southwest German Basin, Hauffiopteryx altera sp. nov., differring from H. typicus in the morphology of the arrangement of cranial elements surrounding the external nares. A phylogenetic analysis recovers the German and English material referred to H. typicus as sister taxa, suggesting that these are indeed conspecific. H. typicus forms a monophyletic group with H. altera and a specimen from the Pliensbachian of Switzerland previously referred to Leptonectes tenuirostris but consistent with H. typicus. We conclude that Hauffiopteryx represents a valid genus, defined by a set of synapomorphies from both the skull and postcranium. Parsimony analysis recovers Hauffiopteryx as sister taxon to Stenopterygius + Ophthalmosauridae.
Erin E. Maxwell. Staatliches Museum für Naturkunde, Rosenstein 1, 70191 Stuttgart, Germany, erin.maxwell@smns-bw.de
Dirley Cortés. Redpath Museum, McGill University, 859 Sherbrooke St. W., Montreal QC H3A 0C4, Canada. Smithsonian Tropical Research Institute, Balboa-Ancón 0843-03092, Panamá, Panamá. dirley.cortes@mail.mcgill.ca
Keywords: Hauffiopteryx; new species; Posidonienschiefer Formation; Germany; Toarcian; Early Jurassic
Final citation: Maxwell, Erin E. and Cortés, Dirley. 2020. A revision of the Early Jurassic ichthyosaur Hauffiopteryx (Reptilia: Ichthyosauria), and description of a new species from southwestern Germany. Palaeontologia Electronica, 23(2):a31. https://doi.org/10.26879/937
palaeo-electronica.org/content/2020/3078-revision-of-hauffiopteryx
Copyright: June 2020 Palaeontological Association.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/041F166A-28FC-4ED6-AD7C-DF64EF4EF1A7
INTRODUCTION
The Posidonienschiefer Formation (Posidonia Shale) of southwestern Germany is considered one of the classic Mesozoic marine fossil lagerstätte, and has yielded thousands of exceptionally preserved fish and reptile remains (Urlichs et al., 1994). The fossil marine reptiles, in particular, have garnered a great deal of research attention. The most abundant and best-preserved of these are the ichthyosaurs, represented in museum collections by hundreds of specimens including examples with fossilized soft-tissues, gastric contents, and embryos preserved inside the body cavity (Hauff, 1921; Böttcher, 1989, 1990). The first ichthyosaur formally named from the Posidonia Shale was Temnodontosaurus trigonodon, named over 175 years ago (Theodori, 1843), and the most recently erected species still regarded as valid were named almost 90 years ago (Stenopterygius uniter von Huene, 1931 and Hauffiopteryx typicus [von Huene, 1931]). Ongoing research has emphasized questions pertaining to paleobiology, including intraspecific variation (e.g., Maxwell, 2012a), ontogeny (e.g., Dick and Maxwell, 2015), diet (e.g., Dick et al., 2016), and prevalence of skeletal pathologies (e.g., Pardo-Pérez et al., 2019). The discovery of new species following so many years of intensive research effort was thought to be unlikely.
The monotypic genus Hauffiopteryx (Maisch, 2008) is the most recently named and least distinctive ichthyosaur genus from the Posidonienschiefer Formation. The genus has a convoluted history. Hauffiopteryx typicus (von Huene, 1931) was initially referred to Stenopterygius as S. hauffianus / S. hauffianus forma typica due to its small size and similarities in forelimb morphology (von Huene, 1922, 1931); the unusual morphology of the pelvic girdle was interpreted as an abnormality (McGowan, 1978). Maisch (2008) recognized Hauffiopteryx as distinct from Stenopterygius based on the small, round temporal fenestra, extensive gastralia, and medially unfused pelvic girdle. He considered Hauffiopteryx to be nested within Thunnosauria, potentially forming a sister group to Stenopterygius + Ophthalmosauridae. This hypothesis was supported by subsequent cladistic analyses (Fischer et al., 2016; Moon, 2017). However, Hauffiopteryx has also been recovered in a more basal position, as sister group to Thunnosauria (Ji et al., 2016).
Maisch (2008) initially identified seven specimens referable to Hauffiopteryx typicus: one from the Toarcian of the UK, five from southwestern Germany, and one from Luxembourg. We were unable to examine the latter skull personally but consider most likely referable to the more abundant genus Stenopterygius based on the apparent exclusion of the prefrontal by the lacrimal from the posterior edge of the external nares (Godefroit, 1994: pl. 3a). Two additional specimens from the Toarcian of the UK were later referred (Caine and Benton, 2011). The specific referral of BRLSI M 1399, part of the original referred material cited by Maisch (2008), was later questioned following CT-scanning and detailed description, as it failed to form a monophyletic group with H. typicus in phylogenetic analysis (Marek et al., 2015). This raised questions of provincialism and endemism among Early Jurassic ichthyosaurs that can only be resolved by restudy of the German material.
In addition to the material from the Toarcian of Germany and the UK, a small skull from the Pliensbachian of Switzerland (NMO 26575) referred to Leptonectes tenuirostris was described as varying from the typical morphology observed in Hettangian-Sinemurian specimens of L. tenuirostris by a series of features such as small size, shorter rostrum, participation of the prefrontal in the external narial opening, and notching of the forefin elements (Maisch and Reisdorf, 2006). These characters are now known to differentiate Hauffiopteryx and Leptonectes, thus meriting reevaluation of the taxonomic affinities of NMO 26575. Problematically, L. tenuirostris was partially scored based on NMO 26575 in the analysis of Moon (2017), and possibly other analyses as well.
Here, we provide a complete anatomical description of Hauffiopteryx from the Southwest German Basin. We re-examined the five specimens cited by Maisch (2008), and included five additional specimens from the collections of the SMNS, GPIT, and the Werkforum Dotternhausen. This study led to the recognition of a second species of Hauffiopteryx from southwestern Germany, Hauffiopteryx altera sp. nov. We also reconsider the affinities of NMO 26575 from the Early Jurassic of Switzerland described as Leptonectes tenuirostris by Maisch and Reisdorf (2006), which shares many morphological similarities with H. typicus.
MATERIAL
We re-examined the five specimens cited by Maisch (2008) as referable to Hauffiopteryx typicus: GPIT 1491/4 (bed number εII 4, Holzmaden), MHH 9 (bed number εII 4/5, Holzmaden), SMNS 51552 (εII 3, Zell unter Aichelberg), SMNS 80225 (εII 4, Schlierbach), and SMNS 81965 (εII 4, Schlierbach). We also studied an additional six specimens that we considered to be referable to Hauffiopteryx based on the diagnosis of Maisch (2008): SMNS 81367 (a small skeleton in left lateral view lacking the pelvic girdle and anterior caudal region, from bed number εII 4 of Schlierbach, Baden-Württemberg), SMNS 80226 (a partial skeleton in ventral view with pectoral girdle, gastralia and ribs preserved but lacking the hind limbs, from bed number εII 4 of Schlierbach, Baden-Württemberg), SMNS 81962 (an almost complete skeleton in ventral view, including a partial skull, pectoral girdle and limbs, from bed number εII 3, Ohmden), GPIT/RE/12905, a moderately compressed skull from an unknown locality and horizon of the Posidonienschiefer Formation, and FWD-129 (Dotternhausen-Dormettingen, εII4/5), a lightly compressed skull, pectoral girdle and anterior vertebral column preserved in a concretion, previously considered to be a juvenile of Eurhinosaurus longirostris (Jäger, 2005). We also restudied NMO 26575, a three-dimensional skull and some associated postcranial elements from the upper Pliensbachian of Hauenstein, Switzerland, previously referred to Leptonectes tenuirostris but varying in several characteristics now more suggestive of Hauffiopteryx Maisch, 2008, such as small size and closely packed forelimb elements with anterior notches (Maisch and Reisdorf, 2006).
Institutional Abbreviations
BRLSI, Bath Royal Literary and Scientific Institution, Bath, UK; FWD, Werkforum, Dotternhausen-Dormettingen, Baden-Württemberg, Germany; GPIT, Palaeontological Collections of Tübingen University, Tübingen, Germany; MHH, Museum Hauff, Holzmaden, Germany; NMO, Naturmuseum Olten, Switzerland; SMNS, Staatliches Museum für Naturkunde Stuttgart, Germany.
SYSTEMATIC PALEONTOLOGY
ICHTHYOPTERYGIA Owen, 1859
ICHTHYOSAURIA Blainville, 1835
PARVIPELVIA Motani, 1999
HAUFFIOPTERYX Maisch, 2008
Type species. Hauffiopteryx typicus (von Huene, 1931)
Revised generic diagnosis. A small- to mid-sized ichthyosaur, 2-3 m in length, with relatively short and slender antorbital rostrum; slight overbite present (unlike Eurhinosaurus, Excalibosaurus in which a substantial overbite is present); prefrontal participating in margin of external naris (unlike Eurhinosaurus, Leptonectes, Stenopterygius); nasals with a greater contribution to the dorsal midline of the rostrum anterior to the external narial opening than the premaxillae (unlike Stenopterygius); prefrontal with larger external exposure on dorsal skull roof than the postfrontal (unlike Stenopterygius); parietal foramen situated anterior to the supratemporal fenestra (situated at the same level as the anterior edge of the supratemporal fenestra in Stenopterygius); supratemporal fenestra small and circular in dorsal view (similar to Eurhinosaurus but unlike Stenopterygius); supratemporal palmate in posterodorsal view (similar to Eurhinosaurus, unlike Stenopterygius); teeth with smooth enamel (unlike Suevoleviathan); gastralia extending posteriorly to the thirty-fifth presacral vertebra (unique feature among Toarcian ichthyosaurs); rib tuberculum and capitulum widely separated in the dorsal region (unlike Temnodontosaurus, Suevoleviathan, Eurhinosaurus); glenoid end of scapula anteriorly expanded (unlike in Temnodontosaurus); coracoid with anterior notch, foramen between humerus, radius, and ulna absent (unlike Suevoleviathan, Temnodontosaurus trigonodon); anterior digit in both fore- and hindfins with notches on the anterior edge (unlike in Suevoleviathan); proximal limb elements angular and forming a tightly articulated mosaic; ischium and pubis thin and styloidal, fused laterally but widely separated medially (autapomorphic); fibula larger than the tibia; hindfin digits converging distally.
Hauffiopteryx typicus (von Huene, 1931)
Figure 1, Figure 2, Figure 3, Figure 4, Figure 5
Lectotype. GPIT 1491/4; a complete skeleton with partial preserved soft-tissue outline in left lateral view. GPIT 1491/4; a complete skeleton with partial preserved soft-tissue outline in left lateral view (Figure 1A).
Geographical distribution. Baden-Württemberg, Germany; Ilminster (UK) (Maisch, 2008); Solothurn, Switzerland (see discussion for details).
 Stratigraphic distribution. Late Pliensbachian, margaritatus Zone; lower Toarcian, tenuicostatum Zone, semicelatum Subzone- serpentinum Zone, exaratum Subzone (SWGB: beds εII3- εII4/5). Note that according to the specimen label, SMNS 51552 originates from bed εII3, rather than εII2 as stated elsewhere (Maisch, 2008). Peak abundance occurs in εII4, where material referred to Hauffiopteryx represents a substantial percentage of all small ichthyosaurs recovered (Maxwell and Vincent, 2016).
Stratigraphic distribution. Late Pliensbachian, margaritatus Zone; lower Toarcian, tenuicostatum Zone, semicelatum Subzone- serpentinum Zone, exaratum Subzone (SWGB: beds εII3- εII4/5). Note that according to the specimen label, SMNS 51552 originates from bed εII3, rather than εII2 as stated elsewhere (Maisch, 2008). Peak abundance occurs in εII4, where material referred to Hauffiopteryx represents a substantial percentage of all small ichthyosaurs recovered (Maxwell and Vincent, 2016).
Emended diagnosis. Hauffiopteryx typicus can be distinguished from other species of Hauffiopteryx by a triradiate lacrimal forming the posterior border of the external narial opening, deepest lateral exposure of the maxilla located ventral to the posterior half of the external narial opening, a large, rounded orbit, and nasals with pronounced dorsal inflection at the anterior orbital margin.
Referred specimens. Of the material examined, GPIT 1491/4, GPIT/RE/12905, MHH 9, SMNS 51552, and SMNS 80226 can be confidently referred to H. typicus (Figure 1, Figure 2, Figure 3). The English material referred to Hauffiopteryx is also consistent with H. typicus, as is NMO 26575 from the Pliensbachian of Switzerland (see discussion for details).
DESCRIPTION
The description is based on GPIT 1491/4, GPIT/RE/12905, MHH 9, SMNS 51552, and SMNS 80226; selected measurements can be found in Appendix 1. Variation in the shape of the posterior premaxilla and anterior lacrimal in GPIT/RE/12905 is attributed to differences in preservation and intraspecific variation between this and the other skulls; these variants are clearly noted in the description. The English and Swiss material is excluded from the description.
 Premaxilla. The premaxilla is gracile and dentigerous, decreasing rapidly in depth anteriorly (Figure 2A-2C). Posteriorly, a subnarial process extends ventral to the anterior portion of the external narial opening. A robust, extensive supranarial process is absent; however, in GPIT/RE/12905 a slender process is present (Figure 3A). It is shorter than the subnarial process, and extends less than one quarter of the length of the external narial opening. Although apparently absent in GPIT 1491/4, MHH 9, and SMNS 51552, the quality of preservation of this region in these specimens does not preclude the erosion of such a delicate bony structure through either taphonomic processes or preparation. The premaxillary fossa is deepest at its midpoint, becoming rapidly shallower posteriorly. The premaxillary rostrum is short in all diagnostic material, with prenarial segment ratios of ~0.49 (GPIT/RE/12905), 0.48 (MHH 9), ~0.42 (SMNS 80226), and ~0.37 (GPIT 1491/4). The rostral tips (both premaxillary and mandibular) of SMNS 51552, often cited as an example of Hauffiopteryx with a long rostrum (Maisch, 2008), are of problematic authenticity; as preserved the prenarial segment = 0.46.
Premaxilla. The premaxilla is gracile and dentigerous, decreasing rapidly in depth anteriorly (Figure 2A-2C). Posteriorly, a subnarial process extends ventral to the anterior portion of the external narial opening. A robust, extensive supranarial process is absent; however, in GPIT/RE/12905 a slender process is present (Figure 3A). It is shorter than the subnarial process, and extends less than one quarter of the length of the external narial opening. Although apparently absent in GPIT 1491/4, MHH 9, and SMNS 51552, the quality of preservation of this region in these specimens does not preclude the erosion of such a delicate bony structure through either taphonomic processes or preparation. The premaxillary fossa is deepest at its midpoint, becoming rapidly shallower posteriorly. The premaxillary rostrum is short in all diagnostic material, with prenarial segment ratios of ~0.49 (GPIT/RE/12905), 0.48 (MHH 9), ~0.42 (SMNS 80226), and ~0.37 (GPIT 1491/4). The rostral tips (both premaxillary and mandibular) of SMNS 51552, often cited as an example of Hauffiopteryx with a long rostrum (Maisch, 2008), are of problematic authenticity; as preserved the prenarial segment = 0.46.
 Maxilla. The maxilla in H. typicus is short, and does not extend more than a few millimeters anterior to the external narial opening in lateral view (Figure 2A-2C, Figure 3A). The dentigerous portion continues posterior to the external narial opening. The maxilla forms the ventral edge of the external narial opening in most specimens, but is excluded in GPIT/RE/12905 by contact between the premaxilla and lacrimal. An ascending process posterior to the narial opening is absent. The orbital process of the maxilla is moderately elongated, but does not reach the orbital mid-point and is shorter than the suborbital process of the lacrimal. Anteriorly, the maxilla is dorsally overlapped by the premaxilla. Posteriorly, it contacts the jugal and lacrimal.
Maxilla. The maxilla in H. typicus is short, and does not extend more than a few millimeters anterior to the external narial opening in lateral view (Figure 2A-2C, Figure 3A). The dentigerous portion continues posterior to the external narial opening. The maxilla forms the ventral edge of the external narial opening in most specimens, but is excluded in GPIT/RE/12905 by contact between the premaxilla and lacrimal. An ascending process posterior to the narial opening is absent. The orbital process of the maxilla is moderately elongated, but does not reach the orbital mid-point and is shorter than the suborbital process of the lacrimal. Anteriorly, the maxilla is dorsally overlapped by the premaxilla. Posteriorly, it contacts the jugal and lacrimal.
Nasal. The nasals are exposed dorsally over approximately two thirds of the antorbital rostrum (Figure 2A, 2C). The presence of an internasal foramen could not be evaluated in the extremely compressed material, but an internasal depression was certainly present medial to the external nares. The nasal forms the dorsal edge of the external narial opening and contacts the prefrontal at the posterodorsal edge of the external narial opening. On the dorsal skull roof, the nasals contact the prefrontal laterally and the frontal posteriorly. A small, superficial contact between the nasal and the postfrontal, overlapping the prefrontal, may occur posterolaterally (Figure 2C).
Lacrimal. The lacrimal is small, with a very short subnarial process in most skulls (Figure 2A-2C). It usually participates in only the posteroventral corner of the external narial opening. However, in GPIT/RE/12905 the subnarial process of the lacrimal is slender but elongate, extending over more than half the total length of the external narial opening to contact the subnarial process of the premaxilla (Figure 3A); we consider this to be intraspecific variation, as documented in other Early Jurassic ichthyosaur species (see discussion). The suborbital process of the lacrimal articulates ventrally with the jugal and is relatively elongate. The suborbital process is separated from the lateral portion of the element by a ridge, forming part of the circumorbital area. The ascending process contacts the prefrontal via an interdigitating suture, and does not contact the nasal.
Jugal. The jugal is a relatively slender, deeply bowed element. It is overlapped by the postorbital posterolaterally, excluding the ventral postorbital from the posterior edge of the orbit (Figure 2A-2C, Figure 3A). The dorsal process of the posterior jugal is narrow and mediolaterally compressed, lacking pronounced anteroposterior expansion. Anteriorly, the jugal narrows, and slots between the lacrimal and maxilla. It does not reach the external narial opening.
Prefrontal. The prefrontal forms a large narial process, and participates extensively in the posterior portion of the external narial opening (Figure 2A-2C). It forms approximately 50% of the dorsal orbital margin. The prefrontal extends medially to contact the frontal, but may be overlapped by processes of the postfrontal and the nasal.
Frontal. The frontals are relatively narrow and slightly convex, tapering anteriorly. They form the external anterior and lateral margins of the parietal foramen (Figure 2A, 2C).
Parietal. The parietal forms approximately half of the anterior edge of the supratemporal fenestra (Figure 2A, 2C). The medial process of the supratemporal excludes the parietal entirely from the posteromedial edge of the fenestra, best seen in GPIT/RE/12905. The interparietal suture is straight. A weak ridge separates the posterior lamina from the dorsal surface of the parietal. A weakly developed ridge lateral to the parietal foramen runs anterolaterally to posteromedially.
Postfrontal. The postfrontal forms the anterolateral margin of the supratemporal fenestra (Figure 2A, 2C). It interdigitates with the supratemporal along the posterolateral margin of the supratemporal fenestra. The postfrontal overlaps the postorbital along the dorsal edge of the orbit.
Postorbital. The postorbital makes up the posterior edge of the orbit (Figure 2A-2D). It is a narrow semi-lunate element. It contacts the squamosal posteriorly, and the supratemporal dorsally, with a very small anterodorsal contact with the postfrontal. Ventrally, the postorbital is excluded from the orbital margin in all specimens by the ascending process of the jugal.
Supratemporal. The supratemporal is large, forming a posterior lamella with radiating ridges, giving it a scalloped appearance in external view (Figure 2A, 2C). These ridges correspond to the anteromedial and anterolateral processes of the anterior ramus, as well as to small medial and lateral processes of the medial ramus. A well-developed ventral ramus is present. It is not significantly longer than the medial or lateral processes.
Squamosal. The squamosal is well-preserved only in GPIT/RE/12905 (Figure 3). It is exposed on the posterolateral skull and is roughly triangular in shape, with a descending process posterior to the postorbital. Dorsally it contacts the supratemporal.
Quadratojugal. The ventral portion of the quadratojugal is preserved in the lectotype (Figure 2A). In external view, it appears to be excluded from contact with the jugal by the postorbital, but this could be due to taphonomic displacement. The ventral edge is thickened for articulation with the quadrate.
Quadrate. The posterolateral portion of the quadrate is exposed in both MHH 9 and in GPIT 1491/4 (Figure 2A, 2C). The quadrate is laterally concave. The ventral articular end is thickened, and a small occipital lamella is present.
Braincase
Basioccipital. The basioccipital is not well-preserved in the lectotype. It is best-exposed in SMNS 80226 and GPIT/RE/12905 in ventral view (Figure 2D-2E). It has an extensive extracondylar area (ECA), with the portion ventral to the condyle being wider than the lateral ECA. The ventral extracondylar area is anteromedially concave, with small lateral tubers. The condyle itself bears a notochordal pit and is clearly offset from the extracondylar area.
Opisthotic. The right opisthotic is preserved in SMNS 80226 (Figure 2D-2E), in what is interpreted as anterior view. The medial edge bears a notch interpreted as the vagus foramen. The paroccipital process is short. The anterior surface of the opisthotic ventral to the paroccipital process is concave, clearly setting off the process.
Stapes. An element thought to be the left stapes is preserved in SMNS 80226 (Figure 2D). The lateral shaft is slender, and the quadrate articulation is not expanded. The ventral surface of the shaft is more concave than the dorsal surface. The medial head is robust.
Exoccipitals. Exoccipitals are preserved in SMNS 51552 as well as in the lectotype (Figure 2A-2B). These are short, squat elements forming the lateral edges of the foramen magnum. The medial edge is concave. The number of foramina cannot be assessed with confidence.
Prootic, supraoccipital, parabasisphenoid. None of these elements could be identified with confidence in the specimens determined to be diagnostic at the specific level.
Palate
The palate of H. typicus is best-exposed in SMNS 51552 (Figure 2B; see also Maisch, 1998a: fig. 3).
Pterygoid. The pterygoid is preserved in ventral view. The quadrate ramus is preserved posteriorly and is offset from the palatal ramus by a pronounced constriction. The lateral process of the quadrate ramus is more slender than the medial process. The palatal ramus bears a robust postpalatine process.
Palatine. The palatine is present in SMNS 51552; however, due to poor preparation little can be said about either its anatomy or position.
Vomer. The vomer could not be identified in the specimens determined to be diagnostic at the specific level.
Mandible
A small overbite is consistently present (Figure 2A, 2C-2D, 2G); this does not appear to become more pronounced in larger specimens. The lower jaw is bowed ventral to the orbit.
The dentaries are fine and pointed anteriorly, and make up approximately 45% of the length of the mandibular symphysis (Figure 2D, 2G). In lateral view, the dentary fossa is well-developed and deep, but disappears towards the anteriormost tip of the mandible (Figure 2A-2B). Aulacodont tooth implantation is seen throughout the dentary, with the alveolar groove narrowing significantly towards the anterior tip (Figure 2C).
The splenials form the medial surface of the mandible and a substantial portion of the mandibular symphysis. They are most robust at the posteriormost symphysis, thinning rapidly anteriorly in ventral view (Figure 2B, 2D). Posteriorly, they also attenuate in ventral view, but more slowly. At their posterior ends, the splenials are concealed in ventral view in articulated specimens by the medial curvature of the angular, which forms a shelf.
The posterior lateral lower jaw is made up of the surangular and angular. The lateral exposure of the former is much greater than that of the latter. The angular forms the ventral edge of the lower jaw, extending from the retroarticular process posteriorly to approximately the level of the external narial opening. The surangular extends further anteriorly than the angular. Ventral to the orbit, the surangular forms a well-developed surangular fossa. A strong ridge and ventral depression is developed on the lateral surangular contribution to the retroarticular process, presumably indicating a point of muscle attachment.
Prearticular. The prearticular is preserved in SMNS 80226, forming a fine splint of bone in ventral view posterior to and medially overlapped by the splenial.
Articular. The articular is not well-exposed in any of the diagnostic material.
Dentition. The teeth are small and acutely pointed, with the crowns typically showing some lingual curvature. The enamel is smooth (Figure 2F, 2G). Crown height decreases notably towards the anterior tips of the jaws: in MHH 9, for instance, mid-premaxillary tooth crowns are approximately 4.5 mm in height, whereas the anteriormost dentary crowns are approximately 2 mm in height. The tooth roots show strong apicobasal ridges indicative of plicidentine; large quantities of cellular cementum appear to be absent.
Sclerotic ring. The sclerotic ring in the GPIT 1491/4 preserves approximately 16-17 ossicles (poor preservation makes an exact count impossible); 17 ossicles are also preserved in SMNS 51552 and GPIT/RE/12905. In all three specimens, the sclerotic ring appears to fill the orbit, although taphonomic breakage and distortion are moderate to severe in all skulls (least so in GPIT/RE/12905).
Hyoid apparatus. Paired ceratobranchials are ossified but often compressed, making it difficult to assess morphology. Shape differs widely between specimens, with the ceratobranchials being laterally concave in SMNS 51552 (Figure 2B) and laterally convex in SMNS 80226 (Figure 2D). An ossified hyoid corpus is present in SMNS 51552 (Figure 2B).
Postcranium
 Vertebral column. 45-46 presacral vertebrae, 81 preflexural vertebrae, ~3 apical vertebrae, and >55 postflexural vertebrae are preserved in the lectotype, becoming very small towards the end of the caudal fin (Figure 1A). There is some uncertainty in presacral count because the last vertebra with unfused di- and parapophyses is posterior to the position of the pelvic girdle. The atlas and axis are preserved in SMNS 80226 and are exposed in lateral view (Figure 4B). They are completely fused, with a suture persisting on the lateroventral surface. The remaining presacral centra are round in articular view and are deeply amphicoelous. Cervical centra are small, and dimensions increase posteriorly. Separate di- and parapophyses are present in all dorsal vertebrae. Apophyses begin to shift to a mid-centrum position anterior to the tail bend. Apophyses are absent in the apical centra and all postflexural vertebrae. The vertebral column is gently curved along its length, with the points of inflection located in the anterior dorsal region and mid-preflexural region.
Vertebral column. 45-46 presacral vertebrae, 81 preflexural vertebrae, ~3 apical vertebrae, and >55 postflexural vertebrae are preserved in the lectotype, becoming very small towards the end of the caudal fin (Figure 1A). There is some uncertainty in presacral count because the last vertebra with unfused di- and parapophyses is posterior to the position of the pelvic girdle. The atlas and axis are preserved in SMNS 80226 and are exposed in lateral view (Figure 4B). They are completely fused, with a suture persisting on the lateroventral surface. The remaining presacral centra are round in articular view and are deeply amphicoelous. Cervical centra are small, and dimensions increase posteriorly. Separate di- and parapophyses are present in all dorsal vertebrae. Apophyses begin to shift to a mid-centrum position anterior to the tail bend. Apophyses are absent in the apical centra and all postflexural vertebrae. The vertebral column is gently curved along its length, with the points of inflection located in the anterior dorsal region and mid-preflexural region.
In the lectotype, the axial neural spine is much broader than more posterior neural spines at its base, but tapers dorsally (Figure 4A). A substantial facet for the atlantal neural spine is preserved on its anterolateral surface. The atlantal spine itself is not preserved. The anterior dorsal neural spines are rectangular, but become broader and rounded in the posterior 2/3 of the vertebral column (Figure 1A, Figure 4D). Posterior to the pelvic girdle, neural spines become T-shaped, and lose articulation with each other. “Extra-neural processes” (sensu McGowan, 1992), as previously described in Stenopterygius, are present ventral to the posterior end of the dorsal fin and also on several more anterior vertebrae. Although positioned dorsal to the ossified neural spines, these lack any visible connection with the underlying skeletal tissues.
Ribs and gastralia. The cervical ribs are somewhat bicapitate, but the tuberculum and capitulum are less deeply separated than in the dorsal ribs (e.g., Figure 4A, 4C). Cervical and dorsal ribs are deeply grooved on the anterior and posterior surfaces. Caudal ribs are small, holocephalous, and paddle-like. Posterior to the apical vertebrae, ribs are absent - the presence of extensive soft-tissue preservation suggests that this is not an artefact. Chevrons are absent throughout the caudal skeleton.
Gastralia are relatively robust. They are long and thin, tapering at either end. There are two gastralial elements per side. In SMNS 80226, gastralia from the right and left sides are fused, forming a single dorso-ventrally flattened and slightly anteroposteriorly expanded boomerang-shaped bone along the midline (Figure 4C). However, this specimen also shows signs of callus development on many gastralia (Pardo-Pérez et al., 2019); it is unclear whether the formation of a median element may have been caused by trauma rather than representing normal morphology.
Body outline. The dorsal and caudal fins are preserved in the lectotype (Figure 1A). The dorsal fin is triangular in outline, located in the posterior half of the dorsal region and terminating anterior to the pelvic girdle. The caudal fin is semi-lunate in outline, and is symmetrical, with a very high aspect ratio and a strongly concave posterior edge. However, the degree to which preparation may have enhanced this shape is uncertain. The vertebral column extends into the distalmost tip of the ventral lobe.
 Pectoral girdle and forelimb. The interclavicle is T-shaped, with the transverse bar being longer than the posterior stem. The posterior edge of the transverse bar is medially broader than at its lateral ends (Figure 5A). The clavicles are wing-like, tapering laterally, and medially broad (Figure 5A, 5C). The coracoids are large, anteroposteriorly longer than proximodistally wide (Figure 5A, 5C). A small anterior notch is present. The scapular facet is much smaller than the glenoid contribution. A medial facet for the scapula appears to be absent. The scapula itself is divided into a medial blade and a lateral shaft (Figure 5A, 5B). The medial blade is anteriorly expanded, but a well-developed acromion process is absent. The coracoid facet is smaller than the glenoid contribution.
Pectoral girdle and forelimb. The interclavicle is T-shaped, with the transverse bar being longer than the posterior stem. The posterior edge of the transverse bar is medially broader than at its lateral ends (Figure 5A). The clavicles are wing-like, tapering laterally, and medially broad (Figure 5A, 5C). The coracoids are large, anteroposteriorly longer than proximodistally wide (Figure 5A, 5C). A small anterior notch is present. The scapular facet is much smaller than the glenoid contribution. A medial facet for the scapula appears to be absent. The scapula itself is divided into a medial blade and a lateral shaft (Figure 5A, 5B). The medial blade is anteriorly expanded, but a well-developed acromion process is absent. The coracoid facet is smaller than the glenoid contribution.
The humerus is relatively elongate, with the distal end greatly expanded relative to the proximal end (Figure 5). A well-developed humeral head is present only in the lectotype (Figure 5B); in all of the referred specimens the proximal humerus is rather flattened. The lectotype is the only specimen in which the humerus is exposed in dorsal view; unfortunately it is also the most strongly compressed specimen. The dorsal process is small, if present. The deltopectoral crest is robust, located close to the anterior edge and extending over more than half of the total length of the humerus (Figure 5A, 5C). The distal humerus bears a flattened, anteriorly directed facet on its leading edge (“tubercle” of Moon, 2017). Two distal facets are present, for articulation with the radius and ulna. The ulnar facet is larger than the radial facet.
All referred specimens show a similar forelimb configuration, in which the radius, radiale, distal carpal 2, and metacarpal II bear notches on their anterior surfaces (Figure 5B, 5C). In most specimens, the first phalanx of digit II is also notched anteriorly. The radius articulates posteriorly with the ulna, and distally with the radiale and intermedium. The ulna articulates distally with the intermedium and ulnare. A small pisiform posterior to the ulnare is present only in the lectotype (Figure 5B). The paddle is four digits wide; in the lectotype, a series of very small accessory ossicles lies posterior to digit V. The posterior two digits (IV and V) are the longest, with ~10 phalanges. Proximal limb elements are polygonal and tightly packed, whereas more distal elements are rounded and widely spaced. Based on the preserved soft tissue outline, the flipper was elongate and tapering, with digit II positioned close to the anterior edge.
 Pelvic girdle and hind limb. The ilium is not well-preserved in any of the referred specimens. In SMNS 80226, the element inferred to be the ilium is straight, shorter than the ischium and pubis (Figure 6A). The pubis is slender, being straight medially and posteriorly curving laterally to fuse with the ischium. Its medial end appears to be medially flattened. The ischium is the largest element in the pelvic girdle. It is thickened laterally and is triangular in cross-section, with a flat anterior surface. The medial end is also slightly anteroposteriorly expanded, and is gently rounded.
Pelvic girdle and hind limb. The ilium is not well-preserved in any of the referred specimens. In SMNS 80226, the element inferred to be the ilium is straight, shorter than the ischium and pubis (Figure 6A). The pubis is slender, being straight medially and posteriorly curving laterally to fuse with the ischium. Its medial end appears to be medially flattened. The ischium is the largest element in the pelvic girdle. It is thickened laterally and is triangular in cross-section, with a flat anterior surface. The medial end is also slightly anteroposteriorly expanded, and is gently rounded.
The femur is wider distally than proximally. The proximal end is strongly convex. The dorsal process is pronounced and more rounded (Figure 6B-6D). The ventral process is narrower, and extends further distally than the dorsal process (Figure 6A). There are two distal facets, for articulation with the tibia and fibula. The tibial facet is smaller than the fibular facet.
The hind fins are short and rounded (Figure 6B-6E). There are three elements in the zeugopodial row: the tibia, fibula, and a large pisiform that appears to give rise to a postaxial digit (in the sense that all elements in the digit are ossified and decrease in size from proximal to distal). The tibia is smaller than the fibula. The tibia and next three distal elements are notched anteriorly. The astragalus is positioned between the distal tibia and fibula. There are four digits in the metatarsal row, assuming that the posterior digit discussed above can be homologized with the ‘true’ digits. As discussed by Maisch (2008), the digits converge distally and so are of equal length. There are only approximately 4-5 phalanges per digit.
Hauffiopteryx altera sp. nov. Figure 7-Figure 8
zoobank.org/A567A292-2A58-495A-839F-FF8382B49479
v. 2005 Eurhinosaurus longirostris (Mantell, 1851): Jäger, p. 29, fig. 23.
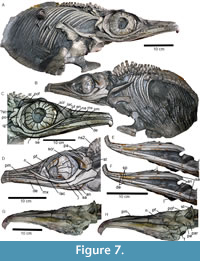 Holotype. Holotype and only referred specimen FWD-129; a skull and partial postcranium preserved in a calcareous concretion (Figure 7-Figure 8; see Table 1 for selected measurements). The bones on the right side of the skull remain in articulation; those on the left side of the skull have suffered some taphonomic displacement.
Holotype. Holotype and only referred specimen FWD-129; a skull and partial postcranium preserved in a calcareous concretion (Figure 7-Figure 8; see Table 1 for selected measurements). The bones on the right side of the skull remain in articulation; those on the left side of the skull have suffered some taphonomic displacement.
Etymology. The specific epithet is derived from the Latin altera, which means different from/other and refers to the anatomical divergence from the type species, H. typicus.
Geographical distribution. Dotternhausen-Dormettingen, Baden-Württemberg, Germany.
Stratigraphic distribution. Posidonienschiefer Formation, lower Toarcian, serpentinum Zone, exaratum Subzone (SWGB: bed εII4/5).
Diagnosis. Hauffiopteryx altera differs from H typicus in: deepest lateral exposure of the maxilla located posterior rather than ventral to the external narial opening; broad, triangular lacrimal excluded from the external narial opening by the descending process of the prefrontal; nasals extending further posteriorly on dorsal skull roof; dorsal exposure of prefrontal substantially greater than that of postfrontal.
DESCRIPTION
Skull
Premaxilla. The premaxilla has been broken and deformed anteriorly (Figure 7). Posteriorly, it is narrow and straight with a deep premaxillary fossa. A supranarial process is absent. The posterior tip of the premaxilla overlaps the maxilla almost to the midpoint and forms the anteroventral edge of the external nares.
Maxilla. The maxilla has a small contact with the ventral edge of the external narial opening, and extends less than one narial length anterior to the nares (Figure 7C-7D). It is deeper posteriorly than anteriorly. Its deepest point in lateral view contacts the anteroventral tip of the prefrontal at the posterior edge of the narial opening (Figure 7C). The ventral edge of the maxilla is long and straight and posteriorly extends only to the anterior edge of the orbit on the right-hand side, being overlapped dorsally by the jugal; on the left the suborbital process is exposed in lateral view and is very long, extending to the orbital midpoint.
Nasal and external narial opening. The external narial opening is 25 mm long (right-hand side). It is simple in shape, with a small posterodorsal notch (Figure 7C). In lateral view, the dorsal surface of the nasal has a weak dorsal inflection anterior to the orbit. The ventral edge of the nasal is deflected ventrally at the level of the external narial opening, and the nasal forms a small ventrolateral shelf dorsal to the posterior narial opening. The nasals are relatively narrow anteriorly, but become broader at the level of the orbit, and are relatively limited in posterior extent. In dorsal view, the right and left nasals diverge posteriorly, creating a V-shaped contact with the frontals. An internasal depression is present, and an internasal foramen is possibly present (Figure 7G-7H); the dorsal surface of each nasal is strongly convex. The nasal contacts the frontal posteriorly and prefrontal laterally.
Lacrimal. The lacrimal is excluded from the external narial opening in lateral view by the prefrontal (Figure 7C-7D). The lacrimal is a blocky element, narrowing anteriorly with a scalene triangle shape in lateral view; it contacts the prefrontal dorsally and the jugal ventrally. The suborbital process is short. The lacrimal does not contact the maxilla but converges with the jugal and the prefrontal anterior to the orbit and posterior to the narial opening. The lacrimal is anteroposteriorly longest at the level of the sclerotic aperture. The anterior tip of the lacrimal appears to be laterally deflected relative to the posterior part of the element, and the portion making up the circumorbital area is more strongly curved than the ventral edge of the element.
Jugal. The jugal is deeply bowed ventral to the orbit. It tapers anteriorly between the lacrimal and the maxilla, extending past the anterior edge of the circumorbital area. Posteriorly, the jugal is overlapped by the postorbital. The posterior ramus of the jugal is offset approximately 120º from the ventral one, being wider and shorter with a flat external surface. The dorsal margin is evenly curved, whereas the ventral one forms a small projection at the posteroventral orbit (Figure 7C-7D).
Prefrontal. The prefrontal is a large element forming the anterodorsal orbital rim, with an anterior process reaching the external narial opening and forming its entire posterior edge. The prefrontal contacts the anterior tip of the jugal (Figure 7C-7D). The body of the prefrontal is massive, being as broad in dorsal view as the nasal (Figure 7G-7H). In lateral view, the anterior portion forming the orbital rim tapers ventrally and is considerably thinner than the posterior portion. Posterodorsally, the prefrontal contacts the postfrontal, parietal, and frontal (medially).
Frontal. The frontals are small and elongate, and are dorsally convex. They enclose the parietal foramen medially (Figure 7G-7H). The frontal is overlapped by the nasal (anteriorly), prefrontal (laterally), and parietal (posteriorly).
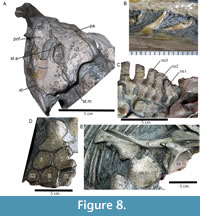 Parietal. The parietal has an irregular external surface (Figure 7G-7H, Figure 8A). There is a depression between the medial supratemporal fenestra and interparietal suture, possibly corresponding to the area lateral to the sagittal eminence. The parietal shelf is large and oriented dorsally. The parietal ridge is weakly developed. Posterolaterally, the parietal has a small process, which contacts the supratemporal posteriorly. The parietal contacts the prefrontal (anteriorly), postfrontal (laterally), and frontal (anterolaterally).
Parietal. The parietal has an irregular external surface (Figure 7G-7H, Figure 8A). There is a depression between the medial supratemporal fenestra and interparietal suture, possibly corresponding to the area lateral to the sagittal eminence. The parietal shelf is large and oriented dorsally. The parietal ridge is weakly developed. Posterolaterally, the parietal has a small process, which contacts the supratemporal posteriorly. The parietal contacts the prefrontal (anteriorly), postfrontal (laterally), and frontal (anterolaterally).
Postfrontal. The postfrontal is anteroposteriorly elongate and has an anteromedial depression in dorsal view (Figure 7G-7H, Figure 8A). Its posteromedial edge encloses the supratemporal fenestra and is overlapped by the supratemporal. The postfrontal becomes narrower anteriorly until it forms a point that overlaps the prefrontal anteriorly. In lateral view, the postfrontal is slightly arched and forms the dorsolateral edge of the posterior skull. The postfrontal extends as far anteriorly as the anterior parietal but is narrower than the latter, and extends anteriorly to the orbital midpoint (Figure 7C).
Postorbital. The postorbital forms the posterior edge of the orbit (Figure 7C). A small dorsal lamella appears to be present. In lateral view, the postorbital is wider than the jugal, but still narrow in relation to its height. The ventral postorbital overlaps the jugal, and is excluded from the posteroventral edge of the orbit by the latter. The dorsal postorbital is overlapped by the postfrontal. The dorsal and ventral ends of the postorbital are tapered, forming a semilunate shape.
Supratemporal. The supratemporal is posteriorly broad and slightly convex dorsally (Figure 7G-7H, Figure 8A). A long anterior ramus excludes the postfrontal from the lateral edge of the supratemporal fenestra. The supratemporal is anteroposteriorly shorter than mediolaterally broad. A descending ramus is present but not clearly exposed. The supratemporal bears four ridges radiating from the posterolateral corner (Figure 8A); we use the term ‘palmate’ to refer to this morphology.
Quadratojugal. The quadratojugal is small, triangular, and articulates along its entire anterior edge with the postorbital (Figure 7C).
Orbit. The sclerotic ring consists of 16 plates. It is 88 mm long and 79 mm high; the sclerotic aperture is 38.5 mm high and 35.5 mm long and is elliptical in shape. The orbit is 96 mm long and 85 mm high and is teardrop-shaped in lateral view when the circumorbital area is also considered. The orbit is bordered by the prefrontal, postfrontal, postorbital, jugal, and lacrimal (Figure 7C-7D).
Quadrate and Palate
There is an element covered by the postorbital in right lateral view, which may be the quadrate based on position and shape.
The palate is not exposed.
Mandible
Dentary. The dentary is a robust element that tapers anteriorly (Figure 7C-7D). The anterior tip is missing. The posterior end forms a thin point, articulating ventrally with the surangular at the level of the anterior orbit. The dentary becomes progressively deeper ventral to the external narial opening until it makes up the entire lateral surface of the lower jaw. The dentary is slightly convex in lateral view along the entire occlusal edge. The dentary fossa is present along its lateral surface.
Angular. The angular is short and does not extend as far anteriorly as the surangular in lateral view (Figure 7D). In ventral view, the posterior end of the angular is slightly expanded with rounded edges, while the anterior end tapers to a sharp point, and disappears from ventral view at approximately the level of the maxilla, just posterior to the symphysis (Figure 7E-7F). The ventral edge of the angular is not strongly curved in lateral view. However, the lower jaw appears slightly bowed ventral to the orbit (Figure 7C-7D).
Splenial. Posteriorly, the splenial contacts the medial surface of the angular, becoming broader in ventral view as the angular thins (Figure 7E-7F). The splenial is thin and convex in ventral view. The posterior end is elongated and pointed. The anterior end is not preserved. The broadest point of the splenial in ventral view occurs at the posterior end of the mandibular symphysis.
Surangular. The surangular is an elongate and large element that forms a significant part of the posterior end of the lower jaw (Figure 7C-7D). It is strongly curved ventral to the orbit. The posterior end is broad and ovate in lateral view, with a sinusoidal dorsal margin, formed in part by a small coronoid process. The anterior contact with the dentary is elongate, almost one-third of the length of the surangular in lateral view. The surangular fossa is deep. Anteriorly, the surangular is excluded in ventral view by the splenial and the dentary at approximately the same point as the mandibular symphysis (Figure 7E-7F).
Prearticular. The prearticular is only preserved on the right side. It is laterally compressed and elongate, positioned medial to the angular and lateral to the posteriormost splenial.
Dentition. The teeth are small and acutely pointed, with smooth, thin enamel lacking macroscopic ornamentation (Figure 8B). There is no pronounced constriction between the crown and root. The roots are not well-enough preserved to assess the presence or absence of apicobasal ridges, however, the root is smooth immediately basal to the crown. Crown height is 4 mm at the midpoint of the dentary. The teeth extend as far posteriorly as the anterior orbital margin.
Hyoid apparatus. The left ceratobranchial is preserved (Figure 7E-7F). An element in the position of the right hyoid is of questionable identity.
Postcranial Axial Skeleton
The atlas-axis complex is not visible. The third cervical centrum (C3) is the first vertebral centrum exposed on the right side. The right and left sides of the atlantal neural spine are unfused, and the atlantal neural spine is much smaller and shorter than the axial neural spine (Figure 8C). The axial neural spine is twice as wide as that of C3 and more posterior dorsal neural spines, with an anterolateral facet for articulation with the atlantal spine (Figure 7D). Postzygapophyses appear to be paired until at least C7. Anterior dorsal centra bear paired apophyses. Vertebrae are taphonomically compressed.
The anteriormost dorsal ribs are bicapitate, but the tuberculum and capitulum are less clearly separated than in more posterior ribs (Figure 7B). Ribs bear an anterodorsal groove; this groove does not extend to the distal half of the rib. Some partial gastralia are preserved immediately posterior to the forelimb (Figure 7A).
Pectoral Girdle
Clavicle. The clavicle is wing-like, broadening medially and concave posteriorly with a thin dorsal process. It is twisted ventrally from the middle of the axis in such a way that the lateral ramus is offset approximately 120º from the medial one (Figure 8E). The medial ramus is more robust, broad, and convex than the lateral ramus. Whereas the medial ramus is slightly curved, the lateral one is flattened on both its dorsal and ventral surfaces.
Scapula. The anterior blade is concave in external view. An anterior expansion is present (Figure 8E). The coracoid facet is smaller than the glenoid facet. The posterior shaft of the scapula is dorsally straight and mediolaterally compressed. The posterior shaft is curved ventrally and slightly flared at its distal end. The ventral edge is proximodistally shorter than the dorsal edge.
Coracoid. The coracoid is saddle-shaped in external view, with the glenoid and scapular facets thickened and projecting ventrally. The coracoid is more or less equidimensional and rounded, and is almost as long as the median stem of the interclavicle (Figure 8E). An anterior notch is present but the posterior notch is reduced or absent. The glenoid facet is slightly rugose and much larger than the scapular facet. The intercoracoid facet is extremely reduced.
Interclavicle. The interclavicle is ‘T’-shaped. The posterior stem is approximately as long as the transverse bar. The transverse bar is relatively wide and convex in external view (Figure 8E). In anterior view, the transverse bar is broad and tapers laterally towards the tips.
Forefin
Humerus. The distal end of the humerus is wider than the proximal end, but not significantly so (Table 1; Figure 8D), and a mid-shaft constriction is present. The proximal anterior surface of the humerus is flattened. The dorsal process lies towards the anterior edge and is relatively small, extending less than half the length of the humeral shaft. In anterior view, it can be seen that the deltopectoral crest extends further distally than the dorsal process. The ulnar facet is longer than the radial facet. The anterior distal edge is slightly squared off, forming a prominent leading edge facet. The ulnar facet is straighter in dorsal view, whereas the radial facet is more concave.
Distal limb elements. The ulna and radius are in broad contact. The radius is notched anteriorly, and bears distal articular facets for the radiale and intermedium; the facet for the former is longer than that of the latter. The ulna is pentagonal in outline, with distal facets for the intermedium and ulnare. The pisiform is absent in FWD-129. The intermedium is pentagonal in outline, and forms a large contact with distal carpal 3. The ulnare is very small and proximodistally short, smaller than the intermedium.
Hauffiopteryx sp., Figure 9
 SMNS 81367, SMNS 80225, SMNS 81962, and SMNS 81965 are referable to Hauffiopteryx sp., but the diagnostic portions of the skull are either not exposed or are damaged (Figure 9).
SMNS 81367, SMNS 80225, SMNS 81962, and SMNS 81965 are referable to Hauffiopteryx sp., but the diagnostic portions of the skull are either not exposed or are damaged (Figure 9).
SMNS 81367 (Figure 9A) is a small individual with the skull preserved in dorsolateral and the postcranium preserved in ventrolateral views. The specimen is clearly referable to Hauffiopteryx based on the small, round supratemporal fenestrae, the palmate morphology of the supratemporal, and the location of the parietal foramen anterior to the anterior edge of the supratemporal fenestra. Extensive reconstruction in the narial region prevents an accurate assessment of morphology and thus specific referral. This specimen was figured as H. typicus by Pardo-Pérez et al. (2019).
SMNS 80225 (Figure 9B) is a poorly preserved specimen prepared in ventrolateral view. The skull has been partially reconstructed. This specimen is referred to Hauffiopteryx sp. based on the number and posterior extent of the gastralia. Fragmentary pelvic elements suggest the absence of a robust ischiopubis, also consistent with Hauffiopteryx. This specimen was considered to be referable to H. typicus by Maisch (2008).
SMNS 81962 (Figure 9C) is preserved in ventral view. The anterior rostrum has been reconstructed. This specimen is referred to Hauffiopteryx based on the reniform outline of the quadrate in lateral view, palmate supratemporal, and the hind limb with digits converging distally.
SMNS 81965 (Figure 9D) is also preserved in ventral view; the rostrum has been reconstructed. This specimen is considered to be referable to Hauffiopteryx based on the extensive development of gastralia, and the medially unfused ischium and pubis. An ossified hyoid corpus is present, a feature thusfar only observed in Hauffiopteryx among ichthyosaurs (Motani et al. 2013). SMNS 81965 was considered to be referable to H. typicus by Maisch (2008).
PHYLOGENETIC ANALYSIS
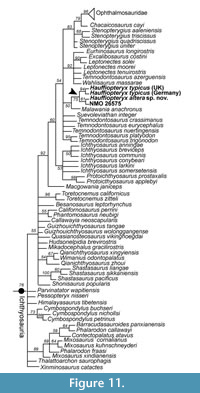
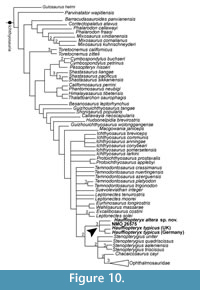 In order to test the relationship between material referred to Haffiopteryx typicus from the UK and Germany, Hauffiopteryx altera sp. nov., NMO 26575, and Leptonectes tenuirostris, we scored these as five separate OTUs in a version of the Moon (2017) Ichthyosauria matrix modified based on Maxwell et al. (2019). The Hauffiopteryx material from England was scored from the literature (Caine and Benton, 2011; Marek et al., 2015); NMO 26575 was scored based on personal observation (EM) and Maisch and Reisdorf (2006). In addition, we added the taxa Protoichthyosaurus prostaxalis and P. applebyi, scored based on Lomax et al. (2017, 2019a, 2019b) and Lomax and Massare (2018). Additional character and scoring changes are outlined in Appendix 2-Appendix 3; the nexus file is provided in Appendix 4. We analyzed the data using maximum parsimony implemented in TNT v. 1.5 (Goloboff and Catalano, 2016) with a New Technology search (100 iterations of the ratchet algorithm, minimum length found 100 times) (Figure 10). Following parsimony analysis, the resulting MPTs were analyzed using the IterPCR script (Pol and Escapa, 2009), and 12 taxa causing instability were pruned from the strict consensus tree. Among neoichthyosaurian taxa, this resulted in Malawania anachronus, Temnodontosaurus eurycephalus, Pervushovisaurus bannovkensis, P. campylodon, and Simbirskiasaurus birjukovi being pruned (see Appendix 2 for complete list). A Bayesian analysis (Figure 11) was executed in Mr. Bayes. Search settings and taxon selection are outlined in Maxwell et al. (2019).
In order to test the relationship between material referred to Haffiopteryx typicus from the UK and Germany, Hauffiopteryx altera sp. nov., NMO 26575, and Leptonectes tenuirostris, we scored these as five separate OTUs in a version of the Moon (2017) Ichthyosauria matrix modified based on Maxwell et al. (2019). The Hauffiopteryx material from England was scored from the literature (Caine and Benton, 2011; Marek et al., 2015); NMO 26575 was scored based on personal observation (EM) and Maisch and Reisdorf (2006). In addition, we added the taxa Protoichthyosaurus prostaxalis and P. applebyi, scored based on Lomax et al. (2017, 2019a, 2019b) and Lomax and Massare (2018). Additional character and scoring changes are outlined in Appendix 2-Appendix 3; the nexus file is provided in Appendix 4. We analyzed the data using maximum parsimony implemented in TNT v. 1.5 (Goloboff and Catalano, 2016) with a New Technology search (100 iterations of the ratchet algorithm, minimum length found 100 times) (Figure 10). Following parsimony analysis, the resulting MPTs were analyzed using the IterPCR script (Pol and Escapa, 2009), and 12 taxa causing instability were pruned from the strict consensus tree. Among neoichthyosaurian taxa, this resulted in Malawania anachronus, Temnodontosaurus eurycephalus, Pervushovisaurus bannovkensis, P. campylodon, and Simbirskiasaurus birjukovi being pruned (see Appendix 2 for complete list). A Bayesian analysis (Figure 11) was executed in Mr. Bayes. Search settings and taxon selection are outlined in Maxwell et al. (2019).
Both Bayesian and parsimony analyses support referral of NMO 26575 and FWD-129 to the genus Hauffiopteryx, distinct from all three species of Leptonectes (Figure 10-Figure 11). Moreover, material referred to Hauffiopteryx typicus from Germany and England formed a monophyletic group. We consider scoring differences between these H. typicus OTUs to be caused by differences in taphonomy (three-dimensional preservation vs. strong lateral compression), variation in the quality of preservation and preparation, and ontogeny, with the British material thought to represent juvenile individuals. Hauffiopteryx formed a sister group to Stenopterygius + Ophthalmosauridae in parsimony analysis (Figure 10); its position relative to Stenopterygius spp. and Leptonectes spp. was unresolved in Bayesian analysis (Figure 11).
DISCUSSION
Comparison of Hauffiopteryx to Other Early Jurassic Ichthyosaurs
 Hauffiopteryx differs from all coeval Toarcian ichthyosaur genera in the extensive participation of the prefrontal in the external narial opening. The prefrontal is excluded in Temnodontosaurus trigonodon, Stenopterygius (Maisch, 1998a); Suevoleviathan (Maisch, 2001), Eurhinosaurus (SMNS 18648), as well as in Leptonectes (McGowan and Motani, 2003). The nasals in Hauffiopteryx are more extensively exposed dorsally anterior to the external narial opening than the premaxillae, unlike in Stenopterygius (e.g., Maxwell, 2012b). As in Eurhinosaurus (e.g., SMNS 18648), the upper temporal fenestrae are small and circular, the supratemporal is broad and palmate in morphology, and the parietal shelf is horizontally oriented. As in Temnodontosaurus trigonodon (Fraas, 1913), the splenial plays a major role in the mandibular symphysis. The teeth of Hauffiopteryx are slender and conical, with smooth enamel unlike in Suevoleviathan, in which the enamel is rugose in texture (Maxwell, 2018). In the postcranium, the gastralia in Hauffiopteryx extend posteriorly at least as far as the thirty-fifth vertebra. This is unique among Toarcian genera: gastralia are unknown in Temnodontosaurus trigonodon and Suevoleviathan; in Eurhinosaurus and Stenopterygius gastralia are present in the anterior- to mid-dorsal region (Buchholtz, 2001). The rib tuberculum and capitulum in the dorsal region are widely separated in Hauffiopteryx, as in Stenopterygius but unlike in Temnodontosaurus trigonodon, Eurhinosaurus, and Suevoleviathan (Maisch and Matzke, 2000). The anterior scapula is expanded and a foramen between the humerus, radius, and ulna is absent, unlike in Temnodontosaurus trigonodon (e.g., SMNS 17560). The elements of the anterior digit in both the fore- and hind limbs bear notches, and the proximal limb elements are angular and tightly interlocking, unlike in Suevoleviathan (Maisch, 1998b). The ischium and the pubis are thin and styloidal, unlike in Temnodontosaurus, Suevoleviathan, and Eurhinosaurus (Maisch and Matzke, 2000), but are widely separated medially, unlike in Stenopterygius in which they are fused both medially and laterally (Maisch and Matzke, 2000).
Hauffiopteryx differs from all coeval Toarcian ichthyosaur genera in the extensive participation of the prefrontal in the external narial opening. The prefrontal is excluded in Temnodontosaurus trigonodon, Stenopterygius (Maisch, 1998a); Suevoleviathan (Maisch, 2001), Eurhinosaurus (SMNS 18648), as well as in Leptonectes (McGowan and Motani, 2003). The nasals in Hauffiopteryx are more extensively exposed dorsally anterior to the external narial opening than the premaxillae, unlike in Stenopterygius (e.g., Maxwell, 2012b). As in Eurhinosaurus (e.g., SMNS 18648), the upper temporal fenestrae are small and circular, the supratemporal is broad and palmate in morphology, and the parietal shelf is horizontally oriented. As in Temnodontosaurus trigonodon (Fraas, 1913), the splenial plays a major role in the mandibular symphysis. The teeth of Hauffiopteryx are slender and conical, with smooth enamel unlike in Suevoleviathan, in which the enamel is rugose in texture (Maxwell, 2018). In the postcranium, the gastralia in Hauffiopteryx extend posteriorly at least as far as the thirty-fifth vertebra. This is unique among Toarcian genera: gastralia are unknown in Temnodontosaurus trigonodon and Suevoleviathan; in Eurhinosaurus and Stenopterygius gastralia are present in the anterior- to mid-dorsal region (Buchholtz, 2001). The rib tuberculum and capitulum in the dorsal region are widely separated in Hauffiopteryx, as in Stenopterygius but unlike in Temnodontosaurus trigonodon, Eurhinosaurus, and Suevoleviathan (Maisch and Matzke, 2000). The anterior scapula is expanded and a foramen between the humerus, radius, and ulna is absent, unlike in Temnodontosaurus trigonodon (e.g., SMNS 17560). The elements of the anterior digit in both the fore- and hind limbs bear notches, and the proximal limb elements are angular and tightly interlocking, unlike in Suevoleviathan (Maisch, 1998b). The ischium and the pubis are thin and styloidal, unlike in Temnodontosaurus, Suevoleviathan, and Eurhinosaurus (Maisch and Matzke, 2000), but are widely separated medially, unlike in Stenopterygius in which they are fused both medially and laterally (Maisch and Matzke, 2000).
Parsimony analysis resolved the position of Hauffiopteryx as sister to Stenopterygius + Ophthalmosauridae (Figure 10). Although poor resolution outside this group increases ambiguity in character reconstruction, this clade is supported by a narrow, tapering anterior jugal, the basioccipital contributing to the floor of the foramen magnum in posterior view, a supraoccipital with parallel exoccipital processes, a caudal region approximately equal in length to the presacral vertebral column, one complete postaxial digit supported by an ulnare with two well-developed distal articular facets, polygonal metacarpals and phalanges, and lateral fusion of the ischiopubis. The strongly grooved ribs with broadly separated tuberculum and capitulum in the mid-dorsal region are also different from all leptonectids (Maisch and Matzke, 2000).
In the skull, greater similarities to leptonectids are present - especially in the dermatocranium. These include the thin rostrum with elongated external narial opening, the small upper temporal fenestra, the enlarged, palmate supratemporal, the horizontally oriented parietal slope, and the slight overbite. However, unexpected similarities with Temnodontosaurus (e.g., SMNS 50000) are also identified in the skull: the anteroposterior orientation and large size of the extracondylar area (ECA) of the basioccipital rather than the ECA being dorsoventrally oriented, such as that of Eurhinosaurus (SMNS 18648) and to a lesser extent Leptonectes tenuirostris and Wahlisaurus (Lomax and Massare, 2012; Lomax, 2016), and the extensive participation of the splenial in the mandibular symphysis (see Fraas, 1913). These differences support an independent derivation of a longirostrine morphotype in Leptonectidae and Hauffiopteryx, as argued by Moon (2017), although unlike Moon we recover T. azerguensis as a leptonectid in the Bayesian analysis (Figure 11).
Maisch (2008) was of the opinion that Hauffiopteryx typicus was characterized by certain unusual ontogenetic changes, namely: 1) strong positive allometry of the antorbital rostrum relative to the skull with increasing body size, and 2) strong positive allometry of the skull relative to presacral length. Unfortunately, the rostrum appears to have been reconstructed/modified in most of the available specimens of H. typicus, with the exception of GPIT 1491/4, GPIT/RE/12905, SMNS 80226, MHH 9, and BRSLI M1399 (see Table 2 for raw measurements). Based on log-transformed data, the antorbital rostrum appears to have grown with weak positive allometry relative to the lower jaw (slope = 1.15), and the lower jaw grew with strong negative allometry relative to the presacral vertebral column, the humerus, and to a lesser extent, the femur (as reported for e.g., Stenopterygius quadriscissus and Ichthyosaurus communis: McGowan, 1973). Thus, at present there is some support for the view that antorbital rostrum of Hauffiopteryx typicus showed a different allometric growth pattern than Ichthyosaurus and Stenopterygius, although there is enough variation present that this pattern is not clearly apparent in the ratios of individual specimens.
Ontogenetic Status of FWD-129 and Generic Referral of Hauffiopteryx altera
FWD-129 shows multiple features consistent with Hauffiopteryx from both the Posidonienschiefer Formation as well as from coeval localities in England, but differs from Stenopterygius, the most abundant and widespread ichthyosaur genus during the Toarcian. Characters supporting the referral of FWD-129 to Hauffiopteryx include participation of the prefrontal in the posterior external narial opening, small, round upper temporal fenestrae situated posterior to the pineal foramen, short nasals that do not contact the parietals, and a forefin with tightly interlocking, polygonal phalanges with a notch on the elements of the leading edge digit (Maisch, 2008; Caine and Benton, 2011). This combination of features is consistent with Hauffiopteryx among all ichthyosaurian genera, and this referral is also supported by both parsimony and Bayesian phylogenetic analysis (Figure 10-Figure 11). However, FWD-129 differs from H. typicus based on some aspects of cranial anatomy.
Maisch (2008) considered individuals of Hauffiopteryx typicus of less than 2.5 m total length to be juveniles. Thus, using Maisch’s criterion, with a total length of 193 cm (Table 2) GPIT 1491/4 would be a juvenile; however, we suspect that the specimen is osteologically mature based on the convex shape of the proximal humerus and closure of the sutures between phalanges (Johnson, 1977). Caine and Benton (2011) estimated total length in BRLSI M1399 at ~150 cm (skull length = 322 mm; humerus = 47 mm [from photo: Caine and Benton, 2011: text-fig. 9]); this specimen has been generally interpreted as a juvenile (Caine and Benton, 2011; Marek et al., 2015).
FWD-129 was initially considered to be a juvenile of Eurhinosaurus longirostris, due to its small size, large orbits, and small temporal fenestra (Jäger, 2005). FWD-129 is approximately equal in size to BRLSI M1399, with a lower jaw at least 280 mm in length and a humerus 46 mm in length (Table 2). The proximal articular surface of the humerus is flat, and the sutures between the limb elements remain open, also strongly supporting a juvenile attribution for this specimen. Relative proportions of the skull to the postcranium (McGowan, 1973), proportions within the skull (especially pertaining to the orbit/sclerotic ring: Fernández et al., 2005), relative tooth size (Dick and Maxwell, 2015), and notching of the anterior limb elements (Maxwell et al., 2014) are known to be ontogenetically influenced in the closely related genus Stenopterygius. However, the characteristics separating FWD-129 from specimens referred to H. typicus, Stenopterygius, and Eurhinosaurus are not expected to vary ontogenetically. This is especially apparent given that FWD-129 is of a similar ontogenetic stage to BRLSI M1399 based on both absolute size and limb ossification, but shows numerous differences in cranial morphology.
Comparison of Hauffiopteryx altera with Hauffiopteryx typicus
The lectotype of H. typicus (GPIT 1491/4) from Holzmaden is severely crushed, whereas FWD-129 is a lightly compressed three-dimensional skeleton. Nevertheless, it is possible to identify ways in which the two skulls differ. In H. typicus, the orbit is very large and round, creating a steep inflection in the nasal in lateral view. In FWD-129, the orbit is more elongate, and in lateral view the nasals are less steeply inflected anterior to the orbit than in H. typicus. SMNS 51552 and MHH 9 (Figure 2B, 2C) are much more similar in shape to the lectotype of H. typicus (GPIT 1491/4; Figure 2A) than to FWD-129 (Figure 7C). Unlike in GPIT 1491/4, MHH 9, and SMNS 51552, the lacrimal of FWD-129 does not separate the prefrontal from the jugal, and is excluded from the external narial opening. The lacrimal of FWD-129 is more robust than in specimens referred to H. typicus. These differences result in a relatively larger distance between the anterior edge of the orbit and the narial opening in FWD-129 than in H. typicus. The maxilla is overlapped dorsally by the lacrimal over almost one third of the posterodorsal surface, and the maxilla forms the ventral edge of much of the external nares in the lectotype, MHH 9, and SMNS 51552. In FWD-129, it is the jugal rather than the lacrimal that overlaps the maxilla posteriorly, and the maxilla is almost entirely excluded from the external narial opening in lateral view.
BRLSI M1399 has a shallower, more elongated maxilla than FWD-129, but the maxilla of FWD-129 has a greater contact with the ventral surface of the premaxilla than that of BRLSI M 1399, with the premaxilla extending well past the midpoint of the external nares in the former specimen. The maxilla of FWD-129 is slightly deeper posteriorly, whereas in the English specimen the maxilla is rather similar in depth along its entire length. The maxilla of FWD-129 contacts the lacrimal and prefrontal posterodorsally, whereas in BRLSI M1399 the maxilla contacts only the lacrimal. The lacrimal of BRLSI M1399 participates posteriorly in the external narial opening (Marek et al., 2015: fig. 1; Caine and Benton, 2011: fig. 9); however, in FWD-129 it is excluded from the external narial opening by the prefrontal. In dorsal view, the prefrontal in FWD-129 is anteroventrally narrower than that of BRLSI M1399. Also, the narial ramus is more anteroposteriorly oblique in relation to the oval orbit. The jugal extends much further anteriorly in FWD-129 than in BRLSI M1399 (Caine and Benton, 2011: fig. 12; Marek et al., 2015: fig. 1). The orbit is also rounder in BRLSI M1399. These features are very similar to the set of characters differentiating FWD-129 from H. typicus, consistent with our interpretation of BRLSI M1399 as referable to H. typicus.
Marek et al. (2015) cited the absence of root striations, the absence of the narial process of the maxilla, the presence of the temporal process of the frontal, wing-like basipterygoid processes, the absence of a ventral notch in the extracondylar area of the basioccipital, and the rod-like ischium/ischiopubis as differing between Hauffiopteryx and BRLSI M1399. However, some of these conclusions are the result of character scoring errors: BRLSI M1399 has a ventral notch in the extracondylar area of the basioccipital (Marek et al., 2015: fig. 10a), and lacks participation of the frontal in the supratemporal fenestra (i.e., processus temporalis of the frontal absent, Marek et al., 2015: fig. 4a). The only character listed that truly differs between H. typicus and BRLSI M1399 is the absence of root striations in the latter; however, Marek et al. (2015) state that the teeth are quite poorly preserved, which may affect character scoring. For that reason, we consider BRLSI M1399 to be referable to H. typicus. Hauffiopteryx typicus from England and Germany formed a monophyletic group in our analyses, to the exclusion of NMO 26575 and Hauffiopteryx altera sp. nov. (Figure 9-Figure 10).
Comparison of Hauffiopteryx typicus to NMO 26575
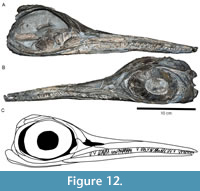 Maisch and Reisdorf (2006) referred a small ichthyosaur from the late Pliensbachian of Switzerland to the much older Rhaetian-Sinemurian species Leptonectes tenuirostris (Figure 12). However, this specimen shares greater similarity with the stratigraphically less far-removed genus Hauffiopteryx than with Leptonectes tenuirostris based on its small size, cranial proportions, significant participation of the prefrontal in the posterodorsal edge of the external nares, and angular, closely packed limb elements with multiple notches along the anterior digit of the forelimb. NMO 26575 is distinct from Hauffiopteryx altera in that the jugal is quite short, and the prefrontal does not exclude the lacrimal from the posterior edge of the external nares. NMO 26575 is more similar to H. typicus in this and most respects, and we consider it to be referable to H. typicus, resulting in a range extension of at least 2 million years for both the genus and the species. One of the key differences between most skulls referred to H. typicus and NMO 26575 is the proximity between the anterior lacrimal and subnarial process of the premaxilla in the latter, excluding the maxilla from participation in the external narial opening. However, the morphology observed in GPIT/RE/12905, in which the anterior lacrimal contacts the subnarial process of the premaxilla to exclude the maxilla from the narial opening, suggests that the anterior process of the lacrimal is variable in length in H. typicus and that NMO 26575 is within the range of variation of this species. Instraspecific variation in this part of the skull has already been noted in other Early Jurassic ichthyosaurs: e.g., in Protoichthyosaurus prostaxalis (Lomax et al., 2019a), and Stenopterygius triscissus (compare Godefroit, 1994: fig. 19; Caine and Benton, 2011: text-fig. 3).
Maisch and Reisdorf (2006) referred a small ichthyosaur from the late Pliensbachian of Switzerland to the much older Rhaetian-Sinemurian species Leptonectes tenuirostris (Figure 12). However, this specimen shares greater similarity with the stratigraphically less far-removed genus Hauffiopteryx than with Leptonectes tenuirostris based on its small size, cranial proportions, significant participation of the prefrontal in the posterodorsal edge of the external nares, and angular, closely packed limb elements with multiple notches along the anterior digit of the forelimb. NMO 26575 is distinct from Hauffiopteryx altera in that the jugal is quite short, and the prefrontal does not exclude the lacrimal from the posterior edge of the external nares. NMO 26575 is more similar to H. typicus in this and most respects, and we consider it to be referable to H. typicus, resulting in a range extension of at least 2 million years for both the genus and the species. One of the key differences between most skulls referred to H. typicus and NMO 26575 is the proximity between the anterior lacrimal and subnarial process of the premaxilla in the latter, excluding the maxilla from participation in the external narial opening. However, the morphology observed in GPIT/RE/12905, in which the anterior lacrimal contacts the subnarial process of the premaxilla to exclude the maxilla from the narial opening, suggests that the anterior process of the lacrimal is variable in length in H. typicus and that NMO 26575 is within the range of variation of this species. Instraspecific variation in this part of the skull has already been noted in other Early Jurassic ichthyosaurs: e.g., in Protoichthyosaurus prostaxalis (Lomax et al., 2019a), and Stenopterygius triscissus (compare Godefroit, 1994: fig. 19; Caine and Benton, 2011: text-fig. 3).
Palaeoecology
The palaeoecology of the morphologically similar genus Stenopterygius is relatively well understood, and includes dietary information for individuals over a wide range of body sizes (Dick et al., 2016), complete ontogenetic series (von Huene, 1922), and embryonic material (Böttcher, 1990). In contrast, Hauffiopteryx is much less abundant. The material listed in the current contribution represents the complete sample of Hauffiopteryx known from publicly accessible collections. Preserved individuals fall into a much narrower size range than Stenopterygius (Table 2 vs. Maxwell, 2012b supplementary table 1) and embryos are unknown, limiting our understanding of ontogenetic variation in this genus. Recognizable prey items are lacking as gastric contents. Thus, how Hauffiopteryx may functionally differ from Stenopterygius is extremely speculative.
Although referral of NMO 26575 to Hauffiopteryx typicus extends the stratigraphic range of the genus and species into the upper Pliensbachian, all finds from the Posidonienschiefer Formation are concentrated in a very narrow stratigraphic range, from the uppermost tenuicostatum Zone to the lowermost serpentinum Zone. This time period corresponds to the onset to the peak of the early Toarcian Oceanic Anoxic Event, an event that profoundly influenced composition and abundance of both vertebrate and invertebrate faunas in the southwest German Basin (e.g., Hauff, 1921; Maxwell and Vincent, 2016). It is thus possible that Hauffiopteryx had a particularly specialized diet or hunting strategy, and its sudden reappearance in - and subsequent disappearance from - the basin was related to changes in prey abundance or habitat (e.g., bottom-water anoxia forcing fish and belemnites into a narrow zone near the sea surface: Ullmann et al., 2014).
CONCLUSIONS
We found no evidence that the English and German material referred to the genus Hauffiopteryx represent separate species; however, we do find evidence that two species of Hauffiopteryx were present in the Early Jurassic of the Southwest German Basin: H. typicus and H. altera sp. nov. These two taxa are differentiated primarily based on characters pertaining to the lacrimal, prefrontal, and jugal. In addition, we refer a specimen previously considered to be Leptonectes tenuirostris to Hauffiopteryx typicus, extending the range of the genus into the Pliensbachian. This result is supported by phylogenetic analysis, which recovers Hauffiopteryx as a sister group to Stenopterygius + Ophthalmosauridae. Hauffiopteryx represents a valid genus, defined by a suite of synapomorphies involving both the skull and postcranium. However, the functional and paleoecological significance of these characters differentiating Hauffiopteryx from the superficially similar genus Stenopterygius are unclear.
ACKNOWLEDGEMENTS
We would like to thank A. Schmid-Röhl (FWD), I. Werneburg (GPIT), R. Hauff (MHH), and D. Steinmann (NMO) for collections access, and C. Gascó Martín for partial preparation of SMNS 80226.DC thanks C. Jaramillo (STRI), the Smithsonian Tropical Research Institute, the Anders Foundation, the 1923 Fund, G.D. and J.W. Johnson, an NSERC CREATE 466283-2015, and an NSERC Discovery Grant to H. Larsson (McGill University, Canada), for funding and support, and J. Pardo-Pérez (SMNS, U. Magallanes, Chile) and C. Gascó Martín (SMNS) for discussion and logistical support. The suggestions of A. Wolniewicz, N. Zverkov, and an anonymous reviewer improved the manuscript.
REFERENCES
Böttcher, R. 1989. Über die Nahrung eines Leptopterygius (Ichthyosauria, Reptilia) aus dem süddeutschen Posidonienschiefer (Unterer Jura) mit Bemerkungen über den Magen der Ichthyosaurier. Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie), 155:1-19.
Böttcher, R. 1990. Neue Erkenntnisse über die Fortpflanzungsbiologie der Ichthyosaurier (Reptilia). Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie), 164:13-51.
Buchholtz, E.A. 2001. Swimming styles in Jurassic ichthyosaurs. Journal of Vertebrate Paleontology, 21:61-73.
Caine, H. and Benton, M.J. 2011. Ichthyosauria from the Upper Lias of Strawberry Bank, England. Palaeontology, 54:1069-1093. https://doi.org/10.1111/j.1475-4983.2011.01093.x
de Blainville, H.M.D. 1835. Description de quelques espèces de reptiles de la Californie: précédée de l’analyse d’un système général d’erpétologie et d’amphibiologie. Nouvelles Annales du Muséum d'Histoire Naturelle, Paris, 4:233-296.
Dick, D.G. and Maxwell, E.E. 2015. Ontogenetic tooth reduction in Stenopterygius quadriscissus (Reptilia: Ichthyosauria): negative allometry, changes in growth rate, and early senescence of the dental lamina. PloS ONE, 10:e0141904. https://doi.org/10.1371/journal.pone.0141904
Dick, D.G., Schweigert, G., and Maxwell, E.E. 2016. Trophic niche ontogeny and palaeoecology of Early Toarcian Stenopterygius (Reptilia: Ichthyosauria). Palaeontology, 59:423-431. https://doi.org/10.1111/pala.12232
Fernández, M.S., Archuby, F., Talevi, M., and Ebner, R. 2005. Ichthyosaurian eyes: paleobiological information content in the sclerotic ring of Caypullisaurus (Ichthyosauria, Ophthalmosauria). Journal of Vertebrate Paleontology, 25:330-337. https://doi.org/10.1671/0272-4634(2005)025[0330:IEPICI]2.0.CO;2
Fischer, V., Bardet, N., Benson, R.B., Arkhangelsky, M.S., and Friedman, M. 2016. Extinction of fish-shaped marine reptiles associated with reduced evolutionary rates and global environmental volatility. Nature Communications, 7:10825. https://doi.org/10.1038/ncomms10825
Fraas, E. 1913. Einunverdrückter Ichthyosaurus-Schädel. Mitteilungen aus dem K. Naturalienkabinett, 79:1-12.
Godefroit, P. 1994. Les reptiles marins du Toarcien (Jurassique inferieur) belgo-luxembourgeois. Mémoires pour servir à l'Explication des Cartes Géologiques et Minières de la Belgique, 39:1-98.
Goloboff, P. and Catalano, S. 2016. TNT, version 1.5, with a full implementation of phylogenetic morphometrics. Cladistics, 32(3):221-238. https://doi.org/10.1111/cla.12160
Hänggi, H. and Reisdorf, A. 2007. Der Ichthyosaurier vom Hauensteiner Nebelmeer. Naturforschende Gesellschaft des Kantons Solothurn Mitteilungen, 2007:7-22.
Hauff, B. 1921. Untersuchung der Fossilfundstätten von Holzmaden im Posidonienschiefer des Oberen Lias Württembergs. Palaeontographica, 64:1-42.
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT, a free program for phylogenetic analysis. Cladistics, 24:774-786. https://doi.org/10.1111/j.1096-0031.2008.00217.x
Huelsenbeck, J. P. and Ronquist, F. 2001. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics, 17:754-755.
Jäger, M. 2005. The Museum of Fossils in the Werkforum: Guidebook of the Exhibition of Jurassic Fossils (third edition). Holcim, Dotternhausen, Germany.
Ji, C., Jiang, D.-Y., Motani, R., Rieppel, O., Hao, W.-C., and Sun, Z.-Y. 2016. Phylogeny of the Ichthyopterygia incorporating recent discoveries from South China. Journal of Vertebrate Paleontology, 36:e1025956. https://doi.org/10.1080/02724634.2015.1025956
Johnson, R. 1977. Size independent criteria for estimating relative age and the relationships among growth parameters in a group of fossil reptiles (Reptilia: Ichthyosauria). Canadian Journal of Earth Sciences, 14:1916-1924.
Lomax, D.R. 2016. A new leptonectid ichthyosaur from the Lower Jurassic (Hettangian) of Nottinghamshire, England, UK, and the taxonomic usefulness of the ichthyosaurian coracoid. Journal of Systematic Palaeontology, 15:387-401. https://doi.org/10.1080/14772019.2016.1183149
Lomax, D.R. and Massare, J.A. 2012. The first reported Leptonectes (Reptilia: Ichthyosauria) with associated embryos, from Somerset, England. Paludicola, 8:263-276.
Lomax, D.R. and Massare, J.A. 2018. A second specimen of Protoichthyosaurus applebyi (Reptilia: Ichthyosauria) and additional information on the genus and species. Paludicola, 11:164-178.
Lomax, D.R., Massare, J.A., and Evans, M. 2019a. New information on the skull roof of Protoichthyosaurus (Reptilia: Ichthyosauria) and intraspecific variation in some dermal skull elements. Geological Magazine, 157:640-650. https://doi.org/10.1017/S0016756819001225
Lomax, D.R., Massare, J.A., and Mistry, R.T. 2017. The taxonomic utility of forefin morphology in Lower Jurassic ichthyosaurs: Protoichthyosaurus and Ichthyosaurus. Journal of Vertebrate Paleontology, 37:e1361433. https://doi.org/10.1080/02724634.2017.1361433
Lomax, D.R., Porro, L.B., and Larkin, N.R. 2019b. Descriptive anatomy of the largest known specimen of Protoichthyosaurus prostaxalis (Reptilia: Ichthyosauria) including computed tomography and digital reconstruction of a three-dimensional skull. PeerJ 7: e6112. https://doi.org/10.7717/peerj.6112
Maisch, M.W. 1998a. Kurze Übersicht der Ichthyosaurier des Posidonienschiefers mit Bemerkungen zur Taxonomie der Stenopterygiidae und Temnodontosauridae. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 209:401-431.
Maisch, M.W. 1998b. A new ichthyosaur genus from the Posidonia Shale (Lower Toarcian, Jurassic) of Holzmaden, SW-Germany with comments on the phylogeny of post-Triassic ichthyosaurs. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 209(1):47-78.
Maisch, M.W. 2001. Neue Exemplare der seltenen Ichthyosauriergattung Suevoleviathan Maisch 1998 aus dem Unteren Jura von Südwestdeutschland. Geologica et Palaeontologica, 35:145-160.
Maisch, M.W. 2008. Revision der Gattung Stenopterygius Jaekel, 1904 emend. von Huene, 1922 (Reptilia: Ichthyosauria) aus dem Unteren Jura Westeuropas. Paleodiversity, 1:227-271.
Maisch, M.W. and Matzke, A.T. 2000. The Ichthyosauria. Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie), 298:1-159.
Maisch, M.W. and Reisdorf, A.G. 2006. Evidence for the longest stratigraphic range of a post-Triassic ichthyosaur: a Leptonectes tenuirostris from the Pliensbachian (Lower Jurassic) of Switzerland. Geobios, 39:491-505. https://doi.org/10.1016/j.geobios.2005.04.005
Marek, R.D., Moon, B.C., Williams, M., and Benton, M.J. 2015. The skull and endocranium of a Lower Jurassic ichthyosaur based on digital reconstructions. Palaeontology, 58:723-742. https://doi.org/10.1111/pala.12174
Maxwell, E.E. 2012a. Unraveling the influences of soft‐tissue flipper development on skeletal variation using an extinct taxon. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, 318:545-554. https://doi.org/10.1002/jez.b.22459
Maxwell, E.E. 2012b. New metrics to differentiate species of Stenopterygius (Reptilia: Ichthyosauria) from the Lower Jurassic of southwestern Germany. Journal of Paleontology, 86:105-115.
Maxwell, E.E. 2018. Redescription of the 'lost' holotype of Suevoleviathan integer (Bronn, 1844) (Reptilia: Ichthyosauria). Journal of Vertebrate Paleontology, 38(2):e1439833. https://doi.org/10.1080/02724634.2018.1439833
Maxwell, E.E., Cortés, D., Patarroyo, P., and Parra Ruge, M.L. 2019. A new specimen of Platypterygius sachicarum (Reptilia: Ichthyosauria) from the Early Cretaceous of Colombia and its phylogenetic implications. Journal of Vertebrate Paleontology, 39: e1577875. https://doi.org/10.1080/02724634.2019.1577875
Maxwell, E.E., Scheyer, T.M., and Fowler, D.A. 2014. An evolutionary and developmental perspective on the loss of regionalization in the limbs of derived ichthyosaurs. Geological Magazine, 151:29-40. https://doi.org/10.1017/S0016756812001070
Maxwell, E.E. and Vincent, P. 2016. Effects of the early Toarcian Oceanic Anoxic Event on ichthyosaur body size and faunal composition in the Southwest German Basin. Paleobiology, 42:117-126. https://doi.org/10.1017/pab.2015.34
McGowan, C. 1973. Differential growth in three ichthyosaurs: Ichthyosaurus communis, I. breviceps, and Stenopterygius quadriscissus (Reptilia, Ichthyosauria). Life Sciences Contributions, Royal Ontario Museum, 93:1-21.
McGowan, C. 1978. Further evidence for the wide geographical distribution of ichthyosaur taxa (Reptilia: Ichthyosauria). Journal of Paleontology, 52:1155-1162.
McGowan, C. 1992. Unusual extensions of the neural spines in two ichthyosaurs from the Lower Jurassic of Holzmaden. Canadian Journal of Earth Sciences, 29:380-383.
McGowan, C. and Motani, R. 2003. Ichthyopterygia. Verlag Dr. Friedrich Pfeil, München.
Moon, B.C. 2017. A new phylogeny of ichthyosaurs (Reptilia: Diapsida). Journal of Systematic Palaeontology, 17:129-155. https://doi.org/10.1080/14772019.2017.1394922
Motani, R. 1999. Phylogeny of the Ichthyopterygia. Journal of Vertebrate Paleontology, 19:473-496.
Motani, R., Ji, C., Tomita, T., Kelley, N., Maxwell, E., Jiang, D., and Sander, P.M. 2013. Absence of suction feeding ichthyosaurs and its implications for Triassic mesopelagic paleoecology. PloS ONE, 8:e66075. https://doi.org/10.1371/journal.pone.0066075
Owen, R. 1859. On the orders of fossil and recent Reptilia, and their distribution in time. Report for the British Association for the Advancement of Science, Aberdeen, 1859:153-166.
Pardo-Pérez, J., Kear, B.P., and Maxwell, E.E. 2019. Palaeoepidemiology in extinct vertebrate populations: factors influencing skeletal health in Jurassic marine reptiles. Royal Society Open Science, 6:190264. https://doi.org/10.1098/rsos.190264
Pol, D. and Escapa, I.H. 2009. Unstable taxa in cladistic analysis: identification and the assessment of relevant characters. Cladistics, 25:515-527. https://doi.org/10.1111/j.1096-0031.2009.00258.x
Ronquist, F. and Huelsenbeck, J.P. 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics, 19:1572-1574. https://doi.org/10.1093/bioinformatics/btg180
Theodori, C. 1843. Über einen kolossalen Ichthyosaurus trigonodon. Gelehrte Anzeigen der königlich bayerischen Akademie der Wissenschaften, 16:906-911.
Ullmann, C.V., Thibault, N., Ruhl, M., Hesselbo, S.P., and Korte, C. 2014. Effect of a Jurassic oceanic anoxic event on belemnite ecology and evolution. Proceedings of the National Academy of Sciences, 111:10073-10076. https://doi.org/10.1073/pnas.1320156111
Urlichs, M., Wild, R., and Ziegler, B. 1994. Der Posidonien-Schiefer und seine Fossilien. Stuttgarter Beiträge zur Naturkunde C, 36:1-95.
von Huene, F.R. 1922. Die Ichthyosaurier des Lias und ihre Zusammenhänge. Gebrüder Borntraeger, Berlin.
von Huene, F.R. 1931. Neue Studien über Ichthyosaurier aus Holzmaden. Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, 42:345-382.
