

 Amphibians and squamate reptiles from the late Miocene of Fălciu (Eastern Romania)
Amphibians and squamate reptiles from the late Miocene of Fălciu (Eastern Romania)
Article number: 25.2.a19
https://doi.org/10.26879/1156
Copyright Paleontological Society, June 2022
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Appendix
Submission: 24 March 2021. Acceptance: 30 May 2022
ABSTRACT
A moderately diverse fossil amphibian and squamate reptile assemblage, consisting of various fragmentary cranial bones, vertebrae and appendicular elements, is reported from the late Miocene (Turolian, MN 11-12) of Fălciu-Prut 1 locality. The fossil assemblage represents a lacustrine taphonomic context, where the skeletal remains reached the burial place after a short transport from a terrestrial environment. The amphibians are represented by salamanders (Chelotriton sp., and Triturus sp.) and anurans (Latonia sp., Pelobates sp., Bufotes cf. viridis and Pelophylax sp.), whereas the squamates by lizards (Lacertidae sp. indet. 1 and 2; Anguinae sp. indet. 1 and 2) and snakes (Scolecophidia indet., Natrix sp., Colubridae indet. and Naja cf. romani). The fauna inhabited probably shrubberies and open woodlands amid mild temperate climate conditions; permanent water sources were present in the area. Zoogeographical links were possible along the northern margin of Paratethys for both the European and Asian faunal representatives. Compared to older and coeval faunas from that region the fauna suggests an environmental shift into a more open environment, resulted from climate aridification. The faunal turnover is highlighted by lack of amphibian groups permanently linked to large, well-oxygenated rivers (cryptobranchid salamanders and proteids) or marshlands (palaeobatrachid frogs) that were still present in older faunas.
Vlad A. Codrea. University Babeș-Bolyai Cluj-Napoca, Department of Geology, 1, Kogălniceanu Str., RO-40084 Cluj-Napoca, Romania. codrea_vlad@yahoo.fr
Marian Bordeianu. University Babeș-Bolyai Cluj-Napoca, Department of Geology, 1, Kogălniceanu Str., RO-40084 Cluj-Napoca, Romania. marian.bordeianu@outlook.com
Márton Venczel. University Babeș-Bolyai Cluj-Napoca, Department of Geology, 1, Kogălniceanu Str., RO-40084 Cluj-Napoca, Romania and Țării Crișurilor Museum, Department of Natural History, Armatei Române 1/A, RO-410087 Oradea, Romania. mvenczel@gmail.com
Key words: aridification; herpetofauna; Moldova region; Paratethys; Turolian
Final citation: Codrea, Vlad A., Bordeianu, Marian, and Venczel, Márton. 2022. Amphibians and squamate reptiles from the late Miocene of Fălciu (Eastern Romania). Palaeontologia Electronica, 25(2):a19. https://doi.org/10.26879/1156
palaeo-electronica.org/content/2022/3639-fossil-herps-from-falciu
Copyright: June 2022 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
Data on the late Miocene herpetofaunas and of other terrestrial vertebrates from the Eastern Dacian basin are extremely scarce being based mainly on scattered, fortuitous finds (Codrea et al., 2017a, 2017b). Distribution of the terrestrial groups of that area was largely influenced by the paleogeographic evolution of the Dacian basin. This foreland basin was shaped during the mid-Miocene collision of the Carpathians and East European Margin (Krézsek and Olariu, 2020), and it was connected to both the Eastern and Central Paratethys seaways (Rögl, 1999). As a result, the migration route for terrestrial vertebrates was possible only north to the shoreline of the Paratethys realm.
The late Miocene (Maeotian) transgression transformed the Carpathians Bend Zone into a vast brackish basin dominated by deltaic sedimentation (Jipa and Olariu, 2009; Palcu et al., 2018, Lazarev et al., 2020). However, an “Intra-Maeotian Event” was also recorded representing a continental phase (Palcu et al., 2018), followed by a transgression and lacustrine sedimentation (Krézsek and Olariu, 2020).
The Maeotian vertebrate locality Fălciu-Prut 1 (hereinafter, abbreviated FP 1) is situated on the territory of Fălciu village (Moldova region, Eastern Romania) on the right bank of the Prut River, just on the border between Romania and Republic of Moldova. It was discovered by one of the authors (VAC) during the recovery of a Quaternary steppe bison cow skull (Bison priscus) that was found by a fisherman on the right bank of Prut River after a flood from 2011. The Maeotian deposits are eroded and mostly covered by the Quaternary loess-like-sediments that are largely exposed in the Prut basin.
The microvertebrates, recovered from the brackish-lacustrine clayish-marls, consist of skeletal remains of urodeles, anurans, lizards, snakes and small mammals. The amphibian and squamate specimens are mostly fragmentary with some signs of erosion indicating that they reached the burial place through transportation from a nearby area. A number of isolated fish remains, consisting of more than one hundred pectoral, dorsal and suborbital fish spines and several hundreds of pharyngeal teeth that resulted from the same sediments, are published by Trif et al. (2021). The samples also contained fragmentary ostrich eggshells that, according to Mikhailov and Zelenkov (2020), should be assigned to the parataxonomic genus Struthiolithus.
The amphibians and squamate reptiles, being ectothermic vertebrates with relatively low mobility, thus indicating closely their environmental preferences (Böhme et al. 2006), are considered adequate for paleoecological reconstructions. In consequence, the discovery of the FP 1 fossil herpetofaunal assemblage may represent an important addition to our knowledge about the late Miocene evolution of these groups on the northern margin of the Eastern Paratethys, revealing the zoogeographical links and paleoecological preferences of the recovered taxa.
In the present paper we: 1) describe the amphibian and squamate remains, 2) compare the assemblage with other late Miocene herpetofaunas and 3) interpret the paleoecological conditions and evaluate the zoogeographical links of the studied fauna.
GEOLOGICAL SETTING
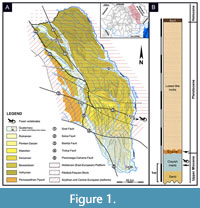 In Moldova region (Eastern Romania), two platforms occur: the Moldavian Platform (in fact, a local name for the south-western region of the East European Platform) and the Scythian Platform (Săndulescu, 1984). Differences between these structural units can be noticed concerning mainly their basements, which is Precambrian in the first one and a presumed infra-Paleozoic in the second. Other differences are also observed in the number of their sedimentary mega-sequences ('mega-cycles' or 'cycles', in Ionesi, 1994). Borehole data revealed that there are three mega-sequences in the Moldavian Platform (late Vendian-Devonian, Cretaceous-Palaeogene, middle-late Miocene) and four in the Scythian one (? pre-Devonian - Devonian, Permian-Triassic, Jurassic-Palaeogene, middle Miocene-Quaternary). However, in the Cenozoic mega-sequence their sedimentary history is similar in concern to lithology and facies distribution (Ionesi, 1994). Due to their monocline structure that is sloping from NW to SE, the deposition/erosion balance in Cenozoic was strongly influenced. In the Moldavian Platform the youngest deposits are Maeotian, whereas in the Scythian Platform this sedimentary history can be followed until the Pliocene and Pleistocene (Figure 1A). The Moldavian Platform exposes dominant deposits of marine-brackish origin, while the exposures on the Scythian one concern mainly continental (fluvial-lacustrine) formations. That means that across Miocene, the marine realm gradually receded, and the land replaced the sea mainly on the easternmost territories (Krézsek and Olariu, 2021). That explains why in the Moldavian Platform the final sedimentary mega-sequence refers only to Middle Miocene (late Badenian) - late Miocene (Maeotian), while in the Scythian Platform it is Middle Miocene (late Badenian) - Pleistocene (Ionesi, 1994).
In Moldova region (Eastern Romania), two platforms occur: the Moldavian Platform (in fact, a local name for the south-western region of the East European Platform) and the Scythian Platform (Săndulescu, 1984). Differences between these structural units can be noticed concerning mainly their basements, which is Precambrian in the first one and a presumed infra-Paleozoic in the second. Other differences are also observed in the number of their sedimentary mega-sequences ('mega-cycles' or 'cycles', in Ionesi, 1994). Borehole data revealed that there are three mega-sequences in the Moldavian Platform (late Vendian-Devonian, Cretaceous-Palaeogene, middle-late Miocene) and four in the Scythian one (? pre-Devonian - Devonian, Permian-Triassic, Jurassic-Palaeogene, middle Miocene-Quaternary). However, in the Cenozoic mega-sequence their sedimentary history is similar in concern to lithology and facies distribution (Ionesi, 1994). Due to their monocline structure that is sloping from NW to SE, the deposition/erosion balance in Cenozoic was strongly influenced. In the Moldavian Platform the youngest deposits are Maeotian, whereas in the Scythian Platform this sedimentary history can be followed until the Pliocene and Pleistocene (Figure 1A). The Moldavian Platform exposes dominant deposits of marine-brackish origin, while the exposures on the Scythian one concern mainly continental (fluvial-lacustrine) formations. That means that across Miocene, the marine realm gradually receded, and the land replaced the sea mainly on the easternmost territories (Krézsek and Olariu, 2021). That explains why in the Moldavian Platform the final sedimentary mega-sequence refers only to Middle Miocene (late Badenian) - late Miocene (Maeotian), while in the Scythian Platform it is Middle Miocene (late Badenian) - Pleistocene (Ionesi, 1994).
The tectonic relationship with the Eastern Carpathians orogen shows that both platforms are thrusted by the orogenic nappes and under their overload the platforms were faulted, dipping as distinct slabs from east to west (Ștefănescu et al., 1985). The boundary between the Moldavian and Scythian platforms refers to a major crustal fault named Bistrița fault, which continues in the Eastern Carpathians orogen (Săndulescu, 1984). Some authors preferred to name this fault by using the locality route that it follows in the Moldova region. Therefore, over time, either the name Fălciu-Plopana fault (Mutihac, 1990) or the name Fălciu-Munteni-Plopana fault (Ionesi, 1994) were wrongly used.
The vertebrate locality of FP 1 is situated on the Scythian Platform, just a few kilometers south to Bistrița fault. It can be found on the easternmost side of the region, right next to the border with the Republic of Moldova, being delimited by the latter one by the Prut River. The outcrop yielding vertebrate fossils represents a lacustrine deposit that is about 1 m thick (Figure 1B). The accumulation of this deposit may be associated with the continental phase of the so called “Intra-Maeotian Event” (Palcu et al., 2018), followed by a late Maeotian transgression, while the lacustrine sedimentation resumed (Krézsek and Olariu, 2021). The vertebrate yielding sediments are represented by brackish-lacustrine clayish-marls that overlay yellow sands (Trif et al., 2021). The top of this succession is represented by a 20-50 cm thick layer of Upper Pleistocene terrace pebble deposit, followed by a 7m loess cover (Figure 1B). The age of the vertebrate assemblage is supported also by the presence of small mammals like “ Proochotona ” cf. eximia Khomenko, 1914. The genus Proochotona has been referred by Čermák (2016) to the genus Ochotona with its oldest European occurrence, represented by O. kalfense, from the early late Miocene (MN 10) of Kalfa (Republic of Moldova) (Delinschi, 2014; Čermák, 2016). The type locality of Ochotona (= Proochotona) eximia is Tarakliya, Republic of Moldova (MN 12) (Khomenko, 1914), while its range (as O. eximia or O. cf. eximia) is restricted to the middle Turolian (MN 11-12) (Delinschi, 2014). The sediments have also yielded a high concentration of the ostracod Cyprideis torosa, which unfortunately is not an important biostratigraphic marker being recorded from the Miocene up to present. However, in the Dacian Basin this species formed massive accumulations in the upper Maeotian when the waters recorded an important desalination due to a regressive trend of the basin scale (Stoica et al., 2013). The fish fauna included among others the genera Carassius, Luciobarbus, Chondrostoma, Leuciscus, Scardinius, Tinca and Silurus (Trif et al., 2021). Taking into account the above data and considerations we attribute the continental age of these deposits to middle Turolian (MN 11-12).
MATERIAL AND METHODS
The studied samples were obtained from about 4.3 tons of clayish marl sediment, collected by two of the authors (VAC and MB) in four field missions (2018-2020). The sediment was sun-dried, hydrogen peroxide (3%) and water were added and subsequently sieved with a 0.3 mm mesh. The micro-vertebrates were sorted out from the remaining sediment using a binocular microscope. The disarticulated bones of salamanders, frogs, lizards and snakes were selected apart from other microvertebrates using taxonomic criteria. Assignment of the fossil remains to various taxa is based on direct comparisons with skeletal parts of recent and fossil taxa or on the identification of diagnostic characters published in relevant literature (see below). The photographs were captured with a Canon EOS 400 digital camera provided with a 60 mm macro lens. All the specimens are housed in the Babeş-Bolyai University Palaeontology-Stratigraphy Museum (= BBUPSM) in Cluj-Napoca.
The anatomical nomenclature follows Ivanov (2008) for caudates, Venczel (2004) for anurans, Venczel and Hír (2013) for lizards, and Szyndlar (1991a, 1991b) for snakes; throughout the paper, the standard orientation system is applied.
SYSTEMATIC PALAEONTOLOGY
CAUDATA Scopoli, 1777
SALAMANDRIDAE Gray, 1825
Genus Chelotriton Pomel, 1853
Chelotriton sp.
Figure 2
Material. UBB V 940/1-2 one fragmentary frontal, one fragmentary dentary; UBB V 939 one left oticooccipital; UBB V 938/1-4 three fragmentary presacral vertebrae, one fragmentary rib.
 Description. The frontal is represented by a small bony fragment preserving a prominent dorsal sculpture consisting of various sized tubercles of pustular type, whereas ventrally, a small part of the endocranial imprint is preserved (Figure 2D-E).
Description. The frontal is represented by a small bony fragment preserving a prominent dorsal sculpture consisting of various sized tubercles of pustular type, whereas ventrally, a small part of the endocranial imprint is preserved (Figure 2D-E).
The oticooccipital is well preserved, except the medial flange of tectum synoticum, which is broken off. The dorsal surface exposes the contact area for the squamosal, situated anterolaterally and that for the parietal placed medially; both areas are delimited by prominent ridges (Figure 2A). The medial part of the ventral surface is rugous representing the contact area for the basisphenoid. The posterior part bears the bipartite atlantal articular surface, consisting of a dorsoposteriorly placed occipital condyle and a shallower ventromedial cotyle to articulate with the atlantal cotyle laterally and with the atlantal odontoid process medially. In ventral view, the large fenestra ovalis (= fenestra vestibuli) projects laterally and slightly ventrally. Anterior to the fenestra ovalis there is a prominent basal process (Figure 2B). In medial view, several foramina are exposed: dorsally, two foramina may represent the endolymphatic foramen (placed anteriorly) and the combined glossopharyngeal-vagus nerve (IX + X) (placed posteriorly). The lower row of foramina may correspond to the facial nerve (VII), placed anteriorly, whereas the trifurcated auditory (= vestibulocochlear) nerve VIII is positioned medially, and the perilymphatic foramen exits posteriorly (Figure 2C).
The dentary is strongly damaged preserving only the medial part of the bone (Figure 2F-G). The labial surface exposes a strongly sculptured and slightly convex lower part and a smooth upper bony lamella representing the dental parapet bearing lingually the insertion surfaces of the small pleurodont teeth. On the lingual surface the Meckel’s canal is widely open posteriorly, whereas it narrows anteriorly; the subdental shelf is narrow and delimited by a sharp bony lamina.
The available vertebrae are opisthocoelous with their condyles short and incompletely delimited from the vertebral centrum (Figure 2I-K). The centrum is short and wide provided with long, double headed transverse processes positioned nearly perpendicular to the axis of the vertebral centrum; the subcentral lamina is wide and concave, pierced by a network of small foramina and bordered medially by a prominent subcentral keel. The prezygapophyses strongly project laterally, whereas the neural laminae in all the specimens are broken off.
The only rib that may be assigned to this taxon consists of a small piece of curved bone bearing a dorsal spine (Figure 2H).
Remarks. The skeleton of the members of Chelotriton is robustly built with some of the cranial bones (e.g., nasal, frontal, parietal, squamosal) bearing a strong sculpture of pustular type. The neural spine of the vertebrae is extremely high bearing also a pustular sculpture; the transverse processes (= rib bearers) are robustly built and project laterally, nearly perpendicular to the main body axis; the ribs are provided with dorsal processes (Ivanov, 2008). The positions of the auditory nerve (VIII) foramina and those of the endolymphatic and perilymphatic foramina in the only oticooccipital specimen are roughly similar to those reported in the specimens of Chelotriton sp. 1, from the early Miocene (MN 4) of Mokrá-Western Quarry, Czech Republic (Ivanov, 2008) and to that of Chelotriton sp., from the middle Miocene (MN 6) of Litke, N-Hungary (Venczel and Hír, 2015). Distribution of these foramina approaches also the condition known in Salamandra (Maddin et al., 2013: fig. 6C), where the perilymphatic foramen is positioned closely posterior to the vestibulocochlear nerve foramina, whereas the much smaller endolymphatic foramen is positioned dorsally to the vestibulocochlear nerve foramina. The sculpture preserved on the fragmentary frontal is different to that reported by Ivanov (2008) and by Venczel and Hír (2015) for Chelotriton, but it approaches the condition reported by Roček (2005) for specimens of Chelotriton sp., from the late Miocene (MN 9) of Rudabánya, N-Hungary. The robustly built vertebrae with short and wide centra, the strongly laterally projected prezygapophyses, the presence of massive and double-headed transverse processes projecting nearly perpendicular to the axis of the vertebral centrum, the short and incompletely separated anterior condyles are typical for the genus Chelotriton. The position and morphology of the dorsal spines on the ribs of Chelotriton is highly variable as reported by Roček (2019) in specimens assigned to Chelotriton paradoxus.
Genus Triturus Rafinesque, 1815
Triturus sp.
Figure 3
Material. UBB V 941 one fragmentary atlas; UBB V 942/1-3 two fragmentary presacral vertebrae, one caudal vertebra; UBB V 943/1-2 one proximal part of a femur, one fragmentary distal femur.
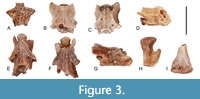 Description. The single atlas preserves the opisthocoelous centrum only. It is provided with two oval atlantal cotyles and the ventrally placed odontoid process; the subcentral surface is strongly sculptured by a network of small foramina. The vertebrae are opisthocoelous, elongated and provided with low and posteriorly bifurcated neural spines; the condyles are large and widened at their ends; the subcentral area is concave and pierced by several foramina of various size; the transverse processes are double headed. The posterior parts of the neural laminae and transverse process are broken off.
Description. The single atlas preserves the opisthocoelous centrum only. It is provided with two oval atlantal cotyles and the ventrally placed odontoid process; the subcentral surface is strongly sculptured by a network of small foramina. The vertebrae are opisthocoelous, elongated and provided with low and posteriorly bifurcated neural spines; the condyles are large and widened at their ends; the subcentral area is concave and pierced by several foramina of various size; the transverse processes are double headed. The posterior parts of the neural laminae and transverse process are broken off.
The two femurs are represented only by fragmentary examples. One of the specimens preserves the proximal part of a femur with a well-developed trochanter and crista trochanterica, whereas the other specimen preserves the distal part of a femur provided with a flattened and widened distal end.
Remarks. The morphology and size of the available specimens approach the condition seen in Triturus, especially those of the group Triturus cristatus. The neural spine is high in the members of Lissotriton, or extremely high in Carpathotriton, however, it is significantly of lower height in Triturus (Venczel, 2008). Nevertheless, in T. marmoratus the neural spine tends to be higher, whereas in the T. cristatus group is of low height (Ivanov, 2008), as is the case with specimens from FP 1. Both femurs represent medium-sized specimens with their proximal and distal ends moderately widened, a condition known in Triturus. In the members of the mostly terrestrial Salamandra and Chelotriton the femur is strongly widened proximally and distally, whereas in the fully aquatic Carpathotriton these extremities always remain slender (Venczel, 2008).
ANURA Rafinesque, 1815
ALYTIDAE Fitzinger, 1843
Genus Latonia Meyer, 1843
Latonia sp.
Figure 4A-B
Material. UBB V 944/1-2 two fragmentary ilia.
 Description. Only the proximal parts of the ilia are preserved representing two specimens of different sizes. The dorsal crest of the ilium is high, and the dorsal protuberance is embedded in the dorsal crest; the dorsal margin of the dorsal protuberance in both specimens is thickened. The preacetabular zone is extremely reduced, whereas the dorsal acetabular expansion and ischiatic process in both specimens is broken off.
Description. Only the proximal parts of the ilia are preserved representing two specimens of different sizes. The dorsal crest of the ilium is high, and the dorsal protuberance is embedded in the dorsal crest; the dorsal margin of the dorsal protuberance in both specimens is thickened. The preacetabular zone is extremely reduced, whereas the dorsal acetabular expansion and ischiatic process in both specimens is broken off.
Remarks. Within the alytids, in the members of Alytes the dorsal crests of the ilium is lacking, whereas Discoglossus and Latonia possess a high and thickened dorsal crest with a relatively small dorsal protuberance that is swollen along the posterodorsal part of the dorsal crest (Bailon, 1999: fig. 19a, j; Venczel and Hír, 2013). In both specimens the angle enclosed by the dorsal protuberance and the base of the dorsal acetabular expansion is extremely wide. In Discoglossus the posterior part of the dorsal protuberance arises steeply (Bailon, 1999: fig. 19a) and in consequence the angle enclosed by the dorsal acetabular expansion, and the dorsal protuberance is less wide than in Latonia (Rage and Hossini, 2000).
PELOBATIDAE Bonaparte, 1850
Genus Pelobates Wagler, 1830
Pelobates sp.
Figure 4C-J
Material. UBB V 945/1-2 one premaxilla, one frontoparietal fragment; UBB V 946/1-2 two fragmentary maxillae; UBB V 947 one fragmentary presacral vertebra.
Description. The premaxilla is well-preserved with moderately high and laterally curved dorsal process and relatively wide pars dentalis (Figure 4C); there are 15 tooth positions preserved; the palatine process is prominent and moderately long. The labial surface of the premaxilla is devoid of primary sculpture.
The frontoparietal preserves the left lateral part of the bone consisting of the laterally tapering processus lateralis superior and the orifice of the canalis arteriae temporalis (Figure 4E-F). The dorsal side is covered by a sculpture consisting of tubercle and irregular crests. The lateral superior process exposes a relatively thin articular surface for the dorsal process of the squamosal to form a fronto-squamosal arch.
The maxillae preserve the posterior part of the bone with a short, wide and mediolaterally flattened pterygoid process; posterior to the latter structure several foramina pierce the media wall of the maxilla (Figure 4G-H). The labial surface bears a primary sculpture consisting of crests and tubercles.
The angulosplenial consists of a small sinuous bone preserving the coronoid process attached to the medial side of the Meckel's groove. The posteriormost part of the angulosplenial, called the pars spatulaeformis praearticularis (Venczel et al., 2013) is not widened (Figure 4D).
The presacral vertebra is procoelous (the condyle is broken off) with elevated neural lamina of imbricate type, pierced by several small foramina; the lateral margins of this elevated part form a bony ridge extending above the postzygapophyses; the neural spine is low and shorter than the posterior margins of the postzygapophyses (Figure 4I-J). The lateral margins of the neural arch preserves in each side the foramen for exit of the spinal nerve. The remnant of the transverse process projects anterolaterally indicating that the vertebra represents a posterior presacral position.
Remarks. The moderately high and laterally curved dorsal process of the premaxilla is reminiscent of Pelobates syriacus (pers. obs.) and in a lesser degree of P. sanchizi (Venczel, 2004: fig. 6I), however, in the latter the dorsal process is tapering distally. In the recent P. fuscus the dorsal process is relatively short and shallowly bent laterally (Roček, 1981: fig. 25a, b). The relatively slender articular surface exposed by the processus lateralis superior of the frontoparietal indicates the presence of a relatively narrow fronto-squamosal arch, which is of variable width in recent P. syriacus, always narrow in the Pliocene P. praefuscus (Syromyatnikova, 2019), but distinctly wider in recent P. cultripes (Roček, 1981: fig. 57) and in the fossil P. sanchizi and P. decheni (Venczel, 2004); P. fuscus lacks a fronto-squamosal arch (Roček, 1981: fig. 57). The outer sculpture of the frontoparietal and maxillae consists of widened pits enclosed by bony crests and rear tubercles, reminiscent of P. syriacus and P. sanchizi. The presence of spinal nerve foramina in the presacral vertebrae of Pelobates is rarely present in P. syriacus (Sanchiz and Młynarski, 1979) and P. sanchizi (Venczel, 2004), but never reported in P. praefuscus (Syromyatnikova, 2019) and P. fuscus.
BUFONIDAE Gray, 1825
Genus Bufotes Rafinesque, 1815
Bufotes cf. viridis (Laurenti, 1768)
Figure 5A-F
Material. UBB V 948 one squamosal; UBB V 949/1-2 two fragmentary presacral vertebrae; UBB V 950/1-3 two fragmentary sacral vertebrae, one fragmentary urostyle; UBB V 951/1-10 ten humeri; UBB V 952/1-6 six ilia.
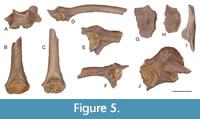 Description. The only squamosal specimen preserves a short end tapering zygomatic process, whereas the posterodorsal process is wide with the distal part somewhat damaged.
Description. The only squamosal specimen preserves a short end tapering zygomatic process, whereas the posterodorsal process is wide with the distal part somewhat damaged.
The presacral vertebrae have short, procoelous centra with flattened condyles and cotyles, whereas the neural lamina is also short, of non-imbricate type, with the neural spine reduced to a low keel. The sacral vertebrae are damaged preserving an anterior cotyle and two posterior condyles for a bicondylar articulation with the urostyle (Figure 5A). The only urostyle preserves the proximal part only; it has two cotyles and relatively robust dorsal crest thickened at its proximal end.
The humeri are preserved as distal fragments representing small-sized individuals (Figure 5B-C). The humeral ball usually is incomplete, placed nearly symmetrically between the medial and lateral epicondyles; there is a small ventral cubital fossa.
The ilia preserve the acetabular region and various portions from the ilial shaft. The acetabular fossa is large and of semicircular shape and with the acetabular rim projecting laterally. In the preacetabular zone there is a deep recess. The dorsal protuberance is prominent and variable in shape with several tuberosities compressed mediolaterally (Figure 5D-F). The ilial shaft lacks a dorsal crest; there is a transverse bony ridge extending from the anterior limit of the acetabular rim to the ventral margin of the ilial shaft. However, that ridge is devoid of a ventral extension to form the so called 'lamina calamita' or calamita ridge (Gómez and Turazzini, 2015).
Remarks. The most typical elements available to document the presence of Bufotes cf. viridis are the ilia. These bones display some variation in the position and morphology of the dorsal protuberance, which is asymmetrical consisting of two or three small tubercles, and positioned above or close to the level of the anterior margin of the acetabular rim. The preacetabular fossa in Bufotes viridis is always well-developed, whereas in Bufo bufo it is lacking (Bailon, 1999: 19l). The occurrence of a prominent bony ridge on the lateral surface of the ilial shaft is variable, but developed in a lesser degree than in the natterjack toad (Epidalea calamita). The presence of the calamita ridge is discernible in Strauchbufo raddei and in the fossil “Bufo” mirus (Syromyatnikova, 2015).
RANIDAE Linnaeus, 1758
Genus Pelophylax Fitzinger, 1843
Pelophylax sp.
Figure 5G-J
Material. UBB V 953/1-2 two fragmentary frontoparietals; UBB V 954/1-2 one premaxilla, one angulosplenial; UBB V 955/1-2 two humeri; UBB V 956 one fragmentary ilium.
Description. Two fragmentary frontoparietal specimens, preserving the posterior halves of the bones, are flattened dorsoventrally with their dorsal surface shallowly concave, and delimited laterally by a low crest; the dorsal surface preserves also several irregular superficial grooves; on the ventral side of the bone, two shallow concave bony surfaces represent the endocranial imprints.
The premaxilla preserves a relatively short and dorsally tapering dorsal process and a prominent, triangular shaped palatine process. The pars dentalis is damaged at its lateral part.
The body of the angulosplenial is sinuous, strongly compressed mediolaterally and provided with a shallow Meckelian groove. The coronoid process is wide, lamellar with its dorsal surface shallowly wrinkled.
The humeri preserve their distal parts only. The humeral shaft bears a relatively well-developed medial crest, and the medial epicondyle is larger than the lateral epicondyle.
The only fragmentary ilium preserves the acetabular region and the proximal part of the ilial shaft. The dorsal protuberance is flush with the dorsal margin of the dorsal crest and there is no thickening of this margin; there is a well-preserved deep supraacetabular fossa.
Remarks. The fragmentary frontoparietals, preserving on their dorsal surfaces a shallow network of grooves, are typical for large specimens of Pelophylax ridibundus; such a sculpture is lacking in the members of Rana (VM pers. obs.). The widened and lamellar coronoid process of the angulosplenial with its dorsal surface wrinkled is also typical for the genus Pelophylax (Bailon, 1999: fig. 9h). The dorsal protuberance of the ilium is prominent and compressed mediolaterally, however, the iliac shaft is broken off distally, and it is not possible to evaluate the height of the complete dorsal crest. The remaining bones are also reminiscent in size and morphology to those of the genus Pelophylax.
SQUAMATA Merrem, 1820
LACERTIDAE Bonaparte, 1831
Lacertidae sp. indet. 1.
Figure 6A-D
Material. UBB V 957/1-4 one fragmentary maxilla, three fragmentary dentaries; UBB V 958 one dentary.
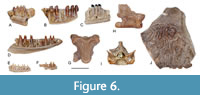 Description. The material is fragmentary, each specimen preserving a various number of teeth of pleurodont type. The outer surface of the only maxillary fragment lacks a labial sculpture. The dentary specimens represent different portions from the dentary. The specimen UBB V 957/2, representing an anterior portion of a dentary, preserves a relatively wide lamina horizontalis delimited ventrally by a narrow and open Meckel’s groove (Figure 6A); the subdental shelf is narrow and moderately deep. The tooth shaft is compressed mesiodistally, whereas the crowns are monocuspid and projected about one third of their height above the dental parapet. The tooth shafts in the specimens representing more posterior portions of the dentary (UBB V 957/3 and 4) are cylindrical, possessing bicuspid tooth crowns with the main cusp positioned distally (Figure 6B-C). The specimen UBB V 958 represents a nearly complete left dentary of a smaller individual (Figure 6D). The horizontal lamina is relatively thin, whereas the subdental shelf is shallow. The tooth shafts are cylindrical and moderately high. The preserved tooth crowns are bicuspid without any sign of striations.
Description. The material is fragmentary, each specimen preserving a various number of teeth of pleurodont type. The outer surface of the only maxillary fragment lacks a labial sculpture. The dentary specimens represent different portions from the dentary. The specimen UBB V 957/2, representing an anterior portion of a dentary, preserves a relatively wide lamina horizontalis delimited ventrally by a narrow and open Meckel’s groove (Figure 6A); the subdental shelf is narrow and moderately deep. The tooth shaft is compressed mesiodistally, whereas the crowns are monocuspid and projected about one third of their height above the dental parapet. The tooth shafts in the specimens representing more posterior portions of the dentary (UBB V 957/3 and 4) are cylindrical, possessing bicuspid tooth crowns with the main cusp positioned distally (Figure 6B-C). The specimen UBB V 958 represents a nearly complete left dentary of a smaller individual (Figure 6D). The horizontal lamina is relatively thin, whereas the subdental shelf is shallow. The tooth shafts are cylindrical and moderately high. The preserved tooth crowns are bicuspid without any sign of striations.
Remarks. The maxilla and dentary specimens, possessing closely spaced and cylindrical tooth shafts and provided with bicuspid tooth crowns are reminiscent of the recent genus Lacerta (e.g., L. agilis, L. viridis or L. trilineata) (Kosma, 2004). The relatively wide and rounded margin of the lamina horizontalis and the posteriorly open and deep Meckel’s groove is also comparable to the genus Lacerta. Compared to the recently reported fossil specimens from the late Miocene (MN 9) of Gritsev, Ukraine, the size and morphology of the larger specimens’ approach to those of Lacerta sp., type I., whereas the smaller specimen (UBB V 958) to that of Lacerta sp., type II, described by Roček (2019). Nevertheless, due to the fragmentary state of the available material a more resolved assignment of the above specimens is not possible.
Lacertidae sp. indet. 2.
Figure 6E-F
Material. UBB V 959/1-4 one fragmentary frontal, one fragmentary postorbital, two fragmentary dentaries.
Description. Both the frontal (a posterior fragment) and the postorbital (proximal part) are extremely fragmentary and covered with an outer sculpture. The dentaries preserve small portions from the middle portion (UBB V 959/3) and posterior portion (UBB V 959/4) of small-sized lacertid specimens (Figure 6E-F). The horizontal lamina is thin and the subdental shelf is shallow. The tooth shaft is cylindrical and moderately high projecting nearly half of their height above the dental parapet. The anterior tooth crowns apparently were monocuspid, those from the middle section bicupsid, whereas the posterior ones are tricupsid.
Remarks. The specimens differ from those of Lacertidae sp. indet. 1., by their smaller size and by the morphology of tooth crown in the posterior teeth displaying the tricuspid condition.
ANGUIDAE Gray, 1825
Anguinae sp. indet. 1.
Figure 6G-J
Material. UBB V 964/1-3 one vertebra, two osteoderms.
Description. In ventral view, the vertebral centrum appears shallowly convex, whereas the medial part of the subcentral area is elevated possessing a flat surface (Figure 6G). The synapophyses are undivided, whereas the interzygapophsyeal ridges weakly developed. The neural spine is relatively high with its anterior portion broken off (Figure 6H). In anterior view, the height of the neural canal is smaller than the dorso-ventral height of the cotyle (Figure 6I).
The osteoderms are subrectangular in shape and thickened in the middle (Figure 6J). The outer surface is sculptured with irregularly distributed grooves and vermicular tuberosities. The smooth area near the margins was covered by other osteoderms from bordering rows; there is no medial crest.
Remarks. The vertebra, provided with a relatively short and wide vertebral centrum, and with robustly built synapophyses is reminiscent of Pseudopus sp. from the late Miocene (MN 9) of Felsőtárkány basin in Northern Hungary (Venczel and Hír, 2013). According to Čerňansky et al. (2018, 2019), in the trunk vertebrae of Pseudopus the neural canal is always smaller than the height of the cotyle. The large-sized and robustly built osteoderm is also typical for the above genus. Abundant remains from the late Miocene (MN 9) of Gritsev have been assigned to P. pannonicus by Roček (2019). Owing to the scarcity of the available material a closer assignment of the specimens is not possible.
Anguinae sp. indet. 2.
Figure 7
Material. UBB V 960/1-4 one fragmentary maxilla, three fragmentary dentaries; UBB V 961/1-10 eight fragmentary presacral vertebrae, two fragmentary caudal vertebrae; UBB V 962/1-4 two vertebrae; two osteoderms; UBB V 963, 200 osteoderms.
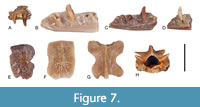 Description. The maxillary and dentary fragments represent small sized individuals. The tooth positions are situated close to each other suggesting closely packed teeth in the maxillae and dentaries. The dental parapet in the dentary is inclined lingually with a relatively wide insertion surface for the tooth bases. The tooth crown appears slightly compressed labiolingually and provided with mesiodistal crests. The subdental shelf is wide and shallow, whereas the horizontal lamina is extremely thin delimiting dorsally the widely open Meckel’s groove. The latter structure is open up to the first tooth position.
Description. The maxillary and dentary fragments represent small sized individuals. The tooth positions are situated close to each other suggesting closely packed teeth in the maxillae and dentaries. The dental parapet in the dentary is inclined lingually with a relatively wide insertion surface for the tooth bases. The tooth crown appears slightly compressed labiolingually and provided with mesiodistal crests. The subdental shelf is wide and shallow, whereas the horizontal lamina is extremely thin delimiting dorsally the widely open Meckel’s groove. The latter structure is open up to the first tooth position.
The vertebrae are relatively small sized with triangle-shape vertebral centrum and slightly convex subcentral area. The pre- and postzygapophyses are oval in shape, the interzygapophyseal ridges reduced, and the neural spine is reduced to a dorsal crest with a more elevated posterior spine. In anterior view (Figure 7H), the height of the neural canal is larger than the height of the cotyle.
The osteoderms are small, thin and elongated, bearing on the outer surface a sculptured area and a more elevated medial crest. Part of the osteodermal surface, covered in living animal by adjacent osteoderms, remains smooth.
Remarks. The presence of mesiodistal crests on the maxillary and dentary teeth, as well as the presence of slender osteoderms, provided with prominent medial crests prevent the association of the fossil material with Anguis or Pseudopus (both lacking mesiodistal crests on the tooth crowns and without elongated osteoderms). The neural canal in Anguinae sp. indet. 2 is always larger than the height of the cotyle, a feature reminiscent of the genus Ophisaurus (Čerňansky et al., 2018, 2019). Nevertheless, more material is needed to document the presence of the latter genus.
SERPENTES Linnaeus, 1758
SCOLECOPHIDIA Duméril and Bibron, 1844
Scolecophidia indet.
Figure 8A-B
Material. UBB V 965/1-2 two fragmentary vertebrae.
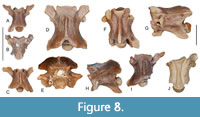 Description. The vertebrae are small sized with their vertebral centrum moderately long (Figure 8A-B). The neural is flattened, devoid of neural spine. The zygosphene in both specimens is damaged. The pre- and postzygapophyses are elongated, whereas the prezygapophyseal process is about the same length as that of the prezygapophysis. The subcentral surface is slightly convex and devoid of haemal keel. The synapophyses are globular and undivided.
Description. The vertebrae are small sized with their vertebral centrum moderately long (Figure 8A-B). The neural is flattened, devoid of neural spine. The zygosphene in both specimens is damaged. The pre- and postzygapophyses are elongated, whereas the prezygapophyseal process is about the same length as that of the prezygapophysis. The subcentral surface is slightly convex and devoid of haemal keel. The synapophyses are globular and undivided.
Remarks. The morphological characters of the trunk vertebrae are shared by the members of Typhlopidae, Leptotyphlopidae and Anomalepididae, therefore closer assignment of the available material is not possible (Szyndlar 1991a).
COLUBRIDAE Oppel, 1811
Genus Natrix Laurenti, 1768
Natrix sp.
Figure 8C-H
Material. UBB V 966/1-25 25 vertebrae, UBB V 967/1-5 five vertebrae.
Description. The centrum of the presacral vertebrae is relatively long, whereas the neural arch is moderately vaulted (Figure 8C-H). The neural spine in most specimens is broken off, and its remnants suggest that it was of relatively low height. The zygosphenal roof is more or less horizontal, whereas the anterior margin is shallowly convex. The apophyses are oval, whereas the prezygapophyseal processes, if preserved, are shorter than the length of the prezygapohyses; the posterior margin of the neural lamina is provided with a prominent epizygapophyseal spine. The subcentral surface is concave delimited laterally by the moderately developed subcentral ridges. The hypapophysis is sigmoid shaped, more prominent and directed posteroventrally in the anterior presacral vertebrae, whereas in the more posterior presacral vertebrae it is hooked backward with a rounded tip. The parapophyseal processes are long, flattened dorsoventrally with blunt anterior tip.
Remarks. The presacral trunk vertebrae display a combination of characters used to diagnose several members of the genus Natrix. Elongated vertebrae are typical for the fossil N. longivertebrata and for the recent N. natrix (Szyndlar, 1991b). In N. longivertebrata the epizygapophyseal spines are usually lacking (Szyndlar, 2005), whereas in those from FP 1 are always prominent. In both, N. natrix and N. longivertebrata the prezygapophyseal processes are moderately long, blunt and flattened dorsoventrally, in contrast part of the vertebrae from FP 1 have short prezygapophyseal processes, whereas others are with long processes as seen in N. rudabanyaensis, known from the late Miocene (MN 9) of Rudabánya, N-Hungary (Szyndlar, 2005). Moreover, the parapophyses of Natrix sp. from FP 1 are obtuse, whereas some of the hypapophyses have their ventral margin flattened (Figure 8F-G), never observed in the above fossil or recent forms.
Colubridae indet.
Figure 8I
Material. UBB V 968/1-3 three fragmentary vertebrae.
Description. The vertebrae are strongly damaged preserving only the ventral part of their centrum. The haemal keel is flattened and more or less parallel sided (Figure 8I). The paradiapophyses are weakly differentiated into parapophyseal and diapophyseal portions.
Remarks. The presence of a flattened haemal keel in the trunk vertebrae of several members of Colubridae (e.g., Coronella, Eirenis, Elaphe, Hierophis, Telescopus) (Szyndlar 1991a: fig. 8), prevent a closer assignment of the available material.
ELAPIDAE Boie, 1827
Genus Naja Laurenti, 1768
Naja cf. romani (Hoffstetter, 1939)
Figure 9
Material. UBB V 969 one venom fang; UBB V 970/1-5 four presacral vertebrae, one caudal vertebra.
Description. The venom fang is slightly curved anteroposteriorly and compressed laterally. On the anterior margin of the venom fang there is a continuous groove and an elongated foramen near the tip of tooth (Figure 9A).
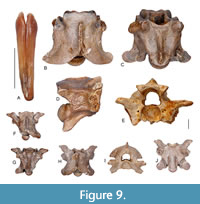 The vertebrae are robustly built with the neural arch moderately vaulted and preserving the remnants of low neural spines. The zygosphenal roof is shallowly convex with variable anterior margin. In the largest specimen there are two small lateral lobes preserved, whereas the medial one is with a medial concavity (Figure 9B-C); in a small specimen the zygosphene is crenate, with a distinct medial lobe (Figure 9E-F). The prezygapophysis is oval-shaped, whereas the prezygapophyseal process is about the same length as the diameter of the prezygapophysis. The hypapophysis is relatively short, robustly built and directed posteroventrally (Figure 9D). The paradiapophyses are well differentiated into parapophyseal and diapophyseal portions and are roughly of equal length. There is an extremely short parapophyseal process. The single caudal vertebra possesses an extremely short prezygapophyseal process; the pleurapophyses are broken off, whereas the haemal processes are short (Figure 9G-H).
The vertebrae are robustly built with the neural arch moderately vaulted and preserving the remnants of low neural spines. The zygosphenal roof is shallowly convex with variable anterior margin. In the largest specimen there are two small lateral lobes preserved, whereas the medial one is with a medial concavity (Figure 9B-C); in a small specimen the zygosphene is crenate, with a distinct medial lobe (Figure 9E-F). The prezygapophysis is oval-shaped, whereas the prezygapophyseal process is about the same length as the diameter of the prezygapophysis. The hypapophysis is relatively short, robustly built and directed posteroventrally (Figure 9D). The paradiapophyses are well differentiated into parapophyseal and diapophyseal portions and are roughly of equal length. There is an extremely short parapophyseal process. The single caudal vertebra possesses an extremely short prezygapophyseal process; the pleurapophyses are broken off, whereas the haemal processes are short (Figure 9G-H).
Remarks. The morphology of the venom fang and vertebrae are closely similar to those of Naja romani described from the late Miocene (MN 9) of Gritsev and Rudabánya, as well as to those known from the late Miocene of Kohfidisch (Szyndlar and Zerova, 1990; Szyndlar, 1991b, 2005). The morphological variations observed in the vertebra from FP 1 (Figure 9B-C) are that on the zygosphenal roof the medial lobe bears a concavity (typically it protrudes anteriorly) and the parapophysis is extremely short, which is usually longer and projecting anteriorly or anteroventrally (Szyndlar, 1991b). More material is needed from the locality of FP 1 in order to investigate if the above morphological variations should represent taxonomically important features.
DISCUSSION
Compared to the highly diversified fossil herpetofauna of Gritsev locality (MN 9) from the area (Appendix 1), the fossil fauna from FP 1 appears as a moderately diverse assemblage of urodeles, anurans and squamate reptiles indicating a lacustrine taphonomic context, the sediments being also rich in freshwater fish of the genera Carassius, Luciobarbus, Chondrostoma, Leuciscus, Scardinius, Tinca and Silurus (Trif et al., 2021). In the oryctocenosis part of the taxa were closely linked to aquatic or periaquatic ecosystems (Triturus, Chelotriton, Latonia, Pelophylax, Natrix), but fully aquatic groups (e.g., Cryptobranchidae, Proteidae, Palaeobatrachidae or Emydidae) were not identified (Böhme et al., 2006). Pelobates probably was nocturnal and preferred loose soil appropriate for digging a burrow in the vicinity of lakes, ponds or rivers; scolecophidian snakes, like the present-day Xerotyphlops vermicularis, known from the Mediterranean region, were also subterranean (Böhme et al., 2006). The remaining taxa, including lacertid and anguid lizards, probably preferred shrubberies and open woodlands and mild temperate climate conditions, whereas the cobra (Naja cf. romani) populated probably various habitats, including forests, rocky terrains and wetlands feeding on frogs, lizards and rodents.
Excepting Chelotriton, Latonia and Naja, the remaining genera of the assemblage are still present in Europe. The salamandrid Chelotriton, resembling morphologically the extant Asian genera Tylototriton and Echinotriton, had survived in Western Europe up to the Pleistocene (Bailon, 1989), whereas a very small population of the alytid frog Latonia nigriventer survived until present in Hula Valley, N-Israel (Biton et al., 2103). The genus Naja, had survived in both the Western and Eastern Mediterranean region up to the late Pliocene (Bailon, 1989; Szyndlar and Zerova, 1990; Rage, 2013).
The herpetofaunal assemblage from the late Miocene (Turolian, MN 11-12) of FP 1 shares a number of common taxa with those known from the late Miocene MN 9 - MN 13 interval of the northern side of Eastern Paratethys. Some relevant published data are available from Eastern Romania (Crețești, MN 9), Republic of Moldova (Bushor 1, MN 9; Cioburciu, MN 11; Cimislia, MN 12) Ukraine (Gritsev, MN 9; Cherevichnoje, MN 12) and Russia (Gaverdovsky, MN 11; Volchaya Balka, MN 11; and Solnechnodolsk, MN 13) (Appendix 1). All included localities were positioned within the main terrestrial faunal corridor extending across the northern margin of the Paratethys marine system between Europe and Asia, whereas the terrestrial links to southern faunas (i.e., Western Mediterranean region, Africa and Middle East) were mostly blocked by the Paratethys Sea. However, the major disparity between the above faunas may be related to their different geological ages and taphonomic context (i.e., karst taphonomic context in Gritsev vs. fluviatile in Crețești, vs. probably salmastric in Gaverdovsky; lacustrine in FP 1 and Volchaya Balka) (Codrea et al., 2017; Tesakov et al., 2017; Roček, 2019). Nevertheless, the intervening changes in the taxonomic nomenclature (e.g., Triturus, Rana, Coluber, Elaphe, Vipera), may affect direct comparisons with the taxon lists that probably were not updated (see Delinschi, 2009). Additionally, in some localities the list of taxa appears incomplete (e.g., in Volchaya Balka the list of lizards and snakes is not available).
The warm and wet climate during the early Vallesian (MN 9) favoured the spread of the fully aquatic cryptobranchids (Andrias and Ukrainurus) (Böhme and Ilg, 2003), proteid salamanders (Mioproteus) and palaeobatrachid frogs (Palaeobatrachus) (Roček, 2019). The mammalian megafauna was dominated by Suoidea, cervids, insectivores, rodents and various carnivores within a closed environment, whereas that of the late Vallesian (MN 10) was dominated by Giraffidae, Cervidae, Rhinocerotidae and Equidae corresponding to an open environment (Koufos, 2006); the middle Turolian (MN 12) environment was dominated by Bovidae, Proboscidea and some carnivorans (Koufos, 2006: fig. 5). The ostrich eggshells present in the samples from FP 1, described under the parataxonomic genus name Struthiolithus by Mikhailov and Zelenkov (2020: fig. 11), also present in the localities of Cioburciu (MN 11) and Cimislia (MN 12) (Delinschi, 2009), suggest the aridification of the paleoenvironment.
The major climate and environmental changes in the early Vallesian (MN 9) - middle Turolian (MN 12) interval seem obvious from the composition of the above herpetofaunas reflecting closely their paleoecologic preferences (Böhme et al., 2006). We may presume that in the oryctocenosis of the FP 1 locality the absence of groups linked permanently to aquatic habitats (Cryptobranchidae, Proteidae and Palaeobatrachidae), still present in the Gritsev assemblage, may have resulted from the missing well-oxygenated large rivers and marshlands from the area. Nevertheless, proteids and palaeobatrachids are still sporadically present in some of the localities from this region in the MN 9 - MN 12 interval (see Appendix 1). Another difference is the more diverse venomous snake assemblage in the herpetofauna of Gritsev, which included cobras (Szyndlar and Zerova, 1990), large and small vipers (Szyndlar, 1991b) and pit vipers (Ivanov, 1999), whereas the geologically younger faunas appear less diverse. The only other locality with cobras, beside Gritsev, is the FP 1 locality, while in the remaining localities only viperid taxa were recorded (see Appendix 1), reflecting the probable environmental shift into a more open environment resulted from aridification.
CONCLUSIONS
The herpetofaunal assemblage from the FP1 fossil locality (late Miocene, Turolian, MN 11-12) represents a lacustrine taphonomic context yielding six amphibians (Chelotriton sp., Triturus sp., Latonia sp., Pelobates sp., Bufotes cf. viridis and Pelophylax sp.) four lizards (Lacertidae sp. indet. 1 and 2; Anguinae sp. indet. 1 and 2) and four different snake taxa (Scolecophidia indet., Natrix sp., Colubridae indet., and Naja cf. romani).
Based on the fossil record from the late Miocene MN 9 - MN 13 interval, the common taxa from the FP1 locality together with those recorded from the northern side of Eastern Paratethys suggest the presence of a wide-ranging faunal migration route between Asia and Europe along the northern margin of Paratethys Sea.
The composition of the FP1 fauna suggests mild temperate climate conditions, a savannah-like landscape with shrubberies and open woodlands, and with some water sources in the area. However, compared to older faunas (e.g., Gritsev, see Appendix 1), the FP1 assemblage is lacking of several amphibian groups preferring well-oxygenated rivers (Cryptobranchidae and Proteidae) and marshlands (Palaeobatrachidae) that may suggest a shift into a more arid paleoenvironment.
The FP 1 fossil herpetofauna represents an important addition to our knowledge on the late Miocene evolution of these microvertebrates, and on the paleoenvironmental reconstruction of the northern side of Eastern Paratethys. Nevertheless, future research should include new material collected from the FP 1 locality, discovery of new localities and revision of the collections listed in the older contributions.
ACKNOWLEDGEMENTS
We thank the three anonymous reviewers for their constructive reviews that helped improving the manuscript. The executive editor, M. Hyžný, and the handling editor, J. Samuels provided valuable help during the editorial process. M. Sutton also provided technical help. MB addresses many thanks to N. Moraru (mayor of Fălciu) and to the Fălciu commune Hall for the granted logistic support to access the country border during the field campaigns from 2018-2020. This research was partially founded by Babeș-Bolyai University grants: AGC 30624, 30778.
REFERENCES
Bailon, S. 1989. Les amphibiens et les reptiles du Pliocène supérieur de Balaruc II (Hérault, France). Palaeovertebrata, 19:7-28.
Bailon, S. 1999. Différentiation ostéologique des anoures (Amphibia, Anura) de France, p. 1-38. In Desse, J. and Desse-Berset, N. (eds.), Fiches d’ostéologie animale pour l’archéologie, Série C: Varia. Centre de Recherches Archéologiques-CNRS, Valbonne, France.
Biton, R., Geffen, E., Vences, M., Cohen, O., Bailon, S., Rabinovich, R., Malka, Y., Oron, T., Boistel, R., Brumfeld, V., and Gafny, S. 2013. The rediscovered Hula painted frog is a living fossil. Nature Communications, 4:1959. https://doi.org/10.1038/ncomms2959
Boie, F. 1827. Bemerkungen über Merrem’s Versuch eines Systems der Amphibien. Erste Lieferung: Ophidier. Isis van Oken, 20:508-566.
Bonaparte, C.L. 1831. Saggio di una distribuzione metodica degli animali vertebrati. Antonio Boulzaler, Roma.
Bonaparte, C.L. 1850. Conspectus Systematum. Herpetologiae et Amphibiologiae. Editio altera reformata, E.J. Brill, Leiden.
Böhme, M. and Ilg, A. 2003. FosFARbase, www.wahre-staerke.com/ (accessed 22 July, 2021).
Böhme, M., Ilg, A., Ossig, A., and Küchenhoff, H. 2006. New method to estimate paleoprecipitation using fossil amphibians and reptiles and the middle and late Miocene precipitation gradients in Europe. Geology, 34:425-428. https://doi.org/10.1130/G22460.1
Čerňanský, A., Syromyatnikova, E.V., and Jablonski, D. 2018. The first record of amphisbaenian and anguimorph lizards (Reptilia, Squamata) from the upper Miocene Solnechnodolsk locality in Russia. Historical Biology, 32:869-879. https://doi.org/10.1080/08912963.2018.1539973
Čerňanský, A., Yaryhin, O., Ciceková, J., Werneburg, I., Hain, M., and Klembara, J. 2019. Vertebral comparative anatomy and morphological differences in anguine lizards with a special reference to Pseudopus apodus. The Anatomical Record, 302:232-257. https://doi.org/10.1002/ar.23944
Codrea, V., Venczel, M., and Ursachi, L., 2017a. Amphibians and squamates from the early Vallesian of Crețești (Vaslui County, E-Romania). Nymphaea, 44:37-56.
Codrea, V., Venczel, M., Ursachi, L., and Rățoi, B., 2017b. A large viper from the early Vallesian (MN 9) of Moldova region (Eastern Romania) with notes on the palaeobiogeography of late Miocene “Oriental vipers”. Geobios, 50:401-411. https://doi.org/10.1016/j.geobios.2017.07.001
Delinschi, A. 2009. Contribution to the study of Maeotian hipparion faunas from the Republic of Moldova. Oltenia Studii şi comunicări Ştiinţele Naturii, 25:391-395.
Delinschi, A. 2014. Late Miocene lagomorphs from the Republic of Moldova. Annales de Paléontologie, 100:157-163. https://doi.org/10.1016/j.annpal.2013.10.004
Duméril, A.M.C. and Bibron, G. 1844. Erpétologie générale ou histoire naturelle complète des Reptiles. Tome sixième. Libraire Encyclopédique de Roret, Paris.
Fitzinger, L.J.F.J. 1843. Systema Reptilium. Fasciculus Primus. Braumüller et Seidel, Wien.
Gómez, R.O. and Turazzini, G.F. 2015. An overview of the ilium of anurans (Lissamphibia, Salientia), with a critical appraisal of the terminology and primary homology of main ilial features. Journal of Vertebrate Paleontology, e1030023. https://doi.org/10.1080/02724634.2015.1030023
Gray, J.E. 1825. A synopsis of the genera of Reptiles and Amphibia, with a description of some new species. Annals of Philosophy, 10:193-217.
Haase, D., Fink, J., Haase, G., Ruske, R., Pécsi, M., Richter, H., Altermann, M., and Jäger, K.-D. 2007. Loess in Europe—its spatial distribution based on a European Loess Map, scale 1:2,500,000. Quaternary Science Reviews, 26:1301-1312.
Hoffstetter, R. 1939. Contribution a l'étude des Elapidae actuels et fossiles et de l'ostéologie des Ophidiens. Archives du Muséum d'Histoire Naturelle du Lyon, 15:1-78.
Ionesi, L. 1994. Geologia unităților de platformă și a Orogenului Nord-Dobrogean. Editura Tehnică, București.
Ivanov, M. 1999. The first European pit viper from the Miocene of Ukraine. Acta Palaeontologica Polonica, 44:327-334.
Ivanov, M. 2008. Early Miocene Amphibians (Caudata, Salientia) from the Mokrá-Western Quarry (Czech Republic) with comments on the evolution of Early Miocene amphibian assemblages in Central Europe. Geobios, 41:465-492. https://doi.org/10.1016/j.geobios.2007.11.004
Jipa, D.C. and Olariu, C. 2009. Dacian Basin. Depositional Architecture and Sedimentary History of a Paratethys Sea. Special Publication No. 3. GeoEcoMar, Bucharest.
Khomenko, J. 1914. La faune méotique du village Taraklia du district de Bendery. Travaux de la Société des Naturalistes et des Amateurs des Sciences Naturellesde Bessarabie, 5:1-55 [in Russian].
Kosma, R. 2004. The dentition of recent and fossil scincomorphan lizards (Lacertilia, Squamata). Systematics, functional morphology, paleoecology. Unpublished PhD Thesis, University of Hannover, Hannover, Germany.
Koufos, G.D. 2006. Palaeoecology and chronology of the Vallesian (late Miocene) in the Eastern Mediterranean region. Palaeogeography, Palaeoclimatology, Palaeoecology, 234:127-145. https://doi.org/10.1016/j.palaeo.2005.01.014
Krézsek, C. and Olariu, C. 2021. Filling of sedimentary basins and the birth of large rivers: The lower Danube network in the Dacian Basin, Romania. Global and Planetary Change 197:103-391. https://doi.org/10.1016/j.gloplacha.2020.103391
Lazarev, S., de Leeuw, A., Stoica, M., Mandic, O., van Baak, C.G.C., Vasiliev, I., and Krijgsman, W. 2020. From Khersonian drying to Pontian “flooding”: late Miocene stratigraphy and palaeoenvironmental evolution of the Dacian Basin (Eastern Paratethys). Global and Planetary Change, 192:103-224. https://doi.org/10.1016/j.gloplacha.2020.103224
Linnaeus, C. 1758. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. 10th Edition. Volume 1. L. Salvii, Stockholm.
Laurenti, J.N. 1768. Specimen Medicum, Exhibens Synopsin Reptilium Emendatum cum Experimentis Circa Venena et Antidota Reptilium Austriacorum. Johann Thomas von Trattnern, Wien.
Maddin, H.C., Venczel, M., Gardner, J.D., and Rage, J.-C. 2013. Micro-computed tomography study of a three-dimensionally preserved neurocranium of Albanerpeton (Lissamphibia, Albanerpetontidae) from the Pliocene of Hungary. Journal of Vertebrate Paleontology, 33:568-587. https://www.jstor.org/stable/42568541
Meyer, v. H. 1843. Summarische Uebersicht der fossilen Wirbelthiere des Mainzer Tertiär-Beckens, mit besonderer Rücksicht auf Weisenau. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefactenkunde, 1843:379-410.
Mikhailov, K.E. and Zelenkov, N. 2020. The late Cenozoic history of the ostriches (Aves: Struthionidae), as revealed by fossil eggshell and bone remains. Earth-Science Reviews, 208:103270. https://doi.org/10.1016/j.earscirev.2020.103270
Mutihac, V. 1990. Structura geologică a teritoriului României. Editura Tehnică, București.
Palcu, D.V., Vasiliev, I., Stoica, M., and Krijgsman, W. 2018. The end of the great Khersonian drying of Eurasia: magnetostratigraphic dating of the Maeotian transgression in the Eastern Paratethys. Basin Research, 31:33-58. https://doi.org/10.1111/bre.12307
Pomel, A. 1853. Catalogue méthodique et descriptif des vertébrés fossiles découverts dans le bassin hydrographique supérieur de la Loire. J. B. Baillières, Paris.
Rafinesque, C.S. 1815. Analyse de la nature ou Tableau de l’univers et des corps organisés. Jean Barravecchia, Palermo.
Rage, J.-C. 2013. Mesozoic and Cenozoic squamates of Europe. Palaeobiodiversity and Palaeoenvironments, 93:517-534. https://doi.org/10.1007/s12549-013-0124-x
Rage, J.-C. and Hossini, S. 2000. Les amphibiens du Miocène moyen de Sansan. In Ginsburg, L. (ed.), La faune miocène de Sansan et son environnement, Mémoires du Muséum National d’Histoire Naturelle, 183:177-217.
Răileanu, V., Tătaru, D., Grecu, B., and Bala, A. 2011. Crustal models in Romania - II. Moldavian Platform and adjacent areas. Romanian Journal of Physics, 57(9–10):1438-1454.
Roček, Z. 1981. Cranial anatomy of frogs of the family Pelobatidae Stannius, 1856, with outlines of their phylogeny and systematics. Acta Universitatis Carolinae, Biologica, 1980:1-164.
Roček, Z. 2005. Late Miocene Amphibia from Rudabánya. Palaeontographia Italica, 90:11-29.
Roček, Z. 2019. A contribution to the herpetofauna from the late Miocene of Gritsev (Ukraine). Comptes Rendus Palevol, 18:817-847. https://doi.org/10.1016/j.crpv.2019.07.003
Rögl, F. 1999. Mediterranean and Paratethys. Facts and hypotheses of an Oligocene to Miocene paleogeography (short overview). Geologica Carpathica, 40:339-349.
Săndulescu, M. 1984. Geotectonica României. Editura Tehnică, București.
Sanchiz, B. and Młynarski, M. 1979. Remarks on the fossil anurans from the Polish Neogene. Acta Zoologica Cracoviensia, 24:175-188.
Scopoli, G.A. 1777. Introductio ad historiam naturalem, sistens genera lapidium, planatarum, et animalium hactenus detecta, caracteribus essentialibus donata, in tribus divisa, subinde ad leges naturae. Gerle, Prague.
Ștefănescu, M., Polonic, P., Alexandrescu, G., Popescu, I., Peltz, S., Ionescu, F., Niculin, M., Popescu-Brădet, L., and Constantinescu, P. 1985. Elaborarea profilelor, scara 1: 200 000 pentru completarea imaginii geologice a teritoriului R.S. România. A13: Secțiune geologică Tăul Zăului-Voiniceni-Ocna de Sus-Frumoasa-Dărmănești-Capăta-Bârlad. Institutul de Geologie și Geofizică, București.
Stoica, M., Lazăr, I., Krijgsman, W., Vasiliev, I., Jipa, D., and Floroiu, A. 2013. Paleoenvironmental evolution of the East Carpathian foredeep during the late Miocene-early Pliocene (Dacian Basin; Romania). Global and Planetary Change, 103:135-148. https://doi.org/10.1016/j.gloplacha.2012.04.004
Szyndlar, Z. 1991a. A review of Neogene and Quaternary snakes of Central and Eastern Europe. Part I. Scolecophidia, Boidae, Colubrinae. Estudios Geologicos, 47:103-126.
Szyndlar, Z. 1991b. A review of Neogene and Quaternary snakes of Central and Eastern Europe. Part II. Natricinae, Elapidae, Viperidae. Estudios Geologicos, 47:237-266.
Szyndlar, Z. 2005. Snake fauna from the Late Miocene of Rudabánya. Palaeontographia Italica, 90:31-52.
Szyndlar, Z. and Zerova, G.A. 1990. Neogene cobras of the genus Naja (Serpentes: Elapidae) of East Europe. Annalen des Naturhistorischen Museums in Wien, A91:53-61.
Syromyatnikova, E.V. 2015. A new species of Bufo (Amphibia, Anura) from the Miocene of Russia. Russian Journal of Herpetology, 22:281-288. https://doi.org/10.30906/1026-2296-2015-22-4-281-288
Syromyatnikova, E.V. 2019. Redescription of Pelobates praefuscus Khosatzky, 1985 and new records of Pelobates from the late Miocene-Pleistocene of Eastern Europe. Historical Biology, 31:888-897. https://doi.org/10.1080/08912963.2017.1402015
Trif, N., Bordeianu, M., and Codrea, V. 2021. A Maeotian (late Miocene) freshwater fish-fauna from Romania. Palaeoworld, 31:140-152. https://doi.org/10.1016/j.palwor.2021.01.004
Venczel, M. 2008. A new salamandrid amphibian from the Middle Miocene of Hungary and its phylogenetic relationships. Journal of Systematic Palaeontology, 6:41-49. https://doi.org/10.1017/S1477201907002283
Venczel, M., Codrea, V., and Fărcaș, C. 2013. A new palaeobatrachid frog from the early Oligocene of Suceag, Romania. Journal of Systematic Palaeontology, 11:179-189. https://doi.org/10.1080/14772019.2012.671790
Venczel, M. and Hír, J. 2015. Lissamphibians and squamate reptiles from the early middle Miocene of Litke, Northern Hungary. Geobios, 48:491-504. https://doi.org/10.1016/j.geobios.2015.09.001
Wagler, J. 1830. Natürliches System der Amphibien, mit vorangehender Classification der Säugthiere und Vogel. Ein Beitrag zur vergleichenden Zoologie. J.G. Cotta, München, Stuttgart and Tübingen.
