

 A new Turolian species Lophocricetus cimishliensis (Rodentia, Zapodidae) from the Republic of Moldova
A new Turolian species Lophocricetus cimishliensis (Rodentia, Zapodidae) from the Republic of Moldova
Article number: 17.3.35A
https://doi.org/10.26879/423
Copyright Society for Vertebrate Paleontology, October 2014
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 1 August 2013. Acceptance: 19 September 2014
{flike id=923}
ABSTRACT
A new species of Lophocricetus, L. cimishliensis, from three Late Miocene localities from the Republic of Moldova is described. All three Late Miocene localities (Chimishliya, Gura Galbene and Gradishte,) are located along the Kogylnik River, between the Dniester and Prut Rivers, Chimishliya District, southern Moldova. They yield diverse mammalian fossils, including several lagomorphs, rodents and insectivores: Parasorex socialis von Meyer, 1865; Alilepus laskarewi Khomenko, 1914; Euroxenomys minutum (Meyer, 1838); Myomimus dehmi (De Bruijn, 1966) / maritsensis De Bruijn et al. 1970 - group of species; Vasseuromys cf. pannonicus (Kretzoi, 1978); Kowalskia progressa Topachevski and Scorik, 1992; Apodemus aff. barbarae (Van de Weerd, 1976); etc.), which are characteristic in the Turolian age. Lophocricetus cimishliensis differs from the species of this genus by smaller sizes, better developed mesocone/mesoconid and style/stylids. Comparison of the new species with other species of Lophocricetus from Eastern Europe and Asia allows reconstructing their phylogenetic interrelationships during the Late Miocene of Europe. Our data suggest two possible scenarios: this species could descend from an East European ancestor or from an Asian population.
Andrian Delinschi, National Museum of Ethnography and Natural History of Moldova, 82 Kogălniceanu Str., 2009, Chișinău, Republic of Moldova. andriandelinschi@yahoo.com
Key words: Rodentia; Zapodidae; Lophocricetus; systematics; Turolian; Republic of Moldova
Final citation: Delinschi, Andrian. 2014. A new Turolian species Lophocricetus cimishliensis (Rodentia, Zapodidae) from the Republic of Moldova. Palaeontologia Electronica Vol. 17, Issue 3;35A; 13p. https://doi.org/10.26879/423
palaeo-electronica.org/content/2014/923-new-dipodid-from-moldova
http://zoobank.org/5F766775-E6C6-47CB-A718-A43C6EAEEBF1
INTRODUCTION
Lophocricetus Schlosser, 1924, is a common genus in the Asian and Eastern Europe Late Miocene fossil record. It was originally described on fossil material from the Late Miocene Chinese localities Ertemte 1 and Harr Obo, as zapodid rodents of large to medium size, marked by concave occlusal teeth surfaces (Schlosser, 1924). Schlosser (1924) included this taxon in the subfamily Microtinae . Later Schaub (1934) made a revision, including Lophocricetus in Cricetidae, and this was supported by Stehlin and Schaub (1951), Gromov (1962) and Vorontzov (1967). Later, Savinov (1970) described a new subfamily, Lophocricetinae with the single genus Lophocricetus, related to Dipodidae. Topachevsky et al. (1984) for the first time described Lophocritetus in Eastern Europe with three new species: L. sarmaticum, L. maeoticus and L. complicidens . The first two species were described based on a single tooth. Therefore, later Zazhigin et al. (2002) synonymized L. sarmaticum with L. complicidens and considered L. maeoticus as a junior synonym of L. minuscilus .
Zazhigin et al. (2002) included Lophocricetus into the family Dipodidae and divided it in two subgenera: Lophocricetus s.str. including L. minuscilus Savinov, 1977 (MN10-11 Kazakhstan and Ukraine), L. complicidens Topachevsky et al. 1984 (MN10, Ukraine), L. vinogradovi Savinov, 1970 (MN12-13, Kazakhstan), L. grabaui Schlosser, 1924 (MN13, China), L. reliquus (MN13, Mongolia); Paralophocricetus, including P. pussilus , Schaub, 1934 (MN13-14, China and Mongolia), P. intermedius (MN13, Mongolia), P. progressus (MN14, Eastern Siberia); P. afanasievi Savinov, 1970 (MN13, Kazakhstan), P. sibiricus (MN14, Western Siberia), P. ultimus (MN14-15, Western Siberia).
Qiu et al. (2008), accepted the systematics proposed by Zazhigin et al. (2002), but they included Lophocricetus into Zapodidae and completed the list of Lophocricetus s.s. with the species “gansus” and “saraicus”, initially described by Zheng (1982) and Zazhigin et al. (2002) as Heterosminthus . Unfortunately Qiu did not justify the usage of Zapodidae, for which reason I continue to attribute Lophocricetus to Dipodidae.
In Republic of Moldova, Lungu (1981) reported the first zapodids from Buzhor (MN9), including two new genera Bujoromys (wrongly related to Muridae) and Sarmatosminthus . Later, Topachevsky et al. (1984) validated genera Sarmatosminthus, which has been attributed to Lophocricetinae . Revising the Eastern European and Asian dipodids, Zazhigin and Lopatin (2000) concluded that both Bujoromys and Sarmatosminthus are synonyms of Heterosminthus . In a recent work Lungu and Rzebik-Kowalska (2011), do not support the taxonomy proposed by Zazhigin and Lopatin (2000) and continue to assign the name Sarmatosminthus for the zapodids from Buzhor and Keinar (MN10). The fossil collections at Tiraspol State University (Kisinev) allow redescription of the material from Keinar (M1, m2) as Lophocricetus sp., based on presence of following characters: strong development of ridges, the protocone is connected to the paracone and the hypocone joins the metacone on M1.
MATERIAL AND METHODS
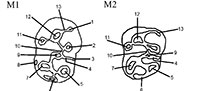
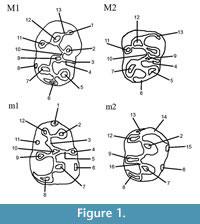 A main part of the fossil material, described here as Lophocricetus, was collected during fieldwork in 2000-2012 from the Late Miocene deposits of the Republic of Moldova. The fossil materials were extracted from sediments by dry screening. They are stored in the paleontological collections of the National Museum of Ethnography and Natural History of Moldova (abbreviated: NMENHM). The teeth measurements are given in millimeters. The measurements were done by digital microscope LEICA DVM 5000 in the Institute for Geoscience, Eberhard Karls University Tübingen, Germany. The images were performed on scanning electron microscope in same institute. The dental nomenclature and measurements follow Qiu (1985) and Zazhigin and Lopatin (2000) (Figure 1).
A main part of the fossil material, described here as Lophocricetus, was collected during fieldwork in 2000-2012 from the Late Miocene deposits of the Republic of Moldova. The fossil materials were extracted from sediments by dry screening. They are stored in the paleontological collections of the National Museum of Ethnography and Natural History of Moldova (abbreviated: NMENHM). The teeth measurements are given in millimeters. The measurements were done by digital microscope LEICA DVM 5000 in the Institute for Geoscience, Eberhard Karls University Tübingen, Germany. The images were performed on scanning electron microscope in same institute. The dental nomenclature and measurements follow Qiu (1985) and Zazhigin and Lopatin (2000) (Figure 1).
GEOLOGICAL SETTINGS
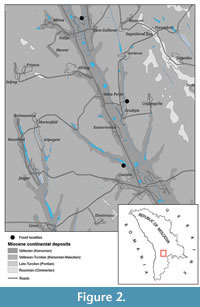 The material was collected from the Balta Formation (Barbot de Marny, 1869) (named Balta-Paun in Ionesi et al., 2005), Chimishliya District, Republic of Moldova (Figure 2).
The material was collected from the Balta Formation (Barbot de Marny, 1869) (named Balta-Paun in Ionesi et al., 2005), Chimishliya District, Republic of Moldova (Figure 2).
The sediments of the Balta Formation are common in the central and southern areas on Prut-Dniester interfluve. According to Khubka (1967, 1968), the Balta Formation is built by Late Miocene continental deposits. Lithologic it is represented by clay and sand, with cyclic structure. Each cycle starts with middle grain sand, with coarse sand lenses and rudite where most fossil material was found. The middle part is composed of medium to fine sand, with oblique and cross lamination. The top of the cycle is composed of silt and scaly clay. The thickness of each cycle ranges from 4 to 20 m. Overall, 11 to 12 cycles are recorded from the Prut-Dniester interfluve, while only 3 to 4 rhythms are present in the Kogylnik Valley. Genetically, each cycle begins with alluvial riverbed deposits followed by a transition from the riverbed to meadow deposits, and finally meadow and lacustrine deposits. The cross laminations are typical for these deposits. Each cycle is situated above the eroded surface of the previous cycle. Cyclic structure of these deposits, expressed by vertical changes of granulometric content, is related to the periodic change of the level of the erosion base of the fluvial network, leading to increased fluvial erosion (gravel and coarse sand lenses ), or alternatively, to the decreasing erosion (clays).
SYSTEMATIC PALEONTOLOGY
Order RODENTIA Bowdich, 1821
Family DIPODIDAE Fischer von Waldheim, 1817
Subfamily LOPHOCRICETINAE, Savinov, 1970
Genus LOPHOCRICETUS Schlosser, 1924
Type species Lophocricetus grabaui Schlosser, 1924
Diagnosis of the genus. The upper molars of Lophocricetus are generally characterized by the presence of the main (protocone, paracone, mesocone metacone, hypocone) and the secondary (anterostyle, protostyle, mesostyle, posterostyle) cusps, as well as lophs like anteroloph, endoloph, mesoloph and posteroloph. The lower molars of Lophocricetus are characterized by the presence of the main (anteroconid, protoconid, metaconid, hypoconid, entoconid), secondary cusps (anterostylid, ectostylid, posterostylid) and lophids (anterolophid, ectolophid, posterolophid).
The protostyle on M1 is generally strong and connected to protocone in all Lophocricetus species.
The anteroconid on m1 is generally weak or absent and is isolated or connected to metaconid in all Lophocricetus species. A strong mesoconid is triangular in shape and is fused with the long ectomesolophid in majority of Lophocricetus species. More detailed in Schlosser (1924, p. 41), Savinov (1970, p. 102), Qiu (1985, p. 42) and Zazhigin et al. (2002, p. 64).
Lophocricetus cimishliensis sp. nov.
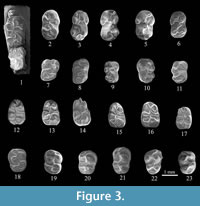
Figure 3.1-23
2009 - Lophocricetus minuscilus Delinschi, p. 393
http://zoobank.org/03B8FA7A-3C40-4568-8021-13756B8807BB
 Type locality. Gura Galbene, Chimishliya District, Republic of Moldova.
Type locality. Gura Galbene, Chimishliya District, Republic of Moldova.
Holotype. MNEIN Loph/GG-1, jaw fragment with P4-M2. National Museum of Etnography and Natural History of Moldova.
Type horizon. Middle grained sand, with coarse sand lenses and rudite, upper part of the Balta Formation
Geological age. Middle Turolian, Late Miocene, MN12.
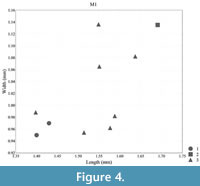
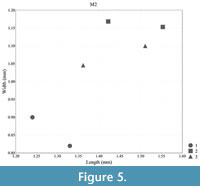
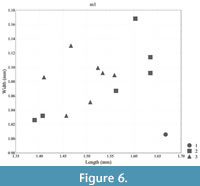
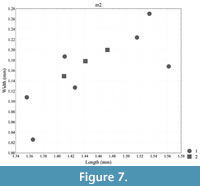
Measurements: see Table 1, Figure 4, Figure 5, Figure 6, and Figure 7
Diagnosis: small sized species of Lophocricetus with well-developed protostyle on M1 and M2 and strong stylids on m1-m2. The protocone on M1 are well developed, and in most teeth, is connected to small anterostyle through weak anteroloph, the mesoloph in most cases is short, the endoloph connected to paracone, the ridge-like posteroloph, usually is connected to the metacone; M2 with single anteroloph, long mesoloph, long posteroloph in some cases is connected to metacone; m1 in most cases with isolated weak anteroconid, usually strong ectomesolophid, ectostylid and posterostilyd, the hypoconid connected to entoconid or entoconid-mesoconid ridge conection; m2 protoconid connected with metaconid or anterolophid, anterostylid and protostylid are presented or not.
 Differential diagnosis. Lophocricetus cimishliensis sp. nov. differs from:
Differential diagnosis. Lophocricetus cimishliensis sp. nov. differs from:
Lophocricetus minuscilus by 1. better developed of lingual posteroloph on M1; 2. anteroconid on m1 is small to medium sized in most cases isolated; 3. better developed mesosconid on m1; 4. strong developed posterolophid on m1; 5. better developed stylids on m1-m2.
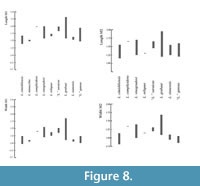
Lophocricetus complicidens by 1. smaller size (Figure 8, Figure 9); 2. better development of mesoloph on M1; 3. lack of connection between the paracone and mesocone on M2; 4. in some cases the hypoconid is connected directly to mesoconid; 5. on m2 posterolophid is connected to the labial part of hypoconid (on L. complicidens this connection is in lingual part of hypoconide, or posterolophid is isolated).
 Lophocricetus vinogradovi by 1. smaller size (Figure 8, Figure 9); 2. less developed anteroloph; 3. better developed mesoloph; 4. more lingual posteroloph ( L. vinogradovi usually has double branch); 5. presence of mesocone on M2; 6. usually isolated anteroconid on m1 (in L. vinogradovi, anteroconid is usually connected to metaconid).
Lophocricetus vinogradovi by 1. smaller size (Figure 8, Figure 9); 2. less developed anteroloph; 3. better developed mesoloph; 4. more lingual posteroloph ( L. vinogradovi usually has double branch); 5. presence of mesocone on M2; 6. usually isolated anteroconid on m1 (in L. vinogradovi, anteroconid is usually connected to metaconid).
Lophocricetus reliquus by 1. smaller size (Figure 8, Figure 9); 2. better developed mesoloph; 3. less developed of anterostyle; 4. shorter lingual branch of posteroloph on M1.
Lophocricetus grabaui by 1. smaller size (Figure 8, Figure 9); 2. more elongate M1 and m1; 3. weaker development of protostyle; 4. strongly shifted backward protostyle towards protocone; 4. presence of the mesoloph on M1.
Lophocricetus xianensis by 1. weaker developed protostyle; 2. endoloph joining protocone; 3. posteroloph connected to metacone on M1; 4. larger posterior endosinus on M2 (on L. xianensis posterior endosinus is smaller); 5. presence of styles and stylids.
 “Lophocricetus”gansus: smaller size (Figure 8, Figure 9), weaker developed anterocone, lack of connection between hypocone and posteroloph.
“Lophocricetus”gansus: smaller size (Figure 8, Figure 9), weaker developed anterocone, lack of connection between hypocone and posteroloph.
“Lophocricetus”saraicus: smaller size (Figure 8, Figure 9), always lack of connection between hypocone and posteroloph on M1.
Description
M1 is elongated with relatively prominent cusps and weak crests. The protostyle is well developed, strongly shifted backward and always connected to protocone through a weak crest. The anterostyle in most teeth is connected to protocone (at two teeth from Gradishte, the anterocone is isolated and anteroloph lack), through weak anteroloph. The protocone is joined to large paracone. In nine out of twelve teeth, the anterior endoloph connects mesocone to paracone and to protocone-paracone yoke in three ( sensu Qiu et al., 2008). The mesocone is relatively well developed, most mesolophs are short. In one tooth (Figure 3.5) mesoloph is long and ends with a small mesostyle at the labial corner of the tooth. The posterostyle ( sensu Qiu, 1985) or hypostyle ( sensu Qiu et al., 2008) is missing. The ridge-like posteroloph, in ten of twelve determinable specimens, is connected to the metacone and to the hypocone-metacone yoke, in two. In one strong worn tooth from Chimishliya posterior endosinus is closed in fosette (Figure 3.2).
 M2 is longer than wide, and slightly wider anteriorly than posteriorly. In five teeth the protostyle is well developed and connected to protocone, in one tooth (Chimishliya) the protostyle is very weak, like a small projecting spur attached to the protocone. The anteroloph is single in all cases. The weak anterior endoloph is present in three cases, connected to the paracone. In four of six determinable specimens the small mesocone is present. The long mesoloph reaches to the labial edge of the tooth and ends with a small mesostyl in three cases. The narrow and long posteroloph is connected to hypocone, and in three from six determinable specimens also to metacone.
M2 is longer than wide, and slightly wider anteriorly than posteriorly. In five teeth the protostyle is well developed and connected to protocone, in one tooth (Chimishliya) the protostyle is very weak, like a small projecting spur attached to the protocone. The anteroloph is single in all cases. The weak anterior endoloph is present in three cases, connected to the paracone. In four of six determinable specimens the small mesocone is present. The long mesoloph reaches to the labial edge of the tooth and ends with a small mesostyl in three cases. The narrow and long posteroloph is connected to hypocone, and in three from six determinable specimens also to metacone.
The m1 is relatively elongated. The anteroconid varies in its development: it is well developed as a separated cusp (nine teeth), poorly developed but also separated (three teeth), or poorly developed, joining the metaconid (two teeth). The protoconid is isolated in a single tooth (Figure 3.15), otherwise joining to the strong ectolophid that connects the protoconid, metaconid and mesoconid. In a single tooth (Figure 3.13), a rudimentary ridge between the protoconid and mesoconid can be observed. A small cusp (mesostilyd?) is present in one tooth from Gura Galbene (Figure 3.12). The well-developed mesoconid is triangular in shape, fused with a long ectomesolophid, which ends with an ectostylid. The hypoconid, is connected to entoconid (Figure 3.12), mesoconid (Figure 3.13) or is isolated (Figure 3.15). Strong developed posterolophid varies in its configurations: separated in one tooth, joins only to hypoconid in seven teeth and joint to hypoconid and posterior part of entoconid in another six. The well-developed posterostylid connects to hypoconid in seven teeth, but is separated in other six and is missing in one tooth.
 The m2 shows distinct variation in outline. The labial arm of the anterolophid is long and turns posterolabially. Connection between the labial arm of anterolophid and protoconid can be seen in five teeth, and is lacking in another fives. In two teeth (Figure 3.18-19), a fine and low lingual arm of anterolophid is observed. The protoconid is connected to metaconid (six teeth) or to metaconid-anteroconid (four teeth) by a crest, variable in outline. The hypoconid is always connected to the entoconid by the posterior ectolophid. The posterolophid joins the hypoconid in five teeth and is connected to entoconid with closed posterior ectosinusid in another five. The anterostylid can be seen in a form of a small separated cusp in five cases. The posterostylid can be seen in three cases.
The m2 shows distinct variation in outline. The labial arm of the anterolophid is long and turns posterolabially. Connection between the labial arm of anterolophid and protoconid can be seen in five teeth, and is lacking in another fives. In two teeth (Figure 3.18-19), a fine and low lingual arm of anterolophid is observed. The protoconid is connected to metaconid (six teeth) or to metaconid-anteroconid (four teeth) by a crest, variable in outline. The hypoconid is always connected to the entoconid by the posterior ectolophid. The posterolophid joins the hypoconid in five teeth and is connected to entoconid with closed posterior ectosinusid in another five. The anterostylid can be seen in a form of a small separated cusp in five cases. The posterostylid can be seen in three cases.
DISCUSSION
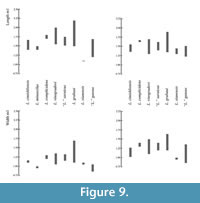 Biometry. In terms of molar length and width, Lophocricetus cimishliensis is generally smaller than most known representatives to species of Lophocricetus (Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, and Figure 9). Lophocricetus cimishliensis is close to L. minuscilus from Cherevichnoe (Ukraine) and Petropavlovsk 1A (Kazakhstan) (Topachevsky et al., 1984; Zazhigin et al., 2002) in morphology. However, the number of specimens of L. minuscilus available for study is very small, with the four M1 - m1 from Petropavlovsk 1A, and a single m1 from Cherevichnoe. Therefore, it is difficult to conclude about the relationships of these two species. Lophocricetus cimishliensis is slightly smaller than L. xianensis (Figure 8, Figure 9) from Loc.19, Lantian County, China (Qiu, 2008).
Biometry. In terms of molar length and width, Lophocricetus cimishliensis is generally smaller than most known representatives to species of Lophocricetus (Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, and Figure 9). Lophocricetus cimishliensis is close to L. minuscilus from Cherevichnoe (Ukraine) and Petropavlovsk 1A (Kazakhstan) (Topachevsky et al., 1984; Zazhigin et al., 2002) in morphology. However, the number of specimens of L. minuscilus available for study is very small, with the four M1 - m1 from Petropavlovsk 1A, and a single m1 from Cherevichnoe. Therefore, it is difficult to conclude about the relationships of these two species. Lophocricetus cimishliensis is slightly smaller than L. xianensis (Figure 8, Figure 9) from Loc.19, Lantian County, China (Qiu, 2008).
The sample of Lophocricetus reliquus from Hirghis-Nur, Mongolia (Zazhigin et al., 2002) is very scanty, making any comparison difficult. Nevertheles, M1 of L. reliquus is slightly larger than that of M1 of L . cimishliensis, moreover, size of M2 of L. reliquus is within the limits of M2 of L.cimishliensis .
The length values of Lophocricetus complicidens from its type locality Novoelizavetovka, Ukraine (Topachevski et al., 1984) as well as the width measurements of M1 - 2 and m1-2 clearly show larger sizes of L. complicidens (Figure 8, Figure 9).
Size differences of Lophocricetus grabaui from Ertemte and Harr Obo, China (Qiu, 1985), “L” . gansus from Builstyn Khudag, Mongolia (Daxner-Hock, 2001), L. vinogradovi from Gusinyi Perelet, Kazakhstan and “L” . saraicus (Figure 8, Figure 9) from the Olkohon Island, Irkutsk, Russia (Zazhigin et al., 2002) is present in Figure 8, Figure 9.
Biostratigraphic Assignations of Species of Lophocricetus
Lophocricetus is a typical representative to genera of the Upper Miocene faunas from Asia and Eastern Europe. The oldest records of Lophocricetus are considered L. xianensis from Loc. 19, Lantian County, China (MN10, Qiu, 2008), and L. minuscilus from Petropavlovsk 1A, Kazakhstan, (MN10, Savinov, 1977; Zazhigin et al., 2002).
Description of L. minuscilus from Cherevichnoe (Ukraine, MN12) is based on only a single m1, therefore it is difficult to make an advanced comparison regarding its relationships with other taxa. Zazhigin et al. (2002) considered this species typical for the Late Vallesian of Kazakhstan, but according to Nesin and Nadachovski (2001) and Nesin and Storch (2004) Cherevichnoe is considered as a Middle Turolian locality. Thereby, it is necessary to revise of Cherevichnoe specimen, or extend the stratigraphic distribution of L. minuscilus till Middle Turolian. Most of the other members of the genus have been recorded from Turolian deposits, including L. complicidens (MN11, Topachevsky et al. 1984), “L.” gansus (MN11, Daxner-Hock, 2001), L. vinogradovi (MN12, Savinov 1970), “L.” saraicus (MN12, Zazhigin et al. 2002), L. reliquus (MN12, Zazhigin et al., 2002), L. cimishliensis sp. nov. (MN12). The latest, geologically youngest species of Lophocricetus is considered L. grabaui from Ertemte and Harr Obo, Inner Mongolia China (MN13-14, Qiu, 1985).
Phylogenetic Relationships of Lophocricetus cimishliensis with Other Species of Lophocricetus
Based on the presence or absence of posterostyle on M1-2 the genus Lophocricetus can be divided into Lophocricetus sensu stricto and Paralophocricetus (Zazhigin et al., 2002). The specimen from the Chimishliya District has an indistinct posterostyle, which allows it to be attributed to Lophocricetus sensu stricto.
The developed protostyle located more posteriorly to the center of the protocone on M1 of Lophocricetus cimishliensis (a diagnostic feature of Lophocricetus (Schlosser, 1924)) brings the species close to L . minuscilus from Petropavlovsk 1A (Kazakhstan) and L. complicidens from Cherevichnoe, beside this L. cimishliensis can be excluded from L. vinogradovi, L. grabaui and L. xianensis .
Anteroloph on M2 (another important feature of Lophocricetus ), on L. cimishliensis, is single in all teeth, this is common for L. complicidens, “L” . saraicus, L. grabaui, “L.” gansus, and differs in L. vinogradovi and L xianensis . M2 of L.minuscilus a nd L. reliquus is unknown.
Lophocricetus cimishliensis has relatively strong and short mesolophs (also an important taxonomic character among Lophocricetus species.) on M1 and M2. This character also makes the new species similar to L. minuscilus and L. complicidens, excluding any similarity with L. grabaui, L. reliquus, L. vinogradovi, and L. xianensis .
A connection between posterolophid and the main cusps of m1 on Lophocricetus, shows distinct variation. Thus, posterolophid can connect just hypoconid like in L. grabaui, L. complicidens, L. minuscilus, L. vinogradovi, “L” . saraicus, or connected with hypoconid and posterior part of entoconid or isolated like in L. cimishliensis, “L” . gansus, and L. xianensis .
The development of stylids on m1-2 (a diagnostic feature of Lophocricetus ) can occur either in form of separated cusps or can be joined to the main cusps as in L. cimishliensis, L. complicidens, L. vinogradovi, “L” . gansus (few cases), L. grabaui, or can be indistinct in L. xianensis, L. minuscilus ?.
The presence of Lophocricetus cimishliensis in the Turolian fauna of the Republic of Moldova, indicate two possible scenarios regarding their ancestry: 1. Lophocricetus is considered to have descended from Heterosminthus (Zazhigin et al., 2002; Qiu, 2008), so L. cimishliensis may have descended from an East European ancestor and can form a phylogenetic line like: Heterosminthus gabuniai Lungu, 1981(= Sarmatosminthus gabuniai in Lungu and Rzebik-Kowalska, 2011), (Buzhor, MN9) - Lophocricetus sp. (= Heterosminthus sp. in Lungu and Rzebik-Kowalska, 2011), (Keinar, MN10) - Lopocricetus cimishliensis (Chimishliya, Gura Galbene, Gradishte, MN12); 2. It may have descended from an Asian population, which later migrated into Eastern Europe: L. minuscilus (Kazakhstan, MN10) - L. minuscilus (Ukraine, MN12) - L. cimishliensis (Republic of Moldova, MN12).
The tectonic events and geographical changes during the Neogene of Europe likely provided geographical barriers to dispersal, as well as concurrent land bridges allowing faunal interchange. As an example, similarity between Lophocricetus cimishliensis and L. minuscilus from the Vallesian of Kazakhstan can be considered (Savinov, 1977, Zazhigin et al., 2002).
CONCLUSIONS
The assemblage with Lophoricetus from the Chimishliya District (Chimishliya, Gura Galbene, Gradishte) is the first record of this genus from the Turolian deposits of the Republic of Moldova. Lophocricetus cimishliensis from the Chimishliya District is smaller than the majority of the Lophocricetus species, shearing comparable sizes with L. minuscilus . The molars of this new species distinctly differ from those of all other Lophocricetus species, in the constant presence of a well-developed mesocone and short mesoloph on M1, and strong stylids on m1-2.
The study of Lophocricetus cimishliensis promises interesting results on evolution and distribution of Lophocricetus, as well as on biostratigraphy and faunal evolution in Eastern Europe. This genus is of peculiar interest, since it did not penetrate into Western Europe, due to orographic, climatic, biological or other barriers.
Among the other Upper Miocene representatives of zapodids in the Republic of Moldova Heterosmintus gabuniai occurs abundantly (= Sarmatosminthus gabuniai in Lungu and Rzebik-Kowalska, 2011) in Buzhor (MN9), whereas Lophocricetus sp. (= Heterosminthus sp. in Lungu and Rzebik-Kowalska, 2011) is present with two teeth in Keinar (MN10).
ACKNOWLEDGEMENTS
I would like to thank M. Sinitsa (Kiev) and V.S. Zazhigin (Moscow) for their criticism and continuous encouragement. Also, many thanks to colleagues from the Paleontological collection, University of Tubingen, Germany: M. Böhme, J. Prieto and D. Vasilyan for their help, as well as valuable comments and critical remarks. Thanks are also due to V. Codrea (Cluj-Napoca) and A. Ciornei for the improvement of English grammar and I. Rosca for cartographic assistance. Many thanks are also given to N. Volontir, Head of Department of General Geography, Tiraspol State University Moldova, for making possible work on the collection of Prof. Lungu.
REFERENCES
Barbot de Marny, N. 1869. Geological Essay of the Khersonian District. Demakova, B., St. Petersburg.
Daxner-Höck, G. 2001. New zapodids (Rodentia) from Oligocene-Miocene deposits in Mongolia. Part I. Senckenbergiana lethaea, 81:359-389.
Delinschi, A. 2009. Contribution to the study of maeotian Hipparion faunas from the Republic of Moldova. Oltenia Journal for Studies in Natural Sciences, XXV/2009:391-395.
Gromov, I. 1962. Order Rodentia, 472-507 In Base of Paleontology. Mammalia. Nauka, Moskow. (In Russian)
Ionesi, L., Ionesi, B., Rosca, V., Lungu, A., and Ionesi, V. 2005. Middle and Upper Sarmatian from the Moldavian Plate . Romanian Academy Press. (In Romanian)
Khubka, A. 1967. Lithology, facies and stratigraphy of continental deposits of Upper Miocene of Moldavian SSR and adjacent areas of Ukrainian SSR . PhD Thesis summary. Odessa State University, Ukraine. (In Russian)
Khubka, A. 1968. On the condition of formation Balt deposits on the territory of Moldavia. Reports of the Academy of Sciences of the URSS, 180:25-36. (In Russian)
Lungu, A. 1981. Hipparion fauna of the Middle Sarmatian from Moldavia (Insectivora, Lagomorpha, Rodentia ). Izdatelistvo ”Shtiintsa”. Kisinev. (In Russian)
Lungu, A. and Rzebik-Kowalska, B. 2011. Faunal assemblages, stratigraphy and taphonomy of the Late Miocene localities in the Republic of Moldova . Polish Academy of Sciences, Kraków.
Nesin, V.A. and Nadachowski, A. 2001. Late Miocene and Pliocene small mammal faunas (Insectivora, Lagomorpha, Rodentia) of southeastern Europe. Acta Zoologica Cracoviensia, 44:107-135.
Nesin, V.A. and Storch, G. 2004. Neogene Murinae of Ukraine (Mammalia, Rodentia). Senckenbergiana lethaea, 84:351-365.
Qiu, Z.D. 1985. The Neogen mammalian faunas of Erteme and Harr Obo in Inner Mongolia (Nei Mongol), China. - 3. Jumping mice - Rodentia: Lophocricetinae. Senckenbergiana lethaea, 66:39-67.
Qiu, Z.D., Zheng, S.H., and Zhang, Z.Q. 2008. Sciurids and Zapodids from the Late Miocene Bahe Formation, Lantian, Shaanxi. Vertebrata PalAsiatica, 46:111-123.
Savinov, P. 1970. Jerboas (Dipodidae, Rodentia) from Neogene of Kazakhstan. Materials on the evolution of terrestrial vertebrates . Nauka:91-134. (In Russian)
Savinov, P. 1977. New representat of Lophocricetinae from North Kazakhstan. Meso-Cenozoic flora and fauna from North-Western of Kazakhstan . Alma-Ata, 7:27-32. (In Russian)
Schaub, S. 1934. Über einige fossile Simplicidentaten aus China und der Mongolei. Abhandlungen der schweizerischen palaeontologischen Gesellschaft, 54:1-40.
Schlosser, M. 1924. Tertiary vertebrates from Mongolia. Paleontologica sinica, Series C, 1:1-119.
Stehlin, H.G. and Schaub, S. 1951. Die Trigonodontie der simplicidentaten Nager. Schweizerische Palaeontologische Abhandlungen, 67:1-385.
Topachevsky, V.A., Scorik, A.F., and Rekovetz, L.I. 1984. The Oldest Lophocricetinae (Rodentia, Dipodidae) of South-Western European Part of the USSR. Vestnik Zoologii 4:32-39. (In Russian)
Vorontzov, N. 1967. Evolution of the digestive system of rodents (Muridae) . Novosibirsk. (In Russian)
Zazhigin, V.S. and Lopatin A.V. 2000. The history of the Dipodoidea (Rodentia, Mammalia) in the Miocene of Asia: 1. Heterosminthus (Lophocricetinae). Paleontological Journal, 3:90-102. (In Russian)
Zazhigin, V.S., Lopatin, A.V., and Pokatilov, A.G. 2002. The history of the Dipodoidea (Rodentia, Mammalia) in the Miocene of Asia: 5. Lophocricetus (Lophocricetinae). Paleontological Journal, 2:62-75. (In Russian)
Zheng, S.H. 1982. Middle Pliocene micromammals from the Tianzhu Loc. 80007 (Gansu Province). Vertebrata PalAsiatica, 20:138-147. (In Chinese with English abstract)
Holotype. MNEIN Loph/GG-1, jaw fragment with P4-M2. National Museum of Etnography and Natural History of Moldova.
