

 Early Pleistocene freshwater fishes of Copăceni (Dacian Basin, southern Romania)
Early Pleistocene freshwater fishes of Copăceni (Dacian Basin, southern Romania)
Article number: 23(1):a02
https://doi.org/10.26879/1014
Copyright Paleontological Society, January 2020
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 30 July 2019. Acceptance: 12 January 2020.
ABSTRACT
This paper describes the first early Pleistocene freshwater fish assemblage from the Dacian Basin, Romania. The fossil sample, consisting mainly of isolated teeth, supports the presence of a rich and taxonomically diverse fish assemblage: 17 taxa were identified, belonging to 10 genera of the families Cyprinidae, Salmonidae, Siluridae, Esocidae, and Percidae. The ecological affinities of the identified taxa suggest the early Pleistocene fishes from Copăceni lived in a large lowland mesophitic river. The faunal composition is comparable to that of other southeast European assemblages, however, it includes more thermophilic taxa, suggesting the Dacian Basin represented an early Pleistocene refugium in a cooling post-Pliocene environment. The taxonomical composition of the fossil assemblage suggests a pre-Pleistocene dispersion of freshwater taxa took place in the areas surrounding the Euxinian Basin.
Ştefan Vasile. University of Bucharest, Faculty of Geology and Geophysics, Research Centre for the Geology of Coal Deposits and Environment Protection, 1 Nicolae Bălcescu Ave., Bucharest 010041, Romania, yokozuna_uz@yahoo.com
Oleksandr Kovalchuk. National Museum of Natural History of the National Academy of Sciences of Ukraine, Department of Paleontology, 15 Bohdan Khmelnytsky St., Kyiv 01030 Ukraine, biologiest@ukr.net
Alexandru Petculescu. Romanian Academy, “Emil Racoviță” Institute of Speleology, 13-15 Calea 13 Septembrie, Bucharest 050711, Romania, alexpet@gmail.com
and Romanian Institute of Science and Technology, 3 Virgil Fulicea st., Cluj-Napoca 400022, Romania.
Márton Venczel. Țării Crișurilor Museum, 1/A Calea Armatei Române, Oradea 410087, Romania, mvenczel@gmail.com
Keywords: fish assemblage; diversity; environment; distribution; Pleistocene; Europe
Final citation: Vasile, Ştefan, Kovalchuk, Oleksandr, Petculescu, Alexandru, and Venczel, Márton. 2020. Early Pleistocene freshwater fishes of Copăceni (Dacian Basin, southern Romania). Palaeontologia Electronica, 23(1):a02. https://doi.org/10.26879/1014
palaeo-electronica.org/content/2020/2901-pleistocene-fishes-of-romania
Copyright: January 2020 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
The Dacian Basin is represented by a sedimentary unit resulted from the fragmenting of the Paratethys during the Middle Miocene. It is bordered by the Carpathians to the north and west, by the Balkans to the south, and by the folded units of Northern Dobrogea to the east (e.g., Jipa and Olariu, 2009; Andreescu et al., 2013). Although some connections to the neighbouring Pannonian and Euxinian basins still existed, since its separation as an individual basin, the water salinity in the Dacian Basin gradually changed from normal marine to brackish, and it became freshwater during the late Pliocene (Jipa and Olariu, 2009; Andreescu et al., 2013). The late Pliocene and early Pleistocene sedimentary units (Andreescu et al., 2013) consist of alluvial and fluvial rudites and arenites, except for a silty unit, the “Copăceni beds”, that will be discussed in detail below.
The Plio-Pleistocene freshwater deposits of the Dacian Basin yielded mammalian fossil remains that were reported from many localities (Athanasiu, 1915; Apostol, 1965, 1968, 1972, 1976, 1981; Apostol and Stancu, 1968; Apostol and Vicoveanu, 1970; Feru et al., 1978, 1979, 1983; Terzea and Boroneanț, 1979; Apostol and Cacoveanu, 1980; Terzea, 1981, 2005; Rădulescu and Samson, 1986, 1990, 2001; Rădulescu et al., 1989, 1993, 1999, 2003; Lister and van Essen, 2003; Markov and Spassov, 2003; Popescu, 2004; Croitor and Popescu, 2011; Vasile et al., 2012, 2013a). Non-mammalian vertebrate fossils described from the Dacian Basin are exceedingly rare. The only two assemblages with reptile and amphibian remains are Tetoiu (Voicu et al., 2015) and Copăceni (Vasile et al., 2013b, 2015, 2016, 2018), both of early Pleistocene age. However, the taxa reported from these sites are still to be described in detail. The Plio-Pleistocene fish assemblages of the Dacian Basin are poorly known, as are other freshwater fossil fish assemblages of Romania, irrespective of age (for a review, see Trif and Codrea, 2018).
The early Pliocene fishes (local Dacian age) of the Dacian Basin are known by otoliths (rare gobiid teeth are also present) belonging to Gobiidae and Sciaenidae. These families include brackish and freshwater species, in agreement with the palaeoenvironment supported by the brackish invertebrates known from successions in western Dobrogea (at Oltina, in south-eastern part of the basin) and Oltenia (several sites in the west of the basin including Corabia, Scoarța, and Ișalnița) (Pană, 1977, 1982, 1995; see also Bannikov et al., 2018 for a revision of otoliths).
A few isolated pharyngeal teeth, assigned to cyprinid fishes, such as Tinca and Leuciscus (Pană, 1982), document a transition from brackish to freshwater environments, towards the end of the early Pliocene (the late Dacian local age). Additional fish material was recently described from the late Pliocene (the Romanian local stage) of Podari, consisting of isolated teeth, cranial bones, vertebrae, and fin rays, identified as belonging to Esox sp., Silurus sp., Scardinius sp., and Tinca sp. (Trif et al., 2016), suggesting a typical freshwater environment was already present in the Dacian Basin at the end of the Pliocene.
GEOLOGICAL SETTING
 The fossil material described in this paper was collected from the sediments cropping out at the left bank of Argeș River, at Copăceni (Ilfov County), a village located about 20 km southwards of Bucharest (Figure 1). In a couple of recent and comprehensive reviews of the Plio-Pleistocene stratigraphic units of the Dacian Basin (Andreescu et al., 2011, 2013), the sedimentary succession from Copăceni is provisionally left unformalized, being referred to as the “Copăceni beds”, a unit that, due to its silty-clayey lithology, stands out from the lower Pleistocene Frătești Formation represented by sands and pebbles. The same authors suggest that the “Copăceni beds” might represent a distinct formation, or the basal sub-unit of the lithologically similar Coconi Formation (a unit known from boreholes in the area surrounding Bucharest, informally known as “the Marly Complex”, term proposed by Liteanu, 1952).
The fossil material described in this paper was collected from the sediments cropping out at the left bank of Argeș River, at Copăceni (Ilfov County), a village located about 20 km southwards of Bucharest (Figure 1). In a couple of recent and comprehensive reviews of the Plio-Pleistocene stratigraphic units of the Dacian Basin (Andreescu et al., 2011, 2013), the sedimentary succession from Copăceni is provisionally left unformalized, being referred to as the “Copăceni beds”, a unit that, due to its silty-clayey lithology, stands out from the lower Pleistocene Frătești Formation represented by sands and pebbles. The same authors suggest that the “Copăceni beds” might represent a distinct formation, or the basal sub-unit of the lithologically similar Coconi Formation (a unit known from boreholes in the area surrounding Bucharest, informally known as “the Marly Complex”, term proposed by Liteanu, 1952).
There are, however, some contradictions regarding the age of the Coconi Formation. Andreescu et al. (2013: figure 2, figure 6) considered it to be a middle Pleistocene unit, accumulated during the Netindavian, the early Dinogetian local age, without offering any arguments for this assignment, whereas Andreescu et al. (2011) pointed out that the mollusk assemblage is irrelevant for dating this unit, but the presence of Prolagurus pannonicus and Allophaiomys pliocaenicus in a borehole sample from Fierbinți (Rădulescu et al., 1997) supports the late early Pleistocene age of the Coconi Formation. In spite of agreeing to this conclusion, Andreescu et al. (2011: figure 2) depicted the Coconi Formation as largely Dinogetian (i.e., middle Pleistocene).
The “Copăceni beds” are considered to have accumulated during the late early Pleistocene (late Argedavian local age = Uzunian sub-age), based on the mollusk assemblage (Andreescu et al., 2011), and due to the presence of Mimomys savini, Allophaiomys pliocaenicus, and Lagurodon arankae (Andreescu et al., 2011; Știucă et al., 2012; Vasile et al., 2015). Taking this information into account, and pending research on the small mammalian assemblage of the Coconi Formation, we consider the “Copăceni beds” (1.1-1.0 Ma) as representing the basal member of a lower Pleistocene Coconi Fm., in which only the upper part might be of middle Pleistocene age.
The sedimentary succession near Copăceni (Figure 1.3), in the area of the Children’s Park, appears to be continuous, but several distinct beds can be identified. The base of the succession is not accessible, being covered by the Argeș River. Only the uppermost 20-30 cm of a silty mudstone layer, rich in carbonate concretions (hence referred herein as the concretionary layer) is accessible when the river level is low, during late summer and early autumn. This layer is not very rich in large vertebrate fossils, but it yielded several proboscidean and cervid bones. The concretionary layer is covered by fine sands, 15-20 cm thick, rich in unionid bivalve and microgastropod shells (referred herein as the lower coquina). The lower coquina is covered by silty mudstones (referred herein as the intermediate mudstones), which are 150-180 cm thick, and rich in large mammalian remains including proboscideans, rhinocerotids, and bovids. The uppermost layer, covered by modern soil, consists of 15-30 cm of fine sands with gastropod shells and a few rhinocerotid and bovid remains (referred herein as the upper coquina).
Mollusk fossil remains are very common in the lower and upper coquinas. Andreescu et al. (2011) listed bivalves Psudosturia caudata, Bogatschevia cf. scutum, B. sturi, B. cf. tamanica, Wenziella ex gr. wilhelmi, Potomida ovata, Margaritifera cf. arca, Crassiana pseudodavilai, Unio cf. chasaricus, and U. kujalnicensis, but these taxa have not been described in detail so far.
Large mammalian remains can be found in situ or scattered along the river banks, after intense bank erosion occurs during autumn and spring, when the waters of the Argeș River are the highest. These large mammalian remains drew attention on the fossiliferous potential of the site. Proboscidean molar and postcranial bone fragments are the most common findings at the site, and were described in detail. Most of the proboscidean material was assigned to Mammuthus meridionalis, but some specimens were suspected as belonging to Elephas antiquus (Vasile et al., 2013a). Incipient study of the rhinocerotid material suggests that they belong to Stephanorhinus etruscus (Flintașu, 2016; Flintașu et al., 2019), whereas other large specimens were assigned to the cervid Praemegaceros pliotarandoides, and to the bovid Leptobos vallisarni. A single coprolite suggests the presence of a hyaenid carnivore (Știucă et al., 2012; Vasile et al., 2015).
Following the discovery of large mammalian remains, screen-washing of sediments was used in order to identify biostratigraphically-relevant small mammals. Isolated molars were assigned to Mimomys savini, Allophaiomys pliocaenicus, and Lagurodon arankae, but some specimens similar in morphology to Desmana cf. radulescui, Allactaga sp., and a castorid (Trogontherium cf. dacicum) were also mentioned (Știucă et al., 2012; Vasile et al., 2015).
Ectothermic vertebrates were also found during screen-washing, and preliminarily assigned to fishes (Squalius sp., Rutilus sp., Scardinius sp., Carassius sp., Tinca sp., and Silurus sp.), amphibians (Rana sp., Bufo sp., Pelobates sp., Pelophylax sp., Triturus sp., and Lissotriton sp.), lizards (Lacerta sp.), ophidians (Natrix tessellata, and Coronella sp.), but, except for the fish taxa, described in this paper, none of the groups mentioned above were so far described in detail (Vasile et al., 2013a, b, 2015, 2016, 2018).
MATERIAL METHODS
Approximately 1500 kg of sediment were processed in order to obtain small vertebrate remains, being evenly collected from the three main lithological horizons described above: the lower coquina, the intermediate mudstone, and the upper coquina (Figure 1.3).
The sediment collected from the strata cropping out along the Argeș River was dried to remove water contributing to the rock’s cohesion. The sediment was subsequently soaked into water containing a small amount (ca. 5 %) of acetic acid, in order to dissolve the carbonate matrix of the mudstone. The soaked sediment was screen-washed, using a battery of two sieves, with 2 mm and 0.75 mm mesh sizes. The concentrate resulted by removal of the silty and clayey fractions was sorted under the binocular, and fossil remains were extracted.
The total number of processed fish remains is 130 specimens. Of these, 17 (13.1 %) are determined to species, 94 (72.3 %) to the genus, and 19 (14.6 %) up to the family level. Isolated pharyngeal teeth and their fragments numerically predominate among the fish remains in the collection, while jaw teeth, pharyngeal bone fragments, fin rays, and vertebrae centra are less numerous. Cranial elements (premaxilla, dentary, opercle) are represented by a single bone each. Specimens that could be identified taxonomically at least to family level were inventoried and stored in the collection of the “Emil Racoviță” Institute of Speleology, Bucharest.
The systematic affiliation was determined under the generally accepted methodology using the comparative osteological collection of the Department of Paleontology, National Museum of Natural History of the National Academy of Sciences of Ukraine (Kyiv). Ichthyological taxonomy used in the paper follows Nelson et al. (2016), and Froese and Pauly (2019). The names of skeletal elements are consistent with the nomenclature of Lepiksaar (1994) and Radu (2005). Terminology for pharyngeal teeth follows Rutte (1962) and Sytchevskaya (1989). Palaeoecological analysis was conducted using the actualistic approach.
Abbreviations. Co-P, Copăceni-Park Site; ISER, “Emil Racoviță” Institute of Speleology, Bucharest; NISP, number of identified specimens.
RESULTS OF THE STUDY
Following screen-washing procedures, the fossil specimens belonging to actinopterygian fishes proved to be among the most abundant. Most of the specimens consist of isolated teeth, making taxonomical assignment difficult at the species level. Following thorough morphological and morphodimensional comparison to extant fishes and to fossil material, most specimens were determined to generic level, whereas some material was only identified to family level.
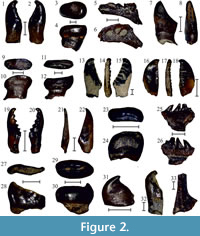 Leuciscus sp. is represented by eight isolated pharyngeal teeth (ISER Co-P 32, 50, 55a-b, 73, 80a-c), both complete and fragmented. Slender pharyngeal teeth have elongated and high crowns, cylindrical pedicle, arched or almost straight back, and small blunt hook at the tip. The grinding surface is narrow, with an elevated wrinkled edge (Figure 2.1-2.2).
Leuciscus sp. is represented by eight isolated pharyngeal teeth (ISER Co-P 32, 50, 55a-b, 73, 80a-c), both complete and fragmented. Slender pharyngeal teeth have elongated and high crowns, cylindrical pedicle, arched or almost straight back, and small blunt hook at the tip. The grinding surface is narrow, with an elevated wrinkled edge (Figure 2.1-2.2).
Ten isolated molariform pharyngeal teeth (ISER Co-P 4, 28a-d, 65a-e) in the material from Copăceni were assigned as belonging to Rutilus cf. frisii (Nordmann, 1840). They are quite large (up to 4.4 mm in height), with fungiform, laterally compressed crown and straight tooth back. The hook at the tip is absent, and the convex grinding surface is eroded. The tooth belly is rounded, hanging above the neck, while the pedicle is oval in cross-section (Figure 2.3-2.4).
Rutilus sp. is represented by one dentary fragment with rounded symphysial edge (ISER Co-P 7), one pharyngeal bone fragment (ISER Co-P 6), and 10 isolated pharyngeal teeth (ISER Co-P 30, 34, 37, 40, 44, 59, 81a-d). The dorsal edge of the dentary is narrow and forms a sharp ridge. The high pharyngeal bone is represented by a small fragment. There was two-line dentition (three teeth in the main row, and two pharyngeal teeth in the second row), but only compressed pedicles are preserved on the bone surface. The cavernous surface is narrow, with three subtriangular caverns having rounded edges (Figure 2.5-2.6). Isolated pharyngeal teeth have laterally compressed, wide and low molariform crowns without the hook at their tips. The pedicle is suboval in cross-section. The wide grinding surface is slightly concave and obliquely cut towards the rounded and narrow belly. There is a clearly defined tooth neck (Figure 2.7-2.12).
Seven isolated pharyngeal teeth (ISER Co-P 26a-g) are similar in morphology to those of Scardinius ponticus Kovalchuk, 2014, described from the late Miocene and Pliocene of Ukraine (Kovalchuk, 2014, 2017). These teeth have high and laterally compressed crowns. The narrow and curved tooth back rises upwards, expanding in the rounded hook, which is separated by the distinct furrow from the grinding edge. The grinding surface is long and flattened, being almost parallel to the tooth back. The anterior grinding edge is sculpted by eight convex cogs, separated from each other by protruding ridges. The small belly is laterally compressed. Tooth pedicle is cylindrical in cross-section and narrowed at the base (Figure 2.13-2.15).
There are three other pharyngeal tooth fragments representing the genus Scardinius in the processed material (ISER Co-P 70a-b, 79). These specimens differ from other known species of this genus in the presence of an enormous number of cogs (9 and 13) at the anterior grinding edge. Tooth back is straight, with a small hook at the tip (Figure 2.16-2.18). Pending more complete material, that might present additional features to support the description of a distinct species, we provisionally assign these specimens as belonging to Scardinius sp. Copăceni. In addition, there are nine pharyngeal teeth of Scardinius sp. (ISER Co-P 54a-c, 62, 71a-d, 78) with a blunt hook at the tip, six cogs at the grinding surface, straight or slightly curved back, rounded belly, and clearly pronounced neck (morphology exemplified by specimen ISER Co-P 54b, Figure 2.19-2.20).
Chondrostoma sp. is represented by nine isolated pharyngeal teeth (ISER Co-P 35, 36, 48, 72a-d, 82a-b). The cultriform teeth have elongated, compressed, and oblique crowns, while the hook at the tip is absent. The grinding surface is narrow and smooth. The cylindrical pedicle is short, tooth back is almost straight, with a distinct neck and rounded belly (Figure 2.21-2.22).
Remains belonging to Barbus sp. are quite numerous in the material from Copăceni. There are three small pharyngeal bone fragments with one or two preserved teeth (ISER Co-P 53a-b, 66), as well as seven isolated pharyngeal teeth (ISER Co-P 31, 38, 47, 51, 61, 75a-b). The latter have a cylindrical pedicle, straight back, high spatulate crowns with a small blunt hook at the tip, and wide grinding surface. The tooth belly is small and convex, and the neck is absent or weakly expressed (Figure 2.23-2.24). A small fragment of ceratotrichium (ISER Co-P 69) with a distinct external edge was identified as belonging to Barbinae gen. et sp. indet. (Figure 2.25-2.26).
Carassius sp. is represented by two pharyngeal teeth (ISER Co-P 5, 43) having a laterally compressed crown, arcuate back, and rounded belly. The medial surface is concave. The slightly curved and narrow grinding surface is surrounded by low edges (Figure 2.27-2.28).
Nine isolated pharyngeal teeth, both complete and fragmented, in the Copăceni sample were assigned to Tinca sp. (ISER Co-P 29, 42, 57, 58, 64, 74a-c, 77). Their crowns are low (up to 2.0 mm in height) and flattened (3.6-4.0 mm in width). The tooth neck is not clearly expressed, with an arched back and a weak widened hook at the tip. The tooth belly is slightly convex, with narrow, laterally compressed grinding surface. The grinding edge is obliquely bevelled towards the tooth belly. A groove at the anterior edge of the grinding surface is indented by transverse wrinkles and forms a keel-form curl at the crown surface. The pedicle is oval (Figure 2.29-2.30).
Two pharyngeal teeth (ISER Co-P 49, 60) with a narrow gracile crowns, small thin hook at the tip, widened middle part of the grinding surface and rounded pedicle (Figure 2.31), resemble those in representatives of the genus Abramis, and therefore were assigned to Abramis sp.
Squalius sp. is represented by two isolated pharyngeal teeth (ISER Co-P 33, 46) with high elongated crowns, cylindrical pedicle and almost straight back (Figure 2.32).
It is not possible to clearly identify the systematic affiliation of some fish remains from Copăceni closer than to family level due to their poor preservation. Those are one small opercular fragment (ISER Co-P 2) with lenticular-shaped articular facet located near the upper edge of the bone, well-developed striae, smooth external surface and concave internal one; a pharyngeal bone fragment (ISER Co-P 16, Figure 2.33), as well as two pharyngeal tooth fragments (ISER Co-P 55a-b). These specimens were left here as Cyprinidae gen. et sp. indet.
 Silurus sp. is represented by three complete and fragmented pectoral spines (ISER Co-P 10, 15, 23) and one cleithrum fragment (ISER Co-P 24). All these bones belonged to young individuals. The small pectoral spine is gracile, oval in cross-section, with longitudinal striations along its shaft. The articular head of the spine is compact and flattened, with a finely porous surface. The medial and lateral margins of the shaft have numerous small and smoothed weak denticles starting almost from the articular head. The denticles on the medial margin are smaller and located more densely, whereas those on the lateral margin are larger and irregular, their tips are directed proximally. There is a deep and elongated articular facet for attaching the pectoral spine to the medial part of the cleithrum (Figure 3.1-3.4). Other catfish remains (e.g. cranial bone fragment ISER Co-P 14 (Figure 3.5), three isolated trunk vertebrae ISER Co-P 12a-b, 17) are partially broken. The centra are compressed anteroposteriorly and dorsoventrally, with almost flattened surface. We assign these fossils as belonging to Siluridae gen. et sp. indet.
Silurus sp. is represented by three complete and fragmented pectoral spines (ISER Co-P 10, 15, 23) and one cleithrum fragment (ISER Co-P 24). All these bones belonged to young individuals. The small pectoral spine is gracile, oval in cross-section, with longitudinal striations along its shaft. The articular head of the spine is compact and flattened, with a finely porous surface. The medial and lateral margins of the shaft have numerous small and smoothed weak denticles starting almost from the articular head. The denticles on the medial margin are smaller and located more densely, whereas those on the lateral margin are larger and irregular, their tips are directed proximally. There is a deep and elongated articular facet for attaching the pectoral spine to the medial part of the cleithrum (Figure 3.1-3.4). Other catfish remains (e.g. cranial bone fragment ISER Co-P 14 (Figure 3.5), three isolated trunk vertebrae ISER Co-P 12a-b, 17) are partially broken. The centra are compressed anteroposteriorly and dorsoventrally, with almost flattened surface. We assign these fossils as belonging to Siluridae gen. et sp. indet.
Remains of salmonid fishes (Salmonidae gen. et sp. indet.) are represented by eight complete and partially broken vertebrae (ISER Co-P 1, 3, 11a-d, 13a, 26a). Their centra are small (0.5-5.0 mm in diameter, mean 2.8 mm), with deeply concave, subrectangular anterior surface and diamond-shaped posterior one. One centrum is hexagonal in shape. The aperture of the notochord is displaced dorsally. There is a network of narrow interlaced cords at the ventral surface of the centra, as well as open lattice-shaped structure (longitudinal flattened bone strips and rounded pores) at their lateral surface (Figure 3.6-3.17).
There are numerous isolated teeth of pikes in the Copăceni sample. Most of them (ISER Co-P 25, 41, 45a-b, 52, 63a-g, 76a-d) are elongated and slender jaw teeth of different size (up to 10 mm) having two sharp edges. Their cross-section is interiorly smooth and exteriorly convex (morphology exemplified by specimens ISER Co-P 25 and 52, illustrated in Figure 3.18-3.19, and Figure 3.20-3.21, respectively). Another six pike teeth (ISER Co-P 27, 42, 45c, 63h, 68, 76e) are smaller and significantly differ in morphology. Their base is circular, and the crown is slightly or moderately curved (Figure 3.22-3.26). These teeth most probably come from the palatine. In addition, there are three small vertebrae (3-4 mm in diameter; ISER Co-P 18, 19, 39) with deeply concave and circular anterior and posterior surfaces, as well as clearly expressed annual rings. Lateral surface of these vertebrae is perforated by the large rectangular pores with rounded edges (Figure 3.27-3.30). All these specimens were assigned as belonging to Esox sp. A clearer identification is not possible due to their poor preservation (see Kovalchuk et al., 2017).
Perca sp. is represented in the Copăceni sample by one complete spiny ray of the first dorsal fin (ISER Co-P 9). The fin ray (Figure 3.31-3.33) is narrow and long (16.2 mm), pointed at the tip. Its base is widened, there are small lateral pits from both sides, the anterior surface is sculpted by flattened ridges, while the posterior one has sharp edges. The proximal part of the anal fin ray (ISER Co-P 13) with a wide base and two circular articulation surfaces was also assigned to Perca sp. We identified a small fragment of premaxilla (ISER Co-P 67, Figure 3.34-3.35) and one broken vertebra centrum (ISER Co-P 22) as belonging to Percidae gen. et sp. indet.
DISCUSSION
The early Pleistocene freshwater fish assemblage of Copăceni consists of 17 species within 12 genera belonged to the families Cyprinidae, Siluridae, Salmonidae, Esocidae, and Percidae. Cyprinids predominate in the number of identified species over all other families representing by individual taxa. It should be noted that isolated pharyngeal teeth are difficult or even impossible to identify to lower taxonomic levels because of the high morphological variability depending on their position on the pharyngobranchial bone, ontogeny, wear, and fishes’ diet (Tadajewska, 1998; van den Hoek Ostende et al., 2015). Therefore, our identifications of the cyprinid remains from Copăceni up to the generic level should be considered as tentative.
All processed skeletal elements are morphologically identical to those in living genera and almost all of them - to those in extant species. The majority of the remains were those of pike Esox sp. (NISP = 24; 18.3 %). The roach Rutilus sp., barbel Barbus sp., Pontic roach Rutilus cf. frisii, and rudd Scardinius sp. were of minor importance numerically (NISP = 12, 11, 10, 9, respectively). Other species are recorded by a fewer number of identified specimens. The minimum number of remains in the Copăceni sample belong to the bream Abramis sp., crucian carp Carassius sp., and perch Perca sp. (namely, two bones for each species).
Certain fish taxa listed in the faunal assemblage of Copăceni are also known from other Pleistocene localities in Europe. For example, remains of the roach, bream, pike and perch were identified among the osteological material from Greenlands Pit (Schreve et al., 2002) and West Runton (Böhme, 2010), both Middle Pleistocene in age. The ichthyofauna similar in species composition is known from the Stuttgart Wilhelma Bärengehege (Böttcher, 1994) and Rabutz (Böhme and Ilg, 2003) in Germany. Pawłowska (1963) noted the presence of Rutilus, Abramis, Carassius, Esox, and Perca in the interglacial deposits of Barkowice near Sulejów in Poland. All fish species identified in Copăceni are common in Pleistocene-aged assemblages of Ukraine - Lysa Gora 1 (Rekovets et al., 2014) and Medzhybizh (Kovalchuk, 2017). The fish species list of Copăceni is similar to those for the early Pleistocene Yayladag Site in Turkey (Vasilyan et al., 2014), and the middle Pleistocene Bobovek locality in Slovenia (Gaudant, 1978).
With so few occurrences of freshwater fish assemblages, spread across the entire timespan of the Pleistocene, palaeobiogeographic reconstructions remain difficult to obtain. Nonetheless, each new occurrence adds some information to the existing data. The early Pleistocene deposits from Copăceni were tentatively correlated, based on mollusk and rodent assemblages, to the 8th terraces of the Prut and Dniester rivers (Andreescu et al., 2011, 2013). Although the composition of the mollusk assemblage from Copăceni is only tentative, the correlation between mammalian assemblages from Copăceni and those from the early Pleistocene fluvial deposits of Ukraine and the Republic of Moldova (e.g., Matoshko et al., 2004) support this conclusion. The composition of the fish assemblage from Copăceni is quite similar to those of the Nogaiskian ichthyofaunal assemblage of southeastern Europe (Kovalchuk, 2017). The sedimentology of the deposits from the Dacian Basin (Andreescu et al., 2011, 2013) and that of the coeval units from of Southern Moldova and Ukraine (Matoshko et al., 2004; Nadachowski et al., 2006) suggests, however, that during the early Pleistocene most of the basins were filled with fluvial sediments, and no continuous lake that would ensure dispersion throughout the region existed. A probable isolation of the fish faunas from the Dacian Basin is also supported by a higher diversity of cyprinids, as opposite to the case recorded from Nogaisk, where cyprinid diversity decrease was linked to a general decrease of temperature compared to the late Pliocene (Kovalchuk, 2017). The Dacian Basin may have thus represented a refugium for thermophilous taxa during the early Pleistocene.
A transfer of taxa between the Dacian Basin and the Moldo-Ukrainian area north of the Black Sea seems more probable to have occurred during the late Pliocene. The presence of a large freshwater lake in the Dacian Bain, continuous from west to east, is supported by the mollusk faunas and otolith assemblage (Pană, 1982). Evidence of connectivity to other freshwater basins to the east is not very well documented, but the study of mollusk faunas (Pană and Müntz, 2003: figure 3) suggest a corridor existed during the late Pliocene between the area north-east of the Dacian Basin and the area of southern Republic of Moldova and southern Ukraine (i.e., using the terms of Pană and Müntz, 2005, a connection across the Bârlad Depression between the Pericarpathian Depression and the Predobrogean Depression). The freshwater fish assemblage reported from the late Pliocene of Podari (Trif et al., 2016), albeit not very diverse, includes similar taxa to the late Shirokinian ichthyofaunal assemblages of southeastern Europe (Kovalchuk, 2017), supporting the idea that some connectivity between the two areas existed prior to the end of the Pliocene. More evidence in this respect could be provided by further research in eastern Romania, where rich mammalian faunas of early Pliocene age were described from Berești and Mălușteni, with freshwater fishes mentioned from the same deposits (Simionescu, 1932a, b). A revision of the material mentioned from these sites is needed.
Blanck et al. (2007) postulated that freshwater fishes can use a large diversity of habitats. It is difficult to provide a clear palaeoecological interpretation of the Copăceni assemblage based on such a poor material due to a wide ecological diversity within genera (Vasilyan et al., 2014).
Dace (Leuciscus), roach (for instance, Rutilus frisii), nase (Chondrostoma), and salmonids (e.g., trout Salmo trutta) prefer moderate to fast-flowing streams and rivers with and rock or gravel bottom (Kottelat and Freyhof, 2007). Barbel (Barbus) is also a rheophylic and lithophilous fish (Kotlik et al., 2004; Britton and Pegg, 2011). It inhabits moderately flowing waters with a sandy or gravel bottom. The occurrence of the rudd (Scardinius), crucian carp (Carassius), and tench (Tinca) indicates the presence of well-developed underwater vegetation. These fish tend to feed in areas with a large supply of macrophytes (Nordstrom, 2011) and prefer standing to slowly flowing waters and muddy bottom (Moreno Rendón et al., 2003).
The catfish (Silurus) inhabits large and medium-sized lowland rivers and backwaters. This fish is an impressive predator with a wide range of food items (Copp et al., 2009). The pike (Esox) is a cool water fish with a wide environmental tolerance (Casselman and Lewis, 1996). It inhabits shallow and moderately productive waters (Craig, 2008; Harvey, 2009). The perch (Perca) also prefers the areas with slowly flowing water and thickets of vegetation (Eklöv, 1997).
Summing up the palaeoecological evidence, it can be assumed that the Copăceni fish assemblage lived in a large lowland mesotrophic river. There were rapids with fast-flowing, well-aerated water over sandy or pebbly riverbeds, areas with slowly flowing water, muddy or fine-sandy bottom and thickets of underwater vegetation, as well as backwaters. The Copăceni fish assemblage was adapted to temperate or warm climate. This is indicated by the predominance of thermophilic elements in its composition and a relatively small portion of cryophilic forms.
CONCLUSIONS
The freshwater fish fauna from the early Pleistocene of Copăceni is the first Pleistocene ichthyofaunal assemblage described from the Dacian Basin, southern Romania. It shows high taxonomic diversity, including 17 taxa comprised in 10 genera belonging to the families Cyprinidae (the best represented), Siluridae, Salmonidae, Esocidae, and Percidae. The faunal composition is comparable to that of other southeast European assemblages, but it includes more thermophilic taxa, suggesting the Dacian Basin represented an early Pleistocene refugium in a cooling post-Pliocene environment. Comparison between the late Pliocene and early Pleistocene fish assemblages of the Dacian Basin and other ichthyofaunas of southeastern Europe, north of the Euxinian Basin, suggests a connection between the two areas existed during the late Pliocene, allowing for freshwater fish taxa interchange. The ecological affinities of the identified taxa show that fishes from Copăceni lived in a large lowland mesophitic river, with parts of rapid waters flowing over pebbly or sandy riverbeds, but also areas of muddy waters, rich in vegetation.
ACKNOWLEDGEMENTS
The authors dedicate this paper to the memory of Dr. E. Știucă (“Emil Racoviță” Institute of Speleology, Bucharest) and his initiative to search for vertebrate fossils at Copăceni. O. Flintașu, R. Ilie, M.O.B. Săvescu, S. Radu, and P. Răileanu are thanked for their help in collecting and transporting sediment samples. We express our sincere thanks to two anonymous reviewers for their constructive suggestions and fruitful advice. This work was supported by a grant of the Romanian Ministry of Research and Innovation, CNCS - UEFISCDI, project number PN-III-P1-1.1-PD-2016-0848, within PNCDI III (to Ș.V.). The study also benefited from financial support from grants PCE 197/2016, PCCF 16/2018, and from the EEA-Norway Grant #0126 (KARSTHIVES) (all to A.P.).
REFERENCES
Andreescu, I., Codrea, V., Enache, C., Lubenescu, V., Munteanu, T., Petculescu, A., Știucă, E., and Terzea, E. 2011. Reassessment of the Pliocene/Pleistocene (Neogene/Quaternary) boundary in the Dacian Basin (Eastern Paratethys), Romania. Oltenia. Studii și comunicări. Științele Naturii, 27:197-220.
Andreescu, A., Codrea, V., Lubenescu, V., Munteanu, T., Petculescu, A., Știucă, E., and Terzea, E. 2013. New developments in the Upper Pliocene-Pleistocene stratigraphic units of the Dacian Basin (Eastern Paratethys), Romania. Quaternary International, 284:15-29. https://doi.org/10.1016/j.quaint.2012.02.009
Apostol, L. 1965. Sur la presence d’ Anancus arvernensis (Croizet et Jobert) et d’ Archidiskodon meridionalis Nesti sur le territoire de Curtea de Argeș. Travaux du Muséum d’Histoire Naturelle “Grigore Antipa”, 5:455-462.
Apostol, L. 1968. Particularité morphologiques des molaires de proboscidiens fossils quaternaires de Roumanie, conserves dans la collection du Musée d’Histoire Naturelle “Grigore Antipa”. Travaux du Muséum d’Histoire Naturelle “Grigore Antipa”, 9:581-616.
Apostol, L. 1972. Les mammifères pléistocènes de la zone de Prundu, district d’Ilfov. Travaux du Muséum d’Histoire Naturelle “Grigore Antipa”, 12:427-438.
Apostol, L. 1976. L’étude morphométrique des mammifères fossils quaternaires de la Plaine Roumaine et leur distribution paléozoogéographique. Travaux du Muséum d’Histoire Naturelle “Grigore Antipa”, 17:341-375.
Apostol, L. 1981. Mammifères pléistocènes de la zone de București et de ses alentours. Travaux du Muséum d’Histoire Naturelle “Grigore Antipa”, 23:305-312.
Apostol, L. and Cacoveanu, I. 1980. L’étude de restes fossils quaternaires d’Eléphantidés, de Bovidés et de Cervidés conserves dans les musées des villes d’Alexandria et de Roșiorii de Vede (Département de Teleorman, Roumanie). Travaux du Muséum d’Histoire Naturelle “Grigore Antipa”, 22:587-607.
Apostol, L. and Stancu, R. 1968. Resturi de Archidiskodon meridionalis (Nesti) descoperite în județul Argeș. Studii și Comunicări, Istorie - Științele Naturii, 1:163-176.
Apostol, L. and Vicoveanu, D. 1970. L’étude des eléphantidés, des rhinocéridés et de bovidés des depots quaternaires de la vallée inférieure du Bîrlad, existants au Musée de Tecuci (Dép. Galatzi). Travaux du Muséum d’Histoire Naturelle “Grigore Antipa”, 10:365-382.
Athanasiu, S. 1915. Resturile de mamifere pliocen superioare de la Tulucești. Anuarul Institutului Geologic al României, 6:408-415.
Bannikov, A.F., Scwarzhans, W., and Carnevale, G. 2018. Neogene Paratethyan croakers (Teleostei, Sciaenidae). Rivista Italiana di Paleontologia e Stratigrafia, 124:535-571. https://doi.org/10.13130/2039-4942/10696
Blanck, A., Tedesco, P.A., and Lamouroux, N. 2007. Relationships between life-history strategies of European freshwater fish species and their habitat preferences. Freshwater Biology, 52:843-859. https://doi.org/10.1111/j.1365-2427.2007.01736.x
Böhme, M. 2010. Ectothermic vertebrates, climate and environment of the West Runton Freshwater Bed (early Middle Pleistocene). Quaternary International, 228:63-71. https://doi.org/10.1016/j.quaint.2010.06.021
Böhme, M. and Ilg, A. 2003. Database of Vertebrates: Fossil Fishes, Amphibians, Reptiles and Birds (fosFARbase) Localities and Taxa from the Triassic to the Neogene. www.wahre-staerke.com. Accessed 24-03-2019.
Böttcher, R. 1994. Niedere Wirbeltiere (Fische, Amphibien, Reptilien) aus dem Quartär von Stuttgart. Stuttgarter Beiträge zur Naturkunde B, 215:1-75.
Britton, J.R. and Pegg, J. 2011. Ecology of European barbel Barbus barbus: implications for river, fishery, and conservation management. Reviews in Fisheries Science, 19:321-330. https://doi.org/10.1080/10641262.2011.599886
Casselman, J.M. and Lewis, C.A. 1996. Habitat requirements of northern pike (Esox lucius). Canadian Journal of Fisheries and Aquatic Science, 53:161-174. https://doi.org/10.1139/f96-019
Copp, G.H., Britton, J.R., Cucherousset, J., García-Berthou, E., Kirk, R., Peeler, E., and Stakènas, S. 2009. Voracoius invader or benign feline? A review of the environmental biology of European catfish Silurus glanis in its native and introduced ranges. Fish and Fisheries, 10:252-282. https://doi.org/10.1111/j.1467-2979.2008.00321.x
Craig, J.F. 2008. A short review of pike ecology. Hydrobiologia, 601:5-16. https://doi.org/10.1007/s10750-007-9262-3
Croitor, R. and Popescu, A. 2011. Large-sized ruminants from the Early Pleistocene of Leu (Oltenia, Romania) with remarks on biogeographical aspects of the “Pachycrocuta event”. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, 261:353-371. https://doi.org/10.1127/0077-7749/2011/0180
Eklöv, P. 1997. Effects of habitat complexity and prey abundance on the spatial and temporal distributions of perch (Perca fluviatilis) and pike (Esox lucius). Canadian Journal of Fisheries and Aquatic Sciences, 54:1520-1531. https://doi.org/10.1139/f97-059
Feru, M., Radulesco, C., and Samson, P. 1978. Biostratigraphie (Micromammifères) des dépôts plio-pléistocènes de la zone de Slatina (dép. d’Olt). Travaux de l’Institut de Spéologie “Émile Racovitza”, 17:117-134.
Feru, M., Radulesco, C., and Samson, P. 1979. Biostratigraphie (Micromammifères) des dépôts plio-pléistocènes du domaine gétique de la Dépression valaque. Travaux de l’Institut de Spéologie “Émile Racovitza”, 18:141-169.
Feru, M.U., Rădulescu, C., and Samson, P. 1983. Succession des mammifères plio-pléistocènes dans le Bassin Dacique (Roumanie). Anuarul Institutului de Geologie și Geofizică, 59:161-167.
Flintașu, O. 2016. Evaluarea taxonomică a resturilor de rinoceri din Pleistocenul inferior de la Copăceni, p. 27-30. In Lorinț, C.R. (ed.), Lucrările celui de-al XIV-lea Simpozion Național Studențesc “Geoecologia”, Petroșani.
Flintașu, O., Vasile, Ș., and Dumitrache, I.L. 2019. Taxonomical analysis of the early Pleistocene rhinocerotid fossil remains from Copăceni (Dacian Basin, southern Romania), p. 26. In Blanco, F., Carro Rodríguez, P.M., Domínguez García, A.C., Gamboa, S., Linares Ruiz, M., Mateos-Carralafuente, J.R., Menéndez, I., Martín Perea, D.M., MartínezMonleón, M., Martínez Nebreda, S., Navalón, G., Porras, L., Prieto, I., Püschel, T.A., Ruiz-Galván, A. (eds.), Book of Abstracts of the 4th IMERP, Cuenca.
Froese, R. and Pauly, D. Editors. 2019. FishBase. World Wide Web electronic publication. http://www.fishbase.se, version (04/2019).
Gaudant, J. 1978. Sur la présence de Leuciscus leuciscus (L.) (Poisson téléostéen, Cypriniformes) dans le pléistocène de Bobovek, près de Kranj (Slovénie, Yugoslavie). Razprave Slovenska Akademija Znanosti in Umetnosti, Rzred za Prirodoslovne Vede, 4:48-57.
Harvey, B. 2009. A biological synopsis of northern pike (Esox lucius). Canadian Manuscript Report of Fisheries and Aquatic Sciences, 2885:i-v+1-31.
Jipa, D.C. and Olariu, C. 2009. Dacian Basin. Depositional Architecture and Sedimentary History of a Paratethys Sea. Geo-Eco-Marina, Special Publication 3.
Kotlik, P., Bogutskaya, N.G., and Ekmekçi, F.G. 2004. Circum Black Sea phylogeography of Barbus freshwater fishes: divergence in the Pontic glacial refugium. Molecular Ecology, 13:87-95. https://doi.org/10.1046/j.1365-294X.2003.02021.x
Kottelat, M. and Freyhof, J. 2007. Handbook of European Freshwater Fishes. Publications Kottelat, Cornol, Switzerland.
Kovalchuk, O.M. 2014. A new extinct carp fish species (Teleostei, Cyprinidae) from the Late Neogene of Southeastern Europe. Vestnik Zoologii, 48:411-418. https://doi.org/10.2478/vzoo-2014-0049
Kovalchuk, O.M. 2017. Regional fish-based biostratigraphy of the Late Neogene and Pleistocene of southeastern Europe. Vestnik Zoologii, 51:375-392. https://doi.org/10.1515/vzoo-2017-0045
Kovalchuk, O.M., Wilson, M.V.H., and Grande, T. 2017. A review of Neogene and Quaternary pikes of southeastern Europe and a new species from the early Pleistocene of Nogaisk, Ukraine. Acta Palaeontologica Polonica, 62:121-135. https://doi.org/10.4202/app.00311.2016
Lepiksaar, J. 1994. Introduction to Osteology of Fishes for Paleozoologists. University Press, Göteborg.
Lister, A.M. and van Essen, H. 2003. Mammuthus rumanus (Ștefănescu) the earliest mammoth in Europe, p. 47-52. In Știucă, E. and Petculescu, A. (eds.), Advances in Vertebrate Paleontology “Hen to Panta” - a tribute to Constantin Rădulescu and Petre Mihai Samson. Romanian Academy, “Emil Racoviţă” Institute of Speleology, Bucharest.
Liteanu, E. 1952. Geologia zonei orașului București. Studii Tehnice și Economice, Seria E, Hidrogeologie, 1:1-83.
Markov, G.N. and Spassov, N. 2003. Primitive mammoths from Northeast Bulgaria in the context of the earliest mammoth migrations in Europe, p. 53-58. In Știucă, E. and Petculescu, A. (eds.), Advances in Vertebrate Paleontology “Hen to Panta” - a tribute to Constantin Rădulescu and Petre Mihai Samson. Romanian Academy, “Emil Racoviţă” Institute of Speleology, Bucharest.
Matoshko, A.V., Gozhik, P.F., and Danukalova, G. 2004. Key Late Cenozoic fluvial archives of Eastern Europe: the Dniester, Dnieper, Don and Volga. Proceedings of the Geologists’ Association, 115:141-173. https://doi.org/10.1016/S0016-7878(04)80024-5
Moreno Rendón, P., Martín Gallardo, J., García Caballos, E., Peréz, R., and Escudero García, J.C. 2003. Determination of substrate preferences of tench, Tinca tinca (L.), under controlled experimental conditions. Journal of Applied Ichthyology, 19:138-141. https://doi.org/10.1046/j.1439-0426.2003.00469.x
Nadachowski, A., Mirosław-Grabowska, J., David., A., Tomek, T., Garapich, A., Pascaru, V., Obadă, T., and Szyndlar, Z. 2006. Faunal assemblages and biostratigraphy of several Pliocene sites from Moldova. Courier Forschungsinstitut Senckenberg, 256:249-259.
Nelson, J.S., Grande, T.C., and Wilson, M.V.H. 2016. Fishes of the World (5th Edition). John Wiley & Sons, Inc., Hoboken, New Jersey.
Nordstrom, K. 2011. Tinca tinca. Fish, 423:1-11.
Pană, I. 1977. Problems arisen by the appearance of the new otolith species and their stratigraphical implications, examples of the Dacian otoliths from S. Dobrudja. Analele Universității din București, Geologie, 26:105-124.
Pană, I. 1982. Otholites Dacien supérieur (Parscovien) d’Olténie. Analele Universității din București, Geologie, 31:33-42.
Pană, I. 1995. Otholites daciens, p. 427-445. In Marinescu, F. and Papaianopol, I. (eds.), Chronostratigraphie und Neostratotypen. 9, Pliozän PL1, Dazien. Ed. Academiei Române, Bucharest.
Pană, I. and Müntz, K. 2005. Date noi asupra dacianului de pe Valea Sărățelului (sectorul Valea Verzii), p. 266-272. In Pană, I. (ed.), O viață dedicată profesiei de geolog. Etapa I: 1955-1974. Editura Universitară “Ion Mincu”, Bucharest.
Pawłowska, K. 1963. Ichtiofauna łupkow interglacjalnych (Masovien I) z Barkowic Mokrych koło Sulejowa. Acta Palaeontologica Polonica, 8:475-493.
Popescu, A. 2004. Sur le gisement paléontologique Leu (Mammalia, Pléistocène). Acta Palaeontologica Romaniae, 4:369-372.
Radu, V. 2005. Atlas for the identification of bony fish bones from archaeological sites. Studii de Preistorie, Supplementum 1:1-77.
Rădulescu, C. and Samson, P. 1990. The Plio-Pleistocene mammalian succession of the Olteț Valley, Dacic Basin, Romania. Quartärpaläontologie, 8:225-232.
Rădulescu, C. and Samson, P. 1986. Les mammiferes du Pleistocene inferieur d’Izvoru (department d’Olt, Roumanie). Quartärpaläontologie, 6:157-171.
Rădulescu, C., Samson, P.-M., and Știucă, E. 1989. Pliocene (Lower Romanian) micromammals in the Dacic Basin. Miscellanea Speologica Romanica, 1:313-326.
Rădulescu, C. and Samson, P. 2001. Biochronology and evolution of the Early Pliocene to Early Pleistocene mammalian faunas of Romania. Bollettino della Società Paleontologica Italiana, 40:285-291.
Rădulescu, C., Enciu, P., and Petculescu, A. 1997. On the presence of Prolagurus and Allophaiomys in the Dacic Basin. Travaux de l’Institut de Spéologie “Émile Racovitza”, 36:119-133.
Rădulescu, C., Samson, P.-M., Știucă, E., Enciu, P., and Popescu, A. 1993. New fossil mammal associations of the Jiu Valley. A contribution to the Early-Late Pliocene biostratigraphy of the Dacic Basin, Romania. Travaux de l’Institut de Spéologie “Émile Racovitza”, 32:95-105.
Rădulescu, C., Samson, P.-M., Știucă, E., Petculescu, A., Enciu, P., and Popescu, A. 1999. Micromamiferele romaniene din vestul Bazinului Dacic. Oltenia. Studii și comunicări. Științele Naturii, 15:26-35.
Rădulescu, C., Samson, P.-M., Petculescu, A., and Știucă, E. 2003. Pliocene large mammals of Romania. Coloquios de Paleontología, Volumen Extraordinario, 1:549-558.
Rekovets, L., Čermák, S., Kovalchuk, O., Prisyazhniuk, V., and Nowakowski, D. 2014. Vertebrates from the Middle Pleistocene locality Lysa Gora in Ukraine. Quaternary International, 326-327:481-491. https://doi.org/10.1016/j.quaint.2013.10.016
Rutte, E. 1962. Schundzähne von Süsswasserfischen. Palaeontographica Abteilung, 120:165-212.
Schreve, D.C., Bringland, D.R., Allen, P., Blackford, J.J., Gleed-Owen, C.P., Griffiths, H.I., Keen, D.H., and White, M.J. 2002. Sedimentology, palaeontology and archaeology of late Middle Pleistocene River Thames terrace deposits at Purfleet, Essex, UK. Quaternary Science Reviews, 21:1423-1464. https://doi.org/10.1016/S0277-3791(01)00100-7
Simionescu, I. 1932a. Les vertébrés pliocèns de Berești. Buletinul Societății Române de Geologie, 1:215-228.
Simionescu, I. 1932b. Vertebratele pliocene de la Mălușteni (Covurlui). Publicațiunile Fondului Vasile Adamachi, 9:83-151.
Știucă, E., Petculescu, A., Vasile, Ș., and Tiță, R. 2012. Macro- and micromammal faunas associated with Mammuthus (Archidiskodon) meridionalis in the Lower-Middle Pleistocene from Copăceni (Ilfov County, Romania), p. 76-77. In Murariu, D., Adam, C., Chişamera, G., Iorgu, E., Popa, L.O., Popa, O.P. (eds.), Annual Zoological Congress of “Grigore Antipa” Museum, 21-23 November 2012, Bucharest.
Sytchevskaya, E.K. 1989. Presnovodnaja ihtiofauna neogena Mongolii. Trudy Sovmestnoj Sovetsko-Mongolskoj Paleontologicheskoj Ekspeditsii, 39:1-144. (In Russian)
Tadajewska, M. 1998. Pharyngeal teeth and shape of the ossa pharyngea inferiora during development of Abramis brama and Blicca bjoerkna (Cyprinidae). Cybium, 22:123-147.
Terzea, E. 1981. Remarques sur la biostratigraphie du Pliocène du sud de la Plaine Romaine (Zone de Turnu-Măgurele). Travaux de l’Institut de Spéologie “Émile Racovitza”, 20:113-126.
Terzea, E. 2005. Ansambluri de mamifere pliocene din Bazinul Dacic (România) și semnificația lor ecologică și paleoclimatică. Oltenia. Studii și comunicări. Științele Naturii, 21:9-13.
Terzea, E. and Boroneanț, V. 1979. Découverte d’une faune de Mammifères pliocènes à Ciuperceni (dèp. de Teleorman). Remarques sur deux espèces inconnues en Roumanie. Travaux de l’Institut de Spéologie “Émile Racovitza”, 18:171-184.
Trif, N. and Codrea, V. 2018. Critical overview on the odontological researches of the Mesozoic and Cenozoic fish from Romania. Brukenthal. Acta Musei, 13:497-516.
Trif, N., Vasilyan, D., and Popescu, A. 2016. Fossil fish remains from the Podari, SW of Romania. The first report. Oltenia. Studii şi Comunicări. Ştiinţele Naturii, 32:12-19.
van den Hoek Ostende, L.W., Gardner, J.D., van Bennekom, L., Alçiçek, M.C., Murray, A.M., Wesselingh, F.P., Alçiçek, H., and Tesakov, A. 2015. Ericek, a new Pliocene vertebrate locality in the Çameli Basin (southwestern Anatolia, Turkey). Palaeobiodiversity and Palaeoenvironments, 95:305-320. https://doi.org/10.1007/s12549-015-0202-3
Vasile, Ș., Panaitescu, D., Știucă, E., and Virág, A. 2012. Additional proboscideans fossils from Mavrodin (Teleorman County, Romania). Oltenia. Studii și Comunicări. Științele Naturii, 28:211-218.
Vasile, Ș., Petculescu, A., and Venczel, M. 2015. Taxonomic diversity of the vertebrate assemblages from Copăceni - a new reference site for the Lower Pleistocene of the Dacian Basin, Romania, p. 75. In Jagt, J.W.M., Hebda, G., Mitrus, S., Jagt-Yazykova, E.A., Bodzioch, A., Konietzko-Meier, D., Kardynał, K., Gruntmejer, K. (eds.), European Association of Vertebrate Palaeontologists, XIII Annual Meeting, 8-12 July 2015, Opole, Poland.
Vasile, Ș., Știucă, E., and Panaitescu, D. 2013a. First find of elephantid remains from the Pleistocene of Copăceni (Ilfov County, Romania). Oltenia. Studii și Comunicări. Științele Naturii, 29:42-51.
Vasile, Ș., Știucă, E., and Venczel, M. 2013b. First find of ectothermic vertebrates from the Pleistocene “Copăceni beds” (southern Romania), p. 90-91. In Țabără, D. (ed.), Ninth Romanian Symposium on Paleontology, Iași.
Vasile, Ș., Venczel, M., and Petculescu, A. 2016. New data on the early Pleistocene vertebrate assemblages from Copăceni (Dacian Basin, southern Romania), p. 35. In Holwerda, F., Madern, A., Voeten, D., van Hateren, A., Meijer, A., den Ouden, N. (eds.), XIV Annual Meeting of the European Association of Vertebrate Palaeontologists, 6-10 July 2016, Haarlem, The Netherlands.
Vasile, Ș., Venczel, M., and Petculescu, A. 2018. Early Pleistocene amphibians from Copăceni (Dacian Basin, southern Romania), 154-155. In Popa, L.O., Adam, C., Chişamera, G., Iorgu, E., Popa, L.O., Popa, O.P. (eds.), International Zoological Congress of “Grigore Antipa” Museum, Bucharest.
Vasilyan, D., Schneider, S., Bayraktutan, M.S., and Şen, S. 2014. Early Pleistocene freshwater communities and rodents from the Pasinler Basin (Erzurum Province, north-eastern Turkey). Turkish Journal of Earth Sciences, 23:1-15. https://doi.org/10.3906/yer-1307-16
Voicu, F.-A., Venczel, M., Petculescu, A., and Vasile, Ș. 2015. Taxonomic diversity of early Pleistocene snakes from the Tetoiu Formation (Dacian Basin, Romania) - preliminary results, p. 120-121. In Bucur, I.I., Lazăr, I., Săsăran, E. (eds.), Tenth Romanian Symposium on Paleontology, 16-17 October 2015, Cluj-Napoca.
