

 Approaching the Khersonian Crisis: Fish otoliths from the upper Bessarabian (middle Sarmatian s.l.; Late Miocene) of Jurkine (Kerch Peninsula, Crimea)
Approaching the Khersonian Crisis: Fish otoliths from the upper Bessarabian (middle Sarmatian s.l.; Late Miocene) of Jurkine (Kerch Peninsula, Crimea)
Article number: 26.2.a31
https://doi.org/10.26879/1300
Copyright Paleontological Society, August 2023
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 27 April 2023. Acceptance: 27 July 2023.
ABSTRACT
After its separation from the world ocean, the Paratethys formed a large inland marine water body during the Serravallian and Tortonian (Middle to Late Miocene) with a diverse environmental setting that underwent multiple fast and short-lived ecological changes. The trapped bony fish fauna either adapted to the rapid ecological changes (forced endemism) or perished. A major event during the late Sarmatian s.l. (Khersonian; late Tortonian; Late Miocene) was the Khersonian Crisis in the Eastern Paratethys, which is postulated to have led to the demise of stenohaline marine fishes in the basin. A rich fully marine otolith-based fish fauna has recently been described from the middle Bessarabian (middle Sarmatian s.l.; early Tortonian) of Jurkine, Kerch Peninsula, Crimea. A small otolith-based fauna from a higher part of the Bessarabian in the Jurkine section is described. It is greatly depleted in faunal composition and shares only a single species with the older, stenohaline marine fauna: the clupeid Maeotichthys wilhelmi. A second, new clupeid is described as Maeotichthys salebrosus n. sp. The highly diverse and dominant association of the Gobiidae, a family that has shown to be particularly well positioned to adapt to the rapid ecological changes in the basin, is not present anymore. Instead, the Gobiidae are represented only by a single taxon of the Benthophilini (tadpole gobies): Scythogobius spissus n. gen., n. sp. We interpret this drastic change in the faunal composition of fishes, which is also mirrored in benthic foraminifera, as an indication of spreading suboxic sea bottom conditions in the basin (including the Jurkine section), probably as a harbinger of the approaching Khersonian Crisis.
Werner W. Schwarzhans. Zoological Museum, Natural History Museum of Denmark, Universitetsparken 15, 2100 København, Denmark; and Ahrensburger Weg 103, 22359 Hamburg, Germany. Corresponding author. wwschwarz@aol.com
http://orcid.org/0000-0003-4842-7989
Andriy Bratishko. Faculty of Natural Sciences, Luhansk Taras Shevchenko National University, Koval St. 3, Poltava, Ukraine, 36023 and BugWare, Inc., 1615 Village Square Blvd, Ste. 8, Tallahassee, Florida 32309, U.S.A. andriybratishko@gmail.com
Yuliia V. Vernyhorova. Department of Stratigraphy and Paleontology of Cenozoic Deposits, Institute of Geological Sciences, National Academy of Sciences of Ukraine, O. Honchar Str. 55-b, Kyiv, Ukraine, 01054; and Department of Invertebrate Fauna and Systematics, Schmalhausen Institute of Zoology of NAS of Ukraine, B. Khmelnitsky Str. 15, Kyiv 01054, Ukraine. july.vern@gmail.com
Keywords: new genus; new species; Ukraine; Eastern Paratethys; otoliths, Gobiidae
Final citation: Schwarzhans, Werner W., Bratishko, Andriy, and Vernyhorova, Yuliia V. 2023. Approaching the Khersonian Crisis: Fish otoliths from the upper Bessarabian (middle Sarmatian s.l.; Late Miocene) of Jurkine (Kerch Peninsula, Crimea). Palaeontologia Electronica, 26(2):a31.
https://doi.org/10.26879/1300
palaeo-electronica.org/content/2023/3922-approaching-khersonian-crisis
Copyright: August 2023 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
https://zoobank.org/29D45563-17AC-4364-979D-4F9876CD7D7D
INTRODUCTION
A rich otolith-based fish fauna has recently been described from the middle Bessarabian (basal Tortonian, Late Miocene) of the Jurkine section on the Kerch Peninsula, Crimea, and was discussed in comparison to paleoenvironmental reconstructions based on analysis of the benthic fauna and the lithology (Bratishko et al., 2023). This fish fauna was shown to be the result of a forced endemic evolution in the restricted marine environment of the Eastern Paratethys and was considered to be mainly represented by stenohaline marine fishes. It was postulated that this faunal association might represent the last highly diverse fish community to be expected in the Eastern Paratethys prior to the Khersonian Crisis and that the crisis caused the demise of all stenohaline fishes in the basin and only euryhaline lineages would have survived. The Khersonian, being the latest regional substage of the Sarmatian s.l. in the Eastern Paratethys, was a time of rapid changes of sea level and environments in the basin including both hypersaline and phases with freshwater input (e.g., Belokrys, 1976; Palcu et al., 2021, but see Šujan et al., 2022).
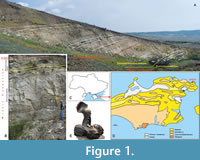 During the evaluation of microsamples collected across the entire outcrop sequence at Jurkine, it was found that the Khersonian strata were devoid of aragonitic fossils, probably due to dissolution caused during diagenesis or by subaerial weathering. However, a few otoliths were found in strata about 16 m below the base of the Khersonian and 7 to 8 m above the interval that yielded the rich otolith-based fish fauna described in Bratishko et al. (2023) (i.e., microfauna samples #20 and #21; Figure 1-Figure 2). The purpose of this article is to describe the new otolith specimens obtained and discuss what they reveal about environmental changes affecting the fish fauna upon approaching the Khersonian substage. A comprehensive analysis of the environmental development is presented for the Sarmatian Sea of the Kerch Peninsula and its adjacent area during the Bessarabian and Khersonian.
During the evaluation of microsamples collected across the entire outcrop sequence at Jurkine, it was found that the Khersonian strata were devoid of aragonitic fossils, probably due to dissolution caused during diagenesis or by subaerial weathering. However, a few otoliths were found in strata about 16 m below the base of the Khersonian and 7 to 8 m above the interval that yielded the rich otolith-based fish fauna described in Bratishko et al. (2023) (i.e., microfauna samples #20 and #21; Figure 1-Figure 2). The purpose of this article is to describe the new otolith specimens obtained and discuss what they reveal about environmental changes affecting the fish fauna upon approaching the Khersonian substage. A comprehensive analysis of the environmental development is presented for the Sarmatian Sea of the Kerch Peninsula and its adjacent area during the Bessarabian and Khersonian.
MATERIAL AND METHODS
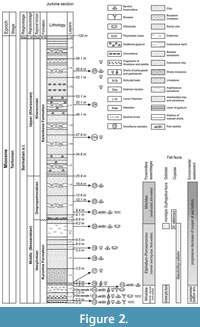 Ten otolith specimens belonging to four species are described in the following. They were obtained from microfauna samples #20 and #21 (Figure 2) at the transition of the Vasylivkian to the Dnipropetrovskian regional horizons in the upper Bessarabian. The bulk samples were processed for foraminifera using a 0.063 mm mesh. The holotypes are registered with the National Museum of Natural History of the National Academy of Sciences of Ukraine (NMNH), Kyiv, in the Department of Monographic Collection under the collection registrations NMNH ГКН (MBA - main book of acquisition) 5960 079 to 080. All other specimens are deposited at the Senckenberg Forschunginstitut und Naturmuseum (SMF), Frankfurt/Main, under the collection registrations SMF PO 101.337 to 340.
Ten otolith specimens belonging to four species are described in the following. They were obtained from microfauna samples #20 and #21 (Figure 2) at the transition of the Vasylivkian to the Dnipropetrovskian regional horizons in the upper Bessarabian. The bulk samples were processed for foraminifera using a 0.063 mm mesh. The holotypes are registered with the National Museum of Natural History of the National Academy of Sciences of Ukraine (NMNH), Kyiv, in the Department of Monographic Collection under the collection registrations NMNH ГКН (MBA - main book of acquisition) 5960 079 to 080. All other specimens are deposited at the Senckenberg Forschunginstitut und Naturmuseum (SMF), Frankfurt/Main, under the collection registrations SMF PO 101.337 to 340.
The otoliths were photographed with a Canon EOS 1000D mounted on a Wild M400 photomacroscope that was remotely controlled from a computer. Individual images of every view of the objects taken at a ranging field of depth were stacked using the Heliconfocus software from Heliconsoft (Kharkiv, Ukraine). Adjustment of exposure and contrast and retouching were completed in Adobe Photoshop when necessary to improve the images without altering any morphological features. All otoliths are shown from the inner face (if not annotated otherwise) of the right side and mirror imaged when necessary.
We used the morphological terminology established by Koken (1884) with amendments by Chaine and Duvergier (1934) and Schwarzhans (1978), and we applied the morphometrics for gobies as established in Schwarzhans (2014). The abbreviations used are as follows: OL, otolith length; OH, otolith height; OT, otolith thickness; OsL, ostium length; CaL, cauda length; SuL, sulcus length; and SL, standard length (in fish).
The foraminifera assemblage was described from the entire section in Bratishko et al. (2023). Here, the paleoenvironments of the middle-late Sarmatian s.l. of the Kerch Peninsula and its adjacent areas are analyzed based on the composition of the fauna (mollusks, foraminifers, and fish otoliths) and certain paleogeographic features from the data base of YV.
SYSTEMATIC PALAEONTOLOGY
(by Schwarzhans and Bratishko)
Division TELEOSTEI Müller, 1846
Order ANGUILLIFORMES Regan, 1909
Family OPHICHTHIDAE Rafinesque, 1815
Genus indet.
Ophichthidae indet.
Figure 3A-D
Material. One specimen, SMF PO 101.337, Jurkine, Kerch Peninsula, Crimea, microfauna sample #20, upper Vasylivkian, Bessarabian, middle Sarmatian s.l.
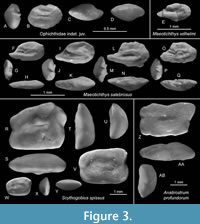 Description. The single specimen is very small (0.5 mm in length), oval in outline, and thick (OL:OH = 1.4; OH:OT = 1.4). All rims are regularly curved without prominent angles, except for the dorsal rim, which shows a small indentation at its middle.
Description. The single specimen is very small (0.5 mm in length), oval in outline, and thick (OL:OH = 1.4; OH:OT = 1.4). All rims are regularly curved without prominent angles, except for the dorsal rim, which shows a small indentation at its middle.
The strongly convex inner face shows a slightly supramedian, short, narrow, somewhat deepened, and anteriorly opened sulcus (OL:SuL = 1.45). The ostium is slightly wider than the very narrow cauda and of about equal length (OsL:CaL = 1.2). The dorsal field shows a distinct depression above the cauda and the posterior part of the ostium; the ventral field is smooth without ventral furrow. The outer face is smooth and very strongly convex except for a slight concavity behind the middle of the otolith.
Discussion. This tiny but nevertheless highly characteristic otolith presumably stems from a juvenile fish. Its allocation to the family Ophichthidae is tentative, and only when larger specimens have been found can a conclusive interpretation be given.
Order CLUPEIFORMES Bleeker, 1859
Family CLUPEIDAE Rafinesque, 1810
Genus MAEOTICHTHYS Bratishko and Schwarzhans, 2023
Maeotichthys wilhelmi (Djafarova, 2006)
Figure 3E
2006 Otolithus (Osmeridarum) wilhelmi - Djafarova, 2006, plate 2, figure 3.
2023 Maeotichthys wilhelmi (Djafarova, 2006) - Bratishko and Schwarzhans (in Bratishko et al., 2023; see there for further references): figure 4J-AA.
Material. Two specimens, SMF PO 101.338, Jurkine, Kerch Peninsula, Crimea, microfauna sample #20, upper Vasylivkian, Bessarabian, middle Sarmatian s.l.
Discussion. Both specimens are fragmented, but one specimen could be reconstructed from the fragments retrieved and is shown digitally assembled in Figure 2E. Maeotichthys wilhelmi differs from the parallel occurring M. salebrosus n. sp. described below in the thinner appearance and the deeper ventral rim. Maeotichthys wilhelmi is widely distributed in the middle Sarmatian s.l. (Bessarabian) of the Eastern Paratethys and is also known from the early Sarmatian s.l. (Volhynian) of Bulgaria (Strashimirov, 1984, as revised in Bratishko et al., 2023).
Maeotichthys salebrosus n. sp.
Figure 3F-Q
zoobank.org/02CD4720-EAEB-4F55-91B3-B8506A333089
?1968 Otolithus (Clupea) fimbriosus - Suzin, 1968, plate 18, figure 4 [name not available according to ICZN article 13.1.1].
Holotype. Figure 3I-K, NMNH ГКН 5960 079, Jurkine, Kerch Peninsula, Crimea, microfauna sample #21, lower Dnipropetrovskian, Bessarabian, middle Sarmatian s.l.
Paratypes. Four specimens, SMF PO 101.339, Jurkine, Kerch Peninsula, Crimea, microfauna sample #20, upper Vasylivkian, Bessarabian, middle Sarmatian s.l.
Name. From salebrosus (Latin) = rugged, referring to the rugged appearance of the otoliths.
Diagnosis. Robust, thick otoliths with long rostrum (18-25% of OL). OL:OH = 1.65-1.8; OH:OT = 1.6-2.0. Ventral rim relatively shallow, irregular, rugged. Inner face flat with marked crista superior and inferior and bulged area behind sulcus.
Description. The otoliths of M. salebrosus are small, reach 1.25 mm in length (holotype 1.15 mm), and are relatively thick and robust. The rostrum is long and variably pointed or rounded; an antirostrum is not clearly developed, and the excisura is wide and not deep, more like a broad concavity. The dorsal rim is low, irregularly curved, and can be smooth (Figure 3F, I) or coarsely undulating (Figure 3L). The ventral rim likewise is irregularly curved and relatively shallow; only rarely is the postventral region somewhat expanded (Figure 3L).
The inner face is almost flat in the horizontal direction but slightly curved in the vertical direction. It bears a moderately long and distinctly deepened sulcus that shows equally long and wide ostium and cauda, which are indistinctly marked by a variable indentation of the dorsal sulcus margin or a deepening of the ostium toward the cauda; OL:SuL = 1.3-1.4; OsL:CaL = 1.3. The crista superior and inferior are well developed, ridge-like (best seen on anterior views in Figure 3J, M, P), and the area behind the caudal tip is more or less strongly bulged. The dorsal depression is distinct and relatively large; a ventral depression or furrow underlies most of the crista inferior. The outer face is variably convex and relatively smooth. The otolith rims are thick, contributing to the overall thick appearance of the otoliths.
Discussion. Maeotichthys salebrosus differs from M. wilhelmi in the more robust, rugged appearance and the relatively shallow ventral rim (vs. ventral rim expanded along the postventral rim and recessed below the rostrum). A specimen figured by Suzin (1968) as Clupea fimbriosus from the middle Sarmatian of the Crimea may represent the same species based on the rather shallow ventral rim and not M. wilhelmi as suggested by Bratishko et al. (2023); nevertheless, the name is not available according to the regulations of the ICZN. Maeotichthys salebrosus occurs in parallel in the late Vasylivkian but not in earlier, restricted marine sediments. We therefore assume that M. salebrosus was probably adapted to a more brackish marine environment, based, however, on a limited number of specimens.
Order GOBIIFORMES Jordan and Evermann, 1896
Family GOBIIDAE Cuvier, 1816
Genus SCYTHOGOBIUS n. gen.
zoobank.org/46A20282-5726-46BB-A6CE-3557EF595087
Type species. Scythogobius spissus n. sp.
Name. Named after the Scyths, early historic inhabitants of the Crimean Peninsula.
Diagnosis. A fossil otolith-based genus of the family Gobiidae with the following combination of characteristics. The otoliths are moderately elongate (OL:OH = 1.25-1.4) and robust with a flat to moderately convex inner face and a strongly convex outer face (OH:OT = 1.7-2.2). The ventral rim of the otolith is nearly straight, and the anterior and posterior rims are almost vertical with minor indentations at about the level of the anterior tip of the sulcus and the posterior tip of the sulcus. The postdorsal projection is short and not bent outward. The sulcus is relatively small, shallow, smoothened sole-shaped, and only slightly inclined. A subcaudal iugum is not developed.
Discussion. Scythogobius is interpreted as a member of the Ponto-Caspian endemic clade Benthophilini. It resembles otoliths of the extant species of Benthophilus, Caspiosoma, and Anatirostrum in the reduced sulcus morphology and the absence of a subcaudal iugum. In all these genera, however, the sulcus morphology is further altered in shape to a nearly oval sulcus outline and a w-shaped ventral sulcus margin. Moreover, these extant otoliths are usually thinner with a less strongly convex outer face, except for Anatirostrum. Anatirostrum is a deepwater endemic monospecific genus in the southern Caspian Sea Basin and probably represents an autogenic endemic speciation. An otolith of the extant A. profundorum (Berg, 1927) is depicted in Figure 3Z-AB. Its otoliths differ from Scythogobius in the deepened sulcus (vs. shallow) and the smaller size of the sulcus (OL:SuL = 2.75 vs. 1.75-2.15). We therefore consider Scythogobius to be unrelated to Anatirostrum and instead to represent an early endemic lineage during the Bessarabian that did not persist to the present-day Ponto-Caspian goby association.
Species. Scythogobius is a monospecific genus with S. spissus from the Late Miocene (Bessarabian) of Crimea.
Scythogobius spissus n. sp.
Figure 3R-Y
zoobank.org/C5B4CFB5-08F9-42AC-ABD3-B92E2605B755
Holotype. Figure 3R-T, NMNH ГКН 5960 080, Jurkine, Kerch Peninsula, Crimea, microfauna sample #20, upper Vasylivkian, Bessarabian, middle Sarmatian s.l.
Paratypes. Two specimens, SMF PO 101.340, same data as holotype.
Name. From spissus (Latin) = compact, thick, referring to the thick appearance of these otoliths.
Diagnosis. See generic diagnosis (monospecific genus).
Description. The otoliths are moderately large, compact with a subrectangular outline up to 3.35 mm in length (holotype). The dorsal rim is low, straight, flat, nearly horizontal, and slightly ascending to an obtuse postdorsal angle. The ventral rim flat and horizontal. The anterior rim is nearly vertical with a small indentation at the level of the tip of the sulcus in large specimens and straight in small ones (Figure 3W). The posterior rim is vertical or slightly inclined outwardly toward a short, obtuse postdorsal projection, which could be an ontogenetic effect. The postdorsal projection is not bent outward. All rims are smooth.
The inner face is flat or moderately convex (compare Figure 3S and 3Y). The sulcus is centrally positioned, shallow, mostly small (probably widened by erosion in Figure 3V); OL:SuL = 1.75-2.15. The sulcus shape is reduced and rounded sole-type with a low, almost leveled ostial lobe. There is no subcaudal iugum. The dorsal depression is variable but mostly shallow and indistinct. There is no ventral furrow discernable. The outer face is strongly convex, smooth.
DISCUSSION
Summary of the Regional Paleoenvironmental Setting in the Sarmatian Sea During the Bessarabian and Khersonian
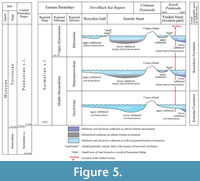
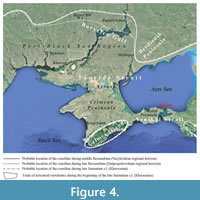 During the Sarmatian s.l., the area of the Kerch Peninsula (Figure 4-Figure 5) constituted the western part of the Yenikal Strait, which was located southeast of the Borysfen Gulf and the Tauride Strait and separated the Crimea Island from the Caucasian Island (according to Kolesnikov, 1940). In this area, the Sarmatian Sea was not deep in the sense of having oceanic depths. It was mainly a shelf sea with a shallow, highly variable coastal zone (upper sublittoral) along the periphery of the basin and a deep shelf zone (lower sublittoral) in the central region (Figure 5) (Kolesnikov, 1940; Molyavko and Pidoplichko, 1955; Belokrys, 1963, 1966, 1976; Didkovskiy, 1964; Maisuradze, 1980). The Borysfen Gulf presumably had an average depth of about 15-20 m, and it probably did not exceed 40-50 m at its deepest part (e.g., Molyavko and Pidoplichko, 1955; Belokrys, 1963, 1966). Only the Black Sea area of the Pontian Basin had greater depths during the Sarmatian s.l. (Kolesnikov, 1940; Popov et al., 2004).
During the Sarmatian s.l., the area of the Kerch Peninsula (Figure 4-Figure 5) constituted the western part of the Yenikal Strait, which was located southeast of the Borysfen Gulf and the Tauride Strait and separated the Crimea Island from the Caucasian Island (according to Kolesnikov, 1940). In this area, the Sarmatian Sea was not deep in the sense of having oceanic depths. It was mainly a shelf sea with a shallow, highly variable coastal zone (upper sublittoral) along the periphery of the basin and a deep shelf zone (lower sublittoral) in the central region (Figure 5) (Kolesnikov, 1940; Molyavko and Pidoplichko, 1955; Belokrys, 1963, 1966, 1976; Didkovskiy, 1964; Maisuradze, 1980). The Borysfen Gulf presumably had an average depth of about 15-20 m, and it probably did not exceed 40-50 m at its deepest part (e.g., Molyavko and Pidoplichko, 1955; Belokrys, 1963, 1966). Only the Black Sea area of the Pontian Basin had greater depths during the Sarmatian s.l. (Kolesnikov, 1940; Popov et al., 2004).
 During the middle of the Bessarabian (Vasylivkian regional horizon), there was a wide middle shelf zone with a diverse and rich benthic fauna typical for the middle Sarmatian s.l., with, for example, common species of Mactra, Obsoletiforma, Plicatiforma, and Barbotella among mollusks and Meandroloculina, Quinqueloculina, Triloculina, Flintina, Elphidium, and Porosononion among foraminifera (Kolesnikov, 1940; Didkovskiy, 1964; Belokrys, 1963, 1976; Maisuradze, 1980; Nevesskaya et al., 1986; Nevesskaya and Sokolov, 1986; Paramonova, 1994; Popov et al., 2004; Maisuradze and Koiava, 2011; Popov et al., 2019). This faunal association lived along the periphery of the Borysfen Gulf in the Tauride Strait, and also in the western part of the Yenikal Strait (i.e., on the western terrain of the Kerch Peninsula) (Figure 4-Figure 5) (Kolesnikov, 1940; Molyavko and Pidoplichko, 1955; Didkovskiy, 1964; Belokrys, 1966, 1976; Didkovskiy and Kulichenko, 1975; Bratishko et al., 2023). At the same time in the eastern part of the Kerch Peninsula and on the Taman Peninsula (eastern part of the Yenikal Strait) (Figure 4-Figure 5), the basin was deeper (lower sublittoral) with a periodically increasing oxygen deficiency near the sea bottom, which is reflected in the composition of the fauna (e.g., Arkhangelsky et al., 1930; Kolesnikov,1940; Nevesskaya and Sokolov, 1986; Popov et al., 2016).
During the middle of the Bessarabian (Vasylivkian regional horizon), there was a wide middle shelf zone with a diverse and rich benthic fauna typical for the middle Sarmatian s.l., with, for example, common species of Mactra, Obsoletiforma, Plicatiforma, and Barbotella among mollusks and Meandroloculina, Quinqueloculina, Triloculina, Flintina, Elphidium, and Porosononion among foraminifera (Kolesnikov, 1940; Didkovskiy, 1964; Belokrys, 1963, 1976; Maisuradze, 1980; Nevesskaya et al., 1986; Nevesskaya and Sokolov, 1986; Paramonova, 1994; Popov et al., 2004; Maisuradze and Koiava, 2011; Popov et al., 2019). This faunal association lived along the periphery of the Borysfen Gulf in the Tauride Strait, and also in the western part of the Yenikal Strait (i.e., on the western terrain of the Kerch Peninsula) (Figure 4-Figure 5) (Kolesnikov, 1940; Molyavko and Pidoplichko, 1955; Didkovskiy, 1964; Belokrys, 1966, 1976; Didkovskiy and Kulichenko, 1975; Bratishko et al., 2023). At the same time in the eastern part of the Kerch Peninsula and on the Taman Peninsula (eastern part of the Yenikal Strait) (Figure 4-Figure 5), the basin was deeper (lower sublittoral) with a periodically increasing oxygen deficiency near the sea bottom, which is reflected in the composition of the fauna (e.g., Arkhangelsky et al., 1930; Kolesnikov,1940; Nevesskaya and Sokolov, 1986; Popov et al., 2016).
Toward the end of the Bessarabian (Dnipropetrovskian regional horizon), the coastal, upper sublittoral zone was also located along the periphery of the basin but was subject to fluctuating coast lines and riverine discharge (e.g., the northern margin of the Peri-Black Sea Region, the Molochna River Basin; see figure 8 in Belokrys, 1976) (Figure 5). It is characterized by a rich mollusk fauna: for example, a large number of mollusks with spiny shells such as Plicatiforma fittoni fittoni (d’Orbigny, 1845), Obsoletiforma obliquoobsoleta (Kolesnikov, 1929), sharp-keeled morpho-types such as Mactra (Sarmatimactra) fabreana (d’Orbigni, 1844), and the appearance of Barbotella intermedia (Radovanović and Pavlović, 1893), Gibbula af. minuta (Sinzow, 1880), Calliostoma cordieriana (Orbigny, 1844) (see Belokrys, 1963, 1966, 1976). However, the mollusk fauna is already sharply depleted at a distance of 30-40 km from the coast (Figure 5) and is represented almost entirely by small Mactra (Sarmatimactra) podolica Eichwald, 1930, Obsoletiforma obsoleta nefanda (Kolesnikov, 1935), and Inaequicostata suessi (Barbot de Marny, 1869). The deposits of these areas are dolomitized (see Belokrys, 1963, 1966, 1976; Vernyhorova 2014, 2016, and references therein). Foraminifera near the coast and further out into the basin were represented by few tests of certain species: Quinqueloculina consobrina var. sarmatica (Gerke and Isaeva, 1952), Q. carinata (d’Orbigny, 1850), Porosononion subgranosus (Egger, 1857), Elphidium macellum (Fichtel and Moll, 1798), E. aculeatum (d’Orbigny, 1846) (Didkovskiy, 1964; Didkovskiy and Kulichenko, 1975; Maisuradze, 1980; Nevesskaya and Sokolov, 1986; Maisuradze and Koiava, 2011). In contrast to the northern Crimean Peninsula and Syvash area (Figure 4), the Yenikal Strait on the Kerch Peninsula (Figure 4-Figure 5) accumulated calcareous non-dolomitized clays, and assemblages of small mollusks and small foraminifers were highly depleted in diversity (see Belokrys, 1976; Vernyhorova 2014, 2016, and references therein; Bratishko et al., 2023). The basin in the northern part of the Kerch Peninsula, including the area around Jurkine, became deepened. As a result, the low-oxygenated deeper-water environment with depleted foraminiferal fauna also encroached into the Jurkine region during the Dinpropetrovskian (Figure 5) (Vernyhorova et al., 2012; Vernyhorova, 2014; Bratishko et al., 2023). Small areas of land were formed as a result of folding and emergence from the sea (Bratishko et al., 2023).
At the beginning of the Khersonian, the Borysfen Gulf, the Tauride Strait, and the western part of the Yenikal Strait had the same environmental conditions as at the late Bessarabian (Dnipropetrovskian regional horizon) (Figure 5). Dolomitic clays were deposited in the coastal shallow zone in the north and east and the intermediate Syvash zone and calcareous non-dolomitic clays along the Yenikal Strait (on the Kerch Peninsula) (see Molyavko and Pidoplichko, 1955; Didkovskiy and Kulichenko, 1975; Belokrys, 1976). A sharp depletion of the faunal diversity is observed everywhere. Mollusks are represented by only a few species like Mactra (Chersonimactra) caspia (Eichwald, 1841), and M. (Chersonimactra) bulgarica (Toula, 1892) (Didkovskiy and Kulichenko, 1975; Vernyhorova 2014, 2016 and references therein). The coastline had a complex structure and was characterized by numerous bays, estuaries, peninsulas, and islands (see “small lands” in Figure 4), which sometimes connected into elongated sandy spits on which terrestrial vertebrates could exist (Molyavko and Pidoplichko, 1955). Thus, in the Lower Dnipro region (near the cities of Berislav and Tyaginka in the Kherson region and Yuryevka city in the Mykolaiv region) (Figure 4), deposits at the beginning of the Khersonian are composed of clays, marls, sands, and conglomerates of up to 2 m thickness and contain remains of terrestrial vertebrates (horses, proboscides, rhinoceroses, giraffes, gazelles, etc.) (Molyavko and Pidoplichko, 1955; Molyavko, 1960; Didkovskiy and Kulichenko, 1975; Nevesskaya and Sokolov, 1986). Foraminifera are almost absent, except occasionally a small number of representatives of Ammonia, Elphidium, Porosononion, Entosolenia, and Bolivina (Didkovskiy, 1964; Bogdanowich, 1965; Didkovskiy and Kulichenko, 1975; Maisuradze, 1980; Nevesskaya and Sokolov, 1986; Maisuradze and Koiava, 2011; Bratishko et al., 2023).
Faunal Evaluation
The otoliths presented here from the higher part of the Bessarabian sequence (microfauna samples #20 and #21) in the Jurkine section are composed of 10 specimens and represent only four species. The faunal composition, however, differs drastically from the rich and diverse restricted marine fish fauna observed in the earlier Bessarabian sequence (Vasylivkian; interval with microfauna samples #13 to #15) from Jurkine as described in Bratishko et al. (2023). The main difference is that the rich gobiid association is gone, and instead, a single taxon (Scythogobius spissus) represents the Gobiidae. Furthermore, no representatives of the rich and diverse stenohaline endemic fish fauna have been found. Finally, the Clupeidae, the other main faunal component in this small assemblage, contains the only persistent species from the lower sequence and, in addition, a new species not present in the lower part of the section.
A comparable observation has been made with the change of the composition in benthic foraminifera. The middle Bessarabian (Vasylivkian) in the Jurkine section is rich and diverse in foraminifera at the same levels where we observed a diverse otolith-based fish fauna (Bratishko et al., 2023). This foraminiferal assemblage is also characterized by a maximal diversity in endemism with Miliolidae dominating (Bratishko et al., 2023). However, a drastic change in foraminiferal diversity and composition occurs shortly above that level toward the end of the Bessarabian (microfauna samples #16 to #20, the latter of which, and sample #21, provided the otoliths described here). Several gradations of changes in the foraminiferal diversity are observed. The Miliolidae epifaunal assemblage (e.g., Quinqueloculina, Dogielina, Meandroloculina, and Sarmatiella genera) have large, thick-walled tests (oxic indicators) and were thriving in the earlier part of the Vasylivkian (lower part of the Jurkine section: microfauna samples #12-14; see Bratishko et al., 2023). Together with the species-rich demersal fish assemblage, the miliolids required a good oxygenation of the water for their living habitat (after Kaiho, 1994; Murray, 2006). Microfauna sample #15 represents a transitional stage with reduced miliolid contents and the beginning of the predominance of the elphidiid assemblage but still the same fish association. The miliolid-dominated foraminiferal assemblage abruptly terminated in the upper part of the Vasylivkian (microfauna samples #16-20), giving way to a benthic foraminifer association dominated by species of the genera Elphidium and Porosononion with large, thick-walled tests (see Bratishko et al., 2023). This change in the foraminiferal association may indicate a shallowing of the basin toward the end of the Bessarabian (e.g., Bogdanovich, 1964; Maisuradze, 1980) and a slight decrease in oxygen content at the sea bottom since it contains some infaunal foraminifera species that are indicators for suboxic B conditions (according to Kaiho, 1994, 1999; Murray, 2006). Higher in the section, in the Dnipropetrovskian (microfauna samples #21-23), another sharp change occurred in the foraminifer composition (Bratishko et al., 2023): a re-appearance of the Miliolidae assemblage with prevalence of Quinqueloculina consobrina var. nitens (Reuss in Bogdanowicz, 1952). Q. angustioris (Bogdanowicz, 1952), Sarmatiella ex gr. subtilis Bogdanowicz, 1952, which were found together with certain Elphidium and Porosononion species: Elphidium reginum (d’Orbigny, 1846), E. macellum (Fichtel et Moll, 1798), Elphidium sp.1, and Porosononion subgranosus subgranosus Egger in Bogdanowich (1960). However, in contrast to the Miliolidae assemblage of the Vasylivkian time, this assemblage is characterized by the small size of tests and low abundance in the samples. Such a benthic foraminiferal composition indicates oxygen deficiency (suboxic A foraminifera indicators) at the bottom water (after Kaiho, 1994; Murray, 2006) during the Dnipropetrovskian time. Overall, by the end of the Bessarabian, the environmental conditions for the benthic fauna deteriorated toward oxygen deficiency in the bottom waters over most of the Kerch Peninsula.
The otolith-based ichthyofauna shows a similar abrupt change in composition to that of mollusks and foraminifers discussed above, and the change in composition is therefore likely to be related to the same changes in environment. Relatively few otoliths have been retrieved from microfauna samples #20 and #21, and hence no quantitative assessment can be made of the changes of the fish fauna. Nevertheless, certain qualitative observations can be made based on the changes in the composition of the families Gobiidae and Clupeidae albeit on a small number of specimens of these families in the upper Bessarabian.
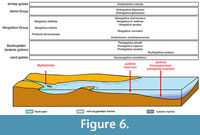
The Gobiidae are the main demersal group of fishes in the Sarmatian Sea. They are remarkable for their rapid adaptation to the changing environments in the Paratethys until today and in fact benefited from reduced competition from other teleost groups that could not adapt and became extinct in the basin (Bratishko et al., 2023). The otoliths obtained from the lower section at Jukine equivalent to microfauna samples #12 to #15 contained 12 gobiid species accounting for about 70% of the entire fauna (Bratishko et al., 2023). None of these species have been identified in microfauna samples #20 or #21. Instead, the only gobiid found in these samples was Scythogobius spissus belonging to a lineage within Benthophilini not present in the lower levels. We interpret this drastic change in gobiid composition as a reaction to the deterioration of oxygen levels near the sea bottom. In such a scenario, Scythogobius spissus would have been a species adapted to a deeper environment with suboxic conditions at the sea bottom, similar perhaps to the extant Anatirostrum profundorum that lives in the deep South Caspian Sea. The suboxic sea bottom environment was apparently not suitable for the many other gobiid species found in the lower Jurkine section characterized by a shallower position in the Sarmatian Sea and with a well-oxygenated bottom environment. It is possible that a similar situation existed laterally and contemporaneously to the lower Jurkine level in the deeper part of the Yenikal Strait further to the east (Figure 4-Figure 5). Otoliths have been described previously also from a location near Mykhailivka (Bratishko et al., 2017), which corresponds to a transitional freshwater-brackish-marine position during the Dnipropetrovskian. This assemblage was also dominated by otoliths of the Gobiidae, but again with a different composition of species. The emerging picture of the endemic gobiid evolution during the Bessarabian is that of a specific adaptation to the different environments in the Sarmatian Sea (Figure 6). Specific adaptations and endemisms occurred in a transitional freshwater-brackish water environment, in the well-oxygenated shallow marine environment and in a deeper suboxic environment (Figure 6).
 The Clupeidae are more common in microfauna samples #20 and #21 than in the lower part of the Jurkine section and include a new species, Maeotichthys salebrosus, along with M. wilhelmi that is also known from the lower section. Clupeids are epipelagic fishes and are therefore less responsive to changes in oxygen level near the sea bottom. Ponto-Caspian clupeids have also shown to be tolerant to changes in salinity levels and are one of two fish families that have survived and adapted through forced endemic evolution in the Ponto-Caspian Basin. We therefore interpret the occurrence of a new species of Maeotichthys in the Dnipropetrovskian in the Jurkine section as a stratigraphic signal rather than an ecological one.
The Clupeidae are more common in microfauna samples #20 and #21 than in the lower part of the Jurkine section and include a new species, Maeotichthys salebrosus, along with M. wilhelmi that is also known from the lower section. Clupeids are epipelagic fishes and are therefore less responsive to changes in oxygen level near the sea bottom. Ponto-Caspian clupeids have also shown to be tolerant to changes in salinity levels and are one of two fish families that have survived and adapted through forced endemic evolution in the Ponto-Caspian Basin. We therefore interpret the occurrence of a new species of Maeotichthys in the Dnipropetrovskian in the Jurkine section as a stratigraphic signal rather than an ecological one.
Microfauna samples #20 and #21 are already close to the onset of the Khersonian Crisis. The Khersonian Crisis (late Sarmatian s.l.) is a time interval in the Eastern Paratethys characterized by rapidly changing macro-environments and sea level changes. Hypersaline conditions alternated with freshening of the water in the basin. The extent of the sea level changes and the environmental implications are currently under debate (compare Popov et al., 2019; Palcu et al., 2021; Šujan et al., 2022), but the effects on the biota trapped in the Eastern Paratethys were dramatic. Bratishko et al. (2023) postulated that during the Late Sarmatian Extinction Event (LSEE) near the base of the Khersonian, all stenohaline fishes became extinct in the Eastern Paratethys and only euryhaline lineages survived. In the case of the Gobiidae, it is mostly the lineages of euryhaline gobies observed at Mykhailivka that seemed to have continued, while the stenohaline goby lineages of the lower Jurkine section (Vasylivkian) apparently terminated with the LSEE. The Clupeidae are epipelagic fishes that have the capability to adapt to brackish water environments, as demonstrated by their endemism during the earlier Karaganian Crisis (Baykina and Schwarzhans, 2017) and their thriving in today’s Caspian Sea. The Ophichthidae are mostly marine fishes, but a few brackish and freshwater lineages exist as well (Froese and Pauly, 2023).
We therefore interpret the observed faunal change and depletion of diversity as having been related to environmental changes from a well-oxygenated to a suboxic environment. Perhaps these changes already foreshadow effects related to the beginning of the Khersonian Crisis. The establishment and broadening of suboxic sea bottom conditions would point to reduced circulation in the water column, which in turn could have been caused by emplacement of a freshwater surface water lens from a nearer or more intense riverine discharge into this region of the basin. Such a development would have suppressed suitable environments for stenohaline marine fishes and favored the spread of euryhaline lineages of the gobies as well as gobies adapted to suboxic conditions and clupeid lineages tolerant to varying epipelagic environments.
CONCLUSION
The small assemblage of otoliths obtained from microfauna samples #20 and #21 above the previously studied rich assemblage described in Bratishko et al. (2023) shows a dramatic shift in composition that is most likely due to changes in the ecological environment - for instance, toward suboxic bottom water conditions upon the approach to the Khersonian Crisis. This study also shows the drastic changes in fish faunal composition that occurred in the enclosed Paratethys across a short time range. The faunal differences may also represent an expression of the ecological diversity of the forced endemic evolution that took place in the Eastern Paratethys after the separation from the world ocean in the Middle Miocene (Serravallian).
The rapid change of the fish community observed in the Jurkine section may also serve as an indication of the kind of faunal changes to be expected once further Neogene sediments of the Eastern Paratethys become available for otolith extraction and study. The unraveling of the complex fish evolution in the Eastern Paratethys is still in the early stages. We sincerely hope that the currently difficult accessibility of prospective localities will improve in the future and allow for further sampling and research.
ACKNOWLEDGEMENTS
The authors a thankful to the following colleagues and institutions for allowance to extract otoliths of extant museum voucher specimens of the Gobiidae for comparison purposes: S. Tessier (CMNFI, Ottawa), F. Krupp (SMF, Frankfurt/Main), E. Vasilieva (ZMMGU, Moscow), J. Nielsen, and P. Møller (ZMUC, Copenhagen).
REFERENCES
Arkhangelsky, A.D., Blokhin, A.A., Menner, V.V., Osipov, S.S., Sokolov, M.I., and Chepikov, K.R. 1930. Kratkiy ocherk geologicheskogo stroeniia i neftianykh mestorozhdeniy Kerchenskogo poluostrova [A short review of the geological structure and oil deposits of the Kerch Peninsula]. Transactions of the Geological and Prospecting Service of the USSR, 13:1-146. (In Russian)
Baykina, E.M. and Schwarzhans, W.W. 2017. Review of “ Clupea humilis ” from the Sarmatian of Moldova and description of Moldavichthys switshenskae gen. et sp. nov. Swiss Journal of Palaeontology, 136:141-152.
https://doi.org/10.1007/s13358-016-0121-6
Belokrys, L.S. 1963. Stratigraphiya i fauna molliuskov sarmatskikh otlozheniy Borisfenskogo zaliva [Stratigraphy and Mollusk Fauna of the Sarmatian Deposits of the Borysphen Bay]. Summary of the PhD thesis in Geological and Mineralogical Sciences. Institute of Geological Sciences of the Academy of Sciences of UkrSSR, Kyiv. (In Russian)
Belokrys, L.S. 1966. K biostratigrafii nizhnesarmatskikh otlozheniy Borisfenskogo zaliva [Biostratigraphy of the lower Sarmatian deposits of Borysphen Bay]. Proceedings of Kyiv University, Series Geology and Exploration, 2:46-68. (In Russian)
Belokrys, L.S. 1976. Sarmat yuga USSR [The Sarmatian of the South of the Ukrainian SSR], p. 3-21. In Barg, I.M. and Nosovskiy, M.F. (eds.), Stratigraphy of the Northern Black Sea Region and the Crimea. Dnepropetrovsk. (In Russian)
Bogdanowich, A.K. 1965. Stratigraphicheskoye i fatsialnoye raspredeleniye foraminifer v miocene Zapadnogo Predkavkaziya i voprosy ikh genezisa [The stratigraphic and facies distribution of foraminifers in the Miocene of the Western Ciscaucasia and questions of their genesis]. Trudy KFVNII, 19:300-350. (In Russian)
Bratishko, A., Kovalchuk, O., and Schwarzhans, W. 2017. Bessarabian (Tortonian, late Miocene) fish otoliths from a transitional freshwater-brackish environment of Mykhailivka, Southern Ukraine. Palaeontologia Electronica, 20.3.44A:1-13.
https://doi.org/10.26879/769
Bratishko, A., Schwarzhans, W., and Vernyhorova, Y. 2023. The endemic marine fish fauna from the Eastern Paratethys reconstructed from otoliths from the Miocene (middle Sarmatian s.l.; Bessarabian) of Jurkine (Kerch Peninsula, Crimea). Rivista Italiana di Paleontologia e Stratigrafia, 129:111-184.
https://doi.org/10.54103/2039-4942/18877
Chaine, J. and Duvergier, J. 1934. Recherches sur les otolithes des poissons. Etude descriptive et comparative de la sagitta des Teléostéens. Actes de la Societé Linnéenne de Bordeaux, 86:5-256.
Didkovskiy, D.Ya. 1964. Biostratigrafiia neogenovyh otlozheniy yuga Russkoi platformy po faune foraminifer [Biostratigraphy of the Neogene deposits in the south of the Russian Platform based on the foraminifera fauna]. Summary of DSc thesis in Geological and Mineralogical Sciences. Institute of Geological Sciences of the Academy of Sciences of UkrSSR, Kyiv. (In Russian)
Didkovskiy, V.Ya. and Kulichenko, V.G. (eds.). 1975. Stratigrafiya USSR. Tom 10. Neogene [Stratigraphy of UkrSSR. Vol. 10. Neogene]. Naukova dumka, Kyiv. (In Russian)
Djafarova, J.D. 2006. Otolity neogena Azerbaidjana [Neogene otoliths of Azerbaijan]. Nafta, Baku. (In Russian)
Froese, R. and Pauly, D. 2021. FishBase. World Wide Web electronic publication, accessed December 2022.
https://www.fishbase.se/search.php
Kaiho, K. 1994. Benthic foraminiferal dissolved-oxygen index and dissolved-oxygen levels in the modern ocean. Geology, 22:719-722.
https://doi.org/10.1130/0091-7613(1994)022<0719:BFDOIA>2.3.CO;2
Kaiho, K. 1999. Effect of organic carbon flux and dissolved oxygen on the benthic foraminiferal oxygen index (BFOI). Marine Micropaleontology, 37:67-76.
https://doi.org/10.1016/S0377-8398(99)00008-0
Koken, E. 1884. Über Fisch-Otolithen, insbesondere über diejenigen der norddeutschen Oligocän-Ablagerungen. Zeitschrift der Deutschen Geologischen Gesellschaft, 36:500-565.
Kolesnikov, V.P. 1940. Verkhniy miocen [Upper Miocene], p. 1-687. In: Neogen SSSR. Stratigrafiya SSSR [Neogene of the USSR. Stratigraphy of the USSR], vol. 12. Academy of Sciences of the USSR, Moscow-Leningrad. (In Russian)
Maisuradze, L.S. 1980. K paleobionomicheskoi istorii pozdnego miocena Chernomorsko-Kaspiiskogo basseina [On the palaeobionomic history of the Late Miocene of the Black Sea-Caspian Basin]. Metsniereba, Tbilisi. (In Russian)
Maisuradze, L. and Koiava, K. 2011. Biodiversity of Sarmatian Foraminifera of the Eastern Paratethys. Bulletin of the Georgian National Academy of Sciences, 5:143-151.
Molyavko, G.I. 1960. Neogen pivdnya Ukrainy [Neogene of southern Ukraine]. Academy of Sciences of the UkrSSR, Kyiv. (In Ukrainian)
Molyavko, G.I. and Pidoplichko, I.G. 1955. Do paleogeografii prychoronomorskykh stepiv pivdnia URSR v neogeni i antropogeni [Palaeogeography of the steppes of the Northern Black Sea region in the south of the UkrSSR during the Neogene and Anthropogene]. Geological Journal, 15:9-25. (In Ukrainian)
Murray, J.W. 2006. Ecology and Applications of Benthic Foraminifera. Cambridge University Press, UK.
https://doi. org/10.1017/CBO9780511535529
Nevesskaya, L.A., Goncharova, L.A., Ilyina, l.B., Paramonova, N.P., Popov, S.V., Babak, E.V., Bagdasarjan, K.G., and Voronina, A.A. 1986. Istoriya neogenovykh molliuskov [The history of Neogene molluscs]. Trudy Paleontologicheskogo Instituta Akademii Nauk SSSR, 220:1-120. (In Russian)
Nevesskaya, L.A. and Sokolov, B.S. (eds.). 1986. Stratigrafiya SSSR. Neogenovaya sistema [Stratigraphy of the USSR. Neogene system]. Nedra, Moscow. (In Russian)
Palcu, D.V., Patina, I.S., Şandric, I., Lazarev, S., Vasiliev, I., Stoica, M., and Krijgsman, W. 2021. Late Miocene megalake regressions in Eurasia. Scientific Reports, 11:11471.
https://doi.org/10.1038/s41598-021-91001-z
Paramonova, N.P. 1994. Istoriya sarmatskikh i akchagylskikh dvustvorchatykh mollkuskov [History of Sarmatian and Akchagylian bivalves). Transactions of the Palaeontological Institute, 260:1-212. (In Russian)
Popov, S.V., Rögl, F., Rozanov, A.Y., Steininger, F.F., Shcherba, I.G., and Kovać, M. (eds.). 2004. Lithological-paleogeographic maps of Paratethys. 10 maps late Eocene to Pliocene. Courier Forschungs-Institut Senckenberg, 250:1-46.
Popov, S.V., Rostovtseva, Yu.V., Fillippova, N.Yu., Golovina, L.A., Radionova, E.P., Goncharova, I.A., Vernyhorova, Yu.V., Dykan, N.I., Pinchuk, T.N., Iljina, L.B., Koromyslova, A.V., Kozyrenko, T.M., Nikolaeva, I.A., and Viskova, L.A. 2016. Paleontology and stratigraphy of the Middle - Upper Miocene of Taman Peninsula. Part 1. Description of key-sections and benthic fossil groups. Paleontological Journal, 50:1-168.
https://doi.org/:10.1134/S0031030116100014
Popov, S.V., Rostovtseva, Y.V., Pinchuk, T.N., and Patina, I.S. 2019. Oligocene to Neogene paleogeography and depositional environments of the Euxinian part of Paratethys in Crimean-Caucasian junction. Marine and Petroleum Geology, 103:163-175.
https://doi.org/10.1016/j.marpetgeo.2019.02.019
Schwarzhans, W. 1978. Otolith-morphology and its usage for higher systematical units, with special reference to the Myctophiformes s.l. Mededelingen Werkgroep Tertiaire Kwartaire Geologie, 15:167-185.
Schwarzhans, W. 2014. Otoliths from the middle Miocene (Serravallian) of the Karaman Basin, Turkey. Cainozoic Research, 14:35-69.
Strashimirov, B. 1984. Otolity ot dolnia sarmat na Severnaia Bulgaria. Palaeontology, Stratigraphy and Lithology, 20:15-41. (In Bulgarian)
Šujan, M., Hudáčková, N., and Magyar, I. 2022. How to drain a megalake: Comments on a study by Palcu et al. (2021). Scientific Reports, 11:11471. https://doi.org/10.31223/X5865V (preprint).
Suzin, A.V. 1968. Otoliths, p. 74-77. In Zhizhchenko, B.P. (ed.), Micropaleontologicheskiye metody stratigraficheskikh postroeniy v neftegazonosnykh oblastiah [Micropalaeontological methods of stratigraphic research in oil-and-gas bearing regions]. Nedra, Moscow. (In Russian)
Vernyhorova, Y.V. 2014. Lito- i biofatsialni osoblyvosti neogenovykh vidkladiv Kerchenskogo pivostrova [Litho-and biofacies features of the Neogene deposits of the Kerch Peninsula]. Collection of scientic works of the Institute of Geological Sciences NAS of Ukraine, 7:126-171. (In Ukrainian)
Vernyhorova, Y.V. 2016. Stratygrafichna skhema neogenovykh vidkladiv Krymskogo pivostrova [Stratigraphic scheme for the Neogene deposits of the Crimean Peninsula]. Stratigraphy and Paleontology. Heolohiia ta Rudonosnist Ukrainy, 2:59-106. (In Ukrainian)
Vernyhorova, Y.V., Fikolina, L.A., and Obsharskaya, N.N. 2012. Strukturno-fatsialnoye rayonirovaniye neogenovykh otlozheniy Kerchenskogo polyostrova [Structural and facies zonation of the Neogene deposits of the Kerch Peninsula]. Geological Journal, 3:74-94. (In Russian)
