

 Moderate extinctions and slow recovery of non-marine teleost fishes across the Cretaceous–Paleogene boundary, with a systematic appraisal of early Paleocene teleost fishes from Saskatchewan, Canada and Montana, USA
Moderate extinctions and slow recovery of non-marine teleost fishes across the Cretaceous–Paleogene boundary, with a systematic appraisal of early Paleocene teleost fishes from Saskatchewan, Canada and Montana, USA
Article number: 28.2.a28
https://doi.org/10.26879/1559
Copyright Society of Vertebrate Paleontology, July 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 7 April 2025. Acceptance: 23 June 2025.
ABSTRACT
Teleost fishes from the lowermost Paleocene Tullock Member of the Fort Union Formation of Garfield County, Montana, USA, and the Ravenscrag Formation of southern Saskatchewan, Canada, are described and compared with assemblages from the latest Cretaceous Hell Creek Formation of Garfield County, Montana, to analyze the faunal dynamics of freshwater teleost fishes during the Cretaceous-Paleogene mass extinction event. A minimum of 18 early Paleocene operational taxonomic units (OTUs) are recognized with 10 of these being present in the earliest Paleocene (Puercan 1 North American Land Mammal “age”) localities and eight being range-through taxa that are inferred to be present based on their presence in lower and higher horizons. An estimated 27% of the OTUs present during the end of the Cretaceous of the northern Western Interior of North America did not extend into the earliest Paleocene. Further extinctions occurred in successive subintervals of the early Paleocene, resulting in a total loss of 56% of latest Cretaceous taxa by the last subinterval of the early Paleocene (Puercan, Pu3). Rather than a recovery to the levels of richness seen during the latest Cretaceous, a stepwise pattern of decline in taxonomic richness occurred throughout the early Paleocene. Comparison of the early Paleocene assemblages with those of the late Paleocene assemblages from Alberta and Saskatchewan, Canada, indicates that taxonomic richness remained low through the Paleocene. This pattern differs from the one seen in mammals, for which taxonomic richness exceeded latest Cretaceous levels by the Pu3.
Donald B. Brinkman. Royal Tyrrell Museum of Palaeontology, Box 7500, Drumheller, AB, Canada, T0J 0Y0; and Department of Biological Sciences, University of Alberta, Edmonton, Alberta, Canada (corresponding author). don.brinkman@gov.ab.ca
Julien D. Divay. Royal Tyrrell Museum of Palaeontology, Box 7500, Drumheller, AB, Canada, T0J 0Y0. jddivay@gmail.com
David G. DeMar, Jr. Department of Paleontology, Burke Museum of Natural History, and Culture, University of Washington, Seattle, Washington 98195, USA; and Department of Paleobiology, National Museum of Natural History, Smithsonian Institution, Washington, DC, 20560, USA. ddemar@uw.edu
Gregory P. Wilson Mantilla. Department of Paleontology, Burke Museum of Natural History, and Culture, University of Washington, Seattle, Washington 98195, USA; and Department of Biology, University of Washington; Seattle, Washington, 98195, USA. gpwilson@uw.edu
Keywords: Tullock; Ravenscrag; Paleocene; Cretaceous-Paleogene mass extinction; Esox; Percomorpha
Final citation: Brinkman, Donald B., Divay, Julien D., DeMar, David G. Jr., and Wilson Mantilla, Gregory P. 2025. Moderate extinctions and slow recovery of non-marine teleost fishes across the Cretaceous–Paleogene boundary, with a systematic appraisal of early Paleocene teleost fishes from Saskatchewan, Canada and Montana, USA. Palaeontologia Electronica, 28(2):a28.
https://doi.org/10.26879/1559
palaeo-electronica.org/content/2025/5566-teleost-fishes-across-the-cretaceous-paleogene-boundary
Copyright: July 2025 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Few mass extinction events have attracted as much attention from the public and scientific community as that marking the end of the Cretaceous. Study of the pattern of extinctions across the Cretaceous-Paleogene (K-Pg) boundary and the subsequent biotic recovery during the Paleocene has been central to the development of our understanding of mass extinctions and their impact on the evolutionary history of paleocommunities (e.g., Twitchett, 2006; Brusatte et al., 2015). Our understanding of the pattern of changes among non-marine paleocommunities during this mass extinction has, in large part, resulted from studies of the fossil record from the Hell Creek region in northeastern Montana, USA (Archibald, 1996, 2011; Hartman et al., 2002; Wilson et al., 2014b; Fastovsky and Bercovici, 2016). One pattern that has emerged from these studies is that aquatic communities incurred proportionally fewer extinctions than terrestrial communities did (Bryant, 1987a, 1989; Archibald and Bryant, 1990; Sheehan and Fastovsky, 1992; Robertson et al., 2013). However, within aquatic communities, K-Pg survival varied among higher-level taxonomic groups. For example, freshwater elasmobranchs did not persist across the K-Pg boundary in North America (except for Myledaphus; Wynd et al., 2020), whereas basal actinopterygians were hardly affected–only the large-bodied vidalamiine amiid Melvius went extinct (Bryant, 1987b, 1989; Archibald and Bryant, 1990). Recent studies that have incorporated both finer taxonomic resolutions and relative abundance data have shown that the end-Cretaceous mass extinction event affected some aquatic taxa more than initially recognized. For example, among salamanders and albanerpetontids, the post-K-Pg “survival” fauna from the lowermost Tullock Member of the Fort Union Formation was taxonomically depauperate and highly uneven compared to late Maastrichtian assemblages (Wilson et al., 2014a). Frogs also show a decline in diversity following the K-Pg extinction event (Gardner and DeMar, 2013; Mercier et al., 2014). Turtles were also impacted by the mass extinction event. Holroyd et al. (2014) reported that in northern Montana only 18 of the 24 genus-level lineages present in the upper Maastrichtian are present in the lower Paleocene, giving a 75% survivorship record. Subsequent description of the turtle Saxochelys (Lyson et al., 2019) from the Hell Creek Formation decreases this survivorship record to 60%.
Teleost fishes are an important yet poorly understood component of the non-marine aquatic paleocommunities of the latest Cretaceous and early Paleocene. In marine environments, the diversification of crown teleosts appears to be directly related to the extinction of fishes at the end of the Cretaceous (Friedman, 2009; Schwarzhans and Stringer, 2020; Schwarzhans et al., 2024). The most prominent casualties of marine teleost fishes across the K-Pg boundary are large-bodied predatory fish, including pachycormids, pachyrhizodontids, ichthyodectiforms, enchodontids, and cimolichthyids. Their extant ecological analogues, such as scombroids (tunas and mackerels), xiphioids (billfishes), sphyraenids (barracudas), and carangoids (jacks and dolphinfishes) respectively, first appeared in the early Paleogene (Patterson, 1993), suggesting that they originated during a radiation of teleost fishes immediately following the K-Pg extinction event (Cavin, 2002). The K-Pg extinction event also marked a profound change in the structure of global marine fish communities; there was a dramatic increase in the relative abundances of ray-finned fishes in Paleocene paleocommunities based on the relative abundance of shark denticles and ray-finned fish teeth in deep-sea sediments (Sibert and Norris, 2015).
The detailed pattern of extinction and survival of non-marine teleost fishes across the K-Pg boundary is much less well understood. Archibald and Bryant (1990) found a 33% extinction of teleost genera and species in the Hell Creek Formation and Tullock Member of the Fort Union Formation of northeastern Montana. However, this was based on only six taxa, far fewer than the 20 that are currently recognized in the late Maastrichtian of the northern Western Interior of North America (Brinkman et al., 2021). More recently, a study of the differences in teleost assemblages of two localities that bracket the K-Pg boundary in the Denver Basin, Colorado, USA, provided a detailed understanding of the impact of the K-Pg extinction event on teleost fishes in that area (Wilson et al., 2024). They found that taxonomic richness was reduced, largely as a result of the loss of rare morphotypes. Also, they documented a change in relative abundance patterns, with some taxa that are rare in the Cretaceous assemblage becoming dominant in the Paleocene, and the Paleocene assemblage being of low evenness compared to the Cretaceous assemblage.
 Here, we document the diversity of teleost fishes from the early Paleocene of the northern Western Interior of North America based on isolated elements from six vertebrate microfossil localities from the Tullock Member of the Fort Union Formation of Garfield County, Montana, USA, and one from the Ravenscrag Formation of southern Saskatchewan, Canada (Figure 1). The inclusion of the locality form Saskatchewan increases the geographic region covered but is included because it documents the Pu2 interval, which is not yet known from the Hell Creek regions. Comparison of these early Palaeocene fish assemblages with those of the late Maastrichtian Hell Creek Formation of Montana described previously by Brinkman et al. (2014, 2021) provides an understanding of both the effect of the K-Pg mass extinction event on teleost fishes in this region and the subsequent changes in richness and abundance during the approximately first one million years following the event.
Here, we document the diversity of teleost fishes from the early Paleocene of the northern Western Interior of North America based on isolated elements from six vertebrate microfossil localities from the Tullock Member of the Fort Union Formation of Garfield County, Montana, USA, and one from the Ravenscrag Formation of southern Saskatchewan, Canada (Figure 1). The inclusion of the locality form Saskatchewan increases the geographic region covered but is included because it documents the Pu2 interval, which is not yet known from the Hell Creek regions. Comparison of these early Palaeocene fish assemblages with those of the late Maastrichtian Hell Creek Formation of Montana described previously by Brinkman et al. (2014, 2021) provides an understanding of both the effect of the K-Pg mass extinction event on teleost fishes in this region and the subsequent changes in richness and abundance during the approximately first one million years following the event.
GEOLOGY
In North America, the lower Paleocene approximately corresponds to the Puercan (Pu) North American Land Mammal “age” (NALMA). The Puercan is divided into three subintervals (Pu1, Pu2, Pu3) based on successive first appearances of mammalian taxa. Early Puercan (Pu1) assemblages are of particular interest because they document the biota immediately after the K-Pg bolide impact. In this study, we sampled three Pu1 localities, all from the Tullock Member of the Fort Union Formation (hereafter Tullock Member) of Garfield County, Montana: McKeever Ranch 1 (UCMP locality V72210), Worm Coulee 1 (UCMP locality V74111), and Morales 1 (UCMP locality V77128). All three localities are preserved in channel facies. Worm Coulee 1 and Morales 1 were deposited during the Paleocene portion of Chron 29r (Swisher et al., 1993; LeCain et al., 2014; Sprain et al., 2018), and between two coals: the IrZ coal, which lies immediately above the K-Pg boundary layer, and the Hauso Flats Z (HFZ) coal. High-resolution 40Ar/ 39Ar geochronology performed on tephras preserved within these coal beds revealed that these Pu1 assemblages are confined to the first ca.70 ka after the K-Pg boundary (Renne et al., 2013; Sprain et al., 2015, 2018). McKeever Ranch 1 is bracketed below by the IrZ and above by another lignite that is younger than HFZ (Wilson Mantilla et al., 2021). Thus, McKeever Ranch 1 is likely younger than Worm Coulee 1 and Morales 1, probably within 105-139 ka after the K-Pg boundary.
Middle Puercan (Pu2) assemblages are not presently known from the Tullock Member. Localities that were previously thought to be of Pu2/3 age are now considered to be from the Pu3 subinterval (Sprain et al., 2018), including the three localities sampled here: Howard’s Biscuit Butte 2 (UCMP locality V81029), Yellow Sand Hill 8 (UCMP locality V75194), and Garbani 13-NW Harley’s High (UCMP locality V73080). All of these are preserved within channel facies that are bracketed by the W coal and Y coal, which have estimated ages of 65.741 Ma and 65.118 Ma, respectively (~300-900 ka after the K-Pg boundary; Sprain et al., 2015, 2018). The age of these localities within this interval varies because Yellow Sand Hill is likely slightly younger than the Garbani-13 and Howard’s Biscuit Butte 2 (Weaver et al., 2022).
Samples of teleost fishes from the Ravenscrag Formation of southern Saskatchewan were available from a single locality, the Pine Cree Park locality, which is located north-east of Eastend, Saskatchewan in Pine Cree Provincial Park (Figure 1). Fossils from Pine Cree Park occur in fine- to very fine-grained sandstone along with siltstone clasts and the broken shells of mollusks and has been interpreted as part of a channel deposit (Redman et al., 2015). This locality was discovered by Loris Russell in 1971. Mammal specimens from the locality were described by Russell (1974) and Scott et al. (2016). Study of the locality by Redman et al. (2015) and Scott et al. (2016) provisionally considered the locality to be Pu2 in age based on Pu2 mammals.
MATERIALS AND METHODS
This study is based on material from vertebrate microfossil localities that originally were bulk sediment sampled and underwater screen washed in the search for mammals of Puercan age. As is typical for such localities, fishes are represented by disarticulated, isolated elements. The combined taxonomic/morphotype approach adopted by Brinkman et al. (2013, 2014, 2017, 2021) in studies of fishes from the Milk River Formation of Alberta, Canada, the Grand Staircase/Escalante region of Utah, and the Hell Creek Formation of Montana, USA, is used to ensure that all available material is incorporated into the analysis of the diversity of teleosts present.
Interpretations of the diversity of teleosts are based primarily on abdominal centra and tooth-bearing elements. Morphologically distinctive elements that could not be identified but were judged to be from taxonomically distinct fishes, rather than a result of variation within a taxon, were given alpha-numeric designations and treated as operational taxonomic units (OTU) in the faunal analyses. Whereas morphologically distinct dentaries generally can be taxonomically distinct, vertebral elements must be evaluated more cautiously because of variation along the vertebral column. The range in variation along the vertebral column in extant teleosts provides a framework in which to evaluate whether distinct morphotypes are from different regions of the vertebral column or are from taxonomically distinct groups. Acanthomorph and ostariophysan teleosts provide challenges because of the high degree of variation along the column in members of these groups. To ensure that interpretations of the taxonomic diversity of acanthomorphs was not artificially inflated by this variation the first abdominal vertebra was emphasized in defining distinct morphotypes. More posterior abdominal centra were associated with the first centrum morphotypes based on shared or transitional morphological features. Taxonomic units based on more posterior abdominal centra were recognized only when they displayed morphological features that clearly separated them from all the first abdominal centra that were observed.
To identify the elements present, comparisons were made with osteological specimens of extant taxa. The comparative collections of the University of California Museum of Paleontology (Berkeley, California, USA), Royal Ontario Museum (Toronto, Ontario, Canada), Canadian Museum of Nature (Ottawa, Ontario, Canada), University of Michigan (Ann Arbor, Michigan, USA), and Royal Tyrrell Museum of Palaeontology (Drumheller, Alberta, Canada) provided a broad range of comparative specimens of recent fishes. A list of recent osteological specimens used for comparison is given in Brinkman et al. (2021, appendix 1).
To aid in the identification of acanthomorph tooth-bearing elements, µCT scans were used to investigate the pattern of canals within the elements. Two specimens, UCMP 191569 from locality V73080, and UALVP 58820 from the Pine Cree locality, were scanned at the University of Alberta using a SkyScan 1172 scanner using a voltage of 70V and a current of 129µA, and voxel size of 8.52 µm. The internal structure of the element was reconstructed through interpolated slice-by-slice segmentation using Amira 2019.4.
To document the morphological variation within the taxonomic units recognized, an extensive series of photographs of the elements described here is included. Specimens were whitened with ammonium chloride before photography to emphasize surface relief and were photographed with a digital still camera attached to a Wild MC3 microscope.
Comparison of faunal assemblages preserved in different localities or formations are based on both presence/absence and relative abundance data. Badgley (1986) concluded that, when formerly articulated material has been widely dispersed and has accumulated as isolated specimens, the minimum number of identifiable elements of a taxon is the best basis for documenting differences in the relative abundance of taxa between sites. Because the fossil assemblages included in this study accumulated under generally similar taphonomic conditions, major differences in relative abundance of taxa in localities being compared are interpreted as a reflection of differences in relative abundance of taxa in the original source communities, rather than differences in taphonomic processes. Brinkman (2008) argued that the biases introduced by the taphonomic processes can be further minimized by focusing on taphonomically similar elements. Because centra from different kinds of teleosts are both taphonomically similar and more abundant than tooth-bearing elements, only centra were used to quantify the difference in relative abundance of teleosts at localities being compared. The counts of centra of each OTU recognized here from the localities examined are listed in Table 1.
To assess potential effects of the mass extinction on the teleost assemblages leading up to and across the K-Pg boundary, we combined three datasets for quantitative analysis: two derived from Garfield County, northeastern Montana and one from southwestern Saskatchewan, for quantitative analysis. The first dataset was gathered from the Hell Creek Formation as presented in Brinkman et al. (2021, table 2) and the second from the Tullock Member as assembled here (Table 1). These two datasets comprise a total of 3,348 vertebral centra from 13 localities that span most of the stratigraphic thicknesses of the Hell Creek Formation (all Lancian) and the lower half of the Tullock Member (Pu1 and Pu3). These datasets were analysed to estimate the local teleost diversity of northeastern Montana. The third data set was compiled from the Pine Cree Park locality (Pu2) of the Ravenscrag Formation and was combined with the local Montana datasets to fill the temporal gap (i.e., Pu2) between the Pu1 and Pu3 localities of the Tullock Member. All three datasets with a combined total of 5,486 centra were analysed to estimate regional teleost diversity. For each locality we calculated: i) the raw number of OTUs; ii) the standing richness (i.e., the number of OTUs that first or last appear in a horizon or that range through a horizon); iii) the standing richness of local survivors (i.e., the number of OTUs that first appear in the Hell Creek and last appear in the Tullock Member); and iv) the expected number of OTUs in 124-specimen subsamples. See Table 1 and Brinkman et al. (2021, table 2) for the OTU nomenclature. Expected richness with 95% confidence intervals was calculated using Analytic Rarefaction version 1.3 (Holland, 2003) and only for the 10 best-sampled localities with more than 125 specimens. The three excluded localities (UCMP localities V77130 [N=51], V75194 [N=61], and V81029 [N=79]) are considered poorly sampled on account of their rarefaction curves failing to reach an asymptotic phase. We also calculated the relative abundances of the higher-level taxonomic groupings of teleosts per Brinkman et al. (2021, figure 20) to track any potential changes across the K-Pg boundary and to the Pu3 subinterval.
All specimens from the Tullock Member of the Fort Union Formation used in this study are housed in the University of California Museum of Paleontology (UCMP). Specimens from the Pine Cree Park locality are housed in the Royal Saskatchewan Museum (RSM) and the University of Alberta Laboratory for Vertebrate Palaeontology (UALVP).
SYSTEMATIC PALEONTOLOGY
Division TELEOSTEOMORPHA Arratia, 2001
Subdivision TELEOSTEI Müller, 1844 (sensu Patterson and Rosen, 1977)
Cohort ELOPOMORPHA Greenwood et al., 1966
Order ALBULIFORMES sensu Forey, Littlewood, Ritchie, and Meyer, 1996
Suborder ALBULOIDEI sensu Forey, Littlewood, Ritchie, and Meyer, 1996
Family PHYLLODONTIDAE Dartevelle and Casier, 1943
Genus PHYLLODUS Agassiz 1839
Phyllodus paulkatoi Estes and Hiatt, 1978
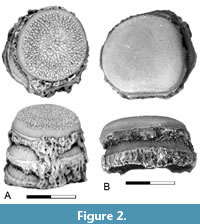
Figure 2
 1969b cf. Paralbula casei, Estes et al., p. 11
1969b cf. Paralbula casei, Estes et al., p. 11
1969a Phyllodus toliapicus, Estes, p. 319-321, fig. 1A-C
1978 Phyllodus paulkatoi, Estes and Hiatt, p. 1-10. fig. 1-2
1989 Phyllodus paulkatoi, Bryant, p. 26
2021 Phyllodus paulkatoi, Brinkman et al., p. 941, fig. 3B-C
Voucher specimen. From Pu3 localities of the Tullock Member of the Fort Union Formation: UCMP 191569 from locality V73080: two isolated teeth and two partial tooth-plates, one with three stacked teeth and the other with two stacked teeth.
Description. Phyllodus paulkatoi is represented by both isolated teeth and partial tooth-plates from UCMP locality V73080. In the partial tooth-plates, the teeth are arranged in stacks with the replacement teeth directly below the functional teeth, a defining feature of the genus (Estes, 1969a) (Figure 2A-B). The teeth are low, circular in outline, and have a wide-open base. Surface texture varies, with some teeth having a rugose surface (Figure 2A) and others being smooth (Figure 2B).
Remarks. Phyllodus paulkatoi was described by Estes and Hiatt (1978) based on partial basibranchial tooth-plates from the Upper Cretaceous Hell Creek Formation and the lower Paleocene Tullock Member. Phyllodus, along with the late Campanian taxon Paralbula, are members of the Phyllodontidae, an extinct group of albuloid fish in which the flattened, button-like teeth of the basibranchial are arranged in multiple sets of replacement teeth. In Phyllodus the teeth are superimposed in stacks, whereas in Paralbula the teeth are alternating or irregularly arranged. In the Puercan localities examined here, Phyllodus was only observed in localities of Pu3 age and only in the Montana region.
Subdivision OSTEOGLOSSOMORPHA Greenwood et al., 1966
Order indet.
Genus CORIOPS Estes, 1969b
Coriops sp.
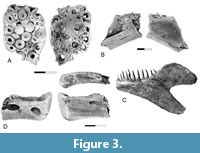
Figure 3A
 1969b Coriops amnicolus, Estes, p. 7-9, plate 4
1969b Coriops amnicolus, Estes, p. 7-9, plate 4
1989 Coriops amnicolus, Bryant, p. 25
1990 Teleost D, Brinkman, p. 44, fig. 4
2005 Coriops, Neuman and Brinkman, p. 174-176, fig. 9.6A
2010 Coriops, Larson, Brinkman, and Bell, p. 1165, fig. 5A
2013 Coriops, Newbrey et al., fig. 3F-I
2013 Coriops, Brinkman et al., p. 207-209, fig. 10.11A-B
2021 Coriops, Brinkman et al., p. 944-945. fig. 6A
Voucher specimens. From Pu3 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 191562 from locality V73080, nearly complete basibranchial element of large size.
Description. The presence of Coriops in the early Paleocene is documented by a nearly complete basibranchial from the Pu3 interval of the Tullock Member (Figure 3A). As in the basibranchials of Coriops from the upper Maastrichtian, blunt teeth cover the occlusal surface of the element, and the ventral surface is covered by a lacy network of bone. The element differs from Cretaceous specimens in that fewer teeth are present, and the teeth in the center of the element are larger than teeth near the lateral edges.
Remarks. Coriops is widespread in the Late Cretaceous. It is represented by tooth-bearing elements in both the late Campanian and late Maastrichtian and is often the most abundant teleost present. Centra referred to Coriops by Neuman and Brinkman (2005) were subsequently shown to be characteristic of a larger group that includes both Coriops and Lopadichthys and are described separately below.
Genus LOPADICHTHYS Murray, Zelenitsky, Brinkman, and Neuman, 2018
Lopadichthys sp.
Figure 3B
2018 Lopadichthys, Murray et al., p. 1-38
Voucher specimens. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: RSM 1954-114, posterior half of dentary.
Description. The presence of Lopadichthys in the Pine Cree Park locality is documented by a single, partial dentary. This specimen consists of the posterior end of the dentary, including the posterior end of the tooth row (Figure 3B). Two rows of teeth, mostly broken, are present, with the teeth of the lateral row being relatively larger than those of the medial row. Both rows of teeth are located on the dorsal edge of the dentary and the broken bases of the teeth face dorsally. The lateral surface of the dentary is smooth and gently convex. The sensory canal is located near its ventral edge. Sensory canal pores are relatively small. The depth of the dentary increases posteriorly, indicating that the complete dentary was likely to have been triangular in lateral view.
Remarks. Lopadichthys colwellae was named based on three articulated skeletons preserved on a single slab from the upper Paleocene Paskapoo Formation of Alberta, Canada, and a series of isolated dentaries that had previously been referred to Joffrichthys (Murray et al., 2018). A dentary from the Pine Cree Park locality (Figure 3B) is referred to Lopadichthys based on similarities with the dentary of Lopadichthys colwellae from the upper Paleocene (Figure 3C). In both, the external surface is smooth and gently convex, the sensory canal is located near the ventral edge of the dentary, and a tall coronoid process is present. However, the presence of two rows of teeth of the Pine Cree Park specimen differentiates it from the dentary of Lopadichthys colwellae.
A dentary from the Lance Formation with similarities to Lopadichthys was described by Brinkman et al. (2021) and referred to as aff. Lopadichthys. This dentary differs from the Paleocene specimens of Lopadichthys in being more curved when seen in occlusal view and in having a concave ventral edge when seen in lateral view. However, the similarities in shape and development of the tooth rows suggest that they are phylogenetically related, and that Lopadichthys originated during the Cretaceous.
Coriops or Lopadichthys
Figure 4A-D
 1990 Teleost D, Brinkman and Eberth, p. 44-45, fig. 1
1990 Teleost D, Brinkman and Eberth, p. 44-45, fig. 1
2001 Teleost D, Peng, Russell, and Brinkman, p. 18, plate 4, fig. 7-9
2002 Morphoseries IIA-1, Brinkman and Neuman, p. 144-146, fig. 4.1-4.11
2005 Coriops, Neuman and Brinkman, p. 174-176, fig. 9.6B-D, 9.8D
2010 Coriops, Larson, Brinkman, and Bell, p. 1165, fig. 5A
2013 Coriops, Newbrey et al., fig. 3
2013 Coriops, Brinkman et al., p. 207-209, fig. 10.11
2014 Coriops, Brinkman et al. p. 252-253, fig. 4
2018 Coriops, Murray et al., fig. 12D-F
2019 Coriops, Brinkman, p. 115-119, fig. 3
2021 Coriops or Lopadichthys, Brinkman et al., p. 945-947, fig. 7A-E
Voucher specimens. From Pu1 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP V276782 from locality V77128, 10 centra; UCMP 276783 from locality V77128, 103 centra; UCMP 276784 from locality V77128, four centra; UCMP 230711 from locality V72210 approximately 15 centra; UCMP 230710 from locality V72210 one centrum; UCMP 230661 from locality V74111, five centra; UCMP 230662 from locality V74111, two centra.
Localities. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: RSM 1954-95, 121 abdominal centra; UALVP 58818, (uncounted) abdominal centra.
Localities. From Pu3 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 230689 from locality V81029, six centra; UCMP 191567 from locality V73080, four centra.
Description. Osteoglossomorph centra like those of Coriops and Lopadichthys described by Brinkman et al. (2021) are present in Pu1, Pu2, and Pu3 localities. As in the Late Cretaceous samples, distinct anterior, mid, and posterior regions of the abdominal vertebral column can be recognized. The centra from the mid-abdominal region (Figure 4B-C) are shorter than wide and round or slightly higher than wide in end view. The neural arches are autogenous, and the articular surfaces for the neural arch are large, oval pits that extend for the full length of the centrum. A mid-dorsal pit is present between the neural arch articular pits. This pit is also oval and about half the size of the neural arch articular pits. Parapophyses are vertical flanges that extend ventrolaterally from the centrum. The base of the parapophysis is located anterior to the middle of the centrum and extends from the lateral edge of the neural arch articular pit to the ventral edge of the centrum. The parapophyses are laterally or ventro-laterally oriented. A rib articular pit is present posterior to the parapophysis. A mid-ventral pit is present.
The anterior abdominal centra (Figure 4A) are similar to the mid-abdominal centra in having autogenous neural arches and a mid-dorsal pit between the neural arch articular pits. They differ from the mid-abdominal centra in that the parapophyses are shorter and are oriented more strongly ventrally than in centra from more anterior positions in the vertebral series. These processes originate from lower down on the side of the centrum; therefore, they do not extend dorsally to the neural arch articular pit. The ventral surface of the centrum between the parapophyses is pierced by a series of small foramina rather than a single large mid-ventral pit.
Posterior abdominal centra (Figure 4D) differ from the mid-abdominal centra in having neural arches fused to the centrum. A ridge extends from the neural arch to the tip of the parapophyses. As in the more anterior centra, a mid-ventral pit is present.
Remarks. Osteoglossomorph centra from the upper Campanian Belly River Group were referred to Coriops by Neuman and Brinkman (2005). However, these were shown to differ little from those of the Paleocene genus Lopadichthys by Murray et al. (2018). Brinkman et al. (2021) concluded that the similarity in the centra of these genera should be considered as evidence that they are members of a higher-level group of osteoglossomorphs. Additionally, because Brinkman (2019) noted the similarity of this morphology to centra of Phareodus as documented by Divay and Murray (2016b) from the Eocene Bridger Formation, this larger group of osteoglossomorphs likely also includes some Paleogene osteoglossids. Because such a group is currently not recognized, they will be referred to below as centra of Coriops/Lopadichthys type.
Centra of Coriops/Lopadichthys type are widely distributed in both the upper Maastrichtian and lower Paleocene formations. They are generally of high relative abundance in upper Maastrichtian localities (Brinkman et al., 2021, table 2). Their relative abundance varies in the Paleocene but is consistently a lower percentage than in the Cretaceous. Based on counts of centra (Table 1), they are most abundant at UCMP locality V77128 (Pu1) where they constitute 19% of teleost abdominal centra present. In contrast, they are absent at locality V75194 (Pu3), which might be a sampling bias (only 61 centra in total; Table 1).
Genus OSTARIOSTOMA Schaeffer, 1949
Ostariostoma sp.
Figure 3D
1991 Ostariostoma, Grande and Cavender, p. 405-416
2005 Teleost unidentified dentary #4, Neuman and Brinkman, p. 176; fig. 9.7D
2017 Ostariostoma, Brinkman et al., p. 19-23, figs. 9-10
2021 Ostariostoma, Brinkman et al., p. 947, fig. 6C
Voucher specimens. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Formation, Saskatchewan: UALVP 58819, one dentary.
Description. A single dentary like that referred to Ostariostoma by Brinkman et al. (2017) was recovered from the Pine Cree Park locality (Figure 3D). This specimen, which consists of the anterior end of the element, preserves two sensory canal pores. As in the Cretaceous dentaries, these are exceptionally large. A series of tooth bases is present on the occlusal surface of the dentary, indicating that a single row of relatively large teeth was present.
Remarks. Ostariostoma is one of the few teleosts from the Upper Cretaceous or lower Paleocene of the Western Interior of North America represented by an articulated skeleton (Schaeffer, 1949; Grande and Cavender, 1991). The single specimen, which is from the upper Maastrichtian to lower Paleocene Livingston Group in Montana, is preserved as an impression in a hard mudstone. Fine detail is present in the natural mould, allowing for detailed descriptions based on latex peels taken from the original specimen (Grande and Cavender, 1991). A dentary from the upper Santonian Milk River Formation was referred to Ostariostoma by Brinkman et al. (2017) based on comparison with a cast of the original articulated specimen. The Milk River dentary was similar to the dentary of the type specimen in the presence of a single row of large teeth and of three large sensory canal pores in the lower half of the elements (Brinkman et al., 2017, figure 9). Dentaries of Ostariostoma are widely distributed in the Cretaceous, being present in the upper Santonian Milk River Formation (Brinkman et al., 2017, figures 9-10), the upper Campanian Belly River Group (Neuman and Brinkman, 2005, figure 9.7D), and the upper Maastrichtian Lance Formation (Brinkman et al., 2021). The dentary present in the Pine Cree Park locality shows little difference from the Cretaceous specimens.
Superorder CLUPEOMORPHA Greenwood, Rosen, Weitzman, and Myers, 1966
Order ELLIMMICHTHYIFORMES Grande, 1985
Family ARMIGATIDAE Murray and Wilson, 2013
Genus DIPLOMYSTUS Cope, 1877
Diplomystus sp.
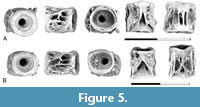
Figure 5
1997 Teleost Q, Eberth and Brinkman, p. 58
 2002 Morphoseries IIB-2 Brinkman and Neuman, p. 149-150, fig. 7
2002 Morphoseries IIB-2 Brinkman and Neuman, p. 149-150, fig. 7
2013 Diplomystus sp. Brinkman et al., p. 213, fig. 10.15
2016a Diplomystus, Divay and Murray, p. 2-4, fig. 2
2019 Diplomystus, Brinkman, p.120-121, fig. 6
Voucher specimens. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: RSM 1594-35, one centrum; RSM 1594-36, one centrum; RSM 594-104, two centra.
Description. The presence of Diplomystus in the Pine Cree Park assemblage is documented by four centra from the posterior abdominal region of the vertebral column. As in the specimens of Diplomystus from the Wasatch Formation described by Divay and Murray (2016a, figure 2), the neural arch articular pits are shallow depressions restricted to the anterior half of the centrum (Figure 5). These surfaces are separated by a mid-dorsal pit that is triangular, with the posterior end being widest. This pit is bordered laterally by a ridge that extends posteriorly from the neural arch articular pits and ends in a low postero-dorsal process. Parapophyses are preserved in place, partially fused to the centrum. A broad area above the parapophyses is formed by a coarse network of ridges. The lateral surface of the parapophyses is also formed by a network of ridges. Ventrally a variable number of antero-posteriorly oriented ridges are present, with one specimen having three closely spaced ridges (Figure 5A) and the second having a mid-ventral pit (Figure 5B). A similar range of variation is present in specimens from the Upper Cretaceous of Utah (Brinkman et al., 2013, figure 10.15) and from the early Eocene of Wyoming (Divay and Murray, 2016a, figure 2E-F), so this likely reflects variation along the column.
Remarks. Diplomystus is well represented by articulated specimens from both Cretaceous and Eocene localities (Grande, 1984; Bagils, 2004). Isolated centra from the lower Eocene Bitter Creek locality of the Wasatch Formation, Wyoming, were described by Divay and Murray (2016a), and these allowed isolated centra of Diplomystus from Cretaceous vertebrate microfossil localities to be recognized. Based on the distribution of centra, Diplomystus is widespread in non-marine beds of the Upper Cretaceous of the Western Interior of North America. It is present in the Cenomanian Dakota Formation and the Santonian portion of the John Henry Member of the Straight Cliffs Formation, both of Utah (Brinkman et al., 2013), and the upper Campanian Belly River Group of Alberta (Brinkman, 2019).
Superorder OSTARIOPHYSI Sagemehl, 1885 (sensu Fink and Fink, 1996)
Order GONORYNCHIFORMES Berg, 1940
gen. et sp. indet. type H
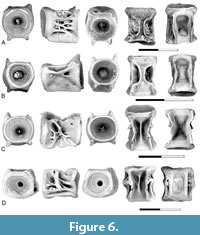
Figure 6
2002 Morphoseries IIIA-1, Brinkman and Neuman, p. 150-151, fig. 8.1-8.4
 2005 Morphoseries IIIA-1 (teleost indeterminate), Neuman and Brinkman, p. 180, fig. 9.8G
2005 Morphoseries IIIA-1 (teleost indeterminate), Neuman and Brinkman, p. 180, fig. 9.8G
2013 Genus et sp. indet. type HvB, Brinkman et al., p. 225, fig. 10.26
2014 Genus and species indet H-vB, Brinkman et al., p. 261, fig. 10C
2017 ? Ostariostoma, Brinkman et al., p. 21, fig. 10-11
2019 Gonorynchiformes gen. et sp. indet. type H, Brinkman, p. 127-129, fig. 11
2021 Gonorynchiformes gen. et sp. indet. type H, Brinkman et al., p. 947-949, fig. 8A-C
Voucher specimens. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: RSM 1594-8, one anterior centrum; RSM 1594-10, one first abdominal centrum (centrum type HvB); RSM 1594-11, one anterior centrum (centrum type HvB); RSM 1594-5, one mid-dorsal centrum (centrum type HvA); RSM 1594-6, one abdominal centrum (centrum type HvA); RSM 1594-7, one abdominal centrum (centrum type HvA); RSM 1594-93, 33 abdominal centra (centrum type HvA); RSM 1594-92, 21 first abdominal centra (centrum type HvB).
Description. Gonorynchiform indet. type H is represented by two distinctive centrum morphotypes, one, initially referred to as type HvB, was interpreted as a first abdominal centrum, and the second, initially referred to as type HvA, was interpreted as representing more posterior abdominal centra (Brinkman et al., 2017, 2021; Brinkman, 2019).
The first abdominal centrum (Figure 6A-B) corresponds to the morphology of those from the Upper Cretaceous identified as such by Brinkman et al. (2014, 2017) and Brinkman (2019), and other gonorynchiform first centra from the Late Cretaceous (Divay et al., 2020, figure 6A) and early Eocene (Divay and Murray, 2016a, figure 4A) in being longer than wide and having a distinctive sub-rectangular mid-ventral pit. Neural arches are not fused to the centrum. The neural arch articular pits are characteristic, long, oval pits that are separated from each other by a robust bar of bone. Parapophyses are absent, and no distinct parapophyseal pit is present. The sides of the centrum vary from having a series of struts extending between the ends of the centrum (Figure 6A) to being smooth (Figure 6B).
The more posterior abdominal centra (Figure 6C-D) are similar to the first abdominal centra in being longer than wide and having a sub-rectangular mid-ventral pit. In some centra, the pit is constricted at about mid length, as in the first abdominal centrum. Neural arches are fused to the centrum and a narrow mid-dorsal ridge extends for the length of the centrum between the neural arch bases. The development of the parapophyses varies, likely because of variation along the vertebral column. On specimens interpreted as being from a more anterior position on the vertebral column (Figure 6C), parapophyses are autogenous and distinct parapophyseal pits are present on the lateral surface of the centrum. A ridge extends from the neural arch to the dorsal edge of the parapophyseal pit. On specimens interpreted as being from a more posterior position (Figure 6D) parapophyses are fused to the centrum. These extend from the neural arch to the ventral edge of the centrum.
Remarks. Gonorynchiform gen. et sp. indet. type H was first recognized by Brinkman et al. (2013) based on material from the Turonian Smoky Hollow Member of the Straight Cliffs Formation, Utah. Two centrum morphotypes, initially referred to as centrum type HvA and type HvB, were recognized as representing different positions in a single vertebral series. Later, Brinkman et al. (2017) recognized that centrum type HvB matched the centrum identified as a gonorynchiform first abdominal centrum by Divay and Murray (2016a, figure 4A-B), indicating affiliations with that teleost group.
Based on a comparison of the centra of teleost Gonorynchiform type H and the centra partially visible in the type specimen of Ostariostoma, Brinkman et al. (2017) suggested that gonorynchiform indet. type H was synonymous with Ostariostoma. This was supported by a similarity in distribution of the centra of Gonorynchiform indet. type H and the dentaries referred to Ostariostoma. If correct, this would imply that Ostariostoma was a member of the Gonorynchiformes rather than the Osteoglossomorpha as generally accepted (Grande and Cavender, 1991; Hilton, 2002; Wilson and Murray, 2008; Lavoué, 2016). However, pending the discovery of additional specimens that confirm the association of the dentaries referred to Ostariostoma and the centra of gonorynchiform indet. type H, these are retained as distinct operational taxonomic units.
Centra of gonorynchiform indet. type H are widely distributed in the Upper Cretaceous of the Western Interior, being present in the Turonian Smoky Hollow Member of the Straight Cliffs Formation (Brinkman, 2013), the Santonian Milk River Formation (Brinkman et al., 2017), the upper Campanian Belly River Group (Brinkman, 2019), and the upper Maastrichtian Hell Creek and Lance formations (Brinkman et al., 2021). They are fairly common at the Pine Cree Park locality (60 specimens) but only constitute about 3% of all teleost abdominal centra present; none are present in the Montana localities (Table 1). The variable abundances of this taxon may reflect local environmental conditions rather than regional patterns of distribution because it has a patchy distribution in the Cretaceous: it is abundant in the Milk River Formation and Belly River Group of Alberta but rare in the Hell Creek Formation (Brinkman et al., 2014, 2017, 2021).
Genus NOTOGONEUS Cope, 1885
Notogoneus sp.
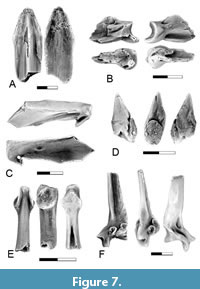
Figure 7-Figure 8
1997 Teleost R, Eberth and Brinkman, p. 57
 2002 Morphoseries IIIA-2, Brinkman and Neuman, p. 151, fig. 8.5-8.8
2002 Morphoseries IIIA-2, Brinkman and Neuman, p. 151, fig. 8.5-8.8
2019 Notogoneus, Brinkman, p. 126-129, fig. 10
2020 Notogoneus, Divay et al., p. 1-18, figs. 2-9
2021 Notogoneus, Brinkman et al., p. 949, fig. 8D
Voucher specimens. From Pu1 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276776 from locality V77128, three centra; UCMP 276777 from locality V77128, one centrum; UCMP 276778 from locality V77128, one centrum; UCMP 230713 from locality V72210, one first abdominal centrum; UCMP 230716 from locality V72210, one centrum; UCMP 230706 from locality V77210, one centrum; UCMP 230717 from locality V72210, one centrum.
Localities. From Pu3 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 230690 from locality V81029, one centrum; UCMP 230691 from locality V81029, one centrum; UCMP 230686 from locality V81029, basioccipital; UCMP 230729 from locality V75194, two centra; UCMP 191562 from locality V73080, three centra.
Localities. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: cranial elements: RSM 1594-78, anterior end of vomer; RSM 1594-80, anguloarticular; RSM 1594-81, retroarticular; RSM 1594-77, basibranchial; RSM 1594-82, one dentary; RSM 1594-100, two basibranchials; RSM 1594-99, one dentary; RSM 1594-98, one anguloarticular; RSM 1594-101, approximately 40 basioccipitals; UALVP 58823, one basioccipital.
Vertebral elements: RSM 1594-29, one centrum; RSM 1594-30, one centrum; RSM 1594-31, one centrum; RSM 1594-32, one centrum; RSM 1594-103, approximately 328 centra; RSM 1594-102, one centrum of large size; UALVP 58814, one centrum of large size.
Appendicular skeletal elements: RSM 1594-79, one pectoral fin ray base; RSM 1594-83, one pectoral fin ray base; RSM 1594-97, two pectoral fin ray bases.
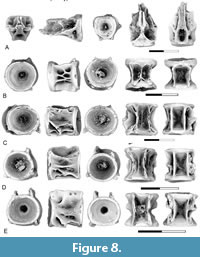 Description. Notogoneus is represented in the lower Paleocene localities of Saskatchewan and Montana by both cranial elements and elements of the axial and appendicular skeletons (Figure 7-Figure 8). The cranial elements include the vomer, basioccipital, anguloarticular, retroarticular, dentary, and basibranchials.
Description. Notogoneus is represented in the lower Paleocene localities of Saskatchewan and Montana by both cranial elements and elements of the axial and appendicular skeletons (Figure 7-Figure 8). The cranial elements include the vomer, basioccipital, anguloarticular, retroarticular, dentary, and basibranchials.
The vomer (Figure 7A) is represented by its anterior end. It is like that of the Cretaceous specimens described by Divay et al. (2020, figure 2A-B) in being an elongate, flattened element with a triangular anterior end and a slightly rounded ridge extending longitudinally on its dorsal surface.
The basioccipital (Figure 8A) is like that described by Divay and Murray (2016a, figure 3A) and by Divay et al. (2020, figure 5E) in that the posterior articular surface is higher than wide. Dorsally, a pair of ridges extends forward from the exoccipital articular surface forming a bi-lobed mid-dorsal ridge. Ventrally, a large circular mid-ventral pit is present at the posterior end of the basioccipital. A narrow flat surface extends anterior to the pit. This surface is depressed relative to the more lateral areas of the basioccipital.
The dentary is edentulous, and although it is incomplete, the preserved base of the ventral process, by the symphysis, suggests that the complete dentary would have been deeply forked, with ventral and coronoid processes joining anteriorly (Figure 7C). It has a smaller symphysis than dentaries of Notogoneus from the Late Cretaceous described by Divay et al. (2020, figure 3A), but is otherwise morphologically identical, including in the placement of the only visible foramen, immediately beneath the oral margin on the medial surface of the coronoid process.
The anguloarticular (Figure 7B) is a short, relatively robust element with a well-defined concave articular surface facing posteriorly for the articulation with the quadrate. The ventral surface has a shallow triangular articular surface for the articulation with the retroarticular. It does not significantly differ from the Upper Cretaceous anguloarticular described by Divay et al. (2020, figure 3E).
The retroarticular (Figure 7D) is a short, spine-like element with a circular articular surface for contact with the postero-ventral surface of the anguloarticular. As was described for Late Cretaceous Notogoneus elements (Divay et al., 2020), despite being adjacent to the articulation of the mandible with the quadrate, the retroarticular does not seem to have been included in this articulation.
The basibranchial (Figure 7E) is a rod-like bone with circular lateral facets for articulation with the anterior basihyals at the anterior end of the bone. It differs from the Upper Cretaceous specimens illustrated by Divay et al. (2020, figure 5A) in that the articular surface for the basihyals is more nearly circular. This may reflect a morphological difference between the Late Cretaceous and Paleocene taxa, or this may result from preservation differences.
Centra (Figure 8B-E) are referred to Notogoneus based on comparison with isolated centra of Notogoneus from the lower Eocene Wasatch Formation described by Divay and Murray (2016a) and from the upper Campanian Dinosaur Park Formation described by Divay et al. (2020). Both first abdominal centra (Figure 8B) and more posterior abdominal centra (Figure 8C-E) are present in our assemblages. The first abdominal centrum is sub equal in width and length. Neural arches are autogenous. Neural arch articular pits have a distinctive sub-oval shape and are separated from one another by a narrow bar of bone. Dorsal processes extend from the posterior end of these pits. One to three variably developed buttresses are present on the lateral surface of the centrum. Parapophyses are absent. A large, rectangular mid-ventral pit is present, bordered by sharp ridges.
More posterior abdominal centra (Figure 8C-E) are holospondylous, with parapophyses and neural arches fused to the centrum. The neural arches are located close together and extend the full length of the centrum. A narrow mid-dorsal ridge is present between the bases of the neural arch. The parapophyses are ventro-laterally oriented processes restricted to the anterior half of the centrum. A sharp-edged ridge extends along the ventro-lateral edge of the centrum from the parapophyses to the posterior end of the centrum. A second major ridge extends between the ends of the centrum mid-way along its side. This ridge generally originates above the parapophyses (Figure 8D-E), but on some it contacts the parapophyses (Figure 8C). The ridge varies in thickness, likely because of variation along the column. Ventrally a rectangular mid-ventral pit bordered by sharp, parallel ridges is present.
The appendicular skeleton is represented by pectoral fin ray bases (Figure 7F). Fin ray bases were identified by Divay et al. (2020) based on comparison with this element in an articulated specimen of Notogoneus from the Green River Formation (see Divay et al., 2020, figure 8E). They are recognizable based on the characteristic bifurcation of the distal tip of the ventral process and the presence of a circular facet on the posterior surface of the fin ray base. As with the specimens described by Divay et al. (2020), these are from an anterior position on the fin, as indicated by the markedly concave, saddle-shaped articular head for direct articulation with the scapula.
Remarks. Notogoneus is one of the few genera of non-marine teleosts represented by articulated specimens in both Upper Cretaceous and Paleogene beds of North America (Grande and Grande, 1999; Grande et al., 2022). In addition, it is represented in the Upper Cretaceous Dinosaur Park Formation by isolated elements from vertebrate microfossil localities (Divay et al., 2020) that were identified by comparison with articulated Notogoneus osculus specimens from the Green River Formation and recent skeletons of other genera within the Gonorynchiformes, including the chanid Chanos chanos, and the gonorynchid Gonorynchus forsteri. Notogoneus is present but rare in the upper Maastrichtian (Brinkman et al., 2021). Elements of Notogoneus from the lower Paleocene show only minor differences from the corresponding elements from the Dinosaur Park Formation. The major difference in the cranial elements is the proportions of the dentary, which appear more elongate in the Puercan-age specimens. Centra of Notogoneus from the Puercan localities have generally more gracile lateral buttresses than centra from the Dinosaur Park Formation.
Notogoneus has a patchy distribution in both the Upper Cretaceous and the lower Paleocene. In the Upper Cretaceous Belly River Group, it is abundant in a series of fine-grained deposits in the Onefour area of southern Alberta representing very slow-moving waters on the lateral floodplains but is rare outside of this area and environment (Divay et al., 2020). It is also present but rare in the Hell Creek Formation (Brinkman et al., 2021). In the lower Paleocene, it is only present in the Pine Cree Park locality, where it is moderately abundant (Table 1).
Series OTOPHYSI Garstang, 1931 (sensu Rosen and Greenwood, 1970)
Order indet.
Gen. et sp. indet. U3/BvD
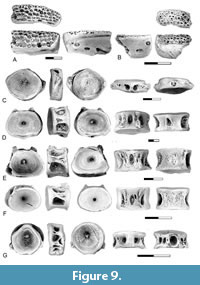
Figure 9
2013 Otophysi gen et sp. indet. type U-3/BvD, Brinkman et al., p. 215-219, figs. 10.19A-C, 10.20A-C, 10.21A-C
 2014 Otophysi gen et sp. indet. Brinkman et al., p. 255-256, fig. 7A-D
2014 Otophysi gen et sp. indet. Brinkman et al., p. 255-256, fig. 7A-D
2017 Otophysi genus et sp. indet U-3/BvD, Brinkman et al., p. 30-33, figs. 16-17, 18B
2019 Otophysi genus et sp. indet U3/BvD, Brinkman, p. 129-131, figs. 12-13B
2021 Otophysi gen. et sp. indet. U3/BvD, Brinkman et al., p. 949-951, fig. 9
Voucher specimens. From Pu1 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276732 from locality V77128, one partial centrum of type U3; UCMP 276740 from locality V77128, one partial centrum; UCMP V276731, from locality V74111, one partial centrum.
Localities. From Pu3 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 191569 from locality V73080, four dentaries; UCMP 191566 from locality V73080, approximately 44 centra of type U3; UCMP 191564 from locality V73080, approximately 43 post-Weberian abdominal centra; UCMP 191569 from locality V73080, four dentaries (tooth bearing elements of amiids and other teleosts included in this specimen number); UCMP 230688 from locality V81029, three anterior Weberian centra of intermediate size; UCMP 230692 from locality V81029, eight post-Weberian abdominal centra; UCMP 230706 from locality V75194, one anterior Weberian centrum; UCMP 230707 from locality V75194, three anterior Weberian centra.
Localities. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: RSM 1594-33, one abdominal centrum; RSM 1594-34, one abdominal centrum.
Description. The presence of Otophysi indet. type U3/BvD in the lower Paleocene is documented by dentaries, centra from the Weberian apparatus, and post-Weberian abdominal centra.
Dentaries of Otophysi indet. type U3/BvD were recovered from the Pu3 localities of Montana (Figure 9A-B). These are similar to Cretaceous dentaries described by Brinkman et al. (2013, figure 10.19) in having a tooth-bearing surface that is convex in cross section and covered by a pad of closely spaced teeth with the teeth arranged in loose rows and showing no variation in size across the tooth row. A series of three sensory canal pores is present on the dentary. The anterior-most sensory canal pore is located just posterior to the symphysis and is in a relatively more dorsal position than the other foramina. The more posterior pores are located near the ventral edge of the dentary.
The first Weberian centrum (Figure 9C, G) is greatly foreshortened; the anterior surface is nearly flat, and the posterior surface is shallowly concave. A pair of small circular pits separated by a rounded bar of bone is present on the dorsal surface. Parapophyses are fused to the centrum but are broken so their full length is not known. The bases of each process forms vertically oriented ridges on the sides of the centrum. A distinctive circular fossa is present on the ventral surface of the centrum. In specimens from the Pu1 interval, this fossa is large, with a diameter that is greater than half the length of the centrum (Figure 9G). In specimens from the Pu3 localities in Montana the fossa is much smaller (Figure 9C).
Abdominal centra (Figure 9D-F) were initially referred to as centrum type BvD and were not recovered from the Pu1 localities but are abundant in the Pu3 localities of Montana. They are also present, although rare, in the Ravenscrag Formation of Saskatchewan. The centrum is low and wide. Neural arches are fused to the centrum and a deep mid-dorsal pit is present between the bases of the neural arches. Parapophyses on anterior to mid-abdominal centra are autogenous and large parapophyseal pits are present on the ventro-lateral sides of these centra. In the posterior abdominal centra, the parapophyses are fused to the centrum. The ventral surface is covered by a network of antero-posteriorly oriented ridges. An elongated, slit-like mid-ventral pit is present in some specimens (Figure 9D-E) but absent on others (Figure 9F). The variable development of this pit is interpreted as a result of variation along the vertebral column, with the pit being present on centra from the anterior portion of the abdominal series.
Remarks. Otophysi indet. type U3/BvD was first recognized by Brinkman et al. (2013) based on specimens from the Upper Cretaceous of Utah. Otophysi indet. type U3/BvD was included in the Otophysi by Brinkman et al. (2013) because one centrum type included in this taxon, initially referred to as centrum type U3, was identified as part of the Weberian apparatus based on comparison with the cyprinid Cyprinus carpio. The second centrum morphotype included in this taxon, initially referred to as centrum type BvD, was interpreted as representing the post-Weberian abdominal centra. Dentaries referred to Otophysi indet. type U3/BvD by Brinkman et al. (2013, 2017) also supported the otophysan relationships of Otophysi indet. type U3/BvD because they had similarities with those of some catfishes. The presence of a Weberian apparatus was further supported by the identification of the third Weberian centrum by Brinkman et al. (2017). Brinkman et al. (2021) described a centrum morphologically transitional between the third Weberian centrum and abdominal centra that may be a fourth Weberian centrum. Brinkman et al. (2013) suggested that Otophysi indet. type U3/BvD was more closely related to catfish than to other otophysans because of similarities in the dentaries. However, the recovered Weberian centra resemble those of cypriniforms more than those of siluriforms, and the possibility that the features of the dentary shared with catfishes are present in basal members of the Otophysi cannot be excluded. Therefore, Otophysi indet. type U3/BvD may also be a basal member of the Otophysi, or perhaps a basal cypriniform with a siluriform-like dentary.
Teleost gen. et sp. indet. type U3/BvD is widespread in the Cretaceous, being present in the Turonian to the upper Campanian of southern Utah (Brinkman et al., 2013), the Milk River Formation (Brinkman et al., 2017), the Belly River Group (Brinkman, 2019), and the Hell Creek, Lance, and Scollard formations (Brinkman et al., 2014, 2021). The presence of centra and dentaries of Otophysi indet. type U3/BvD from the lower Paleocene show that this taxon crossed the K-Pg boundary.
Two distinct variants of teleost gen. et sp. indet. type U3/BvD are present in the lower Paleocene. In Pu1 assemblages, it is represented by anterior Weberian centra that are similar to those from the Late Cretaceous in that the circular fossa on the ventral surface of the centrum is large (Figure 9G). The anterior Weberian centra from the Pu3 localities of the Tullock Member differ in that the circular fossa is much smaller (Figure 9C). Dentaries and abdominal centra of Otophysi indet. type U3/BvD from the Pu3 localities of Montana also differ from their Late Cretaceous counterparts. The dentaries differ in the arrangement of the sensory canal pores. In the dentaries from the Pu3 interval, the anterior-most sensory canal pore is located just posterior to the symphysis, and it is more dorsally positioned compared to the more posterior pores, which are located near the ventral edge of the dentary (Figure 9A-B). In the dentaries from Cretaceous localities, the anterior-most pore in the series is located further posteriorly and is not raised relative to the more posterior pores (Brinkman et al., 2013, figure 10.19; Brinkman et al., 2017, figure 16B). The abdominal centra from the Pu3 localities of Tullock Member differ from their Cretaceous counterparts in the shape of the parapophyseal articular pits and the development of the surface of the centrum dorsal to this pit. In the centra from the Tullock Member, the parapophyseal pits are oval in lateral view and are separated from the neural arch articular pit by a broad, flat surface (Figure 9D-F). This differs from the condition in the Cretaceous specimens where the pits are more rectangular in shape and abut against the neural arch articular pits so are separated from them by only a sharp ridge (Brinkman et al., 2013, figure 10.21B-C; Brinkman, 2019, figure 12C-D). The abdominal centra from the Pine Cree Park locality are like those of the younger (Pu3) localities of Montana.
Based on these morphological features the Pu1 and Pu3 morphotypes of teleost gen. et sp. indet. type. U3/BvD appear to be taxonomically distinct. To recognize this, the taxon from the Cretaceous and Pu1 deposits is designated Otophysi indet. type U3/BvD (K morph), and the taxon present in the Pu2 and Pu3 subintervals is designated type U3/BvD (P morph).
The distribution of centra of teleost indet. U3/BvD (P morph) in the Pu2 and Pu3 localities shows a strong paleobiogeographic pattern. It is moderately abundant in some of the localities in Montana (up to 28%) but very rare in the Pine Cree Park locality in Saskatchewan (<<0.1%; Table 1). This mirrors the tendency for elements of teleost indet. U3/BvD (K morph) to show a strong latitudinal pattern of distribution in the Upper Cretaceous (Brinkman et al., 2013, 2017), being more abundant in more southerly localities, suggesting that this latitudinal pattern extended into the early Paleocene.
Cohort EUTELEOSTEI (sensu Arratia, 1999)
Family ESOCIDAE Rafinesque, 1815
Genus ESTESESOX Wilson et al., 1992
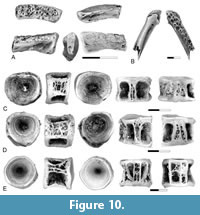
Estesesox sp.
Figure 10
1964 Platacodon (in part), Estes, p. 51-53, fig. 26
 1992 Estesesox foxi, Wilson et al., p. 819-846, figs. 2-3, 5, 7-8
1992 Estesesox foxi, Wilson et al., p. 819-846, figs. 2-3, 5, 7-8
1997 Teleost N, Eberth and Brinkman, p. 57
2001 Teleost indet. Peng, Russell, and Brinkman, p. 18, Plate 4, fig. 12-13
2002 Morphoseries IB-1, Brinkman and Neuman, p. 141-143, fig. 2-1 to 2-14
2005 Morphoseries IB-1, Neuman and Brinkman, p. 176, fig. 9.8B
2010 Esocoidea, Larson, Brinkman, and Bell, p. 1165, fig. 5C
2013 Salmoniform, Brinkman et al., p. 223-225, fig. 10.25
2014 Salmoniform, Brinkman et al., p. 257-259, fig. 9
2017 Estesesox, Brinkman et al., p. 33-34, fig. 19
2019 Esocidae, Brinkman, p. 133-136, figs. 15-16
2021 Estesesox, Brinkman et al., p. 951-952, fig. 10
Voucher specimens. From Pu1 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276730 from locality V74111, anterior end of dentary; UCMP 276728 from locality V74111, one palatine; UCMP 276729 from locality V77128, one palatine; UCMP 230659 from locality V74111, one abdominal centrum; UCMP 230660 from locality V74111, one abdominal centrum; UCMP 230708 from locality V72210, one abdominal centrum.
Description. The presence of Estesesox in the Pu1 is documented by a dentary, palatines, and centra. The dentary (Figure 10A) is similar to specimens of Estesesox from the Upper Cretaceous in the presence of multiple rows of teeth on the anterior end of the dentary. At least three rows of teeth are present. The teeth are all small and show little variation in size across the tooth row. The symphysis is tall and narrow, consisting of a vertical flange that forms the ventral half of the anterior end of the dentary.
The palatine (Figure 10B) is similar to the palatines from the upper Maastrichtian described by Wilson et al. (1992) and Brinkman et al. (2021). It is narrow, and only two rows of teeth are present.
Centra like those from the upper Maastrichtian that were referred to Estesesox by Brinkman et al. (2021) are also present in Pu1 localities (Figure 10C-E). As in esocids generally, these are simple spools with large pits for the neural arches and parapophyses. A mid ventral ridge is present. Centra from the anterior and posterior abdominal regions of the vertebral column are present. In centra from a more anterior position, the neural arch articular pits extend the full length of the centrum (Figure 10C-D). In ventral view, the medial edges of the parapophyseal articular pits converge towards the anterior end of the centrum. In centra from the posterior region of the vertebral column, the neural arch articular pits do not extend to the posterior end of the centrum (Figure 10E). In ventral view, the medial edges of the parapophyseal articular pits are sub-parallel to one another. They differ from posterior abdominal centra from Cretaceous localities in that the parapophyseal and neural arch articular pits are separated from one another by a surface formed by a network of bone ridges in the Puercan material, whereas these pits are separated by a single longitudinal ridge of bone in Cretaceous elements.
Remarks. Esocids were first recognized in the Late Cretaceous by Wilson et al. (1992) based on dentaries that had been previously referred to the acanthomorph Platacodon by Estes (1964). Wilson et al. (1992) recognized that the teeth of the dentaries had distinctive, C-shaped bases, a derived feature of esocids associated with the presence of depressible teeth. Two genera were named, Estesesox and Oldmanesox. These differ in that Oldmanesox has a single row of teeth with the teeth being larger and more widely spaced, whereas Estesesox has several rows of teeth at the anterior end of the dentary. Oldmanesox was represented by only a few specimens from the Dinosaur Park Formation, but Estesesox was abundantly represented by specimens from both the Dinosaur Park and the Hell Creek formations. In a later description of esocid dentaries from the Hell Creek Formation, Brinkman et al. (2014) recognized three taxa based on variation in the number of rows of teeth at the anterior end of the dentary. However, this variation was subsequently reinterpreted as variation related to ontogeny (Brinkman et al., 2021), and the esocid dentaries from the upper Maastrichtian were included in a single species of Estesesox.
Esocid centra from the Upper Cretaceous were first described by Brinkman and Neuman (2002). In a later study of the fishes from the Hell Creek Formation, three centrum morphotypes were recognized, which were assumed to be taxonomically distinct (Brinkman et al., 2014). However, based on a study of variation along the vertebral column in extant species of Esox these three morphs were reinterpreted as representing different regions along the vertebral column (Sinha et al., 2019).
Wilson et al. (1992) also described two kinds of esocid palatines from the Upper Cretaceous. One, from the upper Campanian Dinosaur Park Formation, had a broad tooth-bearing surface with multiple rows of teeth. The second, from the upper Maastrichtian Hell Creek Formation, was narrow and had only two rows of teeth. Wilson et al. (1992) referred the broad palatine to Estesesox and interpreted the narrow palatine as evidence for a second esocid. However, Brinkman et al. (2021) suggested that the late Campanian and late Maastrichtian representatives of Estesesox may differ in the palatine, so recognized only a single esocid in the late Maastrichtian assemblages.
Estesesox is represented in the lower Paleocene by dentaries, palatines, and centra. The dentaries and palatine do not differ from their upper Maastrichtian counterparts. The centra differ only in the presence of a broader area between the neural arch and parapophyseal pits on the posterior abdominal centra. Because only three vertebral centra are available from the lower Paleocene (Table 1), the full range of variation along the vertebral column is not represented, and the possibility that the centra being compared come from different regions of the column cannot be eliminated.
Genus ESOX Linnaeus, 1758
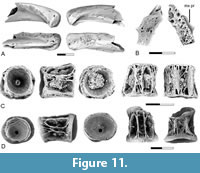
Esox sp.
Figure 11
 Voucher specimens. From Pu1 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276775 from locality V77128, three centra; UCMP 276774 from locality V77128, five centra; UCMP 276773 from locality V77128, eight centra; UCMP 230702 from locality V72210, anterior end of a dentary; UCMP 230701 from locality V72210, one palatine; UCMP 230659 from locality V74111, anterior abdominal centra (not counted); UCMP 230660 from locality V74111, one anterior abdominal centrum.
Voucher specimens. From Pu1 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276775 from locality V77128, three centra; UCMP 276774 from locality V77128, five centra; UCMP 276773 from locality V77128, eight centra; UCMP 230702 from locality V72210, anterior end of a dentary; UCMP 230701 from locality V72210, one palatine; UCMP 230659 from locality V74111, anterior abdominal centra (not counted); UCMP 230660 from locality V74111, one anterior abdominal centrum.
Localities. From Pu3 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 191574 from locality V73080, three partial dentaries; UCMP 191575 from locality V73080, one palatine; UCMP 191563 from locality V73080, one posterior abdominal centrum; UCMP 191269 from locality V80101, one dentary; UCMP 230727 from locality V75194, one posterior abdominal centrum; UCMP 230727 from locality V75194, one posterior abdominal centrum; UCMP 230687 from locality V81029, one posterior abdominal centrum.
Localities. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: RSM 1594-60, one dentary; RSM 1594-62, one palatine; RSM 1594-26, four centra; UALVP 58817, one anterior abdominal centrum; RSM 1594-27, six posterior abdominal centra; RSM 1594-28, one posterior abdominal centrum; UALVP 58817, centra (not counted).
Description. The dentaries of Esox sp. (Figure 11A) are similar to those of the Cretaceous esocids, Estesesox and Oldmanesox in general shape but differ from those genera and are like extant species of Esox in that the shelf medial to the tooth row is broader and the symphyseal region is reduced. They are like extant species of Esox and to Oldmanesox in having a single row of teeth but differ from the latter genus in that the teeth are relatively larger.
The palatine (Figure 11B) differs from those of the Cretaceous esocids and is similar to the extant species of Esox in having a maxillary process, which is a distinct lateral process near the anterior end of the element for articulation with the maxilla. Grande et al. (2004) recognized the presence of a maxillary process on the palatine is a feature in which extant species of Esox differ from more basal esocids. Also, the shape of the anterior articular surface suggests that it articulated with both the maxilla and premaxilla, a feature in which extant species of Esox differs from more basal esocids (Grande et al., 2004).
Centra of Esox from the lower Paleocene localities were identified through comparison with extant species of the genus as described by Grande et al. (2004) and Sinha et al. (2019). In Esox, abdominal centra have a ventral groove or pit, in contrast with the esocid centra from the Upper Cretaceous, which have a mid-ventral ridge. Thus, the presence of esocid centra from the lower Paleocene with a ventral groove in Pu1 localities (Figure 11C-D) further supports the presence of Esox during that time. Centra in which the groove is bordered by a pair of bulges (Figure 11C-D) are interpreted as anterior abdominal centra. Parapophyseal pits are equal in size to the neural arch articular pits and are located on the side of the centrum, therefore they are not visible in ventral view.
Remarks. The presence of Esox in the Puercan localities is supported by characteristic features of the dentaries, palatine, and centra that were interpreted as diagnostic for the genus by Grande et al. (2004). The dentaries are like those of Esox, in having a reduced symphysis, a single row of large teeth, and broad shelf medial to the tooth row on the dentary. The palatines are like those of Esox in the presence of a maxillary process. Centra are similar to those of extant species of Esox in the presence of a ventral groove on the abdominal centra (Grande et al., 2004; Sinha et al., 2019).
The presence of centra of Esox in Pu1 localities suggests that the genus first appears immediately following the K-Pg mass extinction event. Previously, the earliest record of the genus was E. tiemani, which is represented by an articulated skeleton from the upper Paleocene Paskapoo Formation (Wilson, 1980, 1984). Because Esox is not present in any of the Upper Cretaceous localities examined, its appearance could either be a result of immigration into the area during the Pu1 interval or a rapid diversification event following the K-Pg boundary. The first of these hypotheses might be the most parsimonious given how little time there was following its first appearance after the K-Pg boundary (within the first 70 ka of the Paleocene).
Superorder ACANTHOMORPHA Rosen, 1973 (sensu Stiassny, 1986)
Order PERCOPSIFORMES Berg, 1940
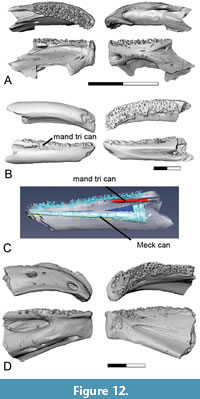
aff. Lindoeichthys Murray et al., 2019
Figure 12A
2005 Acanthomorph dentary, Neuman and Brinkman, p. 176, fig. 9.7E-G
 2010 Acanthomorpha, Larson et al., p. 1167-1168, fig. 6H
2010 Acanthomorpha, Larson et al., p. 1167-1168, fig. 6H
2013 Acanthomorpha gen et sp. indet., Brinkman et al., p. 227, fig. 10.28B
2014 Percopsiformes gen et sp. indet., Brinkman et al., p. 262, fig. 13
2019 Percopsiformes, Murray et al., p. 8, fig. 6
2021 Percopsiformes gen. et sp. indet., Brinkman et al., p. 952-953, fig. 11A-B
Voucher specimens. From Pu1 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276765 from locality V77128, one dentary.
Localities. From Pu3 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 191569 from locality V73080, 10 dentaries.
Localities. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: RSM 1594-59, one dentary; RSM 1594-116, four dentaries.
Description. The presence of a percopsiform in the early Paleocene with affinities to the Late Cretaceous genus Lindoeichthys is documented by dentaries (Figure 12A). As in the dentary of Lindoeichthys described by Murray et al. (2019) and the percopsiform dentaries from the upper Maastrichtian described by Brinkman et al. (2021), the sensory canal is open except for a narrow bridge that forms the posterior border of a single circular pore located at the anterior end of the dentary. The tooth-bearing surface is broad and covered by a shagreen of tiny teeth. The bases for these teeth show little variation in size across the width and length of the tooth-bearing surface.
Remarks. Percopsiforms are represented in the upper Maastrichtian and lower Paleocene by articulated skeletons of two genera, Lindoeichthys from the upper Maastrichtian of Alberta (Murray et al., 2019) and McConichthys from the lower Paleocene of Montana (Grande, 1988). Based on comparison with the dentary of an articulated specimen of Lindoeichthys, the percopsiform affinities of isolated dentaries that had previously been referred to the group by Brinkman et al. (2014) were confirmed. In both the isolated dentaries and the articulated specimen of Lindoeichthys, the dentary is deep, and the sensory canal is open except for a bridge that passes across it near its anterior end. McConichthys differs in having an elongate narrow dentary with the sensory canal located in a groove on the ventral surface of the dentary (Grande, 1988). The presence of percopsiform dentaries similar to those of Lindoeichthys in their proportions and in the presence of an open sensory canal with a bridge passing across it near its anterior end documents the presence of this or a closely related taxon in the lower Paleocene.
In the Puercan localities, dentaries of aff. Lindoeichthys were recovered from both Pu1 and Pu2 intervals. It was not present in assemblages from the Pu3 localities of Montana.
aff. McConichthys Grande, 1988
Figure 12 B-C
Voucher specimens. From Pu3 locality of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 191569 from locality V73080 two dentaries.
Localities. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: RSM 1594-55, one dentary; RSM 1594-118, one dentary.
Description. The presence of a percopsiform with affinities to McConichthys is documented by a dentary that is like that of the type species in being rod-like (Figure 12B-C). The teeth are loosely arranged in rows and show no variation in size across the surface of the tooth-pad. The tooth-pad widens slightly towards the anterior end of the bone. A narrow shelf is present medial to the tooth-pad. The inner surface of the dentary has a broad groove extending partway along the element. The external surface is convex and smooth. The internal structure of the element was investigated using micro-CT scans of specimen UCMP 191569, which allowed the canals within the element to be digitally reconstructed. The Meckelian canal extends for the length of the dentary (Figure 12C: Meck can). A slit-like mandibular trigeminal canal passes under the tooth-pad near the posterior end of the bone (Figure 12B-C: mand tri can). The sensory canal is not enclosed within the dentary.
Remarks. The dentary here referred to aff. McConichthys is included in the Acanthomorpha because the size and arrangement of the tooth bases on the tooth-bearing surfaces is similar to those of aff. Lindoeichthys and “Priscacara”. In its proportions, this dentary is similar to that of the percopsiform McConichthys longipinnis as illustrated by Grande (1988, figure 3) in being elongate and narrow. Also, as in McConichthys the sensory canal is not enclosed within the dentary. However, it differs from the dentary of McConichthys in lacking a groove for the sensory canal on the ventral surface of the element.
Dentaries of aff. McConichthys are present both from the Pu2 Pine Cree Park locality and the one of the Pu3 localities of Montana, V73080.
Order PERCIFORMES Bleeker, 1859
Suborder ?MORONOIDEI Smith and Craig, 2007 (sensu Whitlock, 2010)
Family ?MORONIDAE Jordan, 1923 (sensu Whitlock, 2010)
Genus “PRISCACARA” Cope, 1877
“Priscacara” sp.
Figure 12D
2014 Priscacara, Brinkman et al., p. 262, fig. 14
2021 Priscacara, Brinkman et al., p. 953, fig. 11C-D
Voucher specimens. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: RSM 1594-57, one dentary; RSM 1594-115, six dentaries; UALVP 58821, one dentary.
Description. The tooth-bearing surface of the dentaries (Figure 12D) is covered by a pad of tiny teeth showing little variation in the size of the tooth-bases across the surface. The surface is widest near the symphysis and narrows posteriorly. The mandibular sensory canal is enclosed in a bony tube. Two relatively large pores for the sensory canals are present near the ventral edge of the dentary. A large depression is present on the lateral surface above the sensory canal in the posterior half of the dentary. The length of this depression is about equal to the length of the anterior toothed portion of the dentary.
Remarks. The presence of “Priscacara” in the lower Paleocene is documented by dentaries from the Pine Cree locality. We here refer to “Priscacara” in scare quotes because of the concern over the monophyly of the genus expressed by some authors, who consider “Priscacara” to be a form genus uniting various superficially similar fishes rather than a natural taxon (see Grande, 2001). Furthermore, we tentatively place “Priscacara” in the Moronidae because this classification was proposed relatively recently by Whitlock (2010). However, we acknowledge the considerable uncertainty over the taxonomic placement of this potentially polyphyletic genus (see Brinkman et al., 2021 for details). Of most relevance to this matter, “Priscacara” has been generally accepted as a perciform since its initial description (Cope, 1877).
“Priscacara” was reported in the Hell Creek Formation of Montana by Brinkman et al. (2014, 2021) based on dentaries that matched those of articulated specimens of “Priscacara” from the Green River Formation. Subsequently, isolated dentaries of “Priscacara” from the lower Eocene Wasatch Formation were described by Divay and Murray (2016a) providing additional data on the morphology of the dentary in this genus that supports the identification of the Hell Creek specimens. “Priscacara” dentaries from the Pine Cree Park locality differ little from specimens from the Hell Creek and Wasatch formations, indicating that this taxon continued through the K-Pg mass extinction event, although it appears to have been locally extirpated from the study area during the Pu1 interval.
Acanthomorpha indet.
Figure 13B-C
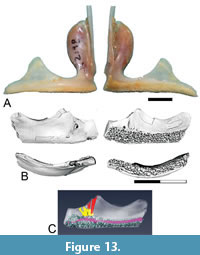 Voucher specimens. From Pu3 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 191569 from locality V73080 one premaxilla; UCMP 230726 from locality V75194, one premaxilla.
Voucher specimens. From Pu3 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 191569 from locality V73080 one premaxilla; UCMP 230726 from locality V75194, one premaxilla.
Localities. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: RSM 1594-50, one premaxilla; RSM 1594-51, one premaxilla; RSM 1594-52, one premaxilla; RSM 1594-53, one premaxilla; RSM 1594-54, one premaxilla; RSM 1594-118, six premaxillae; UALVP 58820, one premaxilla.
Description. Flattened tooth-bearing elements that bear a pad of teeth that are loosely arranged in longitudinal rows and show no variation in size across the surface of the tooth-pad similar to the acanthomorph dentaries described above are tentatively identified as premaxillae of an acanthomorph (Figure 13B-C). This identification is based on a similarity to the premaxilla of Holocentrus rufus (Figure 13A). The element is plate-like and, seen in lateral view, the occlusal edge is straight, and the dorsal edge is concave, only preserving the broken base of the postmaxillary process in its posterior end, and an anterior dorsal protuberance, which, based on comparison with the premaxilla of Holocentrum rufus, would have been the base of a single anterior process that would have divided to form the ascending and articular processes.
The tooth-pad forms a broad surface along the straight edge of the element, with a broad flat surface above this. As with the dentary, the tooth-pad is slightly wider towards the anterior end of the bone. In occlusal view, the element is strongly curved. In lateral view, a pair of foramina is present near the anterior end of the bone. To understand the significance of these foramina, micro-CT scans of specimen UALVP 58820 were undertaken and the internal canals reconstructed (Figure 13C). The foramina are openings of canals that pass through the anterior end of the element (shown in yellow in Figure 13C). An additional canal extends along the length of the tooth row, with branches going to the base of the teeth (shown in pink in Figure 13C), likely for the rostral/maxillary branch of the trigeminal nerve. A pair of canal-like spaces also extend dorsally near the anterior end of the premaxilla (shown in red in Figure 13C). These closely approach but are not connected to the canals that pass through the end of the dentary, so are likely spaces within the bone, rather than canals through which vessels or nerves passed. Except for these canals and spaces, the element is solid in cross section.
Remarks. The tooth-bearing element tentatively identified as a premaxilla is thought to be from an acanthomorph because the size and arrangement of teeth is like that of the acanthomorph dentaries described above. The possibility that this element is a dentary is rejected because a Meckelian canal is absent. The identification of the elements as a premaxilla is based on the curvature of the element as seen in occlusal view, the presence of preserved bases for the ascending and articular processes anteriorly, and postmaxillary process posteriorly, and the absence of a Meckelian canal.
Tooth-bearing elements similar to these premaxillae are absent in Cretaceous localities. Within the lower Paleocene, these elements are present only in the Pu2 and Pu3 intervals but are more abundant in the Pu2 Pine Cree Park locality than in the Tullock localities. Thus, they are likely associated with one of the acanthomorphs present in that locality. However, the variation in morphology of premaxillae in acanthomorphs is not well documented, so it is not possible to associate these with centra or dentaries also present in the locality.
Acanthomorph Centrum Morphotypes
Acanthomorph centra have long been recognized as morphologically distinct from other teleosts in their possession of a suite of derived characters mainly related to the articulation of the first vertebral centrum with the skull, and of more posterior abdominal centra with one another. The first centrum has a tripartite anterior articular surface with separate surfaces for the basioccipital and exoccipitals. Zygapophyseal articulations are present between the centra and in the anterior region of the vertebral column the ribs articulate on the lateral surface of the neural arch rather than lower on the centrum itself (Rosen and Patterson, 1969; Grande, 1988). A survey of the morphology of the first abdominal centrum in extant acanthomorphs (Murray and Brinkman, 2023) demonstrated that this element is highly variable in extant acanthomorphs and confirmed that it can provide reliable information on diversity of acanthomorphs within an assemblage. Because acanthomorph centra are abundant in the Puercan localities and distinct morphotypes can be recognized, they provide independent evidence regarding the diversity of acanthomorphs in these assemblages.
Acanthomorph centrum type HC-1
Figure 14
2014 Acanthomorph centrum type HC-1, Brinkman et al., p. 263-264, fig. 15C
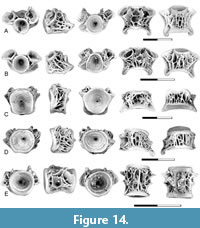 2021 Acanthomorph centrum type HC-1, Brinkman et al., p. 954-955, fig. 12
2021 Acanthomorph centrum type HC-1, Brinkman et al., p. 954-955, fig. 12
Voucher specimens. From Pu1 locality of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276745 from locality V77128, two first abdominal centra.
Localities. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: RSM 1594-47, one first abdominal centrum; RSM 1594-48; one first abdominal centrum; RSM 1594-49, one first abdominal centrum; RSM 1594-84, 52 first abdominal centra; RSM 1594-18, one anterior abdominal centrum; RSM 1594-19, one mid abdominal centrum; RSM 1594-20, one mid abdominal centrum; RSM 1594-21, one anterior abdominal centrum; RSM 1594-22, one mid abdominal centrum; RSM 1594-87, 807 mid abdominal centra; RSM 1594-88, (uncounted) anterior abdominal centra; RSM 1594-89, 547 abdominal centra.
Description. Acanthomorph centrum type HC-1 is characterised by the presence of a first abdominal centrum with the following features: 1) the exoccipital articular surfaces do not contact one another above the basioccipital articular surface; 2) the neural arch is fused to the centrum; 3) a network of fibers is present on the dorsal surface of the centrum between the bases of the neural arch; and 4) a distinct mid-ventral pit is absent (Figure 14A-B). The sides of the centrum are formed by a coarse network of interconnected ridges that are mainly oriented longitudinally.
The more posterior abdominal centra of acanthomorph centrum type HC-1 (Figure 14C-E) share with the first centrum the presence of a network of bone fibers extending between the ends of the centrum dorsally between the neural arch bases. Also, as with the first abdominal centrum, a mid-ventral pit is absent or weakly developed. Anterior abdominal centra type HC-1 are short and have particularly well-developed zygapophyses (Figure 14C-D). The abdominal centra from a more posterior position are longer and have a distinct rib articular pit on the side of the centrum (Figure 14E).
Remarks. Acanthomorph centrum type HC-1 was first recognized in the upper Maastrichtian by Brinkman et al. (2014) based on centra from the Hell Creek Formation. Subsequently this morphology was also reported from the Lance and Scollard formations (Brinkman et al., 2021). Variation is present in the relative length of the first abdominal centrum; specimens from the lower Paleocene are relatively short compared to specimens from the Lance Formation, although whether this variation is indicative of anagenesis is unclear at this time. In other features, they are indistinguishable.
The first abdominal centrum of acanthomorph type HC-1 is similar to centra from the Eocene Wasatch Formation of Wyoming and from the Eocene-Oligocene Cypress Hills Beds of southwestern Saskatchewan identified as aff. Amblyopsidae by Divay and Murray (2015, figure 6F; 2016a, figure 6A-C), and thus are likely from this or a related group of Percopsiformes. They are also like the first abdominal centrum of Percopsiformes in that the neural arch is fused to the centrum, one of the features that Divay and Murray (2015, 2016a) considered diagnostic of the group.
Acanthomorph centrum type HC-1 is present in both the Pu1 and Pu2 intervals but does not occur in the Pu3 localities of Montana. However, because it is likely from a member of the Amblyopsidae, which are extant percopsiform fishes, we can infer that these were present through the entire Paleocene. It is rare in the Pu1 interval, with only two first abdominal centra present in localities of this age. In contrast, it is the most abundant kind of teleost centrum in the Pine Cree Park locality (66%; Table 1).
Acanthomorph centrum type HC-2
Figure 15
2014 Acanthomorph first abdominal centrum type HC-2, Brinkman et al., p. 263-264, fig. 15A
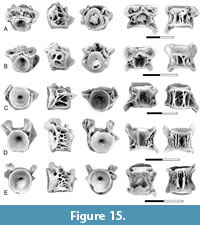 2021 Acanthomorph first abdominal centrum type HC-2, Brinkman et al., p. 955, fig. 13
2021 Acanthomorph first abdominal centrum type HC-2, Brinkman et al., p. 955, fig. 13
Voucher specimens. From Pu1 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276746 from locality V77128, approximately 146 first abdominal centra; UCMP 276747 from locality V77128, two first abdominal centra; UCMP 276748 from locality V77128, two first abdominal centra; UCMP 276733 from locality V77128, seven first abdominal centra; UCMP 276734 from locality V77128, one first abdominal centrum; UCMP 276734 from locality V77128, one first abdominal centrum; UCMP 276749 from locality V77128, approximately 96 abdominal centra; UCMP 276750 from locality V77128, two abdominal centra; UCMP 276751 from locality V77128, two abdominal centra; UCMP 230643 from locality V74111, one large first abdominal centrum; UCMP 230644 from locality V74111, one large first abdominal centrum; UCMP 230645 from locality V74111, one small first abdominal centrum; UCMP 230646 from locality V74111, one small first abdominal centrum; UCMP 230647 from locality V74111, (uncounted) large abdominal centra; UCMP 230647 from locality V74111, one large abdominal centrum; UCMP 230649 from locality V74111, (uncounted) small abdominal centra; UCMP 230650 from locality V74111, one small abdominal centrum; UCMP 230703 from locality V72210, (uncounted) first abdominal centra; UCMP 230704 from locality V72210, (uncounted) abdominal centra.
From Pu3 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 191571 from locality V73080, approximately 67 first abdominal centra; UCMP 191570 from locality V73080, approximately 125 abdominal centra; UCMP 230693 from locality V81029, (uncounted) abdominal centra of small size; UCMP 230694 from locality V81029, (uncounted) abdominal centra of small size; UCMP 230615 from locality V81029, (uncounted) abdominal centra of small size; UCMP 230696 from locality V81029, one first abdominal centrum of small size; UCMP 230697 from locality V81029, (uncounted) abdominal centra of large size; UCMP 230698 from locality V81029, (uncounted) abdominal centra of large size; UCPM 230699 from locality V81029, (uncounted) abdominal centra of large size; UCMP 230732 from locality V74194, (uncounted) abdominal centra; UCMP 230733 from locality V74194, (uncounted) abdominal centra; UCMP 320795 from locality V74194, (uncounted) abdominal centra.
Localities. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: RSM 1594-37, one first abdominal centrum; RSM 1594-38, one first abdominal centrum; RSM 1594-39, one first abdominal centrum; RSM 1594-40, one first abdominal centrum; RSM 1594-41, one first abdominal centrum; RSM 1594-141, 45 first abdominal centra; RSM 1594-12, one mid abdominal centrum; RSM 1594-13, one mid abdominal centrum; RSM 1594-14, one mid abdominal centrum; RSM 1594-15, one mid abdominal centrum; RSM 1594-16, one mid abdominal centrum; RSM 1594-17, one mid abdominal centrum; RSM 1594-86, 111 abdominal centra; UALVP 58810, (uncounted) first abdominal centra; UALVP 58811, (uncounted) abdominal centra.
Description. Acanthomorph centrum type HC-2 was described by Brinkman et al. (2013) based on centra from the Hell Creek Formation. Both first abdominal and more posterior abdominal centra of type HC-2 are present in the Puercan localities examined here.
In the first abdominal centrum of acanthomorph centrum type HC-2 from the Paleocene localities, as in specimens from the Hell Creek Formation, the exoccipital articular surfaces contact one another above the basioccipital articular surface. As in acanthomorph centrum type HC-1 the neural arches are fused to the centrum, but rather than a network of bone, a mid-dorsal pit is present between the bases of the neural arches (Figure 15A-B). The sides of the centrum are formed by a network of relatively coarse bone ridges generally oriented antero-posteriorly between the ends of the centrum with smaller ridges connecting these. A distinct mid-ventral pit is present, typically bordered by enlarged bony ridges.
More posterior abdominal centra referred to acanthomorph centrum type HC-2 share with the first centrum the presence of mid-dorsal and mid-ventral pits with the mid-ventral pit bordered by distinct ridges (Figure 15C-E). Also, as with the first centrum, the sides of these centra are formed by a relatively coarse network of bony ridges with larger ridges extending between the ends of the centrum and smaller ridges connecting these. Variation is present in the development of ridges on the side of the centrum. Some have relatively few, robust ridges that are widely spaced (Figure 15C). Others have ridges that are distinctly finer and more closely spaced, resulting in numerous small pores evenly distributed along the side of the centrum (Figure 15D-E). These variations are likely of taxonomic importance, but intermediates are present, therefore distinct morphotypes are not recognized here.
Remarks. Acanthomorph centrum type HC-2 is the most abundant teleost morphotype in all the Tullock Member localities examined (Table 1). Similar centra from the Wasatch Formation were attributed by Divay and Murray (2016a) to aff. Centrarchidae or “Priscacara.” Thus, these centra may be associated with the dentaries of “Priscacara” described above. Centrum type HC-2 is different from these Eocene taxa from the Wasatch in lacking the well-defined, rounded dorsal articular pits for the neural arches, which are very distinctive in the Eocene specimens (Divay and Murray, 2016a, figures 7C-D, 9A). Therefore, even if correctly associated with the dentaries referred to “Priscacara”, this taxon is likely distinct from the Eocene taxon.
Acanthomorph indet, type HC-4
Figure 16
2014 Acanthomorph centrum type HC-4, Brinkman et al., p. 264, fig. 15E
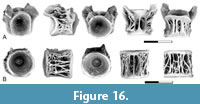 2021 Acanthomorph centrum type HC-4, Brinkman et al., p. 955-957, fig. 15
2021 Acanthomorph centrum type HC-4, Brinkman et al., p. 955-957, fig. 15
Voucher specimens. From Pu3 locality in the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 191570 from locality V73080, three centra (centra of acanthomorph HC-2 also included in this catalogue number).
Description. Acanthomorph indet, type HC-4 was recognized by Brinkman et al. (2014) based on abdominal centra that had a single well-developed mid-dorsal ridge extending between the ends of the centrum. Later, a first abdominal centrum from the upper Maastrichtian Lance Formation of Wyoming was tentatively associated with the abdominal centra (Brinkman et al., 2021), but first abdominal centra of this morphology were not recovered in the Puercan localities, where acanthomorph centra type HC-4 are only represented by more posterior abdominal centra (Figure 16). As in the Cretaceous specimens described by Brinkman et al. (2014, 2021), in specimens of HC-4 from the Puercan localities a well-developed mid-dorsal ridge extends between the ends of the centrum. Variation is present in the thickness of the ridge, with some specimens having a robust ridge that expands at its posterior end (Figure 16A) and others having a slenderer ridge that shows no distinct expansion (Figure 16B). This likely reflects variation along the vertebral column, with the more robust ridge being present in more anterior centra. As in centrum type HC-1, a mid-ventral pit is poorly developed or absent.
Remarks. Acanthomorph indet, type HC-4 has a distinctive morphology that cannot be directly associated with any specific group of extant acanthomorphs and may therefore represent an archaic group that is restricted to the Late Cretaceous and early Paleocene. It appears to be broadly similar to the morphology of percomorphs and may therefore represent a related taxon. However, this taxonomic affiliation is only tentative, and, as before (Brinkman et al., 2021), we leave this type as representing an indeterminate acanthomorph. Centrum type HC-4 is present in both the Hell Creek Formation and the Puercan-age localities of the Tullock Member, but it is rare in both intervals (Table 1).
Acanthomorph indet. type HC-5
Figure 17
2021 Acanthomorph centrum type HC-5, Brinkman et al., p. 957-958, fig. 16
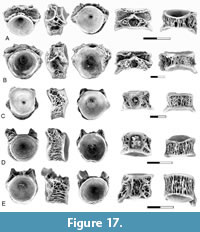 Voucher specimens. From Pu1 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276752 from locality V77128, 12 first abdominal centra; UCMP 276753 from locality V77128, two first abdominal centra; UCMP 276754 from locality V77128, two first abdominal centra; UCMP 276755 from locality V77128, 57 abdominal centra; UCMP 276756 from locality V77128, two abdominal centra; UCMP 276757 from locality V77128, two abdominal centra; UCMP 230644 from locality V74111, two first abdominal centra; UCMP 230646 from locality V74111, one first abdominal centrum; UCMP 230648 from locality V74111, three abdominal centra.
Voucher specimens. From Pu1 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276752 from locality V77128, 12 first abdominal centra; UCMP 276753 from locality V77128, two first abdominal centra; UCMP 276754 from locality V77128, two first abdominal centra; UCMP 276755 from locality V77128, 57 abdominal centra; UCMP 276756 from locality V77128, two abdominal centra; UCMP 276757 from locality V77128, two abdominal centra; UCMP 230644 from locality V74111, two first abdominal centra; UCMP 230646 from locality V74111, one first abdominal centrum; UCMP 230648 from locality V74111, three abdominal centra.
Description. Acanthomorph indet. type HC-5 was recognized by Brinkman et al. (2021) based on both the first and more posterior abdominal centra. Both regions of the vertebral column are represented in the Puercan samples. The first abdominal centrum (Figure 17A-B) is similar to that of acanthomorph centrum type HC-2 in that the exoccipital articular facets meet above the anterior articular surface for the basioccipital, the neural arches are fused to the centrum, and in that a deep mid-dorsal pit extends between the bases of the neural arches. It differs in being distinctly shorter, in that the lateral and ventral surfaces of the centrum are formed by numerous fine ridges of bone extending longitudinally between the ends of the centrum, and in the absence of a mid-ventral pit. More posterior abdominal centra referred to Acanthomorph indet. type HC-5 (Figure 17C-E) share with the first abdominal centrum the presence of numerous fine ridges of bone extending between the ends of the centrum and the presence of a weakly developed or absent mid-ventral pit. The ridges bordering the mid-ventral pit, when present, are not significantly more robust than those on the lateral surfaces of the centrum (Figure 17C-D). Variation is present in the length of the centrum, with more anterior centra being similar to the first abdominal centrum in being very short (Figure 17B-C). More posterior abdominal centra are about as long as they are wide (Figure 17E).
Remarks. Acanthomorph indet. type HC-5 is distinct in the foreshortening of the first few abdominal centra and in the absence or weak development of a mid-ventral pit. Centra of acanthomorph indet. type HC-5 are present but rare in the upper Maastrichtian Hell Creek, Lance, and Scollard formations (Brinkman et al., 2021). In the lower Paleocene, they are restricted to two Pu1 localities, where they sometimes occur in high specimen abundances (UCMP loc. V77128; Table 1).
Acanthomorph indet. type HC-5 show a significant size range, with the diameter of the posterior end of the largest centra exceeding 5 mm, making it one of the largest teleost fish in the Puercan localities of Montana and Saskatchewan.
Affiliations of acanthomorph indet. type HC-5 are unknown. It is unlikely that Acanthomorph indet. type HC-5 are those of aff. McConichthys because they have a different stratigraphic distribution: Acanthomorph indet. type HC-5 are restricted to the upper Maastrichtian and Pu1 intervals, while dentaries of aff. McConichthys were only recovered in the Pu2 and Pu3 intervals. Acanthomorph indet. type HC-5 is broadly similar to the morphology of percomorphs, but none of the extant and fossil taxa we have examined correspond to the finer details of this morphology or to its most distinctive characteristics, such as the greatly shortened anterior abdominal centra. It is therefore likely that Acanthomorph indet. type HC-5 is from an unidentified percomorph.
Acanthomorph indet. type AvE
Figure 18
2019 Acanthomorph centrum type AvE, Brinkman, p. 141, fig. 20
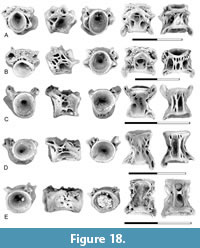 Voucher specimens. From Pu1 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276742 from locality V77128, eight first abdominal centra; UCMP 276743 from locality V77128, one first abdominal centrum; UCMP 276744 from locality V77128, two first abdominal centra; UCMP 276736 from locality V77128, approximately 81 abdominal centra; UCMP 230642 from locality V74111, 16 centra.
Voucher specimens. From Pu1 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276742 from locality V77128, eight first abdominal centra; UCMP 276743 from locality V77128, one first abdominal centrum; UCMP 276744 from locality V77128, two first abdominal centra; UCMP 276736 from locality V77128, approximately 81 abdominal centra; UCMP 230642 from locality V74111, 16 centra.
Localities. From the Pu2 Pine Cree Park locality, Ravenscrag Formation, Saskatchewan: RSM 1594-42, one first abdominal centrum; RSM 1594-43, one first abdominal centrum; RSM 1594-44, one first abdominal centrum; RSM 1594-45, one first abdominal centrum; RSM 1594-46, one first abdominal centrum; RSM 1594-85, nine first abdominal centra; RSM 1594-23, one abdominal centrum; RSM 1594-24, one abdominal centrum; RSM 1594-25, one abdominal centrum; RSM 1594-90, 22 abdominal centra.
Description. Acanthomorph indet. type AvE was recognized by Brinkman (2019) based on centra from the upper Campanian Belly River Group of Alberta. The first abdominal centrum of acanthomorph type AvE is similar to type HC-2 in that the exoccipital articular surface meet above the surface for the basioccipital, the neural arches are fused to the centrum, a deep pit is present between the base of the neural arches, and a well-developed mid-ventral pit bordered by robust ridges is present. It differs from type HC-2 and all other centrum morphotypes recognized here in the presence of deep lateral pits bordered anteriorly by a buttress that extends laterally from the anterior articular surface (Figure 18A-B).
Abdominal centra of type AvE (Figure 18C-E) differ from those of types HC-1 and HC-2 in being more elongate and in having relatively smooth lateral and ventral surfaces.
Remarks. Acanthomorph type AvE was recognized based on first abdominal centra with distinctive pits in the lateral surface of the centrum bordered anteriorly by a buttress that extends laterally from the anterior edge of the centrum (Brinkman, 2019). Its presence in the Puercan is documented by centra showing this feature. The lower Paleocene specimens referred to Acanthomorph indet. type AvE differ from the upper Campanian specimens in that the articular surfaces for the exoccipitals meet above the articular surface for the basioccipital.
In the Upper Cretaceous, Acanthomorph indet. type AvE is present only in the upper Campanian Belly River Group of Alberta (Brinkman, 2019). Thus, although the taxon represented by these centra is not known from the upper Maastrichtian Hell Creek Formation, that Acanthomorph indet. type AvE was recovered in our Puercan samples is evidence that it extended across the K-Pg boundary.
Because no dentaries can be associated with Acanthomorph indet. type AvE, this centrum morphology is considered evidence for a distinct taxonomic unit, referred to as acanthomorph indet. type AvE, in faunal lists (Brinkman, 2019; this study). The affinities of the taxon represented by this morphology are difficult to determine more precisely. Similar first centra, with lateral pits and smooth bone surfaces, were recovered from the Wasatch Formation and attributed to an indeterminate centrarchid or “Priscacara” by Divay and Murray (2016a, figure 9A), but these differed from acanthomorph indet. type AvE in having a pair of distinctive, circular dorsal pits. However, because of distinctive similarities in all other features, it is likely that the taxon represented by acanthomorph indet. type AvE centra was also an unidentified perciform.
Centra from Teleosts of Uncertain Taxonomic Affiliations
As in vertebrate microfossil assemblages from elsewhere within the Western Interior, centra and tooth-bearing elements are present that are taxonomically distinct but currently cannot be identified below the level of Teleostei. Despite their uncertain status, they help characterize the vertebrate assemblages preserved at the individual vertebrate microfossil localities and the geographic and stratigraphic distribution patterns that are present. These elements are described here as distinct morphotypes and are treated as distinct operational taxonomic units.
Teleost indet. type U-4
Figure 19
2013 Genus et sp. indet. type U-4, Brinkman et al., p. 221, fig. 20.23
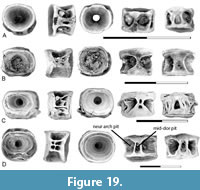 2014 Genus and species indet. U-4, Brinkman et al., p. 259-260, fig. 10A-B
2014 Genus and species indet. U-4, Brinkman et al., p. 259-260, fig. 10A-B
2019 Teleost centrum morphotype U-4, Brinkman, p 145, fig. 23
2021 Genus and species indet. U-4, Brinkman et al., p. 959-960, fig. 18
Voucher specimens. From Pu1 localities of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276758 from locality V77128, five abdominal centra; UCMP 276759 from locality V77128, one abdominal centrum; UCMP 276760 from locality V77128, one abdominal centrum; UCMP 230709 from locality V72210, one abdominal centrum; UCMP 230654 from locality V74111, two abdominal centra.
Localities. From Pu3 locality of the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP191563 from locality V73080, two centra.
Description. Teleost indet. type U-4 is represented by centra that are small, simple spools with deep neural arch and parapophyseal pits and a shallowly excavated to nearly flat anterior surface (Figure 19). Centra are wider than high and may appear sub-rectangular in anterior view (Figure 19C). Neural arch articular pits are closely spaced. In some specimens, these pits are separated by a robust bar of bone (Figure 19A-B), whereas in others, a single, well-defined mid-dorsal pit is present (Figure 19D). Parapophyseal pits are large, oval pits about equal in size to the neural arch articular pits. Occasionally, the parapophyses are preserved in place (Figure 19C). A mid-ventral pit is generally present (Figure 19A, C-D).
Remarks. Teleost indet. type U-4 is widespread in the Cretaceous, occurring in the Cenomanian to Campanian of Utah (Brinkman et al., 2013), the Santonian to upper Campanian of Alberta (Brinkman et al., 2017), and the upper Maastrichtian Hell Creek, Lance, and Scollard formations (Brinkman et al., 2021), all formations of the Western Interior. In the Puercan localities examined here, it was present in both the Pu1 and Pu3 intervals of Montana but was not observed in the Pine Cree Park locality of Saskatchewan. No dentaries can be associated with the centra of teleost indet. type U-4. The taxonomic relationships of the fish represented by teleost indet. type U-4 centra are unknown, although this distinctive morphology is very different to that of the extant teleosts we have examined, suggesting that this is an archaic and extinct taxon.
Teleost indet. type PT-1
Figure 20
2021 Teleost indet. type PT-1, Sinha et al., p. 6, fig. 6.
 Voucher specimens. From Pu1 localities in the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276761 from locality V77128, approximately 32 abdominal centra; UCMP 276762 from locality V77128, three abdominal centra; UCMP 276763 from locality V77128, three abdominal centra; UCMP 230712 from locality V72210, five centra; UCMP 230651 from locality V74111, seven centra; UCMP 230652 from locality V74111, one centum.
Voucher specimens. From Pu1 localities in the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 276761 from locality V77128, approximately 32 abdominal centra; UCMP 276762 from locality V77128, three abdominal centra; UCMP 276763 from locality V77128, three abdominal centra; UCMP 230712 from locality V72210, five centra; UCMP 230651 from locality V74111, seven centra; UCMP 230652 from locality V74111, one centum.
Localities. From Pu3 localities in the Tullock Member of the Fort Union Formation, Garfield County, Montana: UCMP 230728 from locality V75194, one centrum. UCMP 191565 from locality V73080, two centra.
Description. Teleost indet. type PT-1 is represented by centra that have multiple ridges of bone extending between the ends of the centrum, which are interconnected with a multitude of transverse ridges, giving a fine, lacy texture to the surface of the centrum (Figure 20). Neural arches and parapophyses are fused to the centrum. On the anterior centra, the neural arch extends for the full length of the centrum (Figure 20A). On more posterior abdominals, the neural arch is restricted to the anterior end of the centrum (Figure 20B-E). The parapophyses are blunt, laterally directed processes located at mid height near the anterior end of the centrum. The anterior-most centrum recovered here (Figure 20A) is without distinct pits dorsally or ventrally. On the more posterior centra, a slit-like mid-dorsal pit is generally present, extending for the length of the centrum between the neural arches. Ventrally, a pair of elongate pits separated by a narrow mid-ventral ridge is present. In end view the centrum is round. Centra from an anterior position along the vertebral series (Figure 20A-B) are short and have a flat or slightly excavated anterior end. More posterior centra are more elongate and have a more distinctly excavated anterior end.
Remarks. Teleost indet. type PT-1 was recognized by Sinha et al. (2021) based on centra from the late Paleocene (Tiffanian) beds of the Ravenscrag Formation of Saskatchewan, Canada. Centra of teleost indet. type PT-1 are easily recognizable because of the lacy texture of the lateral surface of the centrum and the flattened anterior end. They were interpreted as evidence for a distinct kind of teleost fish, rather than a regional variant from along the vertebral column of another teleost present in the Ravenscrag assemblage, because the variation in the lengths of the centra and in the positions of the parapophyses indicate that multiple regions of the column are present. Specimens from the Puercan localities described here show a similar range of variation that can be attributed to variation along the vertebral column. Relationships of the teleost represented by these centra are unknown.
Teleost indet. type PT-1 is present in localities of both Pu1 and Pu3 age in Montana but does not occur in the Pine Cree locality of Saskatchewan. In Montana, it is abundant in the Pu1 interval and rare in the Pu3 interval (Table 1). The morphology of teleost centrum indet. type PT-1 is different from that of all of our comparative material, and it has no clear Late Cretaceous precursors. Thus, it was likely an immigrant of unknown taxonomic affinity, which entered the area after the K-Pg mass extinction event.
RESULTS
Patterns of Teleost Richness and Extinction Across the K-Pg Boundary
A general understanding of the changes in taxonomic richness and patterns of extinction across the K-Pg boundary and through the subsequent lower Paleocene can be obtained by comparing the assemblages present in the Lancian and the successive Puercan subintervals. The stratigraphic distribution of the teleost fishes of the early Paleocene described here, along with assemblages from the late Maastrichtian and late Paleocene (Tiffanian NALMA) intervals, is shown in Table 2. Twenty operational taxonomic units (OTUs) were recognized in the upper Maastrichtian by Brinkman et al. (2021). In addition, two OTUs that were not recovered in Maastrichtian assemblages are inferred to have been present during this time based on their occurrences in older and younger intervals (i.e., range-through occurrences). These are the clupeomorph Diplomystus and acanthomorph indet. type AvE. These are inferred to be present during the upper Maastrichtian because they are present in both the upper Campanian Belly River Group of Alberta (Brinkman, 2019) and localities of Pu1 and Pu2 age (this study). With the inclusion of these OTUs, a minimum of 22 taxa would have been present in the late Maastrichtian of the northern Western Interior.
In the localities sampled here, 10 OTUs are present in the Pu1 interval. Six of the 22 late Maastrichtian OTUs listed in Table 2 do not extend into the Paleocene, giving an extinction rate of 27%. These non-survivors are the indeterminate elopiform, the large-bodied hiodontid, both species of Wilsonichthys, acanthomorph type HC-3, and teleost indet. type RvB. Five of these are rare in the upper Maastrichtian formations, each being represented by fewer than 10 specimens in a sample of more than 1,900 specimens from 10 localities spread across Wyoming, Montana, and southern Alberta (Brinkman et al., 2021, table 2). These rare taxa are the elopiform (represented by two specimens), the large hiodontid (represented by four specimens), teleost indet. type RvB (represented by five specimens), and acanthomorph type HC-3 (represented by nine specimens). The loss of these rare taxa is consistent with McKinney’s (1997) observation that rarity is a good predictor of extinction across multiple time scales.
The decrease in richness resulting from these extinctions is countered by the first appearances of two new taxa during the Pu1. One of these, Esox, is a member of a group present in the Cretaceous, the Esocidae, so its origin may be a result of in situ speciation during the earliest Paleocene (Pu1). The second, teleost indet. type PT-1, is of unknown relationships and has no clear Cretaceous precursors, so it likely immigrated into northeastern Montana. In addition, a minimum of eight other taxa based on centra can be inferred to be present during the Pu1 interval based on their occurrence in older and younger beds (taxa based on dentaries such as Ostariostoma and “Priscacara” are not included in this estimate of diversity because it is likely that they are also represented by OTUs based on centra). These range-through taxa are Phyllodus, Coriops, aff. Coriops/Lopadichthys, Diplomystus, gonorynchiform indet. type H, acanthomorph indet. type HC-4, acanthomorph indet. type HC-6, and at least one member of the Hiodontidae. The first six of these are present in both the late Maastrichtian and Pu2 and/or Pu3 subintervals. Hiodontids have not yet been recovered from Paleocene localities, but a member of the Hiodontidae can be inferred to have been present because this extant family first appears in the Cretaceous. Acanthomorph indet. type HC-6 is a range-through taxon because it has similarities to Mioplosus, an acanthomorph from the early Eocene Green River Formation (Brinkman et al., 2021). With the addition of these taxa, 18 teleost taxa would have been present during the Pu1 interval. Thus, although the extinction rate at the end of the Cretaceous is 27%, taxonomic richness of the Pu1 assemblage (including range-through taxa) is only 18% lower than late Maastrichtian levels.
The teleost assemblage from the single Pu2 locality sampled here, the Pine Cree Park locality, includes 13 OTUs, six of which are acanthomorphs. However, three of the acanthomorphs are represented only by jaw elements, and three are represented only by centra. Because it is possible that the jaw elements and centra are associated, together these elements are considered evidence for only three kinds of acanthomorphs. Thus, a minimum of 10 taxa are present in the Pu2 subinterval. Three of the taxa that crossed the K-Pg boundary, Otophysi indet. type U3/BvD (K morph), Estesesox, and acanthomorph indet. type HC-5, did not extend into the Pu2 subinterval. Of all taxa recovered in our full sample, none have their first appearance in the Pu2 subinterval. However, seven taxa can be inferred to have been present during this time based on their occurrences in younger and older beds. These range-through taxa are Phyllodus, Coriops, a member of the Hiodontidae, acanthomorph indet. type HC-4, acanthomorph indet. type HC-6, teleost indet. type U-4, and teleost indet. type PT-1. With the addition of these taxa, teleost assemblages of the Pu2 interval would have included a minimum of 17 taxa. Thus, if these range-through taxa are included, the taxonomic richness of the Pu2 interval is slightly less than that of the Pu1 interval.
Teleost assemblages from the localities of Pu3 age include 11 OTUs. These include four acanthomorphs, two of which are represented by jaw elements and two by vertebral elements. Because the jaw elements may be associated with the vertebral elements, these elements are considered evidence for only two kinds of acanthomorphs for the estimates of taxonomic richness. Thus, a minimum of nine OTUs are present in the three Pu3 localities that were sampled. Three of the Cretaceous OTUs that are present in the Pu2 locality do not extend into the Pu3 or younger intervals. These are Ostariostoma, teleost indet. type H, and acanthomorph indet. type AvE. As a result, only nine of the 15 Cretaceous OTUs that crossed the K-Pg boundary extended into the Pu3 subinterval for a total loss of 56% of late Maastrichtian taxa prior to the Pu3 interval. No OTUs recovered in our total sample first appeared in the Pu3 subinterval. However, five taxa can be inferred to have been present during this time based on their occurrence in older and younger beds. These are Diplomystus, Notogoneus, acanthomorph indet. type HC-1, acanthomorph indet. type HC-6, and a member of the Hiodontidae. With the addition of these range-through taxa, a minimum of 14 taxa would have been present during the Pu3 subinterval, which is a lower taxonomic richness than the 18 and 17 taxa of the Pu1 and Pu2 subintervals, respectively. Thus, extinction/extirpation outpaced teleost diversification/immigration during the early Paleocene of the northern Western Interior of North America following the end-Cretaceous mass extinction.
A more precise understanding of the changes in taxonomic richness through the late Maastrichtian and early Paleocene are obtained by comparing the local fossil assemblages from the Hell Creek Formation and Tullock Member of Garfield County, Montana. These localities have been placed within a high-resolution chronostratigraphic framework based on their position relative to the K-Pg boundary (e.g., LeCain et al., 2014; Moore et al., 2014; Sprain et al., 2015, 2018; Weaver et al., 2022). To alleviate any potential taphonomic bias introduced by including counts of teeth and tooth-bearing elements in this analysis, only OTUs represented by centra were included in the analysis.
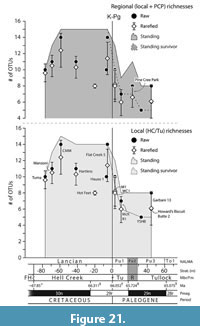 As shown in the bottom graph in Figure 21, raw teleost richness fluctuates from between eight and 14 OTUs through the Hell Creek Formation. Raw richness peaks at 14 OTUs in both the lower and upper thirds of the Hell Creek at locality CMM (-59 m) and Flat Creek 5 (-5.2 m), respectively. Above Flat Creek 5 and across the K-Pg boundary, raw richness declines with only between five and 10 OTUs present at each locality in the Tullock Member. In this unit, peak richness occurs just above the K-Pg boundary at the Morales 1 site (2.9 m above the boundary). Accounting for variation in sample sizes via rarefaction analysis reveals an overall similar pattern to that observed for the raw richness. Notably, the richness declines across the K-Pg boundary is reinforced and statistically robust on account of the largely non-overlapping 95% confidence intervals (Figure 21; Table 3).
As shown in the bottom graph in Figure 21, raw teleost richness fluctuates from between eight and 14 OTUs through the Hell Creek Formation. Raw richness peaks at 14 OTUs in both the lower and upper thirds of the Hell Creek at locality CMM (-59 m) and Flat Creek 5 (-5.2 m), respectively. Above Flat Creek 5 and across the K-Pg boundary, raw richness declines with only between five and 10 OTUs present at each locality in the Tullock Member. In this unit, peak richness occurs just above the K-Pg boundary at the Morales 1 site (2.9 m above the boundary). Accounting for variation in sample sizes via rarefaction analysis reveals an overall similar pattern to that observed for the raw richness. Notably, the richness declines across the K-Pg boundary is reinforced and statistically robust on account of the largely non-overlapping 95% confidence intervals (Figure 21; Table 3).
Standing richness (i.e., the number of OTUs that first or last appear in a horizon or that range through a horizon) shows a similar pattern but lacks the steep decline in the middle of the Hell Creek (i.e., from CMM [-59 m] to Hot Feet [-19.5 m]). As with raw teleost richness, standing richness declined across the K-Pg boundary. Between 10 and 15 OTUs co-occur in the Hell Creek assemblages (mean=13.4, median=14, mode=14), whereas only between eight and 12 do so in the Tullock (mean=9.2, median=8, mode=8). However, as discussed above, teleost richness of the Tullock was buoyed by immigration (teleost indet. type PT-1) and/or possible in situ speciation (i.e., Estesesox and Esox, Otophysi indet. type U-3/BvD [K morph] and [P morph]) during the Puercan, which was shown to be an important aspect of the recovery phase for other local vertebrates (e.g., mammals; Clemens, 2010; Wilson, 2013: Wilson et al., 2014a). Richness of the Puercan is further bolstered if taxa based on non-vertebral elements are included. For example, Phyllodus paulkatoi, which is based on teeth and tooth-plates, occurs at the Pu3 Garbani 13-Northwest Harley’s High locality. At that stratigraphic horizon (~45 m above the K-Pg boundary) the raw and standing richnesses thus would increase by one from eight to nine OTUs (not shown in Figure 21).
The standing richness curve of the local K-Pg survivors (see “standing survivor” richness in the bottom graph in Figure 21) emphasizes how the first appearances of OTUs in the lower half of the Tullock affects the perceived level of extinctions across the K-Pg boundary and during the Puercan. Only eight of the 14 upper Hell Creek OTUs make it across the boundary into the Pu1 interval for an extinction rate of 43%. The three OTUs known only from the lower Hell Creek (i.e., acanthomorph indet. type HC-3, teleost indet. types RvB, Gonorynchiformes indet. type H) are not included in this estimate as they may have gone extinct (at least locally) prior to the K-Pg boundary. When including the one range-through occurrence (i.e., the OTU that occurs in the Hell Creek and Pu3 of the Tullock but not the Pu1; acanthomorph indet. type HC-4), the extinction rate drops to 29%. During the Pu1 and Pu3 subintervals additional extinctions (Estesesox, acanthomorph indet. type HC-1 and HC-5, Otophysi indet. type U-3/BvD [K morph]) and an extirpation (acanthomorph indet. type AvE) occurred, with only five Cretaceous OTUs (acanthomorph indet. type HC-2 and HC-4, teleost indet. type U-4, Coriops/Lopadichthys, and Notogoneus) remaining at the top of the studied section (Table 1) for a total loss of 64% of upper Hell Creek OTUs in the local section of northeastern Montana, an estimate that again is like that calculated for the northern Western Interior (56%).
To have a more complete view of the early Paleocene, we can incorporate the Pu2 Pine Cree assemblage into our quantitative analysis, although we acknowledge the biogeographic variable that this might add to the results. The overall regional richness pattern is similar to the local pattern but with slightly elevated standing richnesses. Three Pine Cree Park taxa are not found in the Tullock Member, including gonorynchiform indet. type H, Diplomystus, and Otophysi indet. type U-3/BvD (P morph). Including these three taxa increases the regional standing richness by one during the Lancian (middle and upper thirds of the Hell Creek), one during the Pu1, and by three during the Pu2. The Pu2 occurrence of Gonorynchiformes indet. type H lowers the local extinction rate of 29% (four out of 14) to a regional extinction rate of 27% (four out of 15), which is equal to what we calculated above for the northern Western Interior. Standing survivor richness only increased by one during the Pu1 and Pu2 subintervals but ultimately dropped to only five taxa in the Pu3.
Changes in Teleost Relative Abundances
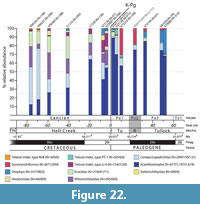 Wilson et al. (2024) found that, in the Denver Basin, the relative abundances of freshwater teleosts from two fossil assemblages (one below and one above the K-Pg boundary) changed, with the Paleocene assemblage being dominated by fewer, highly abundant morphotypes. The sites studied here also show that significant changes in the relative abundances of the major teleost groups were associated with the local declines in teleost richness across the K-Pg boundary and through the Puercan. These changes are illustrated in Figure 22 following the methodology of Brinkman et al. (2021, figure 20). Several patterns are apparent when tracking the assemblages up section through the Hell Creek Formation and lower half of the Tullock Member. These include: i) the decline in richness and relative abundance of osteo-glossomorphs resulting from the extinction of the Wilsonichthyidae and the extirpation of the Hiodontidae across the K-Pg boundary and the declining relative abundances of Coriops/Lopadichthys through the entire section, ii) the increasing relative abundances of the acanthomorphs through the Hell Creek and their overall higher relative abundances in the Tullock (i.e., 12%-64% versus 57%-89%, respectively), iii) the moderate increases in relative abundances of teleost indet. type U-4 through the Hell Creek (from ~1% near the base to 14.9% just below the boundary) and its immediate drop and maintained lower relative abundances in the Tullock localities, iv) the first appearance of teleost indet. type PT-1 in the Tullock, and v) the significantly higher relative abundances of the Otophysi during the Pu3 interval at the top of the section. Another noteworthy result is the ubiquitous higher relative abundance of the gonorynchiform Notogoneus at the McKeever Ranch 1 locality (UCMP loc. V72210) and the Pine Cree locality. At the McKeever Ranch 1 locality, centra of Notogoneus account for 30.5% of the sample and 15.5% at the Pine Cree locality (Table 1). These abundances are significantly higher than from all other localities where the taxon occurs (0.3%-2.5%). The high abundance of Notogoneus may be a result of local paleoecological factors, because it also shows a patchy distribution in the Belly River Group of Alberta, Canada, generally being rare or absent but being of high relative abundance at localities that are characterized by freshwater estuarine conditions on the distal lateral floodplain with extremely low water flow strengths (Brinkman, 2019; Divay et al., 2020). This instance highlights the potential influence of local ecological conditions on the taxonomic diversities of our samples.
Wilson et al. (2024) found that, in the Denver Basin, the relative abundances of freshwater teleosts from two fossil assemblages (one below and one above the K-Pg boundary) changed, with the Paleocene assemblage being dominated by fewer, highly abundant morphotypes. The sites studied here also show that significant changes in the relative abundances of the major teleost groups were associated with the local declines in teleost richness across the K-Pg boundary and through the Puercan. These changes are illustrated in Figure 22 following the methodology of Brinkman et al. (2021, figure 20). Several patterns are apparent when tracking the assemblages up section through the Hell Creek Formation and lower half of the Tullock Member. These include: i) the decline in richness and relative abundance of osteo-glossomorphs resulting from the extinction of the Wilsonichthyidae and the extirpation of the Hiodontidae across the K-Pg boundary and the declining relative abundances of Coriops/Lopadichthys through the entire section, ii) the increasing relative abundances of the acanthomorphs through the Hell Creek and their overall higher relative abundances in the Tullock (i.e., 12%-64% versus 57%-89%, respectively), iii) the moderate increases in relative abundances of teleost indet. type U-4 through the Hell Creek (from ~1% near the base to 14.9% just below the boundary) and its immediate drop and maintained lower relative abundances in the Tullock localities, iv) the first appearance of teleost indet. type PT-1 in the Tullock, and v) the significantly higher relative abundances of the Otophysi during the Pu3 interval at the top of the section. Another noteworthy result is the ubiquitous higher relative abundance of the gonorynchiform Notogoneus at the McKeever Ranch 1 locality (UCMP loc. V72210) and the Pine Cree locality. At the McKeever Ranch 1 locality, centra of Notogoneus account for 30.5% of the sample and 15.5% at the Pine Cree locality (Table 1). These abundances are significantly higher than from all other localities where the taxon occurs (0.3%-2.5%). The high abundance of Notogoneus may be a result of local paleoecological factors, because it also shows a patchy distribution in the Belly River Group of Alberta, Canada, generally being rare or absent but being of high relative abundance at localities that are characterized by freshwater estuarine conditions on the distal lateral floodplain with extremely low water flow strengths (Brinkman, 2019; Divay et al., 2020). This instance highlights the potential influence of local ecological conditions on the taxonomic diversities of our samples.
DISCUSSION
Based on the combined evidence of centra and tooth-bearing elements, a minimum of 18 Operational Taxonomic Units (OTUs) are recognized in the lower Paleocene (Puercan 1-3). One elopomorph, Phyllodus, is represented by teeth and branchial tooth-plates. The possibility that one of the centrum morphotypes recognized here is from Phyllodus is considered unlikely because the centra present in a closely related taxon, Paralbula, which were described by Neuman and Brinkman (2005) and Brinkman (2019), are unlike any of the centra present in the lower Paleocene localities examined. Thus, assuming that centra of Phyllodus and Paralbula were similar, we can be reasonably confident that Phyllodus centra are not present in the Puercan samples because none that resemble those of Paralbula are present.
Three osteoglossomorphs are represented by tooth-bearing elements: Lopadichthys, represented by a dentary, Coriops, represented by a basibranchial tooth-plate, and Ostariostoma, represented by a dentary. The centra of Lopadichthys and Coriops cannot be distinguished from each other, so for these osteoglossomorphs, tooth-bearing elements give a more reliable indication of taxonomic diversity than do centra. The third osteoglossomorph in the assemblage, Ostariostoma, is represented only by dentaries. Based on a similar stratigraphic and biogeographic distribution pattern, the possibility that these are associated with the centra referred to gonorynchiform indet. type H cannot be ruled out. Thus, to minimize a possible inflation of the diversity present, only one of these two taxa is included in estimates of the diversity of teleost fishes present.
The clupeomorph Diplomystus, is represented only by centra. Dentaries of Diplomystus are known from other assemblages (Divay and Murray, 2016a), so the possibility that this taxon is also represented by one or more of the tooth-bearing elements described herein can be excluded.
Two ostariophysans are represented by both dentaries and centra, and two are represented only by centra. The ostariophysans represented by both dentaries and centra are Notogoneus and Otophysi indet. type U3/BvD (P morph). Otophysi indet. type U3/BvD (K morph) is known by dentaries and centra from Cretaceous deposits but only by centra from the Pu1. As discussed above, the fourth ostariophysan, gonorynchiform indet. type H, may be independently represented by tooth-bearing elements referred to Ostariostoma, so only one of these two taxa is included in the estimates of diversity of early Paleocene teleost assemblages. Because centra are more abundant, they are used for estimating the relative abundance and distribution of gonorynchiform indet. type H in the lower Paleocene localities.
Esocids are represented by two taxa: Estesesox and Esox. Both are represented by centra, dentaries, and palatines.
Three acanthomorphs are represented by dentaries and five by centra. However, it is not possible to confidently associate centrum morphotypes with dentaries. Thus, estimates of the total diversity of acanthomorphs in the early Paleocene are based only on centra.
Two kinds of teleosts of unknown relationships are represented by centra, teleost indet. type U-4 and teleost indet. type PT-1. No tooth-bearing elements can be associated with either of these centrum morphotypes.
The list of early Paleocene teleosts recognized here differs from that of Bryant (1989) in not including Platacodon or Paleolabrus. The report of Platacodon in the early Paleocene is not accepted here because this occurrence was based on dentaries that had been referred to this genus by Estes (1964) but were subsequently shown to be from an esocid by Wilson et al. (1992). Currently, Platacodon is represented only by distinctive pharyngeal tooth-plates and isolated pharyngeal teeth. These are widely distributed in upper Maastrichtian localities (Brinkman et al., 2021) but are not present in the localities studied here. Paleolabrus was erected by Estes (1969b) based on tooth-bearing elements and centra. Bryant (1989) noted that some of the tooth-bearing elements had infolded bases, a feature found in Lepisosteus. A subsequent study of fossil and recent lepisosteiforms by Grande (2010) has reinforced the significance of this radial infolding of the base of larger teeth, or “plicidentine”, as an autapomorphy of lepisosteid gars, and unique to members of the family within Actinopterygii, so the tooth-bearing elements with this feature can be confidently identified as being from a member of the Lepisosteidae. Based on the published figures, the centra that Estes (1969b) referred to Paleolabrus are those of Notogoneus (compare Estes, 1969b, plate 3e-f with Divay et al., 2020, figure 7C). The identity of the tooth-plates is uncertain, but the presence of paired vomers suggests it is not a teleost. Thus, Paleolabrus is not included because it appears to be a chimera, including elements from multiple different kinds of fish.
Comparison of Teleost Assemblages of the Early and Late Paleocene
Comparison of fish assemblages from the early Paleocene (this study) with those from the late Paleocene indicates that, rather than a recovery to pre-existing levels of diversity seen during the Late Cretaceous, a further decrease in taxonomic richness occurred throughout the epoch. Late Paleocene teleosts are best known from the Paskapoo Formation of Alberta, Canada, which is Tiffanian in age (Table 2). Teleosts from this formation that are represented by articulated specimens include: i) the osteoglossomorphs Joffrichthys symmetropterus, J. tanyourus, and Lopadichthys lindoei (Li and Wilson, 1996; Murray et al., 2018); ii) the esocid Esox tiemanni (Wilson, 1980); iii) the smelt Speirsaenigma lindoei (Wilson and Williams, 1991); and iv) the percopsiforms Massamorichthys wilsoni and Lateopisciculus turrifumosus (Murray 1994; Murray and Wilson, 1996). In addition to these seven taxa, isolated elements were referred to the Hiodontidae, Cyprinoidea, Gonorynchidae, and Asineopidae by Wilson (1980). The hiodontid element was later referred to Joffrichthys (Li and Wilson, 1996), although the identification of the cyprinoid is still accepted (Murray, 2020). Thus, based on both articulated specimens and isolated elements, 10 kinds of teleosts are present in the Tiffanian Paskapoo Formation.
Additional evidence regarding the diversity of teleost assemblages of Tiffanian age is provided by a vertebrate microfossil assemblage from a locality in the Ravenscrag Formation of Saskatchewan called the Roche Percée locality. The fishes from this locality were described by Sinha et al. (2021). Nine teleosts are present. Thus, although the teleosts present in the Ravenscrag locality cannot be identified with the same level of precision as those from the Paskapoo Formation, the diversity of these two assemblages are similar. Two of the Ravenscrag Formation fishes, teleost indet. type PT-1 and protacanthopterygian indet. type ES-1, had not been recognized in the Paskapoo Formation and are not present in the Puercan localities sampled here. In addition, one indeterminate osteoglossomorph, designated type SO-1, and two indeterminate acanthomorphs designated type RPA-1 and RPA-2, are represented by centra. However, it is possible that the osteoglossomorph and acanthomorphs are from taxa represented by articulated specimens in the Paskapoo Formation, so they are not included as separate OTUs in Table 2. Surprisingly, the taxonomic richness of the Tiffanian interval, with a total of 15 OTUs present, remains relatively low. Thus, rather than a recovery to levels of diversity seen during the Late Cretaceous, taxonomic richness remained low through the early Paleocene and into the late Paleocene.
CONCLUSIONS
Based on our comparisons of late Maastrichtian and early Paleocene teleost assemblages from northeastern Montana and south-central Saskatchewan, the K-Pg mass extinction event had a moderate impact on non-marine teleost fishes with an extinction rate of 27% immediately across the K-Pg boundary. Most of the casualties were taxa that were rare in the late Maastrichtian assemblages. A similar result was recently documented from the Denver Basin in Colorado (Wilson et al., 2024). Although new taxa occurred immediately following the K-Pg boundary, either as a result of immigration or of diversification events, further apparent extinctions occurred during the early Paleocene, and taxonomic richness remained low. This pattern contrasts with the one seen with teleost fishes in marine environments, where the extinction of top predators of the Cretaceous allowed a diversification of crown teleosts, giving rise to new families of fast swimming and piscivorous bony fishes that filled the functional roles vacated by extinction victims (Cavin, 2002; Sibert and Norris, 2015; Ribeiro et al., 2018). Nevertheless, the higher than previously reported level of extinctions of teleost fishes between the Late Cretaceous and the late Paleocene adds to the evidence that the K-Pg event had a substantial influence on shaping today’s aquatic communities, and highlights that the effects of this event are more complex than previously thought.
ACKNOWLEDGEMENTS
This study would not have been possible without the efforts of an army of students, volunteers, and professionals who processed samples from numerous vertebrate microfossil localities. Although the focus of the work was often mammals, all vertebrate fossils were recovered and made available for subsequent faunal studies. Thanks to P. Holroyd for access to specimens in the collections of the UCMP. Thanks to R. McKellar, E. Bamford, and T. Tokayrk for access to specimens in the collections of the Royal Saskatchewan Museum. For access to collections in the UALVP we would like to thank A. Murray. Also, thanks to A. Murray and O. Vernygora of the University of Alberta for CT scanning specimens. For access to recent comparative material, we would additionally like to thank S. Cumbaa and M. Currie (Canadian Museum of Nature), D. Nelson (University of Michigan Museum of Zoology), G. Arratia and A. Bentley (Kansas University), and K. Seymour (Royal Ontario Museum). We also thank two anonymous reviewers who made significant comments that led to the improvement of the paper.
We acknowledge that the fossils in this paper from Montana were collected on lands that are the traditional territory of the Fort Belknap Assiniboine and Gros Ventre Tribes and Fort Peck Assiniboine and Sioux Tribes. Future field trips will be respectful to the original peoples and sovereignty. We thank the McKeever and Engdahl families for land access and support, H.J. Garbani for the site discovery, volunteers from the UCMP, UW Biology, and Burke Museum for field and laboratory work. We gratefully acknowledge W.A. Clemens’ immense contributions to study of the Hell Creek area, his thoughtful mentorship, and his loyal friendship over many decades. Fieldwork to collect fossil and geological data were done with permissions of the US Bureau of Land Management, Montana Department of Natural Resources and Conservation, and oral permission of private landowners (Bob and Jane Engdahl and John and Cathy McKeever). The discovery and initial collecting at the various localities were largely funded by the UCMP and the NSF. Further research was supported by the Hell Creek Project from the Myhrvold and Havranek Charitable Family Fund, UW Biology, and Burke Museum (G.P.W.M.).
REFERENCES
Agassiz, L. [1839] 1843. Recherches sur les poissons fossiles. Vol. 3. Neuchatel and Soleure.
https://www.biodiversitylibrary.org/item/23069#page/76/mode/1up
Archibald, J.D. 1996. Dinosaur Extinction and the End of an Era: What the Fossils Say. Columbia University Press, New York.
https://doi.org/10.1016/s0012-8252(97)81864-8
Archibald, J.D. 2011. Extinction and Radiation: How the Fall of Dinosaurs Led to the Rise of Mammals. The Johns Hopkins University Press, Baltimore, Maryland.
https://doi.org/10.1093/jmammal/gyv087
Archibald, J.D. and Bryant, L.J. 1990. Differential Cretaceous/Tertiary extinctions of nonmarine vertebrates; evidence from northeastern Montana. Geological Society of America Special Papers, 247:549-562.
https://doi.org/10.1130/spe247-p549
Arratia, G. 1999. The monophyly of Teleostei and stem-group teleosts, consensus and disagreements, p. 265-334. In Arratia, G. and Schultze, H.-P. (eds.), Mesozoic fishes 2-systematics and fossil record. Verlag Dr. Friedrich Pfeil, München, Germany.
https://doi.org/10.1671/0272-4634(2001)021[0767:tsgotc]2.0.co;2
Arratia, G. 2001. The sister-group of Teleostei: consensus and disagreements. Journal of Vertebrate Paleontology, 21:767-773.
https://doi.org/10.1671/0272-4634(2001)021[0767:tsgotc]2.0.co;2
Badgley, C. 1986. Counting individuals in mammalian fossil assemblages from fluvial environments. Palaios, 1:328-338.
https://doi.org/10.2307/3514695
Bagils, R.Z. 2004. Basal clupeomorphs and ellimmichthyiform phylogeny, p. 391-404. In Arratia, G. and Tintori, A. (eds.), Mesozoic Fishes 3-Systematics, Paleoenvironments and Biodiversity. Verlag Dr. Friedrich Pfeil, München.
https://doi.org/10.1206/0003-0082(2003)404<0001:roebad>2.0.co;2
Berg, L.S. 1940. Classification of fishes, both Recent and fossil. Travaux de l’Institut Zoologique de l’Académie des Sciences de l’URSS. Moscow, Russia. Translated and reprinted in English, 1947. J. W. Edwards, Ann Arbor, Michigan.
Bleeker, P. 1859. Enumeratio speciorum piscium hucusque in Archipelago Indico observatarum. Acta Societatis Scientiarum Indo-Neêrlandae, 6:1-276.
Brinkman, D.B. 1990. Paleoecology of the Judith River Formation (Campanian) of Dinosaur Provincial Park, Alberta, Canada: Evidence from vertebrate microfossil localities. Palaeogeography, Palaeoclimatology, Palaeoecology, 78:37-54.
https://doi.org/10.1016/0031-0182(90)90203-J
Brinkman, D.B. 2008. The structure of Late Cretaceous (Late Campanian) nonmarine aquatic communities: a guild analysis of two vertebrate microfossil localities in Dinosaur Provincial Park, Alberta, Canada, p. 33-60. In Sankey, J.T. and Baszio, S. (eds.), Vertebrate Microfossil Assemblages, Their Roles in Paleoecology and Paleobiogeography. Indiana University Press, Bloomington, Indiana.
https://doi.org/10.1016/0031-0182(90)90203-j
Brinkman, D.B. 2019. Teleost abdominal centra from the Belly River Group of Alberta, Canada. Paludicola, 12:109-152.
Brinkman, D.B. and Neuman, A.G. 2002. Teleost centra from uppermost Judith River Group (Dinosaur Park Formation, Campanian) of Alberta, Canada. Journal of Paleontology, 76:138-155.
https://doi.org/10.1666/0022-3360(2002)076<0138:tcfujr>2.0.co;2
Brinkman, D.B., Newbrey, M.G., Neuman, A.G., and Eaton, J.G. 2013. Freshwater Osteichthyes from the Cenomanian to Late Campanian of Grand Staircase-Escalante National Monument, Utah, p. 195-236. In Titus, A.L. and Loewen, M.A. (eds.), At the Top of the Grand Staircase, The Late Cretaceous of Southern Utah. Indiana University Press, Indiana.
Brinkman, D.B., Newbrey, M.G., and Neuman, A.G. 2014. Diversity and paleoecology of actinopterygian fish from vertebrate microfossil localities in the Maastrichtian Hell Creek Formation of Montana, p. 247-270. In Wilson, G.P., Clemens, W.A., Horner, J.R., and Hartman, J.H. (eds.), Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas. Geological Society of America Special Paper, 503.
https://doi.org/10.1130/2014.2503(09)
Brinkman, D.B., Neuman, A.G., and Divay, J.D. 2017. Non-marine fishes from the Late Santonian Milk River Formation of Alberta, Canada-evidence from vertebrate microfossil localities. Vertebrate Anatomy Morphology and Paleontology, 3:7-46.
https://doi.org/10.18435/b5pp41
Brinkman, D.B., Divay, J.D., DeMar, Jr., D.G., and Wilson Mantilla, G.P. 2021. A systematic reappraisal and quantitative study of the non-marine teleost fishes from the late Maastrichtian of the Western Interior of North America - evidence from vertebrate microfossil localities. Canadian Journal of Earth Sciences, 58:936-967.
https://doi.org/10.1139/cjes-2020-0168
Brusatte, S.L., Butler, R.J., Barrett, P.M., Carrano, M.T., Evans, D.C., Lloyd, G.T., Mannion, P.D., Norell, M.A., Peppe, D.J., Upchurch, P., and Williamson, T.E. 2015. The extinction of the dinosaurs. Biological Reviews, 90:628-642.
https://doi.org/10.1111/brv.12128
Bryant, L.J. 1987a. Belonostomus (Teleostei: Aspidorynchidae) from the Late Paleocene of North Dakota. Paleobios, 43:1-3.
Bryant, L.J. 1987b. A new genus and species of Amiidae (Holostei; Osteichthyes) from the Late Cretaceous of North America, with comments on the phylogeny of the Amiidae. Journal of Vertebrate Paleontology, 7:349-361.
https://doi.org/10.1080/02724634.1988.10011669
Bryant, L.J. 1989. Non-dinosaurian lower vertebrates across the Cretaceous-Tertiary boundary in northeastern Montana. University of California Publications in Geological Sciences, 134:1-107.
Cavin, L. 2002. Effects of the Cretaceous-Tertiary Boundary Event on Bony Fishes, p. 141-158. In Buffetaut, E. and Koeberl, C. (eds.), Geological and Biological Effects of Impact Events. Springer Science & Business Media, Berlin, Germany.
https://doi.org/10.1007/978-3-642-59388-8_6
Clemens, W.A. 2010. Were immigrants a significant part of the earliest Paleocene mammalian fauna of the North American Western Interior? Vertebrata Palasiatica, 48:285-307.
Cope, E.D. 1877. A contribution to the knowledge of the ichthyological fauna of the Green River shales. Bulletin of the United States Geological Survey of the Territories, 3:807-819.
Cope, E.D. 1885. Eocene paddle-fish and Gonorhynchidae [sic]. American Naturalist, 19:1090-1091.
Dartevelle, E. and Casier, E. 1943. Les poissons fossiles du Bas-Congo et des régions voisines (première partie). Annales du Musée du Congo Belge. A. - Minéralogie, Géologie, Paléontologie, série 3, 2:1-200.
Divay, J.D. and Murray, A.M. 2015. The late Eocene-early Oligocene ichthyofauna from the Eastend area of the Cypress Hills Formation, Saskatchewan, Canada. Journal of Vertebrate Paleontology, 35:1-25.
https://doi.org/10.1080/02724634.2014.956877
Divay, J.D. and Murray, A.M. 2016a. An early Eocene fish fauna from the Bitter Creek area of the Wasatch Formation of southwestern Wyoming, U.S.A. Journal of Vertebrate Paleontology, 36:e1196211.
https://doi.org/10.1080/02724634.2016.1196211
Divay, J. D. and Murray, A.M. 2016b. The fishes of the Farson Cutoff Fishbed, Bridger Formation (Eocene), greater Green River Basin, Wyoming, U.S.A. Journal of Vertebrate Paleontology, 36:e1212867:1-18.
https://doi.org/10.1080/02724634.2016.1212867
Divay, J.D., Brinkman, D.B., and Neuman, A.G. 2020. Late Cretaceous Notogoneus from microvertebrate assemblages of the Dinosaur Park Formation, Campanian of southern Alberta, Canada, and insight into the ecology and evolution of early gonorynchids. Journal of Vertebrate Paleontology, 39(5):e1699801.
https://doi.org/10.1080/02724634.2019.1699801
Eberth, D.A. and Brinkman, D.B. 1997. Paleoecology of an estuarine, incised-valley fill in the Dinosaur Park Formation (Judith River Group, Upper Cretaceous) of Southern Alberta, Canada. PALIOS, 12:43-58.
https://doi.org/10.2307/3515293
Estes, R. 1964. Fossil vertebrates from the Lance Formation, Eastern Wyoming. University of California Publications in Geological Sciences, 49:1-187.
Estes, R. 1969a. Studies on fossil phyllodont fishes: Interrelationships and evolution in the Phyllodontidae (Albuloidei). Copeia, 1969:317-331.
https://doi.org/10.2307/1442082
Estes, R. 1969b. Two new Late Cretaceous fishes from Montana and Wyoming. Breviora, 335:1-15.
Estes, R. and Hiatt, R. 1978. Studies on fossil phyllodont fishes; a new species of Phyllodus (Elopiformes, Albuloidea) from the Late Cretaceous of Montana. PaleoBios, 28:1-10.
Fastovsky, D.E. and Bercovici, A. 2016. The Hell Creek Formation and its contribution to the Cretaceous-Paleogene extinction: A short primer. Cretaceous Research, 57:368-390.
https://doi.org/10.1016/j.cretres.2015.07.007
Fink, S.V. and Fink, W.L. 1996. Interrelationships of the ostariophysan fishes (Teleostei), p. 209-249. In Stiassny, M.L.J., Parenti, L.R., and Johnson, G.D. (eds.), Interrelationships of Fishes. Academic Press, New York.
https://doi.org/10.1016/b978-012670950-6/50012-6
Forey, P.L., Littlewood, D.T.J., Ritchie, P., and Meyer, A. 1996. Interrelationships of Elopomorph fishes, p. 175-192. In Stiassny, M.L.J., Parenti, L.R., and Johnson, G.D. (eds.), Interrelationships of Fishes. Academic Press, New York.
https://doi.org/10.1016/b978-012670950-6/50010-2
Friedman, M. 2009. Ecomorphological selectivity among marine teleost fishes during the end-Cretaceous extinction. Proceedings of the National Academy of Sciences, 106:5218-5223.
https://doi.org/10.1073/pnas.0808468106
Garstang, W. 1931. The phyletic classification of Teleostei. Proceedings of the Leeds Philosophical and Literary Society. Scientific Section, 2:240-260.
Gardner, J.D. and DeMar, Jr, D.G. 2013. Mesozoic and Palaeocene lissamphibian assemblages of North America: a comprehensive review. Palaeobiodiversity and Palaeoenvironments, 93:459-515.
https://doi.org/10.1007/s12549-013-0130-z
Grande, L. 1984. Paleontology of the Green River Formation, with a review of the fish fauna, Second Edition. Geological Survey of Wyoming, Bulletin, 63:1-333.
Grande, L. 1985. Recent and fossil clupeomorph fishes with materials for revision of the subgroups of clupeoids. Bulletin of the American Museum of Natural History, 181:231-372.
Grande, L. 1988. A well preserved paracanthopterygian fish (Teleostei) from freshwater lower Paleocene deposits of Montana. Journal of Vertebrate Paleontology, 8:117-130.
https://doi.org/10.1080/02724634.1988.10011692
Grande, L. 2001. An updated review of the fish faunas from the Green River Formation, the world’s most productive freshwater lagerstätten, p. 1-38. In Gunnell, G.F. (ed.), Topics in Geobiology, Volume 18: Eocene Biodiversity: Unusual Occurrences and Rarely Sampled Habitats. Kluwer Academic/Plenum Publishers, New York, NY.
Grande, L. 2010. An empirical synthetic pattern study of gars (Lepisosteiformes) and closely related species, based mostly on skeletal anatomy. The resurrection of Holostei. American Society of Ichthyologists and Herpetologists Special Publication 6, Supplementary issue of Copeia, 10(2A):1-871.
https://doi.org/10.1643/0045-8511-2010.3.bmi
Grande, L. and Cavender, T.M. 1991. Description and phylogenetic reassessment of the monotypic †Ostariostomidae (Teleostei). Journal of Vertebrate Paleontology, 11:405-416.
https://doi.org/10.1080/02724634.1991.10011412
Grande, L. and Grande, T. 1999. A new species of † Notogoneus (Teleostei: Gonorynchidae) from the Upper Cretaceous Two Medicine Formation of Montana, and the poor Cretaceous record of freshwater fishes from North America. Journal of Vertebrate Paleontology, 19:612-622.
https://doi.org/10.1080/02724634.1999.10011175
Grande, T., Laten, H., and López, J.A. 2004. Phylogenetic relationships of extant esocid species (Teleostei: Salmoniformes) based on morphological and molecular characters. Copeia, 2004:743-757.
https://doi.org/10.1643/cg-04-007r1
Grande, T.C., Wilson, M.V., Reyes, A.V., Buryak, S.D., Wolfe, A.P., and Siver, P.A. 2022. A new, Late Cretaceous gonorynchiform fish in the genus† Notogoneus from drill core of crater-lake deposits in a kimberlite maar, Northwest Territories, Canada. Cretaceous Research, 135:105176.
https://doi.org/10.1016/j.cretres.2022.105176
Greenwood, P.H., Rosen, D.E., Weitzman, S.H., and Myers, G.S. 1966. Phyletic studies of teleostean fishes, with a provisional classification of living forms. Bulletin of the American Museum of Natural History, 131:339-455.
http://hdl.handle.net/2246/1678
Hartman, J.H., Johnson, K.R., and Nichols, D.J. 2002. The Hell Creek Formation and the Cretaceous-Tertiary boundary in the northern Great Plains: An Integrated continental record of the end of the Cretaceous. Geological Society of America Bulletin, Special Paper 361.
https://doi.org/10.1130/SPE361
Hilton, E.J. 2002. Comparative osteology and phylogenetic systematics of fossil and living bony-tongue fishes (Actinopterygii, Teleostei, Osteoglossomorpha). Zoological Journal of the Linnean Society, 137:1-100.
https://doi.org/10.1046/j.1096-3642.2003.00032.x
Holland, S.M. 2003. Analytic Rarefaction 1.3: http://stratigrafia.org/software/
Holroyd, P.A., Wilson, G.P., and Hutchison, J.H. 2014. Temporal changes within the latest Cretaceous and early Paleogene turtle faunas of northeastern Montana, p. 299-312. In Wilson, G.P., Clemens, W.A., Horner, J.R., and Hartman, J.H. (eds.), Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas. Geological Society of America Special Paper, 503.
https://doi.org/10.1130/2014.2503(11)
Jordan, D.S. 1923. Classification of fishes including families and genera as far as known. Stanford University Publications, University Series, Biological Sciences, 3:77-243.
https://doi.org/10.5962/bhl.title.161386
Larson, D.W., Brinkman, D.B., and Bell, P.R. 2010. Faunal assemblages from the upper Horseshoe Canyon Formation, an early Maastrichtian cool-climate assemblage from Alberta, with special reference to the Albertosaurus sarcophagus bonebed. Canadian Journal of Earth Sciences, 47:1159-1181.
https://doi.org/10.1139/E10-00
Lavoué, S. 2016. Was Gondwanan breakup the cause of the intercontinental distribution of Osteoglossiformes? A time-calibrated phylogenetic test combining molecular, morphological, and paleontological evidence. Molecular Phylogenetics and Evolution, 99:34-43.
https://doi.org/10.1016/j.ympev.2016.03.008
LeCain, R., Clyde, W.C., Wilson, G.P., and Riedel. J. 2014. Magnetostratigraphy of the Hell Creek and lower Fort Union Formations in northeastern Montana, p. 137-147. In Wilson, G.P., Clemens, W.A., Horner, J.R., and Hartman, J.H. (eds.), Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas. Geological Society of America Special Paper, 503.
https://doi.org/10.1130/2014.2503(04)
Li., G.-Q. and Wilson, M.V.H. 1996. The discovery of heterotidinae (Teleostei: Osteoglossidae) from the Paleocene Paskapoo formation of Alberta, Canada. Journal of Vertebrate Paleontology, 16:198-209.
https://doi.org/10.1080/02724634.1996.10011308
Linnaeus, C. 1758. Systema Naturae. 10th ed. Regnum Animale, 824 p.
Lyson, T.R., Sayler, J.L., and Joyce, W.G. 2019. A new baenid turtle, Saxochelys gilberti, gen. et sp. nov., from the uppermost Cretaceous (Maastrichtian) Hell Creek Formation: sexual dimorphism and spatial niche partitioning within the most speciose group of Late Cretaceous turtles. Journal of Vertebrate Paleontology, 39(4):e1662428.
https://doi.org/10.1080/02724634.2019.1662428
McKinney, M.L. 1997. Extinction vulnerability and selectivity: Combining ecological and paleontological views. Annual Review of Ecology and Systematics, 28:495-516.
https://doi.org/10.1146/annurev.ecolsys.28.1.495
Mercier, G., DeMar Jr., D.G., and Wilson, G.P. 2014. Frogs and toads (Lissamphibia, Anura) during the end-Cretaceous mass extinction: evidence from the fossil record of northeastern Montana. Program and Abstracts, 74th Annual Meeting, Society of Vertebrate Paleontology, p. 187.
Moore, J.R., Wilson, G.P., Sharma, M., Hallock, H.R., Braman, D.R., and Renne, P.R. 2014. Assessing the relationships of the Hell Creek-Fort Union contact, Cretaceous-Paleogene boundary, and Chicxulub impact ejecta horizon at the Hell Creek Formation lectostratotype, Montana, USA, p. 123-135. In Wilson, G.P., Clemens, W.A., Horner, J.R., and Hartman, J.H. (eds.), Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas. Geological Society of America Special Paper, 503.
https://doi.org/10.1130/2014.2503(03)
Müller, J. 1844. Ueber den Bau und die Grenzen der Ganoiden und über das natürliche System der Fische. Bericht über die zur Bekanntmachung geeigneten Verhandlungen der Akademie der Wissenschaften, Berlin, 1846:117-216. [An 1846 English translation of this paper, translated by J. W. Griffith, can be found in Scientific Memoirs 4(16):499-558.]
Murray, A.M. 1994. Description of two new species of basal paracanthoperygian fishes from the Palaeocene of Alberta, and a phylogenetic analysis of the percopsiforms (Teleostei: Paracanthopterygii). Ph.D. diss. University of Alberta. Edmonton, Alberta.
Murray, A.M. 2020. Early Cenozoic Cyprinoids (Ostariophysi: Cypriniformes: Cyprinidae and Danionidae) from Sumatra, Indonesia. Journal of Vertebrate Paleontology, 40:1-24.
https://doi.org/10.1080/02724634.2020.1762627
Murray, A.M. and Wilson, M.V.H. 1996. A new Palaeocene genus and species of percopsiform (Teleostei: Paracanthopterygii) from the Paskapoo Formation, Smoky Tower, Alberta. Canadian Journal of Earth Sciences, 33:429-438.
https://doi.org/10.1139/e96-032
Murray, A.M. and Wilson, M.V.H. 2013. Two new paraclupeid fishes (Clupeomorpha: Ellimmichthyiformes) from the Upper Cretaceous of Morocco, p. 267-290. In Arratia, G., Schultze, H.-P., and Wilson, M.V.H. (eds.), Mesozoic Fishes 5 - Global Diversity and Evolution. Verlag Dr. Friedrich Pfeil, München.
Murray, A.M., Zelenitsky, D.K., Brinkman, D.B., and Neuman, A.G. 2018. Two new Palaeocene osteoglossomorphs from Canada, with a reassessment of the relationships of the genus † Joffrichthys, and analysis of diversity from articulated versus microfossil material. Zoological Journal of the Linnean Society, 183:907-944.
https://doi.org/10.1093/zoolinnean/zlx100
Murray, A.M., Brinkman, D.B., Newbrey, M.G., and Neuman, A.G. 2019. Earliest North American articulated freshwater acanthomorph fish (Teleostei: Percopsiformes) from Upper Cretaceous deposits of Alberta, Canada. Geological Magazine, 157:1087-1096.
https://doi.org/10.1017/s001675681900132
Murray, A.M. and Brinkman, D.B. 2023. Morphological variation in the first vertebra among acanthomorph fishes-a guide for identifying fossil centra from microvertebrate sites. Vertebrate Anatomy Morphology Palaeontology, 11:42-90.
https://doi.org/10.18435/vamp29392
Neuman, A.G. and Brinkman, D.B. 2005. Fishes of the fluvial beds, p. 167-185. In Currie, P.J. and Kopplelhus, E.B. (eds.), Dinosaur Provincial Park, A Spectacular Ancient Ecosystem Revealed. Indiana University Press, Bloomington, Indiana.
Newbrey, M.G., Brinkman, D.B., Winkler, D.A., Freedman, E.A., Neuman, A.G., Fowler, D.W., and Woodward, H.N. 2013. Teleost centrum and jaw elements from the Upper Cretaceous Nemegt Formation (Campanian-Maastrichtian) of Mongolia and a re-identification of the fish centrum found with the theropod Raptorex kreigsteini, p. 291-303. In Arratia, G., Schultze, H.-P., and Wilson, M.V.H. (eds.), Mesozoic Fishes 5 - Global Diversity and Evolution. Verlag Dr. Friedrich Pfeil, München.
Patterson, C. 1993. Osteichthyes: Teleostei, p. 621-656. In Benton, M.J. (ed.), The Fossil Record 2. Chapman & Hall, London.
Patterson, C. and Rosen, D.E. 1977. A review of the ichthyodectiform and other Mesozoic teleost fishes, and the theory and practice of classifying fossils. Bulletin of the American Museum of Natural History, 158:81-172.
http://hdl.handle.net/2246/1224
Peng, J-H., Russell, A.P., and Brinkman, D.B. 2001. Vertebrate microsite assemblages (exclusive of mammals) from the Foremost and Oldman formations of the Judith River Group (Campanian) of southeastern Alberta: an illustrated guide. Provincial Museum of Alberta Natural History Occasional Paper No. 25:1-54.
Rafinesque, C.S. 1815. Analyse de la nature, ou tableau de l'univers et des corps organisés (No. 177). Palerme, Aux dépens de l'auteur.
Redman, C.M., Gardner, J.D., Scott, C.S., and Braman, D.R. 2015. Geological setting of vertebrate microfossil localities across the Cretaceous-Paleogene boundary in southwestern Saskatchewan, Canada. Canadian Journal of Earth Science, 52:846-862.
https://doi.org/10.1139/cjes-2015-0038
Renne, P.R., Deino, A.L., Hilgen, F.J., Kuiper, K.F., Mark, D.F., Mitchell III, W.S., Morgan, L.E., Mundil, R., and Smit, J. 2013. Time scales of critical events around the Cretaceous-Paleogene boundary. Science, 339:684-687.
https://doi.org/10.1126/science.1230492
Ribeiro, E., Davis, A.M., Rivero-Vega, R.A., Orti, G., and Betancur, R. 2018. Post-Cretaceous bursts of evolution along the benthic-pelagic axis in marine fishes. Proceedings of the Royal Society B, 285:20182010.
https://doi.org/10.1098/rspb.2018.201
Robertson, D.S., Lewis, W.M., Sheehan, P.M., and Toon, O.B. 2013. K-Pg extinction patterns in marine and freshwater environments: The impact winter model. Journal of Geophysical Research: Biogeosciences, 118:1006-1014.
https://doi.org/10.1002/jgrg.20086
Rosen, D.E. 1973. Interrelationships of higher euteleostean fishes, p. 397-513. In Greenwood, P.H., Miles, R.S., and Patterson, C. (eds.), Interrelationships of Fishes. Academic Press, London.
Rosen, D.E. and Patterson, C. 1969. The structure and relationships of the paracanthopterygian fishes. Bulletin of the American Museum of Natural History, 141:357-474.
http://hdl.handle.net/2246/1996
Rosen, D.E. and Greenwood, P.H. 1970. Origin of the Weberian apparatus and the relationships of the ostariophysan and gonorynchiform fishes. American Museum Novitates, 2428:1-5.
http://hdl.handle.net/2246/2638
Russell, L.S. 1974. Fauna and correlation on the Ravenscrag Formation (Paleocene) of southwestern Saskatchewan. Royal Ontario Museum Life Sciences Contribution, 102:1-52.
Sagemehl, M. 1885. Beiträge zur vergleichenden Anatomie der Fische. III. Das Cranium der Characiniden nebst allgemeinen Bemerkungen über die mit einem Weber'schen Apparat versehenen Physostomenfamilien. Morphologisches Jahrbuch, 10:1-119.
Schaeffer, B. 1949. A teleost from the Livingston Formation of Montana. American Museum Novitates, 1427:1-16.
Schwarzhans, W. and Stringer, G.L. 2020. Fish otoliths from the late Maastrichtian Kemp Clay (Texas, USA) and the early Danian Clayton Formation (Arkansas, USA) and an assessment of extinction and survival of teleost lineages across the K-Pg boundary based on otoliths. Rivista Italiana di Paleontologia e Stratigrafia, 126(2):395-446.
https://doi.org/10.13130/2039-4942/13425
Schwarzhans, W.W., Carnevale, G., and Stringer, G.L. 2024. The diversity of teleost fishes during the terminal Cretaceous and the consequences of the K/Pg boundary extinction event. Netherlands Journal of Geosciences, 103:e5.
https://doi.org/10.1017/njg.2024.1
Scott, C.S., Fox, R.C., and Redman, C.M. 2016. A new species of the basal plesiadapiform Purgatorius (Mammalia, Primates) from the early Paleocene Ravenscrag Formation, Cypress Hills, southwest Saskatchewan, Canada: further taxonomic and dietary diversity in the earliest primates. Canadian Journal of Earth Sciences, 53:343-354.
https://doi.org/10.1139/cjes-2015-0238
Sheehan, P.M. and Fastovsky, D.E. 1992. Major extinctions of land-dwelling vertebrates at the Cretaceous-Tertiary boundary, eastern Montana. Geology, 20:556-560.
https://doi.org/10.1130/0091-7613(1992)020<0556:MEOLDV>2.3.CO;2
Sibert, E.C. and Norris, R.D. 2015. New Age of Fishes initiated by the Cretaceous−Paleogene mass extinction. Proceedings of the National Academy of Sciences, 112:8537-8542.
https://doi.org/10.1073/pnas.150498511
Sinha, S., Brinkman, D.B., and Murray, A.M. 2019. A morphological study of vertebral centra in extant species of pike, Esox (Teleostei: Esociformes). Vertebrate Anatomy Morphology Palaeontology, 7:111-128.
https://doi.org/10.18435/vamp29357
Sinha, S., Brinkman, D.B., Murray, A.M., and Krause, D.W. 2021. Late Paleocene Fishes of the Ravenscrag Formation, Roche Percée Area, Southeastern Saskatchewan, Canada. Journal of Vertebrate Paleontology, 41(3):e1957907.
https://doi.org/10.1080/02724634.2021.1957907
Smith, W.M.L. and Craig, M.T. 2007. Casting the percomorph net widely: The importance of broad taxonomic sampling in the search for the placement of serranid and percid fishes. Copeia, 2007:35-55.
https://doi.org/10.1643/0045-8511(2007)7[35:ctpnwt]2.0.co;2
Sprain, C.J., Renne, P.R., Wilson, G.P., and Clemens, W.A. 2015. High-resolution chronostratigraphy of the terrestrial Cretaceous-Paleogene transition and recovery interval in the Hell Creek region, Montana. Geological Society of America Bulletin, 127:393-409.
https://doi.org/10.1130/b31076.1
Sprain, C.J., Renne, P.R., Clemens, W.A., and Wilson, G.P. 2018. Calibration of chron C29r: New high-precision geochronologic and paleomagnetic constraints from the Hell Creek region, Montana. Geological Society of America Bulletin, 130:1615-1644.
https://doi.org/10.1130/b31890.1
Stiassny, M.L.J. 1986. The limits and relationships of the acanthomorph teleosts. Journal of Zoology, 1:411-460.
https://doi.org/10.1111/j.1096-3642.1986.tb00644.x
Swisher III, C.C., Dingus, L., and Butler, R.F. 1993. 40Ar/39Ar dating and magnetostratigraphic correlation of the terrestrial Cretaceous-Paleogene boundary and Puercan Mammal Age, Hell Creek-Tullock formations, eastern Montana. Canadian Journal of Earth Sciences, 30:1981-1996.
https://doi.org/10.1139/e93-174
Twitchett, R.J. 2006. The palaeoclimatology, palaeoecology and palaeoenvironmental analysis of mass extinction events. Palaeogeography, Palaeoclimatology, Palaeoecology, 232:190-213.
https://doi.org/10.1016/j.palaeo.2005.05.019
Weaver, L.N., Tobin, T.S., Claytor, J.R., Wilson Deibel, P.K., Clemens, W.A., and Wilson Mantilla, G.P. 2022. Revised stratigraphic relationships within the lower Fort Union Formation (Tullock Member, Garfield County, Montana, USA) provide a new framework for examining post K-Pg mammalian recovery dynamics. Palaios, 37:104-127.
https://doi.org/10.2110/palo.2021.011
Whitlock, J.A. 2010. Phylogenetic relationships of the Eocene percomorph fishes † Priscacara and † Mioplosus. Journal of Vertebrate Paleontology, 30:1037-1048.
https://doi.org/10.1080/02724634.2010.483534
Wilson, G.P. 2013. Mammals across the K/Pg boundary in northeastern Montana, U.S.A.: Dental morphology and body-size patterns reveal extinction selectivity and immigrant-fueled ecospace filling. Paleobiology, 39:429-469.
https://doi.org/10.1666/12041
Wilson, G.P., DeMar Jr., D.G., and Carter, G. 2014a. Extinction and survival of salamander and salamander-like amphibians across the Cretaceous-Paleogene boundary in northeastern Montana, USA, p. 271-297. In Wilson, G.P., Clemens, W.A., Horner, J.R., and Hartman, J.H. (eds.), Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas. Geological Society of America Special Paper, 503.
https://doi.org/10.1130/2014.2503(10)
Wilson, G.P., Clemens, W.A., Horner, J.R., and Hartman, J.H. 2014b. Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas. Geological Society of America Special Paper, 503.
https://doi.org/10.1130/9780813725031
Wilson Mantilla, G.P., Chester, S.G., Clemens, W.A., Moore, J.R., Sprain, C.J., Hovatter, B.T., Mitchell, W.S., Mans, W.W., Mundil, R., and Renne, P.R. 2021. Earliest Palaeocene purgatoriids and the initial radiation of stem primates. Royal Society Open Science, 8:210050.
https://doi.org/10.1098/rsos.210050
Wilson, J.D., Huang, E.J., Lyson, T.R., and Bever, G.S. 2024. Freshwater fish and the Cretaceous/Palaeogene boundary: a critical assessment of survivorship patterns. Proceedings of the Royal Society B, 291:20241025.
https://doi.org/10.1098/rspb.2024.1025
Wilson, M.V.H. 1980. Oldest known Esox (Pisces: Esocidae), part of a new Paleocene teleost fauna from western Canada. Canadian Journal of Earth Sciences, 17:307-312.
https://doi.org/10.1139/e80-030
Wilson, M.V.H. 1984. Osteology of the Palaeocene teleost Esox tiemani. Palaeontology, 27:597-608.
Wilson, M.V.H. and Murray, A.M. 2008. Osteoglossomorpha: phylogeny, biogeography, and fossil record and the significance of key African and Chinese fossil taxa. Geological Society, London, Special Publications, 295:185-219.
https://doi.org/10.1144/sp295.12
Wilson, M.V.H. and Williams, R.R.G. 1991. New Paleocene genus and species of smelt (Teleostei: Osmeridae) from freshwater deposits of the Paskapoo Formation, Alberta, Canada, and comments on osmerid phylogeny. Journal of Vertebrate Paleontology, 11:434-451.
https://doi.org/10.1080/02724634.1991.10011414
Wilson, M.V.H., Brinkman, D.B., and Neuman, A.G. 1992. Cretaceous Esocoidea (Teleostei): early radiation of the pikes in North American fresh waters. Journal of Paleontology, 66:839-846.
https://doi.org/10.1017/s0022336000020849
Wynd, B.M., DeMar Jr., D.G., and Wilson, G.P. 2020. Euselachian diversity through the uppermost Cretaceous Hell Creek Formation of Garfield County, Montana, USA, with implications for the Cretaceous-Paleogene mass extinction in freshwater environments. Cretaceous Research, 113:104483.
https://doi.org/10.1016/j.cretres.2020.104483
