

 New insights on the anatomy, paleobiology, and biostratigraphy of Xenungulata (Mammalia) from the Paleogene of South America
New insights on the anatomy, paleobiology, and biostratigraphy of Xenungulata (Mammalia) from the Paleogene of South America
Article number: 27.2.a30
https://doi.org/10.26879/1360
Copyright Society of Vertebrate Paleontology, June 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 21 November 2023. Acceptance: 24 May 2024.
ABSTRACT
Xenungulata is a group of early Paleogene mammals endemic to South America that remains poorly understood. The first species described within this order, Carodnia feruglioi, has laid the basis for the homonymous zone, currently positioned between the Peligran (Danian) and Itaboraian (Ypresian) South American Land Mammals Ages. Through the study of new specimens and a revaluation of materials from historical collections, we have identified remains belonging to Carodniidae and Etayoidae xenungulates and assigned them to Carodnia feruglioi and Notoetayoa gargantuai. This has led to a reinterpretation of their dental characteristics, setting the foundations for establishing dental homologies with other native ungulates of South America. Additionally, we introduce Carodnia karuen sp. nov., discovered at the lower Paleocene levels in Cerro Redondo locality in Patagonia, Argentina. The specimen extends the biostratigraphic and biochronological significance of Carodnia as a fossil guide in the Paleocene of southern South America. A paleobiological analysis of these specimens focused on comparing the variation in mandibular morphology and measurements within Carodniidae. This approach has allowed the identification of a potential sexual dimorphism pattern at the generic level for Carodnia. The findings imply that Carodnia feruglioi from the early Paleocene of Patagonia and the early Eocene Carodnia vieirai from Itaboraí, Brazil, probably maintained a sexual variation in mandibular width and height.
Javier N. Gelfo. División Paleontología de Vertebrados, Museo de La Plata. Paseo del Bosque s/n (B1900FWA) La Plata, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Argentina. CONICET, Godoy Cruz 2290, Ciudad Autónoma de Buenos Aires, Argentina. Corresponding author. jgelfo@fcnym.unlp.edu.ar
Guillermo M. López. División Paleontología de Vertebrados, Museo de La Plata. Paseo del Bosque s/n (B1900FWA) La Plata, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Argentina. glopez@fcnym.unlp.edu.ar
Mariano Bond. División Paleontología de Vertebrados, Museo de La Plata. Paseo del Bosque s/n (B1900FWA) La Plata, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Argentina. CONICET, Godoy Cruz 2290, Ciudad Autónoma de Buenos Aires, Argentina. constantino1453@yahoo.com.ar
Keywords: Carodnia; new species; Etayoidae; Paleocene; Eocene; sexual dimorphism; MorphoBank
Final citation: Gelfo, Javier N., López, Guillermo M., and Bond, Mariano. 2024. New insights on the anatomy, paleobiology, and biostratigraphy of Xenungulata (Mammalia) from the Paleogene of South America. Palaeontologia Electronica, 27(2):a30.
https://doi.org/10.26879/1360
palaeo-electronica.org/content/2024/5229-south-american-xenungulata
Copyright: June 2024 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
https://zoobank.org/26FB2C48-D1E1-41FB-A283-983A5367E511
INTRODUCTION
Xenungulata is a group of South American native ungulates (SANU) (Croft et al., 2020), first recorded in the early Paleocene of Patagonia, Argentina, and with a wide distribution through almost all of South America. They are represented by two families (i.e., Carodniidae, Etayoidae) and no more than six species, with inferred body masses varying between 2 and 400 kg (Gelfo et al., 2020). As originally typified by the family Carodniidae, they are characterized by a bilophodont dentition as those present in other SANU such as Pyrotheria, as well as Holarctic groups such as Dinocerata and Perissodactyla (i.e., Tapiridae). The bilophodonty and particularly the similitude of the m3 of Carodnia and Dinocerata (Simpson, 1935a) was considered as an argument to join Pyrotheria and Xenungulata, to that Holarctic group in a non-ungulate clade named Uintatheriamorpha (Schoch and Lucas, 1985; Lucas 1993). However, the possible relationship with Dinocerata was highly questioned and most authors dismissed it (McKenna, 1980; Cifelli, 1993). Later phylogenetic analyses, exclusively based on dental characters, considered xenungulates as the sister group of the pyrotheres Pyrotherium and Carolozittelia (Gelfo et al., 2008). Billet (2009, 2011) proposed to consider Pyrotheria within Notoungulata but did not incorporate Xenungulata in their matrix, and as a result, the dental similarities were overlooked. Muizon et al. (2015) in a large-scale phylogenetic analysis, recovered pyrotheres and xenungulates as sister-groups in most parsimonious trees (MPTs), clearly based on the shared bilophodont structure of their teeth. Despite this similarity, there still exists a high morphological gap to relate both groups, particularly considering the cranial similitudes of pyrotheres and notoungulates.
The genus Carodnia and its type species C. feruglioi Simpson, 1935a, was based on an isolated left m3 from the lower Paleocene beds of the Rio Chico Group at Bajo Palangana locality in Chubut Province (Simpson, 1935a, 1935c). Ctalecarodnia cabrerai Simpson, 1935a, was also described for the same levels, based on an isolated and broken fragment of a molarized left p4 with the trigonid and part of the talonid, an almost complete talonid of a left m1, and talonid and distal part of the trigonid of a right m1 (Simpson, 1935a). Cabrera (1935) studied materials collected by Ing. José Brandmayr in the lower “Pehuenche” strata of Bajo Palangana, found about 20 m above the “Banco Negro” [Black Bank] guide (BN), and attributes five isolated teeth (MLP-PV 34-V-22-8) to Carodnia feruglioi, and a right p4 to Ctalecarodnia cabrerai (MLP-PV 34-V-22-9). These remains include a right m3 for C. feruglioi, of which Simpson (1935a) suggested the possibility that it belongs to the same individual described for the holotype (MGP-PD 29046). Additionally, there are left and right p2 and p3, following Simpson’s (1967) identification, although the p3 was originally considered a pair of left and right p2 (Cabrera, 1935). These elements were never properly described, and only the right p2 and the left p3 were originally drawn by Cabrera (1935:15). Pictures taken from the cast housed at the American Museum of Natural History in New York, USA, were published by Vera et al. (2020) showing a broken and incomplete left p2, with only the distal part of the tooth, which is a cast feature and not the preservation of the original specimen.
The genus Carodnia has a wide paleobiogeographic distribution outside Patagonia with a very conservative dental morphology. Carodnia vieirai Paula Couto, 1952, relies on nearly complete dental and postcranial remains from the lower Eocene levels of the Itaboraí Formation, at São José de Itaboraí, in Río de Janeiro, Brazil (Paula Couto, 1952). Based on it and specimens of C. feruglioi, Paula Couto (1952) created and defined the family Carodniidae, the order Xenungulata, and indicated the junior synonym of Ctalecarodnia respect to Carodnia. In recent times, two new Carodniidae were recognized. Carodnia inexpectans Antoine et al., 2015, came from the lower Eocene levels of the Mogollón Formation, in Quebrada Cabeza de Vaca, Tumbes, northwest Peru, and Rodcania kakan Gelfo et al., 2020, from the Río Loro Formation (late early Paleocene?-early Eocene?) at Aguas Chiquitas locality, Tucumán, Argentina.
A second family of Xenungulata, Etayoidae, was recognized based on Etayoa bacatensis Villarroel, 1987, from the Paleocene-Eocene sediments of the Bogota Formation in Colombia. Notoetayoa gargantuai Gelfo, López, and Bond, 2008, represented by an isolated last lower molar found in the same stratigraphic and geographic provenance of C. feruglioi (i.e., Peñas Coloradas Formation, Chubut province, Argentina) was also added to this family (Gelfo et al., 2008).
This work aims to contribute to the knowledge of Xenungulata, one of the most enigmatic SANU orders through the approach of different confluent topics. Within this context, a novel species of Carodnia is described, and additional specimens from diverse levels and localities in Patagonia are assigned to Carodnia feruglioi and Notoetayoa gargantuai. Several aspects of dental homologies, and the potential significance of Carodnia jaw variation in facilitating paleobiological inferences related to sexual dimorphism, are explored. The xenungulate specimens of Brandmayr’s collection are also studied, illustrated, and reinterpreted; and several aspects of the Carodnia zone (Simpson, 1935b) are here commented on and discussed.
MATERIALS AND METHODS
Several authors (e.g., Bordas, 1935; Feruglio, 1949) have made specific references to the terms “Riochiquense” or “Riochican,” applying them to various concepts, not always related, such as lithostratigraphic or biochronological units, or faunal assemblages. Simpson (1935a) initially defined the Rio Chico Formation based on the Kibenikhoria Fauna, but later expanded the biostratigraphic characterization to include three faunas/zones within this formation (Simpson, 1935c). To avoid potential confusion with these terms, we will use “Riochican” to refer to the SALMA (South American Land Mammal Age), and reserve “Rio Chico” for the specific geological formation or group of lithostratigraphic units previously utilized.
Map imagery of figures was captured and modified from Google Earth Pro, data sourced from SIO, NOAA, U.S. Navy, NGA, GEBCO, and Landsat/Copernicus.
Dental Nomenclature
We follow Gelfo et al. (2008, 2020) for homologies and dental features. The terms “protolophid” and “protocristid” are here considered as the lophid and cristid, respectively, that connect the protoconid and metaconid (Van Valen, 1966) to avoid the term metalophid (Osborn, 1907) used for different structures in several taxa. Also “paralophid” (Stirton, 1941) and “paracristid” (Van Valen, 1966) are considered, respectively, as the lophid and cristid that run mesial from the protoconid. “Hypolophid” is used for the labiolingual lophid extending from the hypoconid to the entoconid. The term “plagioconulid” is used for the small cusp located lingual or mesiolingual to the hypoconid for the cusp mentioned by Vera et al. (2020) as conulid in the m3 of Carodnia feruglioi. The plagioconulid was briefly mentioned in Hershkovitz's (1971) theoretical occlusal pattern of quadritubercular molars, but it was neither adequately defined nor illustrated with any specific taxa. The limited references to this cusp in the existing literature were primarily centered on the variable occurrence of an accessory talonid cusp in primates and artiodactyls (Erfurt and Haubold, 1989; Hershkovitz, 1990; Takai and Anaya, 1996), rather than regarding it as a diagnostic feature for any species. Furthermore, in this context, the plagioconulid is explicitly distinguished from the centroconid, also referred to as mesoconid, as it is developed and associated with the cristid obliqua in several bunodont South American ungulates (e.g., Didolodus multicuspis Ameghino, 1897).
The wear facet in the occlusal surface of premolars and molars is described here following the Modular Wear Facet Nomenclature (Schultz et al., 2018). The following abbreviations are used in the description: pacd-mb (paraconid-mesiobuccal), prcd-h (protoconid-horizontal), prcd-b (protoconid-buccal), prcd-db (protoconid-distobuccal), prld-d (protolophid-distal), prcgd-b (precingulid-buccal), hd-b (hypoconid-buccal), hd-mb (hypoconid-mesiobuccal), and hyld-d (hypolophid-distal). As this system is expandable to other dental elements, we add pocgd-ml (postcingulid mesiolingual).
Institutional Abbreviations
AMNH: American Museum of Natural History, New York, USA; DGM: ex-Divisão de Geologia e Mineralogia, Rio de Janeiro, Brazil; GM: Colecciones paleontológicas del Departamento de Geociencias, Universidad Nacional de Colombia, Bogotá. MGP-PD: Museo de Geologia e Paleontologia, Università degli Studi di Padova, Padua, Italy; MACN: Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina; MLP-PV: División Paleontología Vertebrados, Museo de La Plata, La Plata, Argentina; MPEF-PV: Museo Paleontológico Egidio Feruglio, Trelew, Argentina; UNPSJB-PV: Colección Paleovertebrados, Repositorio Científico y Didáctico. “Dr. Eduardo Musacchio”, Universidad Nacional de la Patagonia San Juan Bosco, Comodoro Rivadavia, Argentina.
GEOGRAPHIC AND GEOLOGICAL SETTINGS
 The newly described specimens presented in this study were collected from three distinct localities within the Chubut province, where the Rio Chico Group is exposed (Figure 1). The initial site, known as Bajo de la Palangana or simply Bajo Palangana, holds a long-standing status as a renowned paleontological site. Situated approximately 40 km north of Comodoro Rivadavia on the Atlantic coast of Chubut province, Argentina, this locality has garnered significant scholarly attention over the years (Feruglio, 1931, 1949; Brandmayr, 1932; Bordas, 1935; Cabrera, 1935; Simpson, 1935a, 1935c; Vera and Krause, 2020). This is an expansive erosional basin at the northeast of Pico Salamanca and the southeast of Pico Chico. Within this basin, the strata of the Rio Chico Group overlay the Salamanca Formation along the coastal cliff, extending westward to underlie the successive strata of the Sarmiento and Chenque formations. In 1935, Simpson delineated three informal members within what was then referred to as the Rio Chico Formation (Simpson, 1933, 1935a). Presently, it is recognized as a group
The newly described specimens presented in this study were collected from three distinct localities within the Chubut province, where the Rio Chico Group is exposed (Figure 1). The initial site, known as Bajo de la Palangana or simply Bajo Palangana, holds a long-standing status as a renowned paleontological site. Situated approximately 40 km north of Comodoro Rivadavia on the Atlantic coast of Chubut province, Argentina, this locality has garnered significant scholarly attention over the years (Feruglio, 1931, 1949; Brandmayr, 1932; Bordas, 1935; Cabrera, 1935; Simpson, 1935a, 1935c; Vera and Krause, 2020). This is an expansive erosional basin at the northeast of Pico Salamanca and the southeast of Pico Chico. Within this basin, the strata of the Rio Chico Group overlay the Salamanca Formation along the coastal cliff, extending westward to underlie the successive strata of the Sarmiento and Chenque formations. In 1935, Simpson delineated three informal members within what was then referred to as the Rio Chico Formation (Simpson, 1933, 1935a). Presently, it is recognized as a group  (Figure 2 and see below). The lowermost unit, characterized by its lack of fossil content, boasts an approximate thickness of 25 m and includes the “Banco Negro Superior” [Upper Black Bank] (BNS) (Feruglio, 1931; Brandmayr, 1932) as its upper boundary (Simpson, 1935b). The middle member, spanning approximately 55 m in thickness, primarily consists of thick cross-bedded red and white sandstones with intercalated clay lenses. Simpson (1935b) designated this interval as the Carodnia zone, which also encompasses the lower mammal level at Cerro Redondo, from where the new species described is coming. The uppermost member is characterized by lenses of white or grey to light pink sandstones interspersed within a predominantly yellowish, greenish, or reddish clay sequence. For this stratum, positioned just below the Casamayor tuff (i.e., Sarmiento Formation), Simpson (1935b) introduced the Ernestokokenia faunal/zone. The latter faunal/zones have been correlated with the Peñas Coloradas and Las Flores formations, respectively, of the Rio Chico Group (Legarreta and Uliana, 1994; Bond et al., 1995). The Peñas Coloradas Formation unconformably overlies the Salamanca Formation and the transitional levels, at the top of which outcrops the BNS identified by Simpson, particularly in the surroundings of Punta Peligro (Raigemborn, 2008; Raigemborn et al., 2010). It is indicative of a fluvial system characterized by low to moderate energy levels. It predominantly consists of fine-grained conglomerate, very fine to very coarse sandstone, and extensive mudstone deposits. These sediments exhibit a composition that includes both volcaniclastic and siliclastic materials, displaying a color spectrum ranging from grey to the distinctive reddish hue observed at the type locality (Raigemborn et al., 2010).
(Figure 2 and see below). The lowermost unit, characterized by its lack of fossil content, boasts an approximate thickness of 25 m and includes the “Banco Negro Superior” [Upper Black Bank] (BNS) (Feruglio, 1931; Brandmayr, 1932) as its upper boundary (Simpson, 1935b). The middle member, spanning approximately 55 m in thickness, primarily consists of thick cross-bedded red and white sandstones with intercalated clay lenses. Simpson (1935b) designated this interval as the Carodnia zone, which also encompasses the lower mammal level at Cerro Redondo, from where the new species described is coming. The uppermost member is characterized by lenses of white or grey to light pink sandstones interspersed within a predominantly yellowish, greenish, or reddish clay sequence. For this stratum, positioned just below the Casamayor tuff (i.e., Sarmiento Formation), Simpson (1935b) introduced the Ernestokokenia faunal/zone. The latter faunal/zones have been correlated with the Peñas Coloradas and Las Flores formations, respectively, of the Rio Chico Group (Legarreta and Uliana, 1994; Bond et al., 1995). The Peñas Coloradas Formation unconformably overlies the Salamanca Formation and the transitional levels, at the top of which outcrops the BNS identified by Simpson, particularly in the surroundings of Punta Peligro (Raigemborn, 2008; Raigemborn et al., 2010). It is indicative of a fluvial system characterized by low to moderate energy levels. It predominantly consists of fine-grained conglomerate, very fine to very coarse sandstone, and extensive mudstone deposits. These sediments exhibit a composition that includes both volcaniclastic and siliclastic materials, displaying a color spectrum ranging from grey to the distinctive reddish hue observed at the type locality (Raigemborn et al., 2010).
An alternative interpretation of the informal members identified by Simpson (1935b) at Bajo Palangana suggests that the lower sterile member should be designated as the Peñas Coloradas Formation (Figure 2). In this reinterpretation, the middle member (i.e., Carodnia zone) and the upper member (i.e., Ernestokokenia zone) are both considered as within the lower and upper segments of the Las Flores Formation, as suggested by (Krause et al., 2017) and further supported by Vera and Krause (2020). In these views, the BNS was considered the probable base of the Las Flores Formation (Vera and Krause, 2020). The correlation of the accurate lithostratigraphic unit with the Carodnia zone hinges upon the necessity for a more precise delimitation of the formations mentioned above. Pending such refinement, we adhere to a conservative interpretation (as advocated by Legarreta and Uliana, 1994; Raigemborn, 2008; Raigemborn et al., 2010). It is worth noting that other remains discussed in this study, recovered by geologist N. Ronconi (MPEF-PV 1872, 1873, 1874), as well as by F.J. Goin (MPEF-PV 564), have been reported to be located within the strata referred as Peñas Coloradas Formation.
The second locality, Cerro Redondo, is a well-known hill positioned approximately 20 km to the north-northeast of Bajo Palangana and nearly 6 km west of Puerto Visser. The new Carodnia species described in this study was found at the stratum identified by Simpson (1935b) as predominantly gray, interspersed with occasional red clays, and punctuated by lenses of white, greenish, or red, coarse, cross-bedded sandstone. This layer is situated approximately 32 to 35 m above the uppermost boundary of the “Banco Negro Inferior” [Lower Black Bank] (BNI). A few fossil mammals were described from this level, among which deserve attention the oldest polydolopid marsupial, Amphidolops yapa, senior synonym of Seumadia yapa Simpson 1935 (Chornogubsky, 2021) and the alleged notonychopid (Bonaparte and Morales, 1997) litoptern Wainka tshotshe Simpson 1935. It is important to note that Simpson (1935b) included this small fauna in the previously mentioned Carodnia zone, despite the absence of shared taxa between Bajo Palangana and Cerro Redondo.
The third locality is referenced in the collection records of UNSJB as “Riochican levels” formerly associated with the now-defunct institution “Tiro Federal Coronel Pringles” (43°16’51.0” S, 65°30’40.2” W) within the Department of Gaiman, Chubut province. This locality appears to correspond to one of the outcrops near Pan de Azúcar Hill, which is presently a restricted area under the control of the Mapuche-Tehuelche community known as “Loma Torta”. While we retain the UNPSJB’s designated collection locality name for this specimen, it is likely synonymous with Simpson’s Pan de Azúcar. The specimen from this locality was recovered from the same level of white to pink arkosic sandstone mentioned by Simpson (Simpson, 1935b; 1935c). Simpson noted the presence of turtle, crocodile, and mammal remains within this level. He conceded that this fauna remained poorly documented but posited that these strata could potentially align with the upper sandstone fauna identified as part of the Ernestokokenia chaishoer zone in the Bajo Palangana region. Following Krause et al. (2017) and Vera and Krause (2020), the equivalent level near the present locality at Pan de Azúcar was identified as part of the Las Flores Formation (Chornogubsky et al., 2023).
SYSTEMATIC PALAEONTOLOGY
Class MAMMALIA Linnaeus, 1758
Order XENUNGULATA Paula Couto, 1952
Family CARODNIIDAE Paula Couto, 1952
Genus CARODNIA Simpson, 1935
Synonymy. Ctalecarodnia Simpson, 1935a: 22, figure 22.
Type species. Carodnia feruglioi Simpson, 1935
Emended diagnosis (after Simpson, 1935a; Paula Couto, 1952; Vera et al., 2020). Carodniid xenungulates with pentadactyl, short and slender limbs, alternated carpal bones, and broad and flat ungual phalanges. Cervical vertebrae short. Complete brachyodont dentition, an anterior extension of the masseteric fossa onto the body of the dentary below the m3, partially hidden in lateral view, by the mandibular ramus. P1-2/1-2 with a main mesial cusp and labiolingually compressed. P3-4 with V-shaped protocone and strong meso-external paracone. M1-2 bilophodont, M3 with well-developed protoloph. No diastema between c-p1-2. The p4 molariform with low and short crested talonid with hypolophid. Trigonid of m1-2 short, formed by a high protolophid and short paracristid, metacristid if present, close to the mesial side of the protolophid; precingulid in m1-2; no paraconid; and labial cingulid on the talonid of m3.
Carodnia feruglioi Simpson, 1935
Synonymy. Ctalecarodnia cabrerai Simpson, 1935a: 24, figure 22. Carodnia cabrerai Paula Couto, 1952: 371.
Holotype. MGP-PD 29046, mandible fragment with left m3.
Hypodigm. MGP-PD 29030a-c type of Ctalecarodnia cabrerai: MGP-PD 29030a, left p4; MPEF-PV 1872, right dentary with m2-3; MPEF-PV 1873, left dentary with p4 and part of m1; MPEF-PV 1874, left dentary with m3; MGP-PD 29030b, left m2? With broken trigonid, and MGP-PD 29030c, right m2? with broken trigonid; MGP-PD 29030d, a fragment of a cusp; MGP-PD 29030e, trigonid fragment of a left m1?; MGP-PD 29033a, left i3?; MGP-PD 29033b, mesial lobe of a left M2?; MGP-PD 29047, isolated right I3; MLP-PV 34-V-22-8: right m3, left and right p2-3; MLP-PV 34-V-22-9, right p4 (originally assigned to Ctalecarodnia cabrerai); UNPSJB-PV 680, a fragment of mandible with left canine and roots of p2-3; UNPSJB-PV 680-1, a fragment of mandible with right m1; UNPSJB-PV 680-2, a fragment of the left mandible with anterior root of m3; UNPSJB-PV 680-3, anterior root of a right m2.
Remarks. We refer here to Carodnia feruglioi a left dentary with p4 and the mesial part of trigonid of the m1 (MPEF-PV 1873), a left dentary with m3 (MPEF-PV 1874,), and a right dentary with m2-3 (MPEF-PV 1872). These specimens were previously described as Carodnia cf. C. feruglioi and are probably of the same individual (Gelfo et al., 2008). They were found by Nestor Ronconi in levels of the Peñas Coloradas Formation, between Puerto Visser and Bahía Bustamante (45°17’19” S, 67°01’29” W) in Chubut province, Argentina. The main difference with materials previously assigned as C. feruglioi is the presence of a postcingulid, which projects from the distolabial side of the hypoconid to the labial cingulid, surrounding the hypoconulid. This feature was also present in the m3 of C. vieirai as mentioned by Paula Couto (1952) and wrongly mentioned as absent in Gelfo et al. (2008) but missing in C. feruglioi and apparently in C. inexpectans (see Antoine et al., 2015: figure 2). Since there are no wear facets present on the distal wall of the talonid in the m3, it can be inferred that there is no functional contact between the distal part of the M3 and the postcingulid of the m3 during the occlusion process. In most mammals, distal wear facets in the m3 are typically confined to the mesial part of the hypoconulid, the postcristid, and the hypocristid, rather than in the postcingulid of the m3 if it is present. Consequently, it is conceivable that the distal wall of the talonid in the m3 remains relatively unaffected by selective pressures associated with chewing, allowing for the passive accumulation of a certain degree of morphological variation, such as the presence or absence of the postcingulid. Based on these dental observations and the overall similitude observed among the three specimens (i.e., MPEF-PV 1872, 1873, 1874) and the holotype of C. feruglioi, we have confidently attributed them to this species.
 The lot MLP-PV 34-V-22-8 belonging to a right m3 left and right p2-3, and MLP-PV 34-V-22-9 right p4 were first described by Cabrera (1935) but interpreting the p3 as P3, and the p4 as Ctalecarodnia cabrerai. Simpson (1967) partially illustrated these specimens correctly, and they were also discussed by Vera et al. (2020). However, the latter discussion was based on incomplete casts and several details of these specimens need to be highlighted considering their possible homologies. The main axis of the p2 runs mesiodistal with the protoconid as the main cusp with two main wear facets prcd-h and prcd-b (Figure 3A-C, G-I). The paracristid projects from it and bifurcates mesially in a strong precingulid which extends in a short portion labially and lingually. A strong distal cristid descends from the protoconid, up to a bifurcation which makes an inverted “Y” in the occlusal view (Figure 3C, I). There is a small bulge or thickening of the distal cristid at the bifurcation point. The short lingual portion of the cristid contacts an entoconid sensu Cabrera (1935). The more labial cristid, here interpreted as cristid obliqua, descends and reaches a high, but not too strong hypoconid at an angle of approximately 90° in labial view (Figure 3C). A postcingulid descends from the hypoconid to the lingual and labial sides. The hypoconulid is a small elevation with a strong wear facet (hd-mb) over the lingual postcingulid, with a soft facet pocgd-ml (Figure 3A, C, I). The p2 structure is comparatively more complex than the p2 of C. vieirai, where no cusp or cristid was identified by Paula Couto (1952) who just did a topographical description. In the p2 of C. vieirai, the protoconid is higher and more transversally compressed, the precingulid is less developed, the distal crest bifurcates in a short distolingual cristid with no entoconid associated, and a more distolabial cristid.
The lot MLP-PV 34-V-22-8 belonging to a right m3 left and right p2-3, and MLP-PV 34-V-22-9 right p4 were first described by Cabrera (1935) but interpreting the p3 as P3, and the p4 as Ctalecarodnia cabrerai. Simpson (1967) partially illustrated these specimens correctly, and they were also discussed by Vera et al. (2020). However, the latter discussion was based on incomplete casts and several details of these specimens need to be highlighted considering their possible homologies. The main axis of the p2 runs mesiodistal with the protoconid as the main cusp with two main wear facets prcd-h and prcd-b (Figure 3A-C, G-I). The paracristid projects from it and bifurcates mesially in a strong precingulid which extends in a short portion labially and lingually. A strong distal cristid descends from the protoconid, up to a bifurcation which makes an inverted “Y” in the occlusal view (Figure 3C, I). There is a small bulge or thickening of the distal cristid at the bifurcation point. The short lingual portion of the cristid contacts an entoconid sensu Cabrera (1935). The more labial cristid, here interpreted as cristid obliqua, descends and reaches a high, but not too strong hypoconid at an angle of approximately 90° in labial view (Figure 3C). A postcingulid descends from the hypoconid to the lingual and labial sides. The hypoconulid is a small elevation with a strong wear facet (hd-mb) over the lingual postcingulid, with a soft facet pocgd-ml (Figure 3A, C, I). The p2 structure is comparatively more complex than the p2 of C. vieirai, where no cusp or cristid was identified by Paula Couto (1952) who just did a topographical description. In the p2 of C. vieirai, the protoconid is higher and more transversally compressed, the precingulid is less developed, the distal crest bifurcates in a short distolingual cristid with no entoconid associated, and a more distolabial cristid.
The p3 of MLP-PV 34-V-22-8 is more molarized with the protoconid as the main cusp but associated labially with a strong and a bit lower metaconid (Figure 3D-F, J-LL). The paracristid could be interpreted as in p2 but there is an incipient paraconid? interrupting their continuity and from it, a mesial cristid contacts the precingulid (Figure 3F). Two small basins are delimited distally to the precingulid, the lingual one, which is also mesial to the base of the metaconid, is larger than the labial one. The mesial wear facets are well developed on the labial side (pacd-mb, prcgd-b, prcd-b, prcd-db) (Figure 3L). The talonid is also better defined than in p2, with a short cristid obliqua, and a large hypoconid, which in both teeth shows a clear hd-mb wear facet. A distal and straight cristid also descends from the protoconid lingually, ending in a small entoconid. The talonid basin is larger than the p2 delimited distally by a transverse postcingulid, which lingually seems to end in a small hypoconulid. This last cusp is close to the entoconid but separated from it by a short lingual opening of the talonid basin.
In contrast to the similar size of p2 and p3 in Carodnia feruglioi and the features mentioned above, the p3 of C. vieirai is smaller than their p2 and differs in the absence of the metaconid and paraconid; also, the lingual distal cristid, is lees projected distally so the talonid basin present a wider lingual opening. Particularly, Paula Couto (1952) mentioned a variable present posterointernal cusp and a hypoconid, united by a smooth ridge.
The p4 (MLP-PV 34-V-22-9) was initially attributed to Ctalecarodnia cabrerai by Cabrera (1935), and it was comprehensively described by him and Vera et al. (2020). Hence, this discussion will focus on providing a few additional comments regarding its homologies. The cristid extending mesially from the metaconid, which does not contact the precingulid, as noted by Cabrera (1935), is here interpreted as the metacristid (Figure 3M-O) as those identified by Vera et al. (2020) in the p4 of MGP-PD 29030a. The protoconid and metaconid are well identified in the protolophid, which is the highest structure of the tooth. The hypolophid is low, transverse, and well differentiated from the postcingulid. The resemblance of this p4, as well as others assigned to C. feruglioi (see Vera et al., 2020) with the p4 of C. vieirai is stronger than those between p2-3. The primary distinctions lie in the labial extension of both the postcingulid and precingulid within the Itaboraian taxon. However, the degree of molarization is nearly identical in both species.
The m3 cusps identification for MLP-PV 34-V-22-8 mostly agrees here with those features referred to by Vera et al. (2020) for the holotype MGP-PD 29046. But, in contrast, the conulid placed mesiolingually to the hypoconid is here identified as plagioconulid (Figure 3P-R). In Carodnia feruglioi this cusp is not easily identified even in specimens with scarce wear, since the hypoconid developed a lingual wear facet (hd-l) very quickly. In these specimens, the plagioconulid was probably rapidly erased by wear and integrated into a single wear facet with the hypoconid. Despite this, the presence of plagioconulid seems to be an important derived feature in the m3 of C. feruglioi.
Assigned materials. MPEF-PV 564 right dentary with the trigonid of the m2; MPEF-PV 8165 right dentary with part of the canine, and premolars and molars alveoli.
Geographic and stratigraphic provenance. Both specimens come from the Peñas Coloradas Formation, Rio Chico Group sensu Raigemborn et al. (2010), in Chubut province. MPEF-PV 564 was found by F.J. Goin in the outcrops located 5 km south of El Gauchito farm (45º13’53.8” S, 67º09’01.9” W). MPEF-PV 8165 was found by one of the authors (JNG) at Bajo Palangana, in a level of red sands partially covered by a fraction of whitish gray conglomerate sands, with a significant degree of weathering at 45º33’28.2” S, 67º17’43.1” W (Figure S1-Figure S2 in Appendix 1).
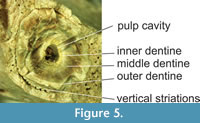
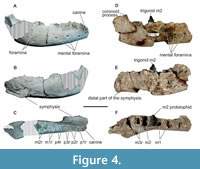 Description and comparisons. MPEF-PV 8165 is a fragmentary right jaw more robust than MPEF-PV 564 (Table 1). It was found in several associated pieces that fit together, plus a more posterior portion with remains of the coronoid crest and distal edge of the m3 alveolus (reconstructed in Figure 4A-C). The rest of the jaw is preserved in two parts, conserved the base of the canine and roots of p1-m2. The canine is transversely fractured practically at the height of the tooth’s neck (Figure 4A, C). The preserved section is rounded (19.24 mm x 17 mm), and three layers of dentine can be observed (Figure 5). While this resembles the different layers of dentin such as hard orthodentine, orthodentine, osteodentine, and vasodentine identified in Xenarthra (Kalthoff, 2011), a microstructural analysis would be necessary to further deepen these types of comparisons. The innermost layer, no more than 0.6 mm wide, surrounds a wide pulp cavity (3.50 mm in diameter) and is only differentiated from the middle layer by its more yellowish coloration. The middle layer is the most homogeneous, compact, and wide (4.56 mm) and is easily differentiated from the outermost portion of dentine by what appears to be a line of fragility. The outermost part does not exceed 1.76 mm in thickness and is well differentiated from the middle layer, and in it, a structure of concentric rings is observed towards the outermost portion. There is practically no preserved enamel, except for a thin layer on some of its edges. Strong vertical striations are observed in the outer part, which also shapes the bony contour of the alveolus. In the canine of Carodnia vieirai, the enamel is thin and corrugated, so, likely, the dentine crenulations observed in this specimen (MPEF-PV 8165) had a similar layer of enamel, which is not preserved. While the direct observation of specimen UNPSJB-PV 680 has failed to verify the presence of the finely grooved enamel layer described for its canine (Vera et al., 2020), it is still probable that this layer was present as observed in C. vieirai. Therefore, the observed grooves in UNPSJB-PV 680 are more likely to be associated with the outer layer of dentine, as reported in the present jaw.
Description and comparisons. MPEF-PV 8165 is a fragmentary right jaw more robust than MPEF-PV 564 (Table 1). It was found in several associated pieces that fit together, plus a more posterior portion with remains of the coronoid crest and distal edge of the m3 alveolus (reconstructed in Figure 4A-C). The rest of the jaw is preserved in two parts, conserved the base of the canine and roots of p1-m2. The canine is transversely fractured practically at the height of the tooth’s neck (Figure 4A, C). The preserved section is rounded (19.24 mm x 17 mm), and three layers of dentine can be observed (Figure 5). While this resembles the different layers of dentin such as hard orthodentine, orthodentine, osteodentine, and vasodentine identified in Xenarthra (Kalthoff, 2011), a microstructural analysis would be necessary to further deepen these types of comparisons. The innermost layer, no more than 0.6 mm wide, surrounds a wide pulp cavity (3.50 mm in diameter) and is only differentiated from the middle layer by its more yellowish coloration. The middle layer is the most homogeneous, compact, and wide (4.56 mm) and is easily differentiated from the outermost portion of dentine by what appears to be a line of fragility. The outermost part does not exceed 1.76 mm in thickness and is well differentiated from the middle layer, and in it, a structure of concentric rings is observed towards the outermost portion. There is practically no preserved enamel, except for a thin layer on some of its edges. Strong vertical striations are observed in the outer part, which also shapes the bony contour of the alveolus. In the canine of Carodnia vieirai, the enamel is thin and corrugated, so, likely, the dentine crenulations observed in this specimen (MPEF-PV 8165) had a similar layer of enamel, which is not preserved. While the direct observation of specimen UNPSJB-PV 680 has failed to verify the presence of the finely grooved enamel layer described for its canine (Vera et al., 2020), it is still probable that this layer was present as observed in C. vieirai. Therefore, the observed grooves in UNPSJB-PV 680 are more likely to be associated with the outer layer of dentine, as reported in the present jaw.
 Posterior to the canine there are seven alveoli in the first portion of the dentary, interpreted here as those of p1 (with a single root), and p2-4, (each of two roots). Our loci interpretation as p1-4, rests in what is observed in Carodnia vieirai, where p1 is small and single-rooted, (Figure 4C). In the occlusal view, the inter-alveolar space between mesial and distal roots of the same premolar is mesiodistally longer than between the alveoli of successive teeth. The labial face of the premolar series is broken.
Posterior to the canine there are seven alveoli in the first portion of the dentary, interpreted here as those of p1 (with a single root), and p2-4, (each of two roots). Our loci interpretation as p1-4, rests in what is observed in Carodnia vieirai, where p1 is small and single-rooted, (Figure 4C). In the occlusal view, the inter-alveolar space between mesial and distal roots of the same premolar is mesiodistally longer than between the alveoli of successive teeth. The labial face of the premolar series is broken.
The preserved portion of the symphysis is large and robust. The upper surface is labiolingually concave and the lower flat. A small foramen opens mesially, like in Carodnia vieirai (DGM 333-M, see Paula Couto, 1952). The more mesial part is missing, and it extends backward distal to the p3 locus. Paula Couto (1952) described for Carodnia vieirai a similar symphysis but in contrast, extended up to the posterior border of p4 in the type specimen DGM 333-M. We could not directly check this specimen, but in another jaw assigned by him to C. vieirai (cast AMNH 49848 of DGM 334-M) the symphysis extends to the distal part of p3 as in MPEF-PV 8165 (see also Paula Couto, 1952: plate 37: 1). This last criterion seems to have been followed by (Muizon et al., 2015) when they coded this character for Carodnia in their matrix. Three oval mental foramina are present below the position of the distal root of p2, p4, and m1 as described for MPEF-PV 564. A rounded fourth smaller foramen is present between the canine and p1, likely corresponding to the one described for C. vieirai beneath the p1 (Paula Couto, 1952). Another similarly rounded foramen is present in the ventral side of the symphysis (see Figure S3 and 3D image available at http://morphobank.org/permalink/?P5193). A small mental foramen was previously mentioned by Paula Couto (1952) for C. vieirai, but positioned on the posterior part of the lower border of the symphysis, beneath the canine, rather than medially as observed in MPEF-PV 8165. Towards the distal portion of the MPEF-PV 8165 dentary, two additional smaller foramina are situated beneath the positions of the m2 roots (Figure 4A). The first is almost vertically aligned with the mental foramina, while the second is located below it but more anteriorly.
The posterior portion of the dentary is fractured distally to the m2, leaving only the ventral edge of the jaw visible at the m3 locus. In this cross-section of the jaw, a relatively large mandibular canal is present at the base. However, at the level of m1, the diameter of the canal is smaller, and in a more central position. The mandibular canal cannot be followed to the back of the jaw since it is broken. The last dentary segment, which bears the distal alveolus of the m3, does not have direct contact with the anterior part.
MPEF-PV 564 represents a fragmentary right jaw, retaining solely the trigonid of m2 and a fragment of the mesial root of m3 (Figure 4D-F). The jaw is broken anterior to the p3, preserving only a severely damaged distal portion of the symphysis. Moving distally along the dentary, a small segment of the coronoid crest is visible on the labial side. Unlike Carodnia vieirai (DGM-333 and DGM-334) and Carodnia inexpectans (Antoine et al., 2015), the extensive damage in the distal part of the jaw hinders the possibility of identifying the anterior foramen of the coronoid canal. In contrast to the alveoli surfaces of the distal root of m2 and m3, those of p3-m1 are broken and eroded, not preserving the real upper margin of the bone. The dentary is robust, with a convex ventral profile below the m2-3 and a straight one mesially. At the m1 position, a small portion of the ventral margin of the dentary is missing. Three mental foramina occur in the mandible, each one located, respectively, below the position of the distal root of p2, the p4 as in Carodnia vieirai (Paula Couto, 1952), and another one under the m1. The mesial fragment of the m2 preserves the trigonid with a clear transverse protolophid with a wear facet that runs through the length of the lophid and slopes distally. Despite the protoconid and the metaconid being almost integrated into the protolophid, they still retain a rounded external surface, making them easily identified in occlusal view. The mesial part of the trigonid is short and there is no trace of paraconid. A blunt paracristid descends abruptly from the mesial border of the protoconid to the base of the tooth and then runs lingually. A precingulid runs from the lingual side mesial to the protoconid base, up to the metaconid but not reaching the labial margin. The precingulid is undulated, thinner and lower on the lingual side, and thicker and higher near the metaconid.
Carodnia karuen sp. nov.
Figure 6A-D
zoobank.org/2705DBC8-B740-4FAD-9859-76DD6357CC5A
Holotype. MLP-PV 90-II-12-121: right jaw fragment somewhat weathered, with m1 nearly complete and p4 with the basal portion and roots partially preserved.
 Etymology. The species name karuen means “old” or “ancient” in the extinct language of the Aónikenk people, who inhabited Patagonia before the formation of the nation-states of Chile and Argentina.
Etymology. The species name karuen means “old” or “ancient” in the extinct language of the Aónikenk people, who inhabited Patagonia before the formation of the nation-states of Chile and Argentina.
Geographic and stratigraphic provenance. Lower fossil level at Cerro Redondo, east of the Chubut province, Argentina (see Simpson, 1935a). Río Chico Group, Peñas Coloradas Formation, lower Paleocene sensu Clyde et al. (2014). Found by J.L. Prado in a Museo de La Plata field trip directed by Rosendo Pascual in 1991.
Diagnosis. Medium-size xenungulate, smaller than Carodnia vieirai and larger than Etayoa bacatensis, and somewhat larger than C. feruglioi. Protolophid of m1 with a strong metaconid and a bit smaller protoconid. Trigonid basin not open lingually but is enclosed by large and well-defined paracristid and metacristid, which are closely pressed against the mesial side of the protolophid in the m1. Precingulid strong and forms a conspicuous shelf as in C. inexpectans but differs from it for being higher and better developed on the lingual side. Hypolophid lower than protolophid, with a strong hypoconid and entoconid. Unlike all other Carodnia species, in which the cristid obliqua is absent in the first molar, C. karuen has a short cristid obliqua extending from the hypoconid up to the base of the distal wall of the trigonid, closer to the lingual side of the protoconid. Also differs from Carodnia feruglioi and C. vieirai for a short entocristid similar to those present in the m3 of C. inexpectans but differing from it in being higher, and almost at the same height as the hypolophid. Broad talonid basin, open lingually.
Description and comparisons. The mesiodistal length of m1 measures 18.21 mm, while the labiolingual dimension is 13.49 mm. The jaw presents two loci preserved, the mesial one with the roots of and part of the enamel of the tooth at the neck is here interpreted as a p4 (Figure 6). The distal one is an m1 with a nearly complete crown structure. The loci assignation rests in the preserved structure of this last tooth, which matches with those present in m1-2 of all Carodnia species. In addition, the roots of the preceding tooth here interpreted as a p4, preserve a distolingual enamel portion of the taloned, which is below the level of the precingulid of the succeeding tooth. This suggests that the more mesial dental piece erupted after the distal one, which agrees with the interpretation of a p4 and m1 for these teeth, following the usual order of eruption in most placental mammals. The lower portion of the jaw is broken, the ramus is robust, high, and at least three times the height of the m1 crown. The lingual side of the dentary is convex, and the labial side is something concave. In contrast to the known jaws of C. feruglioi (UNPSJB-PV 680-1, MPEF-PV 564, and MPEF-PV 8165) there is no foramen in the labial side of the dentary below the mesial root of m1. The p4 is only preserved by a distolingual portion of enamel against the mesial root of the m1, a short piece of dentine at the talonid, and the mesial and distal roots. The roots are mesiodistally compressed and broken on the labial side. The mesial fracture of the jaw exposes the trigonid root of p4, which in frontal view is deep about the high of the preserved dentary.
The m1 is well preserved, with the trigonid somewhat higher, wider, and longer than the talonid. This tooth shows an almost labiolingually oriented bilophodont structure characteristic of Carodniidae. At the mesiolingual side of the trigonid, the enamel is broken and absent from the base of the crown exposing the dentine in the metaconid position. A strong precingulid is sloping down to the labial side and despite being broken, it seems that is not expanded over the labial and lingual side of the tooth. The protolophid like those of the m1 of Carodnia vieirai, is mesially concave in occlusal view, with the cutting plane of the lophid parallel to the one of the hypolophid. The wear facets prld-d and hyld-d are equally developed. The metaconid can be distinguished from the protolophid by a strong enamel edge that bends at the lingual tip of the lophid. The protoconid is not so clearly observed from its occlusal outline. In contrast to C. vieirai or the m2-3 in C. feruglioi and C. inexpectans where no cristids are developed mesial to the protolophid, C. karuen has a conspicuous paracristid and metacristid. The paracristid abruptly descends mesiolingually from the labial end of the protolophid, and progressively bends lingually, to the mesial base of the trigonid. The short metacristid projects mesiolabially from the metaconid almost reaching the lingual end of the paracristid. Mesially the paracristid and metacristid are close together but their contact, closing the trigonid, is inferred since a portion of enamel is missing. At the mesial base of the tooth and below most of the lower part of the paracristid, there is a remnant of a strong precingulid, which leans from the lingual side of the trigonid to a more basal position in the labial side.
The hypolophid is the principal structure of the talonid and with a similar development to other Carodnia species. In the hypoconid sector of the hypolophid, a smooth cristid obliqua descends up to the base of the distal wall of the trigonid more approximately to the protoconid area. The cristid obliqua is smooth and runs mesiolingually, so the short talonid basin, which is open lingually, is also closed labially by it. The entoconid could be identified in the hypolophid by an enlargement at the lingual end, like the one observed in the metaconid but ovoid in outline rather than circular, and with thinner enamel. A very short entocristid runs mesiolingually from the entoconid. The entocristid delimited a short but high lingual edge of the talonid, which in contrast to the protolophid and the hypolophid lacked any wear. Behind the hypolophid base, a short portion of a strong and flat postcingulid is preserved. Even though the cingulid is far from being complete, it appears to have formed a long ledge or step in the distal part of the molar. The anterior root of m1 is stronger than the posterior one, with both roots being mesiodistally compressed and swollen.
There are certain similarities between C. karuen and in general, with the m1 of Carodnia species, and the first molar of the Pyrotheria Carolozittelia tapiroides Ameghino, 1901 (MACN A-10666) and Carolozittelia cf. C. tapiroides (MACN 17985) described by Kraglievich (1957). Both exhibit a bilophodont pattern with a slightly larger protolophid than the hypolophid (usually described in them as “metalophid” and “entolophid”, respectively), the wear facets prld-d and hypld-d, and the presence of enamel crenulations. However, significant differences can be highlighted, particularly with C. karuen, such as the absence in C. tapiroides of a metacristid resulting in a lingually open trigonid, a more robust paracristid in the region of the protoconid, the absence of an oblique cristid, a hypolophid of nearly equal height to the protolophid, and a greatly expanded labial postcingulid forming a large distal ledge. These differences, plus the structure of the third lobe in the m3 of C. tapiroides, set it apart from the typical xenungulate dental pattern of m3 present in Carodniidae and Etayoidae. However, considering a possible phylogenetic link between Xenungulata and Pyrotheria, as explored by some studies (Gelfo et al., 2008; Muizon et al., 2015), C. tapiroides exhibits the highest number of shared dental features among them.
Family ETAYOIDAE Villarroel, 1987
Genus NOTOETAYOA Gelfo, López, and Bond, 2008
Type species. Notoetayoa gargantuai Gelfo, López, and Bond, 2008
Assigned material. UNPSJB PV766 isolated left p2.
 Geographic and stratigraphic provenance. The tooth was found at “Tiro Federal Coronel Pringles” by Mónica Abril in Gaiman locality, Chubut province, and comes from the white to pink arkosic sandstone of the Rio Chico Group. This level was assigned to Las Flores Formations (Chornogubsky et al., 2023).
Geographic and stratigraphic provenance. The tooth was found at “Tiro Federal Coronel Pringles” by Mónica Abril in Gaiman locality, Chubut province, and comes from the white to pink arkosic sandstone of the Rio Chico Group. This level was assigned to Las Flores Formations (Chornogubsky et al., 2023).
Comments. The specimen was preliminarily identified in the collection of UNPSJB as a p2 of Carodnia. Considering the common presence in the Carodnia zone of C. feruglioi and Notoetayoa gargantuai (Gelfo et al., 2008), the similitude of UNPSJB PV766 with the p2 Etayoa bacatensis, the differences with known p2 of C. feruglioi and C. vieirai, and their size, is here assigned to N. gargantuai.
Description and comparisons. The tooth exhibits similarities to the p2 of Etayoa bacatensis (GM-32) than to any other SANU, in terms of its labiolingual compression, presence of two mesiodistally aligned roots, a robust and centrally positioned protoconid, and a relatively short and simplified talonid (Figure 7D-F). These characteristics distinguish it from those of Carodnia feruglioi and C. vieirai. However, it differs from E. bacatensis in its substantially larger size and the absence of cusps in the talonid. The specimen displays a translucent and thin enamel, albeit with surface roughness. Although the roots are missing, there are indications of the presence of two roots, the mesial one larger, as the condition observed in Etayoa (Villarroel, 1987). The tooth is labiolingually compressed, with its main axis oriented mesiodistally (16.25 mm x 8.94 mm) and a strong labial wear facet over the enamel of the distal side, which is similar in position and orientation to the hy-mb wear facet in the p2 of Carodnia feruglioi (MLP-PV 34-V-22-8, Figure 3I), and is here considered homologous to it base on its position since no hypoconid could be identified (Figure 6B-C: hd-b). The protoconid is the only identifiable cusp, slightly rounded at the base, with a flat wear surface (i.e., prcd-h), and the distal and mesial cristids are present. A strong paracristid descends mesially from the apex of the protoconid and in occlusal view, bends a bit lingually before reaching a small elevation from which two structures depart. The first is an almost vertical cristid in the mesial side of the tooth, and the other is a strong and crenulated precingulid that continues in a lingual cingulid and surrounds the base of the protoconid. In lateral view, the paracristid is a high ridge that forms a mesial and convex blade. A distal cristid, partially broken, also descends from the protoconid apex but not with such an abrupt angle as in the paracristid. Distally, it turns lingually down to the base in a postcingulid. The possible contact of the postcingulid and the lingual cingulid cannot be determined since the enamel is broken.
In addition to its larger size, there exist other distinguishing features between the p2 of Notoetayoa and Etayoa bacatensis (Figure 7). These distinctions include a notably more pronounced mesial bifurcation of the paracristid and the presence of a well-developed lingual cingulum in Notoetayoa. Conversely, the distal portion of the p2 in E. bacatensis is wider and displays two cusps as described by Villarroel (1987), a distal and lower cusp, here identified as the hypoconid (Figure 7E-F), and on its lingual side, a faint bump, which, regrettably, could not be corroborated in our photographs or casts. This latter feature may be homologous to the entoconid of the p2 found in Carodnia feruglioi (Figure 3C).
The p2 of Notoetayoa also differs from the p2 of C. feruglioi and C. vieirai. Despite the paracristid in these species being shorter and bifurcates mesially as in Notoetayoa, the precingulid is strong in C. feruglioi and extends in a short portion labially and lingually (Figure 3C), while in C. vieirai (DGM 334) is a sharp ridge with the lingual portion of the precingulid more extended over the base of the protoconid. Probably the vertical cristid here identified in Notoetayoa is homologous to the labial component of the precingulid in Carodnia, but with a modified position consequence of the lingual bend of the paracristid. Other differences with Notoetayoa rest in the further development of the talonid cups in the Carodnia species.
DISCUSSION
The Stratigraphic Position of the Carodnia Zone
The identification of the Carodnia zone not only rests in its paleobiological content but also in the identification of the exact stratigraphic level indicated by Simpson (1935c). Andreis et al. (1975) divided the Rio Chico Formation of Simpson (1933, 1935c) into the Las Violetas and Visser members. The last one recognized as the characteristics of Rio Chico outcrops in Pico Salamanca and Bajo Palangana, where they indicated that the lower contact with the Salamanca Formation is underground but can be observed in the nearby area of Punta Peligro in agreement with the observations made by Brandmayr (1932). This observation aligns with the profiles of Bajo Palangana and Pico Salamanca documented by Feruglio (1949: figures 87, 90), where he specifically noted the presence of the BNS at an elevation of 27 m a.s.l. He correlated this geological feature with the exposures at Punta Peligro, where the “Banco Verde” [Green Bank] (BV) and the BNI are more extensively revealed. Andreis et al. (1975) did not provide a precise stratigraphic location for the Carodnia zone. Their reference was primarily directed towards the presence of fossil bones within the vertebrate assemblage of the Visser Member. Legarreta and Uliana (1994), in their comprehensive restructuring of the Rio Chico unit into a group, designated the lowermost segment as the Peñas Coloradas Formation and attributed the Carodnia zone to it. However, unlike other authors such as Brandmayr (1932), Bordas (1935, 1945), and Simpson (1935c), they did not identify the BNS as a reliable indicator for the presence of Carodnia remains. They also omitted a precise and clear definition of the limits of their Peñas Coloradas new unit. Although in a prior publication, Legarreta et al. (1990) presented a stratigraphic section from north of Comodoro Rivadavia, in which they highlighted the presence of the BNS situated between the lower occurrence of the Gondwanatheria Sudamerica ameghinoi Scillato-Yané and Pascual, 1985 (found in the BNI) and Carodnia. Therefore, based on their earlier work, it can be inferred that they considered the BNS to be either within the Peñas Coloradas Formation or at its base in their subsequent stratigraphic framework. Raigemborn (2008) and Raigemborn et al. (2010) endorse this latter interpretation, based on their observations of the Punta Peligro profile. They indicate that at the base of the Peñas Coloradas Formation, exists an erosive unconformity that has incised into the roof of the BNS, which marks the upper boundary of the “transitional levels” situated above the BV of the Salamanca Formation. Furthermore, they have also identified the so-called “transitional levels” and the BNS beneath the Rio Chico Group, notably in the Las Violetas locality to the north of the basin. Since the publication of Legarreta et al. (1990), the prevailing interpretation has consistently placed the Carodnia zone within the Peñas Coloradas Formation, likely confined to a specific stratigraphic level within this unit (Figure 2). Notably, a U-Pb isotopic date of 61.98 ± 0.041 Myr was obtained from a tuff layer situated in the upper portion of the Peñas Coloradas Formation (Clyde et al., 2014). It is important to note that this tuff layer was identified not at the coast of Chubut province, but rather at Palacio de los Loros and Ormaechea Park, approximately 200 km westward from the coastline (Clyde et al., 2014). These precise dating results led to the correlation of the Carodnia zone with an age of approximately 62 Myr (Woodburne et al., 2014b). This age is notably older than other estimations derived from biochronological inferences (Gelfo et al., 2009; Woodburne et al., 2014a) and is grounded on the stratigraphic position of the Carodnia zone within the Peñas Coloradas Formation.
An alternative interpretation places the Carodnia zone within the lower section of the Las Flores Formation (Krause et al., 2017). In their framework, the Peñas Coloradas Formation is regarded as equivalent to the informal sterile member described by Simpson (1935c), with the BNS serving as its upper boundary (Vera and Krause, 2020). The difference between these two correlations of the Carodnia zone, whether within the Peñas Coloradas or the lower Las Flores formations, lies in the differing interpretations of the outcrops from sea level up to the BNS (Figure 2), the precise delineation of these geological units and their boundaries, as well as variations in the height within the stratigraphic column mentioned in prior literature.
The use of the sea level and guide banks (e.g., BNI, BNS) to stratigraphically locate the mammal fossil levels from Bajo Palangana, place the Carodnia remains between 14 m (Bordas, 1935, 1945; Brandmayr, 1932) and 19-20 m (Cabrera, 1935; Feruglio, 1931) above the BNS (Figure 2). Krause et al. (2017) and Vera and Krause (2020) indicated that Carodnia faunal/zone started 46 m above the base of Rio Chico levels following Feruglio (1949). But to be more accurate, the profile of Bajo Palangana from Feruglio (1949: figure 90) located the Carodnia horizon 46 m above the limit between BV and BNI and indicated that this is 41 m a.s.l. So, the contact between the Salamanca Formation and the BNI at Bajo Palangana is not visible on the coast of Bajo Palangana (Figure S4, Figure S5, Figure S6). Brandmayr (1932) indicated that at the beach southwest of the mouth of the drainage of Bajo Palangana, the BNI almost disappears along a stretch of 1 km. For this reason, Feruglio (1949: figure 90) seems to have correlated the profile of this locality with that of Punta Peligro, where the upper part of Salamanca with its BV and the contact with the BNI outcrop. Despite this, in several works, there is a certain degree of confusion regarding the situation of the BNI in the specific area of Bajo Palangana. Indeed, Simpson, who was carried by Brandmayr to this locality, when located the Carodnia levels indicated that: “In this general area the Salamanca Formation is exposed (along and near the coast, especially at Punta Peligro), capped by the “Banco Negro Inferior” of local workers, here about 8 meters in thickness” (Simpson, 1935c). However, he does not indicate that the BNI does not outcrop at the locality, which was better explained in the previous detailed description of Brandmayr (1932), who stated that the contact between the BV and BNI, is observed in the more northern studied area and Punta Peligro (Table S1).
Stratigraphic references available from Simpson (1935c) merely noted that the Carodnia -bearing levels are situated approximately between 14 to 19 m above the BNS within fine red sandstone strata. These heights correspond to the discoveries of Carodnia remains by Brandmayr and Feruglio respectively. Recently, a lower record was mentioned at 34 m a.s.l. or 7 m above BNS (Gelfo et al., 2021). Brandmayr (1932) provided a more comprehensive description of the level, characterizing it as a part of the “upper Pehuenche”, in a level identified as “h” and composed of clayey, quartzose sandstones, of very fine to medium grain, but with coarse-grained lenses, with clasts reaching 3 cm in diameter (Table S1). Within the lower section, Brandmayr (1932) discerned a pale-green hue, while in the upper segment, reddish colors intermixed with pale green lenticular bands, likely corresponding to the “fine red sandstone” alluded to by Simpson (1935c). Beyond the remnants subsequently attributed to Carodnia, Brandmayr (1932) reported the existence of additional fossils, including bones, notably a scapula (not identified within any of the mentioned collections), and fossilized wood. Other descriptions of this level at Bajo Palangana, (Bordas, 1935, 1945), offer comparatively less comprehensive detail.
The compositional analyses conducted on the Peñas Coloradas Formation within the region (Raigemborn, 2008) and the presence of fossilized wood (Brea and Zucol, 2006) align with the conventional framework, wherein the sterile member identified by Simpson (1935c) as the lower portion of his Rio Chico Formation is integrated into the broader context of the “transitional levels”, extending from the BNI up to the BNS (Raigemborn, 2008; Raigemborn et al., 2010). The question of whether Brandmayr’s (1932) “h” level should be attributed to the Peñas Coloradas or lower levels of the Las Flores Formation is a matter intricately tied to the definition of the lithostratigraphic units involved, specifically their lower and upper boundaries. While this endeavor is beyond the purview of our current study, it is evident that an accurate identification and correlation of all black banks (e.g., BNI, BNS) across the San Jorge Basin is imperative. This becomes especially pertinent when considering the variability in the number of black banks across Patagonia. For instance, in the middle reaches of the Deseado River in Santa Cruz province, only one black guide bank is observed (BN), interpreted as the BNI, incorporating both the BNS and the intervening layer (Roll, 1938). In the Sarmiento Basin, Feruglio (1949) referenced the “Banco Negro Medio” [Middle Black Bank] (BNM) for the lower section of the Rio Chico Formation. However, Feruglio (1949: 28) also mentioned the BNM at 32 meters above the BNI along the north coast from Pico Salamanca, although this might have been an oversight since his and the profiles of Bordas (1945) consistently identified the BNS at this elevation. Regrettably, only the BNI has consistently received recognition from both past and contemporary researchers, largely due to its position overlaying the BV of the Salamanca Formation. As for the other black banks, typically characterized by a similar lithology above and below, a clear determination of their number, interrelation, and distribution across various Patagonian localities remains a crucial task.
At Cerro Redondo, Simpson (1935c) did not make mention of the presence of BNS, instead noting that the first level containing mammal fossils was situated approximately 32 to 33.5 m above the upper boundary of the BNI, equivalent to 36.5 to 38 m above the hill’s base (Figure 2). While Simpson defined the Carodnia zone to encompass the basal faunas of both Bajo Palangana and Cerro Redondo, the absence of the BNS guide level at the latter site, along with the absence of shared taxa between the two locations, introduces some uncertainty regarding the extent of this zone. The only justification offered by Simpson (1935c) for this correlation is based on the similar topographical positioning of these sites and not on the presence of common taxa (e.g., C. feruglioi). It is noteworthy that Carodnia karuen sp. nov. is the first shared genus between the lower levels of Cerro Redondo and Bajo Palangana. Despite these considerations, we should not completely dismiss the possibility of a slightly older age for the lower levels at Cerro Redondo, as suggested by the BNI used as a stratigraphic reference by Simpson (1935b). Nonetheless, the complexities associated with the faults and the intermittent presence of the BNS and BNI guide beds throughout the basin appear to hinder a precise comparison between Bajo Palangana and Cerro Redondo. The presence of C. karuen sp. nov. in the lower levels of Cerro Redondo allows for, at the very least, a partial correlation at the generic level with the Carodnia zone at Bajo Palangana. At a specific level in contrast, several features are more generalized in the dentition of C. karuen sp. nov. than in C. feruglioi. Among them, the better development of the paracristid and metacristid in the trigonid, and the cristid obliqua and entocristid in the talonid, could be interpreted as a more primitive state. On the contrary, C. feruglioi shows a distinct shortening of the trigonid, accompanied by a reduction in the development of both its mesial cristids and those of the talonid. If, indeed, the lower mammalian levels of Cerro Redondo (32-33.5 m above BNI) are a bit older than those of Bajo Palangana, as previously suggested by Simpson (1935c), C. karuen would be the oldest Carodniidae, which already exhibits well-developed protolophid and hypolophid, characteristic of all Carodnia species. These features distinguish it from the basal bunodont pattern that would otherwise facilitate clear phylogenetic relationships with some of the earliest groups within SANU (e.g., Kollpaniinae, Didolodontidae).
In contrast to the debated correlations between the Kibenikhoria and Ernestokokenia chaishoer faunas/zones (Bond and Kramarz, 2021; Vera and Krause, 2020), and their interpretations as Itaboraian and Riochican SALMAs, respectively (Bond et al., 1995; Pascual and Ortiz-Jaureguizar, 2007; Gelfo et al., 2009), the Carodnia zone appears to be delimited, at least within the coastal region of the San Jorge Basin, based on its fossil content. It can be characterized by the first appearance of Xenungulata in Patagonia, represented by the genera Carodnia (C. feruglioi in Bajo Palangana and C. karuen in Cerro Redondo) and Notoetayoa. This zone represents a significant faunal turnover when compared to the earlier Peligran SALMA of the BNI, characterized by a diverse and rich mixture of mammals with various origins (Goin et al., 2022). This includes some taxa linked to Mesozoic Gondwanan lineages (e.g., Monotremata, Dryolestida, and Gondwanatheria), as well as others corresponding to the earliest diversifications of Theria in South America. Notably, these include metatherian (Derorhynchidae and Polydolopimorphia), and the eutherian SANU Notonychopidae and Didolodontidae. It is also noteworthy for the striking Peligran absence of Xenarthra and Notoungulata. In addition to the noticeable shifts in depositional environments between the BNI and the lower Rio Chico Group levels, the presence of Notonychopidae serves as the sole link between these two geographically proximate but biochronologically distinct faunal assemblages. This relation is exemplified at Cerro Redondo by the lesser-known taxon Wainka tshotshe (Simpson, 1935c; Bonaparte and Morales, 1997).
Sexual Dimorphism in Carodnia?
There is a noticeable size variation among mandibles attributed to C. feruglioi and C. vieirai. This difference is particularly evident in the height of the dentary below the molar series, as pointed out by Paula Couto (1952). In the case of specimen DGM 334-M (cast AMNH 49848, Figure S7, Figure S8, Figure S9, Figure S10) assigned to C. vieirai, it was tentatively considered as an immature individual, although this designation lacks explicit justification (Paula Couto, 1952). Presumably, the limited wear on the m3 and the bone size disparity in comparison to the type specimen (DGM 333-M) served as indicative factors for this assessment, a viewpoint shared by Abranches (2012). However, it is important to note that the m3 is a brachydont tooth and is already fully erupted. Additionally, there is no substantial difference in the transverse thickness of the dentary below the m1 when comparing this specimen to the holotype (see Paula Couto, 1952: table 5). The dentary of DGM 334-M is somewhat thicker than the holotype. This observation suggests that the individual represented by DGM 334-M may have already attained its full adult size, making it unlikely to be a juvenile. Paula Couto (1952) mentioned a difference in the height or jaw depth, below the m1, being the dentary of DGM 333-M (holotype) 32% deeper than DGM 334-M (Figure 7, Figure S6, Figure S7, Figure S8, Figure S9).
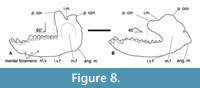 Similar variation in jaw morphology is observed within specimens assigned to C. feruglioi, with notable differences between the jaw fragments MPEF-PV 1872 and MPEF-PV 1874 when compared to MPEF-PV 8165 and MPEF-PV 564. However, since these specimens do not preserve identical portions, direct comparisons are limited to regions distal to m2 (Figure 4, Figure 8; Table 1). While the width of the dentary remains consistent among all specimens, the jaw’s height in the latter two is on average approximately 46% deeper. Despite C. vieirai being a larger species than C. feruglioi, it is noteworthy that both species exhibit specimens displaying these variations, indicating the presence of what appears to be two distinct morphotypes. In one of the morphotypes (A), the jaw is slightly narrower in transverse width but taller or deeper in lateral view. It appears more robust, especially in the case of C. vieirai, where the jaws are nearly fully preserved in both morphotypes, revealing noticeable differences in the mandibular ramus. In the more robust jaws (DGM 333-M) the mandibular ramus is larger and almost rectangular, with a broad and deep masseteric fossa (m.f.) describing an inverted “U”, the incisura mandibulae (i.m.) anteroposteriorly short, demarcated by a prominent processus coronoideus (p. cor.) and a relatively lower processus condylaris (p. con.), which together form an acute angle. In lateral view, the corpus mandibulae is slightly convex ventrally, there is a short concave segment of the margo ventralis (m.v.) below the position of the p3-4 and to the posterior part of the jaw that increases their height up to the incisura vasorum facialium (i.v.f.). There is a wide and convex angulus mandibulae (ang. m.), expanding posteriorly, surpassing a vertical imaginary line distal to the processus condylaris. The processus coronoideus is high and forms an angle of approximately 90° concerning the corpus mandibulae (Figure 8A). It is mesiodistally long and somewhat laterally compressed, in contrast to the much broader and low processus condylaris.
Similar variation in jaw morphology is observed within specimens assigned to C. feruglioi, with notable differences between the jaw fragments MPEF-PV 1872 and MPEF-PV 1874 when compared to MPEF-PV 8165 and MPEF-PV 564. However, since these specimens do not preserve identical portions, direct comparisons are limited to regions distal to m2 (Figure 4, Figure 8; Table 1). While the width of the dentary remains consistent among all specimens, the jaw’s height in the latter two is on average approximately 46% deeper. Despite C. vieirai being a larger species than C. feruglioi, it is noteworthy that both species exhibit specimens displaying these variations, indicating the presence of what appears to be two distinct morphotypes. In one of the morphotypes (A), the jaw is slightly narrower in transverse width but taller or deeper in lateral view. It appears more robust, especially in the case of C. vieirai, where the jaws are nearly fully preserved in both morphotypes, revealing noticeable differences in the mandibular ramus. In the more robust jaws (DGM 333-M) the mandibular ramus is larger and almost rectangular, with a broad and deep masseteric fossa (m.f.) describing an inverted “U”, the incisura mandibulae (i.m.) anteroposteriorly short, demarcated by a prominent processus coronoideus (p. cor.) and a relatively lower processus condylaris (p. con.), which together form an acute angle. In lateral view, the corpus mandibulae is slightly convex ventrally, there is a short concave segment of the margo ventralis (m.v.) below the position of the p3-4 and to the posterior part of the jaw that increases their height up to the incisura vasorum facialium (i.v.f.). There is a wide and convex angulus mandibulae (ang. m.), expanding posteriorly, surpassing a vertical imaginary line distal to the processus condylaris. The processus coronoideus is high and forms an angle of approximately 90° concerning the corpus mandibulae (Figure 8A). It is mesiodistally long and somewhat laterally compressed, in contrast to the much broader and low processus condylaris.
The second morphotype (B) appears to be slightly thicker, but it possesses a more streamlined appearance due to its significantly reduced jaw depth. Notably, in the case of C. vieirai (DGM 334-M), there are additional significant differences when compared to morphotype A of the same species (DGM 333-M). There is a larger and almost rectangular processus coronoideus, mesially projected almost up to the position of the m2, so in lateral view, the angle concerning the corpus mandibulae is conspicuously different to morphotype A, being around 45°. Also differs in the less pronounced incisura mandibulae, which is a long and soft slope from processus coronoideus down to the processus condylaris (Figure 8B). The masseteric fossa and angulus mandibulae are also less developed. The distinctive morphology of morphotype B bears a superficial resemblance to the jaw configuration found in the extant manatee Trichechus manatus Linnaeus, 1758, in contrast to the characteristics observed in morphotype A.
Sexual dimorphism is well-documented in several fossil ungulates, particularly in aspects related to the skull, jaw, canines, and incisors (Gingerich, 1981; Mead, 2000; Mihlbachler, 2007; Chen et al., 2010; Vautrin et al., 2019). However, there is no apparent dimorphism in the size of the canines between the morphotypes identified here for C. vieirai. Instead, there is a noticeable variation in the mesiodistal length of the m2, with DGM 334-M (morphotype B) exhibiting a greater mesiodistal length compared to DGM 333-M (morphotype A). It is not suitable to consider the dental differences in the current discussion of sexual dimorphism without a larger sample to evaluate them. However, the width of the mentioned molars is almost identical (Antoine et al., 2015). The variations observed among the C. vieirai jaws, including the striking differences in the pronounced anterior projection of the processus coronoideus, the size of the diastema between p1 and p2, and the dimensions of the teeth, have already raised questions regarding their possible origin. Among the possible causes, was considered deformation due to taphonomic factors, issues arising during the preparation and restoration processes, and the possibility of the presence of two distinct species or sexual dimorphism (Bergqvist et al., 2008). Although Paula Couto (1952) interpreted some deformation in C. vieirai jaw specimens, at least the similar variation in jaw depth observed in C. feruglioi specimens appears to strengthen the argument against this process. Some of the aspects of jaw morphology in Carodnia can be compared to the sexual dimorphism documented in the North American Miocene rhinoceros Teleoceras major Hatcher, 1894. In this last species a large sample allowed for the documentation of 23 significant characters, among which the depth of the jaw below the m1 was found to be 7.75% greater in males than in females (Mead, 2000). The limited number of remains and their fragmentary state impose statistical limitations on conducting a comprehensive bimodal analysis for morphotypes A and B. Nevertheless, given the evidence presented herein, identifying them as males and females respectively, emerges as one of the plausible hypotheses (Figure 9).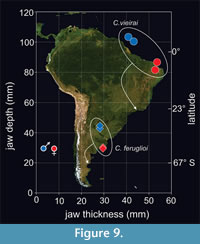 Despite the consistent proportional variation in jaw depth between morphotypes of both species, the difference in thickness is less than 5% among morphotypes of C. feruglioi and around 20% in C. vieirai (see Table 1). Unfortunately, we are unable to elucidate the significance of this variation without a larger, proper sample. Also, no complete jaw specimen of the Patagonian species is available for evaluating the mesial projection of the processus coronoideus observed in morphotype B. A weak point in our argumentation is our lack of knowledge of any other mammal, fossil, or extant, where sexual dimorphism is reflected by this difference in angle in the coronoideus processus of the jaw. Therefore, this characteristic should be considered cautiously until independent evidence supporting or refuting it is available. We are also aware that our limited statistical sample in Carodnia restricts the variables that could be compared with the Teleoceras case. As presented here, the dimorphism observed in Carodnia would exceed more than twice the difference seen in mandibular depth in Teleoceras.
Despite the consistent proportional variation in jaw depth between morphotypes of both species, the difference in thickness is less than 5% among morphotypes of C. feruglioi and around 20% in C. vieirai (see Table 1). Unfortunately, we are unable to elucidate the significance of this variation without a larger, proper sample. Also, no complete jaw specimen of the Patagonian species is available for evaluating the mesial projection of the processus coronoideus observed in morphotype B. A weak point in our argumentation is our lack of knowledge of any other mammal, fossil, or extant, where sexual dimorphism is reflected by this difference in angle in the coronoideus processus of the jaw. Therefore, this characteristic should be considered cautiously until independent evidence supporting or refuting it is available. We are also aware that our limited statistical sample in Carodnia restricts the variables that could be compared with the Teleoceras case. As presented here, the dimorphism observed in Carodnia would exceed more than twice the difference seen in mandibular depth in Teleoceras.
There are alternative explanations to that of possible sexual dimorphism. It could be considered that C. feruglioi and C. vieirai constitute highly variable species and that the individuals found precisely represent the extremes of a range of morphological distribution. Another is that the observed variability within both C. feruglioi and C. vieirai involves the recognition of two distinct species for each, but with minimal or no dental variation: two inhabiting the Paleocene of Patagonia and the others the early Eocene of Itaboraí. In the case of C. feruglioi, all specimens come from the same stratigraphic layer within the Río Chico Group, conventionally dated as late Danian, assuming Peñas Coloradas Formation as the correct stratigraphic level (see above). The minor differences in molar structure, as discussed earlier (see comments on MPEF-PV 1872, 1873, 1874), may not provide substantial support for the existence of a second species. The specimens of C. vieirai, as described by Paula Couto (1952) and considered in the present study, were recovered from a single crevice located near the São José Fault, situated in the southeastern part of the basin. These fossils were originally discovered and excavated in 1949. While another xenungulate specimen, consisting of a left portion of a skull with P4-M3, was mentioned from São José de Itaboraí in previous literature (Souza-Cunha, 1982; Lima and Cunha, 1986), neither its taxonomic assignment nor any comments regarding potential affinities with Carodnia vieirai were provided. Regrettably, this latter specimen has been reported as lost (Bergqvist, 1996), leaving us with only the specimens documented by Paula Couto for analysis. As in the scenario for C. feruglioi, the alternative of classifying the observed variability within C. vieirai as a new taxon would imply the coexistence of two distinct specialized species. These species would share bilophodont molars and exhibit a particular enamel adaptation characterized by the presence of vertical Hunter-Schreger-bands (HSB) in the outer zone and transverse, albeit irregular, HSB in the inner zone (Bergqvist and von Koenigswald, 2017). The recognition of a second taxon will indicate, not only a very close phylogenetic relationship for these Carodnia species but also, the same browser habits and the same niche. In the Itaboraian ecosystem, Carodnia coexisted with other browsers such as the trigonostylopid Astrapotheria Tetragonostylops apthomasi Price and Paula Couto, 1950, the sparnotheriodontid Litopterna Victorlemoinea prototypica Ameghino, 1901, and likely all notoungulates. In this context, all the mentioned ungulates coexisted synchronously and were representative of the Itaboraian locality and SALMA. Using a uniformitarian approach, it becomes challenging to comprehend how they managed to circumvent the competitive exclusion principle (Hardin, 1960). Among African extant savanna ungulates, niche partitioning among browsers primarily hinges on body mass, with a particular focus on their height. This is because they forage at different canopy levels (Skinner and Smithers, 1990; Codron, 2006). However, C. vieirai has been estimated to be approximately 100 cm tall making it the tallest ungulate in the Itaboraí fauna (Bergqvist and von Koenigswald, 2017). Consequently, in contrast to the patterns observed in extant African models, all the browsers in this context likely foraged at the same canopy level. However, this principle must be nuanced and used with caution when extrapolated to the past, as the various variables operating in an ecosystem in the present did not necessarily operate with the same intensity. Counterexamples, such as eight species of rhinos and four proboscideans found in association with Miocene localities in South Asia (Antoine et al., 2013) seem to indicate environmental conditions that accommodated a greater number of large herbivores compared to the present. The main difference with the cases of the morphotypes of Carodnia analyzed here, lies in that if these were different taxa, it seems highly unlikely to have two syntopic species that only differ in jaw depth.
CONCLUSIONS
In this work, and due to the challenges in defining the lithostratigraphic units for the basal part of the Rio Chico Group in Bajo Palangana, we have chosen to maintain a conservative scheme following and adopting for Bajo Palangana the synthesis reached in Raigemborn et al. (2010). The Carodnia zone can be defined as the earliest fossil mammal stratum within the Rio Chico Group. This distinctive zone is characterized by the genus Carodnia and is associated with the presence of Wainka tshotshe, Amphidolops yapa, and Notoetayoa gargantuai. The presence of C. karuen sp. nov. at Cerro Redondo confirms the presence of the genus in the levels correlated by Simpson (1935a) with those of Bajo Palangana, based on stratigraphic positioning. Notably, the Carodnia zone has not only been recognized in these well-known sites but has also been identified in various areas within the San Jorge Gulf Basin. This widespread distribution underscores the genus’s significance as a key indicator in paleontological studies, especially in the basal levels of the Peñas Coloradas Formation, which have traditionally served as reference horizons. This presence distinguishes it from any other SALMA and suggests the possibility of defining a new biocronological unit, using several First Appearance Data (FAD). However, the limited knowledge of the taxa present in these strata and the ongoing debate surrounding the lithostratigraphic units to which the Carodnia zone should be confidently attributed, make it premature - albeit promising - to establish a new post-Peligran and pre-Itaboraian Mammal Age at this time. Further research and consensus about geological aspects are needed in this regard.
The tooth attributed here to Notoetayoa gargantuai represents an isolated element found within the strata from “Tiro Federal” adjacent to the Pan de Azúcar locality in Gaiman. Its significance arises from several factors, including the paucity of taxonomic groups documented at this site, the rarity of specimens attributed to this taxon, and the limited knowledge surrounding the Etayoidae xenungulates. Furthermore, it extends the biochronological range of N. gargantuai beyond the Carodnia zone and, in doing so, broadens our understanding of Xenungulata’s presence in the Riochican SALMA.
The paleobiological inferences presented here regarding the potential sexual dimorphism in C. feruglioi and C. vieirai would benefit from more comprehensive statistical analysis and a larger set of complete specimens. We are aware of the limitations in supporting this hypothesis, given the scarcity of present specimens, and they should be considered not in a definitive manner but rather in this sense until better and greater evidence is available. These considerations should also be extended to assess the applicability of these conclusions to the remaining species within the genus, C. karuen and C. inexpectans. The variations in mandible height and the anterior projection of the coronoid process, as observed in the more complete specimen of C. vieirai (DGM-334), hold notable significance. This becomes particularly intriguing when we consider the distinctive mandibular morphology of C. karuen and the unique labial curvature of the mandibular branch, which suggests a considerable evolutionary adaptability of the jaws among Xenungulata.
ACKNOWLEDGMENTS
We are grateful for the useful critiques and comments provided by P.-O. Antoine and an anonymous reviewer, which helped improve the first version of this manuscript. A special thanks to the Handling Editor, Z. Barkaszi, for his work. We want to thank T. Zanesco, R. Costa, and L.P. Bergqvist for providing us with information related to C. vieirai. We extend our thanks to C.M. Janis due to their comments regarding possible studies of sexual dimorphism; R. S. Gismondi and O. Wilson for the information related to E. bacatensis and C. inexpectans; S. Bargo and M. de Los Reyes from MLP-PV, A. de Sosa Tomas from UNPSJB, E. Ruigómez, and M.E. Pérez from MPEF, and M. Fornasiero from MGP-PD for promptly granting us access to the collections under their care. We are also in debt to M. Abril for sharing valuable information regarding the collections conducted alongside V. Escribano, and to A. Carlini for his generosity in the invitations, both to participate in field-works and the possibility to study fossil specimens. This research was partially supported by UNLP I+D project “Native ungulates from South America and the Antarctic continent: phylogeny, paleobiogeography, and biochronology” - N1016.
REFERENCES
Abranches, C.T.S. 2012. Carodnia vieirai (Mammalia: Xenungulata): uma linhagem evolutiva única? Inferências através de microdesgaste dentário e morfometria geométrica da mandíbula, Universidade Federal do Rio de Janeiro, Rio de Janeiro.
Ameghino, F. 1897. Mammiféres crétacés de l’Argentine (Deuxième contribution à la connaissance de la fauna mammalogique de couches à Pyrotherium). Boletin Instituto Geografico Argentino 18(4-9):406-521.
Ameghino, F. 1901. Notices preliminaries sur des ongulés nouveaux des terrains crétacés de Patagonie. Academia Nacional de Ciencias en Córdoba Boletín, 16:349-426.
Andreis, R.R., Mazzoni, M.M., and Spalletti, L.A. 1975. Estudio estratigráfico y paleoambiental de las sedimentitas terciarias entre Pico Salamanca y Bahía Bustamante, Provincia de Chubut, República Argentina. Revista de la Asociación Geológica Argentina, 30.
Antoine, P.-O., Metais, G., Orliac, M.J., Crochet, J.Y., Flynn, L.J., Marivaux, L., Rajpar, A.R., Roohi G., and Welcomme, J.L. 2013. Chapter 16. Mammalian Neogene Biostratigraphy of the Sulaiman Province, Pakistan, p. 400-422. In Wang, X., Flynn, L.J., and Fortelius, M. (eds.), Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology. Columbia University Press. Columbia University Press.
https://doi.org/10.7312/wang15012-016
Antoine, P.-O., Billet, G., Salas-Gismondi, R., Lara, J.T., Baby, P., Brusset, S., and Espurt, N. 2015. A New Carodnia Simpson, 1935 (Mammalia, Xenungulata) from the Early Eocene of Northwestern Peru and a Phylogeny of Xenungulates at Species Level. Journal of Mammalian Evolution, 22:129-140.
https://doi.org/10.1007/s10914-014-9278-1
Bergqvist, L.P. 1996. Reassociação do pós-crânio às espécies do ungulados da bacia de SJ de Itaboraí (paleoceno), estado do Rio de Janeiro, e filogenia dos “Condylarthra” e ungulados sul-americanos com base no pós-crânio. Universidade Federal do Rio Grande do Sul.
Bergqvist, L.P., Avilla, L.d.S., Porpino, K.O.d., and Abranches, C.T.S. 2008. O enigma de Carodnia vieirai (Mammalia: Xenungulata), da Bacia de S. J. de Itaboraí/RJ (Paleoceno superior) Paleontologia em Destaque. Boletim de Resumos do VI Simpósio Brasileiro de Paleontologia de Vertebrados, Paleontologia em Destaque, (Edição Especial Agosto):41-42.
Bergqvist, L.P. and von Koenigswald, W. 2017. The dentition of Carodnia vieirai (Mammalia: Xenungulata): enamel microstructure and mastication pattern. Palaeontologia Electronica, 20:1-15.
https://doi.org/10.26879/703
Billet, G. 2009. New Observations on the Skull of Pyrotherium (Pyrotheria, Mammalia) and new phylogenetic hypotheses on South American ungulates. Journal of Mammalian Evolution, 17:21-59.
https://doi.org/10.1007/s10914-009-9123-0
Billet, G. 2011. Phylogeny of the Notoungulata (Mammalia) based on cranial and dental characters. Journal of Systematic Palaeontology, 9:481-497.
https://doi.org/10.1080/14772019.2010.528456
Bonaparte, J. and Morales, J. 1997. Un primitivo Notonychopidae (Litopterna) del Paleoceno inferior de Punta Peligro, Chubut, Argentina. Estudios geológicos, 53:263-274.
Bond, M. and Kramarz, A. 2021. Sobre la supuestas ocurrencias compartidas de notoungulados y Astrapoterios en las zonas de Kibenikhoria y Ernestokokenia, Eoceno de Patagonia Central, Argentina. PeAPA, 22(R1):1.
https://doi.org/10.5710/PEAPA.23.03.2022.422
Bond, M., Carlini, A., Goin, F., Legarreta, L., Ortiz-Jaureguizar, E., Pascual, R., and Uliana, M. 1995. Episodes in South American land mammal evolution and sedimentation: testing their apparent concurrence in a Paleocene succession from central Patagonia. VI Congreso Argentino de Paleontología y Bioestratigrafía Actas, p. 47-58.
Bordas, A. 1935. Posición estratigráfica de las capas que contienen restos de mamíferos en el Bajo Palangana (Pico Salamanca), Chubut. Revista de la Sociedad Argentina de Ciencias Naturales, 11:415-422.
Bordas, A. 1945. Notas para el conocimiento de la geologia estratigrafica de algunas zonas de la Patagonia. Anales del Museo de La Patagonia, I:45.
Brandmayr, J. 1932. Informe sobre la región del Bajo Palangana (Pico Salamanca), Yacimientos Petrolíferos Fiscales. Informe inédito, p. 1-12.
Brea, M. and Zucol, A. 2006. Fossil woods of Boraginaceae from the Penas Coloradas Formation (Upper Paleocene), Puerto Visser, Chubut, Argentina. Ameghiniana, 43:139-146.
Cabrera, A. 1935. Estado actual de la cuestión del límite Cretáceo-Terciario en la Argentina desde el punto de vista paleontológico. Instituto del Museo de la Universidad Nacional de La Plata, Obra del Cincuentenario, p. 1-22.
Chen, S., Deng, T., Hou, S., Shi, Q., and Pang, L. 2010. Sexual dimorphism in perissodactyl rhinocerotid Chilotherium wimani from the late Miocene of the Linxia Basin (Gansu, China). Acta Palaeontologica Polonica, 55:587-597.
https://doi.org/10.4202/app.2009.0001
Chornogubsky, L. 2021. Interrelationships of Polydolopidae (Mammalia: Marsupialia) from South America and Antarctica. Zoological Journal of the Linnean Society, 192:1195-1236.
https://doi.org/10.1093/zoolinnean/zlaa143
Chornogubsky, L., Goin, F.J., Ciancio, M.R., Puerta, P., and Krause, M. 2023. Eocene (Ypresian-Lutetian) mammals from Cerro Pan de Azúcar (Gaiman, Chubut Province, Argentina). Historical Biology, 1-11.
https://doi.org/10.1080/08912963.2023.2241054
Cifelli, R. L. 1993. The phylogeny of the native South American ungulates, p. 195-216. In Szalay, F.S., Novacek, M.J., and. McKenna, M.C. (eds.), Mammal Phylogeny, Volume 2: Placentals. Springer Verlag, New York.
Clyde, W.C., Wilf, P., Iglesias, A., Slingerland, R.L., Barnum, T., Bijl, P.K., Bralower, T.J., Brinkhuis, H., Comer, E.E., Huber, B.T., Ibanez-Mejia, M., Jicha, B.R., Krause, J.M., Schueth, J.D., Singer, B.S., Raigemborn, M.S., Schmitz, M.D., Sluijs, A., and Zamaloa, M.d.C. 2014. New age constraints for the Salamanca Formation and lower Rio Chico Group in the western San Jorge Basin, Patagonia, Argentina: Implications for Cretaceous-Paleogene extinction recovery and land mammal age correlations. Geological Society of America Bulletin, 126:289-306.
https://doi.org/10.1130/b30915.1
Codron, D. 2006. The Ecological and Evolutionary Significance of Browsing and Grazing in Savanna Ungulates. University of Cape Town.
Croft, D.A., Gelfo, J.N., and López, G.M. 2020. Splendid innovation: the extinct South American native ungulates. Annual Review of Earth and Planetary Sciences, 48:259-290.
https://doi.org/10.1146/annurev-earth-072619-060126
Erfurt, J. and Haubold, H. 1989. Artiodactyla aus den eozänen Braunkohlen des Geiseltales bei Halle (DDR). Palaeovertebrata, 19:29.
Feruglio, E. 1931. Nuevas observaciones geológicas en la Patagonia Central Contribuciones YPF, Primera Semana de Estudios Geográficos. Buenos Aires, p. 879-920.
Feruglio, E. 1949. Descripción geológica de la Patagonia. Dirección General de Yacimientos Petrolíferos Fiscales, Buenos Aires.
Gelfo, J.N., López, G.M., and Bond, M. 2008. A new Xenungulata (Mammalia) from the Paleocene of Patagonia, Argentina. Journal of Paleontology, 82:329-335.
https://doi.org/10.1666/06-099.1
Gelfo, J.N., Goin, F.J., Woodburne, M.O., and Muizon, C.D. 2009. Biochronological Relationships of the Earliest South American Paleogene Mammalian Faunas. Palaeontology, 52:251-269.
https://doi.org/10.1111/j.1475-4983.2008.00835.x
Gelfo, J.N., García-López, D.A., and Bergqvist, L.P. 2020. Phylogenetic relationships and palaeobiology of a new xenungulate (Mammalia: Eutheria) from the Palaeogene of Argentina. Journal of Systematic Palaeontology, 18:993-1007.
https://doi.org/10.1080/14772019.2020.1715496
Gingerich, P.D. 1981. Variation, sexual dimorphism, and social structure in the early Eocene horse Hyracotherium (Mammalia, Perissodactyla). Paleobiology, 7:443-455.
Goin, F., Gelfo, J., Ortiz-Jaureguizar, E., Forasiepi, A., Candela, A., Vieytes, C., and Cladera, G. 2022. Mamíferos del Banco Negro Inferior, Formación Salamanca, Cuenca del Golfo San Jorge. Relatorio: Geología y Recursos Naturales de la Provincia de Chubut, p. 1029-1053.
Hardin, G. 1960. The competitive exclusion principle: an idea that took a century to be born has implications in ecology, economics, and genetics. Science, 131:1292-1297.
Hatcher, J.B. 1894. A median horned Rhinoceros from the Loup Fork Beds of Nebraska. The American Geologist, 13(3):149-150.
Hershkovitz, P. 1971. Dental Morphology and Evolution. Basic Crown Tooth Pattern and Cusp Homologies of Mammalian Teeth. University of Chicago Press.
Hershkovitz, P. 1990. Titis, New World monkeys of the genus Callicebus (Cebidae, Platyrrhini): a preliminary review. Fieldiana: Zoology, New Series, 55:1-109.
Kalthoff, D.C. 2011. Microstructure of dental hard tissues in fossil and recent xenarthrans (Mammalia: Folivora and Cingulata). Journal of Morphology, 272:641-661.
https://doi.org/10.1002/jmor.10937
Kraglievich, J.L. 1957. Sobre la presencia de Carolozittelia cf. tapiroides Ameghino, en la Fm. La Balsa de Neuquén. Ameghiniana, 1:16-26.
Krause, J.M., Clyde, W.C., Ibañez-Mejía, M., Schmitz, M.D., Barnum, T., Bellosi, E.S., and Wilf, P. 2017. New age constraints for early Paleogene strata of central Patagonia, Argentina: Implications for the timing of South American Land Mammal Ages. Geological Society of America Bulletin, 129:886-903.
https://doi.org/10.1130/b31561.1
Legarreta, L., Uliana, M., and Torres, M. 1990. Secuencias deposicionales cenozoicas de Patagonia Central: sus relaciones con las asociaciones de mamíferos terrestres y episodios marinos epicontinentales. 3 Simposio del Terciario de Chile, p. 135-176.
Legarreta, L. and Uliana, M.A. 1994. Asociaciones de fósiles y hiatos en el Supracretácico-Neógeno de Patagonia: una perspectiva estratigráfico-secuencial. Ameghiniana, 31:257-281.
Lima, M.R.d. and Cunha, F.L.d.S. 1986. Análise palinológica de um nível linhítico da Bacia de São José de Itaboraí, Terciário do Estado do Rio de Janeiro, Brasil. Anais da Academia Brasileira de Ciências, 58:579-588.
Linnaeus, C. 1758. Systema Naturae Per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, Cum Characteribus, Differentiis, Synonymis, Locis. 10th ed. Laurentius Salvius, Stockholm.
Lucas, S.G. 1993. Pantodonts, tillodonts, uintatheres, and pyrotheres are not ungulates, p. 182-194. In Szalay, F.S., Novacek, M.J., and McKenna, M.C. (eds.), Mammal Phylogeny: Placentals. Springer Science & Business Media.
McKenna, M. 1980. Early history and biogeography of South America’s extinct land mammals, p. 43-77. In Ciochon, R.L. and Chiarelli, A.B. (eds.), Evolutionary Biology of the New World Monkeys and Continental Drift. Advances in Primatology. Springer, Boston, MA.
https://doi.org/10.1007/978-1-4684-3764-5_2
Mead, A.J. 2000. Sexual dimorphism and paleoecology in Teleoceras, a North American Miocene rhinoceros. Paleobiology, 26:689-706.
https://doi.org/10.1666/0094-8373(2000)0262.0.CO;2
Mihlbachler, M.C. 2007. Sexual dimorphism and mortality bias in a small Miocene North American Rhino, Menoceras arikarense: insights into the coevolution of sexual dimorphism and sociality in rhinos. Journal of Mammalian Evolution, 14:217-238.
https://doi.org/10.1007/s10914-007-9048-4
Muizon, C.D., Billet, G., Argot, C., Ladevèze, S., and Goussard, F. 2015. Alcidedorbignya inopinata, a basal pantodont (Placentalia, Mammalia) from the early Palaeocene of Bolivia: anatomy, phylogeny, and palaeobiology. Geodiversitas, 37:397-634.
https://doi.org/10.5252/g2015n4a1
Osborn, H.F. 1907. Evolution of Mammalian Molar Teeth: To and from the Triangular Type Including Collected and Revised Researches Trituberculy and New Sections on the Forms and Homologies of the Molar Teeth in the Different Orders of Mammals. Biological Studies and Addresses, 1. Macmillan, New York.
Pascual, R. and Ortiz-Jaureguizar, E. 2007. The Gondwanan and South American episodes: two major and unrelated moments in the history of the South American mammals. Journal of Mammalian Evolution, 14:75-137.
https://doi.org/10.1007/s10914-007-9039-5
Paula Couto, C.d. 1952. Fossil mammals from the beginning of the Cenozoic in Brazil. Condylarthra, Litopterna, Xenungulata, and Astrapotheria. Bulletin of the American Museum of Natural History, 99:355-394.
Price, L.I. and Paula-Couto, C.D. 1950. Vertebrados fósseis do Eoceno na Bacia calcárea de Itaboraí. In Congresso Panamericano de Engenharia de Minas e Geologia Vol. 2, pp. 149-173.
Raigemborn, M.S. 2008. Estudio estratigráfico, sedimentológico y composicional de las sedimentitas del Terciario Inferior (Grupo Río Chico) en el sector Sudoriental del Chubut Extraandino. Universidad Nacional de La Plata.
Raigemborn, M.S., Krause, J.M., Bellosi, E., and Matheos, S.D. 2010. Redefinición estratigráfica del Grupo Río Chico (Paleógeno Inferior), en el norte de la cuenca del golfo San Jorge, Chubut. Revista de la Asociación Geológica Argentina, 67:239-256.
Roll, A. 1938. Estudio geológico de la zona al sur del curso medio del Río Deseado. Boletín de Informaciones Petroleras, 15:66.
Schoch, R. and Lucas, S.G. 1985. The phylogeny and classification of the Dinocerata (Mammalia, Eutheria). Bulletin of the Geological Institutions of the University of Uppsala, 11:31-58.
Schultz, J.A., Menz, U., Winkler, D.E., Schulz-Kornas, E., Engels, S., Kalthoff, D.C., von Koenigswald, W., Ruf, I., Kaiser, T.M., and Kullmer, O. 2018. Modular wear facet nomenclature for mammalian post-canine dentitions. Historical Biology, 30:30-41.
https://doi.org/10.1080/08912963.2017.1302442
Scillato-Yané, G.J.S. and Pascual, R. 1985. Un peculiar Xenarthra del Paleoceno medio de Patagonia (Argentina). Su importancia en la sistemática de los Paratheria. Ameghiniana, 21(2-4):172-176.
Simpson, G.G. 1933. Stratigraphic nomenclature of the early Tertiary of central Patagonia. American Museum Novitates, 644:1-13.
Simpson, G.G. 1935a. Descriptions of the oldest known South American mammals, from the Río Chico Formation. American Museum Novitates, 793:1-25.
Simpson, G.G. 1935b. Early and middle Tertiary geology of the Gaiman region, Chubut, Argentina. American Museum Novitates, 775:1-29.
Simpson, G.G. 1935c. Occurrence and relationships of the Río Chico fauna of Patagonia. American Museum Novitates, 818:1-21.
Simpson, G.G. 1967. The Beginning of the Age of Mammals in South America. Part 2. Systematics: Notoungulata, concluded (Typotheria, Hegetotheria, Toxodonta, Notoungulata incertae sedis), Astrapotheria, Trigonostylopoidea, Pyrotheria, Xenungulata, Mammalia incertae sedis. Bulletin of the American Museum of Natural History, 137:1-260.
Skinner, J. and Smithers, R. 1990. The Mammals of the Southern African Subregion. University of Pretoria, Pretoria, South Africa.
Souza-Cunha, F. 1982. A presença inédita de um mamífero Xenungulata no calcário da Bacia de São José de Itaboraí, RJ. Anais da Academia Brasileira de Ciências, 54:754-755.
Stirton, R. 1941. Development of characters in horse teeth and the dental nomenclature. Journal of Mammalogy, 22:434-446.
https://doi.org/10.2307/1374940
Takai, M. and Anaya, F. 1996. New specimens of the oldest fossil platyrrhine, Branisella boliviana, from Salla, Bolivia. American Journal of Physical Anthropology: The Official Publication of the American Association of Physical Anthropologists, 99:301-317.
https://doi.org/10.1002/(SICI)1096-8644(199602)99:2%3C301::AID-AJPA7%3E3.0.CO;2-0
Van Valen, L.M. 1966. Deltatheridia, a new order of mammals Bulletin of the American Museum of Natural History, 132:1-126.
Vautrin, Q., Tabuce, R., Laurent, Y., Vidalenc, D., and Lihoreau, F. 2019. Intraspecific variation of Eolophiodon laboriense, a basal Lophiodontidae (Mammalia, Perissodactyla) from the early Eocene of Southern France. Geobios, 53:51-63.
https://doi.org/10.1016/j.geobios.2019.02.005
Vera, B. and Krause, J.M. 2020. New insights into the Kibenikhoria and Ernestokokenia faunas and the implications for the early Eocene of Patagonia, Argentina. Journal of Vertebrate Paleontology, 40:e1772801.
https://doi.org/10.1080/02724634.2020.1772801
Vera, B., Fornasiero, M., and Del Favero, L. 2020. New data on Carodnia feruglioi (Carodniidae, Xenungulata) from the early Eocene of Patagonia (Argentina). Ameghiniana, 57:566-581.
https://doi.org/10.5710/AMGH.02.08.2020.3349
Villarroel, C. 1987. Características y afinidades de Etayoa n. gen., tipo de una nueva familia de xenungulata (Mammalia) del Paleoceno medio de Colombia. Comunicaciones Paleontológicas del Museo de Historia Natural de Montevideo, 19(1): 241-253.
Woodburne, M.O., Goin, F.J., Bond, M., Carlini, A.A., Gelfo, J.N., López, G.M., Iglesias, A., and Zimicz, A.N. 2014a. Paleogene land mammal faunas of South America; a response to global climatic changes and indigenous floral diversity. Journal of Mammalian Evolution, 21:1-73.
https://doi.org/10.1007/s10914-012-9222-1
Woodburne, M.O., Goin, F.J., Raigemborn, M.S., Heizler, M., Gelfo, J.N., and Oliveira, E.V. 2014b. Revised timing of the South American early Paleogene land mammal ages. Journal of South American Earth Sciences, 54:109-119.
https://doi.org/10.1016/j.jsames.2014.05.003
