

 Investigation of claims of late-surviving pterosaurs: the cases of the winged dragons of Belon, Aldrovandi, and Cardinal Barberini
Investigation of claims of late-surviving pterosaurs: the cases of the winged dragons of Belon, Aldrovandi, and Cardinal Barberini
Article number: 17.3.41A
https://doi.org/10.26879/461
Copyright Palaeontological Association, November 2014
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 10 February 2014. Acceptance: 24 October 2014
{flike id=967}
ABSTRACT
Here we investigate claims that pterosaurs survived into the sixteenth and seventeenth centuries. In 1557, 1640, and 1651 the European naturalists Pierre Belon, Ulisse Aldrovandi, and Giovanni Faber, respectively, published illustrations of winged, bipedal specimens that had been stuffed and mounted. Some recent young-Earth creationist authors claim that these specimens were recently-killed pterosaurs. The drawings and descriptions are detailed enough to test the pterosaur hypothesis as well as the alternative hypothesis that the specimens are taxidermic composites of parts of different animals. However, before now, no one has investigated these three cases or attempted to test these hypotheses. Here we report an investigation in which these hypotheses are tested. By comparing the specimens with pterosaurs, we found that in all three specimens, all regions of the body are inconsistent with pterosaur anatomy. Comparison with extant animals reveals that Belon’s and Aldrovandi’s dragons are decapitated snakes with attached mammal heads. Their wings are the pectoral fins of flying gurnards (Dactylopterus volitans). Their “legs” are the forelimbs of rabbits or canids in reptile-skin sleeves. The dragon illustrated by Faber and owned by Cardinal Francesco Barberini includes the skull of a weasel (Mustela nivalis), the belly skin of a snake, the dorsal and lateral skin of a lizard, and the tail skeleton of an eel (Anguilla anguilla). These hoaxes now join the list of discredited “proofs” of human-pterosaur coexistence.
Phil Senter. Department of Biological Sciences, Fayetteville State University, 1200 Murchison Road, Fayetteville, North Carolina 28301, U.S.A. psenter@uncfsu.edu
Darius M. Klein. Department of Comparative Literature, Ballantine Hall 914, Indiana University, Bloomington, Indiana 47405, U.S.A. damaklei@umail.iu.edu
Keywords: Pterosauria; young-Earth creationism; Ulisse Aldrovandi; Pierre Belon; Giovanni Faber; hoax
Final citation: Senter, Phil and Klein, Darius M. 2014. Investigation of claims of late-surviving pterosaurs: the cases of Belon’s, Aldrovandi’s, and Cardinal Barberini’s winged dragons. Palaeontologia Electronica 17.3.41A: 1-19. https://doi.org/10.26879/461
palaeo-electronica.org/content/2014/967-late-surviving-pterosaurs
INTRODUCTION
Here we investigate claims that certain taxidermic specimens displayed during the European Renaissance were carcasses of recently-killed pterosaurs. These claims were made by Bill Cooper (1992), John Goertzen (1993, 1998), Dave Woetzel (2006, 2012), and James Gilmer (2011) in order to cast doubt on the separation of humans and pterosaurs by millions of years and, by extension, to cast doubt on evolutionary theory. Although the specimens no longer exist, sixteenth- and seventeenth-century naturalists who examined the specimens described and illustrated them. These descriptions and illustrations contain sufficient detail to allow tests of the pterosaur hypotheses by means of comparison with pterosaur anatomy.
 In 1557 French naturalist Pierre Belon published a book of drawings of organisms that he had seen during a voyage to various parts of the Middle East (Belon, 1557). He included those drawings in a subsequent book in which he described the journey and the people, animals, and plants that he encountered (Belon, 1588). In Egypt he saw “bodies, embalmed and all complete, of certain winged serpents, which had feet, that they say fly from the part of Arabia in Egypt, of which one can see a portrait above...” (Belon 1588, translated from French by P.S.). His drawing, published in both the 1557 and the 1588 books, is reproduced here (Figure 1.1). His mention of flying snakes entering Egypt from Arabia is a reference to a passage by Herodotus in Histories 2.75, 2.76, and 3.107 (Hare, 2010). Herodotus said that, having heard of Egyptian winged snakes, he traveled to a place near Buto in Arabia to ask about them. Buto was a city in the Nile delta, and the ancient Greeks used the term “Arabia” for present-day Arabia plus northeast Egypt up to the Nile (Anthon, 1878). Herodotus did not claim to have seen any of the winged snakes but said that he was told that they fly toward Egypt from Arabia each spring and are killed by ibises before reaching Egypt. This story was repeated by later classical authors (Senter, 2013) and afterward by Renaissance-age encyclopedists (e.g., Gessner, 1589; Aldrovandi, 1640) and was well-known to naturalists in Belon’s day. However, Belon was the first to claim to have seen the animals in Egypt and the first to illustrate one.
In 1557 French naturalist Pierre Belon published a book of drawings of organisms that he had seen during a voyage to various parts of the Middle East (Belon, 1557). He included those drawings in a subsequent book in which he described the journey and the people, animals, and plants that he encountered (Belon, 1588). In Egypt he saw “bodies, embalmed and all complete, of certain winged serpents, which had feet, that they say fly from the part of Arabia in Egypt, of which one can see a portrait above...” (Belon 1588, translated from French by P.S.). His drawing, published in both the 1557 and the 1588 books, is reproduced here (Figure 1.1). His mention of flying snakes entering Egypt from Arabia is a reference to a passage by Herodotus in Histories 2.75, 2.76, and 3.107 (Hare, 2010). Herodotus said that, having heard of Egyptian winged snakes, he traveled to a place near Buto in Arabia to ask about them. Buto was a city in the Nile delta, and the ancient Greeks used the term “Arabia” for present-day Arabia plus northeast Egypt up to the Nile (Anthon, 1878). Herodotus did not claim to have seen any of the winged snakes but said that he was told that they fly toward Egypt from Arabia each spring and are killed by ibises before reaching Egypt. This story was repeated by later classical authors (Senter, 2013) and afterward by Renaissance-age encyclopedists (e.g., Gessner, 1589; Aldrovandi, 1640) and was well-known to naturalists in Belon’s day. However, Belon was the first to claim to have seen the animals in Egypt and the first to illustrate one.
Belon’s winged serpents received much subsequent attention. The German naturalist Conrad Gessner mentioned Belon’s account, reproduced Belon’s drawing in an encyclopedic volume on snakes (Gessner, 1589) (Figure 1.2), and mentioned that the common people called such specimens dragons. The Italian naturalist Ulisse Aldrovandi (1640) also mentioned Belon’s winged serpents and included an illustration of one in a section on winged dragons in his posthumous encyclopedia of snakes, Serpentum et Draconum Historiae (Figure 1.3). The English author Edward Topsell copied Gessner’s copy of Belon’s picture in his own encyclopedic volume on snakes, but did not mention Belon (Topsell, 1608).
Cooper (1992) reproduced Topsell’s copy of Belon’s drawing in an article in Creation Ex Nihilo Technical Journal and cited it as an example of a drawing of a dinosaur. Unaware that the original eyewitness (Belon) had seen the animals up close, in his figure caption he asked, “Could the rather ridiculous looking wings of Topsell’s monster have come about through the original eyewitness account having mistaken from a distance the markings of armour-plating for wings?...The wings apart, note the woodcut’s surprising similarity to a modern reconstruction of a Tyrannosaurus ” (Cooper, 1992). Goertzen corrected Cooper in a letter to the editor in the following issue of the journal, identifying the picture as a copy of Belon’s illustration of a winged specimen that Belon had observed, and implying that it was a pterosaur (Goertzen, 1993). In a subsequent publication, Goertzen identified Belon’s winged animal as the pterosaur Dimorphodon macronyx (Goertzen, 1998). Woetzel (2006) also claimed that Belon’s winged serpents were pterosaurs.
Gilmer (2011) later expressed a unique view on Belon’s flying serpents. He reproduced Aldrovandi’s (1640) copy of Belon’s winged dragon illustration and called it a “land dinosaur”. In his view, the appendages that appeared to be wings were actually a frill that was spread for defensive or aggressive display, as in the frilled lizard (Chlamydosaurus kingii), or were appendages that were “moved like wings to lighten the creature’s body for faster running.”
Aldrovandi had his illustrators produce a series of paintings of which line-drawing copies were included in his books. This series of paintings, which is now called the Tavole di animali, remains unpublished in print, but photos of the entire collection are now posted online (Biblioteca Universitaria di Bologna, 2013). Painting 140 of volume 4 of the Tavole (Figure 1.4) is of a specimen that Aldrovandi described, with an accompanying drawing (Figure 1.5), in Serpentum et Draconum Historiae (Aldrovandi, 1640). According to Aldrovandi’s description, “in the year of our Lord 1600, a true mummified African [ aethiopicus : in reference to lands south of Egypt] dragon was given as gift by Francisco Centensis to the most illustrious Ulisse Aldrovandi...it had, moreover, five prominent and conspicuous protuberances on its back, which were lacking in the dragon of Belon; it also had two feet armed with claws, and was depicted with small ears. The entire body was decorated with green and blackish scales. It bore two wings suitable for flying, and a long and flexible tail, straight with dull yellow scales, such as were visible on the stomach and throat. The mouth was armed with sharp teeth. The lower part of the head was flat next to the small ears. The pupils of the eyes were black with a pale yellow circle. Finally, the two nostrils were visible and open” (Aldrovandi, 1640, translated from Latin by D.K.).
Aldrovandi insisted that his illustrators illustrate specimens directly, depicting what they saw in front of them so as to ensure accuracy rather than taking artistic liberties (Olmi, 2007). The illustration of the specimen that Centensis gave him is therefore likely painted directly from the original specimen. Aldrovandi’s published drawing (Figure 1.3) of Belon’s specimen bears suspicious resemblance to that of the specimen from Centensis (Figure 1.5). This can probably be explained by Aldrovandi’s insistence on depicting actual specimens. Because Belon did not purchase a winged serpent specimen (or, at least, did not record having done so) but simply illustrated one, the illustrator did not have a specimen from Belon to illustrate. In order to draw directly from a specimen, therefore, the illustrator had little choice but to redraw the specimen from Centensis and modify the drawing to match Belon’s illustration. It appears that Aldrovandi’s drawing of Belon’s specimen is really of the Centensis specimen with the five dorsal lumps omitted and with the tail configured to match Belon’s illustration.
Neither Cooper, Goertzen, Gilmer, nor Woetzel specifically mentioned Aldrovandi’s specimen. However, it was included by Aldrovandi in a chapter on winged dragons, which Woetzel (2006) mentioned. Woetzel claimed that the winged animals to which Aldrovandi referred were pterosaurs.
The third alleged pterosaur to be considered here is a specimen owned by Cardinal Francesco Barberini (1597 - 1679), nephew of Pope Urban VIII (reigned 1623 - 1644). The specimen, which Barberini considered a dragon, was a gift from King Louis XIII of France (Bartholin, 1678). Barberini was a patron of certain members of the Lyncean Academy, a scientific association that produced several noteworthy monographs (Freedberg, 2002). One of these was the multi-authored Rerum Medicarum Novae Hispaniae Thesaurus ( Medicinal Treasures of New Spain), on the natural history of Mexico (Lyncean Academy, 1651). Under political pressure (Freedberg, 2002), Lyncean anatomist Giovanni Faber added a description of Barberini’s dragon to Tesoro Messicano (Mexican Treasures), a section in Rerum Medicarum Novae Hispaniae Thesaurus. An English translation by D.K. of Faber’s Latin description of the specimen is given here in Appendix.
Faber’s description of Cardinal Barberini’s dragon included a detailed drawing of the specimen (Figure 1.6). The German Jesuit scholar Athanasius Kircher (1601/2 - 1680) published another drawing of the specimen in the dragon section of his 1664 book Mundus Subterraneus (Subterranean World), a classic of geological literature (Kircher, 1664) (Figure 1.7).
Woetzel (2012) included Faber’s drawing (misidentified as Kircher’s) of Cardinal Barberini’s dragon in a list of several Renaissance-age illustrations that he claimed were depictions of pterosaurs. He claimed specifically that it “resembles the rhamphorhynchoid pterosaurs” and used it to support the idea that live pterosaurs roamed Europe in recent centuries (Woetzel, 2012).
The illustrations and descriptions of the three specimens by Belon, Aldrovandi, and Faber are detailed enough to test the hypothesis that they are pterosaurs. Here, we compare the anatomy of these specimens with pterosaur anatomy to test this hypothesis. We also advance the alternative hypothesis that the three specimens are taxidermic composites and use observations of the anatomy of extant animals to test various hypotheses as to which extant species are represented in the various parts of the three specimens.
COMPARISON WITH PTEROSAURS
 In all three dragons, wing anatomy is very different from that of pterosaurs on several counts. Firstly, in each of the three dragons the wing is shorter than the torso, whereas in a pterosaur the wing is much longer than the torso, usually four or more times the length of the torso (Figure 2). Secondly, Belon’s dragon has six internal struts that radiate from the shoulder (Figure 1.1), Aldrovandi’s has five (Figure 1.4), and according to Faber’s description Bartholin’s dragon has three (Appendix). In contrast, pterosaur wings have only one internal strut, the greatly elongated fourth finger, which is situated along the leading edge of the wing (Figure 2). Thirdly, the wings of the three dragons have scalloped edges, as in a bat (Figure 1), whereas pterosaur wings do not (Wellnhofer, 1991; Unwin, 2006) (Figure 2.3). Fourthly, the wings of Belon’s and Aldrovandi’s dragons have no protruding fingers or claws (Figure 1.1-5), and Barberini’s has two miniscule claws (Figure 1.6-7), whereas a pterosaur wing has three complete, clawed fingers that protrude from the leading edge of the wing (Wellnhofer, 1991; Unwin, 2006) (Figure 2.1-2). Fourthly, the wings of the three dragons are narrow at the base and expand in width distally (Figure 1), whereas pterosaur wings are wide at the base and taper to a point distally (Wellnhofer, 1991; Unwin, 2006) (Figure 2.3). In addition, the wings of Belon’s and Aldrovandi’s dragons originate from the same spot as the root of the “leg” (Figure 1.1-5) whereas in a pterosaur the shoulder and hip are widely separated (Wellnhofer, 1991; Unwin, 2006) (Figure 2.1-2, 2.4). In Barberini’s dragon the root of the wing is posterior to that of the “leg” and posterior to the ribcage (Figure 1.6-7), whereas in pterosaurs the shoulder is at the anterior end of the ribcage, far anterior to the hip (Wellnhofer, 1991; Unwin, 2006) (Figure 2.1-2, 2.4).
In all three dragons, wing anatomy is very different from that of pterosaurs on several counts. Firstly, in each of the three dragons the wing is shorter than the torso, whereas in a pterosaur the wing is much longer than the torso, usually four or more times the length of the torso (Figure 2). Secondly, Belon’s dragon has six internal struts that radiate from the shoulder (Figure 1.1), Aldrovandi’s has five (Figure 1.4), and according to Faber’s description Bartholin’s dragon has three (Appendix). In contrast, pterosaur wings have only one internal strut, the greatly elongated fourth finger, which is situated along the leading edge of the wing (Figure 2). Thirdly, the wings of the three dragons have scalloped edges, as in a bat (Figure 1), whereas pterosaur wings do not (Wellnhofer, 1991; Unwin, 2006) (Figure 2.3). Fourthly, the wings of Belon’s and Aldrovandi’s dragons have no protruding fingers or claws (Figure 1.1-5), and Barberini’s has two miniscule claws (Figure 1.6-7), whereas a pterosaur wing has three complete, clawed fingers that protrude from the leading edge of the wing (Wellnhofer, 1991; Unwin, 2006) (Figure 2.1-2). Fourthly, the wings of the three dragons are narrow at the base and expand in width distally (Figure 1), whereas pterosaur wings are wide at the base and taper to a point distally (Wellnhofer, 1991; Unwin, 2006) (Figure 2.3). In addition, the wings of Belon’s and Aldrovandi’s dragons originate from the same spot as the root of the “leg” (Figure 1.1-5) whereas in a pterosaur the shoulder and hip are widely separated (Wellnhofer, 1991; Unwin, 2006) (Figure 2.1-2, 2.4). In Barberini’s dragon the root of the wing is posterior to that of the “leg” and posterior to the ribcage (Figure 1.6-7), whereas in pterosaurs the shoulder is at the anterior end of the ribcage, far anterior to the hip (Wellnhofer, 1991; Unwin, 2006) (Figure 2.1-2, 2.4).
The heads of the three dragons are different from those of pterosaurs. All three have protruding ear flaps (Figure 1). Numerous pterosaur specimens with soft-tissue impressions are known, and none have ear flaps (Wellnhofer, 1991; Unwin, 2006). The mandible of Barberini’s dragon has a high, prominent coronoid process (Figure 1.5-6). A coronoid process is absent in most pterosaurs (Wellnhofer, 1991; Unwin, 2006) and is small and low in the few pterosaurs that have it (Dalla Vecchia, 2003). Barberini’s dragon also has a heterodont dentition in which teeth are differentiated into incisors, canines, and molariform cheek teeth (Appendix), a situation that is not present in any pterosaur (Wellnhofer, 1991; Unwin, 2006).
The tails of the three dragons are different from those of pterosaurs. All three specimens have long, curled tails (Figure 1), whereas in long-tailed pterosaurs the tail is prevented from bending by a network of thin, bony struts (Wellnhofer, 1991; Unwin, 2006) (Figure 2.5). Such a network is conspicuously absent in the tail skeleton of Cardinal Barberini’s dragon, the only one of the three dragons whose tail skeleton is exposed (Figure 1.6-7). Also, the tail vertebrae of Barberini’s dragon are short relative to their height (Figure 1.6-7), whereas those of long-tailed pterosaurs are much longer than tall (Figure 2.5).
The “legs” of the three dragons are unlike those of pterosaurs. Their “knees” are bent in the wrong direction, revealing that they are actually elbows. These three dragons therefore do not have hindlimbs but instead have forelimbs, and their forelimbs are not incorporated into the wings. Pterosaurs, in contrast, do have hindlimbs, and their wings are forelimbs with patagia (flight membranes).
The illustrations show that the skin of Belon’s and Aldrovandi’s dragons are covered in scales (Figure 1.1-5), as is the skin of Barberini’s dragon according to Faber’s description (Appendix). In contrast, pterosaur skin is not scaly but is covered in short, hairlike filaments (Wellnhofer, 1991; Unwin, 2006).
None of the three dragons is a pterosaur. Their wings, heads, tails, limbs, and skin are inconsistent with pterosaur anatomy.
COMPARISON OF BELON’S AND ALDROVANDI’S DRAGONS
WITH EXTANT ANIMALS
It is important to do more than just demonstrate that the three dragons are not pterosaurs. The countering of incorrect claims regarding specific specimens carries more weight if we can determine what the specimens are. We therefore compared the three specimens to extant animals to determine, as much as possible, what extant species comprised them.
Belon’s and Aldrovandi’s dragons are similar enough to support an inference that they were constructed from the same set of animal parts. They can therefore be considered together. They may even have come from the same region and have been made by the same hoaxer or group of hoaxers. Belon saw his dragons in Egypt, and Aldrovandi’s was said to come from “Aethiopia”.
 On their bellies are transversely elongated ventral scutes (Figure 1.1-4), a feature present in snake skin and absent in the skin of any other animal. The scale rows are diagonal, which is consistent with snake skin (Figure 3). The tails of the dragons also have transversely elongated ventral scutes and have diagonal scale rows (Figure 3.1-2). In contrast, the scale rows on the tails of most lizards other than skinks are vertical, not diagonal (Figure 3.7). We therefore conclude that the neck, torso, and tail of each of the two dragons is that of a snake, to which parts of other animals were attached.
On their bellies are transversely elongated ventral scutes (Figure 1.1-4), a feature present in snake skin and absent in the skin of any other animal. The scale rows are diagonal, which is consistent with snake skin (Figure 3). The tails of the dragons also have transversely elongated ventral scutes and have diagonal scale rows (Figure 3.1-2). In contrast, the scale rows on the tails of most lizards other than skinks are vertical, not diagonal (Figure 3.7). We therefore conclude that the neck, torso, and tail of each of the two dragons is that of a snake, to which parts of other animals were attached.
Aldrovandi described his winged dragon as having green and blackish scales and a yellow belly and tail. His painting shows that the scales were mainly green. Because Egypt does not have bright green snakes, the species of snake may have come from further south. Dendroaspis jamesoni (Jameson’s mamba) is a long (sometimes over 2 m) snake with black-edged green scales, a yellowish belly, and a yellow tail. Its color matches Aldrovandi’s description, but the closest its range currently comes to Egypt is southernmost Sudan (Spawls and Branch, 1995). We therefore are not confident that we can identify the species of snake used.
Both specimens have bulbous torsos, which is not the case in snakes. However, an object inserted down the throat into the digestive tract of a snake can distend the torso without tearing the skin, as in these specimens (Figure 3.8). The difference in shape between the two specimens’ torsos can therefore be explained by a difference in shape between inserted items.
 Several lines of evidence reveal that in each specimen the head of the snake has been replaced by the head of some other animal. First, the snout of a snake does not protrude as far anterior to the eye as is the case with these specimens’ snouts (Figure 4). Second, the tongue of Aldrovandi’s dragon, the only one of the two specimens in which the tongue is illustrated, is not forked (Figure 4.2). Third, in both specimens the dorsal skin of the snout is wrinkled (Figure 4.1-2), which does not happen in snakes. The wrinkling of facial skin is something that happens only in mammals, many of which have loose facial skin that permits changes of facial expression, whereas in non-mammalian vertebrates the skin is firmly and immovably attached to the skull. Both specimens also have pinnae (ear flaps), a feature unique to mammals. Also, both specimens have fleshy cheeks ventral to the eye, another trait unique to mammals. In both specimens the cheeks and the tops of the heads are covered in scales. It is therefore probable that in each case the neck skin of the snake was extended onto and attached to the mammalian heads and cheeks.
Several lines of evidence reveal that in each specimen the head of the snake has been replaced by the head of some other animal. First, the snout of a snake does not protrude as far anterior to the eye as is the case with these specimens’ snouts (Figure 4). Second, the tongue of Aldrovandi’s dragon, the only one of the two specimens in which the tongue is illustrated, is not forked (Figure 4.2). Third, in both specimens the dorsal skin of the snout is wrinkled (Figure 4.1-2), which does not happen in snakes. The wrinkling of facial skin is something that happens only in mammals, many of which have loose facial skin that permits changes of facial expression, whereas in non-mammalian vertebrates the skin is firmly and immovably attached to the skull. Both specimens also have pinnae (ear flaps), a feature unique to mammals. Also, both specimens have fleshy cheeks ventral to the eye, another trait unique to mammals. In both specimens the cheeks and the tops of the heads are covered in scales. It is therefore probable that in each case the neck skin of the snake was extended onto and attached to the mammalian heads and cheeks.
In both dragons the shape of the head is doglike, but the dentition lacks the distinct upper and lower canine teeth that are present in canids, as well as the molariform cheek teeth of canids. Instead, the teeth are all pointed and of similar length, with a slight increase in tooth length anteriorly in Aldrovandi’s painting (Figure 4.2). The only Egyptian mammal with a doglike head and a mouthful of pointed teeth with a gradual increase in tooth size anteriorly is the Egyptian fruit bat (Rousettus aegyptiacus) (Aulagnier et al., 2009) (Figure 4.3). Its post-canine teeth have sharp and procumbent points so that at a glance its dentition looks like a series of fangs that increase in size anteriorly; in this it resembles the dentition in the lower jaw of the painting of Aldrovandi’s dragon. Its ears are also smaller in relative size than those of other Middle Eastern bats, making it a good candidate for the contributor of the heads of the two dragons. All the above points also hold true for the straw-colored fruit bat (Eidolon helvum), which is found south of Egypt along the Red Sea. Also, the size of the head of an adult Egyptian fruit bat is a good fit for the neck of a large snake such as a mamba. Although the tooth counts of fruit bats are lower than that shown in the illustrations of Belon’s and Aldrovandi’s dragons, the overall resemblance is better than that of the head of any other northern African mammal.
 The skin of the two specimens’ wings is supported by internal struts, six in Belon’s dragon and five in Aldrovandi’s (Figure 5). Extant animals with patagia supported by internal struts include only bats and flying lizards (genus Draco). The dragons’ wings do not resemble those of bats. In most bats, the wing has only three internal struts. The first strut is composed of the third metacarpal and finger plus the closely-appressed and fingerless second metacarpal, the second strut is composed of the fourth metacarpal and finger, and the third strut is composed of the fifth metacarpal and finger (Figure 5.9). In some bats the second finger is retained and is separated from the third so that it forms another strut, but even in such cases there are only four struts. Furthermore, in bats the first finger protrudes from the leading edge of the wing, a feature absent in the two dragons.
The skin of the two specimens’ wings is supported by internal struts, six in Belon’s dragon and five in Aldrovandi’s (Figure 5). Extant animals with patagia supported by internal struts include only bats and flying lizards (genus Draco). The dragons’ wings do not resemble those of bats. In most bats, the wing has only three internal struts. The first strut is composed of the third metacarpal and finger plus the closely-appressed and fingerless second metacarpal, the second strut is composed of the fourth metacarpal and finger, and the third strut is composed of the fifth metacarpal and finger (Figure 5.9). In some bats the second finger is retained and is separated from the third so that it forms another strut, but even in such cases there are only four struts. Furthermore, in bats the first finger protrudes from the leading edge of the wing, a feature absent in the two dragons.
The dragons’ wings are also unlike the patagia of flying lizards. In the two dragons, the wing is narrow at the base and wide distally, whereas the patagium of a flying lizard is wider at the base than it is distally.
The Mediterranean fish known as the flying gurnard (Dactylopterus volitans) has pectoral fins with morphology like that of these specimens’ wings (Figure 5.3-5). There are multiple internal struts, the skin can spread as wide between the struts as in the wings of the two dragons, and the fins can be cut into the shapes shown in Belon’s and Aldrovandi’s illustrations. The pectoral fins of a live flying gurnard have brightly colored spots (Figure 5.7), which are absent in Belon’s and Aldrovandi’s dragons, but the spots fade after death, leaving the entire fin a dull brown (Figure 5.3-5).
Sea robins (Triglidae) have spreadable pectoral fins of similar shape (Figure 5.8), but they are too small to have been the source of these two specimens’ wings, and the fins cannot spread as widely between the struts as in Belon’s and Aldrovandi’s dragons (Figure 5.8). The pectoral fins of other bony fishes lack the wide membranes between internal struts that are found in the flying gurnard and sea robins. We therefore conclude that the “wings” of Belon’s and Aldrovandi’s dragons were made from the pectoral fins of flying gurnards.
The limbs of the two specimens are robust, and each terminates in four thick fingers with prominent, curved claws (Figure 1.1-4). Such forelimbs are not found in any circum-Mediterranean reptiles. Local lizards and crocodiles with robust forelimbs have five-fingered hands, and amphibians lack manual claws. Among mammals, robust forelimbs that terminate in four thick fingers are found in members of the Leporidae (the rabbit family), Canidae (the dog family), and Felidae (the cat family). The former two have fingers that are straighter than those of felids and therefore better resemble the fingers of the two dragons.
The scaly covering of the limbs of the two specimens indicates that they were covered in a sleeve of reptile skin. Rabbits, canids, and felids have a dewclaw (a short thumb that does not reach the tip of the forelimb), but its absence in the illustrations can be explained by its having been clipped off, with the reptile-skin sleeve covering the spot where it was removed. It is noteworthy that the illustrations do not show scales or fur covering the fingers. This suggests that the original mammal skin had been removed and that the reptile-skin sleeve did not extend onto the fingers.
We conclude that Belon’s and Aldrovandi’s winged dragons were taxidermic composites. Each was composed of a decapitated snake to which was added a fruit bat’s head, the pectoral fins of a flying gurnard, the forelimbs of a rabbit or canid, and an internal insert that distended the torso.
COMPARISON OF CARDINAL BARBERINI’S DRAGON
WITH EXTANT ANIMALS
Faber’s and Kircher’s illustrations of Cardinal Barberini’s dragon show a clear distinction between canine teeth and molariform cheek teeth, a mammalian trait. They also show a coronoid process on the mandible, another mammalian trait (Figure 1). The dragon’s single, posteriorly curving horn and hooklike, beaked proboscis are absent in all mammals but are known to have been added to a mammalian skull on at least one other fake dragon from the seventeenth century (Senter and Wilkins, 2013). It is therefore safe to dismiss these as fake additions to the skull of Cardinal Barberini’s dragon and not part of the original skull.
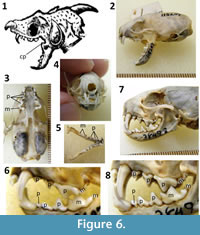 Details of the skull and dentition differ between Faber’s and Kircher’s drawings. The former is detailed and precise in its depiction of the skull and neck, whereas the skull and neck of the latter are more sloppily and less realistically rendered. For example, it includes teeth on the coronoid process (Figure 1.7), a feature present in no known extant or extinct tetrapod. We therefore consider the skull and neck in Faber’s drawing more reliable than those in Kircher’s. In Faber’s drawing, the prominence of the canine teeth, the general size and shape of the cheek teeth, the bulbous shape of the rear of the skull, and the fact that the mandible is only about half the total length of the skull, are consistent with only one mammalian genus: Mustela (weasels) (Aulagnier et al., 2009) (Figure 6). The lower incisors are hidden by the canines in lateral view, which is consistent with the Mustelidae (Aulagnier et al., 2009). The two smallest circum-Mediterranean species of Mustela are M. erminea (stoat) and M. nivalis (common weasel). No other western Eurasian species of Mustela is small enough to match the size of the specimen as described by Faber (Appendix). In both cases, the maximum gape of the jaw matches the gape of Cardinal Barberini’s specimen (Figure 6.2). Faber’s description of the incisors also fits Mustela : the upper incisor next to the canine tooth is larger than the other two upper incisors, and the lower incisors are smaller than the uppers (Appendix) (Figure 6.4).
Details of the skull and dentition differ between Faber’s and Kircher’s drawings. The former is detailed and precise in its depiction of the skull and neck, whereas the skull and neck of the latter are more sloppily and less realistically rendered. For example, it includes teeth on the coronoid process (Figure 1.7), a feature present in no known extant or extinct tetrapod. We therefore consider the skull and neck in Faber’s drawing more reliable than those in Kircher’s. In Faber’s drawing, the prominence of the canine teeth, the general size and shape of the cheek teeth, the bulbous shape of the rear of the skull, and the fact that the mandible is only about half the total length of the skull, are consistent with only one mammalian genus: Mustela (weasels) (Aulagnier et al., 2009) (Figure 6). The lower incisors are hidden by the canines in lateral view, which is consistent with the Mustelidae (Aulagnier et al., 2009). The two smallest circum-Mediterranean species of Mustela are M. erminea (stoat) and M. nivalis (common weasel). No other western Eurasian species of Mustela is small enough to match the size of the specimen as described by Faber (Appendix). In both cases, the maximum gape of the jaw matches the gape of Cardinal Barberini’s specimen (Figure 6.2). Faber’s description of the incisors also fits Mustela : the upper incisor next to the canine tooth is larger than the other two upper incisors, and the lower incisors are smaller than the uppers (Appendix) (Figure 6.4).
Faber’s tooth count--on each side, three upper and three lower incisors, one upper and one lower canine, and three upper and three lower cheek teeth (Appendix)–are consistent with Mustela except that the Mustela has four upper and five lower cheek teeth. Faber calls all the cheek teeth molars (molares), although today’s anatomists distinguish between premolars (adult cheek teeth which replace the deciduous cheek teeth of juveniles) and molars (adult cheek teeth that do not replace deciduous teeth). In Mustela the true upper molar and the second of the two true lower molars are tiny, and each is hidden from view by the much larger preceding tooth unless one detaches the mandible from the skull. It is therefore unlikely that Faber could have seen them. Also, the first lower premolar is pressed against the second in M. nivalis so that the two at first appear to be a single tooth; these two teeth are more clearly demarcated in M. erminea. The tooth count described by Faber is therefore consistent with the appearance of M. nivalis. Also, the depth of the maxilla beneath the eye socket is shallower in M. nivalis than in M. erminea (Figure 6.2, 6.7) and is more consistent with both Faber’s and Kircher’s illustrations. As the smaller of the two species, M. nivalis is also more consistent with Faber’s description of the skull’s size: its length is the width “of two fingers crosswise,” and its width is “the thickness of a thumb” and its lower incisors “are so small that it is necessary to view them with a magnifying lens, if one wishes to discern them” (Appendix). We therefore conclude that the skull of Cardinal Barberini’s dragon is that of M. nivalis.
The skin of Cardinal Barberini’s dragon has transversely elongated ventral scutes, as in snakes. However, Kircher’s illustration shows that the rest of its scales are in transverse rows that are vertical in lateral view (as in some lizards) rather than in diagonal rows (as in snakes). It is therefore likely that the specimen’s belly skin is that of a snake, while its dorsal and lateral skin is that of a lizard. A similar situation was present in another composite “dragon” hoax from Renaissance Europe, in which the torso had the belly skin of a snake but the dorsal and lateral skin of a fish (Senter et al., 2013).
The color of Cardinal Barberini’s dragon’s skin, according to Faber (Appendix), is a mixture of aquamarine, yellowish, and blackish, with more green dorsally and more yellow ventrally. Several lizard species of the appropriate color are present in the circum-Mediterranean area, several of which are in the genus Lacerta (Boulos, 1995; Schleich et al., 1996; Arnold, 2002; Baran et al., 2005; Baha el Din, 2006; Bar and Haimovitch, 2011). The skin covering the neck may be from the same individual as that covering the torso or it may be from a second individual. The range of sizes among appropriately colored, circum-Mediterranean species of Lacerta is consistent with the use of skin from a single individual or two.
 According to Faber, the wings of Cardinal Barberini’s dragon each have three internal struts, and are spotted with a background color of blue on one side and a dark wheatlike color on the other. Both Faber’s and Kircher’s illustrations show a covering of scales (Figure 7.1-2). Two small points that Faber calls claws project anteriorly from each wing. The scaly covering eliminates bony fish fins and bat wings from consideration, because neither is covered in scales. The patagia of flying lizards (Draco) are covered in scales, have ribs as internal struts, can be cut into the shape of the wings of the dragon, and in several species of Draco are spotted (Das, 2010) (Figure 7.3-7). To test whether the two anterior “claws” could be the projecting tips of sliced ribs of a Draco patagium, P.S. cut the right patagium off a dried specimen in his personal collection and found that even at the base of the patagium no two ribs converge closely enough to match the proximity of the claws on the dragon’s wing (Figure 7.4). To test whether the claws could be pieces of the pectoral girdle, P.S. cut the left patagium of the specimen close enough to include part of the body wall (Figure 7.5), and found that no part of the pectoral girdle extended into the cut-off piece. The pectoral girdle is slightly anterior to the anteriormost edge of the patagium where it meets the torso (Figure 7.5). No projecting pieces were present that could be identified as the “claws” of the dragon (Figure 7.6). We therefore could not confirm that the wings of Cardinal Barberini’s dragon were made from cut Draco patagia.
According to Faber, the wings of Cardinal Barberini’s dragon each have three internal struts, and are spotted with a background color of blue on one side and a dark wheatlike color on the other. Both Faber’s and Kircher’s illustrations show a covering of scales (Figure 7.1-2). Two small points that Faber calls claws project anteriorly from each wing. The scaly covering eliminates bony fish fins and bat wings from consideration, because neither is covered in scales. The patagia of flying lizards (Draco) are covered in scales, have ribs as internal struts, can be cut into the shape of the wings of the dragon, and in several species of Draco are spotted (Das, 2010) (Figure 7.3-7). To test whether the two anterior “claws” could be the projecting tips of sliced ribs of a Draco patagium, P.S. cut the right patagium off a dried specimen in his personal collection and found that even at the base of the patagium no two ribs converge closely enough to match the proximity of the claws on the dragon’s wing (Figure 7.4). To test whether the claws could be pieces of the pectoral girdle, P.S. cut the left patagium of the specimen close enough to include part of the body wall (Figure 7.5), and found that no part of the pectoral girdle extended into the cut-off piece. The pectoral girdle is slightly anterior to the anteriormost edge of the patagium where it meets the torso (Figure 7.5). No projecting pieces were present that could be identified as the “claws” of the dragon (Figure 7.6). We therefore could not confirm that the wings of Cardinal Barberini’s dragon were made from cut Draco patagia.
Trade with Australasia had been established by Faber’s day, so P.S. examined specimens of Chlamydosaurus kingii (frilled lizard) to determine whether the dragon’s wings could have been made of frilled lizard frills (Figure 7.8-11). Some specimens have spots on the frill (Figure 7.9), as in Cardinal Barberini’s dragon. The edge of the dorsal half of the frill has small, triangular projections that are similar to the dragon’s “claws” (Figure 7.8-10), so the possibility that all but two of those projections had been cut off a frill to form each of the dragon’s wings was deemed worthy of exploration. However, the frill has no internal struts. It does have three strong creases in its ventral half (Figure 7.8, 7.11), but when the frill is spread the lack of struts in the creases is apparent. Furthermore, the ventral half of the frill, which has the creases, lacks the triangular projections around the edge that otherwise could hypothetically have been the “claws”. We therefore conclude that the wings of Cardinal Barberini’s dragon were not made from frilled lizard frills. We remain baffled as to the identity of the wings.
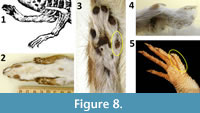 The limbs of Cardinal Barberini’s dragon terminate in five slender, well-defined fingers (Figure 8.1). According to Faber’s description, each finger has a sharp claw, and the finger in the position of the thumb is shorter than the others. The presence of five fingers with sharp claws eliminates many animals from consideration, but it is difficult to eliminate any of the remaining candidates. They include weasels (Figure 8.2-4) and numerous species of shrews and lizards. It is possible that the hoaxer used the forelimbs of the same weasel from which the skull came. The size of the hands of Mustela nivalis relative to its head (Figure 8.2) is similar to that of the dragon (Figure 1.6-7). If a weasel’s hands were used, they were skinned; there is no indication of fur on the hands in the illustrations of the dragon. It is also possible that the hoaxer used the forelimbs of the same lizard from which the skin came. Digital proportions in the hands of Lacerta (Figure 8.5) are similar to those of Cardinal Barberini’s dragon.
The limbs of Cardinal Barberini’s dragon terminate in five slender, well-defined fingers (Figure 8.1). According to Faber’s description, each finger has a sharp claw, and the finger in the position of the thumb is shorter than the others. The presence of five fingers with sharp claws eliminates many animals from consideration, but it is difficult to eliminate any of the remaining candidates. They include weasels (Figure 8.2-4) and numerous species of shrews and lizards. It is possible that the hoaxer used the forelimbs of the same weasel from which the skull came. The size of the hands of Mustela nivalis relative to its head (Figure 8.2) is similar to that of the dragon (Figure 1.6-7). If a weasel’s hands were used, they were skinned; there is no indication of fur on the hands in the illustrations of the dragon. It is also possible that the hoaxer used the forelimbs of the same lizard from which the skin came. Digital proportions in the hands of Lacerta (Figure 8.5) are similar to those of Cardinal Barberini’s dragon.
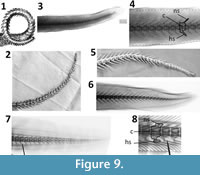 The bodies (centra) of the tail vertebrae of Cardinal Barberini’s dragon are of similar length and height. In contrast, those of small mammals, reptiles, and amphibians are much longer than tall. The vertebrae of fish tails tend to be of similar length and height, but the tail vertebrae of sharks and kin (Chondrichthyes) lack the dorsal and ventral spines (neural spines and hemal spines, respectively) seen on this specimen’s tail vertebrae. Such spines are the norm in bony fishes. We therefore conclude that the tail skeleton is that of a bony fish. In Faber’s illustration the neural and hemal spines point anteriorly, whereas in Kircher’s they point posteriorly. The coiling of the tail evidently confused the eye of one of the two illustrators. The more realistic of the two is the tail in Kircher’s illustration, because posteriorly pointing neural and hemal arches are the norm in vertebrates generally, including bony fishes (Figure 9).
The bodies (centra) of the tail vertebrae of Cardinal Barberini’s dragon are of similar length and height. In contrast, those of small mammals, reptiles, and amphibians are much longer than tall. The vertebrae of fish tails tend to be of similar length and height, but the tail vertebrae of sharks and kin (Chondrichthyes) lack the dorsal and ventral spines (neural spines and hemal spines, respectively) seen on this specimen’s tail vertebrae. Such spines are the norm in bony fishes. We therefore conclude that the tail skeleton is that of a bony fish. In Faber’s illustration the neural and hemal spines point anteriorly, whereas in Kircher’s they point posteriorly. The coiling of the tail evidently confused the eye of one of the two illustrators. The more realistic of the two is the tail in Kircher’s illustration, because posteriorly pointing neural and hemal arches are the norm in vertebrates generally, including bony fishes (Figure 9).
In most bony fishes there is an abrupt decrease in the size of the vertebrae at the base of the tail, unlike the gradual decline in vertebral size down the length of the tail in Cardinal Barberini’s dragon. Eels are exceptional among bony fishes in that their vertebrae exhibit a gradual decline in size down the length of the tail, as in Cardinal Barberini’s dragon. Of the three Mediterranean and European species of eel, Anguilla anguilla (European eel) is the best match to the tail of this dragon (Figure 9.4). The neural and hemal spines of the tail vertebrae of Muraena helena (Mediterranean moray) are shaped differently from those of the dragon (Figure 9.8). The neural and hemal spines of Conger conger (European conger) are relatively much longer than in the dragon (Figure 9.6). We therefore conclude that the tail skeleton of Cardinal Barberini’s dragon is that of Anguilla anguilla.
We conclude that Cardinal Barberini’s dragon was a taxidermic composite. It includes the skull of a common weasel; the belly skin of a snake; the dorsal and lateral skin of one or two individual lizards, possibly of the genus Lacerta ; the tail skeleton of an eel; and “wings” that remain a mystery.
DISCUSSION
To date, no “evidence” for pterosaur-human coexistence has survived scrutiny. Claims of sightings of live pterosaurs in the United States, Cuba, Africa, and Papua New Guinea (e.g., Gibbons and Hovind, 1999; Woetzel, 2006; Stuckwish, 2009; Whitcomb, 2010) are thus far unaccompanied by photographic evidence or physical specimens. An alleged prehistoric painting of a pterosaur in Black Dragon Canyon, Utah, is actually a composite picture of two humanoid figures, two quadrupedal animals, and a horned serpent (Senter, 2012). Two alleged rock paintings of pterosaurs near Alton, Illinois, are fictions created by a nineteenth-century American author, based on now-destroyed rock paintings of wingless creatures (Senter, 2012). Alleged pterosaurs on Greco-Roman coinage are winged snakes (Senter, 2013). Herodotus’ description of the anatomy of Egyptian winged serpents–alleged by some to be pterosaurs--is inconsistent with pterosaur anatomy (Senter, 2013). The “dragon” specimen that Cornelius Meyer exhibited in 1691 is a taxidermic hoax (Senter and Wilkins, 2013). The three specimens investigated here are also taxidermic hoaxes. These three specimens now join the ranks of investigated and falsified “evidence” for human-pterosaur coexistence and against evolutionary theory.
Cardinal Barberini’s dragon was a relatively famous specimen in its day, and it may have been the inspiration for the subsequent hoax perpetrated by Cornelius Meyer in 1691. The latter specimen was adorned with a backward-curving horn and a fake hook on the snout, rare attributes in fake dragons that are both present in Cardinal Barberini’s dragon. As in Cardinal Barberini’s dragon, its skull, ribcage, and tail skeleton were exposed, while its neck was covered in skin, and its tail was coiled. We therefore suggest that it was modeled after Cardinal Barberini’s dragon. Meyer’s dragon was said to have been killed in the marshes near Rome. However, it is but another taxidermic composite hoax, in this case one with the skull of a domestic dog and the forelimbs of a bear (Senter and Wilkins, 2013).
It is also noteworthy that Meyer’s hoax and the three hoaxes investigated here continue to fool people over 300 years later. In all three cases authors writing in the 1990s or the twenty-first century have insisted that the specimens were genuine, recently-killed pterosaurs (Goertzen, 1993, 1998; Woetzel, 2006, 2012) or dinosaurs (Cooper, 1992; Gilmer 2011). These authors are in good company. A plethora of other dragons created by taxidermic hoaxers in Renaissance Europe hoodwinked many prominent citizens and, in some cases, scientists--although not all scientists were fooled (Dance, 1975; Senter et al., 2013). That present-day authors continue to be fooled by hoaxes that were perpetrated centuries ago is a testament to the hoaxers’ skill.
It is noteworthy that some authors identify these specimens as pterosaurs despite their lack of resemblance to pterosaurs in all parts of their anatomy. The tendency to see nonexistent prehistoric animals in artifacts from centuries past is widespread among authors who contend that pterosaurs and dinosaurs coexisted with humans (Senter, 2013). As shown here, such inaccurate interpretations of old artifacts could be corrected by one’s simply taking the time to compare an alleged prehistoric animal with actual examples, e.g., comparing an alleged pterosaur with actual pterosaurs. It could also be alleviated by the study of comparative anatomy, so that parts of extant animals are more readily recognized in taxidermic composites. It is our hope that in future publications, others will heed the lesson of this study and employ these two solutions before publicly jumping to conclusions that are so easily shown to be false.
ACKNOWLEDGMENTS
We would like to thank the following people for their contributions to this project. R. Arrindell provided access to fish specimens at the American Museum of Natural History. M. Arnold and D. Kizirian provided access to reptile specimens at the American Museum of Natural History. C. Potter provided access to mammal specimens at the United States National Museum. D. Lunde, M. Krol, and N. Edmison provided a loan of bat skulls from the United States National Museum. L. Gatens and B. Hess provided access to mammal specimens at the North Carolina Museum of Natural Sciences. G. Hogue provided a loan of eel specimens from the North Carolina Museum of Natural Sciences. J. Woodward provided images of lizard specimens from the Museum of Comparative Zoology. K. Hartel provided x-ray images of eel specimens from the Museum of Comparative Zoology. C. Bennett and an anonymous reviewer provided helpful reviews of this article.
REFERENCES
Aldrovandi, U. 1640. Serpentum et Draconum Historiae. M. Antony Berm, Bologna.
Anthon, C. 1878. A New Classical Dictionary of Greek and Roman Biography, Mythology, and Geography. Harper and Brothers, New York.
Arnold, E.N. 2002. Reptiles and Amphibians of Europe. Princeton University Press, Princeton.
Aulagnier, S., Haffner, P., Mitchell-Jones, A.J., Moutou, F., and Zima, J. 2009. Mammals of Europe, North Africa and the Middle East. A&C Black, London.
Baha el Din, S. 2006. A Guide to the Reptiles and Amphibians of Egypt. The American University in Cairo Press, Cairo.
Bar, A. and Haimovitch, G. 2011. A Field Guide to Reptiles and Amphibians of Israel. Pazbar, Herzliya, Israel.
Baran, I., Ilgaz, Ç., Avci, A., Kumlutaş, Y., and Olgun, K. 2005. Türkiye Amfibi ve Sürüngenleri. Tubitak, Ankara.
Bartholin, T. 1678. De Unicornu. Observationes Novae. Henry Wetstenium, Amsterdam.
Belon, P. 1557. Portraits d’Oiseaux, Animaux, Serpents, Herbes, Arbres, Hommes et Femmes, d’Arabie & Egypte, Observés par P. Belon du Mans. Guillaume Cavellat, Paris.
Belon, P. 1588. Les Observations de Plusieurs Singularités et Choses Mémorables, Trouvées en Grèce, Asie, Judée, Egypte, Arabie & Autres Pays Etrangés. Guillaume Cavellat, Paris.
Biblioteca Universitaria di Bologna. 2013. Le Tavole Acquerellate di Ulisse Aldrovandi. www.filosofia.unibo.it/Aldrovandi/pinakesweb/main.asp (last accessed December 16, 2013).
Bloch, M.E. 1790. Allgemeine Naturgeschichte des Fische. J. Morino, Berlin.
Boulos, I. 1995. Les Amphibiens, les Reptiles et les Oiseaux du Liban. Wizarat al-Zira ‘ah, Beirut.
Cooper, B. 1992. The early history of man--part 4. Living dinosaurs from Anglo-Saxon and other early records. Creation Ex Nihilo Technical Journal, 6:49-66.
Dalla Vecchia, F.M. 2003. An Eudimorphodon (Diapsida, Pterosauria) specimen from the Norian (Late Triassic) of north-eastern Italy. Gortania, 25:47-72.
Dance, P. 1975. Animal Fakes and Frauds. Sampson Low, Maidenhead, United Kingdom.
Das, I. 2010. A Field Guide to the Reptiles of South-east Asia. New Holland, London.
Faber, G. 1651. Dracunculus Monoceros Illustris. Card. Barberini, pp. 816-822. In Lyncean Academy (ed.), Rerum Medicarum Novae Hispaniae Thesaurus. Uncio Vascardi, Rome.
Freedberg, D. 2002. The Eye of the Lynx. University of Chicago Press, Chicago.
Gessner, C. 1589. Schlangenbuch. Froschauer, Zurich.
Gibbons, W.J. and Hovind, K. 1999. Claws, Jaws & Dinosaurs. CSE, Pensacola.
Gilmer, J.E. 2011. 100 Year Cover-up Revealed. We Lived with Dinosaurs! AuthorHouse, Bloomington.
Goertzen, J. 1993. Living dinosaurs. Creation Ex Nihilo Technical Journal, 7:200-201.
Goertzen, J. 1998. The rhamphorhynchoid pterosaur Scaphognathus crassirostris : a “living fossil” until the 17th century?, pp. 253-269. In Walsh, R.E. (ed.), Proceedings of the Fourth International Conference on Creationism. Creation Science Fellowship, Pittsburgh.
Hare, J.B. 2010. Internet Sacred Text Archive. www.sacred-texts.com (last accessed May 20, 2013).
Kircher, A. 1664. Mundus Subterraneus. Johan Johansen and Elias Weyerstraet, Amsterdam.
Lyncean Academy. 1651. Rerum Medicarum Novae Hispaniae Thesaurus. Uncio Vascardi, Rome.
Olmi, G. 2007. Ulisse Aldrovandi, pp. 59-62. In Huxley, R. (ed.), The Great Naturalists. Thames and Hudson, London.
Schleich, H.H., Kästle, W., and Kabisch, K. 1996. Amphibians and Reptiles of North Africa. Koeltz Scientific, Koenigstein.
Senter, P. 2012. More “dinosaur” and “pterosaur” rock art that isn’t. Palaeontologia Electronica, 15(2.22A):1-14.
Senter, P. 2013. Dinosaurs and pterosaurs in Greek and Roman art and literature? An investigation of young-Earth creationist claims. Palaeontologia Electronica, 16(3.25A):1-16.
Senter, P. and Wilkins, P.D. 2013. Investigation of a claim of a late-surviving pterosaur and exposure of a taxidermic hoax: the case of Cornelius Meyer’s dragon. Palaeontologia Electronica, 16(1.6A):1-11.
Senter, P., Hill, L.C., and Moton, B.J. 2013. Solution to a 440-year-old zoological mystery: the case of Aldrovandi’s dragon. Annals of Science, 70:531-537.
Spawls, S. and Branch, B. 1995. Dangerous Snakes of Africa. Ralph Curtis, San Sanibel.
Stuckwish, D. 2009. Biblical Cryptozoology: Revealed Cryptids of the Bible. Xlibris, Bloomington.
Topsell, E. 1608. The History of Serpents. William Taggard, London.
Unwin, D.M. 2006. The Pterosaurs. From Deep Time. Pi Press, New York.
Wellnhofer, P. 1975. Die Rhamphorhynchoidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. Palaeontographica Abteilung A, 148:1-33.
Wellnhofer, P. 1991. The Illustrated Encyclopedia of Pterosaurs. Crescent, New York.
Whitcomb, J.D. 2010. Live Pterosaurs in America. Self-published.
Woetzel, D. 2006. The fiery flying serpent. Creation Research Society Quarterly, 42:241-251.
Woetzel, D. 2012. Chronicles of Dinosauria. Master Books, Green Forest, Arkansas.
