

 The anatomy and diversity of the pterosaurian sternum
The anatomy and diversity of the pterosaurian sternum
Article number: 26.1.a12
https://doi.org/10.26879/1261
Copyright Paleontological Society, April 2023
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 19 December 2022. Acceptance: 28 March 2023.
ABSTRACT
Despite a key role in anchoring major flight muscle groups, the sternum of pterosaurs has attracted relatively little attention in the scientific literature. Here the sterna of more than 60 pterosaur genera are described and compared. Despite the overall conservative nature of the pterosaurian skeleton, the sternum varies greatly within and between clades, and ontogenetically. The sternum appears to have been mostly cartilaginous in juvenile (flying) pterosaurs, which ossified in different patterns in various taxa and even individuals leading at least in part to the variation seen. Comparisons of the pterosaur sternum to other reptiles suggest that it is formed of the clavicles, interclavicle and sternal plated fused into a single unit, though the details of this remain uncertain. Further studies of this major anatomical feature are required given its importance in anchoring flight muscles and therefore locomotion.
David W. E. Hone. School of Biological and Behavioural Sciences, Queen Mary University of London, Mile End Road, London, E1 4NS, UK. d.hone@qmul.ac.uk
Keywords: Pterosauria; anatomy; flight; Pterodactyloidea
Final citation: Hone, David W.E. 2023. The anatomy and diversity of the pterosaurian sternum. Palaeontologia Electronica, 26(1):a12.
https://doi.org/10.26879/1261
palaeo-electronica.org/content/2023/3813-the-sternum-of-pterosaurs
Copyright: April 2023 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
The pterosaurian sternum functions as an anchor and articulations point for the coracoids and as an anchor for major flight muscles (Bennett, 2003). As the forelimbs of pterosaurs are used in flight as well as for launching and terrestrial locomotion, the sternum would have had a critical role in the biomechanics of pterosaurs. The sternum may also have a role in breathing, being moved by the sternal ribs (Claessens et al., 2009; Geist et al., 2014). The sternum should therefore be under strong selection and provide important information on both the biology and phylogeny of pterosaurs, although it has so far had very limited attention in the scientific literature.
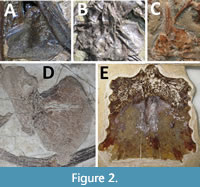
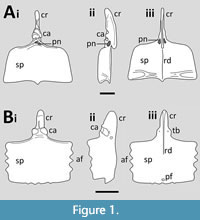 The sternum in pterosaurs consists primarily of a large, thin and convex plate of bone that sits on the anterior part of the chest (Figure 1). This may be anything from square to triangular or sub-circular in shape, and longer than broad or the reverse. In all well-preserved specimens there is an anteriorly directed process called the cristopsine, which varies greatly in shape between taxa, but is often long, thin and dorsoventrally expanded. This serves as an articulation point for the scapulocoracoids, which meet the cristospine on the posteriolateral surface (Bennett, 2001). Unusually in tetrapods, these are often asymmetric with the right articulation anterior to the left. The sternum may have smooth lateral margins or may have a serrated appearance with articulation points along the lateral edges to connect to the sternal ribs (Geist et al., 2014), or these may be fused to the sternum itself. In a few, there is a distally projecting xiphoid process from the middle of the ventral margin of the sternal plate (Jiang et al., 2016). The plate often has a thickened anterior rim and there might be a slight keel on the anterior part of the ventral face. The plate may be pierced by several holes and the cristospine is usually pneumatic with one or more pneumatopores. In short, there is considerable variation in the size, shape and anatomical details of this singular element (and its preservation), despite the often highly conservative nature of pterosaur bauplans (Figure 2).
The sternum in pterosaurs consists primarily of a large, thin and convex plate of bone that sits on the anterior part of the chest (Figure 1). This may be anything from square to triangular or sub-circular in shape, and longer than broad or the reverse. In all well-preserved specimens there is an anteriorly directed process called the cristopsine, which varies greatly in shape between taxa, but is often long, thin and dorsoventrally expanded. This serves as an articulation point for the scapulocoracoids, which meet the cristospine on the posteriolateral surface (Bennett, 2001). Unusually in tetrapods, these are often asymmetric with the right articulation anterior to the left. The sternum may have smooth lateral margins or may have a serrated appearance with articulation points along the lateral edges to connect to the sternal ribs (Geist et al., 2014), or these may be fused to the sternum itself. In a few, there is a distally projecting xiphoid process from the middle of the ventral margin of the sternal plate (Jiang et al., 2016). The plate often has a thickened anterior rim and there might be a slight keel on the anterior part of the ventral face. The plate may be pierced by several holes and the cristospine is usually pneumatic with one or more pneumatopores. In short, there is considerable variation in the size, shape and anatomical details of this singular element (and its preservation), despite the often highly conservative nature of pterosaur bauplans (Figure 2).
 To date, however, the pterosaurian sternum has featured only occasionally in the scientific literature and detailed descriptions are generally lacking. For example, very few characters in major phylogenetic analyses have been based on the sternum (none in Unwin, 2003, one in Kellner, 2003, five in Andres et al., 2014, and five in Vidovic and Martill, 2018), and it has also had little coverage in analyses of pterosaur mechanics (e.g., Bennett, 2003; Chatterjee and Templin, 2004). Only two papers in the last decade make any attempt to compare the sterna of different pterosaur clades. Lü et al., (2011a) provided some very brief comments comparing the sternum of Darwinopterus to other pterosaurs, while Jiang et al. (2016) complete an outline analysis of the sternum shape of 17 monofenestratan taxa. This included members of the wukongopterids, archaeopterodactyloids, pteranodontids and azhdarchoids (plus the genus Haopterus which remains of uncertain placement). Based on this, they suggested that in future phylogenetic analyses, the shape of the sternum should be divided into three characters for which their principal components analysis showed would cover most of the variation seen. These would be “the ratio of the length to width of sternal plate; the position of the lateral margins; and the shape of the posterior margin” (Jiang et al., 2016), though there were no further details about how these characters and their states might be constructed or coded. In short, the sternum remains a greatly overlooked part of pterosaurian anatomy and requires attention.
To date, however, the pterosaurian sternum has featured only occasionally in the scientific literature and detailed descriptions are generally lacking. For example, very few characters in major phylogenetic analyses have been based on the sternum (none in Unwin, 2003, one in Kellner, 2003, five in Andres et al., 2014, and five in Vidovic and Martill, 2018), and it has also had little coverage in analyses of pterosaur mechanics (e.g., Bennett, 2003; Chatterjee and Templin, 2004). Only two papers in the last decade make any attempt to compare the sterna of different pterosaur clades. Lü et al., (2011a) provided some very brief comments comparing the sternum of Darwinopterus to other pterosaurs, while Jiang et al. (2016) complete an outline analysis of the sternum shape of 17 monofenestratan taxa. This included members of the wukongopterids, archaeopterodactyloids, pteranodontids and azhdarchoids (plus the genus Haopterus which remains of uncertain placement). Based on this, they suggested that in future phylogenetic analyses, the shape of the sternum should be divided into three characters for which their principal components analysis showed would cover most of the variation seen. These would be “the ratio of the length to width of sternal plate; the position of the lateral margins; and the shape of the posterior margin” (Jiang et al., 2016), though there were no further details about how these characters and their states might be constructed or coded. In short, the sternum remains a greatly overlooked part of pterosaurian anatomy and requires attention.
The sternum is itself often not preserved in pterosaurs, or is poorly preserved even when the rest of the specimen is present. For example, little more than the cristospine is preserved of the otherwise superbly preserved Anhanguera described by Kellner and Tomida (2000), it is badly preserved in Nurhachius despite being surrounded by better preserved elements (Wang et al., 2005), and the ‘Vienna’ specimen of Aerodactylus (NHMW 1975/1756) has a poorly preserved sternum despite otherwise being complete and articulated and preserving such rare and fragile features as the propatagium and brachiopatagium. A few others are preserved with parts missing, a rough surface texture unlike other elements of a specimen, and even with numerous holes penetrating the main sternal plate (Jiang et al., 2016, figure 1). This collectively suggests that the sternum was often poorly ossified and / or very thin making it unlikely to preserve or to be preserved only poorly, and may have had an extensive cartilaginous component in many taxa. It is as little as 1.5 mm thick in even large taxa such as Quetzalcoathus lawsoni (Padian et al., 2021) and even then is often pneumatic (Elgin and Hone, 2013) and therefore both thin and fragile. Similarly, Elgin and Frey (2011) noted that the pectoral muscles inserting on the sternum would be fleshy and lack ligaments and tendons and with weak sternocostal and other articulations the sternum would likely detach early on in decay. Collectively this would explain the rarity of pterosaur sterna, even in otherwise well-preserved and largely complete specimens.
However, while the sternum of pterosaurs has received little attention and is considered rare, with few available for study (Jiang et al., 2016), an extensive review of the literature suggests that relatively complete and well-preserved sterna are known for over 60 genera taxa, with more preserving at least partial sterna. Almost all major pterosaur family-equivalent clades are represented in this collection including small Triassic animals through to Late Cretaceous giants, and for at least a few species, multiple specimens including juveniles and adults. Thus, most of the known gross diversity of pterosaurs in form and ecology are represented and the sternum, despite the preservational problems (Figure 2), is clearly better known than commonly realised. As such, this is a field worthy of further investigation. Here, every known pterosaur sternum is described, and its structure, ontogeny, function and homology are discussed.
Institutional Abbreviations
BSP, Bavarian State Collection for Palaeontology and Geology, Munich, Germany; CM Carnegie Museum, Pittsburgh, USA; CYGB, Chaoyang Geopark, Chaoyang City, China; IMNH, Iwaki Museum, Fukushima, Japan; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China; JME, Jura Museum Eichstaett, Bavaria, Germany; LF, Lauer Foundation for Paleontology, Science and Education, Wheaton, Illinois, USA; MN, Museu Nacional, Rio de Janerio, Brazil; NHM Natural History Museum, London, UK; NHMW, Natural History Museum, Vienna, Austria; PIN, Palaeontological Institute, Russian Academy of Sciences, Moscow, Russia.
MATERIALS AND METHODS
An extensive survey of the literature was carried out in addition to examining specimens in museum collections and assessment of my own archives of photographs of pterosaur fossils. Every taxon found with any degree of preservation of the sternum was recorded and below each of these is briefly described. Many of the sterna are poorly preserved, not (or only poorly) illustrated, or are not described in anything but very the briefest terms. As a result, many of these following descriptions are necessarily brief since only the limited information of the original scientific literature was available.
Descriptions
Most pterosaur sterna are either preserved in ventral view or have only been figured in ventral view. As a result, the following descriptions are all based on this view unless otherwise stated. The systematic relationships of pterosaurs are controversial with multiple competing, and contradictory, phylogenies in the literature. Here the genera are groups broadly according to the relationships of Zhou et al. (2021) for non-pterodactyloids and Hone et al., (2020) for pterodactyloids, but it should be noted that some taxa do not clearly fall into recognised clades, or are controversial in their placement and some clade names vary in usage by different authors.
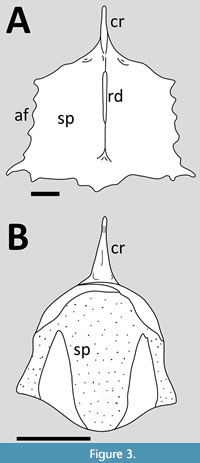 Eopterosauria. The sternum of a subadult Eudimorphodon is near complete and well preserved in ventral view (Wild, 1978, plate 2 and figure 14) (Figure 3A). The sternal plate is approximately square in outline through with a slight triangular part proximally, and it has a slightly wider posterior margin than the proximal one. Both the lateral and posterior margins are sinusoidal with multiple articulation points for costal ribs. The lateral margins show five points each plus the distal corner forms an extended point. The posterior margin is also complex showing five distinct points, with three of them part of a slight posterior expansion of the plate close to the midline. Wild (1978) states that there are eight articulation points on each side, which would match the description here if the elongate corners and the posterior midline point are not included. The cristospine is illustrated as be relatively short and narrow in Wild’s (1978) figure 14, though in his plate 2 he shows it to be considerably longer – this is considered the correct interpretation in comparison to his plate 1. The cristospine is barely connected to the thin keel that runs down most of the face of the sternal plate before bifurcating close to the posterior margin. There are a pair of depressions that sit at the base of the cristospine where it connects to the anteriormost part of the sternal plate, which are the coracoid articulations.
Eopterosauria. The sternum of a subadult Eudimorphodon is near complete and well preserved in ventral view (Wild, 1978, plate 2 and figure 14) (Figure 3A). The sternal plate is approximately square in outline through with a slight triangular part proximally, and it has a slightly wider posterior margin than the proximal one. Both the lateral and posterior margins are sinusoidal with multiple articulation points for costal ribs. The lateral margins show five points each plus the distal corner forms an extended point. The posterior margin is also complex showing five distinct points, with three of them part of a slight posterior expansion of the plate close to the midline. Wild (1978) states that there are eight articulation points on each side, which would match the description here if the elongate corners and the posterior midline point are not included. The cristospine is illustrated as be relatively short and narrow in Wild’s (1978) figure 14, though in his plate 2 he shows it to be considerably longer – this is considered the correct interpretation in comparison to his plate 1. The cristospine is barely connected to the thin keel that runs down most of the face of the sternal plate before bifurcating close to the posterior margin. There are a pair of depressions that sit at the base of the cristospine where it connects to the anteriormost part of the sternal plate, which are the coracoid articulations.
A juvenile specimen referred to Eudimorphodon by Wild (1993) also preserves a sternum in dorsal view (Figure 3B). This is somewhat oval in outline but is broadly similar to that of the subadult described above but with rounded margins and no articulations (Wild, 1993, figure 6; Wellnhofer, 2003, figure 11b). It has a narrowing anterior part, distolaterally expanded corners and with a slight distal expansion in the midline. Wild (1993, figure 6) illustrates this as having two different bone textures with two triangular insertions being different to the rest of the plate. It has at thickened anterior margin, which Wild (1993) says is composed of two separate and overlapping elements. The cristospine is relatively short and sharply pointed with a narrow ridge down the middle.
Renesto (1993) also described an isolated sternum that he referred to Eudimorphodon. Given the recent revisions to a number of specimens formerly identified as being in this clade, (Kellner, 2015; Dalla Vecchia, 2019) this referral must be tentative, but certainly the piece is very similar in form to that described by Wild (1978). The sternum is preserved in ventral view and is incomplete with part of the sternal plate missing (Renesto, 1993; Dalla Vecchia, 2014). As preserved it conforms almost exactly to that described above, though Renesto (1993) notes that there is a deep notch on either side of the cristospine where it meets the sternal plate, which are the articulation for the coracoids, and these are more obvious in this specimen than the specimen described by Wild (1978).
The adult holotype specimen of Seazzadactylus preserves a sternum, though it is covered in places by other elements limiting the available information (Dalla Vecchia, 2019, figure 17). The sternal plate is proximally triangular with slightly concave dorsolateral margins, and then the lateral margins would be parallel to make a more rectangular part. As preserved is it slightly longer than wide though the posterior margin is not visible. The visible lateral margin of one side has a serrated margin with clear points for articulation sternal ribs. Only the base of the cistospine is preserved (Dalla Vecchia, 2019).
Carniadactylus has a sternum preserved in ventral view, but it is mostly hidden behind other elements (Dalla Vecchia, 2009). Is was described by Dalla Vecchia (2009) as being broad and thin, with the anterior part being triangular in outline with a long cristospine that is dorsoventrally short.
A sternum of Austriadraco is both described and illustrated by Wellnhofer, 2003, figures 9 and 11). However, Kellner (2015) correctly identifies this in the text as being a fused fair of frontals (although in the caption of his figure 2e, Kellner identifies this as a sternum), and this identification as cranial elements is followed here.
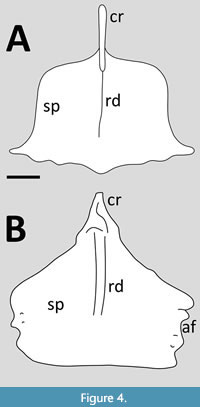 Campylognathoidea. Several specimens of Campylognathoides liasicus are known with preserved sterna, though they mostly show little anatomical information (Figure 4). The cristopine is tall (i.e., it is ventrally expanded), and there is a ridge extending from this into the midline of the sternal plate. According to Padian (2008b), Wiman (1925) described Campylognathoides as having asymmetric coracoid articulations. Padian (2008b) considered the “large, broad quadrangular sternum flared at posterior ends” to be a diagnostic trait of the genus (though noted the flared ends are shared with Eudimorphodon). The best preserved specimen (CM 11424) shows the sternal plate to be roughly square with, as noted, laterally projecting extensions on the distal face, and the posterior margin is scalloped. It is overall remarkable similar to that of Eudimorphodon, only lacking the lateral serrations of the Triassic taxon.
Campylognathoidea. Several specimens of Campylognathoides liasicus are known with preserved sterna, though they mostly show little anatomical information (Figure 4). The cristopine is tall (i.e., it is ventrally expanded), and there is a ridge extending from this into the midline of the sternal plate. According to Padian (2008b), Wiman (1925) described Campylognathoides as having asymmetric coracoid articulations. Padian (2008b) considered the “large, broad quadrangular sternum flared at posterior ends” to be a diagnostic trait of the genus (though noted the flared ends are shared with Eudimorphodon). The best preserved specimen (CM 11424) shows the sternal plate to be roughly square with, as noted, laterally projecting extensions on the distal face, and the posterior margin is scalloped. It is overall remarkable similar to that of Eudimorphodon, only lacking the lateral serrations of the Triassic taxon.
One specimen of C. zitteli is known with a sternum, but it is poorly preserved (Padian, 2008b), which shows a short cristopine, midline ridge and midline extension posteriorly.
Kellner (2015) states that the holotype of Bergamodactlys has “a developed and ossified sternum” but it is not described any further. Wild’s (1978, plate 5) drawing of the specimen shows this to be a small patch of bone close to the dorsal series and no details are apparent.
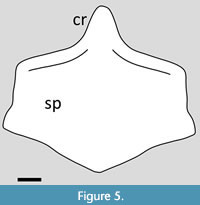 Anurognathidae. There is only one well-preserved sternum known for any anurognathid and that is in Batrachognathus (Ryabinin, 1948). This is pentagonal in shape (Figure 5) with the sternal plate being slightly wider than long. The cristospine is short but broad and semi-circular (Ryabinin, 1948). The lateral and posterior margins are roughly straight and smooth though they are somewhat uneven. Similarly, there is an extra expanse of bone of the posterior left margin such that the two sides are uneven, suggesting incomplete ossification of the sternum and that this would have been larger with a cartilaginous part. The posterior half of the sternal plate has a striated texture. There appears to be a thick and smooth ridge close to the anterior margin and offset from the midline of the sternal plate, but these are most likely the impressions of other bones that lie below the element since they are present on the right side.
Anurognathidae. There is only one well-preserved sternum known for any anurognathid and that is in Batrachognathus (Ryabinin, 1948). This is pentagonal in shape (Figure 5) with the sternal plate being slightly wider than long. The cristospine is short but broad and semi-circular (Ryabinin, 1948). The lateral and posterior margins are roughly straight and smooth though they are somewhat uneven. Similarly, there is an extra expanse of bone of the posterior left margin such that the two sides are uneven, suggesting incomplete ossification of the sternum and that this would have been larger with a cartilaginous part. The posterior half of the sternal plate has a striated texture. There appears to be a thick and smooth ridge close to the anterior margin and offset from the midline of the sternal plate, but these are most likely the impressions of other bones that lie below the element since they are present on the right side.
A second, very poorly preserved sternum is present on the juvenile specimen of Anurognathus. In his detailed description of the specimen, Bennett (2007) described this although the sternum is preserved below the dorsal vertebra as the specimen is in dorsal view. The cristospine is not visible and only parts of the sternal plate can be seen. Bennett suggested that based on the sternal ribs, the plate was as long as four or five of the dorsal vertebrae, and was anteroposteriorly narrow.
Scaphognathidae. Although represented by few taxa and specimens, this clade has a number of well-preserved sterna known (Figure 6). Wellnhofer (1978, figure 8) illustrates the sternum of Scaphognathus in dorsal view. The cristospine is very long (nearly the length of the sternal plate) and tapers anteriorly. Wellnofer (1978) indicates that a depression at the base of the cristospine is a single facet for the articulation of the coracoids, though Bennett (2001) considered these to be asymmetric. The sternal plate is sub-triangular or fan-shaped in outline with some kind of midline rib or depression. There are a pair of holes, one just either side of this midline features around one third of the way down the sternal plate. However, recent photographs under UV light and other regimes (Jäger et al., 2018) show that the sternum was rather longer and more narrow than Wellnhofer illustrated, and that there is no clear evidence of the apparent holes, and that the cristospine appears to be exaggerated in his illustration.
A sub-triangular sternum in ventral view is present on a juvenile specimen of Scaphognathus described by Bennett (2014). This is covered by the ribs and so some details are obscured. It is described as having a thickened anterolateral corner for articulation with a dorsal rib, though there are no clear indications of other articulations on the lateral margins of the sternal plate, and the posterior margin of the plate is thinner (Bennett, 2014). There is no evidence of holes penetrating the sternal plate. A second sternum of a young juvenile Scaphognathus is also known (LF 722) and is broadly like the one described by Bennett (2014). This second specimen is in probable dorsal view, but is broadly asymmetrical in shape with part of the right side missing, but it was likely a fan-shape originally with no apparent cristospine. The posterior margin is thickened, and there are no penetrating holes in the sternal plate.
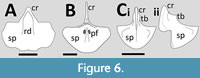 A sternum is preserved in the holotype of Fenghuangopterus (Lü et al., 2010a) (Figure 6A). The cristospine is long and broad, and a rounded ridge runs from the cristospine down on the middle of the sternal plate. Several others are also present at an angle to this one, however, while the middle of these appears to be genuine, the others are most likely the result of underlying ribs. This was described as ‘fan-shaped’ by Lü et al. (2010a), and it is broader than long and with a general convex outline. The sternum is described by Lü et al. (2010a) as having a pair of fenestrae in it, in the same manner as Scaphognathus, but these cannot be seen.
A sternum is preserved in the holotype of Fenghuangopterus (Lü et al., 2010a) (Figure 6A). The cristospine is long and broad, and a rounded ridge runs from the cristospine down on the middle of the sternal plate. Several others are also present at an angle to this one, however, while the middle of these appears to be genuine, the others are most likely the result of underlying ribs. This was described as ‘fan-shaped’ by Lü et al. (2010a), and it is broader than long and with a general convex outline. The sternum is described by Lü et al. (2010a) as having a pair of fenestrae in it, in the same manner as Scaphognathus, but these cannot be seen.
The sternum of Nesodactylus is described and illustrated by Colbert et al. (1969) (Figure 6C) although both Wellnhofer (1978) and Lü et al. (2011) redrew this with a more complete cristospine than is preserved in the original specimen. The sternum is shown to be nearly semi-circular in outline with a straight anterior margin and a smooth margin and is relatively small compared to the cristospine. Colbert et al. (1969) notes that the sternal plate is not flattened but “its two sides extend obliquely upward from the midline”. The cristospine is dorsoventrally tall and simple in morphology with a pair of bulges at the base of this. The lack of any complex anatomy on the ventral margins of the cristospine suggest that this had symmetrical articulations for the coracoids.
The sternal plate of Daohugouopterus is small and triangular in shape, and it is approximately twice as wide as long (Cheng et al., 2015) with smooth lateral margins. The anterior margin as illustrated by Cheng et al. (2015, figure 1) appears to be damaged and it is not well preserved. There is no apparent cistospine preserved and none is mentioned in the description.
Although not described in the literature, photos of specimen PIN 2585/25 of a juvenile of Sordes, show it has a small and triangular plate of bone that is likely an incomplete sternal plate. This is difficult to make out and its very small size suggests it is incompletely ossified.
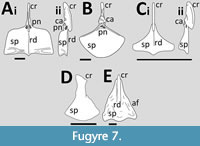 Rhamphorhynchidae. Numerous specimens of Rhamphorhynchus preserve a sternum including both juvenile and adult specimens (Figure 7A-C). Following Bennett (1995), the Solnhofen specimens of Rhamphorhynchus are considered to be a monotypic genus. Colbert et al. (1969) noted that some sterna of specimens of Rhamphorhynchus are proportionally much larger than those of Nesodactlyus but others are similar in size, suggesting ontogenetic changes in size.
Rhamphorhynchidae. Numerous specimens of Rhamphorhynchus preserve a sternum including both juvenile and adult specimens (Figure 7A-C). Following Bennett (1995), the Solnhofen specimens of Rhamphorhynchus are considered to be a monotypic genus. Colbert et al. (1969) noted that some sterna of specimens of Rhamphorhynchus are proportionally much larger than those of Nesodactlyus but others are similar in size, suggesting ontogenetic changes in size.
Small (juvenile) specimens of Rhamphorhynchus (Wellnhofer, 1978, figure 8, ‘R. longicaudus ’) exhibit a sternal plate that is in an inverted kite shape in outline, with slightly concave anterior margins and a slight midline keel (Figure 7C). The margins of the sternal plate are smooth. It has a long cristospine with a ventrally thickened margin. There is a small and circular hole that penetrates the lateral face of the cristospine close to where it meets the sternal plate.
The sternal plate of a specimen of intermediate size (Figure 7B) is illustrated by Wellnhofer (1975, figure 8e), which shows the plate to be roughly semi-circular in outline and shows a somewhat intermediate size and shape between the small morph above and the trapezoid one described below for larger specimens. Another intermediate-sized specimen (Wellnhofer, 1975, figure 8b) in dorsal view shows the presence of a pair of foramina on the dorsal side of the sternal plate, immediately posterior to the cristospine. One of these is positioned in the midline and the other is displaced to the side. Without an illustration of the plate in ventral view it is not clear if these penetrate the plate fully or are pneumatic openings. In dorsal view, the plate is shown to have a thickened anterior margin. The cristospine is similar in size and shape to that of the juvenile specimen and shows asymmetrical articulations for the coracoids.
In large specimens (Wellnhofer, 1978, figure 8, R. muensteri) the sternal plate is closer to a square outline with much more bone in the anterior part of the plate (Figure 7A). There is still a slight midline ridge and also some thickening and raised parts of the plate close to the posterior margins. There are a pair of holes (one each side) of the sternal plate, close to the anterior margin and near the base of the cristospine, though these holes are apparently inconsistently present as they are illustrated in Wellnhofer (1975, figure 8a) but not in Wellnhofer (1978, figure 8). The cristospine is similarly proportioned to that of the juveniles and intermediate-sized animals, though with a proportionally larger hole on the lateral face. This is shown to be circular in figure 8 of Wellnhofer (1978), but elongate and oval in figure 8c of Wellnhofer (1975). Wellnhofer (1978) also includes a dorsal view of the base of the cristospine that clearly shows the asymmetric articulations for the coracoids though these are proportionally smaller than in the intermediate-sized specimens. A still larger specimen of Rhamphorhynchus (JME SOS 4009) is not well preserved but shows a rectangular sternal plate that is wider than long, having a small triangular extension in the middle of the distal face. It has a long and robust cristospine that is ventrally tall and extends posteriorly into a broad ridge running the length of the sternal plate. At least one specimen (Claessens et al., 2009, figure 2d) shows slight expansions on the lateral margin of the sternal plate to contact the sternal ribs.
Bellubrunnus is known from only a single young, but well-preserved, individual (Hone et al., 2012), that has an apparently complete sternum though it is partially covered by other elements. In overall shape it is broadly similar to that of the juvenile Rhamphorhynchus (Figure 7D). The cristospine is long and straight (Hone et al., 2012), and the sternal plate is fan-shaped and a little longer than wide and appears to have slightly thickened anterior margins. Two apparent ridges diverging from the midline across the face of the sternal plate are breaks or distortions based on underlying elements and are not anatomical features.
Wellnhofer (1978) illustrated a sternum of Dorygnathus as being triangular and longer than wide with a convex distal margin and clear articulation points on each side for the sternal ribs. However, the sternum of Dorygnathus is highly variable (Padian, 2008a, figure 20C) varying from triangular to rhombus shaped with varying proportions (Figure 7E). The most unusually shaped of those illustrated by Padian (2008a) was suggested to be misidentified parietals, and that is considered more likely here. Padian (2008a) described several of these sterna briefly, perhaps the most notable of which was one that in contrast to the others had no cristospine, pneumatic foramen or articulation points for sternal ribs. Some specimens are described as having a convex dorsal margin and with a midline keel on the sternal plate and a pneumatic foramen in the centre of the sternum near the anterior margin and with a scalloped posterior margin (Padian, 2008a). The smallest specimen known has a notch at the top of the sternum and a poorly preserved cristospine (Padian, 2008a). One illustrated by Wiman (1925) appears to show facets on the ventral face of the sternal plate, though close to the lateral margins and with a pronounced keel.
The recently named giant rhamphorhynchine Dearc has a small and incomplete sternum preserved (Jagielska et al., 2022). It is unusual in being somewhat heart-shaped but with broad and rectangular lateral ‘wings’ that extend anteriorly. The lateral and posterior margins are nearly straight and the cristospine is missing.
Although Orientognathus was not described as having a sternum (Lü et al., 2015), one may be present. There is a large, thin and somewhat indistinct piece of bone to the side of the middle dorsal vertebrae (Lü et al., 2015, figure 4C). This has some displaced dorsal ribs aligned with it suggesting that they originally connected to the element and it may be a sternal plate. If so, it is broad and thin, but no other details are discernible.
Wukongopteridae. The holotype of Darwinopterus Lü et al. (2010) has a poorly preserved patch of bone near the scaplocoracoids that is likely a sternal plate, though little can be said about it other than it is quite broad and thin. A specimen of Darwinopterus robustodens is preserved in dorsal view and illustrated by Lü et al. (2011a, figure 4m). The sternal plate is roughly triangular in outline with rounded edges and smooth margins. The cristospine is short and robust with a sharp midline edge. Lü et al. (2011a) describe the coracoid articulations as being ‘distinct and slot-like’ but no other details are given.
One specimen of Darwinopterus (Lü et al., 2011b) has been suggested to actually belong to Kunpengopterus by Zhou et al. (2021), although Lü et al., (2011b) never identified to which species it may belong, and the wukongopterids as a whole are in need of clear taxonomic revision. This specimen is notable for being preserved with an egg and can be confidently identified as female having at least reached reproductive maturity. The sternum is difficult to discern in the available photograph (Lü et al., 2011b, figure 1) but is roughly diamond shaped with rounded and smooth edges. The anterior margins are thickened. The base of the cristospine appears to be broad, but no other information can be gained on its morphology.
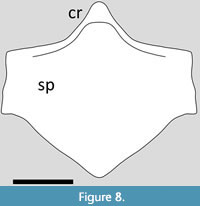 The holotype of Kunpengopterus preserves a near complete sternal plate in dorsal view, though this has disarticulated (Wang et al., 2010). It was described as being ‘heart shaped’ by Wang et al. (2010) and is sub-triangular with smooth and rounded edges, though with slight lateral expansions (Figure 8). Overall, this makes it somewhat intermediate in shape between that of the Darwinopterus of Lü et al. (2011b) and Chanchengopterus described below. The anterior margins are thickened. Only the base of a cristospine is preserved and this is broad.
The holotype of Kunpengopterus preserves a near complete sternal plate in dorsal view, though this has disarticulated (Wang et al., 2010). It was described as being ‘heart shaped’ by Wang et al. (2010) and is sub-triangular with smooth and rounded edges, though with slight lateral expansions (Figure 8). Overall, this makes it somewhat intermediate in shape between that of the Darwinopterus of Lü et al. (2011b) and Chanchengopterus described below. The anterior margins are thickened. Only the base of a cristospine is preserved and this is broad.
Douzhanopterus (Wang et al., 2017) preserves a sternum, though this is broken into two parts. As preserved, there is no cristospine or keel, and the sternal plate is approximately square, with a straight posterior margin but the anterior margin rising towards the midline (Wang et al., 2017) suggesting a more pentagonal shape in the complete sternum.
A sternum is known in an adult Changchengopterus and, although partly covered by other elements, is illustrated (Zhou and Schoch, 2011, figure 2). The sternal plate is roughly triangular in shape, with rounded margins and is rather wider than long. The margins are smooth and lack any costal articulations. Only part of the base of the cristospine is visible and is clearly robust.
One specimen of Wukongopterus preserves a sternum, but this is almost entirely covered by other elements, and the only comment on it by Wang et al. (2009) was that it is well ossified.
The currently unnamed ‘Painten pro-pterodactyloid’ is represented by a single young individual (Tischlinger and Frey, 2013). The sternum is present but almost entirely covered by other elements, and no details are available other than to say that it is clearly thin.
Ctenochasmatidae. The sternum of Elanodactylus is incomplete and broken (Andres and Ji, 2008) and although the images available are not clear, Andres and Ji provide some details. The sternum has straight lateral margins, with at least four clear rib facets one each side, which are more developed anteriorly than posteriorly (Andres and Ji, 2008).
Forfexopterus has a long, narrow and sharply pointed cristospine, which is described as having a medial ridge and is not constricted at the base (Jiang et al., 2016). This has asymmetrical and broad, oval-shaped facets from the coracoids and a pneumatic opening set in a depression at the base of the cristospine (Jiang et al., 2016). The sternal plate is longer than wide and trapezoid in outline with two straight edges on each side followed by a slightly curved posterior edge. The anterior margin is thickened, and there are no rib facets visible (Jiang et al., 2016).
Although there are many known specimens of Pterodactylus (Figure 9A), few seem to preserve even a partial sternum, although the holotype (BSP AS I 739) has one that is partially covered by other elements and another (BSP 1924 V 1) is partially preserved. These match the drawing of Wellnhofer (1978, figure 8) showing the sternal plate to be roughly triangular, though with the two lateral corners being ‘cut off’. The margins are smooth and lack articulations for costal ribs, and there is a thickened rim along the anterior margin (that is also visible in BSP 1924 V 1). The cristospine is long and very narrow and pointed anteriorly. There is a circular hole in the middle of the anterior part of the plate of BSP 1924 V 1, which could simply be damage but given the missing cristospine would sit over this point, this could be the remnants of pneumatopore penetrating the dorsal surface of the cristopsine. It is about the right size, shape and position of a pneumatopore of other pterodactyloids and suggests it was also present in Pterodactylus.
One specimen of Aerodactylus (NHMW 1975/1756) partially preserves a sternum, though this is partly represented by a stain / impression in the matrix. The sternal plate is very similar in shape to that of Pterodactylus being generally triangular with sharp inward turning angles at the lateral margins and smooth edges. Although the cristospine is not easily discernible, it appears to be a short and narrow spike extending anteriorly as in Pterodactylus.
The recently described Balaenognathus has a sternum preserved in dorsal view, but it is mostly hidden by other elements (Martill et al., 2023). However, the sternal plate has a rounded pentagonal outline with a rounded posterior terminus (Martill et al., 2023) and appears to be wider than long and with no apparent thickening at the margins or any articulation points.
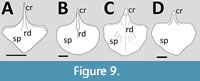 A partial sternum is preserved in Ardeadactylus (Jiang et al., 2016, figure 1F based on Meyer, 1854) (Figure 9C). The sternal plate is shovel-shaped in outline with smooth edges and a strong medial ridge. There are several ridges radiating out from the midline of the plate towards, but not reaching, the edge. The cristospine is broken and shown to be hollow suggesting that it was pneumatic, though the bone walls appear to be relatively thick.
A partial sternum is preserved in Ardeadactylus (Jiang et al., 2016, figure 1F based on Meyer, 1854) (Figure 9C). The sternal plate is shovel-shaped in outline with smooth edges and a strong medial ridge. There are several ridges radiating out from the midline of the plate towards, but not reaching, the edge. The cristospine is broken and shown to be hollow suggesting that it was pneumatic, though the bone walls appear to be relatively thick.
The holotype of Cycnorhamphus has a well-preserved sternum, which is fan-shaped with a semi-circular outline (Bennett, 2013a) (Figure 9B). The cristospine is relatively short and broad and does not appear to project far ventrally. The sternum has smooth margins and appears to have an anteriorly thickened margin.
Huanhepterus is not well illustrated and the only image of the sternum available is an interpretative drawing (Dong, 1982, figure 2). This shows the sternum to be large and nearly diamond-shaped with straight margins and no indication of any cristospine or any articulations for sternal ribs, which fits the short description of the text (Dong, 1982). A cristospine was presumably present originally, but without further information available there is little that can be added.
The holotype specimen of Auroroazhdarcho has a very well-preserved sternum that is seen in dorsal view (Frey et al., 2011). This has a triangular and short cristospine, and the plate has a thickened anterior margin (Figure 9D). Overall, it is ‘shovel shaped’ (Frey et al., 2011) with straight lateral sides, thought the posterior margin appears to be convex on one side and slightly concave on the other. It is described as having three ‘lateral pits’ (Frey et al., 2011) for sternal ribs, which I interpret to mean it has four articulation points.
The sternum of Gegepterus is very poorly preserved and very little information is visible (Wang et al., 2007). It was described as being broad and thin, with the posterior edge convex in shape and at least three articulation points for sternal ribs (Wang et al., 2007).
Although too poorly preserved to be described, fragments of a sternum are present on one specimen of Altmuehlopterus rhamphistinus (BSP.AS.I 745). A more complete one is preserved on a juvenile specimen (LF 2086P). This is seen in ventral view, and is shovel-shaped in outline and has only a small bulge on the proximal margin. This is far too small to articulate with the coracoids suggesting that this is not the cristospine but an expansion of the sternal plate. There is a thickened anterior rim to the plate, and although there is no obvious midline ridge, it does appear to have thinner (or at least different coloured bone) either side of the midline in a pair of approximately oval patches. There are several holes in the sternal plate, but these are irregular and asymmetrically placed and appear to just be damage, though they do show that the plate is very thin (<1 mm, for an animal with a skull length of 120 mm) even on the anterior part of the plate.
There is a sternum known for Prejanopterus (Pereda-Superbiola et al., 2012), a pterosaur of uncertain affinities that may sit close to the Ctenochasmatoidea and so is included here. It was not illustrated by Pereda-Superbiola et al. (2012) but was described as missing the cristospine, with a sternal plate that is longer than wide, being subrectangular in shape with a pronounced keel.
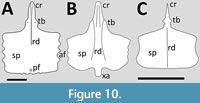 Pteranodontidae. A number of sterna are known for Pteranodon (Bennett, 2001) though many are in poor condition. However, this is to date the best known, and the best illustrated and described pterosaur sternum for any taxon, and can be seen in both ventral and dorsal views (Bennett, 2001) (Figure 10A). The cristopine is quite long and rounded anteriorly and has a distinct step posteriorly where the cristospine expands laterally for the coracoid facets (Bennett, 2001). The facets lie on the lateral and posterior margins of the cristospine and are symmetrical (Bennett, 2001). There is a strong ridge along the ventral face of the cristospine extending onto the anterior half to two-thirds of the sternal plate (Bennett, 2001), which is most clearly seen in Claessens et al. (2009, figure 2e). This ridge shows rugsose muscle scars and is suggested to extend ventrally as a cartilaginous structure (Bennett, 2001). There is also a thin midline ridge on the dorsal side of the cristospine (Bennett, 2001 - which he termed a ‘keel’) until it meets the coracoid facets where it bifurcates and extends around their anterior margins (Bennett, 2001). Posterior to the facets, at the base of the cristospine in dorsal view, is a circular pneumatopore, and dorsal to this in at least some specimens is a small tubercle (Bennett, 2001). On the ventral surface of the base of the cristospine, dorsal to the coracoid facets, are a pair of small tubercles (one each side), which Bennett (2001) suggests may be attachments for sternocoracoid ligaments.
Pteranodontidae. A number of sterna are known for Pteranodon (Bennett, 2001) though many are in poor condition. However, this is to date the best known, and the best illustrated and described pterosaur sternum for any taxon, and can be seen in both ventral and dorsal views (Bennett, 2001) (Figure 10A). The cristopine is quite long and rounded anteriorly and has a distinct step posteriorly where the cristospine expands laterally for the coracoid facets (Bennett, 2001). The facets lie on the lateral and posterior margins of the cristospine and are symmetrical (Bennett, 2001). There is a strong ridge along the ventral face of the cristospine extending onto the anterior half to two-thirds of the sternal plate (Bennett, 2001), which is most clearly seen in Claessens et al. (2009, figure 2e). This ridge shows rugsose muscle scars and is suggested to extend ventrally as a cartilaginous structure (Bennett, 2001). There is also a thin midline ridge on the dorsal side of the cristospine (Bennett, 2001 - which he termed a ‘keel’) until it meets the coracoid facets where it bifurcates and extends around their anterior margins (Bennett, 2001). Posterior to the facets, at the base of the cristospine in dorsal view, is a circular pneumatopore, and dorsal to this in at least some specimens is a small tubercle (Bennett, 2001). On the ventral surface of the base of the cristospine, dorsal to the coracoid facets, are a pair of small tubercles (one each side), which Bennett (2001) suggests may be attachments for sternocoracoid ligaments.
The sternal plate is nearly square in outline and scalloped on both the lateral and posterior margins (Wellnhofer, 1978; Bennett, 2001), although at least some specimens show a smoother lateral margin (e.g., Bennett, 2001, figure 60). The plate is strongly concave in overall shape, although many specimens are preserved as if this was flat due to crushing. It is thinner posteriorly than anteriorly, though apparently still pneumatic even in the posterior part (Bennett, 2001). There are thickened margins along anterior edge of the sternal plate. (Bennett, 2001). Wellnhofer (1978, figure 8) illustrates the plate with a large and circular foramen in the middle of the ventral part of the sternal plate. Posterior to this foramen, Bennett notes that gastralia may be incorporated into the sternum (Bennett, 2001).
The sternum of Nyctosaurus is rectangular in outline and is rather wider than it is long (Figure 10B). The cristospine is long and broad at the base and then narrows anterior to the coracoid articulations before tapering to a point. Williston (1893) described the articulations as being saddle shaped and present on both the dorsal and lateral surfaces of the cristospine. These are illustrated as being symmetrical based on figure in Williston (1897). The sternal plate is thickened anteriorly and shows four lateral projections on each lateral face Williston (1893). The ventral margin in biconcave with a clear midline point that expands distally into a ‘xiphisternal process’ (Williston, 1893), that Williston described as being ‘spatulate and distally rounded’. Wellnhofer (1978) illustrates the sternum as having a broad ridge down the middle of the cristospine and the proximal two-thirds of the sternal plate.
A sternum is preserved in the holotype of Musquizopteryx (Frey et al., 2006). The sternal plate is large and subrectangular, slightly wider than long, and with a long and pointed cristospine (Figure 10C). There is a large foramen at the base of the cristospine, and it has a pair of articulations for the coracoids, but Frey et al. (2006) do not state or show if these are symmetrical or not. Frey et al. (2006) describe the plate as having a concave distal margin, but it is illustrated (Frey et al., 2006, figure 2A) as having a convex one and it appears to be biconcave with a midline point in their figure 3 as seen in Nyctosaurus. The lateral margins are described as being ‘wavy’ (Frey et al., 2006), which suggests scalloping and points for articulation with sternal ribs.
Istiodactyliformes. The early branching istiodacyliform Mimodactylus preserves a sternum in right lateral view (Kellner et al., 2019a). The cristospine is short but broad (Kellner et al., 2019a) and distally expanded, a little like those of azhdarchids but not as hook-shaped (see below). Only half of the sternal late is visible and appears to have been folded in half along the midline or that the second half is simply missing. The visible sternal plate is rectangular, but when complete would have been overall square in shape and probably with a straight lateral edge and a convex posterior margin. The anterior margin was described as being rounded by Kellner et al. (2019a). A midline ridge runs along the cristospine onto the face of the plate, perhaps as far as the posterior margin.
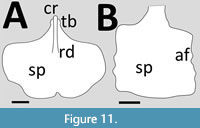 The sternum of Haopterus was described by Wang and Lü (2001) as being fan-shaped, although it might be better considered as semi-circular (Figure 11A). It has a long cristospine, with a strong midline keel that extends onto the anterior half of the sternal plate. The sternal plate itself is about as wide as long and is thin (Wang and Lü, 2001).
The sternum of Haopterus was described by Wang and Lü (2001) as being fan-shaped, although it might be better considered as semi-circular (Figure 11A). It has a long cristospine, with a strong midline keel that extends onto the anterior half of the sternal plate. The sternal plate itself is about as wide as long and is thin (Wang and Lü, 2001).
The sternum of Nurhachius is poorly preserved and was not described in the original paper of Wang et al. (2005). However, Jiang et al. (2016, figure 1K) illustrated this interpreting it as preserving an intact cristospine and only half of the sternal plate being present. As there appears to be an articular facet for the coracoid visible on the cristospine, which would make this preserved in dorsal view. The cristospine appears unusually robust, and this may be crushed and distorted, especially given the presence of only a single facet, although Kellner et al. (2019a) described this as being like that of Mimodactylus. When complete, the sternal plate would be somewhat fan-shaped overall though with straight lateral edges and a convex ventral face. All of these edges are smooth and with no evidence of scalloping or serrations.
The sternum of the large, juvenile istiodactylid Luchibang is large and rectangular in outline being only slightly longer than wide (Hone et al., 2020) (Figure 11B). Only a trace of a cristospine was preserved with a small extension of the middle of the anterior margin of the sternal plate (Hone et al., 2020). The lateral margins of the sternal plate show serrations but the posterior margin of the sternum is straight (Hone et al., 2020).
Although the holotype of Istiodactylus is best known for the well-preserved cranial material, Howse et al. (2001) describe this as having a partial sternum. They describe saddle-shaped articular facets for the coracoids that are asymmetrically arranged, and these can be seen in Plate XL of Hooley’s (1913) description. The piece in question (NHM R 176) is a robust piece of the cristospine that appears to be slightly hooked anteriorly. Seeley (1901, figures 67 and 68)) illustrated and noted that the cristospine was large and dorsoventrally deep and is shown with a distinct ridge along its ventral margin.
Ornithocheiridae. Although Hamipterus is known from multiple specimens preserved in three dimensions, so far there is no description of a sternum and only one partial sternum has been figured (Wang et al., 2014, figure 1). It is not as well preserved as neighbouring elements suggesting that it may be poorly ossified. It is preserved in dorsal view, based on the apparent depression near the base of the cristospine, and there may be a pneumatopore in the depression. The sternal plate is an unusual shape with laterally projecting broad triangles proximally and a semi-circular part distal to this, the maximum width of this is about the same as the anteroposterior length of the plate. The medial parts of the anterior margins are thickened, and the edge of the better preserved left side is smooth. The cristospine is relatively robust and is at least one-third of the length of the sternal plate.
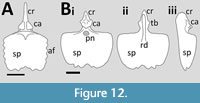 De Buisonjé (1981, figures 5, 8 and 10) illustrated reconstructions of the sternum of Santanadactylus brasilensis in dorsal, lateral and anterior views (Figure 12A). In dorsal view, the sternal plate is roughly square in outline, though with sinusoidal lateral and distal margins. It is slightly dished in shape with a thickened anterior rim and no indication of any pneumatopores. The cristospine is relatively long and tapers anteriorly, and there are clear symmetrical semi-circular facets for the coracoids at the base. In lateral view the cristospine can be seen to be convex in outline with a straight dorsal margin and a slight lateral expansion posterior to the coracoid articulation. A total of six costal ribs are shown to articulate with the sternal plate. (de Buisonjé, 1981, figure 10). In anterior view, the sternal plate is strongly dished and U-shaped with the dorsalmost part of the coracoid articulations being approximately level with the dorsalmost point of the lateral margins of the sternal plate.
De Buisonjé (1981, figures 5, 8 and 10) illustrated reconstructions of the sternum of Santanadactylus brasilensis in dorsal, lateral and anterior views (Figure 12A). In dorsal view, the sternal plate is roughly square in outline, though with sinusoidal lateral and distal margins. It is slightly dished in shape with a thickened anterior rim and no indication of any pneumatopores. The cristospine is relatively long and tapers anteriorly, and there are clear symmetrical semi-circular facets for the coracoids at the base. In lateral view the cristospine can be seen to be convex in outline with a straight dorsal margin and a slight lateral expansion posterior to the coracoid articulation. A total of six costal ribs are shown to articulate with the sternal plate. (de Buisonjé, 1981, figure 10). In anterior view, the sternal plate is strongly dished and U-shaped with the dorsalmost part of the coracoid articulations being approximately level with the dorsalmost point of the lateral margins of the sternal plate.
A sternum is known from the well-preserved subadult specimen of Anhanguera piscator described by Kellner and Tomida, (2000) (Figure 12B). The following description is based on that of Kellner and Tomida (2000) with additional observations from unpublished photos of the specimen. Much of the sternal plate is missing or poorly preserved, but it is sufficient to determine all the major features of the element. The plate is about as wide as it is long and is approximately shovel shaped, although with a somewhat straight distal margin. The lateral edges are smooth, though the distal margin is sinusoidal (and slightly asymmetric). There is a strong and broad midline ridge (the sternal keel of Kellner and Tomida, 2000) that runs from near the distal margin of the sternal plate into the cristospine. In dorsal view, the plate has a distinct depression at the base of the cristospine with a large pneumatic foramen in the centre of this. The cristospine is robust and about half the length of the sternal plate. In dorsal view it has a pair of protuberances (Kellner and Tomida, 2000 refer to these as tubercles) that extend laterally and dorsally from its base, and between these a third and much smaller extension points distally. Anterior to these are the paired and symmetrical articular facets for the coracoids that are broad, flattened and extend laterally. Anterior to these, the spine tapers rapidly and extends ventrally, but with a thick midline ridge. In lateral view, the sternal plate is rather dished, and the cristospine has some odd serrated margins on the anterior tip and on the first (distal) pair of lateral protuberances.
One specimen of Ornithocheirus is illustrated by Wellnhofer (1978, figure 8 - listed as Criorhynchus) though it is a very partial anterior sternal plate with a complete cristospine shown in dorsal view. Little information can be gleaned from what remains of the sternal plate, though it appears to lack any depression or pneumatopore at the base of the cristospine. The preserved cristospine is short and robust, with a slight midline ridge. As Bennett (2001) notes, the preserved cristopsine has symmetrical facets for the coracoids and these are unusual in being mediolaterally elongate.
A very large specimen of Tropeognathus representing the largest known toothed pterosaur has a partial sternum preserved (Kellner et al., 2013). Overall this is very similar to that of other ornithochierids. The sternal plate is mostly complete and shows that it was wider than long with concave anterior margins, convex lateral margins and a somewhat crenelated distal margin. The cristospine is short and robust with an expanded central part around the symmetrical coracoid facets. A large dorsally projecting tubercle as shown on the dorsal face of the cristospine is illustrated by Kellner et al. (2013, figure 6e) though based on the available lateral view of the specimen this may be more of a ridge along the posterior margin of the fact bulge.
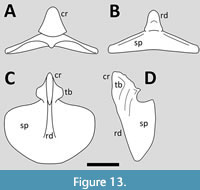 Veldmeijer (2006) described a well-preserved sternum of Coloborhynchus spielbergi that showed only limited deformation and damage, although it has been incompletely prepared and not all of the dorsal surface is visible (Figure 13). The sternal plate is roughly semi-circular in outline although the margins of this are not well preserved. As seen in anterior view, the plate is convex in shape with a thickened anterior margin to this on the dorsal surface. The cristospine is short and triangular and extends onto the ventral surface of the plate as a midline keel that runs for about half the anteroposterior length of the sternal plate. The cristospine has broad lateral expansions for the coracoid articulation facets, which are symmetrical and lie on the lateral and dorsal faces of the spine. A thickened rim runs across the dorsal face of the cristospine, immediately anterior to the margins of the facets. On the dorsal face of the cristospine there is a single medium-sized foramina between and slightly posterior to the facets, and there is a much larger foramen on the anterior part of the sternal plate.
Veldmeijer (2006) described a well-preserved sternum of Coloborhynchus spielbergi that showed only limited deformation and damage, although it has been incompletely prepared and not all of the dorsal surface is visible (Figure 13). The sternal plate is roughly semi-circular in outline although the margins of this are not well preserved. As seen in anterior view, the plate is convex in shape with a thickened anterior margin to this on the dorsal surface. The cristospine is short and triangular and extends onto the ventral surface of the plate as a midline keel that runs for about half the anteroposterior length of the sternal plate. The cristospine has broad lateral expansions for the coracoid articulation facets, which are symmetrical and lie on the lateral and dorsal faces of the spine. A thickened rim runs across the dorsal face of the cristospine, immediately anterior to the margins of the facets. On the dorsal face of the cristospine there is a single medium-sized foramina between and slightly posterior to the facets, and there is a much larger foramen on the anterior part of the sternal plate.
The boreopterid Zhenyuanopterus preserves a sternum (Lü, 2010), though the articulated skeleton is preserved in dorsal view so this is mostly covered by other elements. Lü et al. (2010) describe the sternal plate as being wider than long, but the total length of the element including the cristospine is subequal to the width.
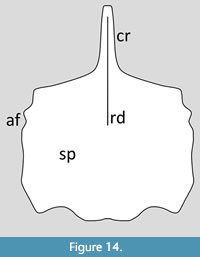 Dsungaripteridae. Young (1973) figured a single sternum from Dsungaripterus, though the photograph of this specimen in Jiang et al. (2016, figure 1N) is rather clearer (Figure 14). The cristopine is long, straight and anteriorly forms a sharp point. There is a sharp midline edge to this than continues down on to the anterior half of the face of the sternal plate. Bennet (2001) notes that the coracoid facets are asymmetric in Dsungaripterus. The sternal plate is overall square in shape, though it is slightly wider than long and as a whole is convex. There are serrations on the lateral margins, though this is somewhat asymmetric being present on the right edge but not the left, probably as a result of poor preservation or incomplete ossification. The posterior margin is biconcave with a single midline point.
Dsungaripteridae. Young (1973) figured a single sternum from Dsungaripterus, though the photograph of this specimen in Jiang et al. (2016, figure 1N) is rather clearer (Figure 14). The cristopine is long, straight and anteriorly forms a sharp point. There is a sharp midline edge to this than continues down on to the anterior half of the face of the sternal plate. Bennet (2001) notes that the coracoid facets are asymmetric in Dsungaripterus. The sternal plate is overall square in shape, though it is slightly wider than long and as a whole is convex. There are serrations on the lateral margins, though this is somewhat asymmetric being present on the right edge but not the left, probably as a result of poor preservation or incomplete ossification. The posterior margin is biconcave with a single midline point.
Azhdarchoidea. An indeterminate and immature azhdarchoid skeleton was described by Elgin and Hone (2013) that preserves a sternum. The cristospine is damaged but appears to be short (Elgin and Hone, 2013), and the sternal plate is slightly longer then broad with a convex posterior margin (the lateral margins are not visible). It is overall strongly convex in shape rather than flat. There are concavities to the lateral faces of the cristospine, which show small foramina and coupled with the loss of the bone cortex on the cristospine showing trabeculae suggests that this element was pneumatic.
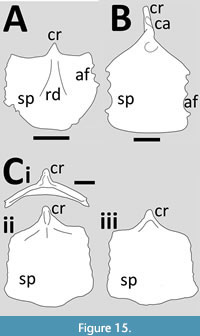 Tapejaromorpha. Cauiajara is known from multiple well-preserved specimens though the sternum has yet to be described in detail, although three have been illustrated (Manzig et al., 2014, figures 6 and 7). It has been described as being semi-circular in shape (Manzig et al., 2014) although this is more like a rectangular plate with a convex posterior margin (Figure 15A). It has convex anterior margins either side of the short and triangular cristospine and several rounded serrations on the lateral faces. Manzig et al.’s figure 7 shows a poorly ossified sternum from an osteologically immature adult (based on the lack of fusion of the carpals). The sternal plate is very thin, wide and may have a sinusoidal lateral margin.
Tapejaromorpha. Cauiajara is known from multiple well-preserved specimens though the sternum has yet to be described in detail, although three have been illustrated (Manzig et al., 2014, figures 6 and 7). It has been described as being semi-circular in shape (Manzig et al., 2014) although this is more like a rectangular plate with a convex posterior margin (Figure 15A). It has convex anterior margins either side of the short and triangular cristospine and several rounded serrations on the lateral faces. Manzig et al.’s figure 7 shows a poorly ossified sternum from an osteologically immature adult (based on the lack of fusion of the carpals). The sternal plate is very thin, wide and may have a sinusoidal lateral margin.
Keresdrakon was described by Kellner et al. (2019b) as an early branching member of Tapejaramorpha. The sternum is broken with the cristospine and the proximalmost part of the sternal plate separate from the main part (Kellner et al., 2019b). It has a long, robust and slightly hook-shaped cristospine (Figure 15B). The dorsal face of the cristospine shows a single asymmetrical articulation facet for the coracoids thus they presumably both articulate against this. The sternal plate is large, flat and square in shape. This has a smooth and convex posterior margin with a slightly concave lateral margin that is scalloped with four clear peaks to articulate with sternal ribs. The dorsal face of the proximal part of the sternum shows a large foramen (Kellner et al., 2019b) which is likely a pneumatopore. There are holes on the distal part of the sternal plate and a serrated posterior margin, but these are the result of poor preservation and / or damage.
An undescribed sternum from Tupuxuara is well-preserved in three dimensions (IMNH 1052). The plate is roughly square in outline with a convex ventral margin. The lateral sides are serrated with at least four articulation points on the left and only three on the right. The whole plate in concave in shape and has a ridge running from the cristospine across the whole length of the ventral face. The cristopsine is narrow, sharply pointed and lacks any obvious depressions to mark the coracoid articulations. On the dorsal surface there is a clear ridge and expansion around the midpart of the cristospine, and at the base there is a deep semi-circular depression.
Wang and Zhou (2002) were the first to illustrate a sternum for Sinopterus dongi, a lthough this shows no details and even the shape of the outline is unclear. Based on a photo of the specimen in question (IVPP V 13363), this can at least be seen to have a partially preserved cristospine that is short and narrow. The sternal plate is not especially thin though the lateral margins are covered by / merge with other elements. Subsequent descriptions of the sternum of this genus by Lü et al. (2006a) and Zhang et al. (2019) were based on different species.
Lü et al. (2006a) describe the sternum of S. dongi as being poorly preserved and less well ossified distally. They say that the lateral margins are parallel to the midline axis (i.e., it is square or rectangular in shape) and there are no facets for sternal rib articulations. However, Lü et al. (2006a, figure 1) show that most of the right margin of the plate is covered by other elements, and the left side does show some crenulations that could be damage or at least conceivably be sternal articulations. The distal margin of the sternal plate is also slightly convex.
The sternum of Sinopterus atavismus is described by Zhang et al. (2019) as being broad and thin with a depression in the middle and being thicker anteriorly than posteriorly. They specifically describe it as having articulations for posterior sternal ribs, though in a reversal of the issue with Lü et al. (2006a) these are not clear at all from the figures (Zhang et al., 2019, figures 1 and 2), and the ragged margins of the poorly preserved sternal plate make this identification uncertain. Zhang et al. (2019) go on to describe the cristopine as being a “short triangle, [that] tapers gradually toward its anterior end, and whose articulations with the coracoids are asymmetric”. Both specimens appear to be osteologically immature based on the lack of fusion of numerous major elements in each, so these apparent differences in sternal articulations may not be the result of ontogeny, though as noted, neither description is well supported by the illustrations, and it is possible that both or neither actually have sternal articulations.
A sternum is preserved in Huaxiapterus, though it is incomplete and partially preserved as an impression (Lü and Yuan, 2005). It was described by Lü and Yuan (2005) as being rectangular in shape and longer than wide. The distal part of the plate appears to be less well preserved and by extension less well ossified given the well-preserved nature of other nearby elements. The cristospine is either not present or covered by other elements.
A sternum is known for one three dimensionally preserved, young specimen (based on the lack of fusion of almost any major elements) of Tapejara wellnhoferi (Eck et al., 2011) (Figure 15C). The sternum is nearly square in outline with a slight expansion distally, which with the base of the cristospine makes it nearly hexagonal with sinusoidal margins and articulations for approximately five costal ribs (Eck et al., 2011, figures 6A-C). In anterior view, the sternal plate is a gentle U-shape with both sides rising dorsally. On the dorsal side of the sternum there is a depression near the anterior under the base of the cristospine and in the depression is a pneumatic foramen. The cristospine is anteroposteriorly very short and dorsoventrally short, too, as was described as being worn by Eck et al. (2011) and is probably not complete.
Recently, a nearly complete subadult specimen of Tupandactylus (Beccari et al., 2021) has been described. The sternum is preserved in three dimensions, though it is partially covered by matrix obscuring some details. The sternum was described as being roughly square-shaped, although the presented CT scan of the specimen suggests that the sternum is longer than wide, although the margins are not well preserved. It was suggested by Beccari et al. (2021) that the sternal plate was originally strongly convex on the ventral face though is now compressed, has been flattened and has a pneumatic opening below the cristospine. The anterior margins of the sternal plate are slightly concave and lead up to a short and incomplete cristospine.
A sternum is preserved for the holotype of Eopteranodon (Lü and Zhang, 2005) though this is very hard to make out in the photos and drawings of the original description. It appears to be large and rectangular, being slightly longer than wide with smooth margins. A referred specimen (Lü et al., 2006b) also preserved a sternum that was rectangular, longer than wide and described as having a short sternal keel, though this could refer to the cristospine or the midline ridge seen in many sterna. Lü et al. (2006) noted that the anterior portion was well preserved, but the distal part was ‘only weakly fossilized’ compared to the anterior part. Both specimens of Eopteranodon show evidence of osteological immaturity such as unfused carpals so this could well represent incomplete ossification of the distal sternum.
Caupedactylus has a partially preserved sternum with no cristospine and the sternal plate preserved in ventral view (Kellner, 2013). The sternal plate is roughly square in outline, with no medial ridge, though there is a mound in the middle of the proximal part, which would have formed the base of the cristospine (based on Kellner, 2013, figure 8). This is described as having several costal articulations on the right side (Kellner, 2013).
The sternum of an indeterminate tapejarid from Brazil is also known (MN 6558-V). The surface of this is poorly preserved and fragmentary, but its overall shape and margins are well-preserved, and it appears to be undistorted apart from some limited crushing. It is slightly longer than wide, but rectangular in overall shape with a near straight anterior margin, six rounded serrations down each side (with the posteriormost being noticeably larger than the others) and a slightly convex ventral margin. The sternal plate has a broad but shallow midline concavity, suggesting that the sternum is preserved in dorsal view but this is difficult to confirm. The anterior part of the plate is damaged with the surface bone missing and shows that the internal part was extensively pneumatic almost to the anterior and lateral margins, and extends anteriorly into the base of the cristospine at least. The cristospine is short, broad and rounded with a slight expansion each side near its base, presumably as part of lateral expansions around the coracoid articulations.
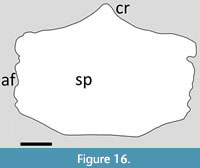 Chaoyangopteridae. The sternum of Jidapterus is preserved in dorsal view and is approximately rectangular in shape and rather wider than long with a wide expansion on the posterior face (Wu et al., 2017) (Figure 16). The cristospine is relatively small and does not project far anteriorly from the sternal plate (Wu et al., 2017). The lateral and posterior margins are slightly irregular, but their nature suggests that this is as a result of poor preservation rather than these being asymmetrical articulations for the costal ribs, though this is a possibility. The sternal plate is thick anteriorly and becomes thinner posteriorly (Wu et al., 2017).
Chaoyangopteridae. The sternum of Jidapterus is preserved in dorsal view and is approximately rectangular in shape and rather wider than long with a wide expansion on the posterior face (Wu et al., 2017) (Figure 16). The cristospine is relatively small and does not project far anteriorly from the sternal plate (Wu et al., 2017). The lateral and posterior margins are slightly irregular, but their nature suggests that this is as a result of poor preservation rather than these being asymmetrical articulations for the costal ribs, though this is a possibility. The sternal plate is thick anteriorly and becomes thinner posteriorly (Wu et al., 2017).
Azhdarchidae. The sternum of Zhejiangopterus is illustrated in a drawing of the specimen by Cai and Wei (1994). The cristospine is shown (Cai and Wei, 1994, figure 5) to be robust and distally is spreads out such that it occupies some of the face of the sternal plate. Although a photo of the specimen is included in the paper, the sternum cannot be made out. The sternum is illustrated as being much longer than wide (Cai and Wei, 1994, figure 5) with broken or incomplete lateral margins and a deep V incised into the posterior margin. In their reconstructed skeleton, however, Cai and Wei (1994, figure 6) illustrate the sternum as being approximately square and slightly longer than wide, with slightly scalloped lateral margins.
Two very fragmentary sterna of Azhdarcho were described by Averianov (2010). These consist of two long and robust, hook-shaped cristospines. A part of the sternal plate is present for one of these, and although hidden under the matrix, is described as having a deeply concave dorsal face (Averianov, 2010). The coracoid articulations on these specimens are asymmetric with the coracoid facts starting on the lateral faces and wrapping around onto the anterior face.
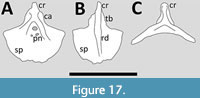 Bennett (2001) noted that the articular facets for the coracoids are asymmetric in Quetzalcoatlus but as there are no descriptions or photographs of the sternum for this taxa, this was all that was known until recently. However, new descriptions of this genus by Padian et al. (2021) and Andres and Langston (2021), that include several partial sterna, have added considerable knowledge of this azhdarchid (Figure 17). Padian et al. (2021) describe a sternum of Quetzalcoatlus lawsoni with a cristospine and partial anterior part of the sternal plate. The plate is thin and its anterior margin projects anteriorly to meet the base of the cristospine, and the plate is strongly concave in anterior view. Andres and Langston (2021) add to this that the plate is posteriolateral margins and that the plate as a whole is somewhat rhombic in outline, though the angles at which the anterolateral and posteriolateral margins meet is rather variable between the three specimens. The plate is also concave in anterior view (Andres and Langston, 2021). The cristospine is short, broad and triangular and the anterior part curves dorsally. On the dorsal face are a pair of broad ridges that run into the anterior margins of the plate, between them is a deep cavity in which lie one, two or three foramina the largest of which is anteriorly positioned and has a small ridge posterior to it and anterior to the other openings when present (Andres and Langston, 2021). A second specimen preserves articular serrations on one side of the plate for up to four sternal ribs (Padian et al., 2021). Both specimens show asymmetrical articulations for the coracoids on the dorsal face of the cristospine with the right anterior to the left, and both are saddle-shaped with an anterolateral orientation (Andres and Langston, 2021).
Bennett (2001) noted that the articular facets for the coracoids are asymmetric in Quetzalcoatlus but as there are no descriptions or photographs of the sternum for this taxa, this was all that was known until recently. However, new descriptions of this genus by Padian et al. (2021) and Andres and Langston (2021), that include several partial sterna, have added considerable knowledge of this azhdarchid (Figure 17). Padian et al. (2021) describe a sternum of Quetzalcoatlus lawsoni with a cristospine and partial anterior part of the sternal plate. The plate is thin and its anterior margin projects anteriorly to meet the base of the cristospine, and the plate is strongly concave in anterior view. Andres and Langston (2021) add to this that the plate is posteriolateral margins and that the plate as a whole is somewhat rhombic in outline, though the angles at which the anterolateral and posteriolateral margins meet is rather variable between the three specimens. The plate is also concave in anterior view (Andres and Langston, 2021). The cristospine is short, broad and triangular and the anterior part curves dorsally. On the dorsal face are a pair of broad ridges that run into the anterior margins of the plate, between them is a deep cavity in which lie one, two or three foramina the largest of which is anteriorly positioned and has a small ridge posterior to it and anterior to the other openings when present (Andres and Langston, 2021). A second specimen preserves articular serrations on one side of the plate for up to four sternal ribs (Padian et al., 2021). Both specimens show asymmetrical articulations for the coracoids on the dorsal face of the cristospine with the right anterior to the left, and both are saddle-shaped with an anterolateral orientation (Andres and Langston, 2021).
Andres and Langston (2021) state that a partial sternum is preserved for a specimen of Montanazhdarcho but this is not mentioned, described or illustrated in either of the two papers that described this taxon (Padian et al., 1995; McGowen et al., 2002).
DISCUSSION
Ontogeny of Pterosaur Sternum
Most taxa described here are represented by only a single sternum (or only one that is well-preserved) but there are some ontogenetic patterns here that point to a change in the sternum during ontogeny for various pterosaurs, and probably reflect a pattern that is true for the clade as a whole. For example, the shapes of the sterna of Eudimorphodon and Rhamphorhynchus (Figure 3 and Figure 7) show that the sternum gets broader over time and generally more elaborate around the margins. Similarly, there is strong variation in the sterna seen in some taxa (e.g., Dorygnathus, Quetzalcoatlus) and in some clades here even when their overall morphology and implied mechanics and ecology are extremely similar (e.g., ornithocheiroids, Figure 12 and Figure 13), which could represent taxonomic differences, but more likely represent changes in ontogeny being represented by animals of different ages being preserved or at least intraspecific variation in rates or degrees of ossification.
Juvenile pterosaurs, including even very young animals, appear to have been capable of powered flight (Hone et al., 2021; Naish et al., 2021) and at least some are highly conservative in their anatomy during ontogeny (Bennett, 1995) and show near isometric growth (Hone et al., 2021). Indeed, Rhamphorhynchus is extremely conservative in its anatomy during development, and I cannot think of another element or skeletal region of pterosaurs that changes a fraction of the degree that the sternum does. Although this genus grows isometrically, it would change in-flight performance as it grew as wing loading and wingspan would change at different rates (Hone et al., 2021). There would be an increase in muscle mass for larger and heavier adults compared to juveniles (Naish et al., 2021), so it is possible that the relative muscle attachments on the sternum might also change during growth and lead to the changing shape of this element.
The alternative to this is that the sternum shows limited ossification in juvenile pterosaurs and extensively ossifies later in ontogeny and that the variation in shape of the preserved specimens (both intra- and some interspecific) is as a result of this. It does seem unlikely that the major downstroke muscles would attach only to the thin and midline sternum seen in a juvenile Rhamphorhynhcus and the very young holotype of Bellubrunnus (Figure 7) or Anurognathus. A sternum that was mostly cartilage would provide a much larger surface area of these muscles, and although muscle attachment sites might change and grow, it also seems unlikely that juveniles would have had a sternum that is one-third or less of the area seen in adults. A cartilaginous expansion would also permit the sternal ribs to reach the sternum if this was considerably broader than as often preserved, and again it is unlikely that in Rhamphorhynchus for example the sternal ribs would simply ‘float’ in the chest cavity and not reach any part of the sternum. Changes in muscle mass as pterosaurs grew could also have been coupled with a change in muscle type as their flight capabilities altered. A move from more slow twitch muscles in juveniles to fast twitch in adults (who needed more power per unit muscle for take-off with greater wing loading) could have resulted in additional strain on the sternum and therefore potentially driving ossification in older and larger animals. The presence of an extensively, or even entirely, cartilaginous sternum in various reptile lineages (Bradley et al., 2019) strongly supports this possibility in pterosaurs.
Bennett (2014) suggests distal and lateral parts of a triangular sternum in a juvenile Scaphognathus were cartilage, and Wild (1993) noted that the juvenile Eudimorphodon specimen has a different surface texture distally. This could have been a result of a different origin of the element implying ossification of different parts at different times. Certainly a number of specimens described here have poorly preserved distal margins (e.g., Eudimorphodon, Forfexopterus, Keresdrakon) suggesting that they were not well ossified at the time of death, and indeed there are a number of pterosaurs whose bones are extremely well-preserved in general but the sternum is either absent or very poorly preserved (e.g., Darwinopterus linglongtaoensis, Musquizopteryx, Keresdrakon, Jidapterus), and supports the idea that it is often not well ossified, even in animals that were not young juveniles and could be considered subadult or even adult (sensu Kellner, 2015).
Finally, the variation seen in the costal articulations also points to ontogenetic changes and that much of the sternum in juveniles might be cartilaginous. In at least some Pteranodon specimens (e.g., YPM 2450), the costal ribs actually fuse to the sternal plate and certainly these would normally articulate with the sternum and likely had a major role in breathing (Claessens et al., 2009 and see below). The variations in the number and form of the costal articulations between the left and right sides of single stern (e.g., Tupuxuara, Anhanguera) indicate that not all of these articulations were of sufficient strength to generate expanded articulations and that not all of them ossified. Similarly, for specimens with narrow sterna anteriorly (e.g., juvenile Rhamphorhynchus) the ribs presumably still articulated with, and were attached to, the sternum in some way or it would not have been able to exert changes on the chest cavity and facilitate breathing. By extension, this implies the possibility that the ribs connected to a broader cartilaginous plate rather than were free floating in the chest bound only by muscles. Geist et al. (2014) also suggested that for larger pterosaurs at least, such a sternal pump would not be viable based on the size and articulations of some of the ribs, and that the sternum would be in a more fixed position in the chest giving an alternative for the importance of these articulations.
Some other ontogenetic changes are also observed in the sterna. While complete adults with well-preserved sterna show the cristospine fully fused to the sternum with no suture apparent, a number of specimens are similarly missing the cristopine (e.g., Luchibang) or represented by little more than the cristospine (Daohugouopterus, Ornithocheirus, Quetzalcoatlus). In some cases, these might relate to broken elements or a poorly preserved sternal plate, and indeed in at least some juvenile animals, the cristospine does appear to be completely part of the sternal plate (e.g., Rhamphorhynhcus). However, in others the separation of these components or the absence of the more robust cristopine but retention of the thinner plate is difficult to explain unless they were not well fused together. A separate cristospine would presumably not be a major issue for flight given that other major elements like the scapulae and coracoids are fully separate elements in juvenile pterosaurs and only fuse later in ontogeny (e.g., Kellner, 2015). While the forces at play in the scaplocoracoids would be rather different to the sternum, a lack of fusion should not a priori imply any issues for powered flight. It does appear that different taxa ossified the sternal plate and the cristospine at different times, and the variation seen and apparent absences in some cases is simply preservation based on different ossification rates.
Homologies of the Pterosaur Sternum
The ancestry of pterosaurs has been controversial and difficult to resolve for an extended period (Hone and Benton, 2007; Bennett, 2013b), but it has recently become clear that pterosaurs are derived archosaurs and lie close to the origin of the Dinosauromorpha (Ezcurra et al., 2020). This provides a strong starting point for establishing the possible homologies of the pterosaur sternum since its unusual morphology is quite unlike that of most other reptiles, although ongoing disagreements over which are the earliest pterosaurs (Baron, 2020) mean this remains a complex issue.
 The sternal complex of most reptiles has a pair of clavicles that contact the scapulae and project towards the midline. Here the clavicles meet the interclavicle, which is an arrow-shaped bone with a pair of rami meeting the laterally positioned clavicles and a medioposterior extension that meets the sternal plate (Figure 18). For early archosaurs the interclavicle may meet and articulate with the scapulae, coracoids, sternal ribs and even the gastralia (Bradley et al., 2019). The clavicles are absent in Crocodylia and in early dinosaurs, (e.g., Tawa - Bradley et al., 2019) the sternal complex consists of a furcula (fused clavicles) anteriorly and paired sternal plates behind (Figure 18B). Therefore, ancestral pterosaur condition might have had both an interclavicle and clavicles or potentially lacked one or the other, while possessing a sternal plate.
The sternal complex of most reptiles has a pair of clavicles that contact the scapulae and project towards the midline. Here the clavicles meet the interclavicle, which is an arrow-shaped bone with a pair of rami meeting the laterally positioned clavicles and a medioposterior extension that meets the sternal plate (Figure 18). For early archosaurs the interclavicle may meet and articulate with the scapulae, coracoids, sternal ribs and even the gastralia (Bradley et al., 2019). The clavicles are absent in Crocodylia and in early dinosaurs, (e.g., Tawa - Bradley et al., 2019) the sternal complex consists of a furcula (fused clavicles) anteriorly and paired sternal plates behind (Figure 18B). Therefore, ancestral pterosaur condition might have had both an interclavicle and clavicles or potentially lacked one or the other, while possessing a sternal plate.
 Based on a juvenile specimen of Eudimorphodon, Wild (1993) suggested that the pterosaur sternum is composed of the interclavicle, clavicles and two sternal plates. In his model, the thickened rim of the anterior part of the sternum would be formed from the two overlapping clavicles, with the interclavicle described as making up the cristospine and the central part of the sternum. Wild (1993) also noted that two parts of the sternum of the juvenile have a different bone texture to the others, and that this was potentially a result of them having different origins. These are broadly triangular sections, on each side of the main sternal plate contacting the posterior margin with a roughly M-shaped sternum surrounding them. Something similar is also seen in the juvenile specimen of Altmulopterus (LF 2086P) described here, which somewhat supports Wild’s contention. Under UV light, this shows a clear difference in colour with two dark patches on the sternal plate, one on each side of the base of a cristospine, and these fade out towards the margins of the element and suggest two separate condensation points and reinforce the idea that the lateral margins may be incompletely ossified (Figure 19).
Based on a juvenile specimen of Eudimorphodon, Wild (1993) suggested that the pterosaur sternum is composed of the interclavicle, clavicles and two sternal plates. In his model, the thickened rim of the anterior part of the sternum would be formed from the two overlapping clavicles, with the interclavicle described as making up the cristospine and the central part of the sternum. Wild (1993) also noted that two parts of the sternum of the juvenile have a different bone texture to the others, and that this was potentially a result of them having different origins. These are broadly triangular sections, on each side of the main sternal plate contacting the posterior margin with a roughly M-shaped sternum surrounding them. Something similar is also seen in the juvenile specimen of Altmulopterus (LF 2086P) described here, which somewhat supports Wild’s contention. Under UV light, this shows a clear difference in colour with two dark patches on the sternal plate, one on each side of the base of a cristospine, and these fade out towards the margins of the element and suggest two separate condensation points and reinforce the idea that the lateral margins may be incompletely ossified (Figure 19).
Even so, Wild’s (1993) figure 6b is not an exact match for his own description and so the possible interpretation of this is confusing. The text says that the differently textured and even coloured inserts are of different origin, but both these and the lateral arms of the ‘M’ of the sternum surrounding them are labelled as the respective left and right sternal plates. Thus either the interclavicle is not restricted to the midline and includes these arms (in which case the figure is wrong) or these arms are part of the sternal plates, in which case it is not clear why these should be a different colour and texture to the rest of the plate.
I suggest that the interclavicle was smaller and instead forms the cristospine alone. This would also then be a separate ossification centre and would explain the apparent lack of fusion of the cristospine onto the sternal plate in young pterosaurs. The clavicles would then form the anterior margin of the sternal plate as described by Wild. The plate itself could have two separate ossification centres from its origin as paired elements (as seen in Eudimorphodon and Altmulopterus), and as these expanded they would meet in the midline and cause the presence of a ridge there, while towards the margins the bones would be thin and uneven in shape matching what is generally seen across multiple specimens.
Structure and Function of the Pterosaur Sternum
Despite its clear importance as a muscle attachment site for major forelimb muscles, the sternum has received relatively little attention in terms of both structure and function. The shape of the sternal plate is similar in a number of groups that lived in similar habitats, and may therefore have had similar functional constraints on their flight and evolutionary pressures associated with different windspeeds or flying in cluttered environments (see following section), and it should be expected that the sternum will be influenced on the size and flight style of pterosaurs given its critical importance in the flight apparatus.
Only Bennett (2003) has provided any detailed assessment of the muscles that may have attached to the sternum and their effect. This work was carried out when the origins of pterosaurs were especially uncertain, and although both major competing hypotheses at the time were considered (pterosaurs close to dinosaurs or as early archosauromorphs), Bennett erred on the side of an earlier branch for Pterosauria. Tokita (2015) also comments on this area, but this largely reproduces Bennett’s patterns of muscles and follows his assumptions of a lizard-crocodilian basis for this. We now understand the pterosaurs are likely very close to the Dinosauria (Ezcurra et al., 2020) and as archosaurs, modern birds and crocodylians should therefore be more informative about their musculature than crocodylians and lepidosaurs. Even so, the obviously highly derived nature of the pterosaur flight apparatus means that any inferences from other taxa living or extinct will have to be tempered by the realities of the anatomy at hand. Pterosaurs evolved muscles that would have functioned for take-off and flapping flight that would be substantially different to any non-flying ancestor. At least some attachment scars are present on the sternum indicating insertions, and many muscle groups in Bennett’s (2003) work were present and / or very similar in both dinosaurs and earlier reptiles anyway. In short, while there may be some issues with some details of Bennett’s reconstructions, they are taken here to be broadly correct. A reassessment of pterosaurian musculature is now due given the recent advances in our understanding of their origins and evolution and discovery or more Triassic species, but it is beyond the scope of this paper to reassess the musculature of the sternum let alone the forelimb as a whole.
The size and extent of the muscle attachments on the sternum are unclear in almost all specimens, but a critical question is whether or not the sternal plate alone sufficed to attach the muscles, or if there was some kind of ventral expansion to increase the area. A midline ‘keel’ has been described for some pterosaur sterna (e.g., Pteranodon, Bennett, 2001; Anhanguera, Kellner and Tomida, 2000) and though a number of specimens do show a raised midline ridge across the ventral face of the sternal plate, this is quite unlike those of birds being only a few millimetres high at most, even in large taxa. It has been suggested that this ridge was the basis for the attachment of a cartilaginous keel as Bennett (2003) notes in Anhanguera at least, it lacks any rugosity or surface structure that would normally indicate the attachment of a large block of cartilage, and the absence of even a minimal keel in numerous taxa would also suggest that if present it was not common in pterosaurs. One Pteranodon specimen (YPM 2546) is described as having a flat ‘top’ (i.e., ventral margin of the keel) and being similar to that seen in domestic chickens (Gallus) (Bennett, 2003). This remains a possibility at least for some and would increase the area of the sternal plate for muscle attachments and would further support the idea of extensive cartilage and incomplete ossification of the sternum as a whole. However, a bony keel is clearly absent in all pterosaurs, and the evidence for even an extensive cartilaginous one is limited to Pteranodon alone and even here its presence is far from certain.
Bennett (2003) commented that there are no muscle attachment scars on the sternal plate. Although major muscle groups would have attached here, they would presumably have been spread out across the plate as a whole, and not terminated in a tendon that attached at a point. Several taxa show a striated middle (e.g., Ardeadactylus, indeterminate azhdarchoid - Elgin and Hone, 2013) or posterior part of the plate (e.g., Fenghuangopterus, Batrachognathus), which may indicate areas of attachment, though could also be linked to ossification as with for example in Keresdrakon where the posterior margin is rough but also incomplete (Figure 15B).
Veldmeijer (2006) stated that the cristospine of Coloborhynchus has multiple muscle attachments and in his figure 20, Veldmeijer did illustrate some striations on the anterodorsal part of the cristospine which are likely muscle scars of some form, but he did not label or identify these. Bennett (2001) also noted that the ventral midline of the cristospine is marked by rugose muscle scars that continue up the anterior end in Pteranodon, also indicating the presence of tendinous muscle attachments and something similar is illustrated for Rhamphorhynchus by Wellnhofer (1975, figure 8a). In general, these are few are far between and even well-preserved sterna may show no clear muscle scars or only faint and indeterminate striations (e.g., Auroroazhdarcho, Anhanguera) and by extension it will likely be difficult to correctly determine the exact arrangement of muscles on the sternum given these limitations.
Bennett (2003) illustrated a long m. sternocleidomastoideus attaching from the anteroventral tip of the cristospine which would turn the head. This would at least explain one pair of these muscle scars, although itself this is interesting since clearly the cristopsine and this attachment is not more exaggerated in long necked and large headed pterodactyloids as seen from the small cristospine in ornithocheirids and azhdarchids compared to e.g., Rhamphoprhynchus (see Figure 7 and Figure 12). It might be expected in such animals that much larger muscles would be needed to move the head and require either a longer or broader cristospine to provide a larger area for attachment but clearly this was not the case. This perhaps points to the optimisation of the stiff necks and supporting ligaments in pterodactlyoids that reduced the need for large supporting muscles even as they got larger. Otherwise the only muscles illustrated for pterosaur sterna by Bennett (2003) was a major attachment for a single and massive m. pectoralis across the sternal plate and cristospine, thought he noted that the m. deltoides clavicularis might also originate on the anterior sternum. This might better match the (limited) data here of multiple possible attachment sites on the cristospine, although Witton (2013) also illustrated the m. sternohyoid-sternothyoid complex as attaching to the anterior cristospine, so this is another possibility.
The cristospine might also be longer than is often seen if it is partly cartilaginous. For example, Luchibang has no apparent cristospine, despite a well-ossified sternal plate and the rest of the skeleton being very well preserved (Hone et al., 2020), so this was presumably composed only of cartilage, and this is also the case in Altmulopterus where the sternal plate is well ossified but the cristopsine is apparently absent. In both cases they are young animals, and perhaps the cristopine ossified later and remained short, but it is possible that the apparent short cristopsines of derived pterodactyloids had a cartilaginous anterior extension and were more similar to those of earlier pterosaurs. This would then change the size and position of attachment points of various muscles and alter their performance and mean that muscles like m. pectoralis could be larger than currently reconstructed. In this particular case it would shift some of the pull of the m. pectoralis anteriorly, which would act against the general progression for pterodactyloids to move to posteriorly (Bennett, 2003) and thus would argue against a longer cristospine, or at least an anterior attachment for this muscle on one.
The asymmetrical articulation of the coracoids of many pterosaur sterna are certainly unusual among vertebrates, and, given the importance of bracing the shoulder girdle for a powered flier this is surely more unusual still (though Bennett, 2001 notes that this is present in some birds). Presumably whatever forces were directed through the coracoids during flapping or maneuvoring in the air would have produced asymmetric strain on the sternum and yet there is no clear indication that this is the case through enlargement of one side over the other. What effect this would have had is unclear or why it is distributed the way that it is. Asymmetry is relatively common in pterosaurs (seen in scaphognathids, campylognathoidids and rhamphorhynchines), and although Howse et al. (2001) state that this was unique in Istiodactylus among pterodactyloids it is also present in ctenochasmatids, dsungariptids, tapejarids and azhdarchids at least (see above). Thus, there is at least no immediately obvious connection to this being linked to the overall size or general ecology (marine or terrestrial environments) of the animals in question.
As seen in Nyctosaurus (Hooley, 1913), there may be a posterior projection from the middle of the posterior edge of the sternal plate in some pterosaurs (Figure 10B). Hooley (1913) does not describe it as an articulation but an extension of the sternum, but Jiang et al. (2016) noted that there is an articulation for this as seen in other taxa (though did not name them), and it is not clear to what taxa or to what articulation Jiang and colleagues were referring. A number of pterosaurs do have a sternal plate that had a posterior margin finishing in a point in the midline (e.g., Chanchengopterus, Ardeadactylus), and this may be similar to some of the articulations seen on the lateral margins for the sternocostal ribs. Even so, it is not at all clear if this is a point of articulation on the posterior margin, and in well-preserved and articulated specimens a bony xiphoid process would surely be present at least occasionally. I suggest therefore, that this is a genuine absence in most cases and potentially all. However, as noted above, if part of the sternum is normally unossified it is possible that pterosaurs often had a cartilaginous xiphoid or other posterior extension of the sternum and this has merely ossified in Nyctosaurus. Given the extreme adaptations to flight attributed to this genus and the presence of features such as ossified tendons in the wings (Witton, 2013) this is a plausible explanation for why it might appear in this clade and not others. In Pteranodon the proximal gastralia may fuse to the posterior part of the sternal plate (Bennett, 2001), though how this might relate to a possible xiphoid process is not obvious. None of this however, addresses the issue of what function the xiphoid process might have held, though it would presumably have been an anchor point for various costal muscles and by extension may have had a role in breathing.
 Similarly, the articulation of the sternal or sternocostal ribs with the sternum would have likely helped in respiration with any movement of the chest facilitating the movement of air through the respiratory system (Claessens et al., 2009). However, different models for the articulation of these ribs have been produced by Bennett (2001) and Claessens et al. (2009) (Figure 20). In the former, the anterior dorsal ribs almost touch the sternum and are separate from this by only a tiny dot of bone that is a very short sternal rib, and posteriorly the sternal ribs are angled from anterodorsally to posterioventrally onto the sternum or the gastralia (Figure 20B). In contrast, Claessens et al. (2009) reconstruct the sternal ribs to be orientated from posteriodorsally to meet the sternum anteroventrally and posteriorly these do not reach the gastralia (Figure 20C). This latter orientation gives a deeper chest to pterosaurs (again, potentially changing muscle insertion angles and orientations) and has the potential for greater movement of the sternum during flight. This does match some well-preserved pterosaur specimens (Claessens et al., 2009, figure 2) that show the articulations of the sternal ribs and in particular does show that they (can) meet the raised points on the lateral margins of the sternum, and implies that these are genuine articulation points. As noted above however, these may be asymmetrical between the left and right sides of the sternum and can be inconsistent in number between specimens of a species as well. Similarly, most Rhamphorhynchus sterna do not show articulation points for the ribs so their presence in even a single specimen shows that the expression of this feature is also variable intraspecifically. This suggests that again there may be varying degrees of cartilage or other supporting tissues at play or that these are not necessarily a clear 1:1 ratio of dorsal ribs and sternal ribs to meet the sternum. As noted above, it would appear that this would have to be the case for things like juvenile Rhamphorhynchus with the anterior sternal plate very narrow (Figure 7C) and must have articulated with the sternal ribs in some manner. Notably, the distance and angle between the scapular and coracoid articulation points different between the two models and with a suitably complete and 3D preserved skeleton it may be possible to determine if either is a more appropriate fit for the scapulocoracoids as a test for these competing models (Figure 20B and C).
Similarly, the articulation of the sternal or sternocostal ribs with the sternum would have likely helped in respiration with any movement of the chest facilitating the movement of air through the respiratory system (Claessens et al., 2009). However, different models for the articulation of these ribs have been produced by Bennett (2001) and Claessens et al. (2009) (Figure 20). In the former, the anterior dorsal ribs almost touch the sternum and are separate from this by only a tiny dot of bone that is a very short sternal rib, and posteriorly the sternal ribs are angled from anterodorsally to posterioventrally onto the sternum or the gastralia (Figure 20B). In contrast, Claessens et al. (2009) reconstruct the sternal ribs to be orientated from posteriodorsally to meet the sternum anteroventrally and posteriorly these do not reach the gastralia (Figure 20C). This latter orientation gives a deeper chest to pterosaurs (again, potentially changing muscle insertion angles and orientations) and has the potential for greater movement of the sternum during flight. This does match some well-preserved pterosaur specimens (Claessens et al., 2009, figure 2) that show the articulations of the sternal ribs and in particular does show that they (can) meet the raised points on the lateral margins of the sternum, and implies that these are genuine articulation points. As noted above however, these may be asymmetrical between the left and right sides of the sternum and can be inconsistent in number between specimens of a species as well. Similarly, most Rhamphorhynchus sterna do not show articulation points for the ribs so their presence in even a single specimen shows that the expression of this feature is also variable intraspecifically. This suggests that again there may be varying degrees of cartilage or other supporting tissues at play or that these are not necessarily a clear 1:1 ratio of dorsal ribs and sternal ribs to meet the sternum. As noted above, it would appear that this would have to be the case for things like juvenile Rhamphorhynchus with the anterior sternal plate very narrow (Figure 7C) and must have articulated with the sternal ribs in some manner. Notably, the distance and angle between the scapular and coracoid articulation points different between the two models and with a suitably complete and 3D preserved skeleton it may be possible to determine if either is a more appropriate fit for the scapulocoracoids as a test for these competing models (Figure 20B and C).
Although the data is generally limited, among archosaurs, articulation points are not unique to pterosaurs, and they are also seen in the early branching ornithischian dinosaur Heterodontosaurus (Radermacher et al., 2021). This also shows spatulate sternal ribs with articular ends as described being very similar to those of both Rhamphorhynchus and to modern crocodylians (Radermacher et al., 2021). In short, these are likely similar for archosaurs as a whole and are probably normal for pterosaurs in general.
In addition to these lateral articulations and the possible xiphoid process in pterosaurs, there are a number of taxa that exhibit similar features on the posterior margin of the sternal plate. As with the lateral points above, these vary intraspecifically and can even be asymmetrically presented on single specimens. Given the fusion of the gastralia into the posterior part of the sternum seen in Pteranodon, and orientation of the sterna ribs, it is not clear what might, if anything, articulate with these crenulations on the posterior margin and if nothing does articulate with them, why they are present. This is a small but unsolved issue.
Pterosaur Sternal Evolution
The intrarelationships of the pterosaurs remain somewhat uncertain, and controversial and multiple competing hypotheses exist for their phylogeny. It is even unclear for some taxa to which major clade they belong, and other clades may join or leave different branches of the pterosaur tree in different analyses. As such, some of the assignments of taxa used throughout this paper remain uncertain or controversial, and thus it is also difficult to discuss.
Two taxa are worthy of special comment here given their uncertain origins and positions, which may cloud the comments below. Firstly, Batrachognathus the anurognathid has a sternum which is surprisingly similar in morphology to that of the wukongopterid Kunpengopterus. The anurognathids are of uncertain phylogenetic position and have been recovered in almost every possible place among non-pterodactyloids (see Hone, 2020). Thus despite showing the anurognathids here as a relatively early branching group, they may be much closer to the wukongopterids, and this apparent strong similarity may be the result of shared ancestry rather than convergence. That said, both are also similar to the sternum of Fenghuangopterus and as all three are animals from terrestrial environments, there is perhaps a functional significance to these similarities. Secondly, Santanadactylus brasilensis while here shown as a derived ornithocheirid (following Veldmeijer, 2006) is also suggested to be of uncertain placement within the Pterodactyloidea with the vertebrae described by de Buisonjé (1981) showing features in common with anhanguerids, tapejarids and Pteranodon (Kellner and Tomida, 2000). The latter is the most pertinent since the sternum of Santanadactylus brasilensis shows some striking similarities with that of Pteranodon and other pteranodontids. Both have elongate cristospines with square-shaped sternal plates that have clearly serrated lateral margins.
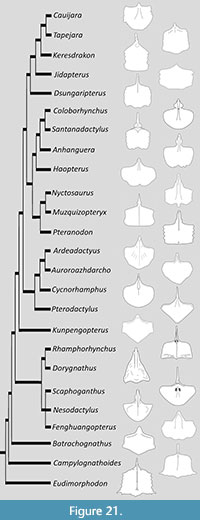 Despite the high degrees of variance in pterosaur sterna described above in various clades, and uncertain phylogenetic positions of some taxa, it is possible to observe some clear clusters of similar forms in some clades and patterns and trends of changes between lineages (Figure 21). Some of the apparent patterns and trends are outlined below.
Despite the high degrees of variance in pterosaur sterna described above in various clades, and uncertain phylogenetic positions of some taxa, it is possible to observe some clear clusters of similar forms in some clades and patterns and trends of changes between lineages (Figure 21). Some of the apparent patterns and trends are outlined below.
Throughout the non-pterodactyloids the cristospine varies in length and may be proportionally short (e.g., Kunpengopterus), mid-sized (e.g., Eudimorphodon, Fenghuangopterus) or long (e.g., Scaphognathus, Rhamphorhynchus). This may also have a broad or narrow attachment to the sternal plate.
The earliest pterosaurs show a sternal plate that is roughly square in adults, but with distinctive expansions in the posterior corners and a clear ridge along the midline of the ventral surface of the sternum, and a serrated posterior margin. The expanded corners are effectively absent after this (there is a slight expansion seen in some, e.g., Rhamphorhynchus, Tapejara, but nothing like to the same extent), and this appears to be restricted to early braches of pterosaur evolution.
Most other non-pterodactyloids have a sternum that is broader than long and with the straight anterior margin to the sternum. However, there is considerable variety in overall shape (e.g., compare the scaphognathids Fenghuangopterus, Nesodactylus and Scaphognathus) and at least some have a posteriorly deep midline expansion to the sternal plate (e.g., Nesodactylus, Kunpengopterus).
Among the pterodactyloids there is a greater range of body sizes, shapes and ecologies represented generally and there is also greater range in sternal morphology. Within the ctenochamatoids, the sternum in generally fan-shaped, though the cristospine may be long and thin or shorter and broader. This morphology is at least partly reflected in the istiodactyliforms which have a broad and fan-shaped sternal plate and might be linked to both groups often occupying coastal environments.
The pteranodontids are generally highly consistent in form with long cristospines and a rectangular sternal plate with serrated lateral margins. Despite the distance in evolutionary history and body size, these are quite similar to those of the earliest pterosaurs, which were also marine animals and may suggest an ecological convergence. The istiodactylids, as noted above, tend to have a more fan-shaped sternum and generally have smooth margins to the sternum with a short cristospine. In the ornithocheirids, the sternum tends to be broader than long with rounded lateral margins. The cristospine is short with broad tubercles at the base and a largemidline ridge from this onto the face of the sternal plate.
The azhdarchoids in general have a relatively square sternum, though notably the tapejarids and azhdarchids have a relatively short cristospine that may be shorter than even that of the ornithocheirds. Other than this, there is some variation in sternal plate shape (e.g., Keresdrakon and Cauijara) though at least some also have a posterior expansion to the plate (e.g., Jidapterus, Cauijara, Tapejara) that is like that seen in other pterosaurs from terrestrial deposits and may represent convergence of ecomorphology.
CONCLUSIONS
The diversity of the pterosaur sterna, both within and between species and clades has been underappreciated to date. This work is only intended to cover and describe the diversity of pterosaur sterna, but it demonstrates that there are various lines of enquiry that, while beyond the scope of this study, should be fruitful and informative in the future into numerous traits present that can be linked to their ancestry, biomechanics and ecology to better understand the life and evolution of the pterosaurs.
The development and ontogeny of the sternum is not well understood. Even the dramatic differences that can be seen between juvenile and adult specimens of Rhamphorhynchus, for example, provide only limited snapshots of what was likely a long and complex (and variable) process of growth and ossification. This is surprising given the importance of the sternum in locomotion and the apparent constancy of other pterosaur elements during growth, and this is an area ripe for further study, though hampered by the limited number of specimens. Similarly, although pterosaur sterna undoubtedly originated from the major sternal elements seen in most reptiles (clavicles, interclavicles, sternal plates) the exact homology of the parts of the pterosaurian sternum are uncertain. The data is limited and unclear and several possibilities exist.
Characters associated with pterosaur sterna have been of only limited use in phylogenetic analyses and should be remedied given the data that is available. Most sterna are poorly preserved and unknown for many taxa, but numerous traits are present and can be scored that would add to our current matrices. Although it is unlikely that well-established pterosaur clades would be rearranged by considering further sternal traits (as for example seen by the inclusion of numerous pelvic characters in work by Hyder et al., 2014) addition of new characters to phylogenetic analyses might well help resolve some problematic and uncertain regions of the tree and provide new topologies that can be further assessed. That said, this must be undertaken with caution given the issues noted above about growth and ossification.
A further assessment of pterosaurian musculature would also be important. Bennett’s (2003, 2008) reconstructions of the forelimb muscles of the pterosaurs Anhanguera and Campylognathoides remain the only detailed work of this kind so far and yet these groups form the basis of pterosaur movements both in flapping in the air and walking on the ground (as well as the transition between the two in take-off). Thus an improved understanding of how the sternum is integrated with the major forelimb muscles is likely to be important for determining the terrestrial and aerial capabilities of different clades.
By extension, this may well be important for better interpreting their ecology. Since the locomotion of flying animals is intimately tied to their foraging styles and environments, determining how different sterna shapes and development patterns are linked to different muscle groups and potentially walking and flight styles will add further to our understanding of the palaeobiology of these animals. Hopefully this work provides a starting point for such research.
ACKNOWLEDGEMENTS
Enormous thanks to S. McDavid without whose superb illustrations this work would be considerably poorer. Thanks to A. Fitch and N. Jagielska discussion of sternal morphology. Thanks to M. Habib for discussions of muscle physiology and ossification. Also thanks to M. Witton for supplying photos and papers, M. McGowen for papers, A. Veldmeijer and E. Endenburg for photos of Tupuxuara, E. Lawlor for photos of Rhamphorhyncus, F. Pinheiro for photos of a tapejarid, N. Jagielska for photos of Dearc, R. Lauer for photographs of Altmulopterus and Scaphognathus, and C. Mehling for generously giving me a rare book on pterosaurs that was very useful. I wish to thank numerous curators who have given me access to specimens over the years which have helped inform this paper: S. Maidment, M. Lamanna, M. Röper, D. Schwartz-Wings, M. Kölbl-Eber, M. Manabe, R. and B. Lauer, O. Rauhut, Z. Fang, and J. Lü. Finally, my thanks to the editor and two anonymous referees for their comments that helped improve this manuscript.
REFERENCES
Andres, B. 2021. Phylogenetic systematics of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea). Journal of Vertebrate Paleontology, 41 (supplement):203-217. https://doi.org/10.1080/02724634.2020.1801703
Andres, B. and Ji, Q. 2008. A new pterosaur from the Liaoning Province of China, the phylogeny of the Pterodactyloidea, and convergence in their cervical vertebrae. Palaeontology, 51:453-469. https://doi.org/10.1111/j.1475-4983.2008.00761.x
Andres, B. and Langston Jr, W. 2021. Morphology and taxonomy of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea). Journal of Vertebrate Paleontology, 41 (supplement):46-202. https://doi.org/10.1080/02724634.2021.1907587
Andres, B., Clark, J., and Xu, X. 2014. The earliest pterodactyloid and the origin of the group. Current Biology, 24:1011-1016. https://doi.org/10.1016/j.cub.2014.03.030
Averianov, A.O. 2010. The osteology of Azhdarcho lancicollis Nessov, 1984 (Pterosauria, Azhdarchidae) from the late Cretaceous of Uzbekistan. Proceedings of the Zoological Institute RAS, 314:264-317.
Baron, M.G. 2020. Testing pterosaur ingroup relationships through broader sampling of avemetatarsalian taxa and characters and a range of phylogenetic analysis techniques. PeerJ, 8:e9604. https://doi.org/10.7717/peerj.9604
Beccari, V., Pinheiro, F.L., Nunes, I., Anelli, L.E., Mateus, O., and Costa, F.R. 2021. Osteology of an exceptionally well-preserved tapejarid skeleton from Brazil: Revealing the anatomy of a curious pterodactyloid clade. PLoS ONE, 16:e0254789. https://doi.org/10.1371/journal.pone.0254789
Bennett, S.C. 1995. A statistical study of Rhamphorhynchus from the Solnhofen Limestone of Germany: year-classes of a single large species. Journal of Paleontology, 69:569-580.
https://doi.org/10.1017/S0022336000034946
Bennett, S.C. 2001. The osteology and functional morphology of the Late Cretaceous pterosaur Pteranodon Part I. General description of osteology. Palaeontographica Abteilung A, 260:1-112.
Bennett, S.C. 2003. Morphological evolution of the pectoral girdle of pterosaurs: myology and function. Geological Society Special Publications, 217:191-215. https://doi.org/10.1144/GSL.SP.2003.217.01.12
Bennett, S.C. 2007. A second specimen of the pterosaur Anurognathus ammoni. Paläontologische Zeitschrift, 81:376-398. https://doi.org/10.1007/BF02990250
Bennett, S.C. 2008. Morphological evolution of the wing of pterosaurs: myology and function. Zitteliana, 28:127-141.
Bennett, S.C. 2013a. The morphology and taxonomy of the pterosaur Cycnorhamphus. Neues Jahrbuch für Geologie und Palaeontologie Abhandlungen, 267:23-41.
https://doi.org/10.1127/0077-7749/2012/0295
Bennett, S.C. 2013b. The phylogenetic position of the Pterosauria within the Archosauromorpha re-examined. Historical Biology, 25:545-563. https://doi.org/10.1080/08912963.2012.725727
Bennett, S.C. 2014. A new specimen of the pterosaur Scaphognathus crassirostris, with comments on constraint of cervical vertebrae number in pterosaurs. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 271:327-348. https://doi.org/10.1127/0077-7749/2014/0392
Bradley, A.B., Burch, S.H., Turner, A.H., Smith, N.D., Irmis, R.B., and Nesbitt, S.J. 2019. Sternal elements of early dinosaurs fill a critical gap in the evolution of the sternum in Avemetatarsalia (Reptilia: Archosauria). Journal of Vertebrate Paleontology, 39:e1700992. https://doi.org/10.1080/02724634.2019.1700992
Cai, Z.-Q. and Wei, F. 1994. [A new pterosaur from the Upper Cretaceous in Linhai, Zhejiang, China]. Vertebrata PalAsiatica, 32:181-194. [in Chinese]
Cau, A., Beyrand, V., Barsbold, R., Tsogtbaatar, K., and Godefroit, P. 2021. Unusual pectoral apparatus in a predatory dinosaur resolves avian wishbone homology. Scientific Reports, 11:14722. https://doi.org/10.1038/s41598-021-94285-3
Chatterjee, S. and Templin, R.J. 2004. Posture, locomotion, and paleoecology of pterosaurs. Geological Society of America, 376:1-64.
Cheng, X., Wang, X., Jiang, S., and Kellner, A.W. 2015. Short note on a non-pterodactyloid pterosaur from Upper Jurassic deposits of Inner Mongolia, China. Historical Biology, 27:749-754. https://doi.org/10.1080/08912963.2014.974038
Claessens, L.P., O’Connor, P.M., and Unwin, D.M. 2009. Respiratory evolution facilitated the origin of pterosaur flight and aerial gigantism. PLoS ONE, 4:e4497. https://doi.org/10.1371/journal.pone.0004497
Colbert, E.H., Brown, B., and Lund, R. 1969. A Jurassic pterosaur from Cuba. American Museum Novitates, 2370:2-26.
Dalla Vecchia, F.M. 2009. Anatomy and systematics of the pterosaur Carniadactylus gen. n. rosenfeldi (Dalla Vecchia, 1995). Rivista Italiana di Paleontologia e Stratigrafia, 115:159-188.
Dalla Vecchia, F.M. 2014. Gli pterosauri triassici. Memorie del Museo Friulano di Storia Naturale, Volume 54. Comune di Udine, Udine, Italy.
Dalla Vecchia, F.M. 2019. Seazzadactylus venieri gen. et sp. nov., a new pterosaur (Diapsida: Pterosauria) from the Upper Triassic (Norian) of northeastern Italy. PeerJ, 7:e7363. https://doi.org/10.7717/peerj.7363
de Buisonjé, P.H. 1981. Santanadactylus brasilensis. Gea, 14:37-48.
Dong, Z.-M. 1982. [A new pterosaur (Huanhepterus quingyangensis gen. et sp. nov.) from Ordos, China]. Vertebrata PalAsiatica, 20:115-121. [in Chinese]
Eck, K., Elgin R.A., and Frey, E. 2011. On the osteology of Tapejara wellnhoferi Kellner 1989 and the first occurrence of a multiple specimen assemblage from the Santana Formation, Araripe Basin, NE Brazil. Swiss Journal of Palaeontology, 130:277-296. https://doi.org/10.1007/s13358-011-0024-5
Elgin, R.A. and Frey, E. 2011. A new azhdarchoid pterosaur from the Cenomanian (Late Cretaceous) of Lebanon. Swiss Journal of Geosciences, 104:21-33. https://doi.org/10.1007/s00015-011-0081-1
Elgin, R.A. and Hone, D.W. 2013. Pneumatization of an immature azhdarchoid pterosaur. Cretaceous Research, 45:16-24. https://doi.org/10.1016/j.cretres.2013.06.006
Ezcurra, M.D., Nesbitt, S.J., Bronzati, M., Dalla Vecchia, F.M., Agnolin, F.L., Benson, R.B., Egli, F.B., Cabreira, S.F., Evers, S.W., Gentil, A.R., and Irmis, R.B. 2020. Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria. Nature, 588:445-449. https://doi.org/10.1038/s41586-020-3011-4
Frey, E., Buchy, M.C., Stinnesbeck, W., Gonzalez, A.G., and Stefano, A. 2006. Muzquizopteryx coahuilensis n. g., n. sp., a nyctosaurid pterosaur with soft tissue preservation from the Coniacian (Late Cretaceous) of northeast Mexico (Coahuila). Oryctos, 6:19-39.
Frey, E., Meyer, C.A., and Tischlinger, H. 2011. The oldest azhdarchoid pterosaur from the Late Jurassic Solnhofen Limestone (early Tithonian) of southern Germany. Swiss Journal of Geosciences, 104:35-55. https://doi.org/10.1007/s00015-011-0073-1
Geist, N.R., Hillenius, W.J., Frey, E., Jones, T.D., and Elgin, R.A., 2014. Breathing in a box: Constraints on lung ventilation in giant pterosaurs. The Anatomical Record, 297:2233-2253. https://doi.org/10.1002/ar.22839
Hone, D.W.E. 2020. A review of the taxonomy and palaeoecology of the Anurognathidae (Reptilia, Pterosauria). Acta Geologica Sinica‐English Edition, 94:1676-1692. https://doi.org/10.1111/1755-6724.14585
Hone, D.W.E. and Benton, M.J. 2007. An evaluation of the phylogenetic relationships of the pterosaurs among archosauromorph reptiles. Journal of Systematic Palaeontology, 5:465-469. https://doi.org/10.1017/S1477201907002064
Hone, D.W.E., Fitch, A.J., Ma, F., and Xu, X., 2020. An unusual new genus of istiodactylid pterosaur from China based on a near complete specimen. Palaeontologia Electronica, 23:a09. https://doi.org/10.26879/1015
Hone, D.W., Ratcliffe, J.M., Riskin, D.K., Hermanson, J.W., and Reisz, R.R. 2021. Unique near isometric ontogeny in the pterosaur Rhamphorhynchus suggests hatchlings could fly. Lethaia, 54:106-112. https://doi.org/10.1111/let.12391
Hone, D.W.E., Tischlinger, H., Frey, E., and Röper, M. 2012. A new non-pterodactyloid pterosaur from the Late Jurassic of Southern Germany. PLoS ONE, 7:e39312. https://doi.org/10.1371/journal.pone.0039312
Hooley, R.W. 1913. On the skeleton of Ornithodesmus latidens; an ornithosaur from the Wealden Shales of Atherfield (Isle of Wight). Quarterly Journal of the Geological Society, 69:372-422.
Howse, S.C.B., Milner, A.R., and Martill, D.M. 2001. Pterosaurs, p. 324-355. In Martill, D.M. and Naish D. (eds.), Dinosaurs of the Isle of Wight. The Palaeontological Association, London, UK.
Hyder, E.S., Witton, M.P., and Martill, D.M. 2014. Evolution of the pterosaur pelvis. Acta Palaeontologica Polonica, 59:109-124. https://doi.org/10.4202/app.2011.1109
Jäger, K.R., Tischlinger, H., Oleschinski, G., and Sander, P.M. 2018. Goldfuß was right: Soft part preservation in the Late Jurassic pterosaur Scaphognathus crassirostris revealed by reflectance transformation imaging and ultraviolet light and the auspicious beginnings of paleo-art. Palaeontologia Electronica, 21(3.3T):1-20. https://doi.org/10.26879/713
Jagielska, N., O’Sullivan, M., Funston, G.F., Butler, I.B., Challands, T.J., Clark, N.D., Fraser, N.C., Penny, A., Ross, D.A., Wilkinson, M., and Brusatte, S.L. 2022. A skeleton from the Middle Jurassic of Scotland illuminates an earlier origin of large pterosaurs. Current Biology, 32(6):1446-1453. https://doi.org/10.1016/j.cub.2022.01.073
Jiang, S., Cheng, X., Ma, Y., and Wang, X. 2016. A new archaeopterodactyloid pterosaur from the Jiufotang Formation of western Liaoning, China, with a comparison of sterna in Pterodactylomorpha. Journal of Vertebrate Paleontology, 36(6):e1212058. https://doi.org/10.1080/02724634.2016.1212058
Kellner, A.W.A. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. Geological Society Special Publications, 217:105-137.
Kellner, A.W.A. 2013. A new unusual tapejarid (Pterosauria, Pterodactyloidea) from the Early Cretaceous Romualdo Formation, Araripe Basin, Brazil. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 103:409-421. https://doi.org/10.1017/S1755691013000327
Kellner, A.W.A. 2015. Comments on Triassic pterosaurs with discussion about ontogeny and description of new taxa. Anais da Academia Brasileira de Ciências, 87:667-689.
https://doi.org/10.1590/0001-3765201520150307
Kellner, A.W.A. and Y. Tomida. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs, 17:1-137.
Kellner, A.W., Campos, D.A., Sayao, J.M., Saraiva, A.A., Rodrigues, T., Oliveira, G., Cruz, L.A., Costa, F.R., Silva, H.P., and Ferreira, J.S. 2013. The largest flying reptile from Gondwana: a new specimen of Tropeognathus cf. T. mesembrinus Wellnhofer, 1987 (Pterodactyloidea, Anhangueridae) and other large pterosaurs from the Romualdo Formation, Lower Cretaceous, Brazil. Anais da Academia Brasileira de Ciências, 85:113-135.
https://doi.org/10.1590/S0001-37652013000100009
Kellner, A.W., Caldwell, M.W., Holgado, B., Dalla Vecchia, F.M., Nohra, R., Sayão, J.M., and Currie, P.J. 2019a. First complete pterosaur from the Afro-Arabian continent: insight into pterodactyloid diversity. Scientific Reports, 9:17875. https://doi.org/10.1038/s41598-019-54042-z
Kellner, A.W., Weinschütz, L.C., Holgado, B., Bantim, R.A., and Sayao, J.M. 2019b. A new toothless pterosaur (Pterodactyloidea) from Southern Brazil with insights into the paleoecology of a Cretaceous desert. Anais da Academia Brasileira de Ciências, 91:e20190768. https://doi.org/10.1590/0001-3765201920190768
Lü, J. 2010. A new boreopterid pterodactyloid pterosaur from the Early Cretaceous Yixian Formation of Liaoning Province, northeastern China. Acta Geologica Sinica‐English Edition, 84:241-246. https://doi.org/10.1111/j.1755-6724.2010.00204.x
Lü, J. and Yuan, C. 2005. New tapejarid pterosaur from western Liaoning, China. Acta Geologica Sinica‐English Edition, 79:453-458. https://doi.org/10.1111/j.1755-6724.2005.tb00911.x
Lü, J.C. and Zhang, B.K. 2005. New pterodactyloid pterosaur from the Yixian Formation of western Liaoning. Geological Review, 51:458-462.
Lü, J.C., Gao, C.L., Liu, J.Y., Meng, Q.J., and Ji, Q. 2006b. New material of the pterosaur Eopteranodon from the Early Cretaceous Yixian Formation, western Liaoning, China. Geological Bulletin of China, 25:565-571.
Lü, J., Liu, J., Wang, X., Gao, C., Meng, Q., and Ji, Q. 2006a. New material of pterosaur Sinopterus (Reptilia: Pterosauria) from the Early Cretaceous Jiufotang Formation, Western Liaoning, China. Acta Geologica Sinica-English Edition, 80:783-789. https://doi.org/10.1111/j.1755-6724.2006.tb00302.x
Lü, J.-C., Fucha, X.H., and Chen, J.M. 2010a. A new scaphognathine pterosaur from the Middle Jurassic of western Liaoning, China. Diqiu Xuebao (Acta Geoscientica Sinica), 31:263-266.
Lü, J.-C., Unwin, D.M., Jin, X.-S., Liu, Y.-Q., and Ji, Q. 2010b. Evidence for modular evolution in a long-tailed pterosaur with a pterodactyloid skull. Proceedings of the Royal Society series B, Biological Sciences, 277:383-389. https://doi.org/10.1098/rspb.2009.1603
Lü, J., Unwin, D.M., Deeming, D.C., Jin, X., Liu, Y., and Ji, Q. 2011a. An egg-adult association, gender, and reproduction in pterosaurs. Science, 331:321-324. https://doi.org/10.1126/science.1197323
Lü, J., Xu, L., Chang, H., and Zhang, X. 2011b. A new darwinopterid pterosaur from the Middle Jurassic of western Liaoning, northeastern China and its ecological implications. Acta Geologica Sinica‐English Edition, 85:507-514.
https://doi.org/10.1111/j.1755-6724.2011.00444.x
Lü, J., Pu, H., Xu, L., Wei, X., Chang, H., and Kundrát, M. 2015. A new rhamphorhynchid pterosaur (Pterosauria) from Jurassic deposits of Liaoning Province, China. Zootaxa, 3911:119-129. https://doi.org/10.11646/zootaxa.3911.1.7
Manzig, P.C., Kellner, A.W., Weinschütz, L.C., Fragoso, C.E., Vega, C.S., Guimarães, G.B., Godoy, L.C., Liccardo, A., Ricetti, J.H., and de Moura, C.C. 2014. Discovery of a rare pterosaur bone bed in a Cretaceous desert with insights on ontogeny and behavior of flying reptiles. PLoS ONE, 9:e100005. https://doi.org/10.1371/journal.pone.0100005
Martill, D.M., Frey, E., Tischlinger, H., Mäuser, M., Rivera-Sylva, H.E., and Vidovic, S.U. 2023. A new pterodactyloid pterosaur with a unique filter-feeding apparatus from the Late Jurassic of Germany. PalZ, 1-42. https://doi.org/10.1007/s12542-022-00644-4
McGowen, M.R., Padian, K., De Sosa, M.A., and Harmon, R.J. 2002. Description of Montanazhdarcho minor, an azhdarchid pterosaur from the Two Medicine Formation (Campanian) of Montana. PaleoBios, 22:1-9.
Meyer, H.V. 1854. Mittheilungen an Professor Bronn. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde, 1854:47-58.
Naish, D., Witton, M.P., and Martin-Silverstone, E. 2021. Powered flight in hatchling pterosaurs: evidence from wing form and bone strength. Scientific Reports, 11:13130. https://doi.org/10.1038/s41598-021-92499-z
Padian, K. 2008a. The Early Jurassic pterosaur Dorygnathus banthensi s (Theodori, 1830). Special Papers in Palaeontology, 80:1-64.
Padian, K. 2008b. The Early Jurassic pterosaur Campylognathoides Strand, 1928. Special Papers in Palaeontology, 80:65-107.
Padian, K., De Ricqles, A.J., and Horner, J.R. 1995. Bone histology determines identification of a new fossil taxon of pterosaur (Reptilia: Archosauria). Comptes Rendus - Academie des Sciences Paris, serie 2 Sciences de la Terre et des Planetes fascicule A, 320:77-77.
Padian, K., Cunningham, J.R., Langston Jr., W., and Conway, J. 2021. Functional morphology of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea). Journal of Vertebrate Paleontology, 41 (supplement):218-251. https://doi.org/10.1080/02724634.2020.1780247
Pereda-Superbiola, X., Knoll, F., Ruiz-Omeñaca, J.I., Company, J., and Torcida Fernández‐Baldor, F. 2012. Reassessment of Prejanopterus curvirostris, a basal pterodactyloid pterosaur from the Early Cretaceous of Spain. Acta Geologica Sinica‐English Edition, 86:1389-1401.
Radermacher, V.J., Fernandez, V., Schachner, E.R., Butler, R.J., Bordy, E.M., Hudgins, M.N., de Klerk, W.J., Chapelle, K.E., and Choiniere, J.N. 2021. A new Heterodontosaurus specimen elucidates the unique ventilatory macroevolution of ornithischian dinosaurs. eLife, 10:e66036. https://doi.org/10.7554/eLife.66036
Renesto, S. 1993. An isolated sternum of Eudimorphodon (Reptilia, Pterosauria) from the Norian (Late Triassic) of the Bergamo Prealps (Lombardy, northern Italy). Rivista Italiana di Paleontologia e Stratigrafia, 99:415-422. https://doi.org/10.13130/2039-4942/8904
Ryabinin, A.N. 1948. [Remarks on a flying reptile from the Jurassic of the Karatau.] Transactions of the Palaeontological Institute, 15:86-93. [in Russian]
Tischlinger, H. and Frey, E. 2013. A new pterosaur with mosaic characters of basal and pterodactyloid Pterosauria from the Upper Kimmeridgian of Painten (Upper Palatinate, Germany). Archaeopteryx, 31:1-13.
Tokita, M. 2015. How the pterosaur got its wings. Biological Reviews, 90:1163-1178.
https://doi.org/10.1111/brv.12150
Unwin, D.M. 2003. On the phylogeny and evolutionary history of pterosaurs. Evolution and Palaeobiology of Pterosaurs. Geological Society Special Publications, 217:139-190.
https://doi.org/10.1144/GSL.SP.2003.217.01.11
Veldmeijer, A.J. 2006. Toothed pterosaurs from the Santana Formation (Cretaceous; Aptian-Albian) of northeastern Brazil. A reappraisal on the basis of newly described material. Unpublished PhD thesis, Utrecht University, Utrecht, the Netherlands.
Vidovic, S.U. and Martill, D.M. 2018. The taxonomy and phylogeny of Diopecephalus kochi (Wagner, 1837) and ‘Germanodactylus rhamphastinus ’ (Wagner, 1851). Geological Society, London, Special Publications, 455:125-147. https://doi.org/10.1144/SP455.12
Wang, X. and Lü, J. 2001. Discovery of a pterodactylid pterosaur from the Yixian Formation of western Liaoning, China. Chinese Science Bulletin, 46:A3-A8. https://doi.org/10.1007/BF02900690
Wang, X. and Zhou, Z. 2002. A new pterosaur (Pterodactyloidea, Tapejaridae) from the Early Cretaceous Jiufotang Formation of western Liaoning, China and its implications for biostratigraphy. Chinese Science Bulletin, 48:16-23. https://doi.org/10.1007/BF03183326
Wang, X., Kellner, A.W., Zhou, Z., and de Almeida Campos, D. 2005. Pterosaur diversity and faunal turnover in Cretaceous terrestrial ecosystems in China. Nature, 437:875-879. https://doi.org/10.1038/nature03982
Wang, X., Kellner, A.W., Zhou, Z., and de Almeida Campos, D. 2007. A new pterosaur (Ctenochasmatidae, Archaeopterodactyloidea) from the lower Cretaceous Yixian Formation of China. Cretaceous Research, 28:245-260. https://doi.org/10.1016/j.cretres.2006.08.004
Wang, X., Kellner, A.W., Jiang, S., and Meng, X. 2009. An unusual long-tailed pterosaur with elongated neck from western Liaoning of China. Anais da Academia Brasileira de Ciências, 81:793-812. https://doi.org/10.1590/S0001-37652009000400016
Wang, X., Kellner, A.W., Jiang, S., Cheng, X., Meng, X., and Rodrigues, T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. Anais da Academia Brasileira de Ciências, 82:1045-1062. https://doi.org/10.1590/S0001-37652010000400024
Wang, X., Kellner, A.W., Jiang, S., Wang, Q., Ma, Y., Paidoula, Y., Cheng, X., Rodrigues, T., Meng, X., Zhang, J., and Li, N. 2014. Sexually dimorphic tridimensionally preserved pterosaurs and their eggs from China. Current Biology, 24:1323-1330. https://doi.org/10.1016/j.cub.2014.04.054
Wang, X., Jiang, S., Zhang, J., Cheng, X., Yu, X., Li, Y., Wei, G., and Wang, X. 2017. New evidence from China for the nature of the pterosaur evolutionary transition. Scientific Reports, 7:42763. https://doi.org/10.1038/srep42763
Wellnhofer, P. 1970. Die Pterodactyloidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. Bayerische Akademie der Wissenschaften, Mathematisch-Wissenschaftlichen Klasse, Abhandlugen, 141:1-133.
Wellnhofer, P. 1975. Die Rhamphorhynchoidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. Palaeontographica, Abteilung A, 148:1-13.
Wellnhofer, P. 1978. Pterosauria. Handbuch der Paläoherpetologie, Teil 19. Gustav Fischer Verlag, Stuttgart, Germany.
Wellnhofer, P. 2003. A Late Triassic pterosaur from the Northern Calcareous Alps (Tyrol, Austria). Geological Society, London, Special Publications, 217:5-22. https://doi.org/10.1144/GSL.SP.2003.217.01.02
Wild, R. 1978. Die Flugsaurier (Reptilia, Pterosauria) aus der Oberen Trias von Cene bei Bergamo, Italien, 17:176-256.
Wild, R. 1993. A juvenile specimen of Eudimorphodon ranzii Zambelli (Reptilia, Pterosauria) from the upper Triassic (Norian) of Bergamo. Rivista del Museo Civico di Scienze Naturali, 16:95-120.
Williston, S.W. 1893. Kansas pterodactyls. Part II. Kansas University Quarterly, 2:79-81.
Williston, S.W. 1897. Restoration of Ornithostoma (Pteranodon). Kansas University Quarterly, 6:35-51.
Williston, S.W. 1902. On the skeleton of Nyctodactylus, with restoration. American Journal of Anatomy, 1:297-305.
Wiman, C. 1925. Über Dorygnathus und andere Flugsaurier. Bulletin Geological Institute Uppsala, 19:23-54.
Witton, M.P. 2013. Pterosaurs. Princeton University Press, Los Angeles, USA.
https://doi.org/10.1515/9781400847655
Wu, W.H., Zhou, C.F., and Andres, B. 2017. The toothless pterosaur Jidapterus edentus (Pterodactyloidea: Azhdarchoidea) from the Early Cretaceous Jehol Biota and its paleoecological implications. PLoS ONE, 12:e0185486. https://doi.org/10.1371/journal.pone.0185486
Young, C.-C. 1973. [Pterosaurs from Wuerho]. Reports of Paleontological Expedition to Sinkiang (II). Pterosaurian Fauna from Wuerho, Sinkiang. Memoirs of the Institute of Vertebrate Palaeontology and Paleoanthropology. Academica Sinica, 11:18-35. [in Chinese]
Zhang, X., Jiang, S., Cheng, X., and Wang, X. 2019. New material of Sinopterus (Pterosauria, Tapejaridae) from the Early Cretaceous Jehol Biota of China. Anais da Academia Brasileira de Ciências, 91 (supplement 2). https://doi.org/10.1590/0001-376520192018756
Zhou, C.F. and Schoch, R.R. 2011. New material of the non-pterodactyloid pterosaur Changchengopterus pani Lü, 2009 from the Late Jurassic Tiaojishan Formation of western Liaoning. Neues Jahrbuch fur Geologie und Palaontologie Abhandlungen, 260(3):265-275. https://doi.org/10.1127/0077-7749/2011/0131
Zhou, X., Pêgas, R.V., Ma, W., Han, G., Jin, X., Leal, M.E., Bonde, N., Kobayashi, Y., Lautenschlager, S., Wei, X., and Shen, C. 2021. A new darwinopteran pterosaur reveals arborealism and an opposed thumb. Current Biology, 31:2429-2436.
https://doi.org/10.1016/j.cub.2021.03.030
