

 On the systematics and phylogenetic nomenclature of the Ornithocheiriformes (Pterosauria, Pteranodontoidea)
On the systematics and phylogenetic nomenclature of the Ornithocheiriformes (Pterosauria, Pteranodontoidea)
Article number: 28.2.a25
https://doi.org/10.26879/20
Copyright Paleontological Society, June 2025
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 7 October 2024. Acceptance: 7 June 2025
ABSTRACT
Ornithocheiriformes (Pterosauria, Pterodactyloidea, Lanceodontia) is the clade comprising all species closer to Ornithocheirus simus than Istiodactylus latidens. This group includes, in a broad sense, all forms usually regarded as close relatives of Ornithocheirus simus, Coloborhynchus clavirostris, and/or Anhanguera blittersdorffi. This group suffers from complex and controversial nomenclatorial problems since the early days of pterosaur science. The recent adherence to the PhyloCode in pterosaur nomenclatorial works provides a basis for a solid standardization of clade names and definitions, even though their stability may sometimes be sensitive to differing phylogenetic hypotheses. The present work aims at integrating/conciliating the convoluted nomenclature of Ornithocheiriformes into both the International Code of Zoological Nomenclature (ICZN) and the International Code of Phylogenetic Nomenclature (PhyloCode), to prevent equivalent clades from having different names under each Code, especially when registered definitions are applied to distinct phylogenetic hypotheses.
R.V. Pêgas. Laboratório de Paleontologia, Museu de Zoologia da Universidade de São Paulo, Av. Nazaré 481, Ipiranga, São Paulo, 04263-000, Brazil. rvpegas.paleo@gmail.com
Keywords: phylogenetic nomenclature; systematics; phylogeny; Pterosauria
Final citation: Pêgas, R.V. 2025. On the systematics and phylogenetic nomenclature of the Ornithocheiriformes (Pterosauria, Pteranodontoidea). Palaeontologia Electronica, 28(2):a25.
https://doi.org/10.26879/20
palaeo-electronica.org/content/2025/5546-systematics-of-ornithocheiriform-pterosaurs
Copyright: June 2025 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
The pterosaurian group Ornithocheiriformes was established as an unranked taxon by Andres (2021) and defined as the largest clade comprising Ornithocheirus simus but not Istiodactylus latidens. These forms are mainly characterized by their elongate jaws bearing numerous elongate, striated teeth (Andres, 2021), apparently well-suited for piscivory (Wellnhofer, 1991; Pêgas et al., 2021; Souza et al., 2023). So far, the group comprises over 30 named species (Andres, 2021; Pêgas, 2024; Pentland et al., 2024). This speciose clade ranges from the Berriasian (Howse and Milner, 1995) to the Turonian (Pentland et al., 2019) and occurs in all continents except Antarctica (Holgado et al., 2019).
Unfortunately, ornithocheiriform systematics suffer from nomenclatural problems that have long needed special attention. For the last 30 years, there have been mainly two parallel ornithocheirifom taxonomic schemes, as follows. Authors who regard Anhanguera blittersdorffi as closer to Tropeognathus mesembrinus than Ornithocheirus simus have used a “restrictive Anhangueridae” that is distinct from Ornithocheiridae (e.g., Kellner, 1989; 2003; Wang and Zhou, 2003; 2012; Rodrigues and Kellner, 2013; Bantim et al., 2014; Aureliano et al., 2014; Pinheiro and Rodrigues, 2017; Holgado et al., 2019; Pêgas et al., 2019; Holgado and Pêgas, 2020; Richards et al., 2021; Jacobs et al., 2024; Silva et al., 2024), while authors who regard that Tropeognathus mesembrinus is closely related to Ornithocheirus simus have used a broad Ornithocheiridae, which also necessarily includes Anhanguera blittersdorffi (and anhanguerids overall) and overwrites the Anhangueridae (e.g., Unwin, 2003; Steel et al., 2005; Elgin and Frey, 2010; Vidovic et al., 2017). These are termed, here, the (1) “restrictive  Anhangueridae” (Figure 1A) and (2) “broad Ornithocheiridae” (Figure 1B) taxonomic schemes. The very foundations of each scheme necessarily lie, respectively, (1) in the exclusion of Ornithocheirus simus from Anhangueridae (which typically includes Tropeognathus mesembrinus, Coloborhynchus clavirostris and Anhanguera blittersdorffi) for the former scheme, and (2) in the inclusion of Anhanguera blittersdorffi in Ornithocheiridae (also including Tropeognathus mesembrinus and Coloborhynchus clavirostris) for the latter. With consistent usage, these two schemes have become well established, stable, and mutually exclusive in the literature.
Anhangueridae” (Figure 1A) and (2) “broad Ornithocheiridae” (Figure 1B) taxonomic schemes. The very foundations of each scheme necessarily lie, respectively, (1) in the exclusion of Ornithocheirus simus from Anhangueridae (which typically includes Tropeognathus mesembrinus, Coloborhynchus clavirostris and Anhanguera blittersdorffi) for the former scheme, and (2) in the inclusion of Anhanguera blittersdorffi in Ornithocheiridae (also including Tropeognathus mesembrinus and Coloborhynchus clavirostris) for the latter. With consistent usage, these two schemes have become well established, stable, and mutually exclusive in the literature.
A decade ago, a redefinition for the Anhangueridae was proposed, as the most inclusive clade containing Anhanguera blitersdorffi but not Ornithocheirus simus (Andres et al., 2014). Similarly, the Ornithocheiridae were redefined as the most inclusive clade containing Ornithocheirus simus but not Anhanguera blitersdorffi (Andres et al., 2014). These definitions have recently been published in accordance with the PhyloCode (Andres, 2021). These redefinitions were proposed under the light of a new phylogenetic hypothesis, herein termed the “restrictive Ornithocheirae” hypothesis (Figure 1C), wherein Anhanguera blittersdorffi and Ornithocheirus simus are closer to each other than to most other members of Ornithocheiriformes; see Andres (2021).
Interestingly, this taxonomic scheme (Andres, 2021) permits the coexistence of both names (Ornithocheiridae and Anhangueridae) under any phylogenetic hypothesis. While these redefinitions work well under the “restrictive Ornithocheirae” hypothesis, they abruptly discontinue the traditional concept of both clade names under any other phylogenetic hypothesis (e.g., Codorniú et al., 2016; Holgado et al., 2019), thereby severely disrupting nomenclatural stability. This fails Article 10.1 of the PhyloCode, which states that “ clade names are generally to be selected in such a way as to minimize disruption of current and/or historical usage (with regard to composition, diagnostic characters, or both) and to maximize continuity with existing literature ” (De Queiroz and Cantino, 2020). Further complicatedly, if applied to alternative phylogenies (e.g. Pêgas, 2024), the registered definition of Anhangueridae (Andres, 2021) would implicate in an overly broad group - by further including Cimoliopteridae, Targaryendraconidae, and Hamipteridae (thus creating an undesirable “broad Anhangueridae”, as well as an undesirable conflict with traditional nomenclature).
Given the issues regarding ornithocheiriform nomenclature, the present work provides a critical revision of ornithocheiriform pterosaur clade names and definitions. The recent implementation of the International Code of Phylogenetic Nomenclature (PhyloCode) (de Queiroz and Cantino, 2020) provides an opportunity to evaluate the utility of previously proposed phylogenetic definitions for established taxon names. All established ornithocheiriform clade names are revised here, aiming at formalizing and standardizing ornithocheiriform nomenclature following the guidelines of the PhyloCode. At the same time, care is taken to preserve consistency with traditional nomenclature based on the International Code of Zoological Nomenclature (ICZN), so as to prevent equivalent taxa from having different names under each Code. Special attention is given to Anhangueridae, with an unrestricted emendation for this clade being presented, along with a new clade name (Anhangueroidea clade nov.), aiming to refer to an apomorphy-based clade that effectively encompasses ornithocheiriforms closer to Anhanguera blittersdorffi than to Ornithocheirus simus (as an alternative to an undesirable “broad Anhangueridae”).
MATERIALS AND METHODS
Phylogenetic Nomenclature
The present work follows the PhyloCode (de Queiroz et al., 2020) as a means of standardizing and stabilizing phylogenetic nomenclature. As such, new clade names, converted clade names, and new unrestricted emendations are presented here following the requirements for establishment as regulated by the PhyloCode (de Queiroz et al., 2020), including presentation of protologues and registration in RegNum, the PhyloCode’s digital database.
Following the guidelines of the PhyloCode, ideal phylogenetic definitions must take into consideration the preservation of traditional usage (Article 10.1). For this purpose, stability across different phylogenetic frameworks is needed, otherwise traditional usage may be lost depending on the preferred framework, requiring the need of unrestricted emendations (Article 15.8 of the PhyloCode). Such stability is best achieved when definitions are carefully constructed to stably refer to equivalent clades across alternative phylogenetic hypotheses (when equivalent clades do exist under different hypotheses; e.g. Note 15.11.1 of the PhyloCode), as well as to prevent name application when intended equivalent clades do not exist in alternative phylogenetic hypotheses (e.g., Articles 11.12 and 11.13 of the PhyloCode). For example, see the works of Madzia et al. (2021), Young et al. (2024), and Leardi et al. (2024).
Phylogenetic Analysis Procedures
The present work is accompanied by a phylogenetic analysis based on the data matrix of Pêgas (2024), which comprises the most comprehensive pterosaur dataset available in the literature so far. The dataset is freely available at the MorphoBank online repository, under project identifier #5720. The analysis follows the same protocol as described by Pêgas (2024), as follows. The analysis was conducted under maximum parsimony, utilizing the software TNT 1.6 (Goloboff and Morales, 2023), and was divided into two steps. New Technology Search was used for the first step (using Sectorial Search, Ratchet, Drift and Tree fusing, default parameters, and random seed = 0). Afterwards, with tree memory space set to maximum, a Traditional Search was performed using trees from RAM (using TBR, collapsing trees after search).
The dataset, based on Pêgas (2024), has been corrected following the observations of Hone et al. (2024) regarding pseudoreplicated data (which were accordingly deleted/corrected) and unnamed states (which were accordingly named). The new characters from Hone et al. (2024), Zhou et al. (2025), and Manitkoon et al. (2025) have been included as well, and further 12 new characters are presented. In total, the dataset comprises 563 characters and 202 in-group operational taxonomic units, representing the most comprehensive phylogenetic dataset for pterosaurs thus far.
For further ornithocheiriform representation, the following taxa were added here: Brasileodactylus araripensis (Kellner, 1984; Veldmeijer et al., 2009), ‘Ornithocheirus’ colorhinus (Rodrigues and Kellner, 2013), the unnamed Kem Kem ornithocheirid (Jacobs et al., 2020), Haliskia peterseni (Pentland et al., 2024), and Akharhynchus martilli (Jacobs et al., 2024). Taxonomy of the genus Anhanguera follows Pinheiro and Rodrigues (2017). Coding for Thapuungaka shawi is herein updated with the inclusion of the referred specimen KK F600 within its hypodigm (Richards et al., 2023). The taxon Saratovia glickmani, newly named and attributed to Targaryendraconia (Averianov, 2025), was unfortunately described too recently to be included in the present analysis.
The species Cearadactylus atrox was excluded, being herein regarded as a junior synonym of Brasileodactylus araripensis (Pêgas and Costa, 2022). Mythunga camara (known only from a fragmentary, mid-section of upper and lower jaws) is also excluded and regarded as a nomen dubium, as its revised diagnosis (Pentland and Poropat, 2019) comprises features that are unfortunately more widespread than previously thought, as follows. Its revised diagnosis comprised a combination of features, which included two proposed autapomorphies (marked with an asterisk): transversely compressed skull; undulating lateral margins of jaw*; three maxillary teeth between last enlarged tooth and nasoantorbital fenestra*; splenial with lingual nutrient foramina; alveolar spacing decreasing posteriorly; and robust, conical, lingually curved and striated teeth. However, undulating lateral margins of the jaw were later shown to be more widespread among anhanguerids, being especially pronounced in some tropeognathines (Pentland et al., 2019, 2022; Holgado and Pêgas, 2020); while three maxillary teeth between the last enlarged tooth and nasoantorbital fenestra are equally present in Tropeognathus mesembrinus (Wellnhofer, 1987; Holgado and Pêgas, 2020), and unclear in other tropeognathines (Pentland et al., 2019, 2022, 2024). The proposed autapomorphies of Mythunga camara can no longer be seen as valid. Furthermore, the combination of a transversely compressed skull; splenial with lingual nutrient foramina; and conical, lingually curved, and striated teeth, describe anhanguerids in general (Wellnhofer, 1985; Kellner and Tomida, 2000); while an alveolar spacing decreasing posteriorly and robust teeth seem general for tropeognathines (Wellnhofer, 1987; Holgado and Pêgas, 2020; Pentland et al., 2022). Therefore, the preserved remains of Mythunga camara do not exhibit diagnostic features that could distinguish it, morphologically, from other tropeognathine anhanguerids, and its holotype should be viewed as an indeterminate tropeognathine.
Institutional Abbreviations
BSPG, Bayerische Staatssammlung für Paläontologie und historische Geologie, Munich, Germany; CAMSM, Sedgwick Museum of Earth Sciences, Cambridge, England, Cambridge, England; FSAC, Faculté des Sciences Aïn-Chock, Casablanca, Morocco; MPPCN, Museu de Paleontologia Plácido Cidade Nuvens, Santana do Cariri, Brazil; MHNS, Museu de História Natural de Sintra, Sintra, Portugal; MN, Museu Nacional/Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil; NHMUK, Natural History Museum, London, England; NSM, Natural Sciences Museum, Tokyo, Japan; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China; IWCMS, Isle of Wight County Museum Service, Brighstone, UK; SMNK, Staatliches Museum für Naturkunde Karlsruhe, Karlsruhe, Germany; SMNS, Staatliches Museum für Naturkunde Stuttgart, Stuttgart, Germany; SMU, Shuler Museum of Palaeontology, Southern Methodist University, Dallas, USA; Uop, University of Portsmouth.
RESULTS AND DISCUSSION
Phylogenetic Analysis Results
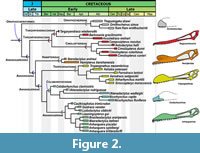
The analysis produced 108 most parsimonious trees, with 2268 steps, ensemble consistency index of 0.345, and ensemble retention index 0.795. The strict consensus tree, focusing on the Ornithocheiriformes, is shown in Figure 2. The remaining portion of the tree is the same as in Zhou et al. (2025). Character/states numbers of the synapomorphies that support each ornithocheiriform clade are given in the Appendix.
SYSTEMATIC PALEONTOLOGY
PTEROSAURIA Owen, 1842
PTERODACTYLOIDEA Plieninger, 1901
PTERANODONTOIDEA Kellner, 2003
ORNITHOCHEIRIFORMES Andres, 2021
ANHANGUEROIDEA new clade name
Registration number. 1057.
Definition. The clade characterized by a reflected palatal tip synapomorphic with that of Anhanguera blittersdorffi Campos and Kellner, 1985; provided that this clade does not include Ornithocheirus simus Owen, 1861. This is an apomorphy-based definition with a qualifying clause.
Reference phylogeny. Present work (Figure 2).
Composition. Anhangueria and Targaryendraconia.
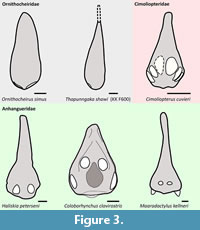 Diagnostic apomorphies. Reflected palatal tip, with first upper alveolus facing markedly anteroventrally (Figure 3, Figure 4); lateral offset of the third alveolus (both jaws) (Figure 5); presence of a palatal ridge; and presence of an individualized mandibular groove.
Diagnostic apomorphies. Reflected palatal tip, with first upper alveolus facing markedly anteroventrally (Figure 3, Figure 4); lateral offset of the third alveolus (both jaws) (Figure 5); presence of a palatal ridge; and presence of an individualized mandibular groove.
 Remarks. This clade is herein intended to comprise a broad clade that includes Anhanguera blittersdorffi and excludes Ornithocheirus simus, by joining all taxa that, similar to the former but unlike the latter, exhibit a reflected palatal tip. This important character, which sets Ornithocheirus simus apart from Coloborhynchus clavirostris, Tropeognathus mesembrinus, and their close relatives (Figure 3, Figure 4), has been left aside in some recent large-scale phylogenetic analyses (Andres et al., 2014; Longrich et al., 2018; Andres, 2021), even though the work of Rodrigues and Kellner (2013) explored this in detail over 10 years ago.
Remarks. This clade is herein intended to comprise a broad clade that includes Anhanguera blittersdorffi and excludes Ornithocheirus simus, by joining all taxa that, similar to the former but unlike the latter, exhibit a reflected palatal tip. This important character, which sets Ornithocheirus simus apart from Coloborhynchus clavirostris, Tropeognathus mesembrinus, and their close relatives (Figure 3, Figure 4), has been left aside in some recent large-scale phylogenetic analyses (Andres et al., 2014; Longrich et al., 2018; Andres, 2021), even though the work of Rodrigues and Kellner (2013) explored this in detail over 10 years ago.
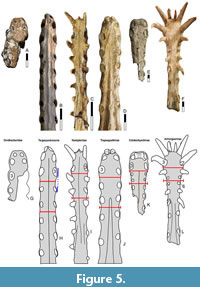 It is important to note that, while all anhangueroids (sensu the present work) exhibit a lateral offset of the third alveolus, this does not necessarily imply in the existence of a proper lateral expansion of the jaw tip as termed here (Figure 5), which is restricted to the Anhangueria (see further below).
It is important to note that, while all anhangueroids (sensu the present work) exhibit a lateral offset of the third alveolus, this does not necessarily imply in the existence of a proper lateral expansion of the jaw tip as termed here (Figure 5), which is restricted to the Anhangueria (see further below).
TARGARYENDRACONIA Pêgas et al. (2019),
converted clade name
Registration number. 1058.
Definition. The most inclusive clade that includes Targaryendraco wiedenrothi (Wild, 1990) but not Anhanguera blittersdorffi Campos and Kellner, 1985, Hamipterus tianshanensis Wang et al., 2014, or Ornithocheirus simus Owen, 1861. This is a branch-based definition.
Reference phylogeny. Present work (Figure 2).
Composition. Cimoliopteridae and Targaryendraconidae.
Diagnostic apomorphies. Jaw anterior ends lateral margins subparallel; dentary symphysis anterior end laterolaterally compressed (about three times alveolar width in total, inclusive of the alveoli); median dentary groove deep and narrow (subequal to half of the alveolar width); bony bar separating the first pair of alveoli thin (half of alveolar width or less; on both jaws); dentition spacing approximately even along jaws; dentition extending for over 70% total jaw length.
Remarks. The present conceptualization of this clade is centered around closer phylogenetic proximity to Targaryendraco wiedenrothi than to Anhanguera blittersdorffi, Hamipterus tianshanensis, or Ornithocheirus simus, irrespective of how inclusive this clade may end up being. According to the present analysis, this clade encompasses two main lineages: targaryendraconids and cimoliopterids. This name and definition are perfectly compatible with alternative phylogenies, even if with a distinct composition. Under other phylogenetic hypotheses, this clade ends up being restricted to (thus being redundant and synonymous with) Targaryendraconidae (see Andres, 2021).
TARGARYENDRACONIDAE Pêgas et al. (2019),
converted clade name
Registration number. 1059.
Definition. The most inclusive clade containing Targaryendraco wiedenrothi (Wild, 1990) but not Cimoliopterus cuvieri (Bowerbank, 1851), Hamipterus tianshanensis Wang et al., 2014, Anhanguera blittersdorffi Campos and Kellner, 1985, or Ornithocheirus simus Owen, 1861. This is a branch-based definition.
Reference phylogeny. Present work (Figure 2).
Composition. Aussiedraco molnari, Barbosania gracilirostris, Targaryendraco wiedenrothi.
Diagnostic apomorphies. Dentary symphysis anterior region with convex occlusal surface; dentary groove extending at the level of the first pair of alveoli (but not to the rostral tip); dentary groove lateral rims conspicuously raised above alveolar margin; enlarged lower first tooth (larger than the second).
Remarks. This clade is intended to encompass all targaryendraconians closer to Targaryendraco wiedenrothi than to Cimoliopterus cuvieri, what may end up being redundant with Targaryendraconia itself under alternative phylogenetic hypotheses. Authors should choose one name over the other in such instances (preferably Targaryendraconia, which is intended as the most general/inclusive one).
Of note, Averianov (2025) recently claimed that the correct name formation, under ICZN rules, should be "Targaryendracoidae", through the addition of “-idae” to the stem of the type genus (Article 29.2 of the Code). However, this does not take into consideration Article 29.3.1 of the Code, which determines that, if “a generic name is or ends in a Greek or Latin word, [...] the stem for the purposes of the Code is found by deleting the case ending of the appropriate genitive singular” (ICZN, 1999). Targaryendraco ends in a Latin word (draco), for which the genitive singular is draconis, hence Targaryendraconidae (and not “Targaryendracoidae”). See for example the agamid subfamily Draconinae Fitzinger (1826), not “Dracoinae”.
CIMOLIOPTERIDAE Pêgas et al. (2019),
converted clade name
Registration number. 1060.
Definition. The most inclusive clade containing Cimoliopterus cuvieri (Bowerbank, 1851) but not Targaryendraco wiedenrothi (Wild, 1990), Hamipterus tianshanensis Wang et al., 2014, Anhanguera blittersdorffi Campos and Kellner, 1985, or Ornithocheirus simus Owen, 1861. This is a branch-based definition.
Reference phylogeny. Present work (Figure 2).
Composition. Aetodactylus halli, Camposipterus nasutus, Cimoliopterus colorhinus comb. nov., Cimoliopterus cuvieri, Cimoliopterus dunni.
Diagnostic apomorphies. Presence of a rhinoid process (“nose-like” anterior protrusion of the rostrum above the first alveoli); palatal ridge anterior end reaching a level between the second and third pair of alveoli; first three pairs of alveoli closely packed together; first pair of alveoli separated by a laminar sheet of bone.
Remarks. The present conceptualization of this clade is centered around the close relationship between Cimoliopterus cuvieri and Camposipterus nasutus, along with any further taxa may be recovered as closer to Cimoliopterus cuvieri than to any of the external specifiers mentioned above, irrespective of how inclusive this clade may end up being. This name and definition are perfectly compatible with alternative phylogenetic hypotheses, even if with a distinct composition. Although restricted to Cimoliopterus, Camposipterus and Aetodactylus under the present phylogenetic hypothesis, this clade may encompass further genera under alternative ones (e.g. Jacobs et al., 2019; Andres, 2021). Of note, 'Ornithocheirus colorhinus’, which was tentatively allocated into Camposipterus by Rodrigues and Kellner (2013) previously, is recognized here as closely related to Cimoliopterus cuvieri, and thus renamed as Cimoliopterus colorhinus comb. nov.
ANHANGUERIA Rodrigues and Kellner (2013),
converted clade name
Registration number. 1061.
Definition. The most inclusive clade that includes Anhanguera blittersdorffi Campos and Kellner, 1985 but not Cimoliopterus cuvieri (Bowerbank, 1851) or Targaryendraco wiedenrothi (Wild, 1990). This is a branch-based definition.
Reference phylogeny. Present work (Figure 2).
Composition. Anhangueridae and Hamipteridae.
Diagnostic apomorphies. Lateral expansion of the anterior jaw tip (forming a rosette comprising 4-5 alveoli); prominent humeral supracondylar process.
Remarks. The present conceptualization of this clade is centered around the hypothesis of monophyly of that subset of ornithocheiriform species that share laterally expanded jaw tips (as originally intended by the nominal authors Rodrigues and Kellner, 2013); to the exclusion of similar taxa that lacks this feature (Figure 5). This name and definition are still compatible with alternative phylogenies, even if with a slightly distinct composition.
Application of this name in some alternative topologies results in a rather restrictive clade that excludes many taxa that exhibit laterally expanded jaw tips (e.g., Andres, 2021). However, it must be noted that these topologies are skewed regarding the optimization of this character, given that it is mistakenly coded as present in taxa such as Ornithocheirus simus, Barbosania gracilirostris, Cimoliopterus cuvieri, and Camposipterus nasutus (see Andres, 2021); all of which actually lack this feature (see Elgin and Frey, 2010; Rodrigues and Kellner, 2013; Pêgas et al., 2019). Alternative phylogenetic datasets require revisions before the optimization of this character can be solidly discussed under the light of the application of the name Anhangueria.
It is worth highlighting that, as originally intended (Rodrigues and Kellner, 2013), the main diagnostic feature of anhanguerians, the so-called “lateral expansion” of the jaw tips, refers to a feature that is distinct from the condition found in Cimoliopterus cuvieri. While this taxon could also, arguably, be described as possessing a “lateral expansion” of the jaw tip, the condition seen in this species (and in targaryendraconians overall) is not the same as that seen in anhanguerians. In targaryendraconians (or, in other words, in non-anhanguerian anhangueroids), a lateral expansion of the third alveolar margin causes the jaw tip to be wider at that region; however, a posterior constriction that effectively delineates a “jaw tip expansion” is lacking (Figure 5). In anhanguerians, in contrast, the first 4-5 alveoli are contained in an anterior jaw tip that is followed by a post-rosette constriction that is most noticeable between the 5th and 6th tooth positions. This latter condition is referred to here as the anhanguerian-type “well-developed lateral expansion of the jaw tip”, being present in anhanguerians and absent in targaryendraconians (Figure 5).
The anhanguerian-like jaw tip expansion can be quantitatively assessed in Table 1 and Table 2 through the “expansion ratio”, calculated as the proportion between a “mid jaw-tip width” (jaw section between the 3rd and 4th pairs of alveoli) and a “posterior jaw-tip width” (jaw section between the 5th and 6th pairs of alveoli). Anhanguerians exhibit noticeable upper jaw expansion ratios of 1.10-1.49 (to the exception of Siroccopteryx moroccensis), while targaryendraconians exhibit expansion ratios of ~1.0 or less (indicating the lack of a constriction). In turn, the lateral offset of the third alveolus (that characterizes anhangueroids overall) can be quantitatively assessed in Table 1 and Table 2 through the “offset ratio”, calculated as the proportion between the maximum width at the third pair of alveoli and at the fifth one. In targaryendraconians there is a very small (yet noticeable) offset of the third alveolus around 1.10, while in anhanguerids this ratio typically surpasses that value (again to the exception of Siroccopteryx moroccensis; Table 1). Note that targaryendraconians exhibit a gentle offset of the third alveolus even though a proper jaw tip expansion is absent (Figure 5; Table 1 and Table 2).
HAMIPTERIDAE Holgado et al. (2019),
converted clade name
Registration number. 1062.
Definition. The most inclusive clade that includes Hamipterus tianshanensis Wang et al., 2014 but not Anhanguera blittersdorffi Campos and Kellner, 1985, Cimoliopterus cuvieri (Bowerbank, 1851), or Targaryendraco wiedenrothi (Wild, 1990). This is a branch-based definition.
Reference phylogeny. Present work (Figure 2).
Composition. Hamipterus tianshanensis and Iberodactylus andreui.
Diagnostic apomorphies. Premaxillary crest anterior region with well-defined curved striae and sulci; premaxillary crest with an anterodorsal expansion.
Remarks. As presently conceptualized, this clade is intended to group species that share a similar premaxillary crest morphology with Hamipterus tianshanensis. This clade is also intended to reflect the sister-relationship between Hamipterus tianshanensis and Iberodactylus andreui as recovered by Holgado et al. (2019) and other recent studies (Pêgas, 2024; Jacobs et al., 2024).
Of note, the synapomorphies that join Hamipterus tianshanensis and Iberodactylus andreui pertain to premaxillary crest expression. Naturally, the use of premaxillary crest morphology in pterosaur systematics requires great caution, given the considerable effects that ontogeny and sexual dimorphism may have in pterosaur crests (e.g., Bennett, 1993; Manzig et al., 2014; Wang et al., 2014; Pinheiro and Rodrigues, 2017; Duque et al., 2020). Luckily, the Hamipterus tianshanensis bonebed provides a rare chance to explore ontogenetic and sexual variations in the premaxillary crest of a pterosaur. The bonebed includes dozens of crested subadult/adult skulls (Wang et al., 20114). Despite considerable variation in crest relative height/robustness, all crests share the same texture (with curved striae and sulci) and overall pattern (all with an anterodorsal expansion, even though this is much more accentuated in inferred males than in inferred females) - see Wang et al. (2014). Similarly, known ontogenetic variations in anhanguerid crests tend to relate to their relative dimensions, but not to overall shape or texture (Pinheiro and Rodrigues, 2017). Therefore, the intraspecific variations seen in Hamipterus tianshanensis and other anhanguerians seem not to affect the proposed diagnosis of Hamipteridae (the shape and texture of the premaxillary crests), which are herein regarded as useful diagnostic features at least for the crested morphs.
ANHANGUERIDAE Campos and Kellner (1985)
{this work}
Registration number. 1063.
Definition. The most inclusive clade that includes Anhanguera blittersdorffi Campos and Kellner, 1985, but not Ornithocheirus simus Owen, 1861, Hamipterus tianshanensis Wang et al., 2014, Cimoliopterus cuvieri (Bowerbank, 1851), or Targaryendraco wiedenrothi (Wild, 1990). This is a branch-based definition. This is also an unrestricted emendation (see the previous definition of Andres, 2021).
Reference phylogeny. Present work (Figure 2).
Composition. Anhanguerinae, Coloborhynchinae, Tropeognathinae.
Diagnostic apomorphies. Presence of frontal and parietal crests; jugal contact with lacrimal in the ventral half of the orbit; broad lacrimal process of the jugal; presence of a sagittal dentary crest; first upper alveoli mostly (or totally) dorsally surpassing second upper alveoli; second upper alveolus anteroposterior axis markedly more anterodorsally inclined than subsequent alveoli; and symmetric sternocoracoid articulations.
Remarks. As mentioned above, the recently registered definitions of Anhangueridae and Ornithocheiridae (Andres, 2021) abruptly discontinue their traditional usages, especially regarding the traditional concept of Anhangueridae (see Introduction). It could be argued that Anhangueridae sensu Andres (2021) is eligible for the petitioning of a restricted emendation, as this definition does not strongly tie this clade name to its current/traditional conceptualization (failing Article 10.1 of the PhyloCode).
However, considering that Anhangueridae sensu Andres (2021) essentially applies to a nested ornithocheiriform group that comprises species closer to Anhanguera blittersdorffi than to either Ornithocheirus simus, cimoliopterids, Hamipterus tianshanensis, or boreopterids; then an unrestricted emendation that redefines the group in such a way would be justifiable and warranted in order to prevent a dramatic shift in clade conceptualization/application between alternative phylogenies (Andres, 2021; Pêgas, 2024). Such unrestricted emendation would fully preserve the application of the clade name between alternative phylogenies (Andres, 2021; Pêgas, 2024), fulfilling the requirements of Article 15.11 of the PhyloCode for an unrestricted emendation proposal (De Queiroz and Cantino, 2020). Also, this unrestricted emendation would create a solid association between this clade name and its historical/current conceptualization (e.g., Kellner, 2003; Rodrigues and Kellner, 2008, 2013; Holgado et al., 2019; Holgado and Pêgas, 2020), including under the reference phylogeny of the original definition (Andres, 2021), thus preserving its usage and nomenclatural stability (and avoiding the need for a restricted emendation).
The present emendation does not necessarily overwrite previous ones (e.g., Andres, 2021). For researchers preferring to follow the original reference phylogeny of Andres (2021), the definitions of Andres (2021) will still be available for usage (PhyloCode Article 15.15; De Queiroz and Cantino, 2020).
As herein conceptualized, this clade is intended to encompass both Anhanguera blittersdorffi and Tropeognathus mesembrinus while at the same time excluding members of Ornithocheiridae and Targaryendraconia, as traditionally and consistently conceptualized (Kellner, 1989, 2003; Wang and Zhou, 2003, 2012; Rodrigues and Kellner, 2013; Bantim et al., 2014; Aureliano et al., 2014; Pinheiro and Rodrigues, 2017; Holgado and Pêgas, 2020; Richards et al., 2021; Pentland et al., 2024; Jacobs et al., 2024).
One of the main synapomorphies herein recovered for the Anhangueridae (sensu the present work) regards the orientation of the second alveolus. In non-anhanguerid anhangueroids, the anteroposterior axis of the second alveolus aligns itself with those of the subsequent alveoli (Figure 4). In contrast, in anhanguerids there is a marked shift in anteroposterior axis orientation between the second alveolus and subsequent ones. While the subsequent ones maintain a constant orientation, in the second one the axis is markedly more anterodorsally inclined relative to the subsequent ones (i.e., the alveolus is markedly anteroventrally oriented, despite not as much as the first alveolus, which is anteroventrally oriented in all anhangueroids). This condition can be clearly seen in well-preserved anhanguerid rostrum remains (Figure 4), as seen in the anhanguerines Anhanguera spp. (Kellner and Tomida, 2000; Fastnacht, 2001), Ludodactylus sibbicki (Frey et al., 2003), and Caulkicephalus trimicrodon (Steel et al., 2005); the coloborhynchines Nicorhynchus capito and Uktenadactylus wadleighi (see Rodrigues and Kellner, 2013; Holgado and Pêgas, 2020); and the tropeognathines Aerodraco sedgwicki, Akharhynchus martilli, Siroccopteryx moroccensis, and Tropeognathus mesembrinus (see Holgado and Pêgas, 2020; Jacobs et al., 2024).
TROPEOGNATHINAE Holgado and Pêgas (2020),
converted clade name
Registration number. 1064.
Definition. The most inclusive clade that includes Tropeognathus mesembrinus Wellnhofer, 1987, but not Anhanguera blittersdorffi Campos and Kellner, 1985 or Coloborhynchus clavirostris Owen, 1874. This is a branch-based definition.
Reference phylogeny. Present work (Figure 2).
Composition. Aerodraco sedgwicki, Akharhynchus martilli, Ferrodraco lentoni, Haliskia peterseni, Siroccopteryx moroccensis, and Tropeognathus mesembrinus.
Diagnostic apomorphies. Lateral expansion of the anterior jaw tip relatively narrow (under 130% the width of the posterior constriction); premaxillary crest anterior margin level with rostrum tip; relatively stout premaxillary teeth (length no more than 4 times width); upper alveoli 4–5 smaller than 3 and 6.
Remarks. This clade is herein conceptualized as an anhanguerid subclade encompassing all taxa closer to Tropeognathus mesembrinus than to either Anhanguera blittersdorffi or Coloborhynchus clavirostris. Under certain phylogenetic analyses, this lineage may be seen as redundant with Tropeognathus mesembrinus itself (Andres, 2021). Still, the inclusion of the Australian Ferrodraco lentoni and the Moroccan Siroccopteryx moroccensis have been greatly supported by other analyses (Holgado and Pêgas, 2020; Richards et al., 2023; Pentland et al., 2024; Jacobs et al., 2024). The inclusion of Haliskia peterseni and Akharhynchus martilli, as recently suggested (Pentland et al., 2024; Jacobs et al., 2024), is also corroborated here. The Cambridge Greensand anhanguerid Aerodraco sedgwickii, previously considered a member of Coloborhynchinae (Holgado and Pêgas, 2020), has been recently recovered as a tropeognathinae instead (Pêgas, 2024). This is corroborated here, with Aerodraco sedgwickii being recovered as a close relative of the Kem Kem Group tropeognathines Siroccopteryx moroccensis and Akharhynchus martilli. See also Holgado (2021) for further considerations.
Thapuungaka shawi has been recovered as a member of Tropeognathinae in previous studies (Richards et al., 2021, 2023; Pêgas, 2024; Pentland et al., 2024). However, it differs from anhangueroids overall in lacking the following features: a dorsally reflected palatal tip, an anteroventrally facing first upper alveolus, a palatal ridge, and a mandibular groove; and further differs from anhanguerians in lacking a proper rostral expansion; and from anhanguerids in lacking an anteroventralized second premaxillary alveolus (see Richards et al., 2021, 2023). It is herein recovered as a closer relative of Ornithocheirus simus instead (see further below).
A subtribe Mythungini has been recently proposed as a subclade within Tropeognathinae (Richards et al., 2023). Given that Mythunga camara is best seen as a nomen dubium as defended here, it is regarded here that this clade name should not be used. Therefore, this clade name is not converted or revised here.
Of note, one of the synapomorphies herein recovered for Tropeognathinae pertain to premaxillary crest morphology: premaxillary crest anterior margin level with rostrum tip. This differs from the typical condition seen in other anhangueroids (e.g., Cimoliopterus, Hamipterus, Anhanguera, Uktenadactylus, Nicorhynchus), which exhibit a recess between the rostrum tip and the premaxillary crest. The presence of such recess is general for all known crested morphs of Hamipterus and Anhanguera, both of which are known from tens of skulls of variable ontogenetic stages and sexual morphs (Wang et al., 2014; Pinheiro and Rodrigues, 2017), and therefore this does not seem to suffer from ontogenetic or sexual influences at least in the crested morphs. The absence of such recess is exclusive to tropeognathines among anhangueroids and seems to be a useful diagnostic feature.
COLOBORHYNCHINAE Holgado et al. (2019),
converted clade name
Registration number. 1065.
Definition. The most inclusive clade that includes Coloborhynchus clavirostris Owen, 1874, but not Tropeognathus mesembrinus Wellnhofer, 1987 or Anhanguera blittersdorffi Campos and Kellner, 1985. This is a branch-based definition.
Reference phylogeny. Present work (Figure 2).
Composition. Coloborhynchus clavirostris, Nicorhynchus capito, Nicorhynchus fluviferox, Uktenadactylus rodriguesae, Uktenadactylus wadleighi.
Diagnostic apomorphies. Lateral expansion of the anterior jaw tip with parallel lateral margins (expansion subquadrangular in shape); dorsal reflection of the palatal tip of ~90°; first upper alveoli completely anteriorized; first upper alveoli strongly elliptical in shape; first upper alveoli do not overlap in level with second upper alveoli; second upper alveolus aligned with third one (in the horizontal plane); third upper alveolus over twice as wide as the fourth one.
Remarks. This clade has already been explored in detail recently by Holgado and Pêgas (2020). It is intended to encompass all anhanguerid species closer to Coloborhynchus clavirostris than to either Anhanguera blittersdorffi or Tropeognathus mesembrinus. The inclusion of Uktenadactylus wadleighi and Nicorhynchus capito is consensual between the present phylogenetic hypotheses and alternative ones (Andres, 2021). The inclusion of Nicorhynchus fluviferox is also supported by the phylogenetic hypothesis of Jacobs et al. (2019, 2020; not analyzed by Andres, 2021); while Uktenadactylus rodriguesae remains to be analyzed under alternative phylogenetic hypotheses. Some hypotheses may also include Siroccopteryx moroccensis in this clade (Holgado et al., 2019; Andres, 2021); but see Jacobs et al. (2019) and Holgado and Pêgas (2020) for criticism to this view.
ANHANGUERINAE Holgado et al. (2019),
converted clade name
Registration number. 1066.
Definition. The most inclusive clade that includes Anhanguera blittersdorffi Campos and Kellner, 1985 but not Coloborhynchus clavirostris Owen, 1874 or Tropeognathus mesembrinus Wellnhofer, 1987. This is a branch-based definition.
Reference phylogeny. Present work (Figure 2).
Composition. Anhanguera blittersdorffi, Anhanguera piscator, Anhanguera spielbergi, Brasileodactylus araripensis, Caulkicephalus trimicrodon, Guidraco venator, Liaoningopterus gui, Ludodactylus sibbicki, Maaradactylus kellneri.
Diagnostic apomorphies. Premaxillary crest anterior base tapered; second premaxillary tooth apex extending anterior to first tooth apex; presence of a post-rosette toothline notch (in lateral view); mesialmost maxillary teeth relatively reduced (smaller than fourth premaxillary tooth and followed by markedly larger mid-maxillary teeth).
Remarks. All ornithocheiriforms exhibit a particular pattern of size heterodonty, wherein the mesialmost maxillary teeth are smaller than mid-maxillary teeth, which are then followed by progressively smaller teeth again (Unwin, 2001; 2003). However, this pattern is not expressed in the same way throughout ornithocheiriforms. In most ornithocheiriforms, the mesialmost maxillary teeth are not reduced relative to the last (fourth) premaxillary tooth; and the size difference between mesialmost and mid-maxillary teeth is ever so slight (e.g., ornithocheirids, targaryandraconians, hamipterids, tropeognathines; Figure 5). Anhanguerines deviate from this. Presence of fifth and sixth upper teeth markedly smaller than fourth and seventh has been a long-held purported synapomorphy of the genus Anhanguera (Kellner, 2003; Pinheiro and Rodrigues, 2017) but is herein recognized as much more broadly distributed. This feature is expressed in the same way in Liaoningopterus gui, Ludodactylus sibbicki, and Guidraco venator. Furthermore, a similar feature is present in Caulkicephalus trimicrodon and Maaradactylus kellneri, wherein fifth, sixth and seventh upper teeth are smaller than fourth and eighth. In other words, all these taxa exhibit relatively reduced mesialmost maxillary teeth, always smaller than the last (fourth) premaxillary tooth, and followed by markedly enlarged maxillary teeth posteriorly (or mid-maxillary teeth); wherein the number of reduced mesialmost maxillary teeth simply varies from 2 (Anhanguera, Liaoningopterus, Ludodactylus, Guidraco) to 3 (Maaradactylus, Caulkicephalus). It is worth noting that the degree of alveolar size variation is ontogenetic, with juveniles exhibiting a gentle (yet present) variation if compared to adults (Duque et al., 2022).
Besides this feature, anhanguerines (sensu the present work) further share a second premaxillary tooth apex extending anterior to first tooth apex, and a post-rosette toothline notch (in lateral view). These two features cannot be found in any other ornithocheiriforms.
ORNITOCHEIRIDAE Burckhardt, 1896
{this work}
Registration number. 1069.
Definition. The most inclusive clade that includes Ornithocheirus simus Owen, 1861, but not Anhanguera blittersdorffi Campos and Kellner, 1985, Hamipterus tianshanensis Wang et al., 2014, Cimoliopterus cuvieri (Bowerbank, 1851), or Targaryendraco wiedenrothi (Wild, 1990). This is an unrestricted emendation.
Reference phylogeny. Present work (Figure 2).
Composition. Ornithocheirus simus, the Kem Kem ornithocheirid, and Thapunggaka shawi.
Diagnostic apomorphies. Rostrum tip with a ram-like anterior surface.
Remarks. Three nodes set Ornithocheiridae apart from Anhangueridae sensu this work (Figure 2). Of note, the Ornithocheiridae sensu this work lack the following seven relevant features: a dorsal reflection of the palatal tip forming a deltoid facet, a lateral offset of the third alveolus, a palatal ridge, and an individualized mandibular groove (synapomorphies of Anhangueroidea); a lateral expansion of the anterior jaw tip forming a rosette (a synapomorphy of Anhangueria); and a second premaxillary alveolus markedly more anteroventrally oriented than subsequent alveoli (a synapomorphy of Anhangueridae). Forcing Ornithocheirus simus as the sister-group of Tropeognathus mesembrinus requires 10 additional steps. Forcing Ornithocheirus simus as the sister-group of Coloborhynchus clavirostris (or of Coloborhynchinae) requires 12 additional steps. Forcing Ornithocheiridae (sensu this work) as the sister-group of Coloborhynchus clavirostris (or of Coloborhynchinae) requires 8 additional steps.
In some phylogenetic analyses, Ornithocheirus simus is recovered as closely related to Coloborhynchus clavirostris and allies (i.e., a member of Coloborhynchinae), based on the following purported synapomorphy: a blunt rostral tip (Andres et al., 2014; Andres, 2021). However, the blunt rostral surface of Ornithocheirus simus is different from that of Coloborhynchus clavirostris, each comprising entirely distinct anatomical regions. The blunt rostral surface (or ram-like rostral surface, as termed here) of Ornithocheirus simus comprises, developmentally speaking, the non-palatal anterior apex of the premaxillae (Owen, 1861; Rodrigues and Kellner, 2013). In contrast, the blunt rostral surface of Coloborhynchus clavirostris comprises the deltoid facet, or in other words, the reflected palatal tip (Owen, 1874; Rodrigues and Kellner, 2013). Therefore, developmentally speaking, the blunt rostral surfaces of Ornithocheirus simus and Coloborhynchus clavirostris are essentially distinct, clearly comprising different anatomical structures (one is not palatal in origin, while the other is), and therefore should not be coded as the same state of the same character.
The Australian species Thapunngaka shawi is herein recovered as a close relative of Ornithocheirus simus for the first time. It is noted here that Thapunngaka shawi exhibits an outstanding resemblance to Ornithocheirus simus (and to the Kem Kem ornithocheirid as well), as revealed by the discovery of KK F600, a partial rostrum tentatively referred to T. shawi (Richards et al., 2023). Unfortunately, this specimen lacks any directly comparable elements to the holotype of T. shawi (KK F494, a partial lower jaw), what lead some authors to question such a referral (Pentland et al., 2024). The present work corroborates that KK F494 and KK F600 represent complementary upper and lower jaws, sharing the following combination of features: matching patterns of alveolus size variation (1 < 2 < 3 > 4 = 5 < 6 > 7 < 8), matching occlusal surfaces (ridgeless palate in KK F494, and grooveless mandible in KK F600), and matching pattern of occlusal margin curvature in lateral view (see Richards et al., 2021; 2023).
Here, the definition of Ornithocheiridae is emended to generate stability in its application relative to the other clade names herein defined (or emended, in the case of Anhangueridae) in the context of the present phylogenetic hypothesis (Figure 2), which are not intended to overlap with Ornithocheiridae in composition. This emendation is intended to preserve stability of the name Ornithocheiridae when applied to multiple phylogenetic hypotheses (e.g., Andres, 2021; Pêgas, 2024), following Article 15.11 of the PhyloCode.
The present emended definitions for Anhangueridae and Ornithocheiridae generate continuity with traditional usage regarding the “restrictive Anhangueridae” scheme (e.g., Rodrigues and Kellner, 2013), though not with the “broad Ornithocheiridae” scheme (Unwin, 2001, 2003). Indeed, applications of these two schemes are mutually exclusive as explained above (see Introduction). Still, traditional usage of a “broad Ornithocheiridae”, or Ornithocheiridae sensu Unwin (2001, 2003), is continuous with present usage of Ornithocheiriformes sensu Andres (2021), which is herein adopted and defended. Therefore, from the viewpoint of traditional nomenclature, it could be said that the present work honors both aforementioned traditional schemes in (1) maintaining the traditional usage and spelling of Anhangueridae (e.g., Kellner, 2003; Rodrigues and Kellner, 2013), as well as (2) elevating the Ornithocheiridae sensu Unwin (2001, 2003) into the Ornithocheiriformes (similarly to how Pteranodontidae sensu Bennett, 1989 has been elevated to Pteranodontoidea sensu Kellner, 2003).
A clade Ornithocheirinae has been recently defined as a node-based clade internally specified by Ornithocheirus simus and Coloborhynchus clavirostris and apparently intended as a subclade of Ornithocheiridae (see Andres, 2021). This clade is correspondent to Ornithocheiriformes in the present phylogenetic hypothesis (see topology in Figure 2) and will thus require an emendation to preserve its application as a subclade of Ornithocheiridae. However, this is deemed unnecessary now, especially considering that, under the context of the present phylogenetic hypothesis, ornithocheirids comprise only three species with poorly resolved relationships.
CONCLUDING REMARKS
Adherence to the PhyloCode in pterosaur systematics provides an opportunity for standardization in ornithocheiriform nomenclature from now on. Still, it is crucial that phylogenetic definitions be proposed aiming at preserving stability in usage/composition, as well as avoiding conflicts with traditional nomenclature. It is hoped that the new emendations, names, and definitions proposed here will provide a stable nomenclatorial scheme fit for a broad range of phylogenetic hypotheses, and mostly non-conflictive with ICZN-based ornithocheiriform taxon names. Finally, the present work urges authors to carefully consider traditional usage as well as alternative phylogenetic hypotheses when constructing phylogenetic definitions, so as to avoid definitions that work well only under certain hypotheses while causing excessive disruption under others.
ACKNOWLEDGMENTS
I thank FAPESP for a postdoctoral fellowship (#2023/11296-0), and two anonymous reviewers for their thoughtful remarks. For access to specimens under their care, I thank U. Cabral, L. Carvalho, and A. Kellner (MN), A. Pinheiro (MPPCN), D. Martill and S. Zouhri (FSAC specimens in UoP), O. Rauhut (BSPG), R. Schoch (SMNS), D. Schreiber (SMNK), M. Day (NHMUK), M. Riley and R. Theodore (CAMSM), M. Munt (IWCMS), S. Jiang and X. Wang (IVPP), M. Manabe, T. Tsuihiji, and C. Sakata (NSM), and T. Alves (MHNS). Thanks also to F. Pinheiro and B. Holgado for several fruitful discussions.
REFERENCES
Andres, B. 2021. Andres, B. 2021. Phylogenetic systematics of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea) . Journal of Vertebrate Paleontology, 41(sup1):203-217.
https://doi.org/10.1080/02724634.2020.1801703
Andres, B., Clark, J., and Xu, X. 2014. The earliest pterodactyloid and the origin of the group. Current Biology, 24(9):1011-1016.
https://doi.org/10.1016/j.cub.2014.03.030
Andres, B. and K. Padian. 2020a. Pterosauria†; pp. 1201-1204. In de Queiroz, K., Cantino, P.D., and Gauthier, J.A. (eds.), Phylonyms. A Companion to the PhyloCode. CRC Press, Boca Raton, FL.
Aureliano, T., Ghilardi, A.M., Duque, R.R., and Barreto, A.M. 2014. On the occurrence of Pterosauria in Exu, Pernambuco (Lower Cretaceous Romualdo Formation, Araripe Basin), Northeastern Brazil. Estudos Geológicos, 24(2):15-27.
https://doi.org/10.18190/1980-8208/estudosgeologicos.v24n2p15-27
Averianov, A.O. 2025. A new ornithocheiran pterosaur from the Upper Cretaceous (Cenomanian) of Saratov, Russia. Anais da Academia Brasileira de Ciências, 97: e20241063.
https://doi.org/10.1590/0001-3765202520241063
Bantim, R.A., Saraiva, A.A., Oliveira, G.R., and Sayão, J.M. 2014. A new toothed pterosaur (Pterodactyloidea: Anhangueridae) from the Early Cretaceous Romualdo Formation, NE Brazil. Zootaxa, 3869(3):201-223.
https://doi.org/10.11646/zootaxa.3869.3.1
Bennett, S.C. 1989. A pteranodontid pterosaur from the Early Cretaceous of Peru, with comments on the relationships of Cretaceous pterosaurs. Journal of Paleontology, 63:669-677.
https://doi.org/10.1017/S0022336000041305
Bennett, S.C. 1993. The ontogeny of Pteranodon and other pterosaurs. Paleobiology, 19(1): 92-106.
https://www.jstor.org/stable/2400773
Bowerbank, J.S. 1851. On the pterodactyls of the Chalk Formation. Proceedings of the Zoological Society of London, 19:14-20.
Campos, D.A. and Kellner, A.W.A. 1985. Panorama of the flying reptiles study in Brazil and South America. Anais da Academia Brasileira de Ciências, 57:453-466.
De Queiroz, K. and Cantino, P. 2020. International code of phylogenetic nomenclature (PhyloCode). 1st Edition. CRC Press, Boca Raton.
https://doi.org/10.1201/9780429446320
Duque, R.R.C., Pinheiro, F.L., and Barreto, A.M.F. 2022. The ontogenetic growth of Anhangueridae (Pterosauria, Pterodactyloidea) premaxillary crests as revealed by a crestless Anhanguera specimen. Journal of Vertebrate Paleontology, 42(1): e2116984.
https://doi.org/10.1080/02724634.2022.2116984
Elgin, R.A. and Frey, E. 2011. A new ornithocheirid, Barbosania gracilirostris gen. et sp. nov. (Pterosauria, Pterodactyloidea) from the Santana Formation (Cretaceous) of NE Brazil. Swiss Journal of Palaeontology, 130:259-275.
https://doi.org/10.1007/s13358-011-0017-4
Fastnacht, M. 2001. First record of Coloborhynchus (Pterosauria) from the Santana Formation (Lower Cretaceous) of the Chapada do Araripe, Brazil. Paläontologische Zeitschrift, 75(1):23-36.
https://doi.org/10.1007/BF03022595
Fitzinger, L. 1826. Neue classification der reptilien nach ihren natürlichen verwandtschaften: nebst einer verwandtschaftstafel und einem verzeichnisse der reptilien-sammlung des K. K. zoologischen museum’s zu Wien. J. G. Heubner, Vienna.
https://doi.org/10.5962/bhl.title.4683
Goloboff, P.A. and Morales, M.E. 2023. TNT version 1.6, with a graphical interface for MacOS and Linux, including new routines in parallel. Cladistics, 39(2):144-153.
https://doi.org/10.1111/cla.12524
Holgado, B. 2021. On the validity of the genus Amblydectes Hooley 1914 (Pterodactyloidea, Anhangueridae) and the presence of Tropeognathinae in the Cambridge Greensand. Anais da Academia Brasileira de Ciências, 93:e20201658.
https://doi.org/10.1590/0001-3765202120201658
Holgado, B. and Pêgas, R.V. 2020. A taxonomic and phylogenetic review of the anhanguerid pterosaur group Coloborhynchinae and the new clade Tropeognathinae. Acta Palaeontologica Polonica, 65(4):743-761.
https://doi.org/10.4202/app.00751.2020
Holgado, B., Pêgas, R.V., Canudo, J.I., Fortuny, J., Rodrigues, T., Company, J., and Kellner, A.W.A. 2019. On a new crested pterodactyloid from the Early Cretaceous of the Iberian Peninsula and the radiation of the clade Anhangueria. Scientific Reports, 9(1):e4940.
https://doi.org/10.1038/s41598-019-41280-4
Jacobs, M.L., Martill, D.M., Ibrahim, N., and Longrich, N. 2019. A new species of Coloborhynchus (Pterosauria, Ornithocheiridae) from the mid-Cretaceous of North Africa. Cretaceous Research, 95:77-88.
https://doi.org/10.1016/j.cretres.2018.10.018
Jacobs, M.L., Martill, D.M., Unwin, D.M., Ibrahim, N., Zouhri, S., and Longrich, N.R. 2020. New toothed pterosaurs (Pterosauria: Ornithocheiridae) from the middle Cretaceous Kem Kem beds of Morocco and implications for pterosaur palaeobiogeography and diversity. Cretaceous Research, 110:104413.
https://doi.org/10.1016/j.cretres.2020.104413
Jacobs, M.L., Smith, R.E., Zouhri, S. 2024. A new ornithocheirid pterosaur (Pterosauria: Ornithocheiridae) from the mid-Cretaceous Ifezouane Formation, Kem Kem Group of Morocco. Cretaceous Research, e2024.106015.
https://doi.org/10.1016/j.cretres.2024.106015
Kellner, A.W.A. 1989. Os répteis voadores do Cretáceo brasileiro. Anuário do Instituto de Geociências, 12:86-106.
https://doi.org/10.11137/1989_0_86-106
Kellner, A.W.A. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. Geological Society, London, Special Publications, 217(1):105-137.
https://doi.org/10.1144/GSL.SP.2003.217.01.1
Kellner, A.W.A. and Tomida, Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), northeastern Brazil. National Science Museum Monograhps, 17:ix-137.
Leardi, J.M., Pol, D., Montefeltro, F., da Silva Marinho, T., Ruiz, J.V., Bravo, G.G., Godoy, P.L., Nicholl, C.S.C., Lecuona, A., and Larsson, H.C. 2024. Phylogenetic nomenclature of Notosuchia (Crocodylomorpha; Crocodyliformes). Bulletin of Phylogenetic Nomenclature, 1(3):44-82.
https://doi.org/10.11646/bpn.1.3.2
Madzia, D., Arbour, V.M., Boyd, C.A., Farke, A.A., Cruzado-Caballero, P., and Evans, D.C. 2021. The phylogenetic nomenclature of ornithischian dinosaurs. PeerJ, 9:e12362.
https://doi.org/10.7717/peerj.12362
Manitkoon, S., Pêgas, R.V., Nonsrirach, T., Warapeang, P., Lauprasert, K., Deesri, U., Tumpeesuwan, S., Wongko, K., and Zhou, X. 2025. First gnathosaurine (Pterosauria, Pterodactyloidea) from the Early Cretaceous of eastern Thailand. Cretaceous Research, 173:e106135.
https://doi.org/10.1016/j.cretres.2025.106135
Manzig, P.C., Kellner, A.W., Weinschütz, L.C., Fragoso, C.E., Vega, C.S., Guimarães, G.B., Godoy, L.C., Liccardo, A., Ricetti, J.H.Z., and de Moura, C.C. 2014. Discovery of a rare pterosaur bone bed in a Cretaceous desert with insights on ontogeny and behavior of flying reptiles. PloS one, 9(8):e100005.
https://doi.org/10.1371/journal.pone.0100005
Molnar, R.E. and Thulborn, R.A. 2008. An incomplete pterosaur skull from the Cretaceous of north-central Queensland, Australia. Arquivos do Museu Nacional, Rio de Janeiro, 65:461-470.
https://biostor.org/reference/248724
Owen, R. 1842. Report on British Fossil Reptiles, Part II: In Report of the Eleventh Meeting of the British Association for the Advancement of Science, 1841:60-204. John Murray, Plymouth.
Owen, R. 1861. Monograph on the fossil Reptilia of the Cretaceous formations. Supplement No. III, Pterosauria (Pterodactylus) and Sauropterygia (Polyptychodon). Palaeontographical Society Monograph, 12:1-25.
Owen, R. 1874. The Fossil Reptilia of the Mesozoic Formations, Part I. Pterosauria (Pterodactylus). [Gault-Lias.]. Monographs of the Palæontographical Society, 27:1-14.
Pêgas, R.V. 2024. A taxonomic note on the tapejarid pterosaurs from the Pterosaur Graveyard Site (Caiuá Group, ?Early Cretaceous of Southern Brazil): evidence for the presence of two species. Historical Biology, 2024:1-22.
https://doi.org/10.1080/08912963.2024.2355664
Pêgas, R.V., Holgado, B., and Leal, M.E.C. 2019. On Targaryendraco wiedenrothi gen. nov. (Pterodactyloidea, Pteranodontoidea, Lanceodontia) and recognition of a new cosmopolitan lineage of Cretaceous toothed pterodactyloids. Historical Biology, 33(8):1266-1280.
https://doi.org/10.1080/08912963.2019.1690482
Pêgas, R.V., Costa, F.R., and Kellner, A.W. 2021. Reconstruction of the adductor chamber and predicted bite force in pterodactyloids (Pterosauria). Zoological Journal of the Linnean Society, 193(2):602-635.
https://doi.org/10.1093/zoolinnean/zlaa163
Pêgas, R.V. and Costa, F.R. 2022. Synonymy between Brasileodactylus araripensis and Cearadactylus atrox (Pterosauria, Anhangueridae; Aptian, Romualdo Formation). Conference Proceedings of the 7th Latin American Congress of Vertebrate Paleontology, Vitória da Conquista, Brazil, p. 546.
Pentland, A.H., and Poropat, S.F. 2018. Reappraisal of Mythunga camara Molnar and Thulborn, 2007 (Pterosauria, Pterodactyloidea, Anhangueria) from the upper Albian Toolebuc Formation of Queensland, Australia. Cretaceous Research, 93:151-169.
https://doi.org/10.1016/j.cretres.2018.09.011
Pentland, A.H., Poropat, S.F., Tischler, T.R., Sloan, T., Elliott, R.A., Elliott, H.A., Elliot, J.A., and Elliott, D.A. 2019. Ferrodraco lentoni gen. et sp. nov., a new ornithocheirid pterosaur from the Winton Formation (Cenomanian-lower Turonian) of Queensland, Australia. Scientific Reports, 9(1):13454.
https://doi.org/10.1038/s41598-019-49789-4
Pentland, A.H., Poropat, S.F., White, M.A., Rigby, S.L., Bevitt, J.J., Duncan, R.J., Sloan, T., Elliot, R., Elliot, H.A., Elliott, J.A., and Elliott, D.A. 2022. The osteology of Ferrodraco lentoni, an anhanguerid pterosaur from the mid-Cretaceous of Australia. Journal of Vertebrate Paleontology, 41(5):e2038182.
https://doi.org/10.1080/02724634.2021.2038182
Pentland, A.H., Poropat, S.F., Duncan, R.J., Kellner, A.W., Bantim, R.A., Bevitt, J.J., Tait, A.M., and Grice, K. 2024. Haliskia peterseni, a new anhanguerian pterosaur from the late Early Cretaceous of Australia. Scientific Reports, 14(1):e11789.
https://doi.org/10.1038/s41598-024-60889-8
Pinheiro, F.L. and Rodrigues, T. 2017. Anhanguera taxonomy revisited: is our understanding of Santana Group pterosaur diversity biased by poor biological and stratigraphic control?. PeerJ, 5:e3285.
https://doi.org/10.7717/peerj.3285
Plieninger, F. 1901. Beiträge zur Kenntnis der Flugsaurier. Palaeontographica, 48:65-90.
Richards, T.M., Stumkat, P.E., and Salisbury, S.W. 2021. A new species of crested pterosaur (Pterodactyloidea, Anhangueridae) from the Lower Cretaceous (upper Albian) of Richmond, north west Queensland, Australia. Journal of Vertebrate Paleontology, 41(3):e1946068.
https://doi.org/10.1080/02724634.2021.1946068
Rodrigues, T., and Kellner, A.W.A. 2013. Taxonomic review of the Ornithocheirus complex (Pterosauria) from the Cretaceous of England. ZooKeys, (308):1.
https://doi.org/10.3897/zookeys.308.5559
Silva, J.L., Bentes, T., Pêgas, R.V., Kellner, A.W., and Costa, F.R. 2024. A new anhanguerid pterosaur specimen from the Romualdo Formation, Araripe Basin (Lower Cretaceous, Northeastern Brazil). Historical Biology, 2024:1-22.
https://doi.org/10.1080/08912963.2024.2372650
Souza, L.G., Pêgas, R.V., Lacerda, M.B., and Riff, D. 2023. Tales of Long Faces: Piscivorous Archosauriformes and the Evolutionary Ways to Form a Fisher. In Ballard, H.W. and Farlow, J.O.O. (eds.), Ruling Reptiles: Crocodylian Biology and Archosaur Paleobiology. Indiana Press, Bloomington.
https://doi.org/10.2307/jj.6047951.13
Steel, L., Martill, D.M., Unwin, D.M., and Winch, J.D. 2005. A new pterodactyloid pterosaur from the Wessex Formation (Lower Cretaceous) of the Isle of Wight, England. Cretaceous Research, 26(4):686-698.
https://doi.org/10.1016/j.cretres.2005.03.005
Unwin, D.M. 2003. On the phylogeny and evolutionary history of pterosaurs. In Buffetaut, E. and Mazin, J.-M. (eds.), Evolution and Palaeobiology of Pterosaurs. Geological Society, London, Special Publications, 217:139-190.
https://doi.org/10.1144/GSL.SP.2003.217.01
Veldmeijer, A.J. 2003. Description of Coloborhynchus spielbergi sp. nov. (Pterodactyloidea) from the Albian (Lower Cretaceous) of Brazil. Scripta Geologica, (125):35-139.
Vidovic, S.U. and Martill, D.M. 2018. The taxonomy and phylogeny of Diopecephalus kochi (Wagner, 1837) and ‘Germanodactylus rhamphastinus’ (Wagner, 1851). Geological Society, London, Special Publications, 455(1):125-147.
https://doi.org/10.1144/SP455.12
Wang, X. and Zhou, Z. 2003. Two new pterodactyloid pterosaurs from the Early Cretaceous Jiufotang Formation of western Liaoning, China. Vertebrata PalAsiatica, 41(01):34.
https://www.vertpala.ac.cn/EN/Y2003/V41/I01/34
Wang, X., Kellner, A.W.A., Jiang, S., and Cheng, X. 2012. New toothed flying reptile from Asia: close similarities between early Cretaceous pterosaur faunas from China and Brazil. Naturwissenschaften, 99:249-257.
https://doi.org/10.1007/s00114-012-0889-1
Wang, X., Kellner, A.W.A., Jiang, S., Wang, Q., Ma, Y., Paidoula, Y., Cheng, X., Rodrigues, T., Meng, X., Zhang, J., and Li, N. 2014. Sexually dimorphic tridimensionally preserved pterosaurs and their eggs from China. Current Biology, 24:1323-1330.
https://doi.org/10.1016/j.cub.2014.04.054
Wellnhofer P. 1985. Neue Pterosaurier aus der Santana-Formation (Apt) der Chapada do Araripe, Brasilien. Palaeontographica. Abteilung A, Paläozoologie, Stratigraphie, 187(4-6):105-182.
Wellnhofer, P. 1987. New crested pterosaurs from the Lower Cretaceous of Brazil. Mitteilungen der Bayerischen Staatssammlung für Palaontologie und Historische Geologie, 27:175-186.
https://doi.org/10.1144/GSL.SP.2003.217.01.05
Wild, R. 1990. Ein Flugsaurierrest (Reptilia, Pterosauria) aus der Unterkreide (Hauterive) von Hannover (Niedersachsen). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 181:241-254
https://doi.org/10.1127/njgpa/181/1990/241
Young, M.T., Wilberg, E.W., Johnson, M.M., Herrera, Y., de Andrade, M.B., Brignon, A., Abel, P., Foffa, D., Fernández, M.S., Vignaud, P., Cowgill, T., and Brusatte, S.L. 2024. The history, systematics, and nomenclature of Thalattosuchia (Archosauria: Crocodylomorpha). Zoological Journal of the Linnean Society, 200(2):547-617.
https://doi.org/10.1093/zoolinnean/zlad165
Zhou, X., Ikegami, N., Pêgas, R.V., Yoshinaga, T., Sato, T., Mukunoki, T., Otani, J., and Kobayashi, Y. 2025. Reassessment of an azhdarchid pterosaur specimen from the Mifune Group, Upper Cretaceous of Japan. Cretaceous Research, 167:e106046.
https://doi.org/10.1016/j.cretres.2024.106046
