

Volume 28.2
May–August 2025
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 28.2 May-August 2025
See all articles in 28.1 January-April 2025
See all articles in 27.3 September-December 2024
See all articles in 27.2 May-August 2024
Interested in submitting a paper to Palaeontologia Electronica?
Click here to register and submit.
 |
||||
 |
||||
Article Search
 Ricardo Araújo
Ricardo Araújo
Huffington Department of Earth Sciences
Southern Methodist University
Dallas, Texas 75275, USA;
Museu da Lourinhã
Rua João de Luís de Moura, 95
2530-158 Lourinhã
Portugal
and Instituto de Plasmas e Fusão Nuclear
Instituto Superior Técnico
Universidade de Lisboa
Portugal
and Museum für Naturkunde
Berlin
Germany
rmaraujo@smu.edu
Ricardo Araújo has completed his doctoral degree at the Southern Methodist University. He is currently a postdoctoral fellow at the Plasma and Nuclear Fusion Institute in the University of Lisbon and at the Museum für Naturkunde studying synchrotron techniques applied to paleontology. The doctoral dissertation focuses on Angolan plesiosaurs, functional morphology, and myological reconstruction in plesiosaurs. He is part of the PaleoAngola project, an international collaborative endeavor that aims to study and collect the vast Angolan paleontological heritage. Ricardo is also a co-leader of the PalNiassa project, a Portuguese-Mozambican cooperation that aims to trigger research and increase dynamism in Mozambican academia, by means of an invaluably important, diverse and undiscovered fossil record. Ricardo also actively collaborates with the Museu da Lourinhã, Portugal, on dinosaur eggs and embryos.

 Fernando Correia
Fernando Correia
Departamento de Biologia
Universidade de Aveiro
Campus Santiago
3810-193 Aveiro
Portugal
fjorgescorreia@sapo.pt
Fernando J.S. Correia (Portugal) is a biologist holding a MS degree in Animal Ecology. He is specialized in Scientific Illustration and Communication. Correia is an Assistant Lecturer in the Department of Biology at the University of Aveiro, he was responsible for the re-introduction of Scientific Illustration discipline in the Portuguese University-level Education. He was also a pioneer in digital techniques applicable to the scientific illustration field in Portugal (1998). Correia is currently the head of the Scientific Illustration Laboratory where he teaches and is responsible for the overall coordination of the Training Course in Scientific Illustration and the Exhibition ExpoDBIO Gallery. He is responsible for more than 75 intensive Science Illustration courses (Portugal and abroad), and was co-founder and first President of GNSI (Guild of Natural Science Illustrators) Chapter - Portugal. He has written more than 85 publications as single authorship including "Mealhada - natures unique" (2011) and "Paleoart - fossils that come to life" (2012) and 30 as co-author, including articles and books, both national and international editions. Correia is an active member of the Association Européenne des Illustrateurs Médicaux et Scientifiques AEIMS (Europe) and the Guild of Natural Science Illustrators / GNSI (USA). He was one of four Portuguese invited to participate in the renowned book "The Guild Handbook of Scientific Illustration" (Elaine Hodges, 2nd ed., 2003).
Correia has participated in more than 70 exhibitions worldwide. His artwork has been honored with several awards and has been part of private and institutional collections worldwide. To view more of his work, visit www.efecorreia-artstudio.com and www.ilustracao-cientifica-uaveiro.com/
FIGURE 1. Three topological hypotheses for the evolution of the pectoral girdle elements from the basal neodiapsid condition to basal eosauropterygian condition are depicted in two-dimensions. Large black dot - glenoid; Small black dot - coracoid foramen; A- anterior margin of the coracoid; M-medial margin of the coracoid; CL - clavicle; INCL - interclavicle; SC - scapula; ST - sternum.
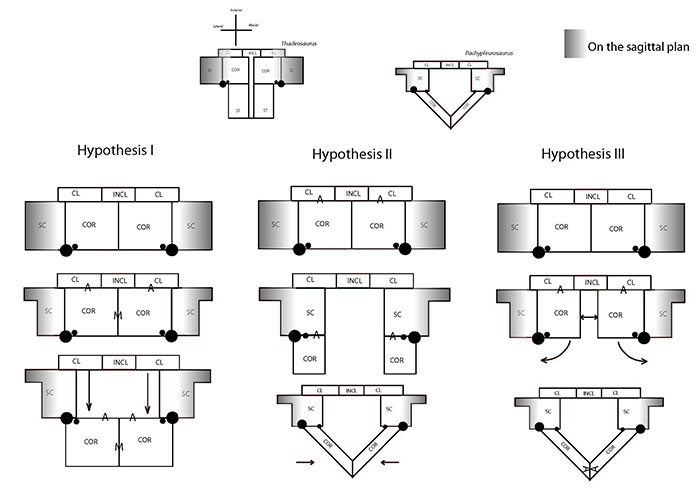
FIGURE 2. Topological homologies underlying the proposed muscle reconstructions, using as a model the nothosaur Ceresiosaurus calcagnii PIMUZ T3983 (left) and the basal plesiosaur Meyerasaurus victor cast of SMNS12478 (right).
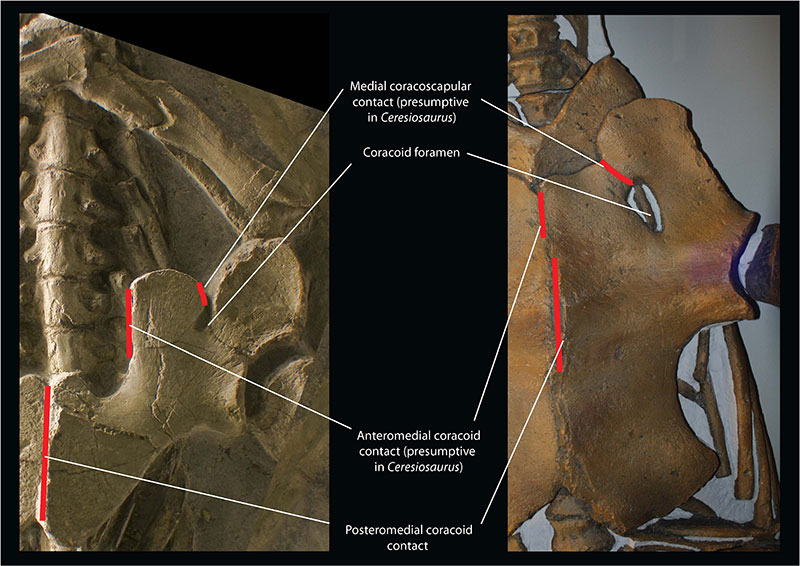
FIGURE 3. Muscle reconstruction of the pectoral girdle musculature of the Eosauropterygia using data from the extant phylogenetic bracket, the fossil record and developmental patterns.
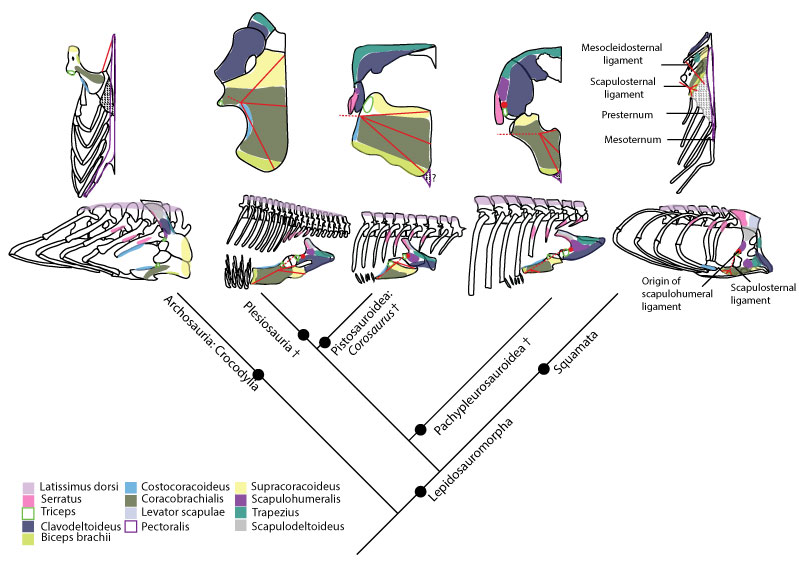
FIGURE 4. Humeral insertions in a basal neodiapsid, Nothosaurus and Cryptoclidus .
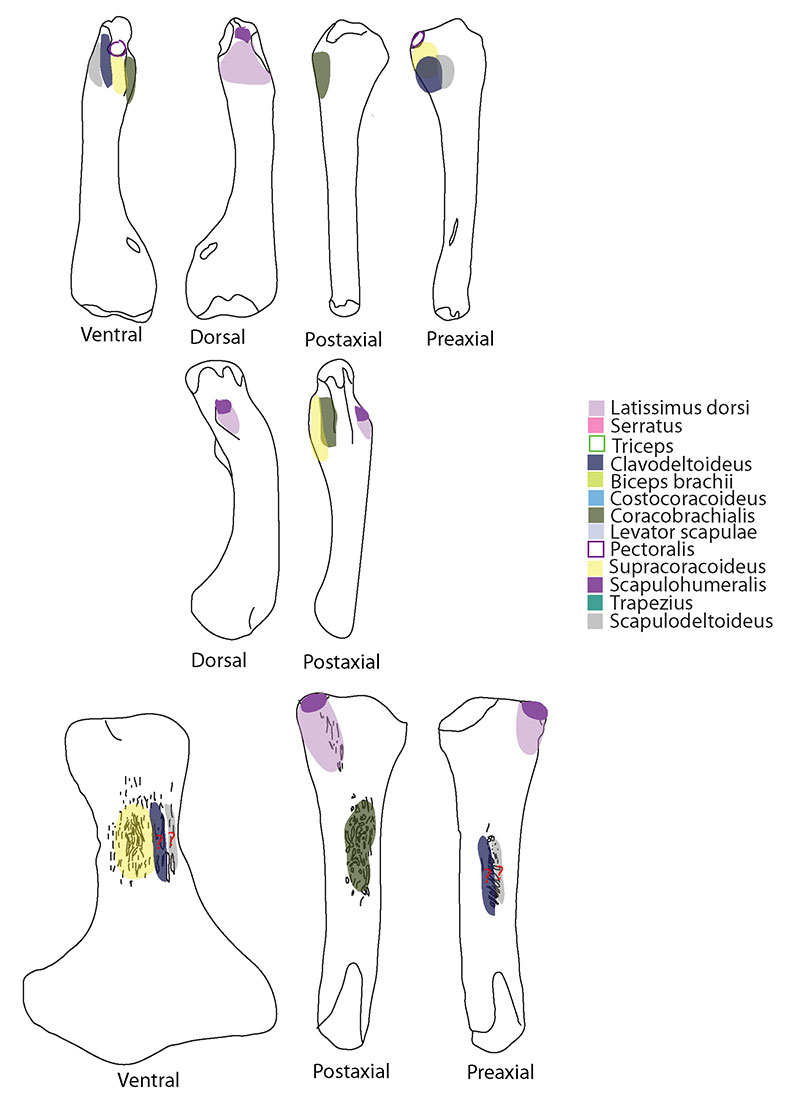
FIGURE 5. M. subcoracoscapularis. Reconstruction for Neusticosaurus 1 in anterior view, 2 in ventral view, 3 in lateral view, 4 in fleshed-out muscle reconstruction and 5 complete skeletal reconstruction in lateral view. Reconstruction for Ceresiosaurus 6 in anterior view, 7 in ventral view, 8 in lateral view, 9 fleshed-out muscle reconstruction and 10 complete skeletal reconstruction in lateral view. Reconstruction for Rhomaleosaurus 11 in anterior view, 12 in ventral view, 13 in lateral view, 14 fleshed-out muscle reconstruction and 15 complete skeletal reconstruction in lateral view.
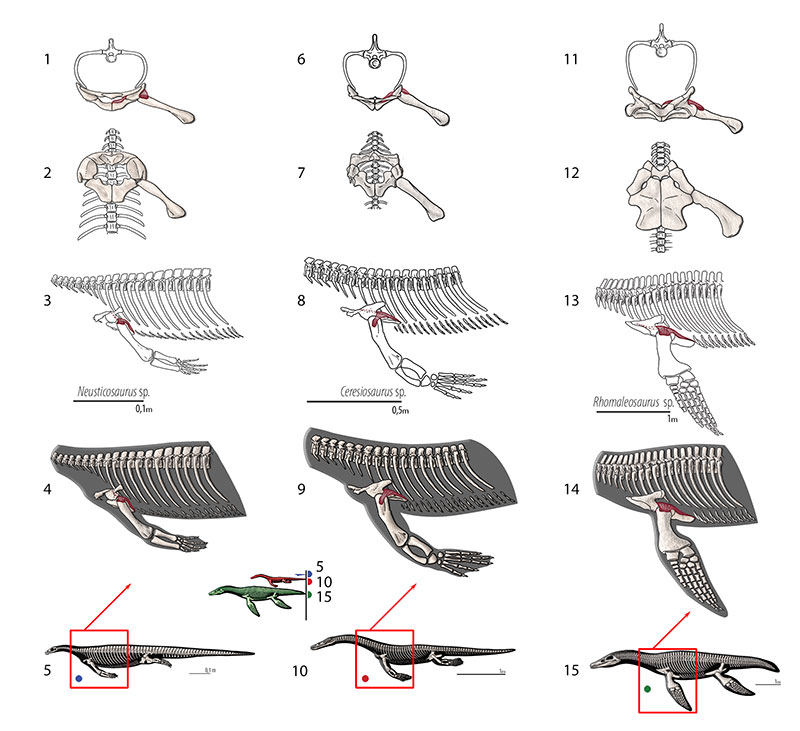
FIGURE 6. M. scapulodeltoideus (= m. deltoideus scapularis, = m. dorsalis scapulae, = m. teres major. Reconstruction for Neusticosaurus 1 in anterior view, 2 in ventral view, 3 in lateral view, 4 fleshed-out muscle reconstruction and 5 complete skeletal reconstruction in lateral view. Reconstruction for Ceresiosaurus 6 in anterior view, 7 in ventral view, 8 in lateral view, 9 fleshed-out muscle reconstruction 10 complete skeletal reconstruction in lateral view. Reconstruction for Rhomaleosaurus and 11 in anterior view, 12 in ventral view, 13 in lateral view, 14 fleshed-out muscle reconstruction and 15 complete skeletal reconstruction in lateral view.
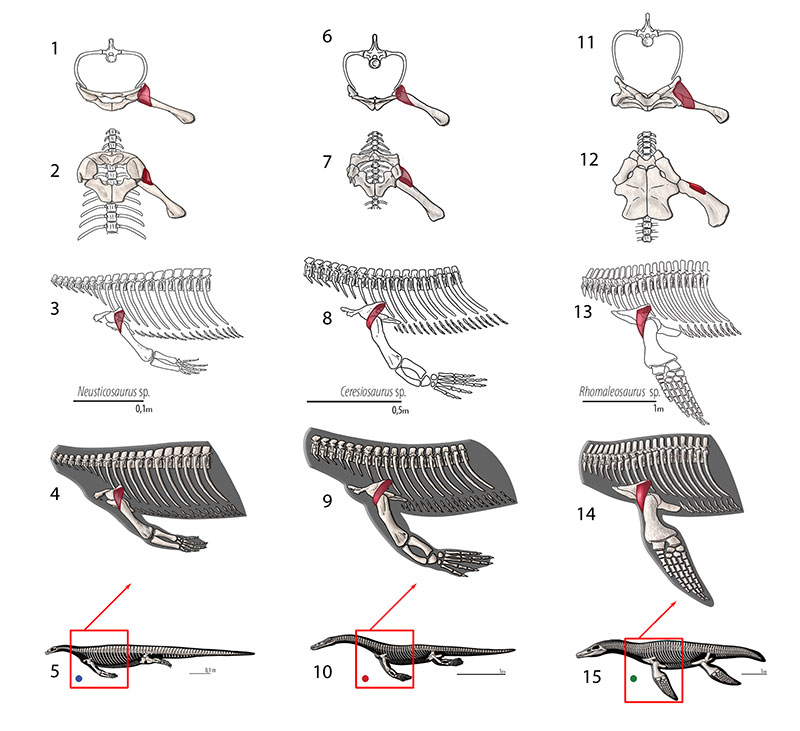
FIGURE 7. M. clavodeltoideus (= m. deltoideus clavicularis, = m. deltoideus). Reconstruction for Neusticosaurus 1 in anterior view, 2 in ventral view, 3 in lateral view, 4 fleshed-out muscle reconstruction and 5 complete skeletal reconstruction in lateral view. Reconstruction for Ceresiosaurus 6 in anterior view, 7 in ventral view, 8 in lateral view, 9 fleshed-out muscle reconstruction and 10 complete skeletal reconstruction in lateral view. Reconstruction for Rhomaleosaurus 11 in anterior view, 12 in ventral view, 13 in lateral view, 14 fleshed-out muscle reconstruction and 15 complete skeletal reconstruction in lateral view.
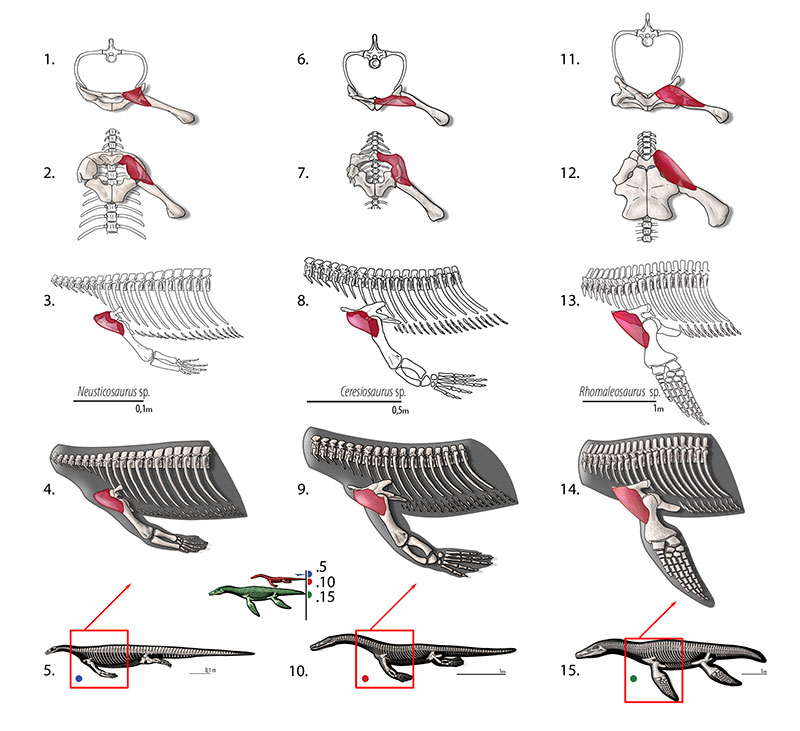
FIGURE 8. M. scapulohumeralis (= m. scapulohumeralis caudalis, = m. scapulohumeralis posterior). Reconstruction for Neusticosaurus 1 in anterior view, 2 in ventral view, 3 in lateral view, 4 fleshed-out muscle reconstruction and 5 complete skeletal reconstruction in lateral view. Reconstruction for Ceresiosaurus 6 in anterior view, 7 in ventral view, 8 in lateral view, 9 fleshed-out muscle reconstruction and 10 complete skeletal reconstruction in lateral view. Reconstruction for Rhomaleosaurus 11 in anterior view, 12 in ventral view, 13 in lateral view, 14 fleshed-out muscle reconstruction and 15 complete skeletal reconstruction in lateral view.
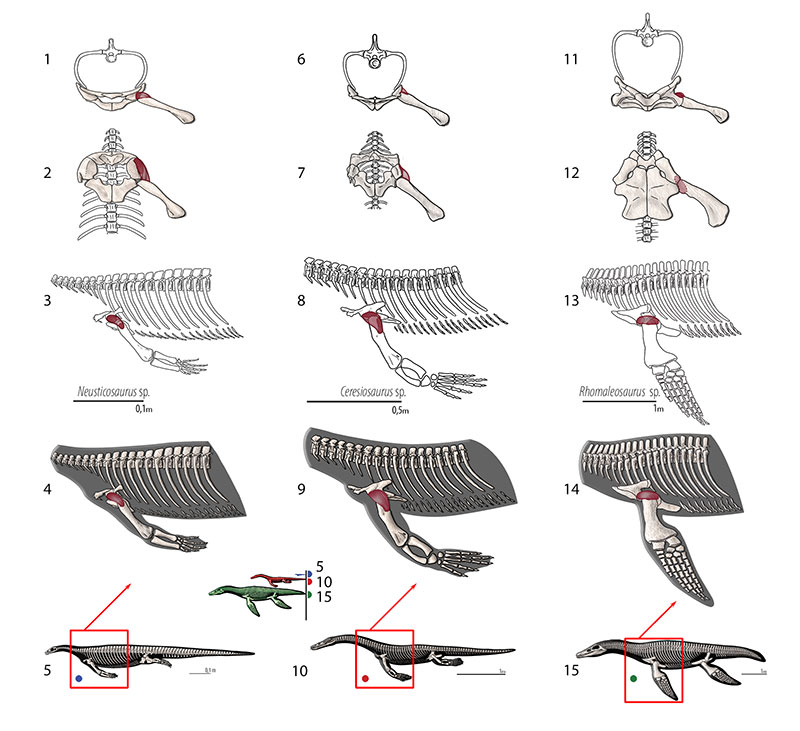
FIGURE 9. M. supracoracoideus (= m. coracobrachialis superior, = m. coracobrachialis brevis dorsalis). Reconstruction for Neusticosaurus 1 in anterior view, 2 in ventral view, 3 in lateral view, 4 fleshed-out muscle reconstruction and 5 complete skeletal reconstruction in lateral view. Reconstruction for Ceresiosaurus 6 in anterior view, 7 in ventral view, 8 in lateral view, 9 fleshed-out muscle reconstruction and 10 complete skeletal reconstruction in lateral view. Reconstruction for Rhomaleosaurus 11 in anterior view, 12 in ventral view, 13 in lateral view, 14 fleshed-out muscle reconstruction and 15 complete skeletal reconstruction in lateral view.
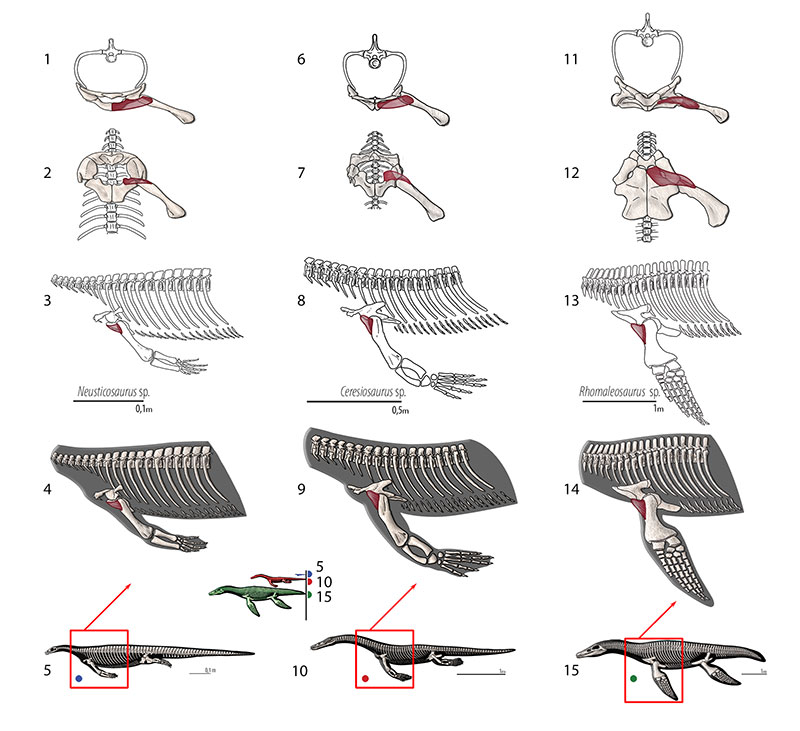
FIGURE 10. M. coracobrachialis longus and brevis. Reconstruction for Neusticosaurus 1 in anterior view, 2 in ventral view, 3 in lateral view, 4 fleshed-out muscle reconstruction and 5 complete skeletal reconstruction in lateral view. Reconstruction for Ceresiosaurus 6 in anterior view, 7 in ventral view, 8 in lateral view, 9 fleshed-out muscle reconstruction and 10 complete skeletal reconstruction in lateral view. Reconstruction for Rhomaleosaurus 11 in anterior view, 12 in ventral view, 13 in lateral view, 14 fleshed-out muscle reconstruction and 15 complete skeletal reconstruction in lateral view.
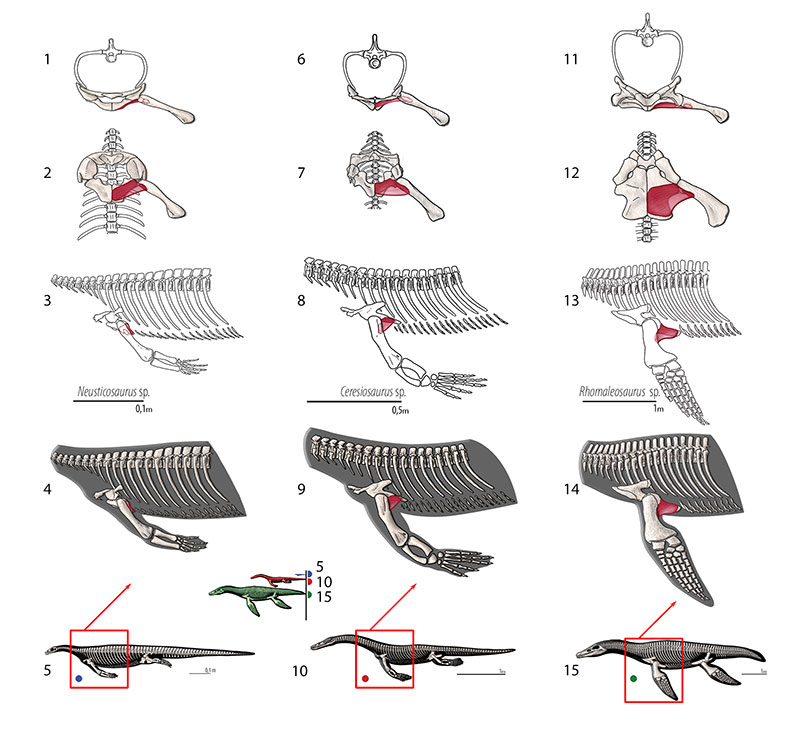
FIGURE 11. M. latissimus dorsi. Reconstruction for Neusticosaurus 1 in anterior view, 2 in ventral view, 3 in lateral view, 4 fleshed-out muscle reconstruction and 5 complete skeletal reconstruction in lateral view. Reconstruction for Ceresiosaurus 6 in anterior view, 7 in ventral view, 8 in lateral view, 9 fleshed-out muscle reconstruction and 10 complete skeletal reconstruction in lateral view. Reconstruction for Rhomaleosaurus 11 in anterior view, 12 in ventral view, 13 in lateral view, 14 fleshed-out muscle reconstruction and 15 complete skeletal reconstruction in lateral view.
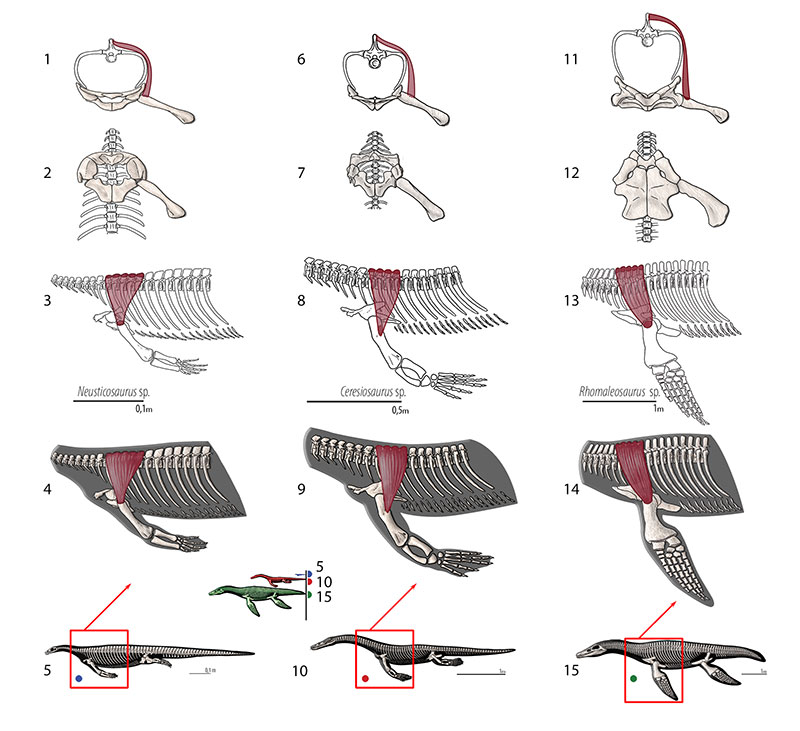
FIGURE 12. M. triceps brachii. Reconstruction for Neusticosaurus 1 in ventral view, 2 in lateral view, 3 fleshed-out muscle reconstruction and 4 complete skeletal reconstruction in lateral view. Reconstruction for Ceresiosaurus 5 in ventral view, 6 in lateral view, 7 fleshed-out muscle reconstruction and 8 complete skeletal reconstruction in lateral view. Reconstruction for Rhomaleosaurus 9 in ventral view, 10 in lateral view, 11 fleshed-out muscle reconstruction and 12 complete skeletal reconstruction in lateral view.
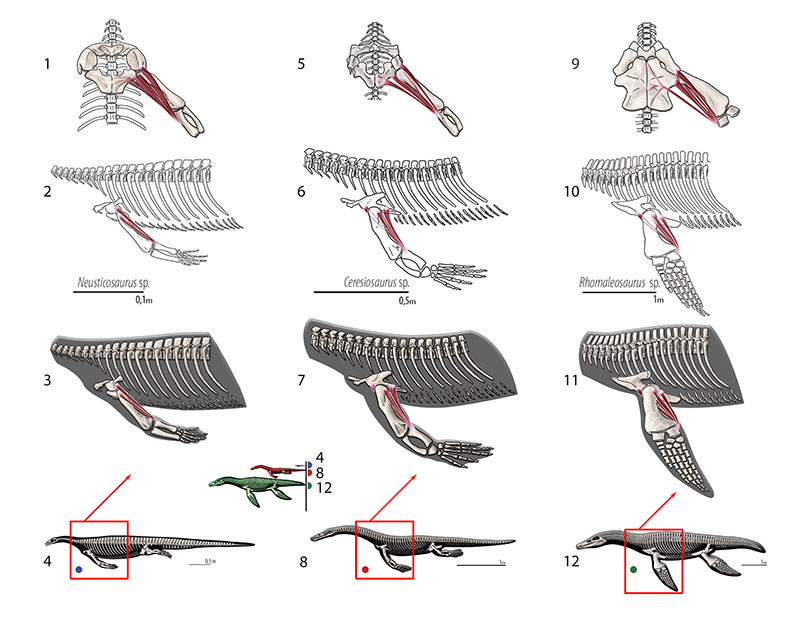
FIGURE 13. M. trapezius . Reconstruction for Neusticosaurus 1 in anterior view, 2 in ventral view, 3 in lateral view, 4 fleshed-out muscle reconstruction and 5 complete skeletal reconstruction in lateral view. Reconstruction for Ceresiosaurus 6 in anterior view, 7 in ventral view, 8 in lateral view, 9 fleshed-out muscle reconstruction 10 complete skeletal reconstruction in lateral view. Reconstruction for Rhomaleosaurus 11 in anterior view, 12 in ventral view, 13 in lateral view, 14 fleshed-out muscle reconstruction and 15 complete skeletal reconstruction in lateral view.
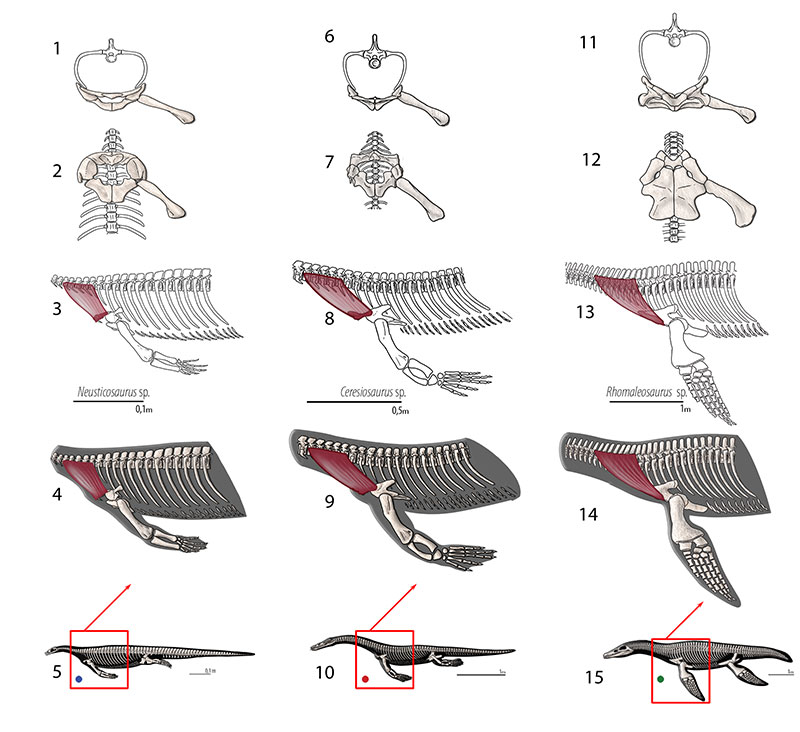
FIGURE 14. M. serratus. Reconstruction for Neusticosaurus 1 in anterior view, 2 in ventral view, 3 in lateral view (orange for Level II inference), 4 fleshed-out muscle reconstruction and 5 complete skeletal reconstruction in lateral view. Reconstruction for Ceresiosaurus 6 in anterior view, 7 in ventral view, 8 in lateral view, 9 fleshed-out muscle reconstruction and 10 complete skeletal reconstruction in lateral view. Reconstruction for Rhomaleosaurus 11 in anterior view, 12 in ventral view, 13 in lateral view, 14 fleshed-out muscle reconstruction and 15 complete skeletal reconstruction in lateral view.
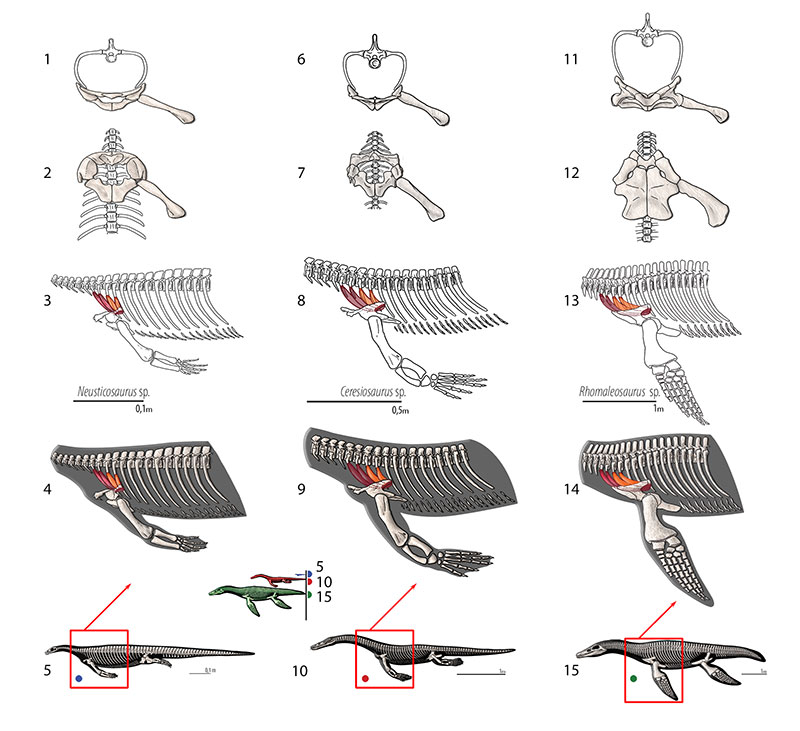
FIGURE 15. M. levator scapulae. Reconstruction for Neusticosaurus 1 in anterior view, 2 in ventral view, 3 in lateral view, 4 fleshed-out muscle reconstruction and 5 complete skeletal reconstruction in lateral view. Reconstruction for Ceresiosaurus 6 in anterior view, 7 in ventral view, 8 in lateral view, 9 fleshed-out muscle reconstruction and 10 complete skeletal reconstruction in lateral view. Reconstruction for Rhomaleosaurus 11 in anterior view, 12 in ventral view, 13 in lateral view, 14 fleshed-out muscle reconstruction and 15 complete skeletal reconstruction in lateral view.
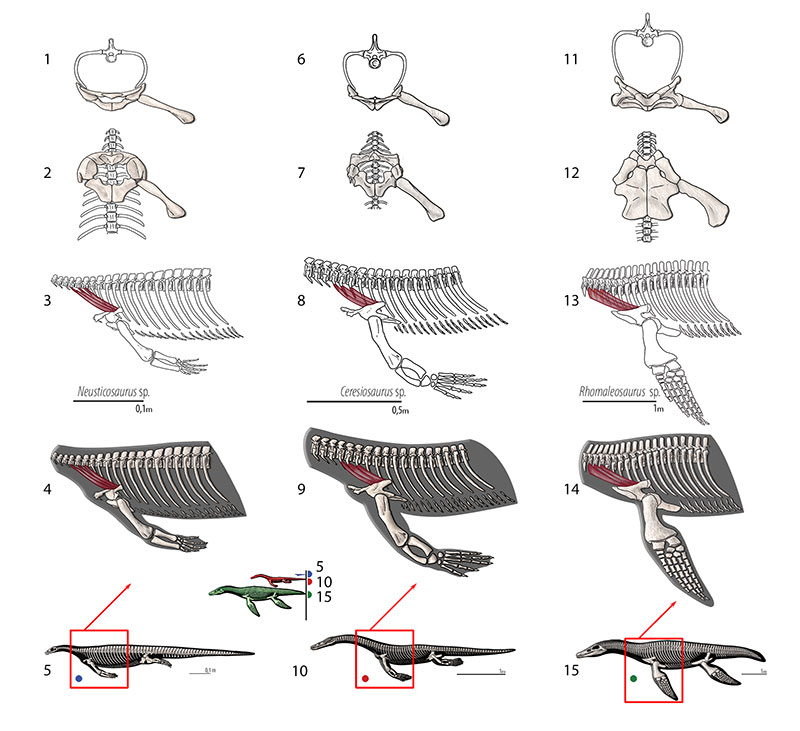
FIGURE 16. M. costocoracoideus. Reconstruction for Neusticosaurus 1 in anterior view, 2 in ventral view, 3 in lateral view, 4 fleshed-out muscle reconstruction and 5 complete skeletal reconstruction in lateral view. Reconstruction for Ceresiosaurus 6 in anterior view, 7 in ventral view, 8 in lateral view, 9 fleshed-out muscle reconstruction and 10 complete skeletal reconstruction in lateral view. Reconstruction for Rhomaleosaurus 11 in anterior view, 12 in ventral view, 13 in lateral view, 14 fleshed-out muscle reconstruction and 15 complete skeletal reconstruction in lateral view.
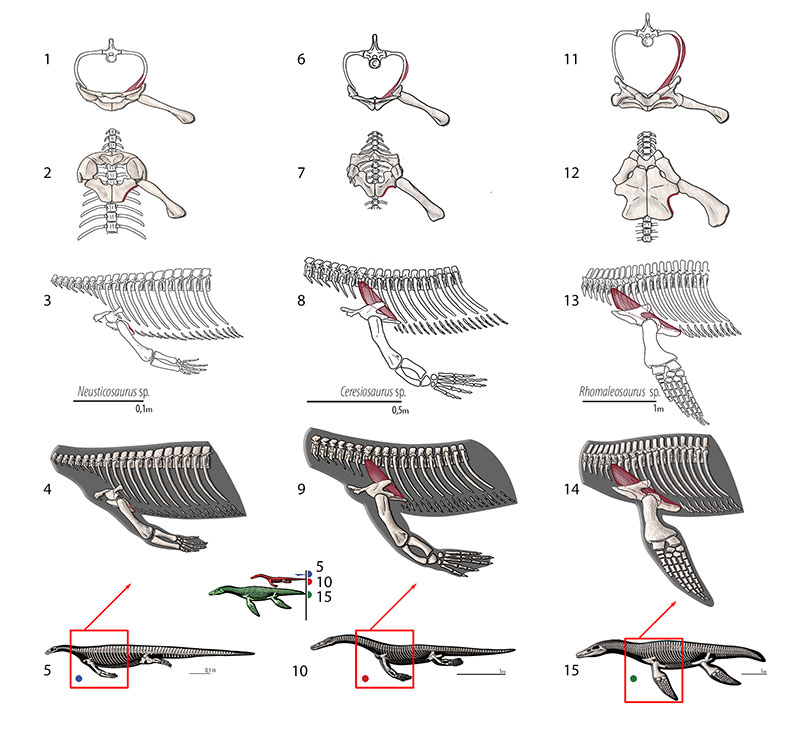
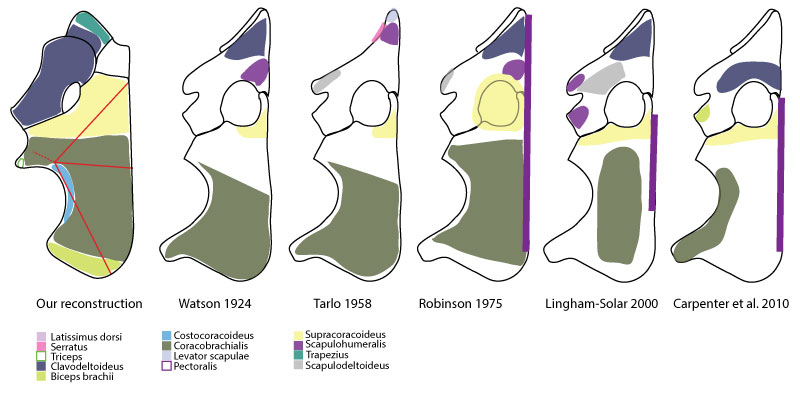
FIGURE 17. Comparison of pectoral girdle musculature reconstruction between this study and previously published reconstructions.
Soft-tissue anatomy of the Plesiosaur pectoral girdle inferred from basal Eosauropterygia taxa and the extant phylogenetic bracket
Plain Language Abstract
Typically to reconstruct soft-tissues (e.g., muscles) in extinct forms it is necessary to compare this tissues with closely related taxa (extant phylogenetic bracket) and assess their osteological correlates (marks of those soft-tissues left on bones). However, that is not enough for plesiosaurians. When plesiosaurians appeared in the fossil record they are already so adapted to a marine lifestyle that it becomes hard to identify the same structures in their terrestrial extant analogues. Therefore, when evolution modified significantly certain structures (such as the pectoral girdle in plesiosaurians, i.e., the bones where the forearm muscles attach to) it becomes hard to establish correct homologous parts. In the case of plesiosaurians due to the adaptation to a marine lifestyle given that their diapsid ancestors were terrestrial. Thus, additionally, we had to look at developmental data and also to several plesiosaurian related taxa (i.e., pachypleurosaurs, nothosaurs). The resulting pectoral girdle muscle reconstructions are significantly different from previous attempts.
Resumen en Español
Anatomía de los tejidos blandos de la cintura escapular de plesiosaurios inferida de taxones de Eosauropterygia basales y del Soporte Filogenético Viviente
Los plesiosaurios eran organismos adaptados secundariamente altamente derivados (si consideramos que los peces están adaptados primariamente) con una larga historia evolutiva, y están estrechamente relacionados con eosauropterigios basales. Los intentos de reconstruir la anatomía de los tejidos blandos pueden ser complicados debido a la falta de especies actuales estrechamente relacionadas, por lo que deben ser consideradas varias líneas de evidencia. Este estudio tiene como objetivo reconstruir la miología de la cintura escapular de los eosauropterigios. La información derivada del método de Soporte Filogenético Viviente (EPB) no fue suficiente para aclarar las inserciones musculares en la cintura escapular de plesiosaurios. Para inferir correctamente las homologías musculares, la información proveniente del Soporte Filogenético Viviente tuvo que ser complementada con información osteológica y del desarrollo, y las transformaciones osteológicas tuvieron que remontarse hasta los neodiápsidos basales del Pérmico. La reconstrucción de la musculatura de la cintura escapular que se presenta es, por lo tanto, significativamente diferente de las propuestas anteriores. Como en los análogos actuales adaptados secundariamente al medio acuático, varios músculos atrofiados (e.g., pectoral, episternocleidomastoideo) y otros se especializaron (e.g., coracobraquial, clavodeltoideo) a fin de alcanzar un papel más relacionado con las rigurosas condiciones de la locomoción subacuática. El subcoracoscapular, escapulodeltoideo, escapulohumeral y supracoracoideo se infieren como estabilizadores glenohumerales. El clavodeltoideo actuó como el principal músculo transportador y el coracobraquial como un músculo retractor importante, posiblemente junto con los latissimus dorsi. Varias cabezas de los tríceps posiblemente se atrofiaron, como en las ballenas, sirviendo principalmente como un estabilizador de la articulación cubital. El trapecio, serrato y elevador de la escápula sirvieron como estabilizadores de la cintura escapular.
PALABRAS CLAVE: miología; Eosauropterygia; neodiápsidos basales; Soporte Filogenético Viviente; cintura escapular; plesiosaurios
Traducción: Enrique Peñalver
Résumé en Français
L'anatomie des tissus mous de la ceinture scapulaire de Plésiosaure déduit à partir de taxons de base d'Eosauropterygia et le support existant phylogénétique
Les plésiosauriens sont des organismes très dérivés secondairement adaptés (si les poissons sont principalement adaptés) avec une longue histoire évolutive, et ils sont étroitement liés avec les éosauroptérygiens basales. Les tentatives pour reconstruire l'anatomie des tissus mous peuvent être compliquées en raison de l'absence d'espèces existantes étroitement liés, ainsi diverses preuves doivent être considérées. Cette étude vise à reconstruire la myologie de la ceinture pectorale d'éosauroptérygiens. L'information tirée de la méthode de fixation phylogénétique existante n'était pas suffisante pour clarifier les attaches musculaires dans la ceinture pectorale de plésiosauriens. Pour conclure correctement les homologies musculaires, les informations de support phylogénétique existant a dû être complétées par des informations développementales et ostéologiques, et les transformations ostéologiques ont dû remonter à des néodiapsides basales du Permien. La musculature reconstruite de la ceinture pectorale présentée ici est donc sensiblement différente des précédentes tentatives. Comme dans les analogues modernes aquatiques secondairement adaptées, plusieurs muscles sont atrophiés (e.g., pectoralis, episternocleidomastoideus) et d'autres sont spécialisés (e.g., coracobrachial, clavodeltoideus) afin d'atteindre un rôle plus influent dans les conditions strictes de locomotion subaquatique. Le subcoracoscapularis, scapulodeltoideus, scapulohumeralis et supracoracoideus sont déduites d'être des stabilisateurs glénohuméraux. Le clavodeltoideus agi comme le principal muscle rapporteur et le coracobrachialis comme un grand muscle écarteur, éventuellement en collaboration avec le latissimus dorsi. Plusieurs têtes du triceps sont éventuellement atrophiées, comme dans les baleines, servant principalement comme un joint stabilisateur cubital. Le trapezius, serratus et levator scapuale servent comme stabilisant de la ceinture pectorale.
MOTS CLÉS: myologie; Eosauropterygia; néodiapsides basales; support phylogénétique existant; ceinture pectorale; plésiosaures
Translator: Kenny J. Travouillon
Deutsche Zusammenfassung
Weichteilanatomie eines Plesiosaurier-Schultergürtels erschlossen aus basalen Eosauropterygia-Taxa und dem Prinzip der phylogenetischen Umklammerung (Extant Phylogenetic Bracket)
Plesiosaurier sind hoch abgeleitete sekundär angepasste Organismen (wenn Fische primär angepasst sind) mit einer langen Evolutionsgeschichte, die eng verwandt mit basalen Eosauropterygiern sind. Ansätze zur Rekonstruktion von Weichteilanatomie sind wegen der fehlenden heute noch lebenden nahen Verwandten schwierig, daher müssen verschiedenen Blicklinien berücksichtigt werden. Diese Studie hat das Ziel die Myologie des Schultergürtels der Eosauropterygia zu untersuchen. Informationen, die mit dem Prinzip der phylogenetischen Umklammerung gewonnen worden waren reichten nicht aus um die Muskelansatzstellen im Schultergürtel von Plesiosauriern zu klären. Um auf die korrekten Muskelhomologien rückschliessen zu können, musste das Prinzip der phylogenetischen Umklammerung mit entwicklungsphysiologischer und osteologischer Information ergänzt werden und osteologische Transformationen mussten bis zu den basalen permischen Neodiapsiden zurückverfolgt werden. Die rekonstruierte Schultergürtelmuskulatur unterscheidet sich daher signifikant von früheren Ansätzen. Wie bei einigen sekundär angepassten aquatischen modernen Entsprechungen atrophierten einige Muskeln (beispielsweise Pectoralis und Episternocleidomastoideus) und andere spezialisierten sich (beispielsweise Coracobrachialis, Clavodeltoideus) um eine maßgebende Funktion innerhalb der stringenten Konditionen subaquatischer Lokomotion zu erreichen. Es wird gefolgert, dass Subcoracoscapularis, Scapulodeltoideus, Scapulohumeralis und Supracoracoideus glenohumerale Stabilisatoren waren. Der Clavodeltoideus war der Hauptprotraktor und der Coracobrachialis der Hauptretraktor, möglicherweise zusammen mit dem Latissimus dorsi. Einige Trizeps-Ansätze atrophierten möglicherweise wie bei Walen und waren damit hauptsächlich Gelenkstabilisatoren. Der Trapezius, Serratus und Levator Scapulae agierten als Brustgürtelstabilisatoren.
Schlüsselwörter: Myologie; Eosauropterygia; basale Neodiapsiden; Extant phylogenetic bracket; Schultergürtel; Plesiosaurier
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
PE: An influential journal
 Palaeontologia Electronica among the most influential palaeontological journals
Palaeontologia Electronica among the most influential palaeontological journalsArticle number: 27.2.2E
July 2024

 A Review of Handbook of Paleoichthyology Volume 8a: Actinopterygii I, Palaeoniscimorpha, Stem Neopterygii, Chondrostei
A Review of Handbook of Paleoichthyology Volume 8a: Actinopterygii I, Palaeoniscimorpha, Stem Neopterygii, Chondrostei