

Volume 28.2
May–August 2025
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 28.2 May-August 2025
See all articles in 28.1 January-April 2025
See all articles in 27.3 September-December 2024
See all articles in 27.2 May-August 2024
Interested in submitting a paper to Palaeontologia Electronica?
Click here to register and submit.
 |
||||
 |
||||
Article Search
 Adrian Marciszak. Department of Paleozoology, Institute of Environmental Biology, Faculty of Biological Sciences, University of Wrocław, Sienkiewicza 21, 50-335 Wrocław, Poland. adrian.marciszak@uwr.edu.pl
Adrian Marciszak. Department of Paleozoology, Institute of Environmental Biology, Faculty of Biological Sciences, University of Wrocław, Sienkiewicza 21, 50-335 Wrocław, Poland. adrian.marciszak@uwr.edu.pl
Adrian Marciszak, born in 1983, is married, has one child. In 2007 completed master thesis entitled "Carnivore mammals (Carnivora, Mammalia) from Żabia Cave". Recently focused on Tertiary carnivore mammals, mostly from territory of Central Europe, but also from Spain and Russia.

 Wiktoria Gornig. Department of Paleozoology, Institute of Environmental Biology, Faculty of Biological Sciences, University of Wrocław, Sienkiewicza 21, 50-335 Wrocław, Poland. wiktoria.gornig@uwr.edu.pl
Wiktoria Gornig. Department of Paleozoology, Institute of Environmental Biology, Faculty of Biological Sciences, University of Wrocław, Sienkiewicza 21, 50-335 Wrocław, Poland. wiktoria.gornig@uwr.edu.pl
Wiktoria Gornig was born in 2014 completed master thesis entitled "Steppe brown bear Ursus arctos priscus Goldfuss, 1818 from Niedźwiedzia Cave in Kletno". Recently she focused on the Sudeten speleoid ursids as an PhD student on the University of Wrocław. Her interests are focused on mammals, especially fossil carnivores, including also recent species. She is also involved in the performance of graphic design, web pages, as well as graphic design of scientific and popular science publications.
 Krzysztof Stefaniak. Department of Paleozoology, Institute of Environmental Biology, Faculty of Biological Sciences, University of Wrocław, Sienkiewicza 21, 50-335 Wrocław, Poland. krzysztof.stefaniak@uwr.edu.pl
Krzysztof Stefaniak. Department of Paleozoology, Institute of Environmental Biology, Faculty of Biological Sciences, University of Wrocław, Sienkiewicza 21, 50-335 Wrocław, Poland. krzysztof.stefaniak@uwr.edu.pl
Krzysztof Stefaniak graduated from the Wroclaw University, Poland in 1986 and received his PhD in 2001. He has been working at the Department of Palaeozoology, Zoological Institute, University of Wroclaw, Poland since 1984. His interests are focused on fossil mammals, especially ungulates, including Neogene and Quaternary deer. His research activity includes also archaeozoology, caves and fossil DNA. He has participated in excavations in karst areas of Poland and the Ukraine for 30 years. He lectures palaeontology, archaeozoology and the history of the formation of the modern fauna and flora during the Quaternary at the University of Wroclaw.
FIGURE 1. Location of Solna Jama Cave in Europe indicated by red star ( 1), in Poland marked by red square ( 2), in the environs of Gniewoszów village indicated by red star ( 3) and map of the site ( 4) after Pulina (1996) and Stefaniak et al. (2009).
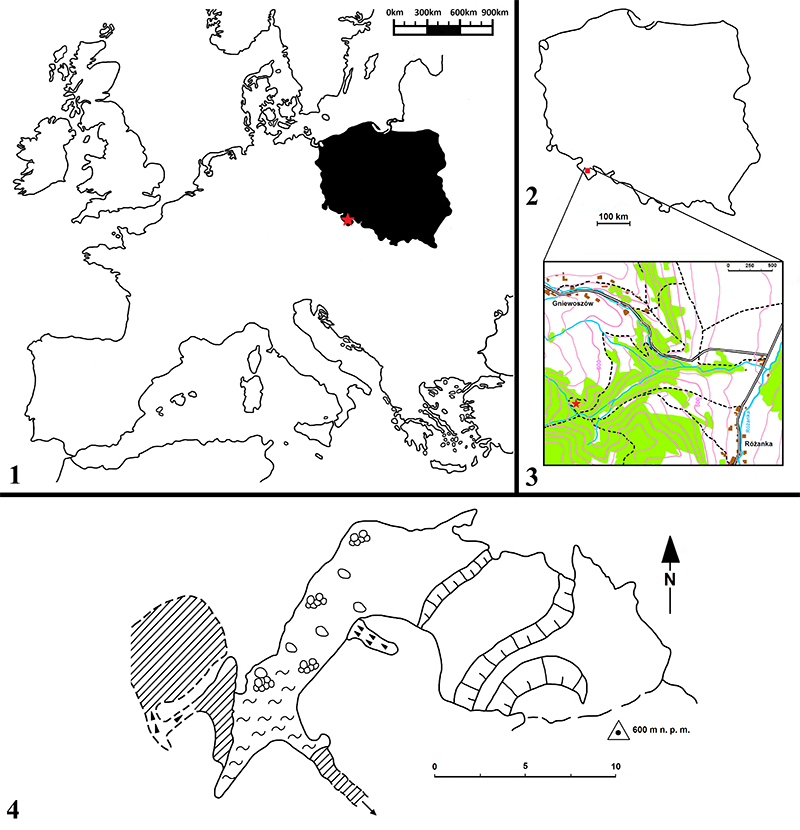
FIGURE 2. Environs of Solna Jama Cave entrance ( 1), showing the fissured crystalline limestones, in which the cave developed. The “Wolverine Niche” ( 2) and the corrosion, phreatic features of the cave passage (3). The “Wolverine Niche” is the place in the cave, where partially preserved skeleton of particularly large Gulo gulo was found (Photo by K. Stefaniak).

FIGURE 3. Ursus arctos priscus m2 (JSJ/Ua/1) from Solna Jama Cave in occlusal view. Note robust build and arctoid shape of tooth, with only slightly expanded, buccal margin of the talonid. Scale bar equals 10 mm.

FIGURE 4. Gulo gulo cranium from Solna Jama Cave (JSJ/Gg/1-1) in dorsal view (left) compared with a cranium from a large modern male from Scandinavia (right) from collection of Natural History Museum University of Wrocław (coll. no. M/500328). Scale bar equals 10 mm.

FIGURE 5. Gulo gulo cranium from Solna Jama Cave (JSJ/Gg/1-1) in ventral view (left) compared with a cranium from a large modern male from Scandinavia (right) from collection of Natural History Museum University of Wrocław (coll. no. M/500328). Note particularly powerful dentition of the individual from Solna Jama Cave in comparison with recent G . gulo. Scale bar equals 10 mm.
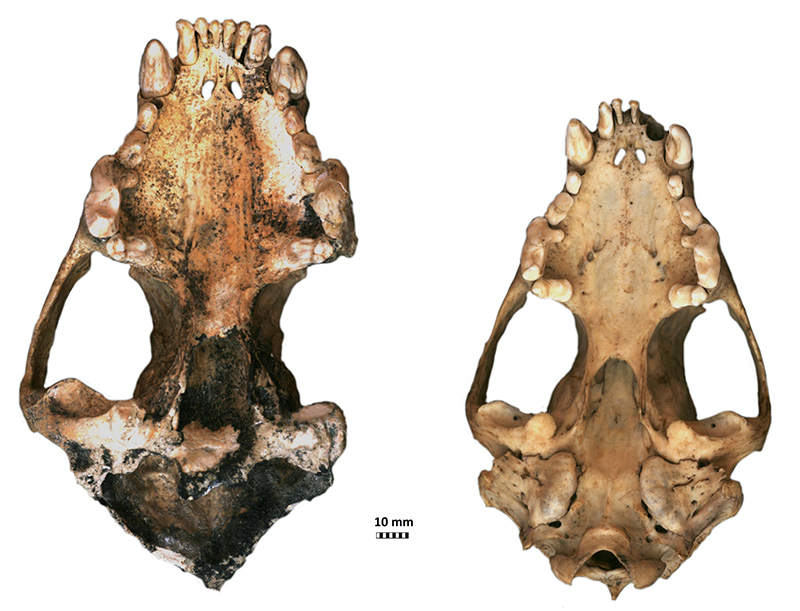
FIGURE 6. Metacarpals (mtcp) and metatarsals (mtts) of Gulo gulo from Solna Jama Cave. From left to right: right mtcp III (JSJ/Gg/1-29), left mtcp V (JSJ/GG/1-31), left mtts II (JSJ/Gg/1-33), left mtts III (JSJ/Gg/1-34), left mtts IV (JSJ/Gg/1-35) and right mtts V (JSJ/Gg/1-36). Scale bar equals 10 mm.
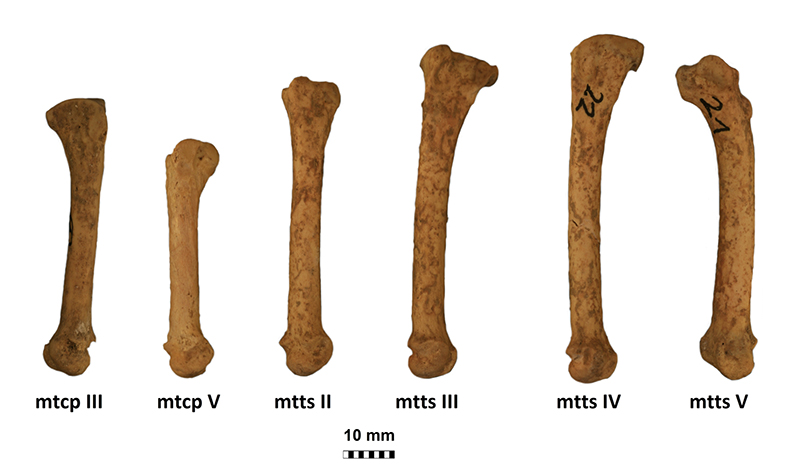
FIGURE 7. Scatter diagram showing the ratio of m1 trigonid breadth (B tri) to m1 talonid breadth (B tal) in the forms of Gulo : G. schlosseri and G. gulo. Data from Döppes (2001): late Pleistocene G. gulo and recent G. gulo ; data from Marciszak (2012): G. schlosseri and late Pleistocene G. gulo (part).
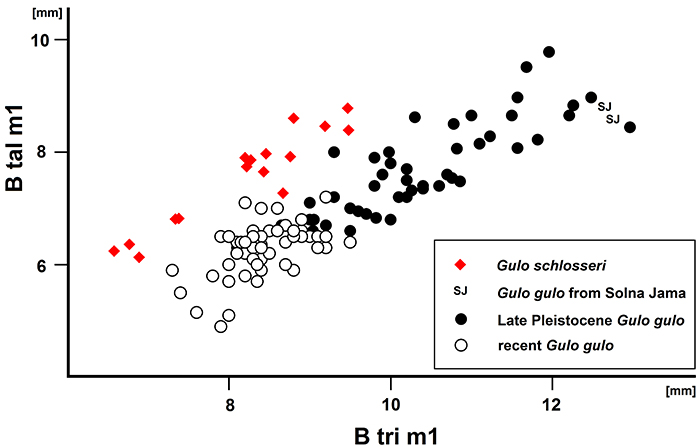
FIGURE 8. Occurrence of members of the genus Gulo in the late Quaternary of Poland (after Marciszak, 2012 and references therein). Red dot: Gulo schlosseri; black dot: G . gulo from late middle Pleistocene ( 3-5) and late Pleistocene-Holocene ( 3-12). Localities: 1, Żabia Cave; 2, Kozi Grzbiet; 3, Biśnik Cave; 4, Cave no. 4 on the Birów Hill; 5, Deszczowa Cave; 6, Nietoperzowa Cave, Ciemna Cave; 7, Mamutowa Cave; 8, Borsuka Cave; 9, Cave in Czarkowa; 10, Niedźwiedzia Cave; 11, Północna Duża Cave, Naciekowa Cave; and 12, Solna Jama Cave.
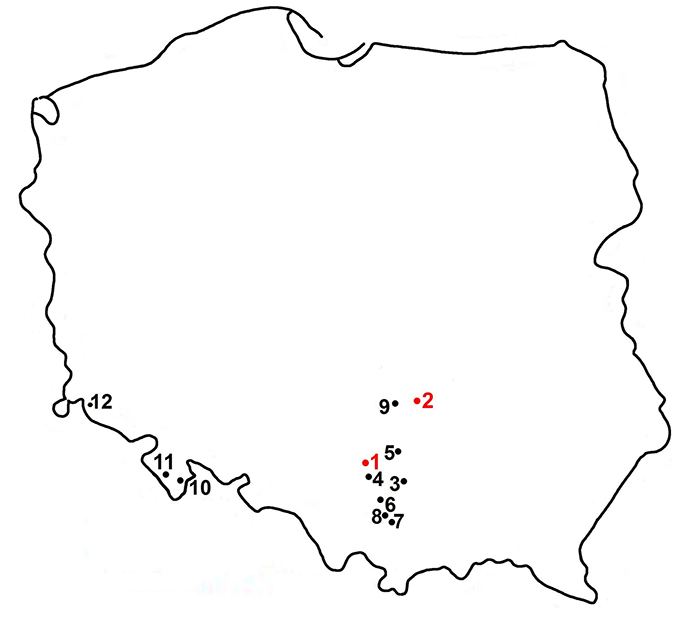
FIGURE 9. Right mandible of Mustela eversmanii (JSJ/Mev/1) from Solna Jama Cave. Note relatively short and robust body mandible and massiveness of the symphysal area. Scale bar equals 10 mm.
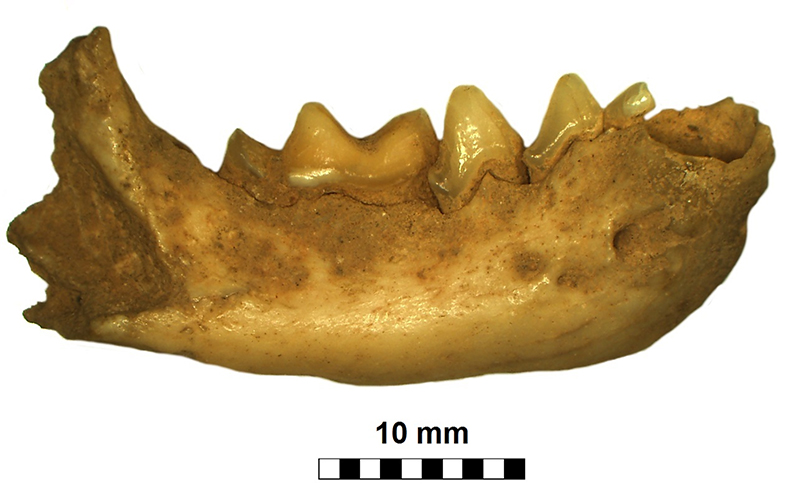
FIGURE 10. Scatter diagram showing the ratio of mandiblar height (measured after m1) to m1 length in fossil and extant Mustela eversmanii and Mu. putorius. Data from Marciszak (2012) and references therein.
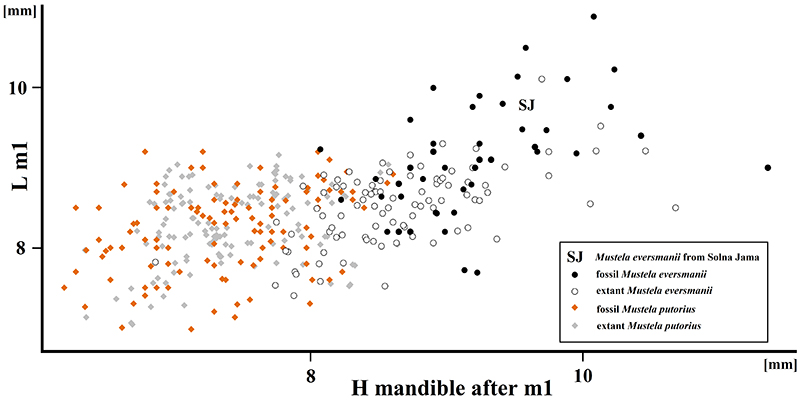
FIGURE 11. Skulls of Mustela nivalis from Poland: recent specimens ( 1-3) and the fossil from Solna Jama Cave ( 4). 1, robust, adult male; 2, adult female; 3, young, adult female; and 4, adult female. Note fully developed sagittal crest in individual from Solna Jama Cave, indicating fully mature age. Scale bar equals 10 mm.

FIGURE 12. Scatter diagram showing the ratio of total calvarium length to zygomatic breadth in extant Mustela nivalis from Poland, compared with the fossil specimen from Solna Jama Cave.
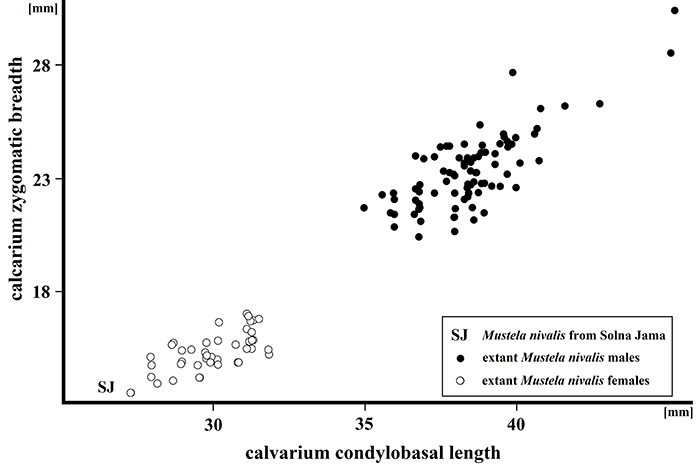
FIGURE 13. Scatter diagram showing the ratio of lower carnassial (m1) length (Lm1) and breadth (B m1) in late Pleistocene and Recent Mustela nivalis from Poland. The Solna Jama Cave specimen displays a moderately large size, with the length of m1 less than 4 mm, typical of the late Pleistocene and postglacial period.
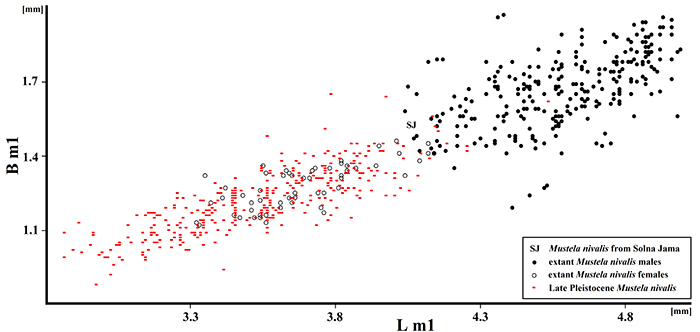
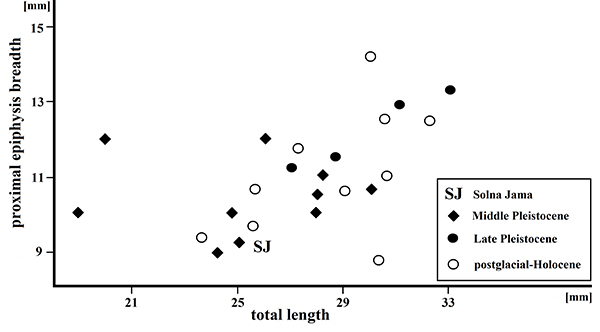
FIGURE 14. Scatter diagram showing the ratio of total length to proximal epiphysis breadth in late pleistocene-holocene Capreolus capreolus phalanx II. Middle Pleistocene locality: Kozi Grzbiet and Miesenheim 1. Late middle and late Pleistocene locality: Weimar Ehringsdorf, Biśnik Cave, Chlupáč Cave and Deszczowa Cave. Postglacial and Holocene locality: Biśnik Cave (uppermost layers), Jasna Strzegowska Cave and Poland in general. Data from Stefaniak (2015) and references therein.
APPENDIX 1.
Measurements of the canid mandible. 1, total length (infradentale to condyle); 2, length of infradentale to angular process; 3, length of infradentale to angular indentation; 4, behind c1 to condyle length; 5, infradentale to anterior margin of masseter fossa; 6, posterior margin of c1 to posterior margin of m3; 7, length of cheek tooth row (anterior margin of p1 to posterior margin of m3); 8, length of premolar row (anterior margin of p1 to posterior margin of p4); 9, length of molar row (anterior margin of m1 to posterior margin of m3); 10, distance between mental foramina; 11, symphysis maximum diameter; 12, symphysis minimum diameter; 13, posterior margin of m3 to condyle length; 14, condyle height; 15, condyle breadth; 16, angular process to coronoid process height; 17, mandible maximum height; 18, mandible body height between p2 and p3; 19, mandible body thickness between p2 and p3; 20, mandible body height between m1 and m2; and 21, mandible body thickness between m1 and m2.
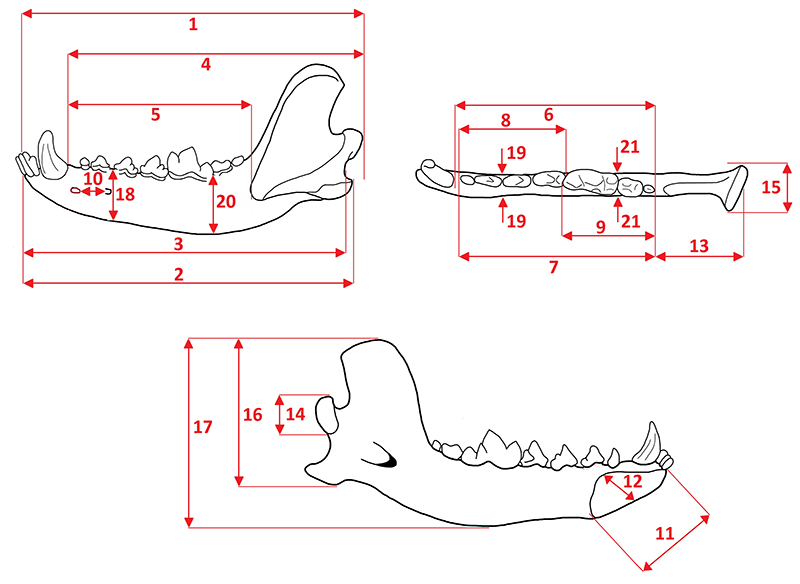
APPENDIX 2.
Measurements (left) and cusp terminology (right) of the ursid lower second molar (m2). 1, total length; 2, trigonid length from lingual side; 3, trigonid length from buccal side; 4, talonid length from lingual side; 5, talonid length from buccal side; 6, trigonid breadth; and 7, talonid breadth.
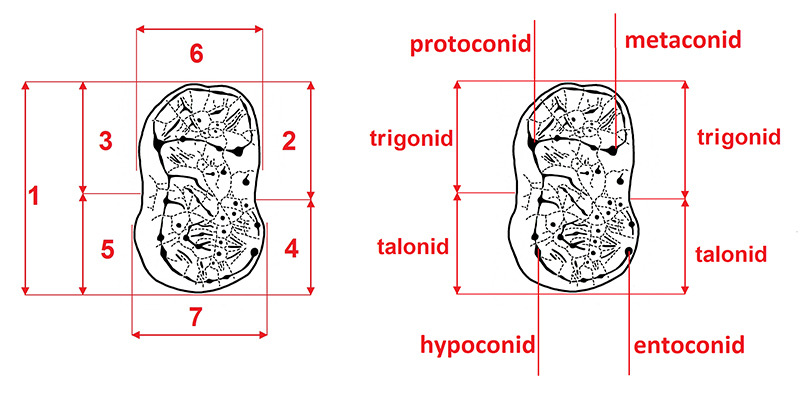
APPENDIX 3.
Measurements of the mustelid cranium. 1, total length (prosthion-acrocranion); 2, condylobasal length (prostion-occipital condyles); 3, basal length (prosthion-basion); 4, viscerocranium length (prosthion-point F); 5, neurocranium length (point F-acrocranion); 6, facial length (prosthion-point F); 7, rostrum length (prosthion-infraorbital); 8, rostrum length (anterior margin of I1 to posterior margin of M1); 9, palatal length (prosthion-staphylion); 10, C1-M1 length on alveoles (anterior margin of alveole of C1 to the posterior margin of alveole of M1); 11, upper premolars row length on alveoles (anterior margin of alveole of P2 to the posterior margin of alveole of P4); 12, breadth at zygomatic arches (zygion-zygion); 13, incisors row breadth (I3-I3 breadth); 14, maximal breadth at the canine alveoli; 15, maximal breadth at the P4 alveoli; 16, maximal breadth at the M1 alveoli; 17, least palatal breadth; 18, least breadth between infraorbital foramens; 19, least breadth between orbits (entorbital-entorbital); 20, frontal breadth (ectorbital-ectorbital); 21, postorbital least breadth (postorbital bar); 22, maximal neurocranium breadth (euryon-euryon); 23, mastoid breadth (otion-otion); 24, maximal breadth of occipital condyles; 25, nasal aperture height; 26, nasal aperture breadth; 27, height of foramen magnum (basionopisthion); 28, breadth of foramen magnum; 29, bullae ossae length; 30, bullae ossae breadth; 31, cranial height (acrocranion-basion); and 32, maximal cranial height (staphylion-frontal).
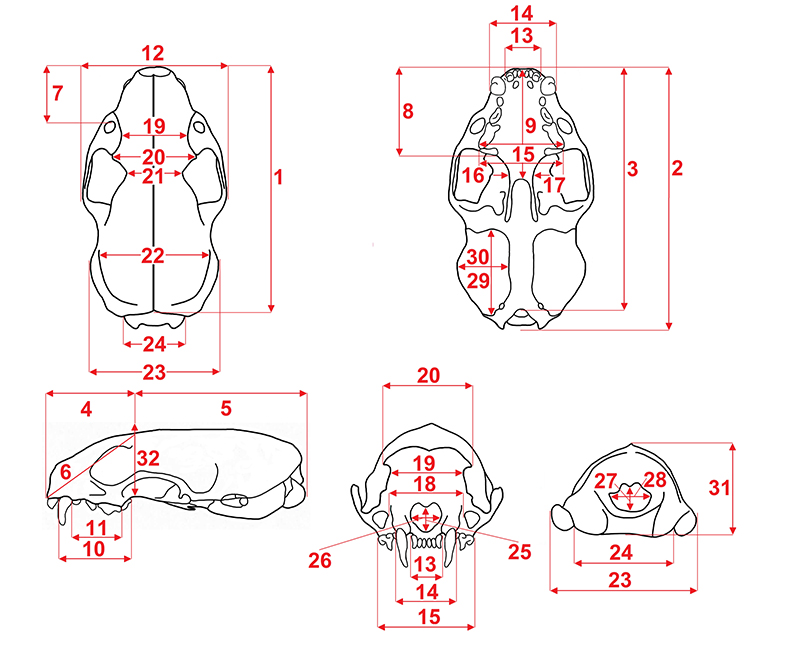
APPENDIX 4.
Measurements of the mustelid mandible. 1, total length (condyle to infradentale); 2, angular process to infradentale length; 3, infradentale to anterior margin of masseter fossa; 4, anterior margin of c1 to posterior margin of m2; 5, cheek teeth row length (anterior margin of p2 to posterior margin of m2); 6, premolars row length (anterior margin of p2 to posterior margin of p4); 7, molars row length (anterior margin of m1 to posterior margin of m2); 8, distance between mental foramens; 9, posterior margin of m2 to condyle length; 10, angular process to coronoid process height; 11, mandible maximal height; 12, body mandible height between p3 and p4; 13, body mandible thickness between p3 and p4; 14, body mandible height between m1 and m2; 15, body mandible thickness between m1 and m2; 16, condyle height; 17, condyle breadth; 18, symphysis maximal diameter; and 19, symphysis minimum diameter.
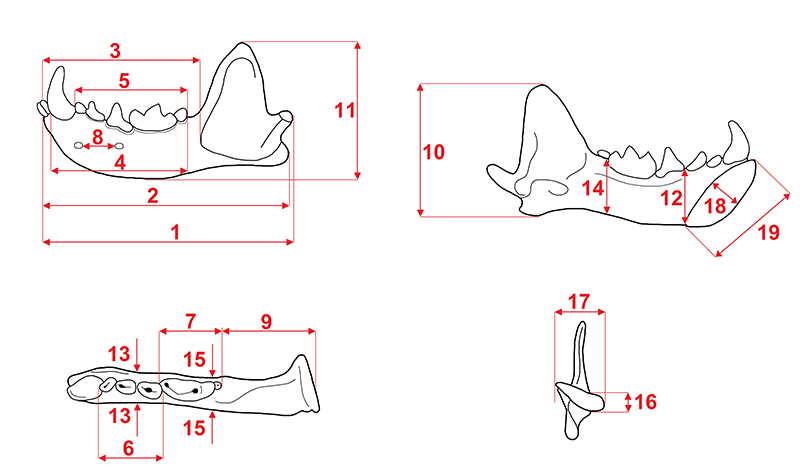
APPENDIX 5.
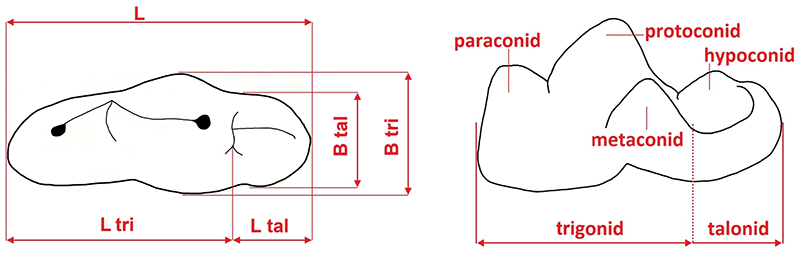
Measurements (left) and cusp terminology (right) of the felid lower carnassial (m1). Abbreviatios: L, total length; L tri, trigonid length; L tal, talonid length; B tri, trigonid breadth; B tal, talonid breadth; tri, trigonid; tal, talonid; par, paraconid; pro, protoconid; met, metaconid; and hyp, hypoconid.
APPENDIX 6.
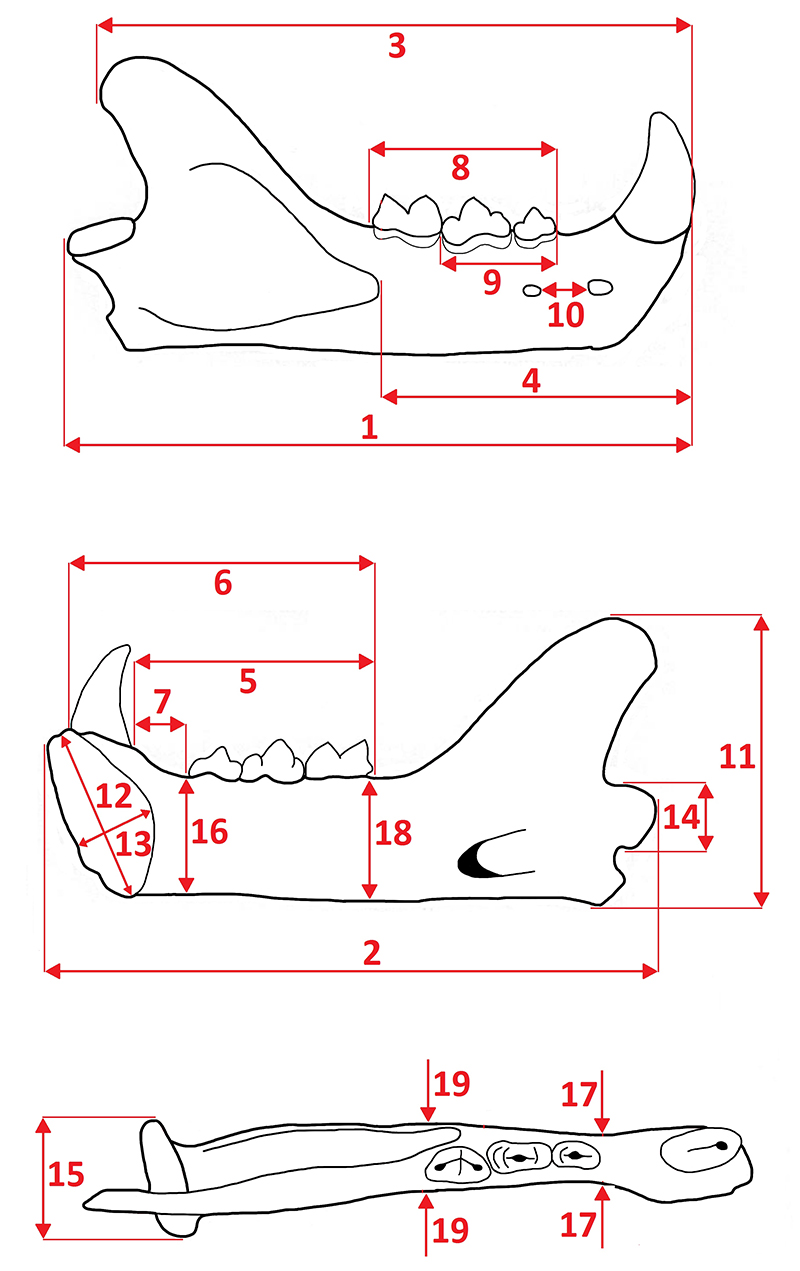
Measurements of the felid mandible. 1, total length (infradentale to condyle); 2, length of infradentale to angular process; 3, infradentale to coronoid process length; 4, infradentale to anterior margin of masseter fossa; 5, anterior margin of c1 to posterior margin of m1; 6, posterior margin of c1 to posterior margin of m1; 7, diastema length; 8, length of cheek tooth row (anterior margin of p3 to posterior margin of m1); 9, length of premolar row (anterior margin of p3 to posterior margin of p4); 10, distance between mental foramina; 11, angular process to coronoid process height; 12, symphysis maximum diameter; 13, symphysis minimum diameter; 14, condyle height; 15, condyle breadth; 16, mandible body height before p3; 17, mandible body thickness before p3; 18, mandible body height behind m1; and 19, mandible body thickness behind m1.
APPENDIX 7.
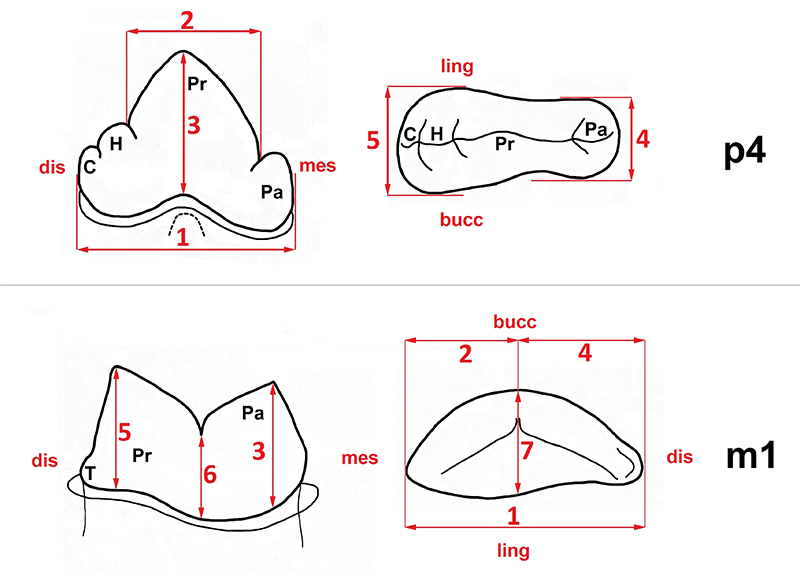
Measurements of the felid p3/p4 and m1 (modified from Schmid, 1940). Left: lateral view; right: occlusal view. Measurements of premolar: 1, total length; 2, protoconid length; 3, protoconid height; 4, anterior breadth; and 5, posterior breadth. Measurements of m1: 1, total length; 2, paraconid length; 3, paraconid height; 4, protoconid length; 5, protoconid height; 6, incisor height; and 7, breadth. Abbreviations: Pa, paraconid; Pr, protoconid; H, hypoconid; T, talonid; C, cingulum; mes, mesial side; dist, distal side; ling, lingual side; bucc, buccal side.
APPENDIX 8.
Measurements (in mm) of Gulo gulo postcranials from Solna Jama Cave. See Appendices 3 and 4 for measured parts.
| Bone part and collection no. | L | pl | pB | mL | mB | dL | dB | B |
| Humerus (JSJ/Gg/1-5) | 15.85 | 14.79 | ||||||
| Radius (JSJ/Gg/1-6) | 134.86 | 18.93 | 12.56 | 11.44 | 8.91 | 26.71 | 17.23 | |
| Mtcp I (JSJ/Gg/1-27) | 36.42 | 9.72 | 9.82 | 4.71 | 5.57 | 7.58 | 8.32 | |
| Mtcp I (JSJ/Gg/1-28) | 47.15 | 10.33 | 8.49 | 5.46 | 6.83 | 10.56 | ||
| Mtcp III (JSJ/Gg/1-29) | 55.38 | 11.98 | 9.91 | 5.71 | 7.16 | 9.72 | 10.94 | |
| Mtcp IV (JSJ/Gg/1-30) | 12.49 | 9.19 | 5.76 | 7.31 | ||||
| Mtcp V (JSJ/Gg/1-31) | 46.98 | 10.08 | 10.96 | 5.39 | 7.06 | 8.84 | 9.34 | |
| Mtts I (JSJ/Gg/1-32) | 5.36 | 6.54 | 7.80 | 8.78 | ||||
| Mtts II (JSJ/Gg/1-33) | 57.36 | 11.16 | 8.17 | 5.82 | 7.56 | 9.26 | 10.64 | |
| Mtts III (JSJ/Gg/1-34) | 64.65 | 14.76 | 11.24 | 6.67 | 8.14 | 9.42 | 11.32 | |
| Mtts IV (JSJ/Gg/1-35) | 68.69 | 14.98 | 10.21 | 6.84 | 7.93 | 9.54 | 11.23 | |
| Mtts V (JSJ/Gg/1-36) | 63.47 | 10.66 | 12.08 | 6.71 | 7.39 | 9.71 | 11.20 | |
| Calcaneus (JSJ/Gg/1-18) | 54.27 | 28.86 | ||||||
| Talus (JSJ/Gg/1-19) | 36.14 | 27.06 | ||||||
| Left pisiform (JSJ/Gg/1-20) | 21.50 | 11.75 | 12.58 | |||||
| Right pisiform (JSJ/Gg/1-21) | 21.68 | 11.69 | 12.92 | |||||
| Navicular (JSJ/Gg/1-22) | 19.97 | 15.47 | ||||||
| Cuboid (JSJ/Gg/1-23) | 15.46 | 15.11 | ||||||
| Cuneiform I (JSJ/Gg/1-24) | 13.67 | 7.59 | ||||||
| Cuneiform II (JSJ/Gg/1-25) | 9.74 | 6.65 | ||||||
| Cuneiform III (JSJ/Gg/1-26) | 16.81 | 9.85 | ||||||
| Phalange I (JSJ/Gg/1-37) | 28.84 | 8.52 | 10.87 | 5.68 | 6.18 | 5.63 | 8.58 | |
| Phalange I (JSJ/Gg/1-38) | 29.60 | 7.94 | 11.22 | 6.19 | 6.56 | 6.13 | 8.80 | |
| Phalange I (JSJ/Gg/1-39) | 29.90 | 8.16 | 10.52 | 5.15 | 6.23 | 6.05 | 8.02 | |
| Phalange I (JSJ/Gg/1-40) | 29.97 | 8.57 | 10.81 | 4.97 | 6.31 | 5.90 | 8.49 | |
| Phalange I (JSJ/Gg/1-41) | 26.39 | 8.32 | 10.99 | 5.36 | 6.52 | 5.89 | 8.59 | |
| Phalange I (JSJ/Gg/1-42) | 26.54 | 8.74 | 11.04 | 5.33 | 6.69 | 6.68 | 8.36 | |
| Phalange I (JSJ/Gg/1-43) | 25.12 | 6.45 | 9.50 | 4.25 | 6.32 | 6.27 | 7.63 | |
| Phalange I (JSJ/Gg/1-44) | 24.68 | 7.82 | 9.67 | 4.61 | 5.51 | 5.82 | 7.43 | |
| Phalange I (JSJ/Gg/1-45) | 30.02 | 9.02 | 11.18 | 5.27 | 6.94 | 6.12 | 8.85 | |
| Phalange I (JSJ/Gg/1-46) | 29.39 | 8.65 | 10.97 | 5.21 | 6.44 | 6.69 | 8.62 | |
| Phalange I (JSJ/Gg/1-47) | 25.61 | 8.05 | 10.80 | 5.25 | 6.66 | 5.89 | 7.96 | |
| Phalange I (JSJ/Gg/1-48) | 26.25 | 8.34 | 10.72 | 4.98 | 6.69 | 5.80 | 7.66 | |
| Phalange I (JSJ/Gg/1-49) | 27.06 | 8.62 | 10.47 | 5.24 | 6.58 | 6.01 | 8.23 | |
| Phalange I (JSJ/Gg/1-50) | 24.42 | 7.14 | 9.29 | 4.80 | 5.95 | 5.86 | 7.11 | |
| Phalange II (JSJ/Gg/1-51) | 17.63 | 8.11 | 8.76 | 4.36 | 5.16 | 6.76 | 6.69 | |
| Phalange II (JSJ/Gg/1-52) | 23.55 | 7.87 | 9.39 | 4.64 | 5.21 | 7.03 | 7.96 | |
| Phalange II (JSJ/Gg/1-53) | 17.71 | 7.51 | 9.01 | 4.71 | 5.61 | 6.42 | 7.49 | |
| Phalange II (JSJ/Gg/1-54) | 18.94 | 7.21 | 8.85 | 4.43 | 5.62 | 6.63 | 7.76 | |
| Phalange II (JSJ/Gg/1-55) | 22.25 | 7.49 | 9.35 | 4.64 | 5.48 | 6.63 | 7.87 | |
| Phalange II (JSJ/Gg/1-56) | 21.85 | 6.41 | 6.31 | |||||
| Phalange II (JSJ/Gg/1-57) | 20.74 | 7.17 | 8.63 | 4.44 | 5.34 | 7.22 | 7.79 | |
| Phalange II (JSJ/Gg/1-58) | 22.38 | 7.42 | 9.32 | 4.69 | 5.38 | 7.09 | 7.81 | |
| Phalange II (JSJ/Gg/1-59) | 22.13 | 7.13 | 9.09 | 4.39 | 6.08 | 6.98 | 7.67 | |
| Phalange II (JSJ/Gg/1-60) | 21.97 | 7.27 | 9.13 | 4.48 | 5.26 | 6.72 | 7.71 |
Large mammals (carnivores, artiodactyls) from Solna Jama Cave (Bystrzyckie Mts, Southwestern Poland) in the context of faunal changes in the postglacial period of Central Europe
Plain Language Abstract
Solna Jama Cave, small cave situated in an abandoned quarry in Southwestern Poland, before the palaeontological exploration was mostly known for its interesting extant invertebrate fauna. Palaeontological exploration started in the 1980s and resulted in 21 mammal forms found during the excavations. Quite surprisingly, the rodent material is relatively sparse. Shrews and bats are represented by two species, and rodents by five forms. The single find of Capreolus capreolus and undetermined large bovid are the only records of Artiodactyla. Carnivores are the most numerous (10 species) mammal taxon and represent at least four families. Besides the fossil material of mammals some fragments snail shells and undetermined bird fragments were found. Two faunal assemblages have been distinguished. The older one, with Ursus ingressus and single, huge m2 of U. arctos priscus, was dated at ca. 40 Ka. The second assemblage of the postglacial age (ca. 12 Ka) was represented by all the recorded mammal species, excluding the bears. The most noteworthy is a find of a skull and partial skeleton of a very large and robust Gulo gulo, which represents the only reliable Polish record of the species from postglacial period. This assemblage included mammals with different ecological preferences. Species associated with forests, like Sciurus vulgaris, Myodes glareolus, Martes martes, Felis silvestris or Ca. capreolus, have been found, together with those that lived on open habitats like Cricetus cricetus or Mustela eversmanii. Also euryoecious species like Arvicola terrestris, Canis lupus, Vulpes vulpes, G. gulo, Meles meles or Mu. nivalis were found in sediments of Solna Jama Cave. Gulo gulo and Mu. eversmanii are regarded as relict, cold-adapted survivors. The palaeoecological analysis showed the existence of taiga type forests together with small streams and meadows in the environs of the cave. Morphometric characteristics of carnivores, showed some adaptation to cool climatic conditions. Solna Jama Cave was situated on one of the few migration corridors crossing the Carpathian arch (Sudety Mts are the western continuation of the Carpathian Mts). The find expands the knowledge of the Sudetes fauna from the Pleistocene/Holocene boundary. In the future more detail excavations are planned, especially in the newly discovered chimney running from the slope of the hill, probably connected with the cave.
Resumen en Español
Grandes mamíferos (carnívoros, artiodáctilos) de la Cueva Solna Jama (Montes Bystrzyckie, Suroeste de Polonia) en el contexto de los cambios faunísticos durante el período post-glacial de Europa central
Los mamíferos de la Cueva Solna Jama resultaron partes provenientes de dos asociaciones faunísticas principales. La más temprana, datada como MIS 3, incluía algunos huesos de Ursus ingressus y un único m2 del enorme Ursus arctos priscus. La posterior, de edad post-glacial (MIS 1), estaba representada por Canis lupus, Mustela eversmanii y Felis silvestris, entre otros. El hallazgo más impresionante, un cráneo y un esqueleto parcial de un Gulo gulo muy grande y robusto, es el único registro documentado con fiabilidad en Polonia de la especie para el período post-glacial. El hallazgo aumenta el conocimiento de la fauna de los Sudetes para el límite Pleistoceno/Holoceno. Las características morfométricas de los carnívoros, el grupo más numeroso respecto a los otros taxones de mamíferos en esta localidad, mostraron cierta adaptación a las condiciones climáticas frías.
Palabras clave: post-glacial; Gulo gulo; calvarium; adaptación; La regla de Bergmann; fauna fósil cavernícola
Traducción: Enrique Peñalver (Sociedad Española de Paleontología)
Résumé en Français
Les grands mammifères (Carnivores, artiodactyles) de la grotte de Solna Jama (Monts Bystrzyckie, sud-ouest de la Pologne) dans le contexte des changements fauniques de la période post-glaciaire en Europe centrale
Les mammifères de la grotte de Solna Jama faisaient partie de deux principaux assemblages fauniques. Le plus ancien, daté du Stade Isotopique Marin 3, incluait quelques os d'Ursus ingressus et une seule m2 de l'énorme U. arctos priscus. L'assemblage plus récent, d'âge post-glaciaire (Stade Isotopique Marin 1), était représenté, entre autres, par Canis lupus, Mustela eversmanii, et Felis silvestris. La trouvaille la plus impressionnante, une tête osseuse et un squelette partiel d'un spécimen de Gulo gulo très grand et robuste, est le seul signalement fiable de cette espèce en Pologne pour la période post-glaciaire. Cette étude améliore la connaissance de la faune des Sudètes à la limite Pléistocène-Holocène. Les caractéristiques morphométriques des Carnivores, qui sont bien plus abondants que les autres taxons de mammifères dans le site, indiquent des adaptations à des conditions climatiques froides.
Mots-clés : post-glaciaire ; Gulo gulo ; calvarium ; adaptation ; loi de Bergmann ; faune fossile de grotte
Translator: Antoine Souron
Deutsche Zusammenfassung
Große Säugetiere (Karnivoren, Ariodactylen) aus der Solna Jama Höhle (Bystrzyckie Gebirge, südwestliches Polen) im Zusammenhang mit Faunenveränderung während der postglazialen Periode in Zentraleuropa
Säugertiere aus der Solna Jama Höhle sind Teil zwei wichtiger Faunenassemblagen. Die frühere, als MIS 3 datiert, beinhaltete wenige Knochen von Ursus ingressus und einen m2 des großen Ursus arctos priscus. Letzterer, mit einem postglazialen Alter (MIS 1) ist zusammen mit Canis lupus, Mustela eversmanii und Felis silvestris repräsentiert. Der eindrucksvollste Fund, ein Schädel und partielles Skelett eines sehr großen und robusten Gulo gulo, stellt den einzigen sicheren Fund dieser Art aus der postglazialen Periode in Polen dar. Der Fund erweitert das Wissen über die Sudeten-Fauna aus der Pleistozän/Holozän Grenze. Morphometrische Merkmale der Karnivoren, welche die Anzahl der anderen Taxa in dieser Fundstelle bei weitem übertrafen, zeigten einige Anpassungen an kühle klimatische Bedingungen.
Schlüsselwörter: postglazial; Gulo gulo; Schädelkalotte; Adaptation; Bergmannsche Regel; fossile Höhlenfauna
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
PE: An influential journal
 Palaeontologia Electronica among the most influential palaeontological journals
Palaeontologia Electronica among the most influential palaeontological journalsArticle number: 27.2.2E
July 2024

 A Review of Handbook of Paleoichthyology Volume 8a: Actinopterygii I, Palaeoniscimorpha, Stem Neopterygii, Chondrostei
A Review of Handbook of Paleoichthyology Volume 8a: Actinopterygii I, Palaeoniscimorpha, Stem Neopterygii, Chondrostei