

Volume 28.3
September–December 2025
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 28.3 September-December 2025
See all articles in 28.2 May-August 2025
See all articles in 28.1 January-April 2025
See all articles in 27.3 September-December 2024
Interested in submitting a paper to Palaeontologia Electronica?
Click here to register and submit.
 |
||||
 |
||||
Article Search
 Juan D. Carrillo. Paläontologisches Institut und Museum, Universität Zürich. Karl-Schmid-Strasse 4, 8006 Zürich, Switzerland and Department of Zoology, University of Cambridge. Downing Street CB2 3EJ, Cambridge, UK. juan.carrillo@pim.uzh.ch
Juan D. Carrillo. Paläontologisches Institut und Museum, Universität Zürich. Karl-Schmid-Strasse 4, 8006 Zürich, Switzerland and Department of Zoology, University of Cambridge. Downing Street CB2 3EJ, Cambridge, UK. juan.carrillo@pim.uzh.ch
Juan Carrillo is a PhD student at the Paleontological Institute and Museum of the University of Zurich, Switzerland and SNF fellow at the Museum of Zoology in Cambridge, UK. His research includes the systematics and paleobiology of south american mammals (native ungulates and rodents)

 Robert J. Asher. Department of Zoology, University of Cambridge. Downing Street CB2 3EJ, Cambridge, UK. r.asher@zoo.cam.ac.uk
Robert J. Asher. Department of Zoology, University of Cambridge. Downing Street CB2 3EJ, Cambridge, UK. r.asher@zoo.cam.ac.uk
Robert Asher is Senior Lecturer and Curator of Vertebrates at the Museum of Zoology in Cambridge, UK. His research interests include development and constraint among southern placental mammals (Afrotheria and Xenarthra), the evolution of endemic African mammals, and the diversification of therian mammals.
APPENDIX 1.
Definitions, measurements (in mm, unless otherwise stated) and equations used (Janis, 1990; Scott, 1990; Tsubamoto, 2014) to estimated the body mass (BM) in kg of MPEF-PV 8166. PE= Percent of error.
| Variable | Acronym | Definition | Type | Dataset | Value | Equation | R2 | % PE | Slope | Intercept | BM | BM + PE | BM - PE |
| Lower molar row length | LMRL | Measured along the base of the teeth | Dental | All ungulates | 75.6 | log BM= 3.265*log(LMRL)-0.536 | 0.941 | 31.9 | 3.27 | -0.536 | 396 | 522 | 269 |
| First lower molar length | FLML | Measured at the occlusal surface of the tooth | Dental | All ungulates | 20.1 | log BM= 3.263*log(FLML)+1.337 | 0.93 | 34.6 | 3.26 | 1.34 | 389 | 524 | 254 |
| First lower molar width | FLMW | Dental | All ungulates | 15.6 | log BM= 2.909*log(FLMW)+2.030 | 0.92 | 38.4 | 2.91 | 2.030 | 316 | 437 | 194 | |
| First lower molar area | FLMA | FLMA= FLML*FLMW | Dental | All ungulates | 313.3 | log BM= 1.553*log(FLMA)+1.701 | 0.93 | 33.2 | 1.55 | 1.7 | 378 | 503 | 252 |
| Second lower molar length | SLML | Measured at the occlusal surface of the tooth | Dental | All ungulates | 22.5 | log BM= 3.201*log(SLML)+1.130 | 0.94 | 31.9 | 3.2 | 1.130 | 289 | 381 | 197 |
| Second lower molar area | SLMA | SLMA= SLML*Second Lower Molar Width | Dental | All ungulates | 343.7 | log BM= 1.563*log(SLMA)+1.541 | 0.94 | 33.5 | 1.56 | 1.54 | 320 | 427 | 213 |
| Third lower molar area | TLMA | TLMA= Third lower molar length *Third lower molar width | Dental | All ungulates | 508.9 | log BM= 1.580*log(TLMA)+1.404 | 0.93 | 33.1 | 1.580 | 1.4 | 479 | 638 | 321 |
| Second upper molar length | SUML | Measured at the occlusal surface of the tooth | Dental | All ungulates | 27.9 | log BM= 3.184*log(SUML)+1.091 | 0.93 | 34.7 | 3.18 | 1.09 | 494 | 666 | 323 |
| Second upper molar width | SUMW | Measured at the occlusal surface of the tooth | Dental | All ungulates | 36.9 | log BM= 3.004*log(SUMW)+1.469 | 0.92 | 38.9 | 3 | 1.47 | 1501 | 2085 | 917 |
| Second upper molar area | SUMA | SUMA=SUML*SUMW | Dental | All ungulates | 1029.5 | log BM=1.568*log(SUMA)+1.277 | 0.94 | 32.7 | 1.57 | 1.28 | 1002 | 1329 | 674 |
| Posterior jaw length | PJL | Measured as the horizontal distance from the back of the jaw condyle to the posterior border of m3 | Crania | All ungulates | 65.3 | log BM= 2.412*log(PJL)+0.031 | 0.93 | 36.5 | 2.41 | 0.03 | 26 | 35 | 16 |
| Maximum width of the mandibular angle | WMA | Measured from the junction of the posterior part of m3 with the jaw to the maximally distant point on the angle of the jaw | Crania | All ungulates | 105.5 | log BM= 2.803*log(WMA)-0.352 | 0.92 | 40.5 | 2.8 | -0.35 | 209 | 293 | 124 |
| Length of the ridge for the masseteric attachment | MFL | Measured from the posterior portion of the jaw glenoid to the most anterior extent of the scar for the origin of the masseter muscle | Crania | All ungulates | 153 | log BM= 2.950*log(MFL)-1.289 | 0.94 | 35.0 | 2.950 | -1.289 | 143 | 193 | 93 |
| Humerus functional length (cm) | H1 | The distance between the most proximal point of the head and the most distal point of the trochlea | Postcranial | All ungulates | 19.85 | log BM= 3.4026*log(H1)-2.3707 | 0.92 | 28 | 3.4026 | -2.37 | 111 | 142 | 80 |
| Humerus trochlear diameter (cm) | H4 | The transverse distance of the distal articular surface | Postcranial | All ungulates | 4.76 | log BM= 2.4815*log(H4)+0.4516 | 0.95 | 22 | 2.4815 | 0.45 | 136 | 166 | 106 |
| Anteroposterior diameter of the femur (cm) | F6 | The transverse diameter of the diaphysis at its midpoint | Postcranial | All ungulates | 3.62 | log BM=2.8210*log(F6)+0.9062 | 0.94 | 23 | 2.8210 | 0.91 | 304 | 373 | 234 |
| Mediolateral diameter of the shaft (cm) | F7 | The anteroposterior diameter of the diaphysis at its midpoint | Postcranial | All ungulates | 2.55 | log BM=2.6016*log (F7)+0.9119 | 0.94 | 26 | 2.6016 | 0.91 | 93 | 117 | 69 |
| Diametro transverso proximal de la tibia (cm) | T2 | The transverse diameter of the tibia’s proximal epiphysis | Postcranial | All ungulates | 5.5 | log BM=2.8491*log(T2)-0.2495 | 0.95 | 21 | 2.8491 | -0.25 | 72 | 88 | 57 |
| Transverse width of tibial trochlea | Li1 | Astragalus | Various mammals | 27.7 | ln BM= 2.789*ln(Li1)+2.078 | 0.98 | 28.83 | 2.79 | 2.08 | 84 | 109 | 60 |
APPENDIX 2.
Selected measurement (in cm) and multivariate equations used (Mendoza et al., 2006) to estimated the body mass in kg (BM) of MPEF-PV 8166. PE= Percent of error.
| Adj. R2 | %MPE | mid PE |
LMRL | LPRL | JMA | JMC | JD | JMB | BM | BM+ mid PE | BM -mid PE |
| 0.98 | 21-25 | 23 | 7.56 | 5.86 | 6.53 | 10.55 | 4.88 | 177 | 217 | 167 | |
| 0.98 | 21-25 | 23 | 7.56 | 5.86 | 6.53 | 10.55 | 4.88 | 11.6 | 184 | 226 | 174 |
FIGURE 1. Geographical and stratigraphical occurrence of MPEF-PV 8166. 1 location of Cañadón Vaca, Chubut, Argentina; 2 Paleogene time table and South American Land Mammal Ages (SALMAs) after (Woodburne et al. 2014a,b); 3 skeletal restoration of Thomashuxleya modified from Simpson (1936); 4 artistic reconstruction of Thomashuxleya externa (by Stjepan Lukac).
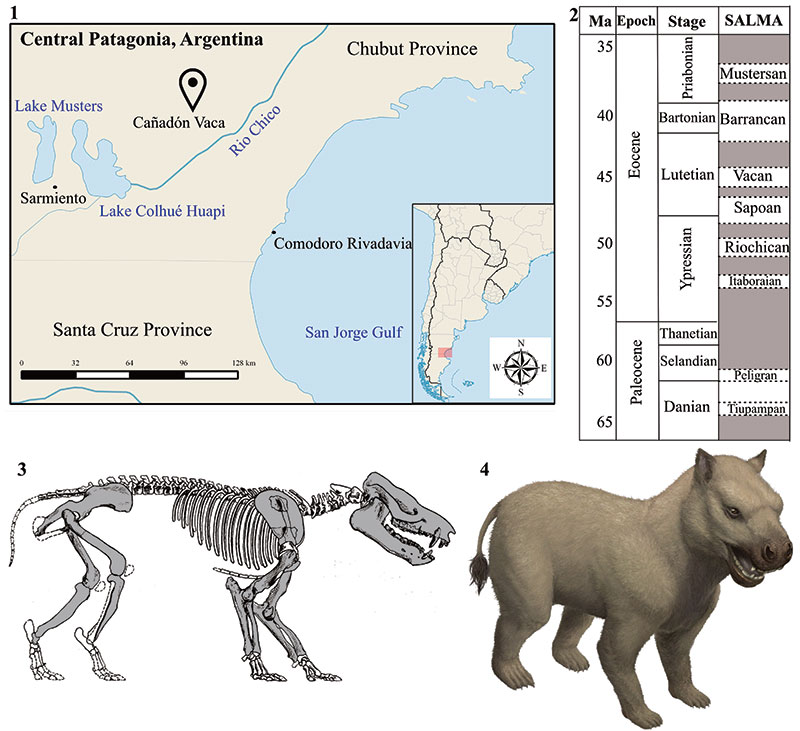
FIGURE 2. Skull of T. externa (MPEF-PV 8166). 1 ventral view; 2 lateral view; 3 occiput in caudal view; 4 detail of upper dentition in occlusal view. Abbreviations are prot=protocone (part of the protoloph), par=paracone (part of the ectoloph), parst=parastyle, met=metacone (part of the ectoloph), metst=metastyle, meph=metaloph, pr.l.f.=primary lingual fold, m.cin=mesial cingulum, l.cin=lingual cingulum.
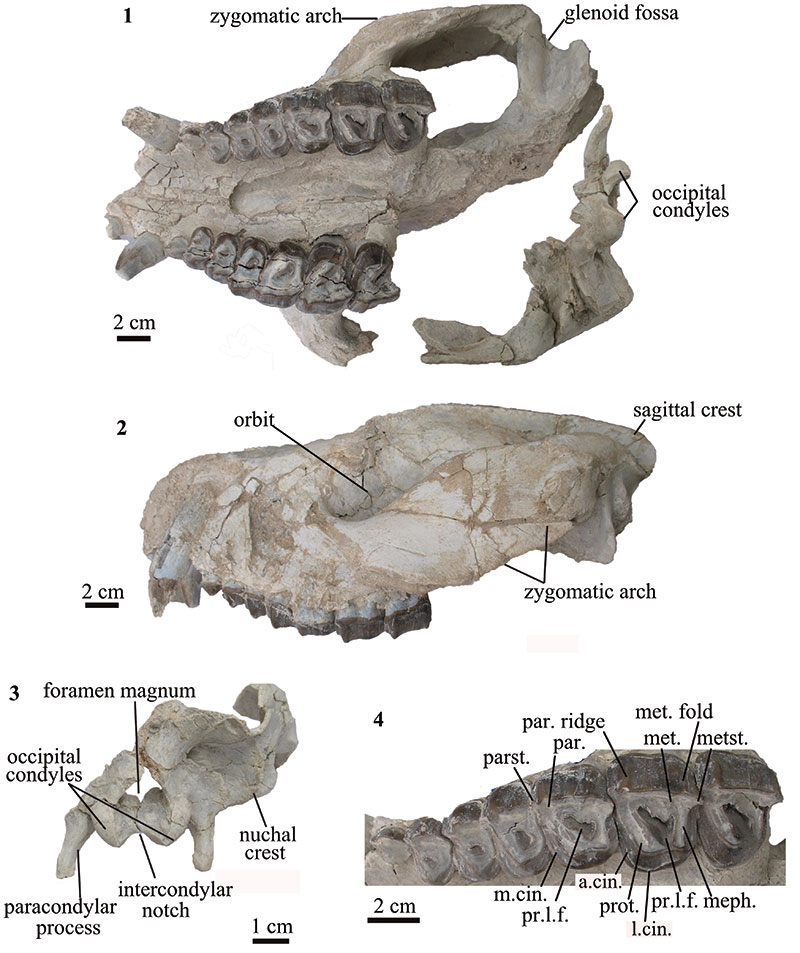
FIGURE 3. Mandible of T. externa (MPEF-PV 8166) in 1 dorsal and 2 lateral views; 3 lower dentition in occlusal view. Abbreviations are tri=trigonid, tal=talonid, prod=protoconid, metd=metaconid, entd=entoconid, hypd=hypoconulid.
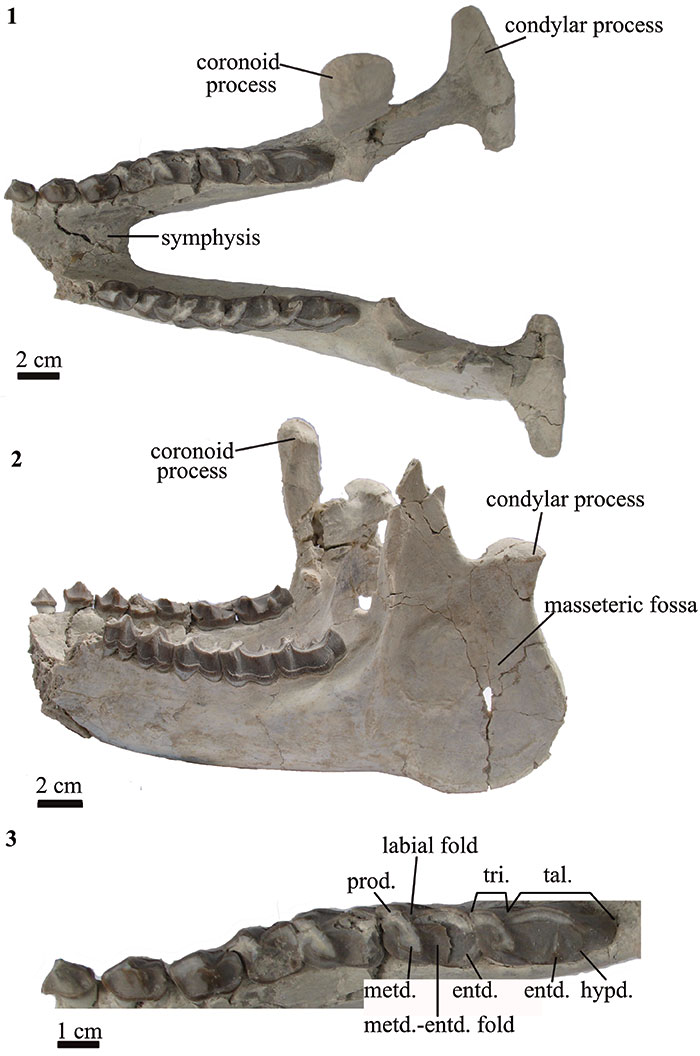
FIGURE 4. Scapulae and humeri of T. externa (MPEF-PV 8166). 1 right and 2 left scapulae in proximal view; 3 left scapula in dorsolateral view; right and left humeri in 4 right and left humeri in anterior (top) and distal (bottom) views; 5 right and left humeri in posterior view.

FIGURE 5. Forelimb of T. externa (MPEF-PV 8166). 1 right and left ulnae in frontal view; 2 left ulna in medial view; right radius in 3 frontal, 4 lateral, and 5 posterior views.
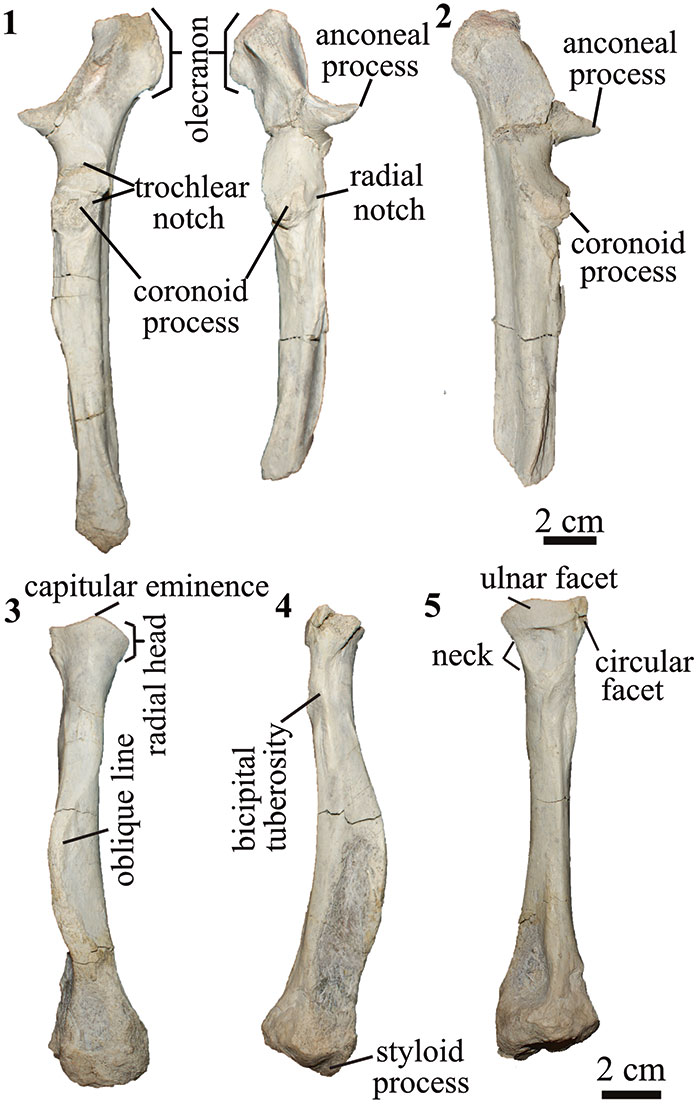
FIGURE 6. Hindlimb of T. externa (MPEF-PV 8166). right pelvis in 1 dorsal and 2 lateral views; 3 right femur in anterior view; 4 right tibia in proximal (top), anterior (middle) and distal (bottom) views; left astragalus in 5 dorsal and 6 plantar views; 7 right navicular in proximal view; 8 navicular and entocuneiform in plantar views.
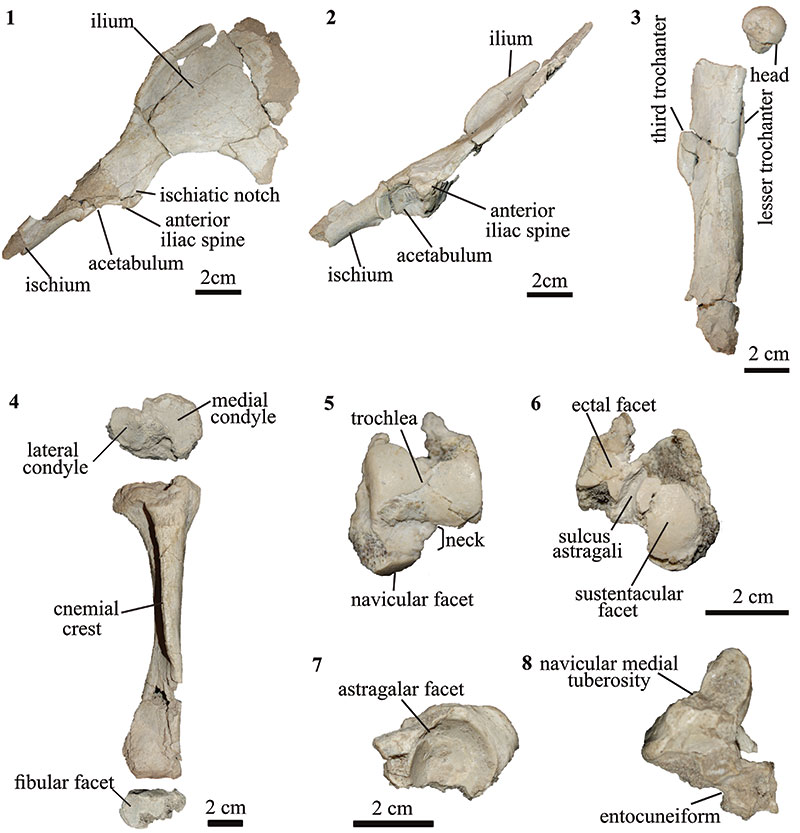
FIGURE 7. Bone histology and microstructure of T. externa (MPEF-PV 8166). 1 Cross section of the midshaft of the right femur; 2 same as 1 after conversion to a binary image (black represents bone and white the cavities); 3 bone histology under linear polarized light, with black arrows pointing the lines of arrested growth (LAGs). OCL= Outer circumferential layer. 4 Bone histology under cross polarized light.

FIGURE 8. Strict consensus of 281 trees, 420 steps in length showing the phylogenetic relationships of Thomashuxleya within Notoungulata based on the morphological dataset of Deraco and García-López (2015). Numbers indicate bootstrap values above 50.
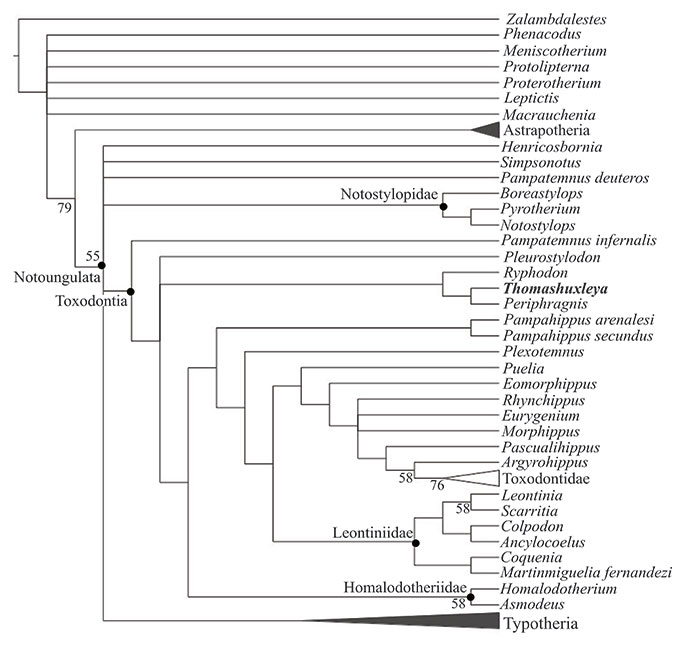
FIGURE 9. Strict consensus of 510 trees, 122374 steps in length from unconstrained parsimony analysis of combined proteomic and morphological data.
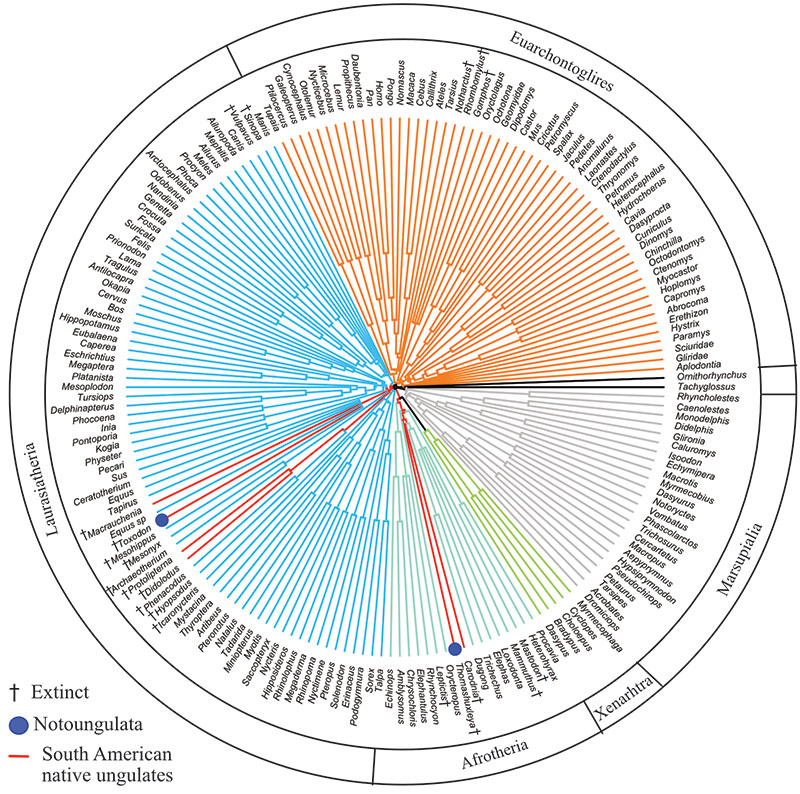
FIGURE 10. Strict consensus of 620 trees, 122391 steps in length from parsimony analysis of combined proteomic and morphological data constraining monophyly of each of two clades (but not both together): Notoungulata (i.e., Thomashuxleya and Toxodon) and Litopterna (i.e., Protolipterna and Macrauchenia).
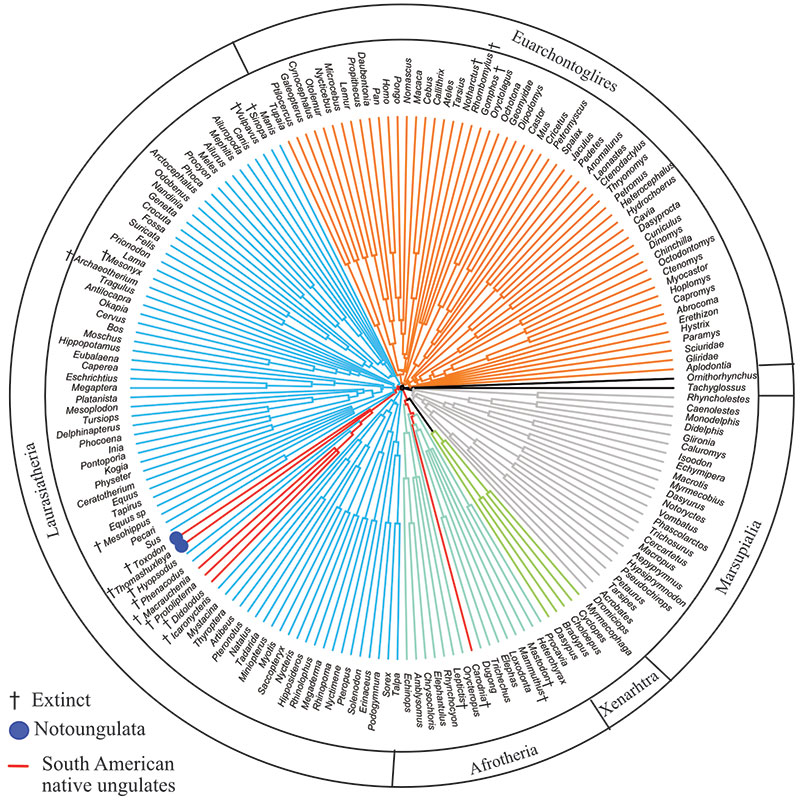
FIGURE 11. Optimal Bayesian tree (i.e., 50% majority rule of post-burn-in trees) of combined proteomic and morphological data. Numbers represent Bayesian posterior probabilities; daggers indicate fossil taxa.
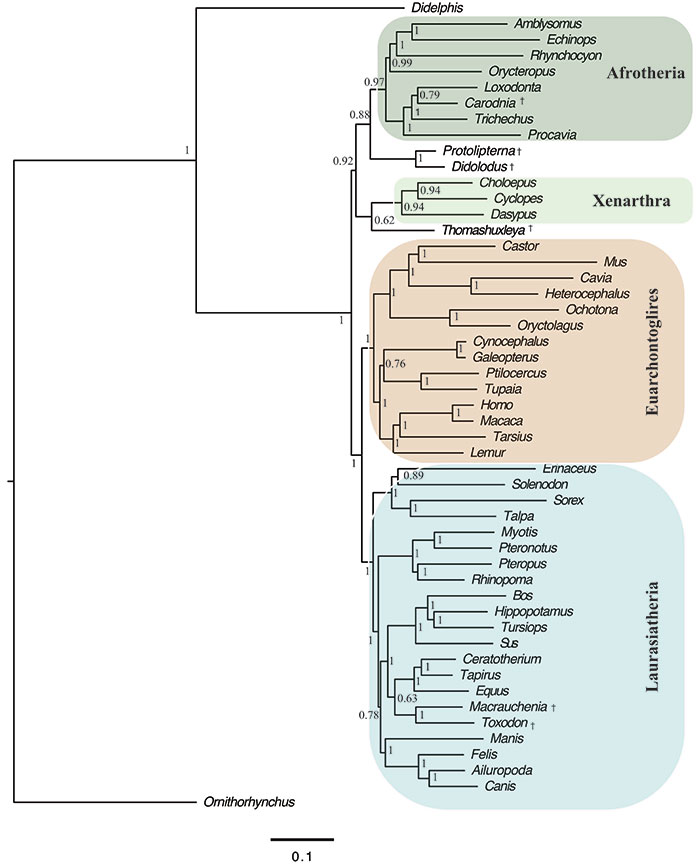
FIGURE 12. Optimal Bayesian tree (i.e., 50% majority rule of post-burn-in trees) of combined proteomic and morphological analysis constraining monophyly of each of two clades (but not both together): Notoungulata (i.e., Thomashuxleya and Toxodon) and Litopterna (i.e., Protolipterna and Macrauchenia). Numbers represent Bayesian posterior probabilities; daggers indicate fossil taxa.
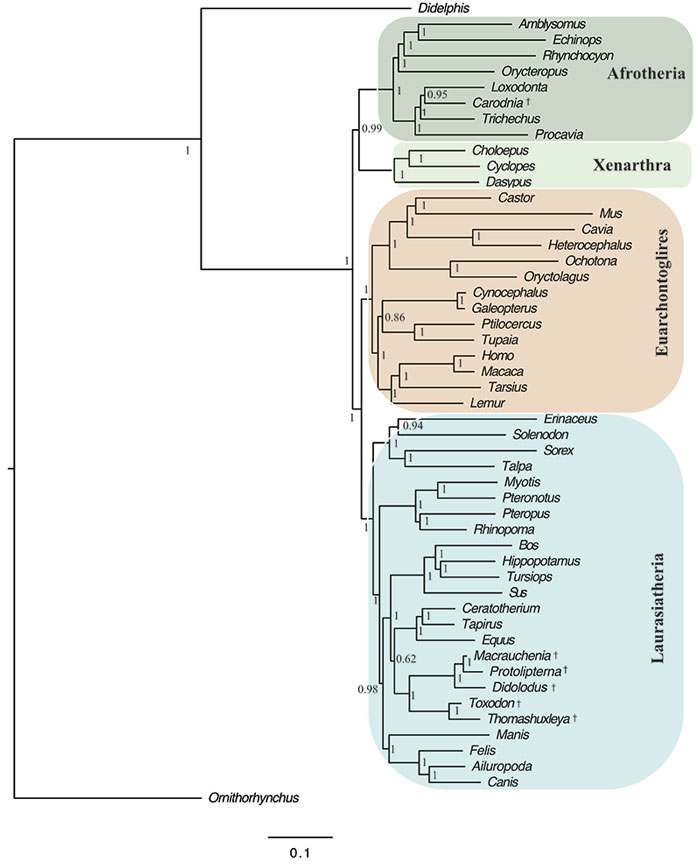
TABLE 1. Best partition scheme for Bayesian analysis as obtained from Partition Finder (Lanfear et al., 2012).
| Best model | Subset partitions | Subset sites |
| JTT+G | TTN | 1-1479 |
| JTT+G | CNR1 | 1480-1814 |
| JTT+G | BCHE | 1815-2146 |
| JTT+G | EDG1 | 2147-2466 |
| JTT+G | RAG1 | 2467-3066 |
| JTT+G | RAG2 | 3067-3215 |
| JTT+G | ATP7A | 3216-3444 |
| JTT+G | TYR1 | 3445-3587 |
| JTT+G | Adora3 | 3588-3698 |
| JTT+G | BDNF | 3699-3885 |
| JTT+G | ADRB2 | 3886-4153 |
| JTT+G | PNOC | 4154-4260 |
| JTT+G | A2AB | 4261-4543 |
| JTT+G | BRCA1, BRCA2 | 4544-5574, 5575-7256 |
| JTT+G | APOB, DMP1 | 7257-7710, 9718-10592 |
| JTT+G | GHR | 7711-8026 |
| JTT+G | VWF | 8027-8417 |
| JTT+G | ENAM | 8418-9717 |
| JTT+G | IRBP | 10593-11010 |
| MTMAM+G | Collagen alpha 1 and 2 | 11011-13038 |
| Standard discrete | Morphology | 13039-16698 |
TABLE 2. Skeletal elements of isotemnid specimens from Cañadón Vaca.
| T. externa MPEF-PV 8166 |
T. externa MNHN CAS 844 |
T. externa MNHN CAS 64 |
T. externa AMNH 28698 |
T. externa AMNH 28447 |
T. externa AMNH 28905 |
T. externa AMNH 28563 |
cf. T. externa AMNH 142463 |
A. distentus AMNH 28906 |
A.distentus AMNH 28647 |
P. similis AMNH 28904 |
P. similis AMNH 28635 |
Isotemnidae indet. AMNH 28690 |
|
| Skull | x | x | x | M1 only | |||||||||
| Mandible | x | x | x | ||||||||||
| Scapula | x | x | x | ||||||||||
| Humerus | x | x | x | x | x | ||||||||
| Ulna | x | x | x | x | x | ||||||||
| Radius | x | x | x | x | x | ||||||||
| Manus | x | x | x | ||||||||||
| Pelvis | x | x | |||||||||||
| Femur | x | x | x | x | |||||||||
| Tibia | x | x | x | x | |||||||||
| Pes | x | x | x | x | x |
TABLE 3. Dental measurements of Thomashuxleya; * =taken from Simpson (1967).
| p1 | p2 | p3 | p4 | m1 | m2 | m3 | |||||||||
| Length | Width | Length | Width | Length | Width | Length | Width | Length | Width | Length | Width | Length | Width | ||
| T. rostrata | MACN 10370* | 13.7 | 11.8 | 17.5 | 12.7 | 18.1 | 14.8 | 20.9 | 16.6 | ||||||
| MACN 10539* | 19.9 | 17.8 | 26 | 19.8 | 26.4 | 19.5 | 37.2 | 16.9 | |||||||
| MACN 10546* | 21.5 | 16.8 | |||||||||||||
| AMNH 28692* | 19.5 | 16.5 | 26 | 16.4 | 28.5 | 18.8 | |||||||||
| AMNH 28764* | 21.8 | 13.7 | |||||||||||||
| Mean | 13.7 | 11.8 | 17.5 | 12.7 | 18.8 | 15.7 | 21.0 | 16.2 | 26 | 18.1 | 27.5 | 19.2 | 37.2 | 16.9 | |
| SD | NA | NA | NA | NA | 1.0 | 1.2 | 0.8 | 1.8 | 0.0 | 2.4 | 1.5 | 0.5 | NA | NA | |
| n | 1 | 1 | 1 | 1 | 2 | 2 | 4 | 4 | 2 | 2 | 2 | 2 | 1 | 1 | |
| T. externa | MACN 10540* | 16.5 | 14.6 | 19 | 17.2 | 21 | 28 | 37 | |||||||
| MACN 10537* | 16.3 | 13 | 19.5 | 14 | 23 | 15.5 | |||||||||
| AMNH 28447* | 14.2 | 11.1 | 16.2 | 11.6 | 18.1 | 13 | 22.0 | 15.6 | 27.5 | 16.5 | 31.6 | 15.8 | |||
| AMNH 28697* | 17.3 | 14.2 | 23.2 | 15.9 | 25.5 | 17.2 | 36 | 16.6 | |||||||
| AMNH 28756* | 14.7 | 10.9 | 15.7 | 12.8 | 16.4 | 14.2 | 21.8 | 16.3 | 25.6 | 17.8 | 32.3 | 16.3 | |||
| AMNH 28698* | 11.0 | 10.0 | 15.1 | 12.6 | 16.7 | 13.6 | 16.5 | 14.6 | 24 | 16.1 | 26 | 17.8 | 37 | 17 | |
| AMNH 28686* | 34.5 | 17.0 | |||||||||||||
| AMNH 28822* | 20 | 16.4 | 23 | 18.0 | |||||||||||
| Mean* | 11 | 10 | 14.7 | 11.5 | 16.3 | 13.2 | 17.3 | 14.4 | 21.6 | 15.7 | 25.5 | 17.1 | 34.7 | 16.5 | |
| SD* | NA | NA | 0.5 | 0.9 | 0.4 | 1.3 | 1.1 | 1.5 | 1.6 | 0.9 | 2.0 | 1.0 | 2.4 | 0.5 | |
| n* | 1 | 1 | 3 | 3 | 4 | 4 | 6 | 6 | 7 | 6 | 7 | 7 | 6 | 5 | |
| MPEF-PV 8166 | 12.8 | 8.3 | 15.4 | 10 | 16.5 | 10.1 | 16.4 | 13.3 | 20.1 | 15.6 | 22.5 | 15.3 | 32.3 | 15.7 | |
|
|
P1 | P2 | P3 | P4 | M1 | M2 | M3 | ||||||||
|
|
|
Length | Width | Length | Width | Length | Width | Length | Width | Length | Width | Length | Width | Length | Width |
| T. rostrata | MACN 10370* | 15.4 | 15.7 | 17.0 | 27.2 | 19.5 | 31.2 | 21 | 35 | ||||||
| MACN 10542* | 29.5 | 41.8 | 33.0 | 44.2 | 28.8 | 40.8 | |||||||||
| Mean* | 15.4 | 15.7 | 17 | 27.2 | 19.5 | 31.2 | 21 | 35 | 29.5 | 41.8 | 33 | 44.2 | 28.8 | 40.8 | |
| SD* | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | |
| n* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| T. externa | MACN 10543* | 24 | 34 | 28.5 | 35.9 | ||||||||||
| AMNH 28699* | 17.5 | 22.1 | 16.8 | 26.4 | 18.5 | 32.1 | 23.8 | 35.8 | 26.7 | 42 | 39.5 | ||||
| AMNH 28757* | 13.8 | 11.5 | 15.2 | 22.1 | 18 | 26 | 25.7 | 27.2 | 39.2 | ||||||
| AMNH 28698* | 10 | 11 | 16 | 18 | 33.2 | 23.9 | 38.7 | 27.2 | 41 | 26 | 41 | ||||
| MNHN CAS 844 | 10.84 | 13.86 | 17.79 | 22.93 | 18.94 | 26.80 | 19.13 | 31.25 | 21.40 | 32.75 | 22.71 | 35.98 | 26.55 | 35.46 | |
| Mean* | 11.5 | 12.1 | 16.8 | 22.4 | 17.4 | 26.4 | 18.5 | 32.2 | 23.8 | 35.3 | 26.5 | 38.8 | 26.3 | 38.7 | |
| SD* | 2.0 | 1.5 | 1.4 | 0.5 | 1.3 | 0.4 | 0.6 | 1.0 | 1.5 | 2.6 | 2.2 | 2.8 | 0.4 | 2.9 | |
| n* | 3 | 3 | 3 | 3 | 4 | 3 | 3 | 3 | 5 | 4 | 5 | 5 | 2 | 3 | |
| MPEF-PV 8166 | 10.2 | 10.5 | 14.0 | 16.2 | 15.0 | 21.6 | 14.6 | 25.9 | 25.2 | 31.3 | 27.9 | 36.9 | 24.0 | 32.0 | |
TABLE 4. Postcranial measurements of MPEF-PV 8166. * Translated from Spanish in Elissamburu (2012).
| Value (mm) |
Acronym | ||||
| Left | Rigth | Scott (1990) |
Elissamburu (2012) |
||
| Scapula | |||||
| Height of the spine | 31.0 | 31.4 | |||
| Humerus | |||||
| Humerus functional length* | 193 | 204 | H1 | LFH | |
| Humerus trochlear diameter* | 52.0 | 43.3 | H4 | DtrH | |
| Maximum length | 211 | 210 | |||
| Maximum diameter of the head | 44.0 | 40.5 | |||
| Maximum diameter of the mayor tuberosity | 46.6 | ||||
| Maximum diameter of the minor tuberosity | 16.6 | ||||
| Maximum widht of the trochlea | 30 | ||||
| Radius | |||||
| Maximum length | 155 | 153 | |||
| Maximum mediolateral width of the medial shaft | 14.1 | 14.0 | |||
| Maximum diameter of the head | 32.0 | ||||
| Minimum diameter of the head | 20.3 | 20.4 | |||
| Maximum diameter of the neck | 20.3 | 19.5 | |||
| Mediolateral width of the distal epiphysis | 36.0 | ||||
| Diameter perpendicular to the maximum width of the distal epiphysis | 31.6 | ||||
| Ulna | |||||
| Anteroposterior diameter of the diaphysis | 28.3 | 28.1 | |||
| Olecraneum height | 64.9 | 63.7 | |||
| Anteroposterior diameter of the oleocranon | 32.6 | ||||
| Proximo-distal length of the trochlear notch | 35.9 | ||||
| Trochlear notch width | 32.6 | ||||
| Width of anconeal process | 20.5 | ||||
| Femur | |||||
| Anteroposterior diameter of the shaft | 25.5 | F7 | DAPF | ||
| Mediolateral diameter of the shaft | 36.2 | F6 | DTF | ||
| Tibia | |||||
| Mediolateral diameter of proximal epiphysis | 55.0 | T2 | DTpT | ||
| Anteroposterior diameter of proximal epiphysis | 44.8 | ||||
| Lateral condyle transverse width | 28.0 | ||||
| Lateral condyle anteroposterior width | 25.0 | ||||
| Astragalus | |||||
| Transverse width of tibial trochlea | 27.7 | Li 1 | |||
| Minimum width of the neck | 14.8 | ||||
| Width sustentacular facet | 17.4 | ||||
| Navicular | |||||
| "Dorsal-plantar" width | 25.6 | ||||
| Astragalar facet transverse width | 24.0 | ||||
| Astragalar facet dorsoventral depth | 25.0 | ||||
| Pelvis | |||||
| Maximum width of the ilium, above the acetabulum | 42.3 | ||||
TABLE 5. Isotemnid astragali measurements in mm.
| Specimen No. | Locality | Maximum length | Trochlear width | Maximum width | Head width |
| MPEF-PV 8166 | Cañadón Vaca | 27.7 | |||
| AMNH 142463 | Cañadón Vaca | 35.9 | 41.4 | ||
| AMNH FM 14501 | "Notostylops beds" | 32.8 | 21.8 | 32.1 | 17.7 |
| AMNH 28690 | Cañadón Vaca | 22 | 12 | 20.6 | |
| AMNH 142464 | Cañadón Vaca | 20.5 | 8.9 | 16.7 | 9.3 |
TABLE 6. Results from the Wilcoxon signed ranks test comparing topologies carried out in PAUP. Meridungulata = constraint with all SANUs within a clade; N/L= Notoungulata (Thomashuxleya, Toxodon) and Litopterna (Macrauchenia, Protolipterna) monophyly constraints for each clade (but not both together); p values greater than 0.05 indicate no difference between optimal and competing topology.
| Topology | Tree length | Rank sums | N | z | p |
| 122374 | (best) | ||||
| N/L | 122391 | 52385.0/--48640.0 | 449 | -0.77 | 0.4390 |
| Meridungulata | 122424 | 214600.5/-193555.5 | 903 | -1.49 | 0.14 |
An exceptionally well-preserved skeleton of Thomashuxleya externa (Mammalia, Notoungulata), from the Eocene of Patagonia, Argentina
Plain Language Abstract
Notoungulata is an enigmatic group of placental mammals that inhabited South America during most of the past 60 million years. Despite more than a century of study, the relationships of nototoungulates to other mammals are not fully resolved and remain a major challenge in paleomammalogy. Here, we describe one of the oldest and most complete skeletons of a notoungulate from the middle Eocene of Patagonia, Argentina, which lived about 45 million years ago. The association of a skull with teeth and other skeletal remains allows us to identify the specimen as Thomashuxleya externa (Isotemnidae), estimate its body mass, describe its bone microanatomy, and examine its relationships to other mammals. This animal likely weighed about 84 ± 24.2 kg. Analysis of microstructure of bones in cross section shows that the new specimen was skeletally mature. In order to infer the position of Thomashuxleya in the evolutionary tree of mammals, we combined sequences from DNA and collagen sequences (known for living and some very recent fossils), and morphological data. This analysis favors a limited number of possible phylogenetic relationships, with possible affinities to even- and odd-toed ungulates, endemic African mammals, or other endemic South American mammals.
Resumen en Español
Un esqueleto excepcionalmente bien preservado de Thomashuxleya externa (Mammalia, Notoungulata), del Eoceno de Patagonia, Argentina
Describimos uno de los esqueletos de notoungulados más antiguos con elementos cráneo-dentales y postcraneales asociados: Thomashuxleya externa (Isotemnidae) de Cañadón Vaca en Patagonia, Argentina (subedad Vaquense de la Edad Mamífero Casamayorense, Eoceno medio). Proveemos estimaciones de masa corporal dadas por distintos elementos del esqueleto, describimos la histología de los huesos, y estudiamos la posición filogenética. Notamos diferencias en las escápulas, húmeros, ulnas y radios del nuevo espécimen en comparación con otros especímenes previamente referidos a este taxon. Estimamos una masa corporal de 84 ± 24,2 kg, mostrando que los notoungulados habían adquirido una gran masa corporal ya en el Eoceno medio. La histología de huesos muestra que el nuevo espécimen era esqueletalmente maduro. El nuevo material soporta la posición de Thomashuxleya como un miembro temprano y divergente de Toxodontia. Dentro de los placentarios, nuestro análisis filogenético de una matriz combinada de ADN, colágeno y morfología favorece únicamente un número limitado de posibles relaciones filogenéticas, pero no puede aún arbitrar entre afinidades potenciales con Afrotheria or Laurasiatheria. Sin restricciones, la parsimonia máxima soporta Thomashuxleya y Carodnia con Afrotheria. Con Notoungulata y Litopterna restringidos como monofiléticos (incluyendo Macrauchenia y Toxodon conocidos por colágenos), estos clados son reconstruidos en el stem de Euungulata (i.e., Perissodactyla y Artiodactyla). El análisis bayesiano no restringido soporta débilmente la posibilidad de que Thomashuxleya sea stem Xenarthra; con Notoungulata y Litopterna restringidos como monofiléticos, los dos clados son recuperados como grupo hermano de Perissodactyla. Datos anatómicos muestreados hasta el momento para Thomashuxleya, combinados con aminoácidos de colágeno para meridiungulados pleistocenos, limitan substancialmente el número de afinidades posibles para las especies endémicas de América del Sur dentro de los mamíferos, aunque aún existe ambigüidad.
Palabras clave: América del Sur; Toxodontia; Litopterna; masa corporal; filogenia
Traducción: Diana Elizabeth Fernández
Résumé en Français
Un squelette exceptionnellement bien préservé de Thomashuxleya externa (Mammalia, Notoungulata) dans l'Éocène de Patagonie, Argentine
Nous décrivons un des plus anciens squelettes de notongulé avec des éléments crâniodentaires et postcrâniens associés : Thomashuxleya externa (Isotemnidae) de Cañadón Vaca en Patagonie, Argentine (sous-âge « Vacan » du Casamayorien, dans l’échelle SALMA des âges des grands mammifères terrestres d’Amérique du Sud, Éocène moyen). Nous fournissons des estimations de masse corporelle obtenues d'après différents éléments du squelette, nous décrivons l'histologie osseuse, et nous étudions sa position phylogénétique. Nous notons des différences au niveau des scapulae, humeri, ulnas, et radius du nouveau spécimen par rapport aux autres spécimens attribués précédemment à ce taxon. Nous estimons une masse corporelle de 84 ±24,2 kg, ce qui montre que les notongulés avaient acquis une masse corporelle importante dès l'Éocène moyen. L'histologie osseuse montre que ce spécimen avait atteint la maturité squelettique. Le nouveau matériel soutient le placement de Thomashuxleya comme un membre basal, divergent tôt au sein des Toxodontia. Parmi les placentaires, notre analyse phylogénétique d'une matrice combinant l'ADN, le collagène, et la morphologie ne soutient qu’un nombre limité de relations phylogénétiques possibles, mais ne peut pas encore trancher entre des affinités potentielles avec les Afrotheria ou les Laurasiatheria. En l’absence de contraintes, le maximum de parcimonie soutient le groupement de Thomashuxleya et Carodnia avec les Afrotheria. Avec une contrainte imposant Notoungulata et Litopterna (incluant Macrauchenia and Toxodon, taxons dont le collagène est connu) comme monophylétiques, ces clades sont reconstruits à la base d'Euungulata (i.e., Perissodactyla et Artiodactyla). En l’absence de contraintes, l'analyse bayésienne soutient faiblement la possibilité que Thomashuxleya soit un xénarthre basal (en dehors du groupe-couronne) ; avec une contrainte imposant Notoungulata et Litopterna comme monophylétiques, les deux clades sont retrouvés comme groupe-frère de Perissodactyla. Les données anatomiques obtenues jusqu'à présent pour Thomashuxleya, combinées aux acides aminés du collagène des méridiongulés pléistocènes, limitent substantiellement le nombre d'affinités possibles pour les espèces endémiques à l'Amérique du Sud au sein des mammifères, même si l'ambiguïté est encore présente.
Mots-clés : Amérique du Sud ; Toxodontia ; Litopterna ; masse corporelle ; phylogénie
Translator: Antoine Souron
Deutsche Zusammenfassung
Ein außerordentlich gut erhaltenes Thomashuxleya externa Skelett (Mammalia, Notoungulata), aus dem Eozän von Patagonien, Argentinien
Wir beschreiben eines der ältesten Notoungulaten-Skelette mit assoziierten craniodentalen und postcranialen Elementen: Thomashuxleya externa (Isotemnidae) aus Cañadón Vaca in Patagonien, Argentinien (Vacanium Unterstufe der Casamayorium SALMA, mittleres Eozän). Wir schätzen das Gewicht mit Hilfe verschiedener Skelett-Elemente, beschreiben die Knochenhistologie und untersuchen die phylogenetische Position. Wir erfassen Unterschiede bei den Scapulae, Humerii, Ulnae und Radii des neuen Stückes im Vergleich mit anderen Stücken, die vorher diesem Taxon zugeschrieben wurden. Wir errechnen ein Gewicht von 84 ± 24.2 kg, und zeigen damit, dass Notoungulaten im mittleren Eozän ein hohes Gewicht erreicht hatten. Die Knochenhistologie zeigt, dass das neue Stück ein ausgewachsenes Skelett hatte. Das neue Material unterstützt die Platzierung von Thomashuxleya als ein frühes, divergentes Mitglied der Toxodontia. Unter den Plazentatieren favorisiert unsere phylogenetische Analyse mit kombinierter DNA, Kollagen und morphologischer Matrix nur eine limitierte Anzahl an phylogenetischen Verwandtschaftsbeziehungen, jedoch kann nicht genau entschieden werden zwischen einer möglichen Zugehörigkeit zu den Afrotheria oder Laurasiatheria. Ohne Constraint unterstützt Maximale Parsimonie Thomashuxleya und Carodnia mit den Afrotheria. Wenn Notoungulata und Litopterna als monophyletisch (mit Macrauchenia und Toxodon bekannt für Kollagene) constraint sind, werden diese Kladen am Stamm zu den Euungulata (i.e., Perissodactyla und Artiodactyla) rekonstruiert. Nicht-constraint unterstützt die Bayesianische Analyse schwach die Möglichkeit dass Thomashuxleya ein Stamm- Xenarthra ist; wenn Notoungulata und Litopterna als monophyletisch constraint sind, werden die zwei Kladen als Schwestertaxa zu den Perissodactyla installiert. Anatomische Daten, die bisher von Thomashuxleya gesammelt und mit Kollagen-Aminosäuren kombiniert wurden, limitieren die Anzahl möglicher Zugehörigkeiten für endemische südamerikanische Arten unter den Säugetieren erheblich, allerdings verbleiben immer noch Unklarheiten.
Schlüsselwörter: Südamerika; Toxodontia; Litopterna; Körpergewicht; Phylogenie
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
PE: An influential journal
 Palaeontologia Electronica among the most influential palaeontological journals
Palaeontologia Electronica among the most influential palaeontological journalsArticle number: 27.2.2E
July 2024

 A Review of Handbook of Paleoichthyology Volume 8a: Actinopterygii I, Palaeoniscimorpha, Stem Neopterygii, Chondrostei
A Review of Handbook of Paleoichthyology Volume 8a: Actinopterygii I, Palaeoniscimorpha, Stem Neopterygii, Chondrostei