

Volume 28.2
May–August 2025
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 28.2 May-August 2025
See all articles in 28.1 January-April 2025
See all articles in 27.3 September-December 2024
See all articles in 27.2 May-August 2024
Interested in submitting a paper to Palaeontologia Electronica?
Click here to register and submit.
 |
||||
 |
||||
Article Search
 Norbert Keutgen. ZFiPBR, Uniwersytet Techniczno-Przyrodniczy Bydgoszcz, ul. Bernardynska 6/8, 85-029 Bydgoszcz, Poland, and University of Natural Resources and Life Sciences DNW, 1190 Vienna, Austria. norbert.keutgen@boku.ac.at
Norbert Keutgen. ZFiPBR, Uniwersytet Techniczno-Przyrodniczy Bydgoszcz, ul. Bernardynska 6/8, 85-029 Bydgoszcz, Poland, and University of Natural Resources and Life Sciences DNW, 1190 Vienna, Austria. norbert.keutgen@boku.ac.at
Norbert Keutgen is a palaeontologist with a special interest in belemnite research, especially of the genera Belemnella and Belemnitella. He received his doctoral degree in Natural Sciences in 1996 on biostratigraphy and paleoecology of the lower Maastrichtian invertebrate fauna in the Maastricht (The Netherlands) – Aachen (western Germany) region. This area has remained his main area of paleontological interest. His current research focuses on the bioclast approach of P.J. Sjeuf Felder and intends to interpret this data with respect to influences of orbital forcing in order to deduce Milankovitch cycles.

 Zbigniew Remin. Faculty of Geology, University of Warsaw, Al. Żwirki i Wigury 93, 02-089 Warsaw, Poland. zbyh@uw.edu.pl
Zbigniew Remin. Faculty of Geology, University of Warsaw, Al. Żwirki i Wigury 93, 02-089 Warsaw, Poland. zbyh@uw.edu.pl
Zbyszek Remin completed his MSc and PhD theses at the Faculty of Geology of the University of Warsaw. His special interests concern the objectification of classification procedures in taxonomical study, as those of fundamental importance for the biostratigraphy and paleobiogeography, by the application of unsupervised Kohonen Neural Networks. He works mainly with upper Cretaceous cephalopod belemnites and ammonites from Europe and North America. Current studies are devoted to the Campanian and Maastrichtian belemnites form the US Atlantic Costal Plain (New Jersey) as well as selected aspects of sedimentological processes at the K-Pg boundary (Cretaceous-Paleogene boundary). Zbyszek is experimenter – he developed a new method of foraminifera extraction by the use of the liquid nitrogen [=LN2] as well as linked enigmatic Neoproterozoic structures with the impression of falling hailstones generated in his lab.
 John W.M. Jagt. Natuurhistorisch Museum Maastricht, de Bosquetplein 6-7, 6211 KJ Maastricht, the Netherlands. john.jagt@maastricht.nl
John W.M. Jagt. Natuurhistorisch Museum Maastricht, de Bosquetplein 6-7, 6211 KJ Maastricht, the Netherlands. john.jagt@maastricht.nl
John Jagt has been curator of palaeontology at the (municipal) museum since 1991, specializing in Late Cretaceous (Santonian-Maastrichtian) and earliest Paleogene (Paleocene) invertebrates such as echinoderms, decapods crustaceans, ammonoid and coleoid cephalopods and some other smaller groups. From time to time, he is also in charge of projects revolving around the excavation, transportation and subsequent preparation of skeletal remains of mosasauroid reptiles from the Maastrichtian type area. A team of around 25 private collectors, from all walks of life, consider the Maastricht museum as their home turf where amassing data on Late Cretaceous and early Paleogene faunas and floras are concerned; John serves as a local co-ordinator of these activities.
FIGURE 1. Map of Europe and Central Asia, illustrating the general distribution of representatives of the Belemnella kazimiroviensis group as a whole, without differentiation into particular species of the group. The origin of holotype specimens of representatives of this group recorded in the literature are as follows: Belemnella kazimiroviensis - Vistula section; Belemnella skolozdrownae - Stevns Klint; Belemnella pensaensis - Saratov/Penza; Belemnella arkhangelskii - Turgay Plateau.
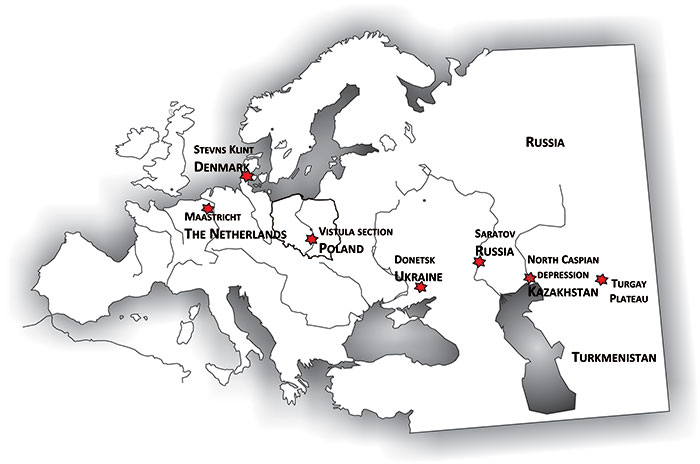
FIGURE 2. 1, location of the Middle Vistula River valley section in the Europe; 2, location in Poland together with the schematic extension of the Cretaceous and pre-Cretaceous deposits in Poland (without the Cenozoic cover); 3, position of Nasiłów along the Vistula River between the villages of Solec nad Wisłą and Kazimierz.
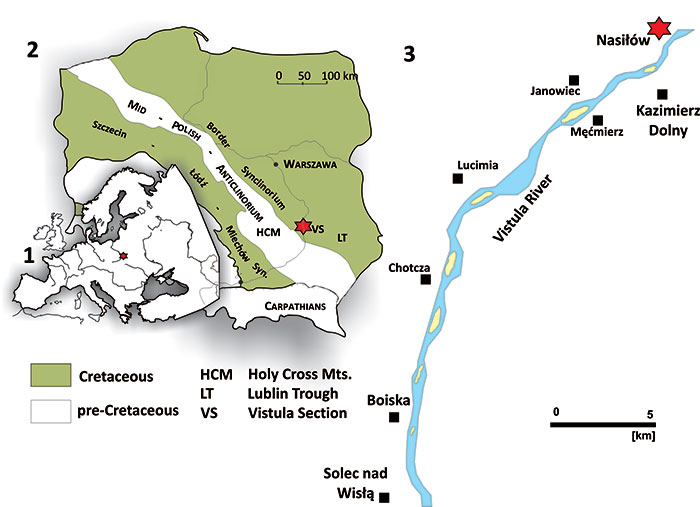
FIGURE 3. 1, Generalised K-Pg section at Nasiłów; 2, Details of the boundary interval and the position of the K-Pg boundary placed at the top of the Kazimierz Opoka; according to Machalski (1998).
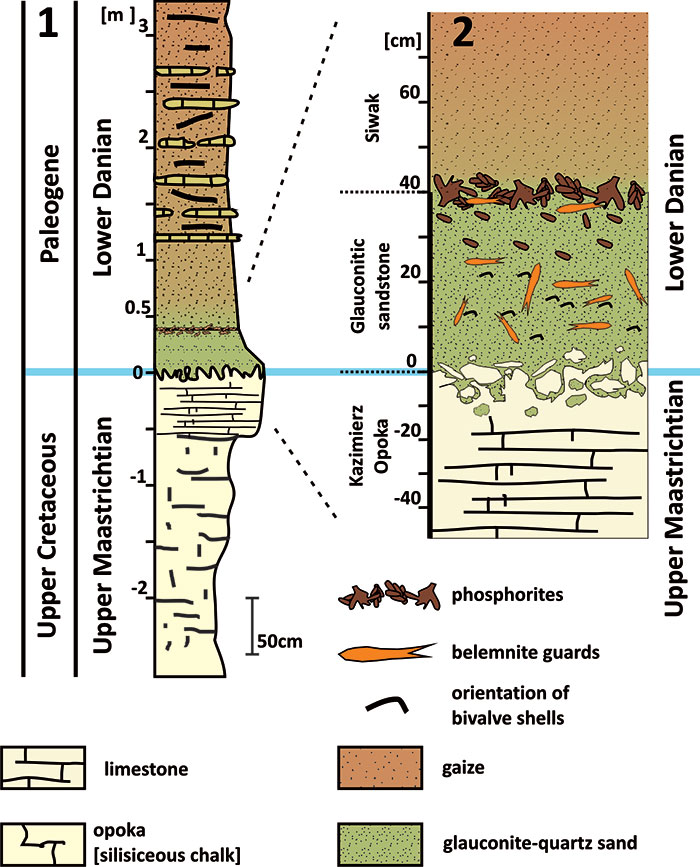
FIGURE 4. 1, Schematic map of southern Limburg (the Netherlands) showing the formerly available outcrops of the Meerssen Member (red stars) (after Jagt et al., 1996) that yielded representatives of the Belemnella kazimiroviensis group; black dots are smaller towns in the area; 2, Position of the Netherlands within Europe (red star).
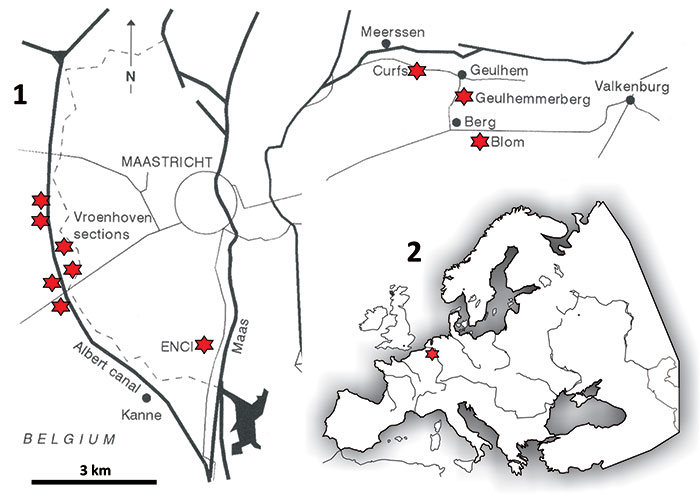
FIGURE 5. Age, lithostratigraphy, general lithology and subdivision into lithostratigraphical horizons and units (IVe-1 to Va-4) of the section formerly exposed at the Curfs-Ankerpoort quarry (Geulhem), with indication (arrow) of the K-Pg boundary, equating with the Berg en Terblijt Horizon at the top of subunit IVf-6 (modified after Jagt and Jagt-Yazykova, 2012). The range of belemnite species shown is that for the entire Maastricht area. R = remanié specimens.
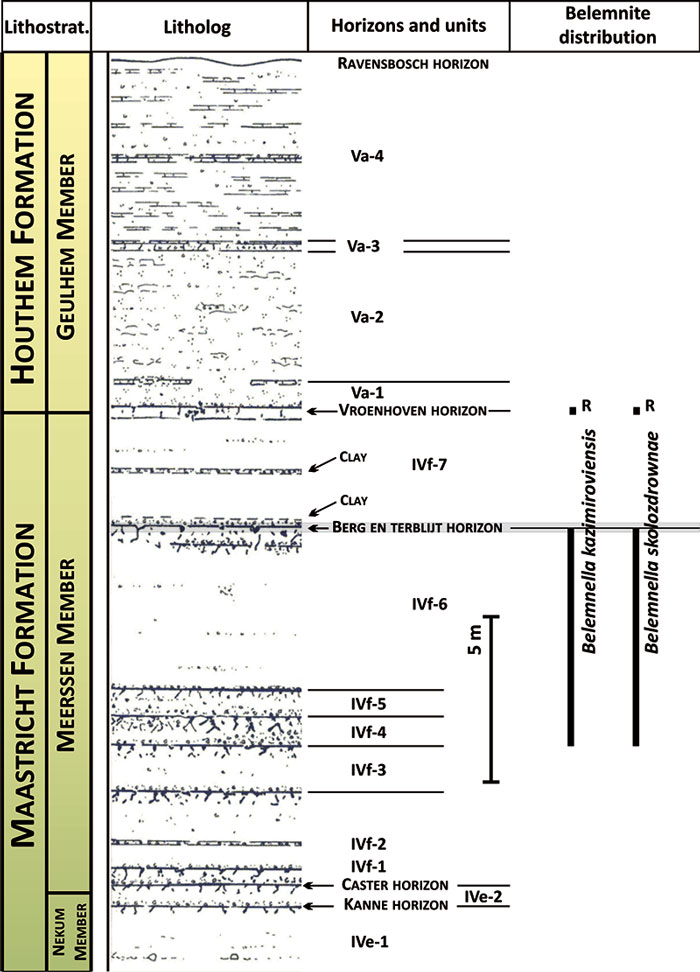
FIGURE 6. Diagram illustrating morphological features of the belemnite guard (lateral view of a longitudinally split guard on the left, ventral view on the right). AA = alveolar angle; FA = fissure angle; SD = Schatzky distance; Ls = length from apex to protoconch; Ds, Dp, Dc = dorso-ventral diameters at different part of the rostrum; Bs, Bp, Bc = lateral diameters at different part of the rostrum (after Keutgen, 1997; Keutgen et al., 2012).
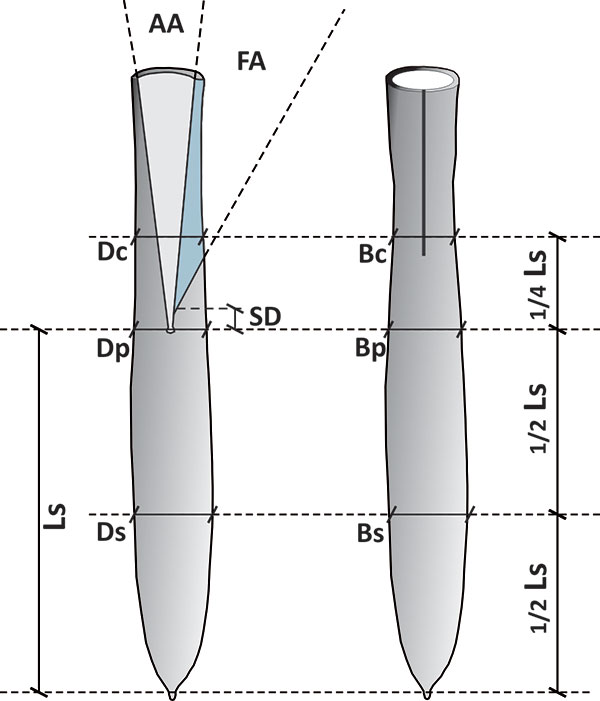
FIGURE 7. Relative distribution of size groups of Bln. gr. kazimiroviensis from the glauconitic sandstone and basal Siwak at Nasiłów (Poland).
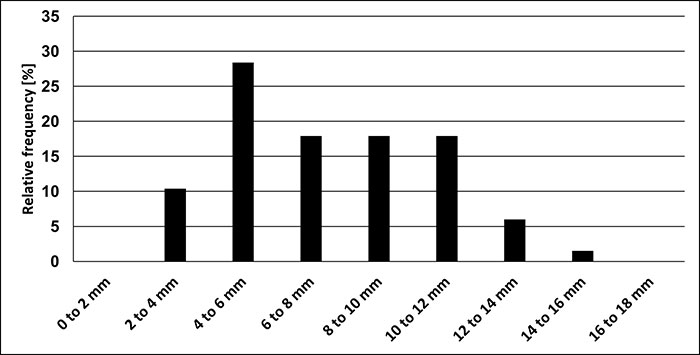
FIGURE 8. Relative distribution of size groups of Bln. gr. kazimiroviensis from the Maastricht area - Geulhemmerberg underground section above the Berg en Terblijt Horizon (the Netherlands) and Albert Canal sections above the Vroenhoven Horizon (Belgium).
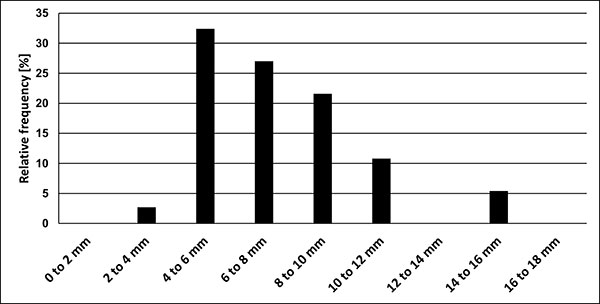
FIGURE 9. Relationship of BI (Birkelund Index) vs Dp (dorso-ventral diameter at the protoconch) (BI = Ls/Dp; compare with Figure 6) of all guards of the kazimiroviensis group from the Middle Vistula River valley, the Maastricht area, Denmark, central and southern Russia and Central Asia, inclusive of data of early ontogenetic stages measured on longitudinally split specimens. The growth relationship is best described by a logarithmic curve, indicating that small (young) specimens are relatively more slender than larger (adult) ones.
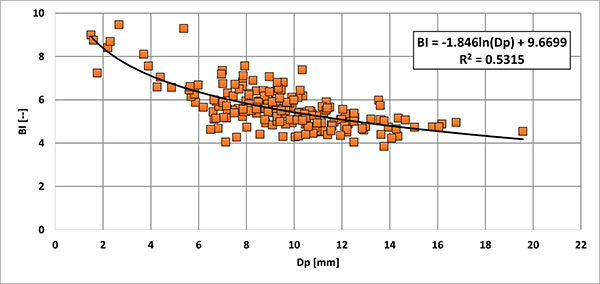
FIGURE 10. Two-dimensional plot Lsn (standardised length form apex to the protoconch) vs. AV (ventral aspect) of guards of Bln. kazimiroviensis and Bln. skolozdrownae from the Kazimierz Opoka (Kongiel Collection) from different localities (Kazimierz, Bochotnica and Nasilów), all except one from levels x and y sensu Pożaryski (1938).
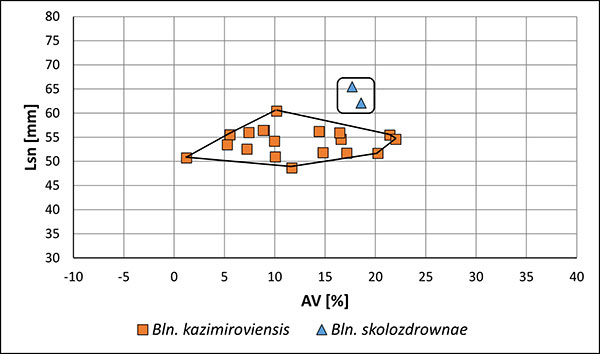
FIGURE 11. Two-dimensional plot for PC 1 and PC 2 derived from the PCA of belemnites of the kazimiroviensis group of the Kazimierz Opoka (Opoka) and the glauconitic sandstone (Greensand) of the Middle Vistula valley, as well as of those from the Maastricht area (the Netherlands, Belgium) (Meerssen).
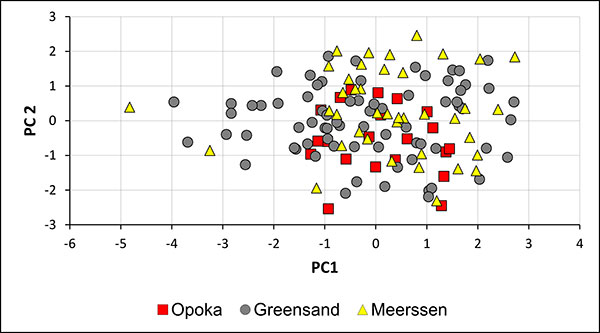
FIGURE 12. Two-dimensional plot Lsn (standardised length form apex to the protoconch) vs. AV (ventral aspect) of guards of Bln. kazimiroviensis and Bln. skolozdrownae from the glauconitic sandstone at Nasilów quarry. The dashed line indicates the variability of both species in the stratigraphically older Kazimierz Opoka.
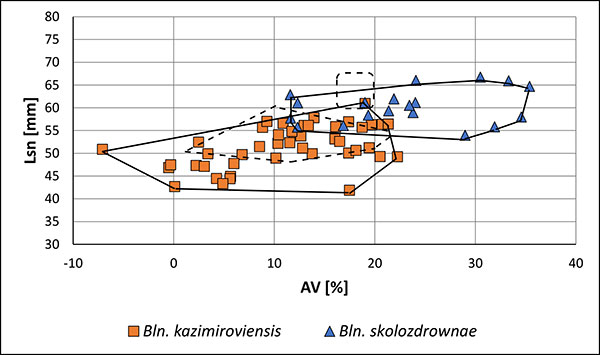
FIGURE 13. Two-dimensional plot Lsn (standardised length form apex to the protoconch) vs. AV (ventral aspect) of guards of Bln. kazimiroviensis from the basal Siwak at Nasilów quarry. The dashed lines indicate the variability of Bln. kazimiroviensis and Bln. skolozdrownae from the stratigraphically older glauconitic sandstone.
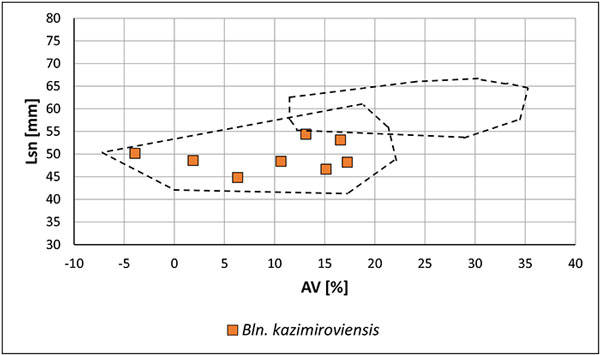
FIGURE 14. Two-dimensional plot Lsn (standardised length form apex to the protoconch) vs. AV (ventral aspect) of guards of Bln. kazimiroviensis and Bln. skolozdrownae from deposits in the Maastricht area (the Netherlands, Belgium). The dashed lines indicate the variability of Bln. kazimiroviensis and Bln. skolozdrownae from the Middle Vistula valley region (Poland).
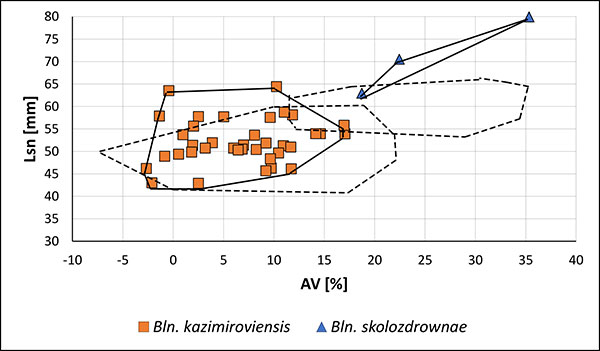
FIGURE 15. Two-dimensional plot Lsn (standardised length form apex to the protoconch) vs. AV (ventral aspect) of guards of Bln. kazimiroviensis from central and southern Russia and Central Asia, as illustrated by Naidin (1975). The dashed lines indicate the variability of Bln. kazimiroviensis and Bln. skolozdrownae from the Maastricht and the Middle Vistula valley regions.
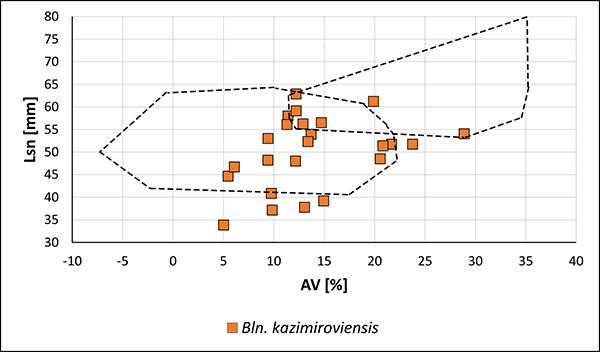
FIGURE 16. Two-dimensional plot Lsn (standardised length form apex to the protoconch) vs. AV (ventral aspect) of guards of Bln. kazimiroviensis and Bln. skolozdrownae from Denmark mentioned in the scientific literature. The holotypes of Bln. kazimiroviensis, proposed by Jeletzky (1951) and Kongiel (1962), are also indicated (blank squares), in addition to the holotype of Bln. skolozdrownae from Denmark. The dashed lines indicate the variability of Bln. kazimiroviensis
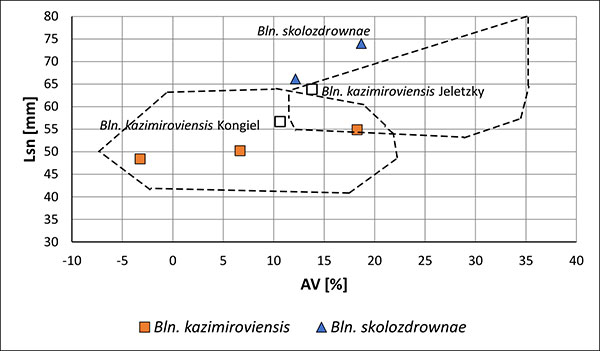
FIGURE 17. Belemnella (Neobelemnella) kazimiroviensis (Skołozdrówna, 1932). 1-4, NHMM JJ 4048/1, former Curfs-Ankerpoort quarry; basal Geulhem Member, Vroenhoven Horizon, + 0.00-0.05 m; 5-8, NHMM JJ 8339, former Blom quarry, Meerssen Member IVf-4; 9-12, NHMM MK 2516, Geulhemmerberg subterranean galleries near Geulhem, top of Meerssen Member (IVf-6); 13-16, MWGUW ZI/69/44, 17-20, ZI/69/47 and 21-24, ZI/69/51, respectively, from the "phosphatic layer" at Nasiłów quarry. All specimens are in natural size in following views: dorsal (1, 5, 9, 13, 17, 21), lateral (2, 6, 10, 14, 18, 22), longitudinally split specimen showing internal features (3, 7, 11, 15, 19, 23), and close-up of alveolar part [x2] with the course of the bottom of ventral fissure - dashed line (4, 8, 12, 16, 20, 24).
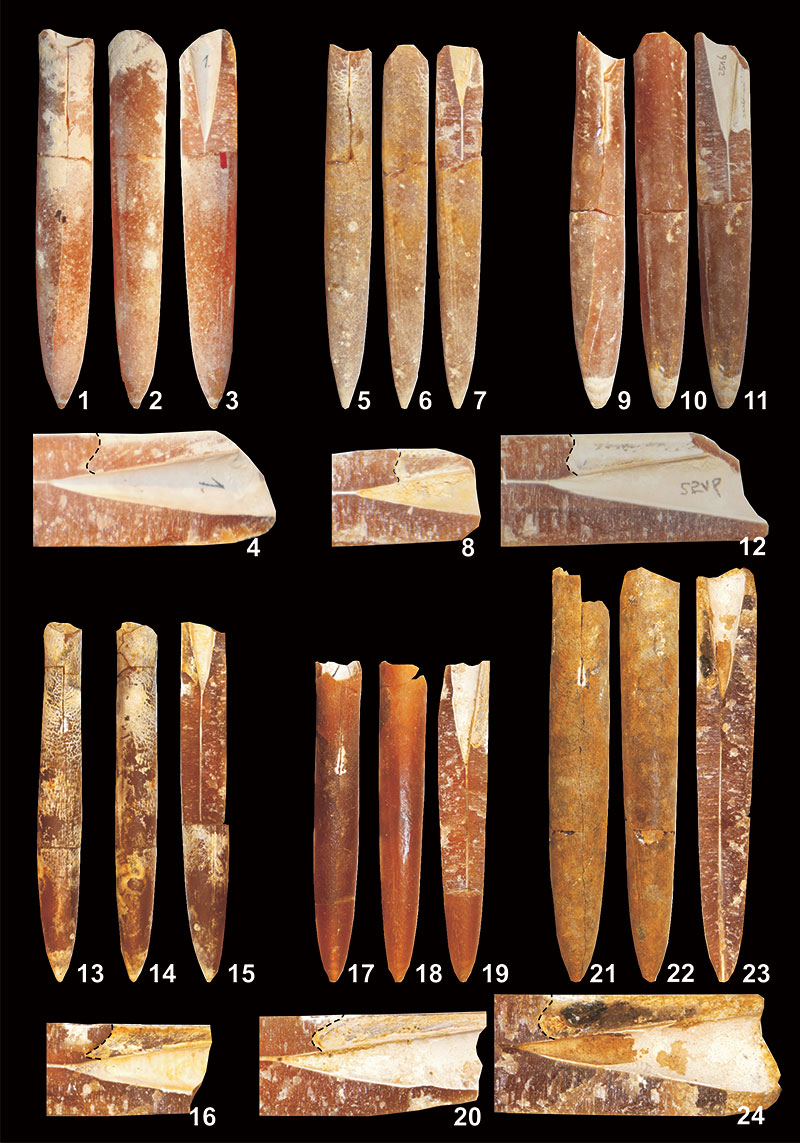
FIGURE 18. Relationship of BI (Birkelund Index) vs Dp (dorso ventral diameter at the protoconch) (BI = Ls/Dp; compare with Figure 6) of guards of Bln. kazimiroviensis from the Middle Vistula valley and the Maastricht area. The growth relationship is best described by a logarithmic curve.
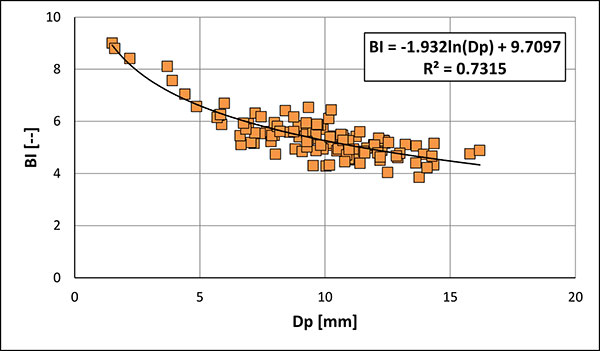
FIGURE 19. Belemnella (Neobelemnella) skolozdrownae (Kongiel 1962). 1-4, NHMM MK 2815, Albert Canal near Vroenhoven (61F-19), uppermost Meerssen Member; 5-8, NHMM JJ 8759, ENCI-HeidelbergCement Group quarry, Maastricht, Meerssen Member, IVf-4 or IVf-5; 9-12, MWGUW ZI/69/70, and 13-16, ZI/69/86 - from the glauconitic sandstone below the "phosphatic layer" at Nasiłów; 17-20, MWGUW ZI/69/60 and 21-24, ZI/69/61 - from the "phosphatic layer" at Nasiłów quarry. All specimens are in natural size in following views: dorsal (1, 5, 9, 13, 17, 21), lateral (2, 6, 10, 14, 18, 22), longitudinally split specimen showing internal features (3, 7, 11, 15, 19, 23), and close-up of alveolar part [x2] with the course of the bottom of ventral fissure - dashed line (4, 8, 12, 16, 20, 24).
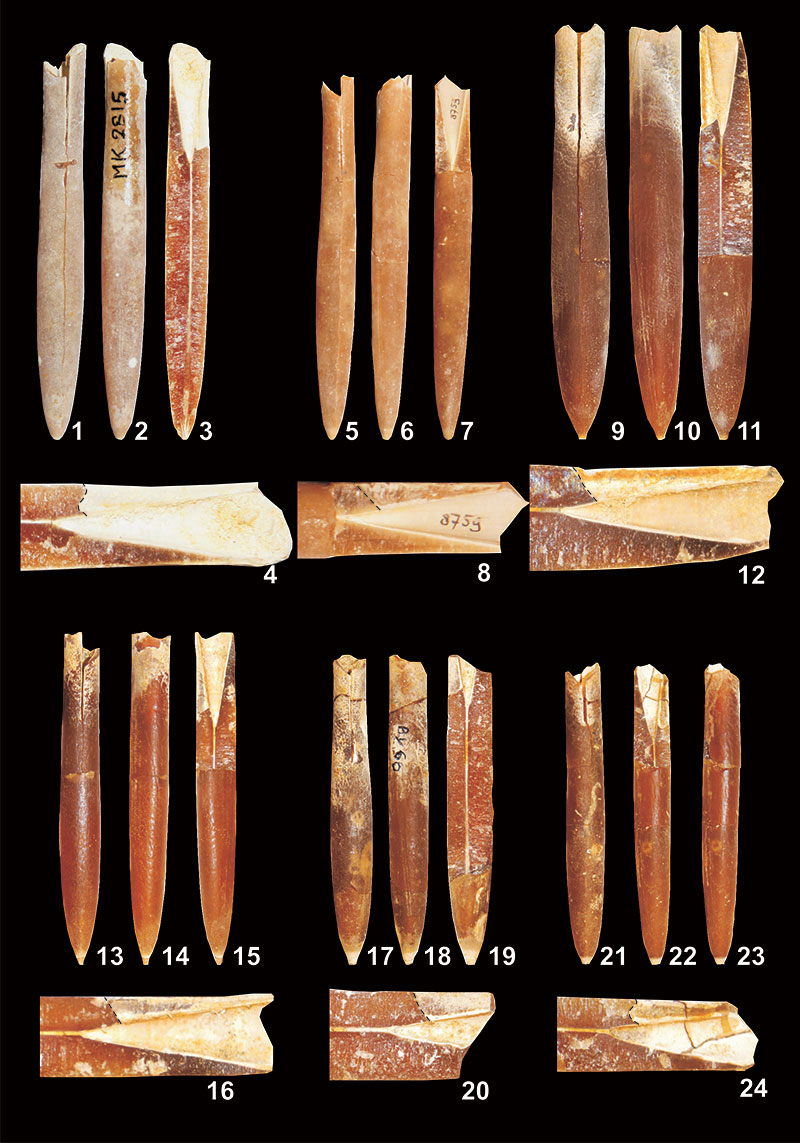
FIGURE 20. Relationship of BI (Birkelund Index) vs Dp (dorso ventral diameter at the protoconch) (BI = Ls/Dp; compare with Figure 6) of guards of Bln. skolozdrownae from the Middle Vistula valley in Poland and the Maastricht area in the Dutch-Belgian border region, in addition to two specimens from Denmark (blank triangles) as illustrated in the scientific literature (Jeletzky, 1951, pl. 7, figure 1; Birkelund, 1957, pl. 6, figure 4).
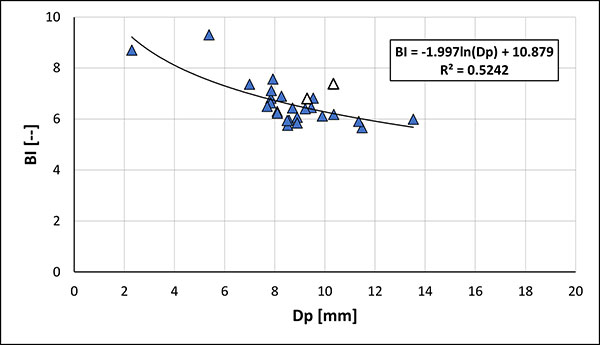
FIGURE 21. An excellently preserved specimen of Bln. kazimiroviensis (left) retaining part of the proostracum, together with a normally preserved guard (right) in the glauconitic sandstone at Nasiłów (field photograph; specimen with proostracum could not be collected completely).

FIGURE 22. Estimation of percentage of belemnites of the kazimiroviensis group remaining in the habitat, calculated from size-group distribution at Nasiłów (Poland) and the Maastricht area (the Netherlands, Belgium).
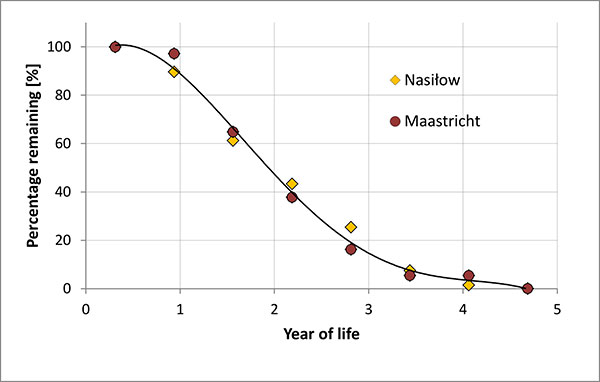
TABLE 1. Distribution of size groups of the stratigraphically youngest populations of Bln. gr. kazimiroviensis from Nasiłów (Poland) and from the Geulhemmerberg and Albert Canal sections near Maastricht (the Netherlands, Belgium). SD: standard deviation, N: number of specimens.
| Nasiłów, glauconitic sandstone - below “phosphatic layer” | Nasiłów, glauconitic sandstone, “phosphatic layer” | Nasiłów, basal Siwak (immediately above "phosphatic layer") | Nasiłów, glauconitic sandstone and basal Siwak | Maastricht, Geuhemmer-berg, above Berg en Terblijt Horizon | Maastricht, Albert Canal, above Vroenhoven Horizon | Maastricht, Geuhemmer-berg & Albert Canal | |
| Mean | 7.8 | 7.4 | 8 | 7.6 | 7.1 | 7.5 | 7.4 |
| Median | 7.9 | 7.4 | 7.2 | 7.8 | 6.7 | 6.8 | 6.8 |
| SD | 2.8 | 3.1 | 4.1 | 3.1 | 3.3 | 2.4 | 2.6 |
| Max | 12.6 | 13.2 | 14.1 | 14.1 | 14.3 | 14.4 | 14.4 |
| Min | 3 | 2 | 3.3 | 2 | 2.5 | 4.7 | 2.5 |
| N | 27 | 31 | 9 | 67 | 10 | 27 | 37 |
TABLE 2. Principal components (PC) co-ordinates of the variables, based on correlations (PCA). See the section Morphometrics of Material and Methods for measurement abbreviations.
| PC 1 | PC 2 | PC 3 | PC 4 | PC 5 | PC 6 | |
| Dp | 0.433 | -0.662 | 0.374 | 0.362 | -0.227 | -0.227 |
| SW | 0.46 | 0.021 | -0.836 | 0.101 | -0.263 | -0.097 |
| AA | -0.148 | 0.764 | 0.171 | 0.585 | -0.107 | -0.104 |
| FA | -0.788 | 0.087 | 0.07 | -0.391 | -0.33 | -0.321 |
| Lsn | -0.778 | -0.304 | -0.094 | 0.266 | -0.261 | 0.392 |
| AV | -0.713 | -0.311 | -0.323 | 0.305 | 0.365 | -0.252 |
TABLE 3. Eigenvalues of correlation matrix and related statistics of the PCA.
| PC | Eigenvalue | Total variance [%] |
Cumulative total variance [%] |
| 1 | 2.16 | 35.97 | 35.97 |
| 2 | 1.22 | 20.32 | 56.29 |
| 3 | 0.99 | 16.44 | 72.73 |
| 4 | 0.8 | 13.35 | 86.08 |
| 5 | 0.44 | 7.38 | 93.46 |
| 6 | 0.39 | 6.54 | 100 |
TABLE 4. Comparison of characteristic parameters of Bln. kazimiroviensis from the Nasiłów area (the Middle Vistula valley, Poland), the Maastricht area (the Netherlands, Belgium) and central and southern Russia as well as Central Asia (for abbreviations see the section Morphometrics of Material and Methods). Data represent means ± standard deviation. Different letters (a or b) within a row indicate significant differences by Tukey HSD test for unequal number of specimens (ANOVA) or Mann-Whitney U test in case the variances were even after data transformation not homogeneously distributed. In the latter case the significance level was corrected according to Bonferroni.
| Nasiłów area - Kazimierz Opoka | Nasiłów area - glauconitic sandstone | Nasiłów area - basal Siwak | Maastricht area | Central and southern Russia; Central Asia | |
| Ls [mm] | 57.2 ± 8.2 a | 52.6 ± 8.8 a | 47.1 ± 8.5 a | 50.9 ± 10.6 a | 53.3 ± 17.2 a |
| Dp [mm] | 11.0 ± 2.3 a | 10.4 ± 2.2 a | 9.3 ± 2.5 a | 9.5 ± 2.3 a | 10.7 ± 3.5 a |
| SW [mm] | 3.3 ± 0.6 a | 3.4 ± 0.8 a | 3.8 ± 0.9 a | 3.5 ± 0.7 a | 3.2 ± 0.7 a |
| AA [°] | 18.5 ± 1.1 ab | 18.5 ± 1.3 ab | 20.0 ± 0.7 a | 18.8 ± 1.4 ab | 18.0 ± 1.8 b |
| FA [°] | 73.4 ± 25.3 a | 73.2 ± 27.2 a | 59.8 ± 28.9 a | 72.9 ± 27.4 a | 63.0 ± 31.5 a |
| BI [--] | 5.3 ± 0.5 a | 5.2 ± 0.6 a | 5.1 ± 0.5 a | 5.4 ± 0.6 a | 5.0 ± 0.6 a |
| Lsn [mm] | 54.0 ± 2.8 a | 51.5 ± 4.6 a | 49.3 ± 3.2 a | 52.3 ± 5.0 a | 50.1 ± 7.9 a |
| AV [%] | 12.1 ± 5.8 ab | 11.1 ± 7.0 a | 9.6 ± 7.6 ab | 6.7 ± 5.4 b | 13.9 ± 6.0 a |
| AL[%] | -4.2 ± 4.4 a | -5.9 ± 6.5 a | -5.0 ± 5.4 a | -10.2 ± 5.4 a | -3.3 ± 9.2 a |
| AVs [%] | 7.4 ± 4.8 ab | 6.4 ± 6.1 ab | 5.8 ± 6.0 ab | 3.1 ± 4.8 b | 9.7 ± 5.7 a |
| ALs [%] | -5.0 ± 4.0 a | -6.2 ± 5.2 a | -6.4 ± 4.2 a | -9.6 ± 3.9 a | -3.6 ± 6.6 a |
TABLE 5. Mean values and standard deviation (SD) of critical characters of Bln. skolozdrownae from the Middle Vistula valley region and the Maastricht area (for abbreviations see the section Morphometrics of Material and Methods). Extreme values of a probably pathological specimen from Maastricht (NHMM 2015 NK001-18) are given in brackets and are not included in the calculation of the means.
| Means ± SD | Observed range | |
| Ls [mm] | 56.9 ± 7.6 | 49.0 - 81.0 |
| Dp [mm] | 8.9 ± 1.6 | 5.4 - 13.5 |
| SW [mm] | 3.1 ± 0.7 | 2.0 - 5.0 |
| AA [°] | 18.8 ± 1.5 | 16 - 21 |
| FA [°] | 107.2 ±20.3 | 70 - 138 |
| BI [--] | 6.4 ± 0.5 | 5.7 - 7.6 (9.3) |
| Lsn [mm] | 61.2 ± 4.1 | 54.0 - 70.5 (79.9) |
| AV [%] | 22.9 ± 7.6 | 11.6 - 35.4 |
| AL[%] | 4.7 ± 5.2 | -3.6 - 15.5 (30.8) |
| AVs [%] | 15.5 ± 5.8 | 6.6 - 30.2 |
| ALs [%] | 0.7 ± 3.5 | -4.9 - 8.4 (22.9) |
The late Maastrichtian Belemnella kazimiroviensis group (Cephalopoda, Coleoidea) in the Middle Vistula valley (Poland) and the Maastricht area (the Netherlands, Belgium) – taxonomy and palaeobiological implications
Plain Language Abstract
Near the end of the Cretaceous Period, during the late Maastrichtian, belemnitellid squid-like cephalopods of the Belemnella kazimiroviensis group appeared in central and western Europe, first in Poland (Middle Vistula River valley) and Denmark, later in the southeast Netherlands and northeast Belgium. This highly successful group probably had its origin in the eastern part of the Russian Platform, close to the beginning of the late Maastrichtian. Representatives of the Belemnella kazimiroviensis group dominated the belemnite record at the end of the Maastrichtian Stage, but became extinct, together with other members of the family Belemnitellidae, close to or at the Cretaceous-Paleogene (K-Pg) boundary.
We have studied the late Late Maastrichtian belemnitellid coleoids of the Belemnella kazimiroviensis group from the two key European sections, i.e., the Middle Vistula valley in central Poland and the Maastricht area in the southeast Netherlands and northeast Belgium and compared this material with material from Denmark, southern and central Russia, and Central Asia. In comparison to former researchers, we recognised only two species within this successful linage: Belemnella (Neobelemnella) kazimiroviensis (Skołozdrówna, 1932) and Belemnella (Neobelemnella) skolozdrownae (Kongiel, 1962). Populations of the kazimiroviensis group in the Nasiłów and Maastricht areas were both dominated by relatively small individuals, which might suggest that a significant number of specimens did not reach the reproductive age or changed habitat during their life cycle. Although part of the studied belemnites came already from the Paleogene deposits, we have found no evidence that they survived the catastrophe that happened some 65 millions of years ago, and this material for the time being must be interpreted as reworked.
Resumen en Español
El grupo Belemnella kazimiroviensis del Maastrichtiense tardío (Cephalopoda, Coleoidea) en el valle del Vístula Medio (Polonia) y el área de Maastricht (Países Bajos, Bélgica) - taxonomía e implicaciones paleobiológicas
Se han estudiado y comparado los Coleoidea del grupo Belemnella kazimiroviensis del Maastrichtiense tardío de secciones clave en el valle del Vístula Medio en Polonia central y del área de Maastricht en el sureste de los Países Bajos y el noreste de Bélgica con materiales de Dinamarca, Rusia meridional y central, y Asia central tal y como han sido figurados en la bibliografía. Se reconocen dos especies: Belemnella (Neobelemnella) kazimiroviensis (Skołozdrówna, 1932) y Belemnella (Neobelemnella) skolozdrownae (Kongiel, 1962). Las poblaciones del grupo kazimiroviensis en las zonas de Nasiłów y Maastricht estaban dominadas por individuos relativamente pequeños, lo que podría sugerir que un número significativo de especímenes no alcanzó la edad reproductiva o cambió el hábitat durante su ciclo de vida. Aunque parte de los belemnites estudiados provinieron de los depósitos del Paleógeno, no hemos encontrado ninguna evidencia que indique que sobrevivieron más allá del límite de K-Pg, por lo que se interpretan como ejemplares reelaborados.
Palabras clave: Cretácico tardío; Maastrichtiense superior; secciones del límite K-Pg; estratigrafía de belemnites; Europa
Traducción: Enrique Peñalver (Sociedad Española de Paleontología)
Résumé en Français
Le groupe Belemnella kazimiroviensis (Cephalopoda, Coleoidea) du Maastrichtien récent de la moyenne vallée de la Vistule (Pologne) et de la région de Maastricht (Pays-Bas, Belgique) – taxinomie et implications paléobiologiques
Les coléoïdes belemnitellidés du groupe Belemnella kazimiroviensis datant du Maastrichtien récent des sections clés de la moyenne vallée de la Vistule, au centre de la Pologne et dans la région de Maastricht au sud-est des Pays-Bas et au nord-est de la Belgique, ont été étudiés et comparés avec du matériel du Danemark, du sud et du centre de la Russie, et d'Asie centrale d'après les illustrations de la littérature. Deux espèces sont reconnues : Belemnella (Neobelemnella) kazimiroviensis (Skołozdrówna, 1932) et Belemnella (Neobelemnella) skolozdrownae (Kongiel, 1962). Les populations du groupe B. kazimiroviensis dans les régions de Nasiłów et Maastricht étaient dominées par des individus relativement petits, ce qui pourrait suggérer qu'un nombre important de spécimens n'atteignaient pas l'âge reproductif ou changeaient d'habitat pendant leur cycle de vie. Bien qu'une partie des bélemnites étudiées viennent de dépôts paléogènes, nous n'avons trouvé aucune preuve qu'elles aient survécu à l'extinction Crétacé-Tertiaire, et elles sont donc interprétées comme remaniées.
Mots-clés : Crétacé récent ; Maastrichtien supérieur ; sections de la limite Crétacé-Tertiaire ; stratigraphie des bélemnites ; Europe
Translator: Antoine Souron
Deutsche Zusammenfassung
Die Belemnella kazimiroviensis Gruppe (Cephalopoda, Coleoidea) aus dem späten Maastricht des Mittleren Vistula Tals (Polen) und des Maastricht Gebiets (Niederlande, Belgien) – Taxonomie und paläobiologische Rückschlüsse
Die belemnitelliden Coleoiden aus dem späten Maastricht der Belemnella kazimiroviensis Gruppe aus zentralen Gebieten des mittleren Vistula Tals in Zentralpolen und dem Maastricht Gebiet im Südwesten der Niederlande und in Nordost Belgien wurden untersucht und mit Material aus Dänemark, Süd-und Zentralrussland und Zentralasien aus der Literatur verglichen. Es wurden zwei Arten erkannt: Belemnella (Neobelemnella) kazimiroviensis (Skołozdrówna, 1932) und Belemnella (Neobelemnella) skolozdrownae (Kongiel, 1962). Die Populationen der kazimiroviensis-Gruppe im Nasiłów und Maastricht Gebiet werden beide von relativ kleinen Individuen dominiert, was möglicherweise darauf hindeutet, dass eine erhebliche Anzahl an Tieren das Fortpflanzungsalter nicht erreichte oder während des Lebenszyklus das Habitat wechselte. Obwohl ein Teil der untersuchten Belemniten aus paläogenen Ablagerungen stammt, haben wir keine Beweise gefunden, dass sie die K/Pg Grenze überlebt haben und so interpretieren wir diese Gegebenheit als aufgearbeitet.
Schlüsselwörter: späte Kreide; oberes Maastricht; K/Pg Grenzbereiche; Belemniten-Stratigraphie; Europa
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
PE: An influential journal
 Palaeontologia Electronica among the most influential palaeontological journals
Palaeontologia Electronica among the most influential palaeontological journalsArticle number: 27.2.2E
July 2024

 A Review of Handbook of Paleoichthyology Volume 8a: Actinopterygii I, Palaeoniscimorpha, Stem Neopterygii, Chondrostei
A Review of Handbook of Paleoichthyology Volume 8a: Actinopterygii I, Palaeoniscimorpha, Stem Neopterygii, Chondrostei