

 Cranial morphology of the Oligocene beaver Capacikala gradatus from the John Day Basin and comments on the genus
Cranial morphology of the Oligocene beaver Capacikala gradatus from the John Day Basin and comments on the genus
Article number: 17.2.25A
https://doi.org/10.26879/419
Copyright Palaeontological Association, June 2014
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 10 July 2013. Acceptance: 25 March 2014
{flike id=722}
ABSTRACT
The cranial morphology of the small Oligocene beaver Capacikala gradatus is described on the basis of a well preserved, nearly complete skull and partial mandibles from the John Day Formation, John Day Fossil Beds, Oregon, USA. The only nearly complete skull known so far from the same area as the type specimen is described here in detail. This is especially appropriate as the type specimen comes from an unknown locality within the John Day Formation and is only a fragmentary skull. The newly described specimen was found between dated marker beds, so that it can be no older than 28.7 Ma, nor younger than 27.89 Ma. Although Capacikala had been named 50 years ago (MacDonald, 1963), it is still not well known. Morphological comparisons are made to other mentioned or illustrated specimens of Capacikala, Palaeocastor and recent Castor; there are similarities and differences to both genera. The findings of the skull is discussed in comparison to the description of the genera Capacikala and Palaeocastor and some characters are revised. A phylogenetic analysis with few selected castorid species was performed, but resulted in poorly supported trees. However, a complete revision of beaver phylogeny and of the characters used is beyond the scope of the paper.
Clara Stefen. Senckenberg Naturhistorische Sammlungen Dresden, Museum of Zoology, Königsbrücker Landstrasse 159, 01109 Dresden, Germany, Clara.Stefen@senckenberg.de
KEY WORDS: Castoridae; Palaeocastorinae; skull; Tertiary
Final citation: Stefen, Clara. 2014. Cranial morphology of the Oligocene beaver Capacikala gradatus from the John Day Basin and comments on the genus. Palaeontologia Electronica Vol. 17, Issue 1;25A; 29p. https://doi.org/10.26879/419
palaeo-electronica.org/content/2014/722-skull-of-capacikala-gradatus
INTRODUCTION
The John Day Basin has long been known to contain an exceptional record of mid Eocene through late Miocene biotas, and has yielded a good record of castorids ranging from Oligocene to early Miocene Palaeocastor through middle Miocene Hystricops and Monosaulax up to late Miocene Dipoides and Castor material (Cope, 1884; Fremd et al., 1994; Hunt and Stepleton, 2004; Samuels and Zancanella, 2011). The most diverse and abundant assemblages are from those strata containing faunas characteristic of the Whitneyan through Arikareean North American Land Mammal Ages (NALMAs) exposed in the Turtle Cove region of Eastern Oregon (Leidy, 1870, 1871; Cope, 1879a; Merriam, 1901; Merriam and Sinclair, 1907; Shotwell, 1968; Fremd et al., 1994, 2000).
Early compiled inventories of the Tertiary faunas of the John Day Region (Merriam and Sinclair, 1907) listed two species of beaver, Steneofiber gradatus and Steneofiber peninsulatus. Both species have been assigned to Palaeocastor by Stirton (1935), who restricted the use of Steneofiber to European specimens. Steneofiber gradatus has been nominated as type species for the new genus Capacikala by MacDonald (1963) whereas Steneofiber peninsulatus is still understood to be a member of Palaeocastor (Xu, 1996; Rybczynski, 2007; Flynn and Jacobs, 2008; Calede, 2013).
The original assignment of both species to the same genus may serve as indicator of the closeness of the two species now assigned to the separated genera Capacikala and Palaeocastor. Martin (1987) erected the subfamily Palaeocastorinae and included the tribe Palaeocastorini with the genera Palaeocastor and Capatanka (understood to be a member of Palaeocastor by McKenna and Bell, 1998), and the tribe Capacikalini with Capacikala and Pseudopalaeocastor, thus setting Capacikala and Palaeocastor even slightly further apart. Korth (2001) retained the subfamily Palaeocastorinae without further tribal differentiation.
The palaeocastorine beavers were restricted to North America and are understood to represent an early, Oligocene-Miocene radiation of fossorial beavers that probably inhabited dry, sandy upland areas. The palaeocastorine beavers as reviewed by Martin (1987) include Capacikala, Capatanka (synonymized to Palaeocastor (McKenna and Bell, 1998)), Euhapsis, Fossorcastor (often understood to belong to Palaeocastor and referred to this genus hereafter), Palaeocastor and Pseudopalaeocastor.
The spiral burrows of the ichnogenus Daemonelix were originally thought to be associated with Palaeocastor (Peterson, 1905, 1906). More detailed analyses showed that three species of beaver were associated with these burrows, in decreasing size: Palaeocastor magnus, P. fossor, and Pseudopalaeocastor barbouri (Peterson, 1905; Martin and Bennett, 1977; Martin, 1994; taxonomic assignment according to Flynn and Jacobs, 2008). Capatanka, Capacikala, and Euhapsis have never been directly associated with Daimonelix. The earliest recognized member of the palaeocastorine beavers, Palaeocastor nebrascensis (Leidy, 1869), dates from the late Oligocene from South Dakota and the group persisted till the end of the Harrison Formation (McDonald, 1963; Martin, 1987). Apparently there are no descendants of this group of fossorial beavers. One of the main differences between them and semiaquatic beavers as pointed out by Stirton (1935) is the flattened enamel face of the incisors. Those however, are understood to be "inconsistent with currently recognized groups" (Flynn and Jacobs, 2008).
Here, a nearly complete skull of Capacikala gradatus from the John Day Basin is described. Comparisons to recent Castor and fossil Palaeocastor and other palaeocastorine beavers are given.
MATERIALS AND METHODS
Material
 In 1983 an NPS contractor, Dr. Hugh Wagner, collected an excellent skull of Capacikala gradatus from a locality within the Sheep Rock Unit of John Day Fossil Beds National Monument (JODA; Appendix 1). The precise locality information including GPS position are on file in the JODA collection. This specimen, HW68338 (subsequently catalogued as JODA 621 at the John Day Fossil Beds National Monument) was accompanied by field notes and a photograph. The latter indicates that the specimen comes from the greyish-green horizon in the middle of the exposure, which represents Turtle Cove Unit H (Figure 1). The ignimbrite Wagner referred to is the Picture George Ignimbrite, recently dated with SCLF techniques at 28.7 Ma. The specimen was in a concretion, and thought to be float; therefore it is possible that it could have possibly originated several meters higher in the column, but below the distinctive welded ash-flow Deep Creek Tuff, which is dated as 27.89 (Albright et al., 2001, 2008).
In 1983 an NPS contractor, Dr. Hugh Wagner, collected an excellent skull of Capacikala gradatus from a locality within the Sheep Rock Unit of John Day Fossil Beds National Monument (JODA; Appendix 1). The precise locality information including GPS position are on file in the JODA collection. This specimen, HW68338 (subsequently catalogued as JODA 621 at the John Day Fossil Beds National Monument) was accompanied by field notes and a photograph. The latter indicates that the specimen comes from the greyish-green horizon in the middle of the exposure, which represents Turtle Cove Unit H (Figure 1). The ignimbrite Wagner referred to is the Picture George Ignimbrite, recently dated with SCLF techniques at 28.7 Ma. The specimen was in a concretion, and thought to be float; therefore it is possible that it could have possibly originated several meters higher in the column, but below the distinctive welded ash-flow Deep Creek Tuff, which is dated as 27.89 (Albright et al., 2001, 2008).
 For comparison, material of Palaeocastor, Capacikala, and Capatanka was studied in the collections of the South Dakota School of Mines and Technology (SDSM, with a focus on the material MacDonald (1963) had mentioned), Los Angeles County Museum (LACM), American Museum of Natural History (AMNH), and the University California Museum of Paleontology (UCMP), including mainly one skull of Palaeocastor nebrascensis UCMP 114635 that was used for detailed comparisons. Material of Castor canadensis used for comparison was studied on different occasions in the Museum of Vertebrate Zoology at the University of California, Berkeley (MVZ) and of Castor fiber in the Senckenberg Natural History Collections Dresden, Museum für Tierkunde (MTD).
For comparison, material of Palaeocastor, Capacikala, and Capatanka was studied in the collections of the South Dakota School of Mines and Technology (SDSM, with a focus on the material MacDonald (1963) had mentioned), Los Angeles County Museum (LACM), American Museum of Natural History (AMNH), and the University California Museum of Paleontology (UCMP), including mainly one skull of Palaeocastor nebrascensis UCMP 114635 that was used for detailed comparisons. Material of Castor canadensis used for comparison was studied on different occasions in the Museum of Vertebrate Zoology at the University of California, Berkeley (MVZ) and of Castor fiber in the Senckenberg Natural History Collections Dresden, Museum für Tierkunde (MTD).
The measurements of skull and mandible were taken with digital calipers to the nearest 0.1 mm, and the dental dimensions were measured with a calibrated reticle under a stereomicroscope. The nomenclature of skull features and foramina is compiled from Freye (1959), Stirton (1965), Wagner (1983), and Wahlert (1977); the description of tooth morphology is based on Stirton (1935).
Phylogenetic Analysis
The phylogenetic relationships within the palaeocastorine beavers had been fairly unresolved (Rybcynski, 2007), and were better resolved by Flynn and Jacobs (2008). The generic assignment of species has not been uniform in the literature (Table 1).
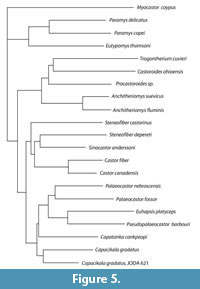
With the description of the presented skull of Capacikala, it seemed worthwhile to check at least with few taxa whether the resolution of a phylogenetic analysis within the palaeocastorine beavers could be enhanced. Only representative beavers were selected for the analysis (Appendix 2). In total 76 morphological characters of the skull, mandible, and teeth (Appendix 3) were chosen. The characters were partially selected from own observations (visits in several collections and additional analysis of published figures) and modified partially from other studies (Xu, 1996; Korth, 2001; Rybczynski, 2007). Recent Myocastor coypus and the early rodents Paramys delicatus, P. copei, and Eutypomys thomsoni were selected as out group taxa. The analysis was conducted using PAUP 4b for windows (Swafford, 1998) with all characters unordered and equally weighted. The bootstrap method with heuristic search and optimality criterion parsimony was selected. One hundred thousand bootstrap replicates were set. The tree resulting from the Neighbor-joining method is illustrated.
SYSTEMATIC PALEONTOLOGY
Order: Rodentia Bowdich, 1821
Family: Castoridae Hemprich, 1820
Genus: Capacikala MacDonald, 1963
Species: Capacikala gradatus (Cope, 1879b)
1879b Steneofiber gradatus Cope, 1879b, p. 63
1907 Steneofiber gradatus Merriam and Sinclair, 1907, p. 185 ff.
1884 Castorgradatus Cope, 1884, p. 839, 844, plate 63, figure 22
1935 Palaeocastor gradatus Stirton, 1935, p. 409, figure 42
1963 Capacikala gradatus, MacDonald, 1963, p. 197
1987 Capacikala gradatus Martin, 1987, p. 88. figure 11D, p. 90, figure 13
1996 Capacikala gradatus Xu, 1996, p. 420
Type: Capacikala gradatus – USNM 7008 (Cranium), according to Stirton (1935, figure 42)
Type Locality: John Day Region, Oregon
Horizon: John Day Formation, Late Oligocene
Description of the Skull
The skull presented herein (JODA 621, Figure 2, Figure 3) is nearly complete, only lacking part of the left zygomatic arch and part of the left pterygoid. Distortion from taphonomic processes is negligible. Although small, the skull seems robust and slightly elongated. The greatest width of the skull is across the zygomatic arch, and is about 44 mm (extrapolated over the broken side) and the length of the skull is ca. 60 mm (Table 2), thus it is slightly longer than wide (the length to width ratio is 1.36).
The skull is slightly inflated dorsally with the greatest extension and thus highest point of the skull just posterior to the orbital constriction and at the level of M1/2. Here is the greatest overall depth of the skull, which is similar to the overall depth at the auditory bullae. In lateral view the auditory bullae extend slightly more ventral than the tooth row, which seem to be the result of the buckling of the skull at the orbital constriction. The tips of the incisors extend ventrally to an imaginary line prolonged straight from the occlusal surface of the tooth rows.
The occipital plate is rectangular to a plane extending from the palate or skull base and tilts slightly anterior dorsally.
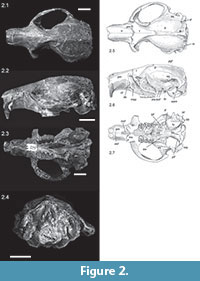 The orbital constriction is just anterior to the middle of the length of the skull, so that it appears to mark the middle of the skull. This narrow orbital constriction gives a kind of "hour glass" impression to the middle of the skull: a triangular shaped narrowing of the frontal posteriorly and an equally sharp but slightly more rounded triangular widening of the (fronto-)parietal posteriorly. The angle between the narrowing frontal and the posteriorly widening parietal at the interorbital constriction is 45°.
The orbital constriction is just anterior to the middle of the length of the skull, so that it appears to mark the middle of the skull. This narrow orbital constriction gives a kind of "hour glass" impression to the middle of the skull: a triangular shaped narrowing of the frontal posteriorly and an equally sharp but slightly more rounded triangular widening of the (fronto-)parietal posteriorly. The angle between the narrowing frontal and the posteriorly widening parietal at the interorbital constriction is 45°.
The frontal crests start as weak crests at the anterior root of the zygomatic arch, but become more prominent only just anterior to the orbital constriction. As the fronto-parietal crests are close to each other here, they form a kind of broad rim, but narrow gradually posteriorly fusing into the nuchal crest (Figure 2.1, 2.5). The junction of the sagittal crest to the lamdoidal crest is strong, broad, and triangular. The incompletely preserved lambdoidal crest is well elevated above the parietal.
The nasals are broadest in the anterior third of the rostrum and narrow posteriorly. The anterior end is broken, but from what is left it can be assumed that they extended further anterior than the incisors thus forming a kind of "roof" over the incisors (Figure 2.2, 2.6). The posterior end of the nasals is a fairly straight line level with the onset of the maxillary part of the zygomatic arch. Nasals and premaxillariess end at about the same level, and the premaxillary bones contribute more to the slightly zigzagging suture with the frontal than the nasals. The maxillary bone does only contribute marginally to the dorsal part of the snout; it meets the premaxillary only laterally at the very beginning of the zygomatic arch. The jugal does not meet the premaxillary. The lacrimal does not have a dorsal component, thus the dorsal part of the root of the zygomatic arch is very simple, consisting of the premaxillary and frontal only, and the maxillary to the lateral side, the zygomatic plate. Laterally on the snout, anterior to the root of the zygomatic arch, there is a slight depression in the premaxillary, but this does not continue into a groove dorsal to the incisor (left maxillary). The scar of the masseter muscle anterior to the zygomatic arch is bordered anteriorly by a slight rim or bulging which seems in diagonal continuation to the crista facialis and masseteric process, although the bulging actually continues anterior to these (Figure 2.4). The maxillary-premaxillary suture starts anterior to the zygomatic arch and extends nearly straight in dorso-ventral direction anterior to the crista facialis. It ends close to the posterior end of the incisive foramen. The crista facialis is nearly straight on the right side and slightly curved anteriorly on the left side of the skull. It ends ventrally in the subtle masseter superficialis process below and posterior to the infraorbital foramen. The infraorbital foramen and the masseter superfacialis process are located in the ventral part of the snout.
 The parietal, interparietal, and dorsal parts of the squamosal show strong rugosities and the cranium is well rounded laterally. The interparietal is triangular with the anterior end in line with the posterior end of the zygomatic arch. The parietal-squamosal sutures are straight to slightly curved but parallel to the sagittal crest, bending medially towards the orbit. The temporal foramen is visible posterior to the external auditory meatus. At the posterior root of the zygomatic arch the jugal and parietal or squamosal form an elongated ridge not quite continuing through to the lambdoidal crest. This contributes to an elongated glenoid fossa. The postglenoid foramina, a large one with a smaller one more anterior, are covered by this squamosal shelf.
The parietal, interparietal, and dorsal parts of the squamosal show strong rugosities and the cranium is well rounded laterally. The interparietal is triangular with the anterior end in line with the posterior end of the zygomatic arch. The parietal-squamosal sutures are straight to slightly curved but parallel to the sagittal crest, bending medially towards the orbit. The temporal foramen is visible posterior to the external auditory meatus. At the posterior root of the zygomatic arch the jugal and parietal or squamosal form an elongated ridge not quite continuing through to the lambdoidal crest. This contributes to an elongated glenoid fossa. The postglenoid foramina, a large one with a smaller one more anterior, are covered by this squamosal shelf.
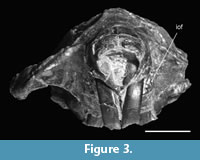
As visible in anterior view the infraorbital foramen is slender, narrow slitlike to slightly oval, bulging slightly more laterally. There is a slight bilateral asymmetry between the right and left infraorbital foramina (Figure 3). The crista facialis covering the infraorbital foramen is nearly straight dorso-ventrally and bulges around the infraorbital foramen. The infraorbital canal continues straight horizontally into the orbit.
Of the orbital foramen the lacrimal foramen is vaguely visible. Similarly the foramen ovale, masticatory and buccal foramen forming a triangle, are well preserved on the left side of the skull.
As visible in ventral view (Figure 2.3, 2.7), small elongated and oval interpremaxillary foramina are present anterior to the incisive foramen. The latter are slender and about one third of the length and right in the middle of the diastema. Their posterior end is about ventral to the masseter superficialis process and slightly posterior to the onset of the zygomatic arch. The maxillary grooves continue from the incisive foramina on to the maxillary. Due to a depression in the maxillary just at the P4 (Figure 2.3, 2.7), probably due to preservation, and a less well-preserved palatine, it is hard to see if the maxillary grooves continue on to the palatine. Only on the right part of the palatine a shallow groove is indicated.
The basioccipital shows no fossa occipitalis, but a depression exists on either side of the narrow crest in the midline. These depressions are bordered by high ridges adjacent to the auditory bullae, from the pterygoid region to the distal end of the auditory bullae. The right pterygoid extends to the bulla like in Castor. Whether it also ends in a hamulus and shows a hole like in Castor can not be discerned. More laterally an extension of the alisphenoid wing, the lateral pterygoid plate (Young, 1937) is present, but its extension and ending is not clearly visible as it is broken. Both form a wide fossa pterygoideus. The posterior end of the palatine in the midline is slightly damaged and level to the posterior end of M2.
There is a single hypoglossal foramen directly adjacent to the condyles in ventral view. Adjacent, projecting under the auditory bulla, is the slit-like and long lacerate foramen. The auditory bullae are mainly oval to slightly kidney shaped, well inflated ventrally, mainly on the medial side, and only slightly compressed laterally. Their greatest extension is parallel to the long axis of the skull, but the highest ridge of the auditory bulla is oriented at an angle of 40-45° to the long axis of the skull. Although the right and left external auditory meatus are slightly damaged it can be assumed that they extended further lateral than the occipital plane.
In posterior view, the supraoccipital shows two ridges, starting at the midline at the lambdoidal crest, but then diverging from each other. Between these arms of the crista occipitalis externa there are two clear, relatively large depressions representing attachment sites for the musculus rectus capitis (Figure 2.4).
The foramen magnum is well rounded; although there is a triangular impression due to the condyles, height and width are subequal. The condyle is similar to Castor, having a diagonal position, with the articular surface mostly ventrally oriented and only a small, tongue-like portion on the occipital plane. Ventrally the articular facets of the condyles do not meet. Neither mastoid nor paroccipital process are prominent and do not extend ventrally further than the condyle.
Description of the Mandible
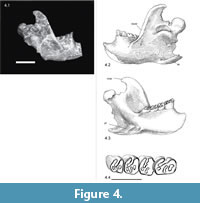 The right and left mandible are present (Figure 4), the left one with a damaged incisor and slightly damaged angular process, the right one with the anterior part missing and a damaged angular process and shelf. The coronoid process and condyles are present in both mandibles, and the angular process and angular shelf (crista pterygoidea of Freye, 1959) can be partially seen in the left mandible. The ventroanterior part of the mandibular bone is missing ventral to the incisors in both mandibles so the position and form of the chin process can not be described. A slight curvature of the mandibular bone between the chin process and the angular shelf can only be assumed. The single foramen mentale is approximately in the middle of the height of the mandibular ramus, and well anterior to the premolar and its alveolar bone (visible in the more fragmentary right mandible Figure 4.1).
The right and left mandible are present (Figure 4), the left one with a damaged incisor and slightly damaged angular process, the right one with the anterior part missing and a damaged angular process and shelf. The coronoid process and condyles are present in both mandibles, and the angular process and angular shelf (crista pterygoidea of Freye, 1959) can be partially seen in the left mandible. The ventroanterior part of the mandibular bone is missing ventral to the incisors in both mandibles so the position and form of the chin process can not be described. A slight curvature of the mandibular bone between the chin process and the angular shelf can only be assumed. The single foramen mentale is approximately in the middle of the height of the mandibular ramus, and well anterior to the premolar and its alveolar bone (visible in the more fragmentary right mandible Figure 4.1).
The linea obliqua ends in a slight depression, the tuberculum mandibulae, in about the middle of the height of the mandibular bone, ventral to the p4/m1 junction. The fossa masseterica is not developed as a deep overall groove, but rather is dominated by the proximal end of the incisor bulging outward labially, forming a knob-like rim. Thus a deep pocket is formed ventral to the coronoid process, similar to that seen in geomyids. The pocket is approximately at the same level as the chewing surface of the teeth and extends in an anterior-posterior direction over two thirds of the ascending ramus, ending at the neck of the condyle. The distal end of the incisor is dorsal to a plane level with the occlusal surface of the cheek teeth.
The condyle is the most posterior end of the jaw if the mandible is placed horizontally on the ventral rim of the mandibular ramus. No part of the linea obliqua of the ascending ramus is particularly enhanced or broadened. The angular process, of which the posterior end can be assumed to be preserved in the left mandible, ends about equally posterior than the condyle. The coronoid process does not extend as far posterior as the angular process and condyle.
In overall shape, the coronoid process is similar to Castor. The tip of the process is curved and points posteriorly and slightly ventrally and is only slightly broadened at the processus muscularis at the top. The condyle is knoblike but slender, with an anterior-posterior extension parallel to the main long axis of the mandible. The angular facet of the condyle on the labial side is tongue-shaped, merging smoothly into the extremely short and stout neck of the condyle, but is clearly bordered by a pronounced rim on the lingual side. Here the articular facet is more elongated in anterior-posterior direction and not extending as far ventrally as on the labial side.
The angular process is characterized by a ventral angular shelf forming a deep fossa pterygoideus lingually showing at least two parts (Figure 4.3). This angular shelf deviates laterally at an angle of nearly 45° to the long axis of the mandible. The orientation of the ventral side of the angular shelf is nearly parallel to the ventral rim of the mandibular ramus, but tilts dorsally towards its distal end. In ventral view the angular shelf is an elongated, slightly irregularly shaped rectangle with rounded edges.
In distal view the coronoid process, condyle and angular process form a clearly zigzagging line: the condyle is lingual to the coronoid process, and the angular process extends lightly further labial than the coronoid process. Thus the condyle is most offset lingually and none of the three processes are in a direct clear line.
Description of the Teeth
Incisors. The upper incisors are nearly orthodont, and triangular in cross section with a smooth and semiflat or very slightly convex only nearly straight enamel face. The cutting blade is straight and chisel shaped. The cross section of the lower incisor is only visible in the alveole of the broken right mandible. It is triangular with a flat enamel surface.
Maxillary Cheek Teeth. The maxillary tooth rows are diverging posteriorly and are tilted slightly labially. P4, M1, and M2 are nearly of the same width, but M3 is narrower (Table 3, Figure 2.7). P4 is rectangular, slightly longer than wide, whereas the M1 and M2 are rather more squared, or if rectangular wider than long. M3 is much smaller and more rounded. The teeth are subhypsodont and of medium to strong wear: the mesoflexus is closing in P4, and closed as mesofossette in all molars. A hypoflexus is still present in P4, M1, and M2, but closed as hypofossette in M3. The outer enamel band is fairly regular and preserved around all teeth except the left P4, where one side is broken.
In the P4, M1, and M2 the mesoflexus/fossette starts perpendicular to the labial side and curves slightly posteriorly towards the linguo-distal side of the tooth. A small parafossette is present adjacent to the tip of the hypoflexus/fossette. In the right P4 the parafossette is more elongated and appears to split into two fossettes. In the left P4 two fossettes are present in the same orientation.
M3 is much smaller, rounded, and strongly worn. There are three small, oval fossettes present, the hypofossette, and mesofossette in line across the tooth and the parafossette anterior to them.
 Mandibular Teeth. The mandibular tooth rows tilt medio-lingually. Like in the maxillary tooth row the premolar is the longest tooth with a rectangular outline. The molars are more squared in outline than the premolar and are rather broader than long (Table 3, Figure 4.4). The m3 is similar in outline, but smaller. The hypoflexids are still open in all teeth, whereas the mesoflexid is only open in the right m1 and closed as mesofossettid in the other teeth. The teeth show a basic beaver pattern of the fossettes with a hypoflexid being opposed by parafossette, mesoflexid/fossettid and metafossettid.
Mandibular Teeth. The mandibular tooth rows tilt medio-lingually. Like in the maxillary tooth row the premolar is the longest tooth with a rectangular outline. The molars are more squared in outline than the premolar and are rather broader than long (Table 3, Figure 4.4). The m3 is similar in outline, but smaller. The hypoflexids are still open in all teeth, whereas the mesoflexid is only open in the right m1 and closed as mesofossettid in the other teeth. The teeth show a basic beaver pattern of the fossettes with a hypoflexid being opposed by parafossette, mesoflexid/fossettid and metafossettid.
In the p4, the hypoflexid extends diagonally on the occlusal surface to near the distal end of the teeth but not to even half of the occlusal width. Directly adjacent to its posterior end is a very small rounded metafossettid. The mesoflexid/fossettid starts in the middle of the lingual side of the tooth and is curved slightly anteriorly, so that it is adjacent to the hypoflexid and extends over more than half of the tooth width. The parafossettid is elongated and oriented diagonally in the anterior part of the tooth.
Phylogenetic Analysis
The phylogenetic analysis with the selected set of taxa left 70 parsimony informative characters and gave a poor result for the bootstrap method with heuristic search clearly grouping in the 50 % consensus trees only both species of the genera Castor and Anchitheriomys and of Paramys together with Euhapsis. The Neighbor- Joining tree (Figure 5) shows the early rodents, Castoroidinae with Anchitheriomys, Castorinae and Palaeocastoridinae. The latter two appear as sister groups here. Within the palaeocastorines Capacikala gradatus and the JODA specimen are the sister group to other palaeocastorine beavers.
DISCUSSION
Morphologic Description and Comparison of Material
The occurrence of this remarkable specimen in a well-studied locality situated between precisely dated marker beds provides the first unequivocal datum for this species in a detailed stratigraphic context. The specimen can be no older than 28.7 Ma, nor can it be younger than the Deep Creek tuff, a marker bed well-dated at 27.89 Ma (Fremd et al., 1994; Albright et al. 2001, 2008); based on proximity to the latter unit it is probably 28 Ma old. Based on this temporally constrained occurrence, it appears that the species occurs in the John Day basin at roughly the same time that it is recognized in the Great Plains, correlative with the beginning of the second division of the Arikareean NALMA, or Ar2.
Comparisons to other material of Capacikala gradatus, Palaeocastor (mainly P. nebrascensis) and P. peninsulatus as depicted by Calede (2013) and recent Castor (based on Castor canadensis from MVZ) are given.
Compared to the few illustrations of some aspects of skull morphology of Capacikala, the present skull differs in the zygomatic plate area from the illustration by Martin (1987, text-figure 11D) in being dorsally flat in the area above the root of the zygomatic arch and from the illustration by Xu (1996, text-figure 6) in lacking a depression dorsal to the incisor. JODA 621 also differs from Capacikala gradatus illustrated in Xu (1996, text-figure 5B) where the infraorbital foramen is indicated further ventrally, a groove is dorsal to the incisor, the incisors are further recurved and a lacrimal is indicated.
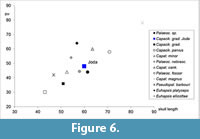 With a width of nearly 60 mm the present skull assigned to Capacikala gradatus is longer than Capacikala from the Wounded Knee, Sharps Formation (Figure 6) and the given range of 50-55 mm for this species (Xu, 1996). It is slightly smaller than several Palaeocastor skulls measured for comparison here, but falls within the size range of the skull length given for Palaeocastor peninsulatus with 50-70 mm and is only slightly longer than the range given for P. nebrascensis with 53-58 mm (Xu, 1996). Only Calede (2013) gives a larger minimum skull length for one specimen of P. peninsulatus (JODA 15820: 72.35 mm).
With a width of nearly 60 mm the present skull assigned to Capacikala gradatus is longer than Capacikala from the Wounded Knee, Sharps Formation (Figure 6) and the given range of 50-55 mm for this species (Xu, 1996). It is slightly smaller than several Palaeocastor skulls measured for comparison here, but falls within the size range of the skull length given for Palaeocastor peninsulatus with 50-70 mm and is only slightly longer than the range given for P. nebrascensis with 53-58 mm (Xu, 1996). Only Calede (2013) gives a larger minimum skull length for one specimen of P. peninsulatus (JODA 15820: 72.35 mm).
For the comparison to Palaeocastor in particular a skull which had been assigned to Palaeocastor by Xu (1996; UCMP 114635) has been used. Although the comparison is focused on this skull, other material especially from the SDSM collection with their taxonomic assignment given by MacDonald (1963) has also been considered here.
The overall similarity on skull shape, size, and different features is great. Capacikala and Palaeocastor share: relatively short skull, broad zygomatic arches; narrow interorbital constriction nearly in the middle of the skull length; shape of the interorbital constriction; course and shape of the fronto-parietal crests and sagittal crest (which of course changes with age as well known for Castor fiber (e.g., Hinze, 1950) and also illustrated for Palaeocastor peninsulatus (Calede, 2013); rugosities on the parietale and interparietale; posteriorly diverging tooth rows; position and shape of incisive foramen; maxillary grooves starting at the posterior end of the incisive foramen (palatine grooves can only be assumed for the present skull of Capacikala, but were observed on other material so also palatal grooves seem to be a shared feature as well); slightly inflated auditory bullae, relatively large; no fossa basioccipitalis but rather a crest in the midline of the basioccipital (as observed in the material studied); maxillary incisors not extending much ventrally. Also the mandible is very similar in general outline, zigzag arrangement of the processes in distal view; knoblike bulging of the posterior end and base of the incisor laterally in the masseteric fossa and the development of angular shelf.
 The orientation and tilt of the angular shelf in the mandible is also very similar in both genera. The ventral surface of the angular shelf is tilted dorsally (angle between a line at ventral rim of mandibular ramus and ventral surface of angular shelf) with about 20° to the long axis of the mandible in Capacikala and with about 23° in Palaeocastor (UCMP 114635), but appears even more tilted dorsally in distal view. The angular shelf projects laterally in an angle (measured in ventral view between a line at ventral rim of mandibular ramus and line in the middle of the angular shelf) of about 35° to the long axis of the mandible in Capacikala, and this angle in Palaeocastor is about 30°.
The orientation and tilt of the angular shelf in the mandible is also very similar in both genera. The ventral surface of the angular shelf is tilted dorsally (angle between a line at ventral rim of mandibular ramus and ventral surface of angular shelf) with about 20° to the long axis of the mandible in Capacikala and with about 23° in Palaeocastor (UCMP 114635), but appears even more tilted dorsally in distal view. The angular shelf projects laterally in an angle (measured in ventral view between a line at ventral rim of mandibular ramus and line in the middle of the angular shelf) of about 35° to the long axis of the mandible in Capacikala, and this angle in Palaeocastor is about 30°.
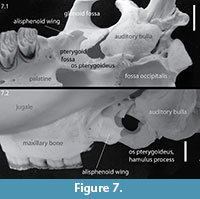
And in both genera a pterygoid fossa is visible. However, the exact extension of the alisphenoid wing, visible as a line level to the middle of the glenoid fossa, and pterygoid bone is not clear due to poor preservation. The pterygoid might well have extended to the bulla as this shows a small anterior projection similar to Castor (Figure 7).
Subtle differences of the present skull from Palaeocastor are the following: Palaeocastor shows a clear depression dorsal to the incisor on the premaxillary; the crista facialis is slightly more diagonally curved in lateral view, and the masseter superficialis process is slightly stronger; slightly more pronounced and longer paroccipital processes; more elongated squamosal crest or shelf, extending from the glenoid fossa nearly into the lambdoidal crest forming the glenoid fossa (lateral view); zygomatic arch clearly more extended dorsoventrally in Palaeocastor (e.g., P. peninsulatus Calede, 2013, figure 2d) than in the preserved JODA skull; palatal notch ends slightly more posteriorly in Palaeocastor (level with anterior part of M3 (Stirton, 1935; MacDonald, 1963; Calede, 2013, figure 1D)). In P. nebrascensis (UCMP 114635) the palate is not complete and could not be assessed. Cope (1884, plate LXIII, figure 18B), however, indicated a palatal notch level with the posterior end of M2 for P. peninsulatus. As nothing about the variability of this feature is known, it is difficult to comment on Cope's observation.
A difference given between Capacikala and Palaeocastor in the literature (Xu, 1996) is the more elongated crista facialis (surrounding infraorbital foramen) in Palaeocastor. From the skulls studied this seems to be true in one case and can not be commented upon in other cases. The upper incisors are very slightly convex, the lower ones have a flat surface. In Palaeocastor there are some subtle differences between species (Calede, 2013). The protrusion of the upper incisors from the alveoles (length of free incisor) is relatively short in the JODA specimen and lingually is as long as the wear facet. The tip of the incisor is flat and therefore chisel shaped. Overall this is similar to other species of Palaeocastor.
Compared to modern Castor the nasomaxillary-frontal suture is slightly more anterior, level with the anterior part of the zygomatic arch and nearly straight and not elongated and pointed. In Castor there is a small praeorbital process just posterior to the maxillary root of the zygomatic arch which is not present in this skull of Capacikala or Palaeocastor. In Castor the orbital constriction is not as narrow and more anterior in the skull than in Capacikala so that the skull does not have the strong hourglass appearance. And therefore the orbit is relatively larger in Capacikala. For Castor it is known that the course and development of the (fronto-)parietal ridges changes with age (Hinze, 1950). However, even in adult recent beavers the sagittal crest itself is narrow, and there is not the development of any broader rim-like structure like in the JODA skull. The zygomatic arch, especially the jugal shows a greater dorso-ventral extension in Castor.
In ventral view the most conspicuous difference is the absence of an occipital fossa in Capacikala. Also, the maxillo-premaxilla suture intersects the incisive foramen more anterior and the auditory bullae extend slightly further ventrally in Capacikala, slightly more so than the molars. The distal end of the palate at the midline is in Castor posterior to the M3, in Capacikala about level with the posterior end of M2. An alisphenoid wing is present in Capacikala and its posterior extension could well have been longer, than in Castor, maybe even to the bulla, but is broken. Also in ventral view the postorbital constriction in Castor is wide, the alisphenoid wing or lateral pteryogoid plate (Figure 7) ends level to the glenoid fossa and thus is positioned anterior to the auditory bullae. In Capacikala the postorbital constriction appears shorter anterior-distally and the glenoid fossa extends further posteriorly, ending posterior to the anterior rim of the auditory bulla.
And compared to Castor there is no crest or rim at the cranial side of the glenoid fossa, but the rim at the base of the zygomatic arch is quite marked in Capacikala.
There are some striking similarities to modern Castor in the lateral view of the snout: there is a broad plate for the attachment of the superficial masseter muscle on the anterior zygomatic plate with a similar relative extension. The curvature of the incisors and their extension or amount of protrusion from the alveoli are similar, as well as the procumbency (Table 4). The same holds for the lack of a marked groove dorsal to the incisor. This feature might need to be checked for variability in modern Castor to be able to assess variability also in fossil taxa of which cranial material is usually rarer. In Castor the crista facials is slightly more obliquely oriented, and a crest extends from it dorsally to the onset of the groove for the masseter muscle. In the JODA specimen the crista facialis is nearly straight and does not extend in marked crests.
A major difference in the mandible is the orientation of the proximal end of the incisors: more lingual in Castor, so that a more pronounced masseteric fossa is visible, and labially causing a bulging in Capacikala. The condyle is broader and more knoblike in Castor and the angular shelf is not as broad. The pterygoid fossa is not as deep in Castor.
Similarities to Castor include the orientation of the occipital plane to the base of the skull, the curvature of the premaxillary and maxillary in the diastema, the grooves on the maxillary bone from the incisive foramen posteriorly and possibly also on the palatine, overall inflation of the skull, the highest point of the skull in lateral view is at the orbit, extension of pterygoid to bulla (although it is not clear if the large foramen is also present in Capacikala), orientation of the crista facialis and masseter superficialis process. The masseter superficialis process is inconspicious in both taxa. The rugosities on the parietal and interparietal are similar, and they also share the (nearly) flat faced incisors (lower incisors are flat and upper ones slightly convex, particularly in young specimens). In both a line extended straight from the occlusal surface of the cheek teeth intersects the wear facet or tip of the incisor.
Comments on the Genus – History and Diagnosis
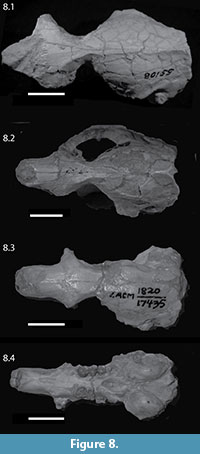 The genus Capacikala was named 50 years ago by MacDonald (1963) and has been included in the revision of palaeocastorine beavers by Martin (1987) (although he does not actually describe Capacikala in the text, but includes it in his proposed phylogeny), and in the overall phylogenetic analyses of Castoridae by Xu (1996), Rybczynski (2007), and Flynn and Jacobs (2008). Despite this work, it seems still poorly understood, not least because a good morphological analysis of the skull is lacking. And although there is abundant material from the Wounded Knee, Sharps Formation (Martin, 1987) and some from Muddy Creek Wyoming (Xu, 1996), most of it is more fragmentary than the skull described herein (Figure 8). Two skulls assigned to Capacikala gradatus have been illustrated, but without detailed descriptions (Cope, 1884, plate 63, figure 22; Stirton, 1935, figure 42). Even though Cope (1884) described a skull of ("Castor") Palaeocastor peninsulatus in detail with comparisons to recent Castor and Steneofiber, he was not as detailed on "Castor" (now considered Capacikala) gradatus. He gives an illustration of a nearly complete skull from the John Day River in Oregon with incomplete zygomatic arches (Cope, 1884, plate 63, figure 22). The skull in this figure is more complete than the type as mentioned and figured by Stirton (1935, figure 42). Cope (1884) indicated the only estimated extent of the zygomatic arches as dotted lines. These are much less expanded than in the JODA skull, indicating Cope's estimate is probably a false reconstruction. This image has been adapted in outline by Martin (1987, figure 13). In dorsal view, the skull illustrated by Cope is similar to the JODA skull in the form of the interorbital constriction, position of the interorbital constriction in relation to skull length, and the form and course of the fronto-parietal and sagittal crest. The snout, however, appears shorter and the postorbital constriction even less extended anterior-posteriorly in Cope's illustration. The skull length given by Cope (1884) of 50 cm is similar to that of a specimen from South Dakota (SDSM 5489).
The genus Capacikala was named 50 years ago by MacDonald (1963) and has been included in the revision of palaeocastorine beavers by Martin (1987) (although he does not actually describe Capacikala in the text, but includes it in his proposed phylogeny), and in the overall phylogenetic analyses of Castoridae by Xu (1996), Rybczynski (2007), and Flynn and Jacobs (2008). Despite this work, it seems still poorly understood, not least because a good morphological analysis of the skull is lacking. And although there is abundant material from the Wounded Knee, Sharps Formation (Martin, 1987) and some from Muddy Creek Wyoming (Xu, 1996), most of it is more fragmentary than the skull described herein (Figure 8). Two skulls assigned to Capacikala gradatus have been illustrated, but without detailed descriptions (Cope, 1884, plate 63, figure 22; Stirton, 1935, figure 42). Even though Cope (1884) described a skull of ("Castor") Palaeocastor peninsulatus in detail with comparisons to recent Castor and Steneofiber, he was not as detailed on "Castor" (now considered Capacikala) gradatus. He gives an illustration of a nearly complete skull from the John Day River in Oregon with incomplete zygomatic arches (Cope, 1884, plate 63, figure 22). The skull in this figure is more complete than the type as mentioned and figured by Stirton (1935, figure 42). Cope (1884) indicated the only estimated extent of the zygomatic arches as dotted lines. These are much less expanded than in the JODA skull, indicating Cope's estimate is probably a false reconstruction. This image has been adapted in outline by Martin (1987, figure 13). In dorsal view, the skull illustrated by Cope is similar to the JODA skull in the form of the interorbital constriction, position of the interorbital constriction in relation to skull length, and the form and course of the fronto-parietal and sagittal crest. The snout, however, appears shorter and the postorbital constriction even less extended anterior-posteriorly in Cope's illustration. The skull length given by Cope (1884) of 50 cm is similar to that of a specimen from South Dakota (SDSM 5489).
 Cope (1884) indicated for C. gradatus that the P4/p4 are the longest teeth in the tooth row and that size decreases regularly to the last tooth. This is also visible at the JODA skull and is shared with other castorid taxa (e.g., Steneofiber, Palaeocastor, Castor). "The middle line of the basioccipital bone is keeled, with a fossa on each side" (Cope, 1884, p. 845) – a feature well visible on the JODA skull as well. The illustration of the ventral view of the skull (Cope, 1884) indicates the presence of maxillary grooves, a palate terminating posteriorly level with the anterior part of M3 and fairly rounded auditory bullae. In this view no clear postorbital constriction can be discerned. Cope (1884) writes that the tympanic meatus is prolonged, which is difficult to support with the illustration or the JODA skull. In the skull illustration by Cope (1884) the os pterygoideus seems to continue from the distal end of the tooth row to the bulla; at least it is probably very long. The extension and form of the alisphenoid wing can not be clearly assessed but seems long as well in the JODA specimen.
Cope (1884) indicated for C. gradatus that the P4/p4 are the longest teeth in the tooth row and that size decreases regularly to the last tooth. This is also visible at the JODA skull and is shared with other castorid taxa (e.g., Steneofiber, Palaeocastor, Castor). "The middle line of the basioccipital bone is keeled, with a fossa on each side" (Cope, 1884, p. 845) – a feature well visible on the JODA skull as well. The illustration of the ventral view of the skull (Cope, 1884) indicates the presence of maxillary grooves, a palate terminating posteriorly level with the anterior part of M3 and fairly rounded auditory bullae. In this view no clear postorbital constriction can be discerned. Cope (1884) writes that the tympanic meatus is prolonged, which is difficult to support with the illustration or the JODA skull. In the skull illustration by Cope (1884) the os pterygoideus seems to continue from the distal end of the tooth row to the bulla; at least it is probably very long. The extension and form of the alisphenoid wing can not be clearly assessed but seems long as well in the JODA specimen.
Cope (1884) mentions also differences to P. peninsulatus: the palate is narrower; the maxillary diastema is relatively shorter; and "the temporal ridges do not unite so early" in C. gradatus.
Even though MacDonald (1963) named the genus Capacikala he did not figure teeth, skull or mandible nor even fragments. His diagnosis for the genus includes small size and a "mastoid breadth equal to or slightly greater than distance from interorbital constriction to posterior edge of cranium at junction of sagittal crest and lambdoidal crest." The latter is supported by the JODA specimen. The specimen is also small in size, but small size is – of course – relative (Figure 6). The skull fragment illustrated by Stirton (1935, figure 42, USNM 7008 and type specimen for Capacikala gradatus) would only allow one to roughly estimate the mastoid breadth.
 The JODA specimen also quite supports MacDonald's (1963) diagnosis of the palatal notch terminating opposite M2 and large, rounded auditory bullae. These expand ventrally slightly more than the tooth rows; but only very slightly so, in overall appearance both extend to about the same level.
The JODA specimen also quite supports MacDonald's (1963) diagnosis of the palatal notch terminating opposite M2 and large, rounded auditory bullae. These expand ventrally slightly more than the tooth rows; but only very slightly so, in overall appearance both extend to about the same level.
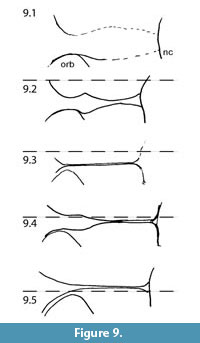
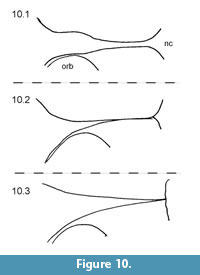
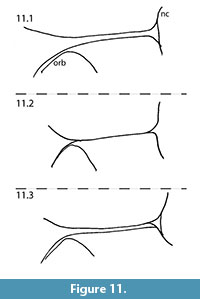
MacDonald (1963) also used the form and course of the fronto-parietal-sagittal crest in the generic diagnosis for Capacikala: "Strongly developed, lyrate, sagittal crest." For comparison with the skull of Capacikala described here, the fronto-parietal-sagittal crest has been sketched from several specimens assigned to Capacikala, Capatanka, and Palaeocastor (Figure 9, Figure 10, Figure 11). Several specimens of Capacikala show a divergence different from a clear linear sagittal crest from the interorbital constriction to the nuchal crest: a lyrate one (Figure 9.4), and one with an irregular, nearly zigzagging course (Figure 9.2). The differences in the fronto-parietal-sagittal crest might be associated with ontogenetic changes with age as indicated above (see Hinze, 1950; Freye, 1959; Calede, 2013). The Capacikala specimen with the lyrate sagittal crest has medium worn teeth, and should therefore be younger than the others with strongly worn teeth. Only for the specimen with irregular shaped crest the wear of the teeth is not clear. In the fewer specimens of Capatanka the variation is slightly less, from a lyrate one (Figure 10.1) to a more or less triangular course with a relative elongate part of a fused sagittal crest. Here the lyrate form, however, is associated with strongly worn teeth and the "triangular" one with medium to strongly worn teeth. The specimens assigned to Palaeocastor show more uniform fronto-parietal-sagittal crests, usually a fusion to form a narrow or broader sagittal crest more or less at the interorbital constriction (Figure 11). All these specimens show strongly or medium to strongly worn teeth. These observations indicate that there is some variation in the form and course of fronto-parietal-sagittal crest, and that there might be more undocumented changes in ontogeny in this feature in fossil beavers – similar to Castor – than realized so far as not all taxa are well represented by material of all individual age classes. In Castor wide and rounded frontoparietal crest in juveniles change to a lyrate form between 1.5-3 years of age and with progressive age form a straight sagittal crest (Freye, 1959).
 Two anterior snout parts of Capacikala have been illustrated: Capacikala sp. (AMNH 64221, Martin, 1987, figure 11D) and Capacikala gradatus (F:AM 64152, Xu, 1996). Both differ from the JODA specimen (Figure 2) in some respects. The first shows a marked bend at the posterior end of the nasals dorsal to the crista facials not present in the JODA specimen. Here the highest point of the skull is further posterior, dorsal to M1/2, and the rise from the tip of the nasals is not as sharp. The curvature of the premaxillary and maxillary bones tapers off towards the tooth row much more gradually in the JODA specimen. Also both illustrations show a differently pronounced shallow groove dorsal to the incisor, not visible in the JODA specimen. As the variability of this feature has neither been studied nor illustrated in detail it is difficult to comment on the importance of this feature.
Two anterior snout parts of Capacikala have been illustrated: Capacikala sp. (AMNH 64221, Martin, 1987, figure 11D) and Capacikala gradatus (F:AM 64152, Xu, 1996). Both differ from the JODA specimen (Figure 2) in some respects. The first shows a marked bend at the posterior end of the nasals dorsal to the crista facials not present in the JODA specimen. Here the highest point of the skull is further posterior, dorsal to M1/2, and the rise from the tip of the nasals is not as sharp. The curvature of the premaxillary and maxillary bones tapers off towards the tooth row much more gradually in the JODA specimen. Also both illustrations show a differently pronounced shallow groove dorsal to the incisor, not visible in the JODA specimen. As the variability of this feature has neither been studied nor illustrated in detail it is difficult to comment on the importance of this feature.
Martin's (1987, p. 74) diagnosis for the subfamily Palaeocastorinae was "beavers with flattened incisors and shortened, rounded tails." His (Martin, 1987, p. 79) diagnosis for the tribe Capacikalini is: "flat-incisored beavers with posteriorly recurved incisors; superficial muscle origins deeply grooved into anterior zygomatic plate; short, deep skulls and double sagittal crests on the interorbital area." The upper incisors of the present skull are slightly convex to nearly flattened, and the lower ones have a flat enamel surface (like in modern Castor the upper incisors are slightly more convex than the lower ones). And in the interorbital area one could argue for a double sagittal crest, even though the fronto-parietal crests fuse to a broad crest slightly more posteriorly. Other specimens of Capacikala do not show double sagittal crests (Figure 9).
Martin's (1987) diagnosis of posteriorly recurved incisors is difficult to address as no clear angle is given. The expression can be related to the angle of which rodent incisor protrude from the jaw, or procumbency of the incisors, long regarded as important for taxonomy and first measured by Thomas (1916, 1919). Incisors which show angles of more than 90°, thus they are "thrown forward", are called proodont, those showing less than 90° ophisthodont, and those showing 90° to the rostrum orthodont. The angle was measured with a goniometer between a line parallel to the cheek teeth row and a tangent "at the midpoint of the incisor" (Landry, 1957, p. 224) and called incisor index (Thomas, 1919) but later termed incisor-rostral angle (Landry, 1957). Lessa and Thaeler (1989) suggested another measurement and calculation of the procumbency index trigonometrically. These authors measure the length of two distances: One from the anterior alveolar rim of P4/p4 (A) and the distal rim of the alveolus of the incisor (B) and one from this latter point to the tip of the incisor (C) in the upper and lower jaw, respectively. The procumbency angle was derived trigonometrically between these lines, but the procumbency ratio was calculated as log(AC/AB) for the upper jaw lower jaw, respectively. Comparative values of procumbency angle for few castorids are given in Table 4. The table indicates that the procumbency of Castor and Palaeocastor simplicidens is similar, that of Capacikala gradatus, Palaeocastor fossor, and Capatanka cankpeopi is similar and slightly larger, and that of P. nebrascensis and P. magnus is the largest. Larger procumbency and enlarged jaw muscles are typical for tooth diggers (Lessa and Thaeler (1989). Samuels and Van Valkenburgh (2009) used geometric morphometrics and inferred chisel-tooth digging for Palaeocastor simplicidens, P. Fossor, and P. magnus, but scratch digging for P. cf. nebrascensis, P. Peninsulatus, and Castor. Capacikala and Capatanka were not included in their study. Just taking procumbency as an indicator one would assume similar behavior using teeth in Castor and Palaeocastor simplicidens and would infer tooth digging for P. nebrascensis, which would not be consistent with the results presented by Samuels and Van Valkenburgh (2009). Therefore, other, more complex functions must play a role in determining incisor procumbency in beavers, and current biomechanical studies will probably help to understand that better. And the procumbency alone may not be totally indicative of type of digging behavior.
The skull described here matches the coarse overall criteria for Palaeocastorinae and Capacikalini given by Martin (1987). Korth (2001) gave a more detailed definition of the Subfamily Palaeocastorinae including synapomorphic features in common with castoroidines and castorines, which can partially be supported with the presented skull (e.g., upper tooth row strongly divergent posteriorly, loss of dP3/dp3, P4/p4 largest cheek tooth, nasal bones extending posteriorly on rostrum to about same level as premaxillaries or cannot be clearly determined like loss of stapedial foramen, posterior palatine foramina within palatine-maxillary suture, palatal surface grooved). The feature in the definition of an incisive foramen less than 30% of total length of the upper diastema is slightly marginal, as the incisive foramen is about 1/3 of the diastema length (Figure 2). The more important autapomorphic features in the definition include a shorter rostrum and broader postorbital constriction and neurocranium than other castorids (which should be addressed, but is beyond the scope of the current paper), flattened enamel surface of incisors (which is shared with other beavers like Castor and Chalicomys), ascending ramus of mandible more anteriorly placed than in other beavers (which is somewhat difficult to assess but actually the ascending ramus crosses m2 at least in Capacikala, Steneofiber, Procastoroides, and Castor).
As indicated in the comparison above, drawing the boundary between Palaeocastor and Capacikala and coming to a consensus on generic assignment of specimens within the palaeocastorine beavers seems to be still difficult (see also Rybcynski, 2007). It is especially difficult, as MacDonald (1963) did not illustrate his new genus Capacikala, the illustration of Capatanka is without indication of sutures of bones, and the definition was fairly coarse and subsequent descriptions offer few new details. So far no general agreement on the generic assignment of all species of palaeocastorine beavers is achieved.
Taking the diagnoses for the discussed genera from Xu (1996) indicates the following differences between Capacikala and Palaeocastor: straight and longer masseter ridge in Capacikala; a lyrate sagittal crest in Capacikala versus a single or double well-developed sagittal crest in Palaeocastor (which is difficult to use as differential feature as there is variability for both genera as shown within this paper and ontogenetic variation known from Castor Hinze, 1950; and Palaeocastor Calede, 2013); maximum width of nasals nearly equal to maximum width of snout (which however is also indicated in Palaeocastor peninsulatus Calede, 2013); internal nares ending level with anterior edge of M3 in Palaeocastor and with M2 in Capacikala (the trait mentioned as end of palate by MacDonald (1963) and supported herein).
For Palaeocastor a skull length to breadth ratio of 1.2 to 1.3 is given but none for Capacikala (Xu 1996). Therefore, it is difficult to compare but the JODA specimen has a skull length-width ratio of 1.36 which is just out the given range of Palaeocastor but not substantially different. The skull length range for Capacikala is given with 43-55 mm and the described skull is somewhat longer with nearly 60 mm.
Similarities between both genera as given by Xu (1996) are: a postzygomatic skull length of about 0.20 to 0.25 of skull length; incisor with flat face; and alignment of the three processes of the mandible in posterior view. As for the incisors the JODA specimen has slightly convex upper incisors not flat ones as given in Xu's diagnosis. But the same holds for Palaeocastor for which also flat faced incisors are given as characteristic but this is not seen in all (compare Calede, 2013). In the JODA specimen the coronoid, condylar, and angular process are not aligned but zigzagging: the condyle being much more lingual than the other two processes which are nearly in one line.
Two mentioned diagnostic features (Xu, 1996) are so common in other castorids that I think they are not very helpful in a diagnosis to differentiate between genera: "P4 complicated in early wear stage, but simpler in late war stage" for Palaeocastor and "Mesoflexus (-felxid) and hypoflexus (-flexid) strongest in dental structures" for Capacikala.
Flynn and Jacobs (2008) revised the diagnoses of the two genera which includes incisors somewhat rounded for Palaeocastor and orthodont incisors for Capacikala which are supported with the JODA specimen.
From this analysis it seems obvious that Capacikala is very similar to Palaeocastor as there are similarities, some of which (paritularly the low position of the infraorbital foramen and the pattern of the mandibular processes in distal view) have been assumed primitive features for all castorids (Korth, 2001). Nevertheless, the shape of the nasals (e.g., compared between the JODA specimen and P. peninsulatus (Calede, 2013, figure 1A), overall skull shape and size; relation of skull length to skull width; proportion of postzygomatic skull length as indicated by Xu (1996); course and shape of fronto-parietal crests and its chaning with age; form of the orbit and interorbital constriction. Differences include the different positions of the distal end of the palate; presence of an interpremaxillary foramen in Capacikla (however also skull fragments in the SDSM assigned to Palaeocastor bear interpremaxillary formanina); subtle differences in the shape of the bulla tympanica; more obvious depression dorsal to the incisor on the maxillary bone; slightly broader maximal dorso-ventral extension of zygomatic arch and the teeth (Stefen, 2010).
Phylogenetic Analysis
As the support for the bootstrap of the presented suggested phylogeny is low, it has to be treated with caution. The three castorid groups Palaeocastorinae, Castorinae, and Castoroidine already indicated by Rybczynski (2007), and Korth (1994, 2001). Contrary to the phylogeny of Rybczynski (2007) the Castorinae appear as sister groups to the Palaeocastorinae and both appear as sister group to the Castoroidinae plus Anchitheriomys. Only Simpson (1945) includes Amblycastor in the Castoroidinae and Xu (1995) lists Anchitheriomys and Trogontherium with Asiatic beavers in the Asiocastorinae. Both indicate some relation or similarity between the discussed taxa. Within the palaeocastorines Capacikala gradatus appears as sister taxon to the other included palaeocastorine beavers. Martin (1987, figure 11) suggested an early separation of Capacikala from other palaeocastorine beavers.
It is not within the scope of the present paper to reanalyze castorid phylogeny nor the relative wealth of different characters. As there are also difficulties to interpret procumbency alone as indicative of specific digging behavior or the size of teeth of different palaeocastorine taxa taxonomically (Stefen, 2010), probable characters need to be examined in relation to checking functional morphological and biomechanic aspects.
ACKNOWLEDGEMENTS
I want to thank the National Park Service for supporting a visit to the John Day Fossil Beds National Monument (JODA) to study the specimen and the DFG for partial funding of the project through travel grants to attend meetings and associated collection visits (2000, 2004). The drawings of the specimen were made by J.P. Lufkin and kindly provided by H. Wagner who was helpful with discussions also. Ted Fremd at JODA was helpful during the visit and supplied data on the stratigraphy and age of the specimen as well as the photographs of the skull and mandible and the basis for Figure 1. R. Dunn, paleobotanist at John Day Fossil Beds, assisted with some stratigraphic figures. Particularly P. Holroyd, UCMP, J. Martin, SDSM, S. Bell and J. Meng, AMNH, and L.Barnes at LACM, and the staff of the MVZ in Berkeley were very helpful with finding material in the collections under their care and giving me access to the material. I also want to thank the Editor and the reviewers for their helpful comments.
REFERENCES
Albright, L.B., Fremd, T., Swisher, C.C., and MacFadden, B.J. 2001. Toward a revision of the chronostratigraphy of the John Day Formation, Oregon. NAPC 2001, Paleobios 21, Supplement to No. 2, p. 22.
Albright, L.B.III, Woodburne, M.O., Fremd, T.J., Swisher, C.C.III, MacFadden, B.J., and Scott, R.G. 2008. Revised Chronostratigraphy and biostratigraphy of the John Day Formation (Turtle Cove and Kimberly Members), Oregon, with implications for updated calibration of the Arikareean North American Land Mammal Age. Journal of Geology, 116:211-237.
Bowdich, T.E. 1821. An analysis of the Natural Classifications of Mammalia for the Use of Students and Travellers. J. Smith, Paris.
Calede, J.J.-M. 2013. Skeletal morphology of Palaeocastor peninsulatus (Rodentia, Castoridae) from the Fort Logan Formation of Montana (early Arikareean): Ontogenetic and paleoecological interpretaions. Journal of Mammalian Evolution, [DOI 10.1007/s10914-013-9231-8.]
Cope, E.D. 1879a. Observations on the faunae of the Miocene Tertiary of Oregon. Bulletin of the U. S. Geological and Geographical Survey of the Territories, 1880:55-69.
Cope, E.D. 1879b. On some characters of the Miocene fauna of Oregon. Proceedings of the American Philosophical Society, 18:63-78.
Cope, E.D. 1884. The Vertebrata of the Tertiary formations of the West. Book 1. Report of the U.S. Geological Survey of the Territories, (F. V. Hayden), 3:1-1009.
Flynn, L.J. and Jacobs, L.L. 2008. Castoridae, p. 89-109. In Janis, C.M., Gunnell, G.F., and Huehn, M.D. (eds.), Evolution of Tertiary Mammals of North America, Vol. 2: Small Mammals, Xenarthrans, and Marine Mammals. Cambridge University Press, Cambridge.
Fremd, T., Bestland, E.A., and Retallack, G.J. 1994. John Bay Basin Paleontology Field Trip Guide and Road Log. Society of Vertebrate Paleontology Annual Meeting, Seattle, Washington.
Fremd, T., Bestland, E.A., and Retallack, G.J. 2000. Eocene and Oligocene paleosols of Central Oregon. Geological Society of America, Special Paper, 344:1-2.
Freye, H-A. 1959. Descriptive Anatomie des Craniums vom Elbe Biber (Castor fiber albicus Matschie 1907). Wissenschaftliche Zeitschrift University Halle, mathematisch-naturwissenschaftliche Reihe, 8:913-962.
Hemprich, W. 1820. Grundriss der Naturgeschichte für höhere Lehranstalten. August Rücker, Berlin.
Hinze, G. 1950. Der Biber. Akademie Verlag, Berlin.
Hunt, R.M. and Stepleton, E. 2004. Geology and paleontology of the upper John Day beds, John Day River Valley, Oregon: lithostratigraphic and biochronologic revision in the Haystack Valley and Kimberly areas (Kimberly and Mt. Misery quadrangles). Bulletin of the American Museum of Natural History, 282:1-90.
Korth, W.W. 1994. The Tertiary Record of Rodents in North America. Plenum Press, New York & London.
Korth, W.W. 2001. Comments on the systematics and classification of the beaver. Journal of Mammalian Evolution, 8:279-296.
Korth, W.W. and Emry, R.J. 1997. The skull of Anchitheriomys and a new subfamily of beavers (Castoridae, Rodentia). Journal of Paleontology, 71: 343-347.
Landry, S.O. 1957. Factors affecting the procumbency of rodent upper incisors. Journal of Mammalogy, 38:223-234.
Leidy, J. 1869. The extinct mammalian fauna of Dakota and Nebraska, including an account of some allied forms from other localities, together with a synopsis of the mammalian remains of North America. Journal of the Academy of Natural Sciences of Philadelphia, 7:341-405.
Leidy, J. 1870. On Vertebrate fossils from the John Day region, Oregon. Proceedings of the Academy of Natural Sciences of Philadelphia, 1870:111-113.
Leidy, J. 1871. Remarks on fossils from Oregon. Proceedings of the Academy of Natural Sciences of Philadelphia, 1871:247-248.
Lessa, E.P. and Thaeler, C.S. 1989. A reassessment of morphological specializations for digging in pocket gophers. Journal of Mammalogy, 70:689-700.
MacDonald, J.R. 1963. The Miocene faunas from the Wounded Knee Area of Western South Dakota. Bulletin of the American Museum of Natural History, 125:139-238.
Martin, L.D. 1987. Beavers from the Harrison Formation (Early Miocene) with a revision of Euhapsis. Dakoterra, 3:73-91.
Martin, L.D. and D.D. Bennett. 1977. The burrows of the Miocene beaver Palaeocastor, Western Nebraska, USA. Palaeogeography, Palaeoclimatology, Palaeoecology, 22:173-193.
McKenna, M. and Bell, S.K. 1998. Classification of Mammals above the Species Level. Columbia University Press, New York.
Merriam, J.C. 1901. A contribution to the geology of the John Day Basin [Oreg.]. Bulletin of University of California Department of Geology, 2:269-314.
Merriam, J.C. and Sinclair, W.J. 1907. Tertiary faunas of the John Day Region. University of California Publications, Bulletin of the Department of Geology, 5:171-205.
Moore, J. 1890a. Concerning a skeleton of the great fossil beaver Castoroides ohioensis. Journal of the Cincinnati Society of Natrual History, 1890:138-169.
Moore, J. 1890b. Description of anew species of gigantic beaver-like rodent. Cincinnati Society of Natural History, 13:26-30.
Mörs, T. and Hulbert, R.C. Jr. 2010. Anchitheriomys Roger, 1898 or Amblycastor Matthew, 1918 (Rodentia, Castoridae) Taxonomic implications of a mandible from the Miocene of Florida. Journal of Vertebrate Palaeontology, 30:1899-1902.
Peterson, O.A. 1905. Description of new rodents and discussion of the origin of Daemonelix. Memoirs of the Carnegie Museum, Pittsburgh, 2:139-191.
Peterson, O.A. 1906. The Miocene beds of western Nebraska and eastern Wyoming and their vertebrate faunae. Annales Carnegie Museum, 4:21-72.
Rybczynscki, N. 2007. Castorid Phylogenetics: implications for the Evolution of Swimming and tree-exploitation in Beavers. Journal of Mammalian Evolution, 14:1-35.
Rybczynski, N., Ross, E.M., Samuels, J.X., and Korth, W. 2010. Reevaluation of Sinocastor (Rodentia: Castoridae) with implications on the origin of modern beavers. PLoS ONE, 5(11):e13990.
Samuels, J.X. and Van Valkenburgh, B. 2009. Craniodental adaptation for digging in extinct burrowing beavers. Journal of Vertebrate Paleontology, 29:254-268.
Samuels, J.X. and Zancanella, J. 2011. An early Hemphillian occurrence of Castor (Castoridae) from the Rattlesnake Formation of Oregon. Journal of Paleontology, 85:930-935.
Schreuder, A. 1929. Conodontes (Trogontherium) and Castor from the Teglian Clay compared with the Castoridae from other localities. Archives du Musée TeylerSeries III, 6:99-321.
Shotwell, J.A. 1968. Miocene mammals of Southeast Oregon. Bulletin of the Museum of Natural History University of Oregon, 14:1-67.
Simpson, G.G. 1945. The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History, 85:1-350.
Stefen, C. 2001. The Barstovian (Miocene) beavers from Stewart Valley, Nevada, and a preliminary discussion of the genus Monosaulax mainly on the basis of tooth morphology. PaleoBios, 21:1-15.
Stefen, C. 2005. Description of the cranial morphology of the Early Miocene beaver Steneofiber castorinus. Neues Jahrbuch für Mineralogie und Paläontologie, 2005:577-596.
Stefen, C. 2010. Morphometric considerations of the teeth of the palaeocastorine beavers Capacikala, Palaeocastor and "Capatanka." Palaeontologica Electronica, 13:2A, 1-35.
palaeo-electronica.org/2010_1/index.html
Stefen, C. and Mörs, T. 2008. The beaver Anchitheriomys from the Miocene of Central Europe. Journal of Paleontology, 82:1009-1020.
Stirton, R.A. 1934. A new species of Amblycastor from the Platybelodon beds, Tung Gur Formation, of Mongolia. American Museum Novitates, 694:1-4.
Stirton, R.A. 1935. A review of tertiary beavers. University of California Publications in Geological Sciences, 23:391-458.
Stirton, R.A. 1965. Cranial morphology of Castoroides. Dr. D.N. Wadia Commemorative Volume Mining and Metallurgical Institute of India, 273-285.
Swafford, D.L. 1998. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4. Sinauer Associates, Sunderland, Massachusetts.
Thomas, O. 1919. The method of taking the incisive index in rodents. The Annals and Magazine of Natural History, 9:289-290.
Wagner, H.M. 1983. The cranial morphology of the fossil beaver Dipoidessmithii (Rodentia: Mammalia). Contributions in Science, Natural History Museum Los Angeles, 346:1-6.
Wahlert, J.H. 1977. Cranial foramina and relationship of Eutypomys (Rodentia, Eutypomyidae). American Museum Novitates, 2626:1-8.
Xu, X.F. 1996. Castoridae, p. 417-432. In Prothero, D.R. and Emry, R.J. (eds.), The Terrestrial Eocene-Oligocene Transition in North America. University Press, Cambridge.
Young, F.W. 1937. Studies of osteology and myology of the beaver (Castor canadensis). Michigan State College Agricultural Experiment Station Memoir, 2:1-84.
