

 What do we know about Neogene bony fishes from Chile? Diversity and biogeographic implications
What do we know about Neogene bony fishes from Chile? Diversity and biogeographic implications
Article number: 28.2.a30
https://doi.org/10.26879/1385
Copyright Palaeontological Association, July 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Appendices
Submission: 4 March 2024. Acceptance: 18 July 2025.
ABSTRACT
Deep climatic and oceanographic transformations in the past have shaped Chile’s modern marine biodiversity. However, unlike some other vertebrates and invertebrates, little is known about the past diversity of bony fishes (Osteichthytes) along this coast during the Neogene. This study aims to summarize paleontological evidence of the diversity of bony fishes in Chile, estimate the completeness of the inventories, and assess biogeographic dynamics. Our results show an inventory of 21 orders, 51 families, 67 genera, and 46 species for the Neogene. Compared to the current diversity (624 genera and 1,198 species), only 3% of the living genera and 20% of living families have fossil counterparts and a single living species has a fossil record. At a generic level, 40% are currently absent in the region, whereas 60% survived. For the genera currently absent in the region, 19% are globally extinct, and 81% are regionally extinct. The changes in the biogeographic dynamics of bony fishes from the past to the present could be related to the intense climatic and oceanographic events that took place throughout the Neogene period. The rarefaction and extrapolation analyses confirm that although the studies on the group have increased in the last years, more taxa could still be found if the sampling effort is increased. Future studies could help us to better understand the effects of climatic and oceanographic changes on the group during the Neogene of Chile.
Pablo Oyanadel-Urbina. Laboratorio de Paleobiología, Centro de Estudios Avanzados en Zonas Áridas, Av. Ossandón 877, Coquimbo, Chile and Área de Investigación y Desarrollo, Therium servicios profesionales limitada, Paleontología y Patrimonio. pablo.oyanadel@ceaza.cl
Jaime A. Villafaña. Departamento de Ecología, Facultad de Ciencias, Universidad Católica de la Santísima Concepción, Concepción 4090541, Chile (corresponding author). jaime.villafana@ceaza.cl
Karina E. Buldrini. Área de Paleontología, Museo Nacional de Historia Natural, Santiago, Chile. karina.buldrini@mnhn.gob.cl
Héctor A. Ramos-Rojas. Laboratorio de Paleobiología, Centro de Estudios Avanzados en Zonas Áridas, Av. Ossandón 877, Coquimbo, Chile and Departamento de Biología Marina, Facultad de Ciencias del Mar, Universidad Católica del Norte, Coquimbo, Chile. hector.ramos.rojas@ceaza.cl
Yusse Hernández-Mora. Laboratorio de Paleobiología, Centro de Estudios Avanzados en Zonas Áridas, Av. Ossandón 877, Coquimbo, Chile and Departamento de Biología Marina, Facultad de Ciencias del Mar, Universidad Católica del Norte, Coquimbo, Chile. yusse.hernandez@ceaza.cl
Martín Chávez-Hoffmeister. Laboratorio de Paleontología, Instituto de Ciencias de la Tierra, Universidad Austral de Chile, Valdivia, Chile and Corporación de Investigación para el Desarrollo de la Paleontología e Historia Natural, CIAHN Atacama, Caldera, Chile. martinchavez@ciahn.cl
Sven N. Nielsen. Laboratorio de Paleontología, Instituto de Ciencias de la Tierra, Universidad Austral de Chile, Valdivia, Chile. sven.nielsen@uach.cl
Jorge A. Campos-Medina. Laboratorio de Paleobiología, Centro de Estudios Avanzados en Zonas Áridas, Av. Ossandón 877, Coquimbo, Chile and Área de Investigación y Desarrollo, Therium Servicios Profesionales Limitada, Paleontología y Patrimonio. jorge.camposmedina@gmail.com
Sandra Rebolledo. Departamento de Antropología, Facultad de Ciencias Sociales, Universidad Alberto Hurtado and Centro de Estudios Históricos y Arqueológicos Aiken, Santiago, Chile.
sanrebolledoc@gmail.com
Marcelo M. Rivadeneira. Laboratorio de Paleobiología, Centro de Estudios Avanzados en Zonas Áridas, Av. Ossandón 877, Coquimbo, Chile and Departamento de Biología Marina, Facultad de Ciencias del Mar, Universidad Católica del Norte, Coquimbo, Chile. marcelo.rivadeneira@ceaza.cl
Keywords: Miocene; Pliocene; fossils; extinction; bony fish
Final citation: Oyanadel-Urbina, Pablo, Villafaña, Jaime A., Buldrini, Karina E., Ramos-Rojas, Héctor A., Hernández-Mora, Yusse, Chávez-Hoffmeister, Martín, Nielsen, Sven N., Campos-Medina, Jorge A., Rebolledo, Sandra, and Rivadeneira, Marcelo M. 2025. What do we know about Neogene bony fishes from Chile? Diversity and biogeographic implications. Palaeontologia Electronica, 28(2):a30.
https://doi.org/10.26879/1385
palaeo-electronica.org/content/2025/5497-neogene-bony-fishes-from-chile
Copyright: July 2025 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Fishes are the oldest living vertebrates on the face of the Earth, with over 500 million years of existence on our planet and more than 32,000 living species (Nelson et al., 2016), accounting for 50% of all known vertebrates (Meléndez, 2008). Their diverse physiological and morphological adaptation strategies allow them to survive in a wide range of environments (Nelson et al., 2016; Froese and Pauly, 2024). Fishes also constitute an important alimentary resource for the planet (Boyd et al., 2022), while fisheries serve as a significant source of information for scientific purposes (Fischer et al., 1995; Seijo and Defeo, 1997).
In Chile, the study of marine living fishes has a long history dating back to the 16th century. Molina (1782) was among the first to compile a list of fishes in the Chilean territory, identifying just 11 taxa. Over time, numerous researchers, including Guichenot (1848), Gay (1854), Philippi (1887a), Reed (1897), and Quijada (1913), among others, contributed to expanding this list. By the mid-twentieth century, Chile’s catalog of living fish species had grown significantly, reaching 348 species, according to Fowler (1945). This count encompassed various types of fish, including lampreys, hagfishes, and bony and cartilaginous fishes. Towards the end of the twentieth century, Pequeño (1989) reported a remarkable increase, with 1,016 species being mentioned as inhabiting Chilean waters. Hüne (2019) added several species in the most recent systematic update, thus identifying 1,307 marine species.
The current biogeographical distribution of bony fishes in Chile results from intricate geological, oceanographic, and climatic processes that unfolded in the past. Certain processes that occurred during the Neogene epoch had an impact on the region’s marine life. Notable events during the Neogene include phases of uplift of the Andes (Victor et al., 2004; Blisniuk et al., 2005), the onset of hyperaridity in the Atacama Desert (Hartley and Chong, 2002), the activation of coastal warm upwelling in the East Pacific during the middle Pliocene (Dekens et al., 2007), and marine cooling through the Humboldt Current System (e.g., Nielsen and Glodny, 2009; Kiel et al., 2023). These events exerted a significant influence on the diversity of marine fauna, encompassing mollusks, marine mammals, birds, and cartilaginous fishes (Rivadeneira and Marquet, 2007; Kiel and Nielsen, 2010; Valenzuela-Toro et al., 2013; Villafaña and Rivadeneira, 2014, 2018; Rivadeneira and Nielsen, 2017; Benites-Palomino et al., 2022; Vermeij et al., 2024). Despite these advancements in understanding other marine taxa, the specific impact of climatic and oceanographic events on the diversity and biogeographic distribution of bony fishes in Chile remains unexplored and requires further investigation.
Fossil bony fishes in Chile have received less attention than their living counterparts, although their study has a long history. The first descriptions of Chilean bony fish fossils are those of Philippi (1887b). Subsequently, numerous studies have expanded our understanding of the fossil record of bony fishes in Chile, spanning from the Paleozoic to the Quaternary (Arratia and Schultze, 1999; Arratia, 2015). However, the primary focus of research within this group has centered on the Mesozoic record of teleost fishes (Arratia et al., 1975; Gasparini, 1979; Bell, 1985; Arratia, 2015; Otero and Suárez, 2022). For the Cenozoic, there are very few reports from the Paleogene (Otero, 2019) but several from the Neogene, particularly from marine environments (Long, 1993; Walsh, 2001; Nolf, 2002; Suárez et al., 2003; Báez, 2006; Gutstein et al., 2008; Carrillo-Briceño, 2011; Pérez, 2017; Oyanadel-Urbina et al., 2018, 2021; Schwarzhans and Nielsen, 2021). In the Quaternary period, research has predominantly focused on remains from archaeological shell middens (e.g., Olguín et al., 2014; Béarez et al., 2016; Morello et al., 2015; Rebolledo et al., 2016, 2021a, b; Labarca et al., 2020; Torres et al., 2022).
While substantial progress has been made in understanding the diversity of both fossil and living fishes in Chile, no studies have hitherto attempted a detailed review and analysis of the group’s historical diversity and biogeographic distribution. In this study, we aim to summarize and analyze the diversity of bony fishes in Chile from the Neogene to the present. We will also explore the potential relationships between changes in diversity over time and the profound climatic and oceanographic events in the region.
MATERIAL AND METHODS
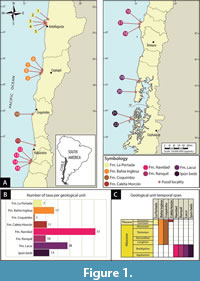 Data. Our dataset of fossil and extant records of bony fishes from Chile was compiled through an exhaustive literature review (Figure 1; Appendix 1 and Appendix 2). This review encompassed a wide range of sources, including journal articles, theses, technical reports, and conference abstracts, ensuring a comprehensive examination of the available information. The information was also complemented with online repositories and museum collection records.
Data. Our dataset of fossil and extant records of bony fishes from Chile was compiled through an exhaustive literature review (Figure 1; Appendix 1 and Appendix 2). This review encompassed a wide range of sources, including journal articles, theses, technical reports, and conference abstracts, ensuring a comprehensive examination of the available information. The information was also complemented with online repositories and museum collection records.
Regarding fossils, our dataset comprises 218 occurrences from different taxonomic levels, sourced from Neogene (23 to 2.6 Ma) localities across Chile. The material of bony fishes examined in this study includes dentaries, skulls, pharyngeal tooth plates, opercular bones, rostra, and otoliths. To ensure data quality and specificity, we excluded fossil records that lacked stratigraphic information and those identified only at higher taxonomic levels (e.g., Teleostei indet.).
Additionally, all taxonomic names were updated in accordance with the most recent studies. The same approach was applied to the assignment of ages for localities and geological formations, specifically for those whose classifications had been revised after the publication of the referenced studies. This refinement ensures the reliability and precision of our database.
For living forms, the data are largely based on the systematic list presented by Hüne (2019). Additionally, we utilized online data as supplementary material from Eschmeyer’s Catalogue of Fishes (Fricke et al., 2024) and FishBase (Froese and Pauly, 2024). Relevant data from the ichthyological collection at the Museo Nacional de Historia Natural, Chile (MNHNCL-ICT) were also incorporated. We exclude extant records reported without specific local information (i.e., those assigned only to the country level). Our compiled list of extant fishes encompasses species inhabiting marine environments and those associated with the sea at some stage of their development, including species that have been introduced into Chilean marine waters.
Analyses. To reconstruct the biogeographic distribution of Neogene fishes at the genus level spanning from the past to the present, we calculated the Lyellian percentages at genus level (Stanley et al., 1980; Pease, 1987), (i.e., the proportion of Neogene genera still living in each region today). Only marine bony fishes were thus considered in the present analyses, excluding all stenohaline freshwater species. The current distribution of genera was obtained from FishBase (Froese and Pauly, 2024) and Hüne (2019). We applied two distinct spatial scales: regional and local. The regional scale examines genera that disappeared from Chile (due to regional extirpation). On the other hand, the local scale considers genera that either contracted or expanded their southern range within Chile from the Neogene to the present. We consider a range shift to have occurred when there is a difference of at least 4º of latitude (the minimum shift observed between fossil localities included in this study) from the southernmost distribution endpoints in the Neogene period and the current distribution. This criterion helped us identify significant changes in the distribution of genera within Chile over time.
We used coverage-based estimations of the genus richness to evaluate and account for possible sampling biases (Chao and Jost, 2012), using the number of sites as a proxy of sampling effort. In addition, we used the Chao 2 extrapolation index as a measure of asymptotic diversity. Analyses were done using the library iNEXT in R (Hsieh et al., 2016).
RESULTS
The Neogene fish fauna of Chile, as documented to date, is composed of 88 different taxa. These fishes are categorized into 21 orders, 51 families, 67 genera, and 46 species (Table 1), with a total of 227 available records (Appendix 1). It is worth noting that 52% of the documented taxa have been identified down to the species level, while 35% have been classified at the genus level only. Notably, when considering the percentage of georeferenced occurrences, two species stand out as the most frequently encountered in the Chilean Neogene: Diogenichthys aguilerai (Myctophidae), accounting for 5% of records, and Citharichthys parvisulcus (Paralichthyidae), representing 4% of records. Furthermore, in terms of individual genera, Diaphus (Myctophidae) boasts the highest number of species at 9%, followed by Lampanyctus (Myctophidae) with 7%. When examining families, Myctophidae and Ophidiidae are the most frequently represented, making up 12% and 7% of the total genera, respectively. Finally, among the various orders, Perciformes and Gadiformes are the most prevalent, accounting for 16% and 13% of the total genera, respectively.
Fossil remains of fishes have been reported from 22 fossil localities, comprising eight geological units: La Portada, Bahía Inglesa, Coquimbo, Caleta Horcón, Navidad, Ranquil, and Lacui formations, and Ipún beds. These deposits are located between Antofagasta (23ºS) and Coyhaique (44ºS) (Figure 1). The Early Miocene stands out as the most well-documented time span in Chile, featuring 62 taxa, mostly identified by the study of otoliths. Following far behind is the Late Miocene, which accounts for 17 taxa, and the Pliocene, with 11 taxa (Table 1), for which only macroscopic skeletal remains are known. It is worth highlighting that the Middle Miocene has only one recorded taxon (Thunnus).
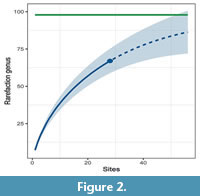 As the number of sites increases, the rarefaction curve rises, and there are no indications of reaching saturation with the 67 known genera (Figure 2). According to the Chao 2 index, the anticipated richness of genera is estimated to be 98, with a 95% confidence interval ranging from 78 to 142 genera. Consequently, the actual number of observed genera falls within a range of 47% to 88% of the expected richness (Figure 2).
As the number of sites increases, the rarefaction curve rises, and there are no indications of reaching saturation with the 67 known genera (Figure 2). According to the Chao 2 index, the anticipated richness of genera is estimated to be 98, with a 95% confidence interval ranging from 78 to 142 genera. Consequently, the actual number of observed genera falls within a range of 47% to 88% of the expected richness (Figure 2).
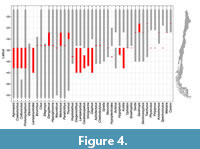
The biogeographic distribution, from the past to the present, exhibits variations across different taxonomic levels. Only 3% of the fish genera that comprise the extant Chilean fauna have been identified in the local fossil record, while 20% of the families and 39% of the orders are also represented in the local record. Among the fossil taxa from Neogene localities that were identified at the species level, only one species still lives along the Chilean coast: Sardinops sagax (Table 1). The remaining species have globally gone extinct. In contrast, 60% of the genera have persisted in the region (Figure 3A and Table 2).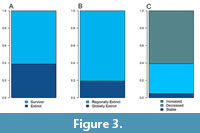 Among the genera that have disappeared from the region, 19% did so as the likely result of global extinctions (e.g., Karrerichthys, Labrodon, Sirembola), while 81% have been extirpated from the region but have survived elsewhere in the world (e.g., Chiloconger, Margrethia, Steindachneria) (Figure 3B and Table 2). Turning our attention to the surviving genera that are still present in Chile (Figure 3C and Table 2), 5% have retained their latitudinal distribution range, including Makaira (Istiophoridae) and Ophidion (Ophidiidae). Meanwhile, 36% have experienced a contraction in their range (Figure 4), including Diogenichthys (Myctophidae), Hypoplectrodes (Serranidae), and Vinciguerria (Phosichthyidae); whereas the remaining 59% of surviving genera have expanded their southern latitudinal range (Figure 4), with notable examples including Anisotremus (Haemulidae), Hippoglossina (Paralichthyidae), and Thunnus (Scombridae).
Among the genera that have disappeared from the region, 19% did so as the likely result of global extinctions (e.g., Karrerichthys, Labrodon, Sirembola), while 81% have been extirpated from the region but have survived elsewhere in the world (e.g., Chiloconger, Margrethia, Steindachneria) (Figure 3B and Table 2). Turning our attention to the surviving genera that are still present in Chile (Figure 3C and Table 2), 5% have retained their latitudinal distribution range, including Makaira (Istiophoridae) and Ophidion (Ophidiidae). Meanwhile, 36% have experienced a contraction in their range (Figure 4), including Diogenichthys (Myctophidae), Hypoplectrodes (Serranidae), and Vinciguerria (Phosichthyidae); whereas the remaining 59% of surviving genera have expanded their southern latitudinal range (Figure 4), with notable examples including Anisotremus (Haemulidae), Hippoglossina (Paralichthyidae), and Thunnus (Scombridae).
DISCUSSION
Taxonomic Composition
 The findings of this study reveal that the taxonomic composition of the fish assemblages in Chile has remained relatively stable since the Early Miocene. Percomorphs were the most diverse group during the Neogene of Chile, accounting for 16% (11 out of 67) of the known genera. This pattern continues today, with perciforms comprising an even larger fraction of the genera inhabiting Chile (40%, 248 out of 624). This is consistent with the fact that the order Perciformes is currently the most diverse group of vertebrates worldwide (Fricke et al., 2024). When examining fish families in Chilean waters, lanternfishes (Myctophidae) exhibit the highest number of genera both in the past (12%, 8 out of 67) and in modern times (4%, 28 out of 624). Diaphus emerges as the most diverse and abundant genus, both in the Neogene and today (Appendix 1 and Appendix 2). Lanternfishes belonging to the genus Diaphus are represented today by 78 valid species that are globally distributed (Froese and Pauly, 2024).
The findings of this study reveal that the taxonomic composition of the fish assemblages in Chile has remained relatively stable since the Early Miocene. Percomorphs were the most diverse group during the Neogene of Chile, accounting for 16% (11 out of 67) of the known genera. This pattern continues today, with perciforms comprising an even larger fraction of the genera inhabiting Chile (40%, 248 out of 624). This is consistent with the fact that the order Perciformes is currently the most diverse group of vertebrates worldwide (Fricke et al., 2024). When examining fish families in Chilean waters, lanternfishes (Myctophidae) exhibit the highest number of genera both in the past (12%, 8 out of 67) and in modern times (4%, 28 out of 624). Diaphus emerges as the most diverse and abundant genus, both in the Neogene and today (Appendix 1 and Appendix 2). Lanternfishes belonging to the genus Diaphus are represented today by 78 valid species that are globally distributed (Froese and Pauly, 2024).
From an ecological perspective, the fossil record of marine fishes from Chile reflects diverse habitats, including coastal, rocky reef, and oceanic environments, and features species that migrate across vast oceanic distances, such as oceanodromous fishes (Nelson et al., 2016). A high number of taxa represented in the fossil record corresponds to forms that are currently linked to deep habitats, like the lanternfishes and sand flounders (Pleuronectiformes: Paralichthyidae) (Froese and Pauly, 2024). However, in the Navidad, Ranquil, Lacui, and other nearby geological formations, soft-bottom fishes and aquatic animals with anadromous/diadromous behaviors have been discovered. In the case of the other geological units, such as the Bahía Inglesa, Coquimbo and La Portada formations, shoal, and pelagic fishes are abundant. In addition, amphidromous fishes associated with rocky reefs can also be found in these units, although in smaller quantities. For instance, in the Horcón Formation, pelagic and oceanodromous fishes are observed, but fishes associated with coastal rocky reefs are more abundant.
Compared to other Neogene localities, the faunal composition of bony fishes from Chile is different from those reported from other regions. In Perú, only seven genera of bony fishes have been reported (De Muizon and DeVries, 1985; Collareta et al., 2015; Landini et al., 2017; Bianucci et al., 2018; Di Celma et al., 2018), which do not belong to the most diverse taxonomic groups in Chile at order and family levels (i.e., perciforms and myctophids). As regards to the fossil record of other regions of the southern Pacific, while Percomorpha is the most diverse order in Chile, the order Gadiformes is the most diverse group of fishes in the Neogene of New Zealand (Schwarzhans, 1980; 2019). Although there exists this difference at the order level, Myctophidae is also the most diverse family in New Zealand as in Chile, this result being dominated by the diverse and abundant early Miocene otolith record of Schwarzhans and Nielsen (2021). This is consistent with the high similarity of faunas between Chile and New Zealand today (Froese and Pauly, 2024). The comparison between the Chilean fauna and other regions in South America is difficult due to the lack of identifications at lower taxonomic levels. For instance, most of the taxa described from nearby regions, such as Peru and Ecuador, are identified at higher taxonomic levels, such as order and family (De Muizon and DeVries, 1985; Carnevale et al., 2011; Collareta et al., 2015) or focus on a particular family (e.g., myctophids; Schwarzhans and Aguilera, 2013). Therefore, interpretations of the differences in terms of faunal composition between the different regions would be pertinent to analyze once more specific data have been described. An increase in the sampling effort could also contribute to the understanding of similarities and differences on the taxonomic composition of bony fishes during the Neogene.
Biogeographic Changes from the Past to the Present
Only seven percent of the bony fish genera from Chile are now globally extinct, a proportion even lower than that observed for chondrichthyans from the eastern Pacific of South America (10%, Villafaña and Rivadeneira, 2018) and tropical America (13%; Carrillo-Briceño et al., 2018). In the case of the surviving genera, the proportion of regional extinction (33%) was similar to that reported for chondrichthyans from the same region (34%; Villafaña and Rivadeneira, 2018). At a local scale, the proportion of genera increasing their southern latitudinal range was higher (61%) than that reported for chondrichthyans (54%; Villafaña and Rivadeneira, 2018). In contrast, the proportion of genera with decreased range was lower (34%), compared to the 46% previously reported for cartilaginous fishes in the region (Villafaña and Rivadeneira, 2018). However, two genera of bony fishes retained their southern latitudinal range from the Neogene to the present in Chile. Overall, the biogeographic distribution did not differ significantly between bony fishes and chondrichthyans from the Neogene of the eastern Pacific.
The biogeographic dynamics of Neogene bony fishes from Chile could be related to the intense climatic and oceanographic changes that occurred in the eastern Pacific off South America (Ibaraki, 1997; Tsuchi, 2002; Dekens et al., 2007). This is consistent with what has been observed in chondrichthyans (Villafaña and Rivadeneira, 2018). For instance, the bigmouth sanddab (Citharichthys gilberti) and the small sanddab (Citharichthys platophrys) are typically found at shallow depths of tropical areas from California to northern Peru (Froese and Pauly, 2024). The presence of typical tropical species during the Neogene was also mentioned by previous studies based on the fossil record of invertebrates and vertebrates, including fishes such as chondrichthyans (Nielsen and Glodny, 2009; Villafaña and Rivadeneira, 2018; Partarrieu et al., 2018; Guicharrousse-Vargas et al., 2021). According to Villafaña and Rivadeneira (2018), the absence of tropical species at the end of the Neogene could be related to the decrease in water temperature during that time. In addition, the loss of habitat caused by the sea level changes (Lambeck and Chappell, 2001; Van der Meer et al., 2017) is also suggested as one of the factors related to biogeographic changes in fishes from South America and Europe (Cione et al., 2007; Pimiento et al., 2017; Villafaña and Rivadeneira, 2019a; Villafaña et al., 2023).
Sampling and Taphonomic Biases
Our results show that the inventory of fossil taxa from the Neogene of Chile is far from being complete. This is reflected by the moderate completeness estimated at the genus level (68%) through the Chao 2 analysis. Also, the percentage of living families with a fossil counterpart (20%) is much lower than the ca. 60-65% estimated globally for Osteichthyes (Foote and Sepkoski 1999). At the same time, taxonomic identification at lower taxonomic levels (i.e., genera and species) is constrained by the lack of available osteological reference collections for living species or by the suboptimal or incomplete preservation of the fossils. This results in that many fossil records are only assigned at the family or genus level (Walsh, 2001; Carrillo-Briceño, 2011; Oyanadel-Urbina et al., 2021). Some remains, such as otoliths, facilitate taxonomic identification due to their diagnostic features enabling greater resolution of the available fish assemblage richness to date (Schwarzhans and Nielsen, 2021), these being the main source of species-level taxonomic identifications. Also, if preserved, otoliths are much more abundant than identifiable parts of the bony skeleton and the two record-types are thus difficult to compare. It is also important to consider the potential taphonomic biases associated with otolith preservation, which could impact the representation of recorded taxa and the composition of the analyzed assemblages (Nolf, 2013; Schwarzhans, 2019). In addition, it is necessary to mention that the results could be influenced by the high number of taxa reported for the Navidad Formation and its equivalents (Early Miocene) compared to the rest of the Neogene. Future paleontological efforts focusing on other Neogene formations in Chile (e.g., Bahía Inglesa and Horcón formations) with records of bony fishes will help to reaffirm the interpretations presented in this work. Therefore, our study indicates that, despite the recent growth of analyses focused on Neogene bony fishes from Chile (Oyanadel-Urbina et al., 2021), the number of taxa should substantially rise with further sampling efforts. Also, additional records from underrepresented stratigraphic intervals (e.g., Middle Miocene) should constrain the timing of faunal change.
Perspective
Understanding taxonomic richness along the current Chilean coast is an endeavor that involves considering variations in the diversity and abundance of bony fishes. This need for a more comprehensive biogeographic framework implies incorporating analyses of under-investigated areas (i.e., the far north and south of Chile) and time spans, including paleontological and archaeological evidence. Once future studies contribute to an increase in fossil fish records for different types of fossils throughout Chile, detailed analyses could be developed that consider potential taphonomic and taxonomic biases in relation to the abundance of certain types of fossils (i.e., otoliths) over others (articulated skeletons). All of these additional analyses and broader spatial-temporal perspectives would greatly contribute to a better understanding of the biogeographic changes of fishes in Chile from the Neogene to the present.
The significant decrease in the abundance of certain coastal species, which has been made evident in recent decades due to human impact, highlights the necessity of comprehending the spatial distribution dynamics of fishes throughout their history, considering local and regional extinction events. Considering the current scenario of anthropogenic global climate change, reflecting upon these dynamics becomes crucial for implementing conservation plans and establishing protected areas.
ACKNOWLEDGEMENTS
This project was partially funded by the Fondo Nacional de Desarrollo Científico y Tecnológico (ANID/FONDECYT #3230610 and #1251475) and ANID/CENTROS REGIONALES (CLAP R20F0008). We thank the reviewers A. Collareta and W. Schwarzhans, as well as the editors J. Samuels and O. Kovalchuk, for their comments and suggestions, which greatly contributed to improving the manuscript.
REFERENCES
Arratia, G. 2015. Los peces osteíctios fósiles de Chile y su importancia en los contextos paleobiogeográfico y evolutivo. Publicación Ocasional del Museo Nacional de Historia Natural, Chile, 63:35–83.
Arratia, G., Chang, A. and Chong, G. 1975. Leptolepis opercularis n. sp. of the Upper Jurassic from Chile. Ameghiniana 12:350–358.
Arratia, G. and Schultze, H.P. 1999 Mesozoic fishes from Chile, pp. 565-593. In Arratia, G. and Schultze, H.P. (eds.), Mesozoic Fishes 2–Systematics and Fossil Record. Verlag Dr. F. Pfeil, München.
Báez, D.A.B. 2006. Estudio paleoambiental de la formación Pisco: localidad Ocucaje. Revista del Instituto de Investigación de la Facultad de Ingeniería Geológica, Minera, Metalúrgica y Geográfica, 9(18):64–69.
Béarez, P., Jackson, D., and Mollaret, N. 2015. Early Archaic fishing (12,600-9,200 cal yr BP) in the Semiarid North Coast of Chile. Journal of Island and Coastal Archaeology 10:133–148.
https://doi.org/10.1080/15564894.2014.940096
Béarez, P., Fuentes-Mucherl, F., Rebolledo, S., Salazar, D., and Olguín, L., 2016. Billfish foraging along the northern coast of Chile during the Middle Holocene (7400-5900 cal BP). Journal of Anthropological Archaeology, 41:85–195.
https://doi.org/10.1016/j.jaa.2016.01.002
Bell, M.C. 1985. The Chinches Formation: an Early Carboniferous lacustrine succession in the Andes northern Chile. Revista Geológica de Chile, 24:29–48.
Benites-Palomino, A., Valenzuela-Toro, A.M., Figueroa-Bravo, C., Varas-Malca, R.M., Nielsen, S.N., Gutstein, C.S., and Carrillo-Briceño, J.D. 2022. A new marine mammal assemblage from central Chile reveals the Pliocene survival of true seals in South America. Historical Biology, 34(11):2205–2217.
https://doi.org/10.1080/08912963.2021.2007528
Bianucci, G., Collareta, A., Bosio, G., Landini, W., Gariboldi, K., Gioncada, A., Lambert, O., Malinverno, E., De Muizon, C., Varas-Malca, R., Villa, I.M., Coletti, G., Urbina, M., and Di Celma, C. 2018. Taphonomy and palaeoecology of the lower Miocene marine vertebrate assemblage of Ullujaya (Chilcatay Formation, East Pisco Basin, southern Peru). Palaeogeography, Palaeoclimatology, Palaeoecology, 511:256–279.
https://doi.org/10.1016/j.palaeo.2018.08.013
Blisniuk, P.M., Stern, L.A., Chamberlain, C.P., Idleman, B., and Zeitler, P.K. 2005. Climatic and ecologic changes during Miocene surface uplift in the Southern Patagonian Andes. Earth and Planetary Science Letters, 230(1-2):125–142.
https://doi.org/10.1016/j.epsl.2004.11.015
Boyd, C.E., McNevin, A.A., and Davis, R.P. 2022. The contribution of fisheries and aquaculture to the global protein supply. Food security, 14(3):805–827.
https://doi.org/10.1007/s12571-021-01246-9
Carrillo-Briceño, J.D. 2011. Significado paleoambiental y paleoecológico de la fauna marina del Plioceno de la Fm. Horcón, Región de Valparaíso, Chile central. Unpublished thesis, Pontificia Universidad Católica de Valparaíso, Valparaíso, Chile.
Carrillo-Briceño, J.D., Carrillo, J.D., Aguilera, O.A., and Sanchez-Villagra, M.R. 2018. Shark and ray diversity in the Tropical America (Neotropics)–an examination of environmental and historical factors affecting diversity. PeerJ, 6:e5313.
https://doi.org/10.7717/peerj.5313
Chao, A. and Jost, L., 2012. Coverage based rarefaction and extrapolation: standardizing samples by completeness rather than size. Ecology, 93(12):2533–2547.
https://doi.org/10.1890/11-1952.1
Cione, A.L., Mennucci, J.A., Santalucita, F., and Hospitaleche, C.A. 2007. Local extinction of shark of genus Carcharias Rafinesque, 1810 (Elasmobranchii, Odontaspididae) in the eastern Pacific Ocean. Andean Geology, 1:139–145.
https://doi.org/10.4067/s0716-02082007000100007
Collareta, A., Landini, W., Lambert, O., Post, K., Tinelli, C., Di Celma, C., Panetta, D., Tripodi, M., Salvadori, P.A., Caramella, D., Marchi, D., and Bianucci, G. 2015. Piscivory in a Miocene Cetotheriidae of Peru: first record of fossilized stomach content for an extinct baleen-bearing whale. The Science of Nature, 102:1–12.
https://doi.org/10.1007/s00114-015-1319-y
De Muizon, C. and DeVries, T.J. 1985. Geology and paleontology of late Cenozoic marine deposits in the Sacaco area (Peru). Geologische Rundschau, 74(3):547–563.
https://doi.org/10.1007/bf01821211
Dekens, P.S., Ravelo, A.C., and McCarthy, M.D. 2007. Warm upwelling regions in the Pliocene warm period. Paleoceanography, 22:1–12.
https://doi.org/10.1029/2006pa001394
Di Celma, C., Malinverno, E., Collareta, A., Bosio, G., Gariboldi, K., Lambert, O., Landini, W., Pierantoni, P.P., Gioncada, A., Villa, I.M., Coletti, G., Muizon de, C., Urbina, M., and Bianucci, G. 2018. Facies analysis, stratigraphy and marine vertebrate assemblage of the lower Miocene Chilcatay Formation at Ullujaya (Pisco basin, Peru). Journal of Maps, 14(2), 257–268.
https://doi.org/10.1080/17445647.2018.1456490
Fischer, W., Krupp, F., Schneider, W., Sommer, C., and Carpenter, K.E. 1995. Guia FAO para la identificación de especies para los fines de la pesca: Pacifico centro-oriental. FAO.
Foote, M. and Sepkoski Jr., J.J. 1999. Absolute measures of the completeness of the fossil record. Nature, 398(6726):415–417.
https://doi.org/10.1038/18872
Fowler, H.W. 1945. Fishes of Chile. Systematic catalog. Apartado de la Revista Chilena de Historia Natural, Part I and II, 1–171.
Fricke, R., Eschmeyer, W.N., and Fong, J.D. 2024. Eschmeuer’s Catalogo of fishes: Genera/species by family/subfamily. http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp). Electronic version accessed 15 December 2024.
Froese R. and Pauly, D. 2024 FishBase. World Wide Web electronic publication, version (12/2024).
https://www.fishbase.org
Gasparini, Z.D. 1979. Comentarios críticos sobre los vertebrados mesozoicos de Chile. Conference Proceedings of the II Congreso Geológico Chileno, Arica, Chile. pp. 15–32.
Gay, C. 1854. Historia física y política de Chile. Zoología. Centro de Investigaciones Diego Barros Arana, Santiago, Chile.
Guicharrousse-Vargas, M., Villafaña, J.A., Carrillo-Briceño, J.D., Oyanadel-Urbina, P., Figueroa, R., Pérez-Marín, J.R., Rivadeneira, M.M., and Kriwet, J. 2021. The First Fossil Record of the Sawshark Pliotrema (Pristiophoridae) from the Neogene of the Southeastern Pacific (Chile). Ameghiniana, 58(2):122–131.
https://doi.org/10.5710/amgh.01.03.2021.3389
Guichenot, A. 1848. Historia física y política de Chile (in Gay, C.). Zoología, 2:328–30.
Gutstein, C.S., Yury-Yañez, R.E., Soto-Acuña, S., Suárez, M.E., and Rubilar-Rogers, D. 2008. Fauna de vertebrados y aspectos tafonómicos del ‘bonebed’ (Mioceno Tardío) de la Formación Bahía Inglesa. Conference Proceedings of the I Simposio de Paleontología en Chile, Santiago, Chile, pp. 102–108.
Hartley, A.J. and Chong, G. 2002. Late Pliocene age for the Atacama Desert: Implications for the desertification of western South America. Geology, 30(1):43–46.
https://doi.org/10.1130/0091-7
Hsieh, T., Ma, K., and Chao, A. 2016. iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods in Ecology and Evolution, 7(12):1451–1456.
https://doi.org/10.1111/2041-210x.12613
Hüne, M. 2019. Lista sistemática actualizada de los peces de Chile. Version 1.4. Fundación Ictiológica. Checklist dataset.
https://doi.org/10.15468/er28jy accessed via GBIF.org on 2024-08-19.
Ibaraki, M. 1997. Closing of the central American seaway and Neogene Coastal upwelling along the Pacific coast of America. Tectonophysics, 281:99–104.
https://doi.org/10.1016/S0040-1951(97)00161-3
Kiel, S. and S.N. Nielsen. 2010. Quaternary origin of the inverse latitudinal diversity gradient among southern Chilean mollusks. Geology, 38:955–958.
https://doi.org/10.1130/g31282.1
Kiel, S., Jakubowicz, M., Altamirano, A., Belka, Z., Dopieralska, J., Urbina, M., and Salas-Gismondi, R. 2023. The late Cenozoic evolution of the Humboldt Current System in coastal Peru: Insights from neodymium isotopes. Gondwana Research, 116:104–112.
https://doi.org/10.1016/j.gr.2022.12.008
Labarca, R., González-Guarda, E., Lizama-Catalán, Á., Villavicencio, N.A., Alarcón-Muñoz, J., Suazo-Lara, F., Oyanadel-Urbina P.A., Soto-Huenchuman, P., Salazar., C., Soto-Acuña, S., and Buldrini, K.E. 2020. Taguatagua 1: New insights into the late Pleistocene fauna, paleoenvironment, and human subsistence in a unique lacustrine context in central Chile. Quaternary Science Reviews, 238:106282.
https://doi.org/10.1016/j.quascirev.2020.106282
Lambeck, K. and Chappell, J. 2001. Sea level change through the last glacial cycle. Science, 292(5517):679–686.
https://doi.org/10.1126/science.1059549
Landini, W., Altamirano-Sierra, A., Collareta, A., Di Celma, C., Urbina, M., and Bianucci, G. (2017). The late Miocene elasmobranch assemblage from Cerro Colorado (Pisco Formation, Peru). Journal of South American Earth Sciences, 73:168–190.
https://doi.org/10.1016/j.jsames.2016.12.010
Long, J.D. 1993. Late Miocene and early Pliocene fish assemblages from the north central coast of Chile. Tertiary Research, 14:117–126.
Meléndez, R. 2008. Peces Marinos, p. 292–297. In Mena, M. and Figueroa-Fernández, A. (eds.), Biodiversidad de Chile Patrimonio y Desafíos. Ocho Libros Editores, Santiago, Chile.
Molina, J.I. 1782. Saggio sulla storia naturale del Chili. Stamperia di S. Tommaso d´Aquino, Bologna.
Morello, F., Calás, E., Torres, J., Borella, F., San Román, M., Martin, F., Contreras, L., Martínez, I., Alfonso-Durruty, M., and Massone, M. 2015. Punta Baxa 7: Sitio arqueológico de la costa norte de Tierra del Fuego, Estrecho de Magallanes (Chile). Magallania, 43:167–188.
https://doi.org/10.4067/s0718-22442015000200009
Nelson, J.S., Grande, T.C., and Wilson, M.V. 2016. Fishes of the World. John Wiley and Sons, Hoboken, New Jersey.
Nolf, D. 2002. Fossil record and paleobiogeography of Steindachneria (Pisces, Gadiformes). Courier Forschungs-Institut Senckenberg, 237:89-95.
Nielsen, S.N. and Glodny, J. 2009. Early Miocene subtropicalwater temperatures in the southeast Pacific. Palaeogeography, Palaeoclimatology, Palaeoecology, 280:480–488.
https://doi.org/10.1016/j.palaeo.2009.06.035
Ojeda, F.P., Labra, F., and Muñoz, A. 2000. Biogeographic patterns of Chilean littoral fishes. Revista Chilena de Historia Natural, 73:625–641.
https://doi.org/10.4067/s0716-078x2000000400007
Olguín, L., Salazar, D., Jackson, D. 2014. Early evidence for open sea navigation and fishing on the Pacific Coast of South America (Taltal, ~7,000 CAL BP). Chungará, 46: 177–192.
Otero, R.A. 2019. Loancorhynchus catrillancai gen. et sp. nov., a new swordfish (Xiphioidei, Blochiidae) from the Middle Eocene of central Chile. PeerJ, 7:e6671.
https://doi.org/10.7717/peerj.6671
Otero, R.A. and Suárez, M.E. 2022. Ray-finned fishes (Actinopterygii) from the lower Maastrichtian of Algarrobo, central Chile. Cretaceous Research, 130:105027.
https://doi.org/10.1016/j.cretres.2021.105027
Oyanadel-Urbina, P.A., Fouquet, N., Villafaña, J.A., Y Mourgues, F.A. 2018. Morphological description of fossils fishes from the Plio-Pleistoceno of the Pampa of Mejillones. Conference Proceedings of the I Congreso Chileno de Paleontología, Punta Arenas, Chile, pp. 301–307.
Oyanadel-Urbina, P., De Gracia, C., Carrillo-Briceño, J.D., Nielsen, S.N., Flores, H., Castelleto, V., Kriwet, J., Rivadeneira, M., and Villafaña, J.A. 2021. Neogene bony fishes from the Bahía Inglesa formation, northern Chile. Ameghiniana, 58(4):345–368.
https://doi.org/10.5710/amgh.26.05.2021.3375
Palacios, C. 2019. Sistemática y Tafonomía de los peces óseos de la Fm. Coquimbo, en el yacimiento paleontológico Los Dedos, Región de Atacama, Chile. Unpublished Bachelor thesis, Universidad de Atacama, Chile.
Partarrieu, D., Villafaña, J.A., Pinto, L., Mourgues, F. A., Oyanadel-Urbina, P.A., Rivadeneira, M.M., and Carrillo-Briceño, J.D. 2018. Neogene ‘horn sharks’ Heterodontus (Chondrichthyes: Elasmobranchii) from the Southeastern Pacific and their paleoenvironmental significance. Ameghiniana, 55(6):651–667.
https://doi.org/10.5710/amgh.19.10.2018.3202
Pease, C.M. 1987. Lyellian Curves and Mean Taxonomic Durations. Paleobiology, 13(4): 484–487.
https://doi.org/10.1017/s0094837300009064
Pequeño, G. 1989. Peces de Chile. Lista sistemática revisada y comentada. Revista de Biología Marina, 24:1–132.
Pérez, J. 2017. Nuevo serránido fósil (Actinopterygii, Perciformes) proveniente del Mioceno temprano de la Región de Arauco, Chile. Unpublished Master thesis, Universidad Austral de Chile, Valdivia, Chile.
Philippi, R.A. 1887a. Historia natural: sobre los tiburones i algunos otros peces de Chile. Anales de la Universidad de Chile, 71:3–42.
Philippi, R.A. 1887b. Los fósiles Terciarios i Cuartarios de Chile. F.A Brockhaus, Leipzig.
Pimiento, C., Griffin, J.N., Clements, C.F., Silvestro, D., Varela, S., Uhen, M.D., and, Jaramillo, C. 2017. The Pliocene marine megafauna extinction and its impact on functional diversity. Nature Ecology and Evolution 1:1100–1106.
https://doi.org/10.1038/s41559-017-0223-6
Quijada, B. 1913. Catálogo Ilustrado y Descriptivo de la Colección de Peces Chilenos y Extranjeros. Boletín del Museo Nacional de Historia Natural, Chile 5:7-130.
https://doi.org/10.54830/bmnhn.v5.1913.853
Rebolledo, S., Béarez, P., Salazar, D., Fuentes, F. 2016. Maritime fishing during the Middle Holocene in the hyperarid coast of the Atacama Desert. Quaternary International 391:3–11.
https://doi.org/10.1016/j.quaint.2015.09.051
Rebolledo, S., Béarez, P., Zurro, D., Santoro, C.M., and Latorre, C. 2021a. Big Fish or Small Fish? Differential Ichthyoarchaeological Representation Revealed by Different Recovery Methods in the Atacama Desert Coast, Northern Chile. Environmental Archaeology, 28(4):270–284.
https://doi.org/10.1080/14614103.2021.1886647
Rebolledo, S., Béarez, P., and Zurro, D. 2021b. Fishing during the early human occupations of the Atacama Desert coast: what if we standardize the data? Archaeological and Anthropological Sciences, 13:1–18.
https://doi.org/10.1007/s12520-021-01387-0
Reed, E.C. 1897. Catálogo de los peces chilenos. Anales de la Universidad de Chile, 98: 653–673.
Rivadeneira, M.M. and Marquet, P.A. 2007. Selective extinction of late Neogene bivalves on the temperate Pacific coast of South America. Paleobiology, 33(3):455–468.
https://doi.org/10.1666/06042.1
Rivadeneira, M.M. and Nielsen, S.N. 2017. Diversification dynamics, species sorting, and changes in the functional diversity of marine benthic gastropods during the Pliocene-Quaternary at temperate western South America. PLoS One, 12(10):e0187140.
https://doi.org/10.1371/journal.pone.0187140
Schwarzhans, W.W. 1980. Fish otoliths from the New Zealand Tertiary. New Zealand Geological Survey Report, 113:1–269.
Schwarzhans, W.W. 2019. Reconstruction of the fossil marine bony fish fauna (Teleostei) from the Eocene to Pleistocene of New Zealand by means of otoliths. With studies of Recent congroid, morid and trachinoid otoliths. Memorie Della Societa Italianà di Scienze Naturali e del Museo di Storia Naturale di Milano, 46:1–326
Schwarzhans, W.W and Aguilera, O. 2013. Otoliths of the Myctophidae from the Neogene of tropical America. Palaeo Ichthyologica, 13:83–150.
https://doi.org/10.1186/s13358-023-00302-5
Schwarzhans, W.W. and Nielsen, S.N. 2021. Fish otoliths from the early Miocene of Chile: a window into the evolution of marine bony fishes in the Southeast Pacific. Swiss Journal of Palaeontology, 140(1):1–62. https://doi.org/10.1186/s13358-021-00228-w
Seijo, J.C. and Defeo, O.C. 1997. Bioeconomía pesquera. Teoría, modelación y manejo FAO, Rome.
Stanley, S.M., Addicott, W.O., and Chinzei, K. 1980. Lyellian curves in paleontology: possibilities and limitations. Geology, 8(9):422-426.
https://doi.org/10.1130/0091-7613(1980)8<422:lcippa>2.0.co;2
Suárez, M.E., Marquardt, C., Lavenu, A., Marincovic, N., and Wilke, H.G. 2003. Vertebrados marinos Neógenos de la Formación la Portada, II Región, Chile. Conference Proceedings of the X Congreso Geológico Chileno, Concepción, p. 9.
Torres, J., Mahé, K., Dufour, J.L., Béarez P., and San Román, M. 2022. Characterizing seasonal fishing patterns and growth dynamics during the Middle and Late Holocene in the Strait of Magellan (Chilean Patagonia): Sclerochronological analysis of tadpole codling (Salilota australis) vertebrae. The Journal of Island and Coastal Archaeology, 17:1–20.
https://doi.org/10.1080/15564894.2020.1755393
Tsuchi, R. 2002. Neogene evolution of surface marine climate in Pacific and notes on related events. Revista Mexicana de Ciencias Geológicas, 19:260–270.
Valenzuela-Toro, A.M., Gutstein, C.S., Varas-Malca, R.M., Suárez, M.E., and Pyenson, N.D. 2013. Pinniped turnover in the South Pacific Ocean: new evidence from the Plio-Pleistocene of the Atacama Desert, Chile. Journal of Vertebrate Paleontology, 33(1):216–223.
https://doi.org/10.1080/02724634.2012.710282
Van Der Meer, D.G., Van Saparoea, A.V.D.B., Van Hinserbengen, D.J.J., Van De Weg, R.M.B., Godderis, Y., Le Hir, G., and Donnadieu, Y. 2017. Reconstructing first-order changes in sea level during the Phanerozoic and Neoproterozoic using strontium isotopes. Gondwana Research, 44:22–34.
https://doi.org/10.1016/j.gr.2016.11.002
Vermeij, G.J., DeVries, T.J., Griffin, M., Nielsen, S.N., Ochoa, D., Rivadeneira, M.M., Salas-Gismondi, R., and Valdovinos, F. 2024. The temperate marine Peruvian Province: How history accounts for its unusual biota. Ecology and Evolution, 14(7):e70048.
https://doi.org/10.1002/ece3.70048
Victor, P., Oncken, O., and Glodny, J. 2004. Uplift of the western Altiplano plateau: Evidence from the Precordillera between 20 and 21 S (northern Chile). Tectonics, 23(4):TC4004.
https://doi.org/10.1029/2003tc001519
Villafaña, J.A. and Rivadeneira, M.M. 2014. Rise and fall in diversity of Neogene marine vertebrates on the temperate Pacific coast of South America. Paleobiology, 40(4):659–674.
https://doi.org/10.1666/13069
Villafaña, J.A. and Rivadeneira, M.M. 2018. The modulating role of traits on the biogeographic dynamics of chondrichthyans from the Neogene to the present. Paleobiology, 44(2):251–262.
https://doi.org/10.1017/pab.2018.7
Villafaña, J.A., Nielsen, S.N., Klug, S., and Kriwet, J. 2019a. Early Miocene cartilaginous fishes (Chondrichthyes: Holocephali, Elasmobranchii) from Chile: diversity and paleobiogeographic implications. Journal of South American Earth Sciences, 96:102317.
https://doi.org/10.1016/j.jsames.2019.102317
Villafaña, J.A., Marramà, G., Hernandez, S., Carrillo-Briceño, J.D., Hovestadt, D., Kindlimann, R., and Kriwet, J. 2019b. The Neogene fossil record of Aetomylaeus (Elasmobranchii, Myliobatidae) from the southeastern Pacific. Journal of Vertebrate Paleontology, 39(1):e1577251.
https://doi.org/10.1080/02724634.2019.1577251
Villafaña, J.A., Rivadeneira, M.M., Pimiento, C., and Kriwet, J. 2023. Diversification trajectories and paleobiogeography of Neogene chondrichthyans from Europe. Paleobiology, 49(2):329–341.
https://doi.org/10.1017/pab.2022.40
Walsh, A.A. 2001. The Bahía Inglesa Formation Bonebed: Genesis and palaeontology of a Neogene konzentrat lagerstätte. Unpublished PhD thesis, University of Portsmouth, United Kingdom.
