

 Life on a Miocene barrier reef – fish communities and environments in the Medobory backreef
Life on a Miocene barrier reef – fish communities and environments in the Medobory backreef
Article number: 27.3.a46
https://doi.org/10.26879/1429
Copyright Palaeontological Association, September 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Tables
Submission: 10 July 2024. Acceptance: 30 August 2024.
ABSTRACT
The late Badenian Medobory barrier reef in western Ukraine offers a unique opportunity to study a backreef fish fauna of the Middle Miocene Central Paratethys Sea mostly by means of otoliths. New sampling at five locations has considerably upgraded the fish fauna, which now comprises 62 species. Nine new species are being described: Umbra euronota n. sp., Scythogobius minimus n. sp., Sarmatigobius cavatus n. sp., Buenia gibba n. sp., Ptereleotris tectus n. sp., Syngnathus vesculus n. sp., Apletodon conwayi n. sp., Palaeolebias winogradskyi n. sp., and Arnoglossus dispar n. sp. Syngnathids and gobiesocids are herewith for the first time identified in fossil otolith assemblages. The new samples also include otoliths from fine fractions down to 0.3 mm mesh size. The fine fraction was extremely rich in small, unidentifiable otoliths of presumably juvenile gobies in certain localities and yielded the small syngnathid and gobiesocid otoliths. The numerous small goby otoliths are thought to represent a fish nursery setting, probably in a seagrass environment. The sampling of rich otolith communities also facilitates the recognition of micro-environments in the Medobory backreef lagoon, such as near-reef (patch reef), deeper lagoon and seagrass meadow faunal compositions that are also largely supported by the evaluation of other co-occurring biota (foraminifera, ostracods, and molluscs).
Werner Schwarzhans. Zoological Museum, Natural History Museum of Denmark, Universitetsparken 15, Copenhagen, 2100 Denmark, and Ahrensburger Weg 103, Hamburg, 22359 Germany. wwschwarz@t-online.de
Oleksandr Klots. Khmelnytskyi city organization of the National Union of Local Historians of Ukraine, Maidan Nezalezhnosti 1, Khmelnytskyi, 29001 Ukraine. o_m_klets@ukr.net
Oleksandr Kovalchuk. Department of Palaeontology, National Museum of Natural History, National Academy of Sciences of Ukraine, Bohdana Khmelnytskoho 15, Kyiv, 01054 Ukraine and Department of Palaeozoology, Faculty of Biological Sciences, University of Wrocław, Sienkiewicza 21, Wrocław, 50-335 Poland and Department of Biology and Biology Teaching Methodology, Faculty of Natural Sciences and Geography, A.S. Makarenko Sumy State Pedagogical University, Romenska 87, Sumy, 40002 Ukraine.(Corresponding author) biologiest@ukr.net
Anastasiia Dubikovska. Department of Biology and Biology Teaching Methodology, Faculty of Natural Sciences and Geography, A.S. Makarenko Sumy State Pedagogical University, Romenska 87, Sumy, 40002 Ukraine and Schmalhausen Institute of Zoology, National Academy of Sciences of Ukraine, Bohdana Khmelnytskoho 15, Kyiv, 01054 Ukraine. oakovska@gmail.com
Tamara Ryabokon. Institute of Geological Sciences, National Academy of Sciences of Ukraine, Olesia Honchara 55b, Kyiv, 01054 Ukraine. tamararyabokon@gmail.com
Volodymyr Kovalenko. Institute of Geological Sciences, National Academy of Sciences of Ukraine, Olesia Honchara 55b, Kyiv, 01054 Ukraine. kovva@ukr.net
Keywords: Medobory barrier reef; otoliths; Gobiidae; Labridae; micro-environment; seagrass community; new species
Final citation: Schwarzhans, Werner, Klots, Oleksandr, Kovalchuk, Oleksandr, Dubikovska, Anastasiia, Ryabokon, Tamara, and Kovalenko, Volodymyr. 2024. Life on a Miocene barrier reef – fish communities and environments in the Medobory backreef. Palaeontologia Electronica, 27(3):a46.
https://doi.org/10.26879/1429
palaeo-electronica.org/content/2024/5322-medobory-backreef-fish-fauna
Copyright: September 2024 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
https://zoobank.org/BD629D9B-2B92-4044-B371-4363A90CEF65
INTRODUCTION
The Medobory barrier reef was a Middle Miocene reef with coralline algae as the main framework builder in western Ukraine and extending into adjacent southern Poland and northern Moldova, located along the eastern margin of the Fore-Carpathian Basin (Górka et al., 2012). The Medobory reef existed for only a brief period, during the late Badenian (13.80-12.65 Ma after Harzhauser et al., 2020). It was a true barrier reef unlike the discontinuous reef bodies in the Middle and Late Miocene of Spain and elsewhere in the Mediterranean (Permanyer and Esteban, 1973; Esteban, 1980). The Medobory barrier reef is remarkable for two reasons: (1) it is believed to have grown near the northern limit of temperature conditions suitable for coralline-algal growth at the time (Górka et al., 2012), possibly enhanced by a warm, shallow environment over the wide shelf; and (2) it was formed after the mid-Badenian ecological crisis marked by the Karaganian event in the Eastern Paratethys and an evaporitic event across large parts of the Central Paratethys (see Peryt, 2006; Harzhauser et al., 2014, 2018 and references therein; Báldi et al., 2017 for more details). Even though the ephemeral marine connectivity existed through the Slovenian Gateway during some time in the late Badenian (Bartol et al., 2014), re-immigration of deep-water and reef-related fishes may have been restricted. The fish communities of the Medobory barrier reef were therefore dominated by endemics that arose from primarily not reef-adapted survivors, chiefly gobies, of the mid-Badenian crisis (Schwarzhans et al., 2022). A rich otolith-based fish fauna has been described from the Medobory backreef by Schwarzhans and Kovalchuk (2022) and subsequently by Schwarzhans et al. (2022). Rich new finds sampled from additional localities have not only raised the count to 62 species including nine new species and 14 in open nomenclature, but also allowed a more diversified picture to be drawn of environmental settings and fish communities in this unique Lagerstätte.
GEOLOGICAL SETTING
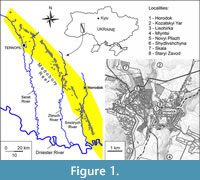 The studied material originates from six localities (Figure 1) including four new ones (Lisohirka, Novyi Pliazh, Skala, and Staryi Zavod) and two that have been investigated previously (Mlyntsi and Shydlivshchyna), fish remains from which were processed and published earlier (Schwarzhans et al., 2022). Mlyntsi (49.1387 N, 26.5981 E) is situated within the Chornovodka River valley near the village of Chornyvody. Shydlivshchyna (49.1717 N, 26.5519 E) is confined to the ravine slope of the Smotrych River north of the city of Horodok. In addition to fish remains, shells of gastropods and bivalves, algae, serpulids, ostracods, bryozoans, and sponges were found in a detrital reef-derived calcarenite (Schwarzhans et al., 2022). Lisohirka (49.1424 N, 26.5645 E) is an abandoned quarry on the northern outskirts of the eponymous village. It occupies the lower part of the right slope of the Smotrych River valley. The fossils were obtained from a layer of unconsolidated uniform reef calcarenite, that is, from within a reef body. A characteristic feature of this layer is the abundance of gastropods up to 3 mm in size as well as a rich assemblage of foraminifera, ostracods, bryozoans, serpulids, remains of sea urchins, and crustaceans. Novyi Pliazh (49.1791 N, 26.5687 E) is confined to the left slope of the Smotrych River valley and represents a natural exposure of detrital calcarenite yielding numerous fish remains. Skala (49.1911 N, 26.5604 E) is an abandoned quarry on the northwestern outskirts of the city of Horodok. Fossiliferous rocks belong to the lowermost part of the upper Badenian sedimentary succession (Pidhirtsi Beds), represented by white quartzitic sands and sandstones in an off-reef position. Staryi Zavod (49.1692 N, 26.5889 E) is located within the city of Horodok, in the steep left slope of the Trostyanets River valley, left tributary of the Smotrych River. The fossils originate from loose, fine-grained reef calcarenite, overlain by a thin (10-30 cm) layer of gravel and underlain by quartz-limestone sands. In the material from this locality are many Cerithioidea shells.
The studied material originates from six localities (Figure 1) including four new ones (Lisohirka, Novyi Pliazh, Skala, and Staryi Zavod) and two that have been investigated previously (Mlyntsi and Shydlivshchyna), fish remains from which were processed and published earlier (Schwarzhans et al., 2022). Mlyntsi (49.1387 N, 26.5981 E) is situated within the Chornovodka River valley near the village of Chornyvody. Shydlivshchyna (49.1717 N, 26.5519 E) is confined to the ravine slope of the Smotrych River north of the city of Horodok. In addition to fish remains, shells of gastropods and bivalves, algae, serpulids, ostracods, bryozoans, and sponges were found in a detrital reef-derived calcarenite (Schwarzhans et al., 2022). Lisohirka (49.1424 N, 26.5645 E) is an abandoned quarry on the northern outskirts of the eponymous village. It occupies the lower part of the right slope of the Smotrych River valley. The fossils were obtained from a layer of unconsolidated uniform reef calcarenite, that is, from within a reef body. A characteristic feature of this layer is the abundance of gastropods up to 3 mm in size as well as a rich assemblage of foraminifera, ostracods, bryozoans, serpulids, remains of sea urchins, and crustaceans. Novyi Pliazh (49.1791 N, 26.5687 E) is confined to the left slope of the Smotrych River valley and represents a natural exposure of detrital calcarenite yielding numerous fish remains. Skala (49.1911 N, 26.5604 E) is an abandoned quarry on the northwestern outskirts of the city of Horodok. Fossiliferous rocks belong to the lowermost part of the upper Badenian sedimentary succession (Pidhirtsi Beds), represented by white quartzitic sands and sandstones in an off-reef position. Staryi Zavod (49.1692 N, 26.5889 E) is located within the city of Horodok, in the steep left slope of the Trostyanets River valley, left tributary of the Smotrych River. The fossils originate from loose, fine-grained reef calcarenite, overlain by a thin (10-30 cm) layer of gravel and underlain by quartz-limestone sands. In the material from this locality are many Cerithioidea shells.
MATERIAL AND METHODS
In total, 114 kg of the fossiliferous rock were processed including 10 kg from Lisohirka, 30 kg from Mlyntsi, 40 kg from Novyi Pliazh, 20 kg from Shydlivshchyna, 14 kg from Staryi Zavod, and 15 kg from the Skala locality (the latter was dry sieved using a mesh size of 1.7 mm).
A single rock sample was taken from the ravine slope in each locality. A slightly different approach was applied to obtain most of the studied material. Sieves at this stage were not used at all. The rock samples were washed in small portions by the vortex (centrifuge) method in a transparent container with water until the silty particles were completely dissolved. Using a large brush, the rock sample rises into the thickness of the vortex by continuous circular movement, and it is intensively washed up to abrupt stop stirring. Large fractions settle faster than small ones. At this moment, it is important to carefully drain such a cloudy water, thus separating the fractions. Then, the rock sample is washed again until the water becomes almost transparent, and the coarse fraction is clearly visible at the bottom of the container. Then the material is dried, and only thereafter it has been sieved. For efficiency and for further visual selection of fossils under a binocular microscope, the rock was sieved and divided into three fractions: large (particles larger than 5 mm), medium (≥1.7 mm) and small (down to 0.3 mm). Otoliths, bone remains, foraminiferans, serpulids, bryozoans, sea urchins, molluscs (gastropods, bivalves), and brachiopods were selected. This collecting approach was used working in the Lisohirka, Novyi Pliazh, and Staryi Zavod localities.
All otoliths were studied with a reflected-light microscope. Photographs were captured with a Canon EOS mounted on the phototube of a Wild M400 photomacroscope. They were taken at regular field-of-depth levels for each view, with the camera being remotely controlled from a computer. The individual photographs of each view were stacked using Helicon Soft’s (Kharkiv, Ukraine) Helicon Focus software. The continuously focused pictures were processed with Adobe Photoshop to enhance contrast, balance exposition, or retouch small inconsistencies, such as sand grains, encrustations, or pigmentation spots insofar as doing so was possible without altering the otolith morphology. All figures show right otoliths except for pleuronectiforms, because they may exhibit side dimorphism. Otherwise, left otoliths have been reversed to facilitate better comparison and are annotated accordingly. All figured otoliths show inner faces unless otherwise annotated.
The specimens considered are housed in the Department of Palaeontology at the National Museum of Natural History, National Academy of Sciences of Ukraine, Kyiv (NMNHU-P, collection PI); and in the Senckenberg Museum, Frankfurt am Main, Germany (SMF, collection PO).
SYSTEMATIC PALAEONTOLOGY
Part 1. Otoliths (by Schwarzhans, Klots, and Kovalchuk)
The otolith-based fossil record from the Medobory backreef localities is summarised in Table 1. The morphological descriptive terminology of otoliths follows Koken (1884) with amendments by Chaine and Duvergier (1934) and Schwarzhans (1978). The classification follows Nelson et al. (2016). We have extensively documented the retrieved otoliths (3177 specimens) from the new samples. Since most species have been recently described in Schwarzhans et al. (2022), however, we only describe new species here and selectively comment on new records, including those in open nomenclature (shown in bold in Table 1).
Class ACTINOPTERYGII Klein, 1885
Subdivision TELEOSTEI Müller, 1846
Order ANGUILLIFORMES Regan, 1909
Family MURAENIDAE Rafinesque, 1810
Muraenidae indet.
Figure 2A-C
Material. 1 otolith, NMNHU-P PI 2595, Novyi Pliazh.
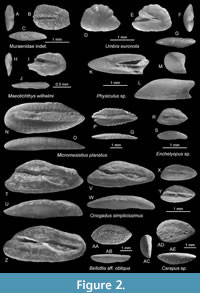 Discussion. The single small otolith of 1.5 mm in length shows the typical otolith shape with a blunt anterior tip and a somewhat tapering posterior tip and sulcus shape of muraenid otoliths. The sulcus is narrow, deep, with slightly wavy margins, anteriorly open and posteriorly terminating at considerable distance from the posterior rim of the otolith. Moray eels are typical fishes in extant reef environments, but their fossil otolith record is extremely scarce with a single unambiguous record from the Neogene of the Caribbean (Aguilera and Rodrigues de Aguilera, 2001).
Discussion. The single small otolith of 1.5 mm in length shows the typical otolith shape with a blunt anterior tip and a somewhat tapering posterior tip and sulcus shape of muraenid otoliths. The sulcus is narrow, deep, with slightly wavy margins, anteriorly open and posteriorly terminating at considerable distance from the posterior rim of the otolith. Moray eels are typical fishes in extant reef environments, but their fossil otolith record is extremely scarce with a single unambiguous record from the Neogene of the Caribbean (Aguilera and Rodrigues de Aguilera, 2001).
Genus ESOCIFORMES Bleeker, 1859
Family Umbridae Bonaparte, 1845
Genus UMBRA Kramer in Scopoli, 1777
Umbra euronota n. sp.
Figure 2D-G
zoobank.org/C90F1413-
Holotype. NMNHU-P PI 2596 (Figure 2D-G), Novyi Pliazh, western Ukraine, late Badenian.
Etymology. From euronotus (Latin) = easterly, referring to the occurrence of the species.
Diagnosis. OL:OH = 1.45. Ventral rim nearly flat and horizontal; dorsal rim domed, broadly crenulated. Rostrum long (17% of OL), inferior, with rounded tip. OL:SuL = 1.35. Cauda short and narrow compared to funnel-shaped ostium; OsL:CaL = 1.8; OsH:CaH = 2.3.
Description. A single, well-preserved and relatively small otolith of 1.65 mm in length. OL:OH = 1.45; OH:OT = 2.5. Its dorsal rim is domed, regularly curved without angles, with a broad, regular crenulation. The ventral rim is flat, horizontal, and slightly undulating. The anterior tip has a massive inferior rostrum (17% of OL, measured from the tip of the rostrum to the deepest notch of the excisura); antirostrum and excisura are minute. The posterior tip is angular, inferior, at about the same level as rostrum but more rounded. The inner face is almost flat with a centrally positioned, deepened sulcus. The sulcus is widely opening anteriorly, terminating distant from the posterior rim (OL:SuL = 1.35). The ostium is funnel-shaped, distinctly longer and wider than small and narrow cauda (OsL:CaL = 1.8; OsH:CaH = 2.3). The dorsal depression is distinct, narrow, and broad triangular; the ventral field lacks the ventral furrow but has a distinct crista inferior below the cauda. The outer face is moderately convex with a broad and smooth postcentral umbo.
Discussion. Nolf (2013) listed 14 otolith-based umbrid species from the European Cenozoic ranging in age from Eocene to Late Miocene and pertaining to three different genera. Nine species are placed in the genera Umbra, Palaeumbra or are generically undefined. Another group of small umbrid otoliths have been placed in the fossil otolith-based genus Mikroumbra Reichenbacher and Weidmann, 1992, but these species do not resemble U. euronota (see Reichenbacher and Weidmann, 1992; Reichenbacher, 1993). Umbra euronota differs from all of them in the relatively short sulcus and the extremely narrow and short cauda, which we consider adequate for a diagnostic definition. The species is morphologically closest to Umbra praekrameri Weinfurter, 1950, from the Late Miocene, Pannonian of Austria. Umbra euronota represents one of the rare freshwater fishes found in the Medobory backreef and is probably derived from the presumed nearby river discharge.
Order Gadiformes Goodrich, 1909
Family Moridae Berg, 1940
Genus PHYSICULUS Kaup, 1858
Physiculus sp.
Figure 2K-M
Material. 1 otolith, NMNHU-P PI 2600, Novyi Pliazh.
Discussion. The single, small otolith of 2.5 mm in length lacks the fragile rear part. It is a typical otolith of the genus Physiculus that differs from P. moravicus (Brzobohatý and Schultz, 1978) from the Badenian of the Central Paratethys (Brzobohatý and Schultz, 1978) in the strongly pointed anterior tip and the smooth dorsal and ventral rims and outer face.
Family Gaidropsaridae Jordan and Evermann, 1898
Genus ENCHELYOPUS Bloch and Schneider, 1801
Enchelyopus sp.
Figure 2R-S
Material. 1 otolith, NMNHU-P PI 2616, Mlyntsi.
Discussion. A single otolith of 1.3 mm in length differs from the ubiquitous Onogadus simplicissimus (Schubert, 1906) (Figure 2T-Z) in being more compressed (OL:OH = 1.8 vs. 2.05-2.55) and thus indicating the presence of a second gaidropsarid fish in the Medobory backreef.
Order Gobiiformes Thacker, 2009
Family Gobiidae Cuvier, 1816
GOBIUS Lineage sensu Agorreta et al., 2013
Genus GOBIUS Linnaeus, 1758
Gobius supraspectabilis Schwarzhans, Brzobohatý, Radwańska and Procházka, 2020
Figure 3H-J
2020a Gobius supraspectabilis; Schwarzhans et al., pl. 4, figs. 10-13.
Material. 4 otoliths, NMNHU-P PI 2631, Staryi Zavod.
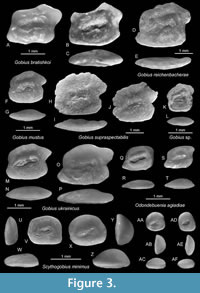 Discussion. Gobius supraspectabilis is easily recognised by its comparatively small sulcus and subcaudal iugum and the intensely and irregularly crenulated dorsal rim. The small sulcus resembles otoliths of the extant of Zosterisessor ophiocepalus (Pallas, 1814) (see Schwarzhans et al., 2020a for figures of otoliths), a genus that is commonly synonymised with Gobius (see Fricke et al., 2024 for more details). It is therefore possible that G. supraspectabilis represents the lineage of Zosterisessor or an extinct clade.
Discussion. Gobius supraspectabilis is easily recognised by its comparatively small sulcus and subcaudal iugum and the intensely and irregularly crenulated dorsal rim. The small sulcus resembles otoliths of the extant of Zosterisessor ophiocepalus (Pallas, 1814) (see Schwarzhans et al., 2020a for figures of otoliths), a genus that is commonly synonymised with Gobius (see Fricke et al., 2024 for more details). It is therefore possible that G. supraspectabilis represents the lineage of Zosterisessor or an extinct clade.
Gobius sp.
Figure 3K-L
Material. 2 otoliths, NMNHU-P PI 2638, Staryi Zavod.
Discussion. Two small, compressed otoliths with a ratio OL:OH just below 1.0 and about 1.0 mm in length are interpreted to represent a further, unidentified Gobius species. The compressed shape is atypical for Gobius otoliths but is occasionally seen in juvenile Gobius otoliths and certain species such as the extant G. vittatus Vinciguerra, 1883 (see Gut et al., 2020; Schwarzhans et al., 2020b for figures of otoliths). Similarly, compressed otoliths are found in representatives of the genus Lesueurigobius, which differ, however, in the large subcaudal iugum, as well as in Proterorhinus showing a morphologically more reduced sulcus.
BENTHOPHILUS Lineage (Benthophilini in Agorreta et al., 2013)
Genus SCYTHOGOBIUS Schwarzhans and Bratishko, 2023 in Schwarzhans et al., 2023
Scythogobius minimus n. sp.
Figure 3U-AF
zoobank.org/89A6D401-
Holotype. NMNHU-P PI 2645 (Figure 3X-Z), Novyi Pliazh, western Ukraine, late Badenian.
Paratypes. 1 otolith, SMF PO 101.346, Novyi Pliazh; 2 specimens, SMF PO 101.347, Lisohirka.
Additional material. 3 otoliths: NMNHU-P PI 2646, 1 specimen, Mlyntsi; 2 specimens, NMNHU-P PI 2647, Lisohirka.
Etymology. From minimus (Latin) = small, referring to the small size of the otoliths.
Diagnosis. Compact, roundish to oval shape; flat inner face and strongly convex, smooth outer face. Sulcus small, narrow, inclined at 18-28° and slightly forward positioned, deepened, simple fusiform shaped and without subcaudal iugum. OL:SuL = 2.2-2.5.
Description. Small, thick, roundish to oval otoliths with sizes up to 1.25 mm in length (holotype). OL:OH = 1.2 in specimens > 1 mm in length (Figure 3U-Z) and 0.9-1.0 in specimens < 0.7 mm in length (Figure 3AA-AF) showing a distinctive allometric ontogeny; OH:OT = 1.6-2.0. Otolith rims are well rounded without prominent angles and smooth; dorsal and anterior rims are regularly rounded, the ventral rim with a relatively low curvature and the posterior rim inclined with somewhat projecting tip at joint of posterior and ventral rims. The inner face is nearly completely flat, smooth, with a small, slightly forward positioned, a simple fusiform, deepened sulcus without the subcaudal iugum. OL:SuL = 2.2-2.5; 18-28° inclination. There is no discernible dorsal depression or ventral furrow. The outer face is strongly convex and smooth.
Discussion. The otoliths of Scythogobius are highly characteristic because of their morphologically reduced otolith and sulcus shapes and the flat inner and strongly convex outer face resulting in a comparatively thick otolith. The genus Scythogobius was established based on S. spissus Schwarzhans and Bratishko, 2023 (Schwarzhans et al., 2023) from the late Bessarabian of the Crimea (Ukraine). Scythogobius minimus differs from S. spissus in the lower index OL:OH at all sizes (0.9-1.2 vs. 1.25-1.4) and the smaller sulcus size (OL:SuL = 2.2-2.5 vs. 1.75-2.15).
Aphia Lineage sensu Agorreta et al., 2013
Genus LESUEURIGOBIUS Whitley, 1950
Lesueurigobius sp.
Figure 4A-I
?2005 “genus Gobiidarum ” sp. 3; Hoedemakers and Batllori, pl. 11, figs. 5-8.
2020b Lesueurigobius sp.; Schwarzhans et al., pl. 2, figs. 20-21.
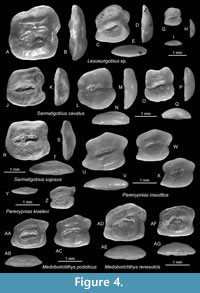 Material. 9 otoliths: 5 specimens, NMNHU-P PI 2648 (Figure 4A-I), Lisohirka; 4 specimens, NMNHU-P 2649, Mlyntsi.
Material. 9 otoliths: 5 specimens, NMNHU-P PI 2648 (Figure 4A-I), Lisohirka; 4 specimens, NMNHU-P 2649, Mlyntsi.
Description. Compressed, massive otoliths up to about 3.0 mm in length; OL:OH = 0.9-0.95; OH:OT = 2.9-3.2. The otolith shape is rectangular with near vertical anterior and posterior rims and horizontal, nearly flat ventral rim. The dorsal rim is high, postdorsally pronounced, with a predorsal concavity in specimens larger than 2 mm. The predorsal angle is high, rounded, not projecting; the postdorsal projection broad but short. The anterior rim sometimes with a small incision at the level of the ostial tip; the posterior rim with a small incision at the level of the caudal tip. All rims are relatively smooth. The inner face is almost flat, with a moderately large, slightly inframedian sulcus. The sulcus is sole-shaped, moderately inclined at 11-18° and has a distinct subcaudal iugum; OL:SuL = 1.8-2.1. The dorsal depression is broad, distinct; the ventral furrow is distinct, close to the ventral rim of the otolith and not joining up to the dorsal depression around the sulcus. The outer face is moderately convex and smooth.
Discussion. The most common Lesueurigobius species in the Badenian of the Paratethys is L. vicinalis (Koken, 1891). Lesueurigobius vicinalis exhibits a broad variability (see Schwarzhans et al., 2020b) and therefore distinction from coeval species such as L. magnijugis Schwarzhans, 2017, and the specimens here denoted as Lesueurigobius sp. relies on relatively subtle features considered to be sufficiently stable. The most characteristic feature of Lesueurigobius sp. is the concavity of the predorsal rim in larger specimens (see also Schwarzhans et al., 2020b). It differs additionally from L. vicinalis in the higher position of the predorsal angle, and from L. magnijugis in the narrower sulcus and the more irregularly curved dorsal rim (vs. expanded and regularly curved).
Genus SARMATIGOBIUS Reichenbacher and Bannikov, 2022
Sarmatigobius cavatus n. sp.
Figure 4J-Q
zoobank.org/DF3FE8DE-
Holotype. NMNHU-P PI 2650 (Figure 4O-Q), Novyi Pliazh, western Ukraine, late Badenian.
Paratypes. 2 otoliths, SMF PO 101.348, same data as holotype.
Etymology. From cavatus (Latin) = excavated, referring to a concave dorsal rim observed in the holotype.
Diagnosis. OL:OH = 0.87-0.97. Ventral rim curved; dorsal rim high, expanded, with flat or concave central portion. OL:SuL = 1.8-2.1; sulcus inclination 5-12°. Cauda narrow; subcaudal iugum distinct.
Description. Thick, high-bodied otoliths up to 1.8 mm in length (holotype 1.4 mm). OL:OH = 0.87-0.97; OH:OT = 2.5-3.0. The anterior rim is nearly vertical, smooth; the posterior rim is nearly vertical or slightly inclined, with a slight concavity at the level of the caudal tip. The dorsal rim is high, with a well-developed postdorsal angle and slightly lower predorsal angle; the middorsal rim is slightly inclined, straight, or concave. All rims are smooth. The inner face is flat, smooth, with a centrally positioned, deepened, sole-shaped sulcus. The latter is relatively short (OL:SuL = 1.8-2.1), with a low inclination (5-12°), with a narrow cauda and a distinct and large subcaudal iugum. The dorsal depression is deep but with indistinct margins, open to the dorsal rim of otolith. The ventral furrow is relatively weak, close to the ventral rim of the otolith, not connected to the dorsal depression around the sulcus. The outer face is smooth and distinctly convex.
Discussion. Reichenbacher and Bannikov (2023) found otoliths in situ and described Sarmatigobius compactus from the Sarmatian of Moldova. They also placed Hesperichthys iugosus Schwarzhans, Brzobohatý and Radwańska, 2020 into Sarmatigobius. Thus, otoliths of Sarmatigobius differ from those of Hesperichthys in the presence of a large subcaudal iugum (vs. no subcaudal iugum) (see also Schwarzhans et al., 2017a). Sarmatigobius cavatus differs from its two congeners in the more compressed shape (OL:OH = 0.87-0.97 vs. 1.0-1.1) and the overall shape of the dorsal rim. It shares with its congeners the deep dorsal depression that reaches close to the dorsal margin of the sulcus and opens to the dorsal margin of the otolith, and the large subcaudal iugum. The specimens show a certain degree of variability in the overall shape, but the curved ventral otolith rim, the convex to flat dorsal rim and the compressed shape remain characteristical.
POMATOSCHISTUS Lineage sensu Agorreta et al., 2013
Genus BUENIA Iljin, 1930
Buenia gibba n. sp.
Figure 5K-S
zoobank.org/9A08D560-
Holotype. NMNHU-P PI 2677 (Figure 5K-M), Novyi Pliazh, western Ukraine, late Badenian.
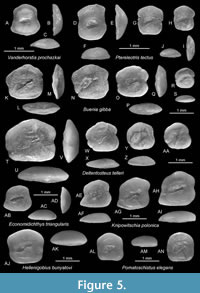 Paratypes. 3 specimens, SMF PO 101.349, Stary Zavod, western Ukraine, late Badenian.
Paratypes. 3 specimens, SMF PO 101.349, Stary Zavod, western Ukraine, late Badenian.
Additional material. 17 otoliths: 2 specimens, NMNHU-P 2678, Lisohirka; 15 specimens, NMNHU-P PI 2679, Staryi Zavod.
Etymology. From gibbus (Latin) = bent, referring to the convex inner face, the main distinguishing to juvenile otoliths of Deltentosteus telleri.
Diagnosis. Nearly quadratic otolith shape; OL:OH = 0.95-1.05. Predorsal angle slightly protruding; dorsal rim highest middorsally. Inner face distinctly convex, smooth. Sulcus steeply inclined, with broad ostium and narrow, tapering cauda; sulcus inclination angle 23-28°. OL:SuL = 1.75-2.2.
Description. Small, thick, compact otoliths with nearly quadrangular shape up to 1.5 mm in length (holotype). OL:OH = 0.95-1.05; OH:OT = 2.6-2.85. The anterior rim is usually inclined upward toward a rounded, slightly protruding predorsal angle. The dorsal rim is curved, highest at or slightly behind the middorsal point. The posterior rim vertical, without a distinct postdorsal projection, usually with a slight concavity at the level of the caudal tip. The ventral rim is flat horizontal. All rims are smooth except dorsal and anterior rims sometimes undulate slightly. The inner face is distinctly convex, increasing in convexity with size, and smooth with a relatively large, shallow, steeply inclined and pronounced asymmetrical otolith. OL:SuL = 1.75-2.2; sulcus inclination angle 23-28°, ostium more steeply inclined than cauda. The ostium is strongly widened with a rounded ostial lobe, the cauda is narrow and tapering. Subcaudal iugum is indistinct and narrow. The dorsal depression is indistinct; the ventral furrow is narrow, relatively faint but long and joining up to a dorsal field around the sulcus. The outer face about as convex as the inner face.
Discussion. Several species occur in parallel with similar looking otoliths such as Deltentosteus telleri (Schubert, 1906), particularly its juvenile specimens (Figure 5W-AA), Hellenigobius bunyatovi (Bratishko, Schwarzhans and Reichenbacher, 2015) (Figure 5AJ-AK) and Pomatoschistus elegans (Procházka, 1900) (Figure 5AL-AN). Buenia gibba differs from all of them in the distinctly convex inner face that is as convex as the outer face (vs. flat or slightly convex, less than outer face). From the juveniles of D. telleri it differs additionally in the rectangular angle at the joint of the posterior and ventral rims (vs. projecting), while larger otoliths of D. telleri are distinctly more elongate (Figure 5T). Buenia gibba differs additionally from Hellenigobius bunyatovi in the vertical posterior rim (vs. dorsally inclined) and the stronger ostial lobe and from Pomatoschistus elegans additionally in being less high bodied (OL:OH 0.95-1.05 vs. 0.9-0.98) and the inclined anterior rim (vs. vertical).
Several Buenia species have recently been recognised or described from the Neogene of the Paratethys and the Mediterranean: B. rudolticensis (Procházka, 1900) from the late Badenian and Sarmatian of the Central Paratethys, Buenia rueckertae (Schwarzhans, 2014) from the Serravallian of the Mediterranean, the extant Buenia affinis Iljin, 1930, from the Messinian and Zanclean of the Mediterranean, Buenia pisiformis Schwarzhans, Agiadi and Carnevale, 2020, from the Tortonian and Messinian of the Mediterranean, and Buenia pulvinus van Hinsbergh and Hoedemakers, 2022, from the Pliocene of the Mediterranean and adjacent NE Atlantic. The otoliths of Buenia pulvinus are mostly more elongate than those in B. gibba (OL:OH = 1.07-1.12 vs. 0.95-1.05) and show regularly curved and not projecting angles. Buenia rudolticensis is usually more highly bodied than B. gibba (OL:OH mostly 0.92-0.95, rarely 1.0 vs. 0.95-1.05) and shows a relatively narrow sulcus (vs. wide ostium and tapering cauda) and a vertical anterior rim (vs. inclined). Buenia rueckertae is similar in otolith shape to B. gibba but more slender (OL:OH = 1.05-1.15 vs. 0.95-1.05), and its cauda is normally wide and not tapering. The otoliths of Buenia pisiformis are more high bodied than those of B. gibba (OL:OH = 0.89-0.97 vs. 0.95-1.05) and show an extremely small sulcus (OL:SuL = 2.6-2.9 vs. 1.75-2.2). The most closely resembling species is the extant B. affinis with a similar index OL:OH of 0.95-1.1, but with a more gently curved dorsal rim and rounded pre- and postdorsal angles compared to B. gibba and normally a wide and not tapering cauda. We consider B. gibba to represent a species adapted to the environment of the Medobory backreef. It likely represents a short-lived endemism in the late Badenian.
Gobiidae indet. erod. and juv.
Material. 1214 otoliths: 105 specimens, NMNHU-P PI 2701, Lisohirka; 1 specimen, NMNHU-P PI 2702, Mlyntsi; 1108 specimens, NMNHU-P PI 2703, Staryi Zavod.
Discussion. A large amount of small gobiid otoliths has been found in some of the localities considered at sizes smaller than 0.8 mm in length, often smaller than 0.5 mm, which probably stem from juvenile or even larval gobiids that cannot be identified up to species or even genus level. Their uneven distribution pattern in various localities is thought to indicate a palaeoenvironmental signal that will be discussed later.
Family Microdesmidae Regan, 1912s
Genus PTERELEOTRIS Gill, 1863
Ptereleotris tectus n. sp.
Figure 5D-J
zoobank.org/B435F4FF-
Holotype. NMNHU-P PI 2704 (Figure 5D-F), Novyi Pliazh, western Ukraine, late Badenian.
Paratypes. 2 otoliths: NMNHU-P PI 2705, 1 specimen, Staryi Zavod; SMF PO 101.350, 1 specimen, Staryi Zavod.
Additional material. 1 otolith, NMNHU-P 2706, Staryi Zavod.
Etymology. From tectus (Latin) = tectiform, roof-shaped, referring to the shape of the dorsal rim resembling a roof.
Diagnosis. OL:OH = 0.78-0.88. Dorsal rim roof-shaped with slightly protruding, similarly high positioned pre- and postdorsal projections. Inner face completely flat. Sulcus small, oval in shape, without subcaudal iugum.
Description. Small, moderately robust otoliths up to 0.9 mm in length (holotype). OL:OH = 0.78-0.88; OH:OT = 3.0. The dorsal rim is high but relatively little bent, highest at its middle and with slightly protruding pre- and postdorsal projections. Anterior and posterior rims are vertical with broad, shallow concavities below pre- and postdorsal projections giving the dorsal rim the characteristic roof shape. The ventral rim is slightly bent, horizontal. All rims are sharp and smooth. The inner face is completely flat with a centrally positioned, small, deepened, oval, unstructured sulcus without the subcaudal iugum. OL:SuL = 2.0-2.3; sulcus inclination angle 13-16°. Dorsal depression is wide, with indistinct margins; the ventral furrow is relatively distinct leading up to the level of the sulcus. The outer face is distinctly convex and smooth.
Discussion. Microdesmid otoliths are small and rare in the fossil record: Microdesmus paratethycus Schwarzhans, 2017 from the late Badenian of Bulgaria and Paroxymetopon alienus Bratishko and Schwarzhans, 2023 from the Bessarabian of Ukraine. For comparison with extant otoliths of Ptereleotris, reference is made to Bratishko et al. (2023).
Order GASTEROSTEIFORMES Goodrich, 1909
Family GASTEROSTEIDAE Bonaparte, 1831
Gasterosteidae indet.
Figure 6A-B
Material. 2 otoliths, NMNHU-P PI 2707, Staryi Zavod.
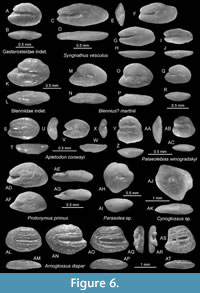 Discussion. Two small otoliths just below 1 mm in length are interpreted to represent an unspecified gasterosteid species indicating a rare freshwater influence in the Medobory backreef. These two small otoliths ressemble Pungitius kornyensis (Schubert, 1902) as figured in Schwarzhans (2017), which is known from the Karaganian of Bulgaria, in the overall otolith shape and the narrow slightly undulating sulcus with its small ostium. The otoliths are characterised by a regular oval outline with a moderately protruding rostrum and a narrow, slightly flexed sulcus with long cauda and short, minimally widened ostium.
Discussion. Two small otoliths just below 1 mm in length are interpreted to represent an unspecified gasterosteid species indicating a rare freshwater influence in the Medobory backreef. These two small otoliths ressemble Pungitius kornyensis (Schubert, 1902) as figured in Schwarzhans (2017), which is known from the Karaganian of Bulgaria, in the overall otolith shape and the narrow slightly undulating sulcus with its small ostium. The otoliths are characterised by a regular oval outline with a moderately protruding rostrum and a narrow, slightly flexed sulcus with long cauda and short, minimally widened ostium.
Order Syngnathiformes Rafinesque, 1810
Family Syngnathidae Rafinesque, 1810
Genus SYNGNATHUS Linnaeus, 1758
Syngnathus vesculus n. sp.
Figure 6C-J
zoobank.org/D66C21A3-
Holotype. NMNHU-P PI 2708 (Figure 6C-E), Staryi Zavod, western Ukraine, late Badenian.
Paratypes. 6 otoliths, SMF PO 101.351, same data as holotype.
Etymology. From vesculus (Latin) = slim, referring to the slender shape of the otolith and the sulcus.
Diagnosis. OL:OH = 1.85-2.25; OL:SuL = 1.5-1.6. Rostrum and antirostrum short, equally long; excisura small. Sulcus narrow, slightly oscillating; ostium deeper than cauda; OsL:CaL = 1.65-2.0.
Description. Small, elongate, and relatively thin otoliths up to 1.45 mm in length (holotype). OL:OH = 1.85-2.25; OH:OT = 1.9-2.3. The anterior tip of the otolith is blunt, with a short, equally long rostrum and antirostrum and a small excisura in-between. Dorsal and ventral rims are shallow, highest at their middle, symmetrical. The posterior rim is rounded or blunt. All rims are smooth. The inner face is nearly flat, smooth, with a slightly supramedian positioned, rather short and narrow sulcus; OL:SuL = 1.5-1.6. The sulcus is slightly oscillating, slightly upward shifted in the rear part of the ostium. The latter is distinctly deeper and longer than a shallow cauda and of equal width; OsL:CaL = 1.65-2.0. The dorsal depression and ventral furrow are not discernible. The outer face is moderately to distinctly convex, smooth.
Discussion. In contrast to relatively commonly found articulated fossil skeletons of syngnathids (see for instance Bannikov, 2010), there are no fossil records of syngnathid otoliths. This discrepancy probably has to do with the robust skeleton of these fishes enhancing their fossilization in contrast to their tiny otoliths, which cannot be expected in the typically used 1 mm mesh frame. Therefore, S. vesculus is the first fossil otolith-based record in the group. All otoliths here recorded have been found in mesh frames smaller than 1 mm, that is, 0.5 mm, and most of the specimens are indeed shorter than 1 mm in length except for the holotype. Extant otoliths of syngnathids have been figured in Lombarte et al. (2006) and Nolf (2013). Syngnathus vesculus resembles otoliths of the extant S. acus Linnaeus, 1758 (see Lombarte et al., 2006 and Nolf, 2013 for figures) but differs in the larger size it can attain and the smooth otolith rims.
Order Blenniiformes Bleeker, 1859
Family Blenniidae Rafinesque, 1810
Genus BLENNIUS Linnaeus, 1758
Blennius? martinii Reichenbacher, Filipescu and Miclea, 2019
Figure 6M-R
2019 Blennius? martinii; Reichenbacher et al., fig. 3I-K.
Material. 3 otoliths, NMNHU-P PI 2711, Staryi Zavod.
Discussion. Three small specimens, the largest (Figure 6M-N) being 0.9 mm in length represent typical blenniid otoliths characterised by the triangular outline, the short sulcus with the anteriorly widened open ostium and the very small, slightly downturned cauda that is separated from the ostium by a distinct narrowing at the collum. They represent Blennius? martinii described by Reichenbacher et al. (2019) from the early Sarmatian of Romania.
Blenniidae indet.
Figure 6K-L
Material. 1 otolith, NMNHU-P PI 2712, Staryi Zavod.
Discussion. A single otolith of 1.15 mm in length resembling Blennius? martinii but differing in lacking the constriction of the sulcus at the collum and the more gently curved ventral rim.
Order GOBIESOCIFORMES Berg, 1937
Family GOBIESOCIDAE Bleeker, 1859
Genus APLETODON Briggs, 1955
Apletodon conwayi n. sp.
Figure 6S-X
zoobank.org/252354B5-
Holotype. NMNHU-P PI 2713 (Figure 6S-U), Mlyntsi, western Ukraine, late Badenian.
Paratype. SMF PO 101.352, Staryi Zavod.
Etymology. Named in honour of Kevin Conway (College Station, Texas, USA) for his many contributions to the understanding of gobiesocid fishes.
Diagnosis. Otolith not exceeding 0.7 mm in length; OL:OH = 1.40-1.45. Rostrum slightly longer than antirostrum. OL:SuL = 1.5; OsL:CaL = 0.95-1.25.
Description. Tiny otoliths maximally reaching 0.7 mm in length (holotype). OL:OH = 1.40-1.45; OH:OT = 2.2-2.3. The dorsal and ventral rims are regularly curved, continuous with a well-rounded posterior rim. The rostrum is distinct with a rounded tip, 8-15% OL, longer than antirostrum; excisura is distinct, variably deep, and sharp. All rims are smooth and thick. The inner face is completely flat in ventral view (Figure 6T, 6W), concave in anterior view because of a broad, depressed sulcus (Figure 6U, 6X). The latter is deepened, anteriorly open, relatively short (OL:SuL = 1.5) divided into about equally long and wide ostium and cauda (OsL:CaL = 0.95-1.25). The sulcus is marked by a narrow, crest-like, and distinct crista superior and inferior along sulcus. Dorsal and ventral depressions are fading away from the sulcus. The outer face is convex and smooth.
Discussion. Gobiesocid fishes or clingfishes are small, cryptic fishes clinging to rocks or sea grass in shallow and littoral water with wave activity. Their otoliths are extremely small, regularly smaller than 1 mm in length and perhaps therefore not described from the fossil record except for a find made in situ (Schwarzhans et al., 2017b) in specimens tentatively attributed to the genus Apletodon from the Sarmatian of the Central Paratethys. These in situ otoliths were up to 0.5 mm in length. Gobiesocid otoliths are characterised by the broad and deepened sulcus with equally wide ostium and cauda and distinct, crest-like cristae surrounding the sulcus (for extant gobiesocid otoliths see Charmpila et al., 2021). Apletodon conwayi represents the first otolith-based species in this group. It differs from Apletodon? sp. found in situ in the Sarmatian (Schwarzhans et al., 2017b) in the more elongate shape (OL:OH = 1.40-1.45 vs. 1.15-1.30) and the relatively wide sulcus.
Order CYPRINODONTIFORMES Berg, 1940
Family CYPRINODONTIDAE Gill, 1865
Genus PALAEOLEBIAS Reichenbacher and Weidmann, 1992
Palaeolebias winogradskyi n. sp.
Figure 6Y-AC
zoobank.org/8FC7AD5B-
Holotype. NMNHU-P PI 2715 (Figure 6Y-AA), Lisohirka, western Ukraine, late Badenian.
Paratypes. 2 otoliths, SMF PO 101.353, same data as holotype.
Etymology. Named in honour of Serhii Winogradsky, a famous Ukrainian microbiologist, discoverer of the process of chemosynthesis, the founder of soil microbiology and ecology, an employee of the Pasteur Institute (France), a foreign member of the Royal Society of London and the last owner of Horodok (Thorton, 1953; Gumeniuk and Kryvyi, 2016; Kryvyi et al., 2022).
Diagnosis. OL:OH = 0.95-1.05; OH:OT = 2.7-2.8. Inner face relatively flat and outer face distinctly convex. Dorsal field wide; dorsal rim rounded. Rostrum short, blunt. Sulcus slightly supramedian; cauda widened with rounded tip.
Description. Small, compact, and high bodied otoliths reaching about 0.6 mm in length (holotype); OL:OH = 0.95-1.05; OH:OT = 2.7-2.8. The dorsal rim is broadly rounded, wide, and slightly crenulated. The ventral rim is moderately deep, regularly curved. The rostrum is short, blunt, and rounded, 7-10% OL; antirostrum and excisura are minute. The posterior tip forming an obtuse inferior angle at the junction of inclined posterior rim and the ventral rim. The inner face is almost flat to slightly convex, with a slightly supramedian sulcus. The latter is moderately deep, relatively short (OL:SuL = 1.3-1.4), divided by a broad and indistinct ventral collum in nearly equally long ostium and cauda. The cauda is slightly widened, with a rounded tip. The dorsal field is wide, with a wide depression. The dorsal depression is well marked toward the sulcus by a distinct crista superior. There is no ventral furrow. The outer face is convex and smooth.
Discussion. Cyprinodontid otoliths are common in the Oligocene and Miocene freshwater sediments of Europe (e.g., Reichenbacher, 1996; Reichenbacher and Weidmann, 1992; Reichenbacher and Prieto, 2006; Reichenbacher et al., 2007, 2019; Reichenbacher and Kowalke, 2009), and extant cyprinodontid otoliths from Europe and the Middle East have also been extensively studied (e.g., Reichenbacher et al., 2007, 2009). A few fossil otoliths have been placed in the extant genus Aphanius Nardo, 1827, and several extinct genera (Prolebias Sauvage, 1874; Palaeolebias Reichenbacher and Weidmann, 1992; Aphanolebias Reichenbacher and Gaudant, 2003). The otoliths from the Medobory backreef resemble most those of the fossil genus Palaeolebias because of its high body, the relatively regularly ventral rim, the short rostrum and posterior tip, and the shape of the sulcus. In fact, Palaeolebias winogradskyi represents the latest record of this genus, which otherwise is primarily known from the Late Oligocene and Early Miocene (Reichenbacher, 1996). Although reatively small it is nevertheless within the range of sizes reported from fossil Prolebias and Palaeolebias species (Reichenbacher and Weidmann, 1992; Reichenbacher, 1993) and show well-developed diagnostic features. Palaeolebias winogradskyi is distinguished from its congeners by the wide dorsal field, regularly rounded dorsal rim, and the rounded caudal tip. The species furthermore represents one of the rare indicators of fresh or brackish water faunal influx.
Order PLEURONECTIFORMES Bleeker, 1859
Family BOTHIDAE Regan, 1910
Genus ARNOGLOSSUS Bleeker, 1862
Arnoglossus dispar n. sp.
Figure 6AL-AT
zoobank.org/A809A94B-
Holotype. NMNHU-P PI 2719 (Figure 6AO-AQ), Staryi Zavod, western Ukraine, late Badenian.
Paratypes. 2 specimens, SMF PO 101.354, Staryi Zavod; 1 specimen, SMF PO 101.355, Skala.
Additional material. 6 otoliths: 2 specimens, NMNHU-P PI 2709, Novyi Pliazh; 3 specimens, NMNHU-P 2720, Staryi Zavod; 1 specimen, Skala.
Etymology. From dispar (Latin) = dissimilar, referring to the large degree of side dimorphism observed in the otoliths of this species.
Diagnosis (left otolith). OL:OH = 1.35-1.45. Dorsal, posterior and ventral rims regularly curved and continuous. Rostrum short, rounded, 5-11% OL. Inner face more convex than outer face. Ostium anteriorly open. OsL:CaL = 1.1-1.3.
Description (left otolith). Small otoliths with a relatively regular oval shape and robust up to a size of 2 mm in length (holotype 1.85 mm). OL:OH = 1.35-1.45; OH:OT = 2.2-2.4. Dorsal, posterior, and ventral rims are regularly curved and continuous. The rostrum is short and rounded, 5-11% OL. Antirostrum and excisura are minute or absent. All the rims are smooth or slightly undulating. The inner face is distinctly convex, with an axially positioned, long, narrow, and deep sulcus. OL:SuL = 1.35-1.45. The ostium is open anteriorly; ostium and cauda are poorly distinguished, the ostium only slightly longer than cauda (OsL:CaL = 1.1-1.3). Circumsulcal depression is narrow and deep, close to the sulcus and separated from it by a sharp, crest-like cristae. The outer face is flat, smooth.
Side dimorphism. Only a single otolith is available of A. dispar from the right side, and it differs in several aspects from those of the left side. The ratio OL:OH is 1.1 (vs. 1.35-1.45). The dorsal rim shows clear pre- and postdorsal angles; the posterior is more slanting than rounded. The sulcus is differentiated in a shorter ostium and longer cauda by an elevated collum (OsL:CaL = 0.9).
Discussion. The bothid otoliths show that the fishes of the genus Arnoglossus underwent a rapid endemic evolution in the Paratethys, particularly in its eastern part, during Badenian and Sarmatian s.l. (Bratishko et al., 2015, 2023; Schwarzhans et al., 2017c). Arnoglossus dispar differs from A. bassanianus (Kramberger, 1883), which has been found with otoliths in situ (Schwarzhans et al., 2017c), in the anteriorly open sulcus and the inner face being more convex than the outer face (vs. outer face being more convex than inner face). Both species occurred in parallel during the late Badenian (A. bassanianus also in the Sarmatian s.s.) whereby A. dispar appears to have been adapted to reefal environments like the Medobory backreef whereas A. bassanianus occurred in clastic sublittoral settings. The later species A. kerichensis Bratishko and Schwarzhans, 2023 and A. scitulus Bratishko and Schwarzhans, 2023 from the Bessarabian of the Eastern Paratethys differ in the longer rostrum, less regularly curved rims and the ostium being distinctly longer than the cauda resulting in a larger ratio OsL:CaL > 1.35, mostly > 1.5 (vs. 0.9-1.3).
Family SOLEIDAE Bonaparte, 1833
Genus PARASOLEA Schwarzhans, Carnevale, Japundžić and Bradić-Milinović, 2017
Parasolea sp.
Figure 6AH-AI
Material. 7 otoliths, NMNHU-P PI 2722, Lisohirka.
Discussion. A few small, high bodied otoliths < 1 mm in length are characterised by a very small sulcus. They likely represent juvenile otoliths of the extinct Paratethyan genus Parasolea.
Family CYNOGLOSSIDAE Jordan and Goss, 1889
Genus Cynoglossus Hamilton, 1822
Cynoglossus sp.
Figure 6AJ-AK
Material. 1 otolith, NMNHU-P PI 2724, Novyi Pliazh.
Discussion. A single otolith of 1.55 mm in length is characterised by a high, pentagonal otolith shape with a deep ventral rim and strong midventral angle, concave postventral section and expanded predorsal lobe. Its inner face is mildly convex and smooth with a poorly discernible sulcus that resembles the “hammer-shaped” outline typical for cynoglossid otoliths, although its termination is relatively rounded instead of shaped like a hammerhead.
Order LABRIFORMES Bleeker, 1859
Family LABRIDAE Cuvier, 1816
Genus CORIS Lacépède, 1801
Coris medoboryensis Schwarzhans, Klots and Kovalchuk, 2022
Figure 7A-H
2022 Coris medoboryensis Schwarzhans, Klots and Kovalchuk; Schwarzhans et al., fig. 8c-d.
 Material. 37 otoliths: 3 specimens, NMNHU-P PI 2725, Lisohirka; 17 specimens, NMNHU-P PI 2726, Mlyntsi; 4 specimens, NMNHU-P PI 2727, Novyi Pliazh; 12 specimens, NMNHU-P PI 2728, Staryi Zavod; 1 specimen, NMNHU-P PI 2729, Skala.
Material. 37 otoliths: 3 specimens, NMNHU-P PI 2725, Lisohirka; 17 specimens, NMNHU-P PI 2726, Mlyntsi; 4 specimens, NMNHU-P PI 2727, Novyi Pliazh; 12 specimens, NMNHU-P PI 2728, Staryi Zavod; 1 specimen, NMNHU-P PI 2729, Skala.
Discussion. Coris medoboryensis was established on two specimens. Many new, additional specimens now give a complete ontogenetic sequence that shows that morphological maturity is reached at a size between 1.5 mm (Figure 7E-F) and 2.0 mm in length (Figure 7B-D).
Genus THALASSOMA Swainson, 1839
Thalassoma vernyhorovae (Schwarzhans, Klots and Kovalchuk, 2022)
Figure 7I-P
2022 Blennius vernyhorovae Schwarzhans, Klots and Kovalchuk; Schwarzhans et al., fig. 8ca-b.
Material. 32 otoliths: 3 specimens, NMNHU-P PI 2730, Lisohirka; 9 specimens, NMNHU-P PI 2731, Mlyntsi; 20 specimens, NMNHU-P PI 2732, Staryi Zavod.
Discussion. Blennius vernyhorovae was established on two specimens. Many new specimens now available show that the species, too, represents a labrid. There are relatively few otoliths in labrids known today with such a deep ventral rim, that is, in the genera Halichoeres Rüppell, 1835, Thalassoma Swainson, 1839, and Xyrichthys Cuvier, 1814. We consider Thalassoma as having the most similar otolith morphology (see Smale et al., 1995; Rivaton and Bourret, 1999; Lin and Chang, 2012, for extant labrid otoliths). Available ontogenetic sequence demonstrates that T. vernyhorovae does not reach the size of Coris medoboryensis (1.7 vs. 2.5 mm in length), and that morphological maturity is reached in T. vernyhorovae between 1.2 mm (Figure 7N) and 1.4 mm in length (Figure 7L-M).
Order SCORPAENIFORMES Garman, 1899
Family LIPARIDAE Gill, 1861
Liparidae indet.
Figure 7W-X
Material. 1 otolith, NMNHU-P 2737, Staryi Zavod.
Discussion. A single, small otolith of 0.85 mm in length resembles extant otoliths of the genus Liparis, and the otolith is therefore tentatively placed in the family Liparidae.
Order PERCIFORMES Bleeker, 1859
Family PRIACANTHIDAE Günther, 1859
Genus PRISTIGENYS Agassiz, 1835
Pristigenys schiecki Schwarzhans, 2010
Figure 7AB-AD
2010 Pristigenys schiecki; Schwarzhans, pl. 75, figs. 10-13.
Material. 15 otoliths: 5 specimens, NMNHU-P PI 2745, Mlyntsi; 4 specimens, NMNHU-P PI 2746, Novyi Pliazh; 4 specimens, NMNHU-P PI 2747, Staryi Zavod; 2 specimens, NMNHU-P PI 2748, Skala.
Discussion. Pristigenys schiecki was so far only known from the Hemmoorian and Reinbeckian of the North Sea Basin, the stratigraphic equivalents of the Ottnangian to Badenian of the Paratethys (Schwarzhans, 2010). Its relative common appearance in the late Badenian of the Medobory backreef environment indicates that the species was more widely distributed than previously perceived. It is not known from the time equivalent strata of the Mediterranean or the NE Atlantic. The cause for the apparent patchy distribution pattern of the species is currently elusive.
Family CEPOLIDAE Rafinesque, 1810
Genus OWSTONIA Tanaka, 1908
Owstonia sp.
Figure 7Y
Material. 1 otolith, NMNHU-P PI 2750, Staryi Zavod.
Discussion. The identification of the cepolid otoliths in the Neogene of Europe is in a catastrophic estate. A review of these otoliths is long overdue but is beyond the scope of this study. The otolith figured here represents the genus Owstonia characterised by compressed, rounded hexagonal otolith shape and a small, reduced ostial colliculum whereby the ostium does not open to the anterior rim of the otolith. This pattern distinguishes otoliths of Owstonia from those of the genus Cepola.
Order Spariformes Bleeker, 1876
Family Sparidae Rafinesque, 1810
Genus DIPLODUS Rafinesque, 1810
Diplodus karrerae Nolf and Steurbaut, 1979
Figure 7AG-AL
1979 Diplodus karrerae; Nolf and Steurbaut, pl. 3, figs. 19-21.
2010 Diplodus karrerae Nolf and Steurbaut, 1979; Schwarzhans, pl. 90, figs. 3-4.
2013 Diplodus karrerae Nolf and Steurbaut, 1979; Schultz, pl. 86, fig. 9a-b.
2014 Diplodus karrerae Nolf and Steurbaut, 1979; Schwarzhans, pl. 6, fig. 4.
2015 Diplodus karrerae Nolf and Steurbaut, 1979; Lin et al., fig. 5.21.
2022 Diplodus karrerae Nolf and Steurbaut, 1979; Brzobohatý et al., pl. 2, fig. V-W.
Material. 20 otoliths: 1 specimen, NMNHU-P PI 2752, Shydlivshchyna; 11 specimens, NMNHU-P PI 2753, Novyi Pliazh; 5 specimens, NMNHU-P PI 2754, Staryi Zavod; 3 specimens, NMNHU-P PI 2755, Skala.
Discussion. The otoliths of Diplodus karrerae are widely distributed in the Middle Miocene of Europe but rarely common. The occurrence of this species in the late Badenian of the Central Paratethys probably indicates a re-immigration from the Mediterranean of this stenohaline marine species. It represents the latest record in the Paratethys.
Part 2. Fish Bones (by Dubikovska and Kovalchuk)
Non-otolith fish remains in the material from the localities studied are considerably less numerous compared to otoliths obtained from the same layers. Almost all of them are represented by isolated teeth, accompanied by a single jaw fragment and one dermal denticle from Lisohirka as well as one vertebra from Skala. Non-otolith remains are also less diverse taxonomically.
Dasyatidae indet. (order Myliobatiformes, class Chondrichthyes) - 1 tooth, NMNHU-P 2771, Shydlivshchyna. The tooth crown bears transversal crest separating the labial and lingual surfaces. The outline of the crown is triangular. Its lingual surface is almost vertical in profile view and concave just below the transversal crest. The labial surface is smooth and curved, with a rounded labial edge. There is a small cusp at the top of the crown, which is crossed in the middle by a small furrow. The hoof-shaped root is shifted posteriorly to the lingual surface, its lobes are well separated, slightly protruding, and triangular in vertical projection. Considering the morphology of the specimen (cf. Cappetta, 2012), it belongs to a female individual of the family Dasyatidae.
Labrodon sp. (family Labridae, order Labriformes, class Actinopterygii) - 1 tooth, NMNHU-P PI 2761, Skala. A small molariform tooth has a triangular outline. Its occlusal surface bears a low central papilla, which is clearly visible in a profile view. This specimen has been identified as Labrodon sp. since it is morphologically like those described and figured in Schultz (1979), Szabó and Kocsis (2020), Szabó et al. (2021) from the Central Paratethys, although we do not exclude the possibility of its belonging to the genus Coris (Carnevale, 2015). The latter is represented in the same layer by an otolith of C. medoboryensis (Schwarzhans et al., 2022).
Trigonodon jugleri (Münster, 1846) (family Labridae, order Labriformes, class Actinopterygii) - 2 teeth: NMNHU-P PI 2762, Skala; NMNHU-P PI 2763, Staryi Zavod. Both anterior teeth are rounded triangle (chisel-like) in shape and share incisiform morphology. The specimens are flattened labiolingually and have an even cutting edge. Their labial surface is vertical and slightly concave, lacking ornamentation (as is the smooth lingual surface). The teeth morphologically resemble those in representatives of the genus Trigonodon, and most probably belong to Trigonodon jugleri (Münster, 1846). This species first appeared in the Early Miocene of Italy and Austria (Schultz and Bellwood, 2004; Marsili et al., 2007), and it was widely distributed within the Central Paratethys during the Middle Miocene (Schultz, 1998; Schultz et al., 2010; Schultz, 2013; Bellwood et al., 2019; Szabó and Kocsis, 2020; Szabó et al., 2021).
Diplodus sp. (family Sparidae, order Spariformes, class Actinopterygii) - 1 tooth, NMNHU-P PI 2764, Lisohirka. The anterior tooth has an incisiform morphology. Its crown is quadrangular, flattened to slightly convex labially, concave lingually, and has a smooth surface. The left cutting edge of the tooth is higher than the right one in lingual view. The root was broken just at the base.
Pshekharus yesinorum Bannikov and Kotlyar, 2015 (family Sparidae, order Spariformes, class Actinopterygii) - 1 tooth, NMNHU-P PI 2765, Skala. A single tooth has a central papilla and probably represents an intermediate morphotype. Its crown is conical, as high as wide, what is a characteristic feature of the genus Pshekharus (see Bannikov and Kotlyar, 2015 for details).
Sparidae indet. (order Spariformes, class Actinopterygii) - 1 jaw fragment, NMNHU-P PI 2766, Lisohirka. 47 teeth: 8 specimens, NMNHU-P PI 2767, Lisohirka; 1 specimen, NMNHU-P PI 2768, Shydlivshchyna; 31 specimens, NMNHU-P PI 2769, Staryi Zavod; 7 specimens, NMNHU-P PI 2770, Skala. There is a rounded opening on the labial side of the jaw. The latter bears one conical (intermediate) tooth and numerous hemispherical (molariform) teeth. Anterior and intermediate teeth are conical, higher than wide, circular in cross-section, and have a rounded pulp cavity. Some of them have a cylindro-conical crown. The apical and basal parts of the latter are separated by a furrow. Molariform teeth with hemispherical or oval crowns are of different sizes.
In addition to the specimens mentioned above, there are some fossils whose precise taxonomic identification is impossible due to their poor preservation or too general morphology (i.e., lacking reliable diagnostic characters). Those are one dermal denticle of a shark (Selachimorpha indet.) NMNHU-P PI 2773, one ray tooth (Batoidea indet.) NMNHU-P PI 2774 from Lisohirka, a single anterior conical tooth of actinopterygian fish NMNHU-P PI 2772 from Shydlivshchyna, as well as one actinopterygian vertebra NMNHU-P PI 2775 from Skala.
Part 3. Other Biota (by Ryabokon, Kovalenko, and Klots)
In addition to the otoliths and other fish skeletal elements described above, rich foraminiferal and ostracod assemblages have also been obtained from the respective rock samples.
Foraminiferan fauna was studied to clarify the stratigraphic position of the samples obtained from the new localities. The list of identified species is presented in Table 2 along with data on the distribution of particular taxa in the Badenian of the Central Paratethys (Cicha et al., 1998; Garecka and Olszewska, 2011; Peryt et al., 2021), Konkian of the Eastern Paratethys (Krasheninnikov et al., 2003; Vernyhorova, 2018; Vernyhorova et al., 2023), and Middle Miocene of the Volyn-Podolia region (Kudrin, 1966; Didkovsky and Satanovskaya, 1970; Goretsky and Didkovsky, 1975; Andreeva-Grigorovich et al., 1996; Gedl and Peryt, 2011; Peryt et al., 2021; Schwarzhans et al., 2022). The studied foraminifers are similar in composition and consist exclusively of benthic forms, including various and numerous miliolids (Quinqueloculina, Triloculina, Pseudotriloculina, Pyrgo, and others) and representatives of the genera Elphidium, Ammonia, Lobatula, Heterolepa, Cibicidoides, Asterigerinata, Globulina, Guttulina, frequent Cancris, Melonis, Reussella, Porosononion, also present Borelis and Sphaerogipsina. All of them are marine species including those strictly associated with reefal environments while brackish taxa are absent. In addition, several permanently motile foraminifers recognised in the studied samples (in particular, Elphidium crispum, E. macellum, Globulina gibba, Nonion depressulum, Reussella spinulosa, Spiroloculina canaliculata, and Triloculina gibba) and representatives of the genera Ammonia, Anomalinoides, Cibicides, Discorbis, Pyrgo, Quinqueloculina, and Rosalina are commonly associated with seagrass or seaweeds (Langer, 1993; Moissette et al., 2007).
The studied foraminifera are characteristic of the Badenian (Middle Miocene, early Serravallian) of the Central Paratethys (Cicha et al., 1998). A specific feature of these assemblages is the presence of some endemic taxa distributed in the Volyn-Podolia region and the Carpathian Foredeep: Nodobaculariella podolica, Sigmomorphina karpatica, Pseudopatellinoides primus, Cibicides menneri, Nonionella ventragranosa, Ammonia galiciana, A. pseudobeccarrii, Elphidium stellans, and E. ukrainicum (Didkovsky and Satanovskaya, 1970; Goretsky and Didkovsky, 1975).
Nearly half of the foraminifera species identified in the studied samples are also common in the Konkian of the Eastern Paratethys, and two-thirds of them (Table 2) are known from the Ternopil and Vyshgorod beds (=upper Badenian) of the Volyn-Podolia region (Kudrin, 1966; Didkovsky and Satanovskaya, 1970; Goretsky and Didkovsky, 1975; Schwarzhans et al., 2022). We therefore conclude that the studied samples date the late Badenian.
Ostracods were recognised in rock samples obtained from Lisohirka, Skala, and Staryi Zavod. The general species list includes 20 taxa (Table 3), of which the most common (i.e., present in the material from all the studied localities) are Aurila cicatricosa and A. novata; less frequent are Cyclocypris regularis, Aurila convexa, Loxoconcha spongiosa, Cytheridea muelleri, and Cnestocythere truncata, while all other species each come from a single locality (Table 3).
Most ostracods from the Middle Miocene of the Medobory backreef preferred normal marine conditions. The presence of Cytheridea muelleri and Leptocythere ex gr. canaliculata in the material from Staryi Zavod and Lisohirka indicates higher than normal salinity (ca. 15-17‰) and warm waters. Only two species (Cyclocypris regularis from Lisohirka and Skala and Cyclocypris laevis from Skala) are indicators of a limited freshwater influx. The presence of five species of the genus Aurila, two species of the genus Loxoconcha, and two species of the genus Xestoleberis in the material from Lisohirka, Skala, and Staryi Zavod (Table 2), in addition to epiphytic foraminifers mentioned above, suggest a seagrass environment (Pisera, 1985; Aiello and Szczechura, 2004; Moissette et al., 2007; Cornée et al., 2009; Forsey, 2016). The studied ostracod assemblages comprise a number of species distributed in the Central Paratethys during the Early Miocene (Kollmann, 1971; Zorn, 1998, 2003; Tunoğlu and Bilen, 2001), Badenian (Paruch-Kulczycka, 1992; Szuromi-Korecz and Szegő, 2001; Aiello and Szczechura, 2004; Zorn, 2004; Szczechura, 2006), Sarmatian (Tóth, 2008), and Pannonian (Stancheva, 1962, 1963), although they are more similar taxonomically to those from the Konkian of the Eastern Paratethys (Schneider, 1953, 1959; Didkovsky, 1959, 1964; Ljuljev, 1967, 1969; Bondar, 2006; Kovalenko, 2013).
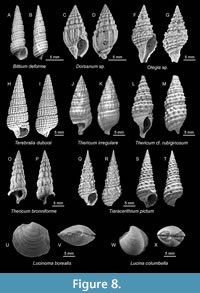 Gastropods are dominated by turitellid forms throughout the sampled localities, particularly species of the genera Terebralia, Thericium, and Tiaracerithium (Figure 8). Terebralia and Tiaracerithium are typical for mudflats (Harzhauser et al., 2023); the other observed genera are widely distributed in sublittoral environments. Lucinid bivalves of the genera Lucina, Lucinoma (Figure 8), and Loripes are common in seagrass environments (e.g., van der Heide et al., 2012; Stanley, 2014) and are common in the material from Novyi Pliazh, Skala, and Horodok being represented by both large (more than 5 mm) and small shells. These molluscs were also observed, albeit rarely, in all other studied localities (Kozatskyi Yar, Lisohirka, Mlyntsi, Shydlivshchyna, and Staryi Zavod). Their distributon and abundance are not entirely congruent with those of foraminifera, ostracods, and fishes, which, however, in combination are more indicative in our opinion than the distribution pattern of the Lucinidae in the Medobory backreef environments.
Gastropods are dominated by turitellid forms throughout the sampled localities, particularly species of the genera Terebralia, Thericium, and Tiaracerithium (Figure 8). Terebralia and Tiaracerithium are typical for mudflats (Harzhauser et al., 2023); the other observed genera are widely distributed in sublittoral environments. Lucinid bivalves of the genera Lucina, Lucinoma (Figure 8), and Loripes are common in seagrass environments (e.g., van der Heide et al., 2012; Stanley, 2014) and are common in the material from Novyi Pliazh, Skala, and Horodok being represented by both large (more than 5 mm) and small shells. These molluscs were also observed, albeit rarely, in all other studied localities (Kozatskyi Yar, Lisohirka, Mlyntsi, Shydlivshchyna, and Staryi Zavod). Their distributon and abundance are not entirely congruent with those of foraminifera, ostracods, and fishes, which, however, in combination are more indicative in our opinion than the distribution pattern of the Lucinidae in the Medobory backreef environments.
FISH LIFE ON THE MEDOBORY BARRIER REEF
Setting of the Scene (Figure 9-Figure 10)
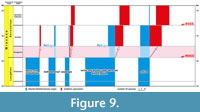 The lower Badenian (sensu Harzhauser et al., 2020; Šegvić et al., 2023) / Tarkhanian (=Langhian) was the time when the Paratethys was broadly re-connected to the World Ocean, that is at least to the Mediterranean in the northwest (Rögl, 1998; Popov et al., 2004; Sant et al., 2019, 2020). This event was related to the Miocene climate optimum (MCO) (Miller et al., 2020) and led to a harmonisation of the biota in the Paratethys with those of the neighbouring seas, called the “early Badenian-build-up-event” (EBBE) in Harzhauser and Piller (2007). The fish fauna is exceptionally well known in the Central Paratethys by means of otoliths (e.g., Radwańska, 1992, and articles cited therein) and exhibits a large degree of congruence with the otolith-based fish fauna in the Mediterranean (Nolf and Brzobohatý, 2004; Schwarzhans and Carnevale, 2024). Time-equivalent otolith associations from the Indian Ocean are not known except for an upper Burdigalian fauna from southern India (Carolin et al., 2023). The lower Badenian otolith association in the Central Paratethys contained an estimated 200+ species (based on cited literature and ongoing research) and was highly diversified, including near-shore, neritic-shelf, open marine mesopelagic and bathybenthic fishes (Figure 9). No otolith associations are known from the Leitha Formation coralline algal reef environment in the Vienna Basin, but skeletal records add to the faunal composition (e.g., Schultz, 2013; Carnevale and Harzhauser, 2013; Carnevale and Collette, 2014; Carnevale, 2015).
The lower Badenian (sensu Harzhauser et al., 2020; Šegvić et al., 2023) / Tarkhanian (=Langhian) was the time when the Paratethys was broadly re-connected to the World Ocean, that is at least to the Mediterranean in the northwest (Rögl, 1998; Popov et al., 2004; Sant et al., 2019, 2020). This event was related to the Miocene climate optimum (MCO) (Miller et al., 2020) and led to a harmonisation of the biota in the Paratethys with those of the neighbouring seas, called the “early Badenian-build-up-event” (EBBE) in Harzhauser and Piller (2007). The fish fauna is exceptionally well known in the Central Paratethys by means of otoliths (e.g., Radwańska, 1992, and articles cited therein) and exhibits a large degree of congruence with the otolith-based fish fauna in the Mediterranean (Nolf and Brzobohatý, 2004; Schwarzhans and Carnevale, 2024). Time-equivalent otolith associations from the Indian Ocean are not known except for an upper Burdigalian fauna from southern India (Carolin et al., 2023). The lower Badenian otolith association in the Central Paratethys contained an estimated 200+ species (based on cited literature and ongoing research) and was highly diversified, including near-shore, neritic-shelf, open marine mesopelagic and bathybenthic fishes (Figure 9). No otolith associations are known from the Leitha Formation coralline algal reef environment in the Vienna Basin, but skeletal records add to the faunal composition (e.g., Schultz, 2013; Carnevale and Harzhauser, 2013; Carnevale and Collette, 2014; Carnevale, 2015).
During the Middle Miocene climate transition (MMCT) and a global cooling pulse at the beginning of the Serravallian (Palcu et al., 2017), the sea level fell (Miller et al., 2020), and the Paratethys became separated from the World Ocean again and divided into the Central and the Eastern Paratethys (Palcu et al., 2017, 2019). This event caused the evaporitic crisis of the middle Badenian in the Central Paratethys and the Karaganian crisis in the Eastern Paratethys, both of which had a severe impact on the Paratethyan biota. The Karaganian crisis in the Eastern Paratethys is thought to have been caused by encroaching surface freshwater influx from the north that led to brackish near-surface waters and strongly reduced oxygenation at depth due to reduced circulation in the water body (Mikerina and Pinchuk, 2014). These changes in turn led to the extinction of deep marine fishes and probably other stenohaline marine fishes in the Eastern Paratethys during the Karaganian crisis (Bratishko et al., 2015, 2023; Schwarzhans et al., 2023). In large parts of the Central Paratethys except for the furthest western part, the coeval middle Badenian salinity crisis (Báldi et al., 2017) also had a severe but probably less dramatic effect on the fish fauna, in combination termed the mid-Badenian extinction event (MBEE) by Bratishko et al. (2023). It seems that endemic evolution took place in both basins (Baykina and Schwarzhans, 2017; Bratishko et al., 2023) (Figure 9). During a short period of the upper Badenian/Konkian (lower Serravallian), the Central and Eastern Paratethys became reconnected again and the Central Paratethys was still connected to the Mediterranean in the west (Bartol et al., 2014). Normal marine conditions returned throughout the Paratethys, and some marine biota probably remigrated into the Eastern Paratethys from the Central Paratethys or the Mediterranean.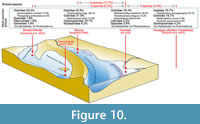 This event was apparently limited in scope. It included only few deepwater fishes (Figure 9) that are mainly found in the westernmost region of the Central Paratethys at Walbersdorf, Austria (ongoing research). There are also indications of a limited amount of meso-and epipelagic endemic speciation having occurred: Gadidae (Paratrisopterus) and Myctophidae (Diaphus) in the Carpathian Foredeep (Schwarzhans and Radwańska, 2022) (Figure 9-Figure 10). Otherwise, fish that have been able to adapt to the foregoing crises were also among those that succeeded most in the adaptation to the new and diverse environments that evolved in the Paratethys (Schwarzhans et al., 2022) (Figure 9). Continued forced endemic speciation transformed the Eastern Paratethys into an evolutionary hotspot during the Serravallian and early Tortonian, at least as far as bony fishes are concerned and despite a subsequent intermittent crisis at the transition of the Badenian to the Sarmatian (Badenian-Sarmatian extinction event = BSEE; Harzhauser and Piller, 2007) when all deepwater fishes finally disappeared from the entire Paratethys (Bratishko et al., 2023; Harzhauser et al., 2024).
This event was apparently limited in scope. It included only few deepwater fishes (Figure 9) that are mainly found in the westernmost region of the Central Paratethys at Walbersdorf, Austria (ongoing research). There are also indications of a limited amount of meso-and epipelagic endemic speciation having occurred: Gadidae (Paratrisopterus) and Myctophidae (Diaphus) in the Carpathian Foredeep (Schwarzhans and Radwańska, 2022) (Figure 9-Figure 10). Otherwise, fish that have been able to adapt to the foregoing crises were also among those that succeeded most in the adaptation to the new and diverse environments that evolved in the Paratethys (Schwarzhans et al., 2022) (Figure 9). Continued forced endemic speciation transformed the Eastern Paratethys into an evolutionary hotspot during the Serravallian and early Tortonian, at least as far as bony fishes are concerned and despite a subsequent intermittent crisis at the transition of the Badenian to the Sarmatian (Badenian-Sarmatian extinction event = BSEE; Harzhauser and Piller, 2007) when all deepwater fishes finally disappeared from the entire Paratethys (Bratishko et al., 2023; Harzhauser et al., 2024).
Marine Fish Life during the Late Badenian/Konkian of the Paratethys (Figure 10-Figure 11)
The basin configuration of the Paratethys became highly fragmented during the late Badenian (Kováč et al., 2007, 2017; Harzhauser et al., 2024), and the environmental settings may have varied considerably across the region. The fish fauna, which was relatively uniform across the Central Paratethys during the early Badenian, showed a much higher degree of disparity during the late Badenian. This observation of biogeographic fragmentation is congruent with the results of a recent study of molluscs from the Central Paratethys (Harzhauser et al., 2024). From a comparison of the late Badenian fish faunas of Borský Mikuláš in the Vienna Basin (Brzobohatý et al., 2022) with those of the deep Carpathian Foredeep at Gliwice, Poland (Śmigielska, 1966), with those of the Medobory backreef of Horodok on the flank of the Carpathian Foredeep (Schwarzhans et al., 2022, and this study), and with those of the Eastern Paratethys in Karaigaly, Kazakhstan (Bratishko et al., 2015), it is clear that the number of shared species is low from one region to the other (Figure 10). In the case of the faunal difference within the Carpathian Foredeep, it is probable that they have to do with the difference in environment, but the differences between the faunas of the Carpathian Foredeep in total, the Vienna Basin, and the Eastern Paratethys are probably geographically driven. Gobies are the dominant group in the shallow-water locations of the Vienna Basin and the Medobory backreef and the second most common group at Karaigaly and even Gliwice, where gadids dominate (Figure 10). The deeper, more open marine environment of the Carpathian Foredeep at Gliwice is remarkable for the abundance of, partly endemic, pelagic taxa (Paratrisopterus of the Gadidae and Myctophidae; Schwarzhans and Radwańska, 2022). The backreef environment of the Medobory barrier reef at Horodok is the only fossil environment known so far in which labrid otoliths constitute a significant component, but labrid skeletons are common in the earlier reef associated Leitha Formation, including a species of Coris (Schultz, 2013). The various extant members of the family Labridae are found in abrasive coastal and reef environments, and seagrass communities. The genera Coris and Thalassoma identified at Horodok are typical for reef environments.
 The abundance and diversity of goby otoliths warrant a comparison of the Medobory backreef community with those of coeval non-reefal Central Paratethys locations and of the Eastern Paratethys (Figure 11). The first observation to be made is that the Eastern Paratethys goby community is relatively lean in species, and only two of the five species described in Bratishko et al. (2015) are also known from the Central Paratethys. Two of the species unique to the Eastern Paratethys represent genera that today constitute a major part of the endemic Ponto-Caspian gobies, namely Neogobius udovichenkoi and Ponticola zosimovichi. The Medobory reef assemblage and the non-reef late Badenian goby association of the Central Paratethys show a high degree of correlation, but eight species are known only from the Medobory backreef (Figure 11). Given how well the non-reef faunas of the Central Paratethys are known, we conclude that these species probably represent gobies that were adapted to reef environments. Some of them, such as Gobius ukrainicus and Medoborichthys podolicus (Table 1), are in fact common in the Medobory localities. In contrast, we classify 11 species as adapted to open marine neritic, not reef-related habitats, many of which are missing from the Medobory localities while others occur only rarely (Figure 11). These species are Gobius mustus, G. reichenbacherae, G. supraspectabilis, and Sarmatigobius iugosus, and they are mostly singular occurrences or few specimens except for G. reichenbacherae. Gobius reichenbacherae is a widespread species in the European seas of the time in open marine neritic environments (Schwarzhans, 2014; Schwarzhans et al., 2020a, 2020b). It may have lived along the Medobory forereef and occasionally entered the backreef environment.
The abundance and diversity of goby otoliths warrant a comparison of the Medobory backreef community with those of coeval non-reefal Central Paratethys locations and of the Eastern Paratethys (Figure 11). The first observation to be made is that the Eastern Paratethys goby community is relatively lean in species, and only two of the five species described in Bratishko et al. (2015) are also known from the Central Paratethys. Two of the species unique to the Eastern Paratethys represent genera that today constitute a major part of the endemic Ponto-Caspian gobies, namely Neogobius udovichenkoi and Ponticola zosimovichi. The Medobory reef assemblage and the non-reef late Badenian goby association of the Central Paratethys show a high degree of correlation, but eight species are known only from the Medobory backreef (Figure 11). Given how well the non-reef faunas of the Central Paratethys are known, we conclude that these species probably represent gobies that were adapted to reef environments. Some of them, such as Gobius ukrainicus and Medoborichthys podolicus (Table 1), are in fact common in the Medobory localities. In contrast, we classify 11 species as adapted to open marine neritic, not reef-related habitats, many of which are missing from the Medobory localities while others occur only rarely (Figure 11). These species are Gobius mustus, G. reichenbacherae, G. supraspectabilis, and Sarmatigobius iugosus, and they are mostly singular occurrences or few specimens except for G. reichenbacherae. Gobius reichenbacherae is a widespread species in the European seas of the time in open marine neritic environments (Schwarzhans, 2014; Schwarzhans et al., 2020a, 2020b). It may have lived along the Medobory forereef and occasionally entered the backreef environment.
Fish Communities and Micro-environments in the Medobory Backreef (Figure 12, Figure 13, Figure 14)
 All eight studied localities are in or in the vicinity of the city of Horodok approximately 10 km behind the crest of the Medobory barrier reef (Figure 1) in the wide backreef lagoon. The biotic composition shows variations indicating subtle differences in micro-environments from one location to another. The largest faunal components in fishes are those that we interpret to be related to reef environments, probably in the vicinity of patch reefs in the backreef lagoon (Figure 12). This component is characterised by several gobiid species that are not known from outside of the Medobory barrier reef and are therefore considered to be reef prone (see above). They are thought to represent mostly new reef adaptations belonging to genera that survived the middle Badenian salinity crisis and took advantage of reduced competitive pressure. Some groups, however, show a possible relationship to reefal gobies from the Indo-West Pacific (Priolepis lineage) or the tropical West Atlantic (Gobiosomatini), but the nature, source, and timing of these assumed relationships are tentative at present and require verification preferably by skeletal finds. In general, fishes are absent of families typical for reefal environments today, for instance of Pomacanthidae, Pomacentridae, or Acanthuridae. The only notable exception is two labrid species of the genera Coris and Thalassoma, which are typically associated with reefs today (Figure 12). The reef-associated fish component varies between 15% in Staryi Zavod and 72% in Mlyntsi (percentages here and later rounded to whole numbers) (Figure 13). In fact, the reef component is greater than 50% in all fish communities except Staryi Zavod and Lisohirka (18%) (Figure 13). The latter is surprising, since the lithology at this outcrop reflects a reefal setting. We assume that the position of Lisohirka was at the edge of a patch reef body and was also populated by fishes from nearby communities that are not reef prone. Such elements at Lisohirka are the Gadidae and Gaidropsaridae, which are shallow marine non-reef-associated families, and elements indicating a seagrass environment in about equal amounts.
All eight studied localities are in or in the vicinity of the city of Horodok approximately 10 km behind the crest of the Medobory barrier reef (Figure 1) in the wide backreef lagoon. The biotic composition shows variations indicating subtle differences in micro-environments from one location to another. The largest faunal components in fishes are those that we interpret to be related to reef environments, probably in the vicinity of patch reefs in the backreef lagoon (Figure 12). This component is characterised by several gobiid species that are not known from outside of the Medobory barrier reef and are therefore considered to be reef prone (see above). They are thought to represent mostly new reef adaptations belonging to genera that survived the middle Badenian salinity crisis and took advantage of reduced competitive pressure. Some groups, however, show a possible relationship to reefal gobies from the Indo-West Pacific (Priolepis lineage) or the tropical West Atlantic (Gobiosomatini), but the nature, source, and timing of these assumed relationships are tentative at present and require verification preferably by skeletal finds. In general, fishes are absent of families typical for reefal environments today, for instance of Pomacanthidae, Pomacentridae, or Acanthuridae. The only notable exception is two labrid species of the genera Coris and Thalassoma, which are typically associated with reefs today (Figure 12). The reef-associated fish component varies between 15% in Staryi Zavod and 72% in Mlyntsi (percentages here and later rounded to whole numbers) (Figure 13). In fact, the reef component is greater than 50% in all fish communities except Staryi Zavod and Lisohirka (18%) (Figure 13). The latter is surprising, since the lithology at this outcrop reflects a reefal setting. We assume that the position of Lisohirka was at the edge of a patch reef body and was also populated by fishes from nearby communities that are not reef prone. Such elements at Lisohirka are the Gadidae and Gaidropsaridae, which are shallow marine non-reef-associated families, and elements indicating a seagrass environment in about equal amounts.
 Many fishes identified in the various localities either are unclear in their environmental preferences or occur primarily in non-reef neritic associations. These two guilds account for between 7% and 21%, and 14% and 30%, respectively (Figure 13). All of them are shallow-water fishes, and while some may occur in the backreef lagoon as occasional visitors (e.g., Gobius mustus, G. reichenbacherae, or G. supraspectabilis), the others, which are more common, probably had a wider distribution pattern that included reef environments. We consider them to be indicative of deeper regions within the backreef lagoon where occasional visitors have been brought in through surge channels and broadly distributed species could easily exchange with off-reef environments.
Many fishes identified in the various localities either are unclear in their environmental preferences or occur primarily in non-reef neritic associations. These two guilds account for between 7% and 21%, and 14% and 30%, respectively (Figure 13). All of them are shallow-water fishes, and while some may occur in the backreef lagoon as occasional visitors (e.g., Gobius mustus, G. reichenbacherae, or G. supraspectabilis), the others, which are more common, probably had a wider distribution pattern that included reef environments. We consider them to be indicative of deeper regions within the backreef lagoon where occasional visitors have been brought in through surge channels and broadly distributed species could easily exchange with off-reef environments.
The new collection activities underlying this study have also used finer sieves down to 0.3 mm diameter (localities Mlyntsi, Novyi Pliazh, Staryi Zavod, Lisohirka, and Skala). As a result, some of these localities yielded many small otoliths well below 1 mm in length, which otherwise are typically not recovered. Most otoliths <1 mm in diameter are from juvenile, maybe even larval Gobiidae that cannot be identified to a generic or species level. In addition, rare occurrences have been observed of pipefishes (Syngnathidae) of the genus Syngnathus and clingfishes (Gobiesocidae) of the genus Apletodon. These fishes have small otoliths in the range of 0.2 mm to just below 1 mm in length and are both typically associated with seagrass meadows (Pollard, 1984). Seagrass meadows in fact often act as nursery grounds and are therefore also a preferred environment for juvenile fishes such as gobies (Heck and Orth, 1980; Jaxion-Harm et al., 2012; Erzad et al., 2020). The proportion of otoliths indicating a potential seagrass environment varies widely from 0% to 52% and is particularly high in Staryi Zavod (52%) and Lisohirka (35%) (Figure 13). Seagrass palaeoenvironments have been commonly recorded from the Middle Miocene of the Central Paratethys (Doláková et al., 2014; Forsey, 2018; Harzhauser et al., 2018; Holcová et al., 2019). Otoliths have occasionally also been used to indicate potential seagrass environments. Brzobohatý et al. (2007) considered the abundance of otoliths of the genus Spondyliosoma in the lower Badenian of Kienberg at Mikulov, Czech Republic, as indicative of a possible seagrass environment. These otoliths, however, are no more present in the upper Badenian. In their palaeoenvironmental assessment of Badenian rocks in the northern Vienna Basin. Harzhauser et al. (2018) mentioned juvenile sea breams (Sparidae) and gobies of the genus Lesueurigobius as potential indicators for seagrass environments. These taxa are here placed in the category of unclear/wide environmental distribution even though they may also be common in seagrass environments. Other possible indicators of seagrass in the Medobory backreef localities are lucinid clams in live position, ostracods of the genera Aurila, Loxoconcha, and Xestroleberis, and certain permanently motile foraminifera (see above). The occurrence of these potential seagrass indicators is congruent with that of the otolith associations, and the indicators occur most diversely at Staryi Zavod and Lisohirka (Figure 13).
Other environmental indicators are rare in the otolith associations. A group of open marine, outer-shelf fishes such as Bellottia (Bythitidae) and Brachydeuterus (Haemulidae) are absent or rare and achieve the highest percentage (8%) in Novyi Pliazh (Figure 13). Their relative abundance could indicate a deeper position in the backreef, possibly not far from a surge channel that may have facilitated the incursion of such outer-shelf faunal elements. Interestingly, Novyi Pliazh is also the only locality with a singular occurrence of a real bathyal fish, an unidentifiable specimen of Physiculus (Moridae). Freshwater fishes are also rare and represented by a species of Umbra (Umbridae), an unidentified Gasterosteidae and Palaeolebias (Cyprinodontidae). They occur at a frequency of one to three specimens per species in the collection clearly indicating that freshwater input did not play a major role in any of the studied locations.
 The resulting picture is that of a wide backreef lagoon at a moderate distance from the crest of the barrier reef (Figure 14). The backreef lagoon probably contained smaller coralline algal and coral patch reefs and common biostromes on the sea floor. However, certain differences in micro-environments can also be observed. A full assessment, however, can only be performed for localities with a complete data set including otoliths sampled from small fractions down to 0.3 mm sieve diameter (Mlyntsi, Novyi Pliazh, Staryi Zavod, Lisohirka, and Skala; Figure 13). Presumed reef-associated fishes formed a major component in all studied localities, often the dominant one. The highest percentage of reef-related fishes was identified in Mlyntsi, which is therefore considered to be located on or close to a patch reef. The same could be true for Shydlivshchyna, from where, however, no small fraction had been sampled, and Lisohirka (Figure 14). The faunal community at Lisohirka is in fact unusual, as it contains a relatively low number of reef-related forms along with many non-reef neritic and widely distributed forms. The lithology indicates a close relation to a reefal environment, while a high percentage of juvenile goby otoliths indicates a seagrass environment. We assume that Lisohirka was situated at the fringes of a patch reef with a nearby seagrass meadow and that the observed fish community therefore represents a mixture of both. Non-reef and deeper neritic/open marine elements might indicate a somewhat deeper position in the backreef lagoon that was easier to populate for these fishes. The presence of usually uncommon open marine faunal elements would indicate the presence of a surge channel through the barrier reef nearby, helping such fishes to occasionally venture into the lagoon. We interpret Novyi Pliazh, Skala, and possibly Kozatskyi Yar to have been in such a position (Figure 14). The fish community of Staryi Zavod is the most divergent of all characterised by an abundance of juvenile goby otoliths, and a few other juvenile otoliths. It is also the only one with pipefish (Syngnathidae) otoliths. From a lithological perspective, Staryi Zavod contains the finest-grained sediment observed in the studied localities. We interpret this location as having been positioned in a seagrass meadow (Figure 14). The faunal associations observed in ostracods, and foraminifera are also congruent with this interpretation but the distribution pattern of lucinid bivalves departs somewhat. It is possible that seagrass occurrences were widely and possibly discontinuously distributed in the Medobory backreef.
The resulting picture is that of a wide backreef lagoon at a moderate distance from the crest of the barrier reef (Figure 14). The backreef lagoon probably contained smaller coralline algal and coral patch reefs and common biostromes on the sea floor. However, certain differences in micro-environments can also be observed. A full assessment, however, can only be performed for localities with a complete data set including otoliths sampled from small fractions down to 0.3 mm sieve diameter (Mlyntsi, Novyi Pliazh, Staryi Zavod, Lisohirka, and Skala; Figure 13). Presumed reef-associated fishes formed a major component in all studied localities, often the dominant one. The highest percentage of reef-related fishes was identified in Mlyntsi, which is therefore considered to be located on or close to a patch reef. The same could be true for Shydlivshchyna, from where, however, no small fraction had been sampled, and Lisohirka (Figure 14). The faunal community at Lisohirka is in fact unusual, as it contains a relatively low number of reef-related forms along with many non-reef neritic and widely distributed forms. The lithology indicates a close relation to a reefal environment, while a high percentage of juvenile goby otoliths indicates a seagrass environment. We assume that Lisohirka was situated at the fringes of a patch reef with a nearby seagrass meadow and that the observed fish community therefore represents a mixture of both. Non-reef and deeper neritic/open marine elements might indicate a somewhat deeper position in the backreef lagoon that was easier to populate for these fishes. The presence of usually uncommon open marine faunal elements would indicate the presence of a surge channel through the barrier reef nearby, helping such fishes to occasionally venture into the lagoon. We interpret Novyi Pliazh, Skala, and possibly Kozatskyi Yar to have been in such a position (Figure 14). The fish community of Staryi Zavod is the most divergent of all characterised by an abundance of juvenile goby otoliths, and a few other juvenile otoliths. It is also the only one with pipefish (Syngnathidae) otoliths. From a lithological perspective, Staryi Zavod contains the finest-grained sediment observed in the studied localities. We interpret this location as having been positioned in a seagrass meadow (Figure 14). The faunal associations observed in ostracods, and foraminifera are also congruent with this interpretation but the distribution pattern of lucinid bivalves departs somewhat. It is possible that seagrass occurrences were widely and possibly discontinuously distributed in the Medobory backreef.
CONCLUSION AND OUTLOOK
New collections from five localities near the city of Horodok in western Ukraine have yielded a significant number of additional fossil fish otoliths from the Medobory backreef. This time, smaller fractions (down to 0.3 mm in diameter) were also sampled for otoliths as well and generated new insights. The principal results are as follows.
1. With the new locations and the sampling of the smaller fraction, the otolith-based fish fauna of the area has now increased to a total of 62 species.
2. Gobioids are the dominant group with 25 species (40.4% of all species) and 78.4% of the specimens.
3. Labridae constitute an important secondary component indicating proximity to reefs. Labrid otoliths are otherwise rare in the fossil record.
4. The small fraction <1 mm in diameter has yielded primarily unidentifiable gobiid otoliths of juveniles and a few new species with primarily small otoliths such as pipefishes (Syngnathidae) and clingfishes (Gobiesocidae).
5. The new sampling allows one to distinguish micro-environments in the composition of the fish communities in the Medobory backreef: (a) a reef-associated environment dominated by supposed reef-related gobies and labrids; (b) a deeper lagoon environment which in addition to faunal reef elements also includes certain off-reef neritic fishes; and (c) an environment within or in the vicinity of seagrass meadows dominated by juvenile gobies, which may have acted as a fish nursery, and with admixture of syngnathids.
Our study documents how new sampling in the Medobory backreef not only significantly contributes to the size of the fauna but also helps to recognise and define palaeoenvironments in this unique setting. The Medobory backreef also shows how fragmented and diversified the fish fauna was in the Paratethys during the early Serravallian (late Badenian/Konkian). We believe that future prospecting for otoliths in the Miocene of the Paratethys, particularly in its eastern part, can provide so much more information about the evolution of fishes in this evolutinary hotspot at that time.
ACKNOWLEDGMENTS
We cordially thank M. Harzhauser and O. Mandic (both Naturhistorisches Museum Wien, Austria) for providing identifications for the figured molluscs. O. Klots thanks leading members of the Winogradsky Club, P. Kryvyi and O. Kotlinskiy, for valuable consultations as well as V. Stolyar, M. Marchenko, and A. Khanenko, for their support, practical help during field studies and in-house processing of materials. The research of O. Kovalchuk was carried out as part of the research projects ‘Development of the biota in the late Cenozoic of Ukraine’ (0120U100451) and ‘The role of particular fish groups in the functioning of late Mesozoic and Cenozoic ecosystems of Eastern Europe’ (0123U102984) funded by the National Academy of Sciences of Ukraine. We are sincerely thankful to M. Harzhauser and two anonymous reviewers whose comments and suggestions significantly improved the original text. We also wish to thank the Handling Editor Z. Barkaszi and the entire PE team for positive and prompt evaluation of our submission.
REFERENCES
Agassiz, J.L.R. 1835. Revue critique des poisson fossiles figurés dans l’Ittiolitologie Veronese. Neues Jahrbuch für Mineralogie, 1835:290–316.
Agorreta, A., San Mauro, D., Schliewen, U., Van Tassell, J.L., Kovačić, M., Zardoya, R., and Rüber, L. 2013. Molecular phylogenetics of Gobioidei and phylogenetic placement of European gobies. Molecular Phylogenetics and Evolution, 69:619–633.
https://doi.org/10.1016/j.ympev.2013.07.017
Aguilera, O. and Rodrigues de Aguilera, D. 2001. An exceptional coastal upwelling fish assemblage in the Caribbean Neogene. Journal of Paleontology, 75:732–742.
https://doi.org/10.1017/s0022336000039767
Aiello, G. and Szczechura, J. 2004. Middle Miocene ostracods of the Fore-Carpathian Depression (Central-Paratethys, southwestern Poland). Bollettino della Società Paleontologica Italiana, 43:11–70.
Andreeva-Grigorovich, A.S., Kulchytsky, Y.O., Gruzman, A.D., Lozynyak, P.Y., Petrashkevich, M.Y., Portnyagina, L.O., Ivanina, A.V., Smirnov, S.E., Trofimovich, N.A., Savytska, N.A., Ponomaryova, L.D., and Shvareva, N.Y. 1996. Scheme of stratigraphy of Neogene deposits of the Western (Central) Paratethys within Ukraine. Paleontological Collection, 31:8–82. (In Ukrainian)
Báldi, K., Velledits, F., Ćorić, S., Lemberkovics, V., Lörincz, K., and Shevelev, M. 2017. Discovery of the Badenian evaporites inside the Carpathian Arc: implications for global climate change and Paratethys salinity. Geologica Carpathica, 68:193–206.
https://doi.org/10.1515/geoca-2017-0015
Bannikov, A.F. 2010. Fossil acanthopterygian fishes (Teleostei, Acanthopterygii), p. 1–244. In Tatarinov, L.P., Vorobyeva, E.I., and Kurochkin, E.N. (eds.), Fossil Vertebrates of Russia and Adjacent Countries. GEOS, Moscow. (In Russian, with English summary)
Bannikov, A.F. and Kotlyar, A.N. 2015. A new genus and species of Early Sarmatian porgies (Perciformes, Sparidae) from the Krasnodar region. Paleontological Journal, 49:627–635.
https://doi.org/10.1134/S0031030115060052
Bartol, M., Mikuž, V., and Horvat, A. 2014. Palaeontological evidence of communication between the Central Paratethys and the Mediterranean in the late Badenian/early Serravallian. Palaeogeography, Palaeoclimatology, Palaeoecology, 394:144–157.
https://doi.org/10.1016/j.palaeo.2013.12.009
Bassoli, G. 1906. Otoliti fossili terziari dell’Emilia. Rivista Italiana di Paleontologia, 12:36–61.
Baykina, E.M. and Schwarzhans, W.W. 2017. Description of Karaganops n. gen. perratus (Daniltshenko 1970) with otoliths in situ, an endemic Karaganian (Middle Miocene) herring (Clupeidae) in the Eastern Paratethys. Swiss Journal of Palaeontology, 136:129–140.
https://doi.org/10.1007/s13358-016-0115-4
Bellwood, D.R., Schultz, O., Siqueira, A.C., and Cowman, P.F. 2019. A review of the fossil record of the Labridae. Annalen des Naturhistorischen Museums Wien A, 121:125–193.
Berg, L.S. 1937. A classification of fish-like vertebrates. Izvestiya Akademii Nauk USSR, Seriya Biologischeskaya, 4:1277–1280. (In Russian)
Berg, L.S. 1940. Classification of fishes, both recent and fossil. Travaux de l’Institut zoologique de l’Académie des sciences de l’URSS, 5:1–517.
Bleeker, P. 1859. Enumeratio specierum piscium hucusque in Archipelago Indico observatarum, adjectis habitationibus citationibusque, ubi descriptiones earum recentiores reperiuntur, nec non speciebus Musei Bleekeriani Bengalensibus, Japonicis, Capensibus Tasmanicisque. Acta Societas Scientarum Indo-Neerlandaises, 6:1–276.
Bleeker, P. 1862. Sur quelques genres de la famille des Pleuronectoïdes. Verslagen en mededeelingen der Koninklijke Akademie van Wetenschappen, 13:422–429.
Bleeker, P. 1876. Atlas ichthyologique des Indes Orientales Neerlandaises, publie sous les auspices du Gouvernement colonial neerlandais. 8. Percoides II (Spariformes). Amsterdam.
Bloch, M.E. and Schneider, J.G. 1801. Systema ichthyologiae iconibus ex illustratum. Post obitum auctoris opus inchoatum absolvit, correxit, interpolavit Johannis Gottlob Schneider, Saxo. Sanderiano Commissum, Berolini [Berlin].
Bonaparte, C.L. 1831. Saggio di una distribuzione metodica degli animali vertebrati. Roma.
Bonaparte, C.L. 1833. Iconografia della fauna italica per le quattro classi degli animali vertebrati. Tomo III. Pesci. Roma.
Bonaparte, C.L. 1845. Specchio generale del sistema ittiologico. Atti della sesta Riunione degli Scienziati Italiani, 6:386–390.
Bondar, О. 2006. Biostratigraphy of Middle-Upper Miocene deposits of Southern Ukraine based on ostracods. Dissertation for obtaining the scientific degree of Candidate of Geological Sciences. Dnipropetrovsk. (In Ukrainian)
Bratishko, A., Schwarzhans, W., Reichenbacher, B., Vernyhorova, Y., and Ćorić, S. 2015. Fish otoliths from the Konkian (Miocene, early Serravallian) of Mangyshlak (Kazakhstan): Testimony to an early endemic evolution in the Eastern Paratethys. Paläontologische Zeitschrift, 89:839–889.
https://doi.org/10.1007/s12542-015-0274-4
Bratishko, A., Schwarzhans, W., and Vernyhorova, Y. 2023. The endemic marine fish fauna from the Eastern Paratethys reconstructed from otoliths from the Miocene (middle Sarmatian s.l.; Bessarabian) of Jurkine (Kerch Peninsula, Crimea). Rivista Italiana di Paleontologia e Stratigrafia, 129:111–184.
https://doi.org/10.54103/2039-4942/18877
Briggs, J.C. 1955. A monograph of the clingfishes (Order Xenopterygii). Stanford Ichthyological Bulletin, 6:i–iv + 1–224.
Brzobohatý, R. and Schultz, O. 1978. Die Fischfauna des Badenien, p. 441–465. In Papp, A., Cicha, I., Seneš, J., and Steininger, F. (eds.), M4, Badenien. Chronostratigraphie und Neostratotypen, 6.
Brzobohatý, R., Nolf, D., and Kroupa, O. 2007. Fish otoliths from the middle Miocene of Kienberg at Mikulov, Czech Republic, Vienna Basin: their paleoenvironmental and paleogeographic significance. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, 77:167–196.
Brzobohatý, R., Zahradníková, B., and Hudáčková, N. 2022. Fish otoliths and foraminifera from the Borský Mikuláš section (Slovakia, middle Miocene, upper Badenian, Vienna Basin) and their paleoenvironmental significance. Rivista Italiana di Paleontologia e Stratigrafia, 128:515–537.
https://doi.org/10.54103/2039-4942/15773
Cappetta, H. 2012. Chondrichthyes (Mesozoic and Cenozoic Elasmobranchii: Teeth), p. 1–512. In Schultze, H.-P. (ed.), Handbook of Paleoichthyology 3E. Verlag Dr. Friedrich Pfeil, München, Germany.
Carnevale, G. 2015. Middle Miocene wrasses (Teleostei, Labridae) from the St. Margarethen (Burgenland, Austria). Palaeontographica, Abteilung A: Palaeozoology - Stratigraphy, 304:121–159.
Carnevale, G. and Harzhauser, M. 2013. Middle Miocene rockling (Teleostei, Gadidae) from the Paratethys (St. Margarethen in Burgenland, Austria). Buletin of Geosciences, 88:609–620.
https://doi.org/10.3140/bull.geosci-1432
Carnevale, G. and Collette, B.B. 2014. † Zappaichthys harzhauseri, gen. et sp. nov., a new Miocene toadfish (Teleostei, Batrachoidiformes) from the Paratethys (St. Margarethen in Burgenland, Austria), with comments on the fossil record of batrachoidiform fishes. Journal of Vertebrate Paleontology, 34:1005–1017.
https://doi.org/10.1080/02724634.2014.854801
Carolin, N., Bajpai, S., Mauryia, A.S., and Schwarzhans, W. 2023. New perspectives on late Tethyan Neogene biodiversity development of fishes based on Miocene (~ 17 Ma) otoliths from southwestern India. PalZ, 97:43–80.
https://doi.org/10.1007/s12542-022-00623-9
Chaine, J. and Duvergier, J. 1934. Recherches sur les otolithes des poissons. Etude descriptive et comparative de la sagitta des Teléostéens. Actes de la Societé Linnéenne de Bordeaux, 86:5–256.
Charmpila, E.A., Wagner, M., and Reichenbacher, B. 2021. First documentation of the otoliths of the species of Gouania (Teleostei: Gobiesocidae) in the Mediterranean Sea. Journal of Fish Biology, 98:1450–1455.
https://doi.org/10.1111/jfb.14646
Cicha, I., Rögl, F., Rupp, C., and Ctyroka, J. 1998. Oligocene–Miocene foraminifera of the Central Paratethys. Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, 549:1–325.
Cornée, J.-J., Moissette, P., Saint Martin, J.-P., Kázmér, M., Tóth, E., Görög, Á., Dulai, A., and Müller, P. 2009. Marine carbonate systems in the Sarmatian (Middle Miocene) of the Central Paratethys: the Zsámbék Basin of Hungary. Sedimentology, 56:1728–1750.
https://doi.org/10.1111/j.1365-3091.2009.01055.x
Cuvier, G. 1814. Observations et recherches critiques sur différentes poissons de la Méditerranée et, à leur occasion, sur des Poissons des autres mers plus ou moins liés avec eux. Bulletin des Sciences, par la Société Philomatique de Paris, 1814:80–92.
Cuvier, G. 1816. Le Règne Animal distribué d'après son organisation pour servir de base à l'histoire naturelle des animaux et d'introduction à l'anatomie comparée. Les reptiles, les poissons, les mollusques et les annélides. Tome II. Deterville, Paris.
Didkovsky, V.Y. 1959. On the microfauna of the sediments of the Konkian horizon of the Ukrainian SSR. Dopovidi Akademii nauk URSR, 4:412–416. (In Ukrainian)
Didkovsky, V.Y. 1964. Biostratigraphy of Neogene deposits in the south of the Russian Platform based on foraminifera fauna. Thesis for obtaining the scientific degree of Doctor of Geological and Mineralogical Sciences. Kyiv. (In Russian)
Didkovsky, V.Y. and Satanovskaya, Z.N. 1970. Paleontological Handbook. Vol. 4. Foraminifera of the Miocene of Ukraine. Naukova Dumka, Kyiv. (In Russian)
Djafarova, J.D. 2006. Neogene otoliths of Azerbaijan. Nafta, Baku. (In Russian)
Doláková, N., Holcová, K., Nehyba, S., Hladilová, Š., Brzobohatý, R., Zágoršek, K., Hrabovský, J., Seko, M., and Utescher, T. 2014. The Badenian parastratotype at Židlochovice from the perspective of the multiproxy study. Neues Jahrbuch Geologie Paläontologie Abhandlungen, 271:169–201.
https://doi.org/10.1127/0077-7749/2014/0383
Erzad, A.F., Hartoko, A., and Muskananfola, M.R. 2020. Seagrass ecosystem as fish larvae habitat and its biomass algorithm from Sentinel-2A satellite at Kemjun Island, Karimunjawa, Indonesia. AACL Bioflux, 13:1511–1521.
Esteban, M. 1980. Tertiary reefs of western Mediterranean with emphasis on Miocene reefs. American Association of Petroleum Geologists Fall Education Conference, Houston.
Forsey, G.F. 2016. Ostracods as proxies for past seagrass: A review. Palaeogeography, Palaeoclimatology, Palaeoecology, 447:22–28.
https://doi.org/10.1016/j.palaeo.2016.01.028
Forsey, G.F. 2018. Seagrass distribution and global biodiversity hotspot during the Badenian (Middle Miocene) of Central Paratethys as indicated by ostracods and other faunal elements. Poster.
https://doi.org/10.13140/RG.2.2.30813.05600
Fricke, R., Eschmeyer, W.N., and Van der Laan, R. (eds.). 2024. Eschmeyer’s catalog of fishes: genera, species, references (Electronic version accessed 03 June 2024).
http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
Garecka, M. and Olszewska, B. 2011. Correlation of the Middle Miocene deposits in SE Poland and Western Ukraine based on foraminifera and calcareous nannoplankton. Annales Societalis Geologorum Poloniae, 81:309–330.
Garman, S. 1899. The Fishes. Reports on an exploration off the west coasts of Mexico, Central and South America, and off the Galapagos Islands by the United States Fish Commission steamer “Albatross” during 1891. Memoirs of the Museum of Comparative Zoology, 24:1–431.
Gedl, P. and Peryt, D. 2011. Dinoflagellate cyst, palynofacies and foraminiferal records of environmental changes related to the late Badenian (Middle Miocene) transgression at Kudryntsi (Western Ukraine). Annales Societalis Geologorum Poloniae, 81:331–349.
Gill, T.N. 1861. Catalogue of the fishes of the eastern coast of North America, from Greenland to Georgia. Proceedings of the Academy of Natural Sciences of Philadelphia, 13 (Suppl.):1–63.
Gill, T.N. 1863. On the gobioids of the eastern coast of the United States. Proceedings of the Academy of Natural Sciences of Philadelphia, 15:267–271.
Gill, T.N. 1865. On a new family type of fishes related to the blennioids. Annals of the Lyceum of Natural History of New York, 8:141–144.
Goodrich, E.S. 1909. Vertebrata Craniata (first faciscle: cyclostomes and fishes). In Lankester, R. (ed.), A treatise on zoology, Part 9. Black, London.
Goretsky, V.O. and Didkovsky, V.Y. 1975. Miocene. Volyn-Podolian plate, p. 84–110. In Bondarchuk, V.G., Didkovsky, V.Y., and Kulichenko, V.G. (eds.), Stratigraphy of the Ukrainian SSR. Vol. 10. Neogene. Naukova Dumka, Kyiv. (In Ukrainian)
Górka, M., Studencka, B., Jasionowski, M., Hara, U., Wysocka, A., and Poberezhskyy, A. 2012. The Medobory Hills (Ukraine): Middle Miocene reef systems in the Paratethys, their biological diversity and lithofacies. Biuletyn Państwowego Instytutu Geologicznego, 449:147–174.
Gumeniuk, G.L. and Kryvyi, P.A. 2016. Serhiy Winogradsky and the city of Horodok (to the 160th anniversary of the birth). Svitohlyad, 5:42–45. (In Ukrainian)
Günther, A. 1859. Catalogue of the fishes in the British Museum. Catalogue of the acanthopterygian fishes in the collection of the British Museum. Gasterosteidae, Berycidae, Percidae, Aphredoderidae, Pristipomatidae, Mullidae, Sparidae. Volume1. British Museum, London.
Gut, C., Vukić, J., Šanda, R., Moritz, T., and Reichenbacher, B. 2020. Identification of past and present gobies: distinguishing Gobius and Pomatoschistus (Teleostei: Gobioidei) species using characters of otoliths, meristics and body morphometry. Contributions to Zoology, 89:282–323.
https://doi.org/10.1163/18759866-bja10002
Hamilton, F. 1822. An account of the fishes found in the river Ganges and its branches. Edinburgh & London.
Harzhauser, M. and Piller, W.E. 2007. Benchmark data of a changing sea - palaeogeography, palaeobiogeography and events in the Central Paratethys during the Miocene. Palaeo, 253:8–31.
https://doi.org/10.1016/j.palaeo.2007.03.031
Harzhauser, M., Peckmann, J., Birgel, D., Draganits, E., Mandic, O., Theobalt, D., and Huemer, J. 2014. Stromatolites in the Paratethys Sea during the Middle Miocene Climate Transition as witness of the Badenian Salinity Crisis. Facies, 60:429–444.
https://doi.org/10.1007/s10347-013-0391-z
Harzhauser, M., Grunert, P., Mandic, O., Lukeneder, P., Gallardo, Á.G., Neubauer, T.A., Carnevale, G., Landau, B.M., Sauer, R., and Strauss, P. 2018. Middle and late Badenian palaeoenvironments in the northern Vienna Basin and their potential link to the Badenian Salinity Crisis. Geologica Carpathica, 69:149–168.
https://doi.org/10.1515/geoca-2018-0009
Harzhauser, M., Kranner, M., Mandic, O., Strauss, P., Siedl, W., and Piller, W.E. 2020. Miocene lithostratigraphy of the northern and central Vienna Basin (Austria). Austrian Journal of Earth Sciences, 113:169–200.
https://orcid.org/0000-0002-4471-6655
Harzhauser, M., Guzhov, A., Landau, B., Kern, A.K., and Neubauer, T.A. 2023. Oligocene to Pliocene mudwhelks (Gastropoda: Potamididae, Batillariidae) of the Eurasian Paratethys Sea - diversity, origins and mangroves. Palaeogeography, Palaeoclimatology, Palaeoecology, 630:111811.
https://doi.org/10.1016/j.palaeo.2023.111811
Harzhauser, M., Landau, B., Mandic, O., and Neubauer, T.A. 2024. The Central Paratethys Sea - Rise and demise of a Miocene European marine biodiversity hotspot. Scientific Reports, 14:16288.
https://doi.org/10.1038/s41598-024-67370-6
Heck, K.L. and Orth, R.J. 1980. Seagrass habitats: the role of habitat complexity, competition and predation in structuring associated fish and motile macroinvertebrate assemblages, p. 449–464. In Kennedy, V.S. (ed.), Estuarine Perspectives. Academic Press, New York.
Hoedemakers, K. and Batllori, J. 2005. Fish otoliths from the Early and Middle Miocene of the Penedis (Catalunya, Spain). Batallerna, 12:105–134.
Holcová K., Dašková J., Fordinál K., Hrabovský J., Milovský R., Scheiner F., and Vacek F. 2019. A series of ecostratigraphic events across the Langhian/Serravallian boundary in an epicontinental setting: the northern Pannonian Basin. Facies, 65:36.
https://doi.org/10.1007/s10347-019-0576-1
Iljin, B.S. 1930. Le système des Gobiidés. Trabajos Instituto Espanol de Oceanografia, 2:1–63.
Jaxion-Harm, J., Saunders, J., and Speight, M.R. 2012. Distribution of fish in seagrass, mangroves and coral reefs: life-stage dependent habitat use in Honduras. Revista de Biología Tropical, 60:683–698.
https://doi.org/10.15517/rbt.v60i2.3984
Jordan, D.S. and Evermann, B.W. 1898. The fishes of North and Middle America: a descriptive catalogue of the species of fish-like vertebrates found in the waters of North America, north of the isthmus of Panama. Part II. Bulletin of the United States National Museum, 47:1241–2183.
Jordan, D.S. and Goss, D.K. 1889. A review of the flounders and soles (Pleuronectidae) of America and Europe. United States Commission of Fish and Fisheries, Report of the Commissioner 14 (for 1886), 2:225–342.
Kaup, J.J. 1858. Uebersicht der Familie Gadidae. Archiv für Naturgeschichte, 24:85–93.
Klein, E.E. 1885. Beiträge zur Bildung des Schadels der Knochenfische. II: Jahreshefte des Vereins für Vaterländische Naturkunde in Würtemberg, 41:107–261.
Koken, E. 1884. Über Fisch-Otolithen, insbesondere über diejenigen der norddeutschen Oligocän-Ablagerungen. Zeitschrift der Deutschen Geologischen Gesellschaft, 36:500–565.
Koken, E. 1891. Neue Untersuchungen an tertiären Fisch-Otolithen. II. Zeitschrift der Deutschen Geologischen Gesellschaft, 43:77–170.
Kollmann, K. 1971. Die Ostracoden der Eggenburger Schichtengruppe Niederösterreichs, p. 605–717. In Steininger, F. and Senes, J. (eds.), Chronostratigraphie und Neostratotypen, vol. 2, Miozän der zentralen Paratethys, M1 Eggenburgien, Die Eggenburger Schichtengruppe und ihr Stratotypus. Bratislava.
Korolyuk, I.K. 1952. Podolian toutras and conditions of their formation. Trudy Instituta geologicheskikh nauk Akademii nauk SSSR, 56:1–136. (In Russian)
Kováč, M., Andreyeva-Grigorovich, A., Bajraktarević, Z., Brzobohatý, R., Filipescu, S., Fodor, L., Harzhauser, M., Nagymarosy, A., Oszczypko, N., Pavelić, D., Rögl, F., Saftić, B., Sliva, L., and Studencka, B. 2007. Badenian evolution of the Central Paratethys Sea: paleogeography, climate and eustatic sea-level changes. Geologica Carpathica, 58:579–606.
Kováč, M., Hudáčková, N., Halásová, E., Kováčová, M., Holcová, K., Oszczypko-Clowes, M., Báldi, K., Less, G., Nagymarosy, A., Ruman, A., Klučiar, T., and Jamrich, M. 2017. The Central Paratethys palaeoceanography: a water circulation model based on microfossil proxies, climate, and changes of depositional environment. Acta Geologica Slovaka, 9:75–114.
Kovalenko, V.A. 2013. Ostracods from the Konkian sediments of the Eastern Black Sea region. Reports of the National Academy of Sciences of Ukraine, 8:105–112. (In Russian, with English summary)
Kramberger, D. 1883. Die jungtertiäre Fischfauna Croatiens II. Beiträge zur Paläontologie Österreich-Ungarns und des Orients, 3:65–85.
Krasheninnikov, V.A., Basov, I.A., and Golovina, L.A. 2003. Eastern Paratethys: Tarkhanian and Konkian regional stages (stratigraphy, micropalaeontology, bionomy, paleogeographical relations). Nauchnyi Mir, Moscow. (In Russian)
Kryvyi, P.A., Gumeniuk, G.L., and Bratchykova, Yu.V. 2022. Serhiy Winogradsky - a great Ukrainian. Infusion & Chemotherapy, 3:57–64. (In Ukrainian, with English summary)
https://doi.org/10.32902/2663-0338-2022-3-57-64
Kudrin, L.N. 1966. Stratigraphy, facies and ecological analysis of fauna from the Paleogene and Neogene deposits of the Fore-Carpathian area. University Press, Lviv. (In Russian)
Lacépède, B.G.E. 1801. Histoire naturelle des poissons. Plassan, Paris.
Langer, M.R. 1993. Epiphytic foraminifera. Marine Micropaleontology, 20:235–265.
https://doi.org/10.1016/0377-8398(93)90035-V
Lin, C.-H. and Chang, C.-W. 2012. Otolith atlas of Taiwan fishes. National Museum of Marine Biology and Aquarium, 12:1–415.
Lin, C.-H., Girone, A., and Nolf, D. 2015. Tortonian fish otoliths from turbiditic deposits in Northern Italy: Taxonomic and stratigraphic significance. Geobios, 48:249–261.
https://doi.org/10.1016/j.geobios.2015.03.003
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus 1. Editio decima, reformata. Laurentii Salvii, Holmia [Stockholm].
Ljuljev, Y.B. 1967. Ostracods and stratigraphy of Miocene deposits of southern Ukraine. Thesis for obtaining the scientific degree of Candidate of Geological and Mineralogical Sciences. Kyiv. (In Russian)
Ljuljev, Y.B. 1969. Konkian ostracods of the Black Sea Basin. Visnyk Kyivskoho universytetu, seriya heolohii, 11:23–28. (In Ukrainian)
Lombarte, A., Chic, Ò., Parisi-Baradad, V., Olivella, R., Piera, J., and García-Ladona, E. 2006. A web-based environment from shape analysis of fish otoliths. The AFORO Database. Scientia Marina, 70:147–152.
Marsili, S., Carnevale, G., Danese, E., Bianucci, G., and Landini, W. 2007. Vertébrés fossiles du Miocène inférieur de la Montagna della Maiella, Italie. Annales de Paleontologie, 93:27–66.
Mikerina, T.B. and Pinchuk, T.N. 2014. Distribution and source of dispersed organic matter in the Karaganian-Konkian-Sarmatian deposits of the Eastern Paratethys. Geology, Geography and Global Energy, 4:20–33.
Miller, K.G., Browning, J.V., Schmelz, W.J., Kopp, R.E., Mountain, G.S., and Wright, J.D. 2020. Cenozoic sea-level and cryospheric evolution from deep-sea geochemical and continental margin records. Science Advances, Review, 6:eaaz1346.
https://doi.org/10.1126/sciadv.aaz1346
Moissette, P., Koskeridou, E., Cornée, J.-J., Guillocheau, F., and Lécuyer, C. 2007. Spectacular preservation of seagrasses and seagrass-associated communities from the Pliocene of Rhodes, Greece. Palaios, 22:200–211.
https://doi.org/10.2110/palo.2005.p05-141r
Müller, J. 1846. Über den Bau und die Grenzen der Ganoiden und über das natürlichen System der Fische. Abhandlungen der königlichen Akademie der Wissenschaften Berlin, 1846:117–216.
Münster, Graf G. zu. 1846. Ueber die in der Tertiär-Formation des Wiener Beckens vorkommenden Fisch-Ueber-reste, mit Beschreibung einiger neuen merkwürdigen Arten. Beiträge zur Petrefacten-Kunde, 7:1–31.
Nardo, G.D. 1827. Prodromus observationum et disquisitionum Adriaticae ichthyologiae Giornale di fisica, chimica e storia naturale, medicina, ed arti, 10:22–40.
Nelson, J.S., Grande, T.C., and Wilson, M.V.H. 2016. Fishes of the World. Fifth edition. John Wiley and Sons, Hoboken.
Nolf, D. 2013. The diversity of fish otoliths, past and present. Operational directorate “Earth and History of Life” of the Royal Belgian Institute of Natural Sciences, Brussels, Belgium.
Nolf, D. and Brzobohatý, R. 2004. Otolithes de poissons su Miocène inferieur Piemontais. Rivista Piemontese di Storia naturale, 25:69–118.
Nolf, D. and Steurbaut, E. 1979. Les otolithes de téléostéens des faluns sallomaciens d’Orthez et de Sallespisse (Miocène moyen d’Aquitaine méridionale, France). Palaeontographica A, 164:1–23.
Pallas, P.S. 1814. Zoographia Rosso-Asiatica, sistens omnium animalium in extenso Imperio Rossico et adjacentibus maribus observatorum recensionem, domicilia, mores et descriptiones anatomen atque icones plurimorum. Academia Scientiarum, Petropolis [Saint Petersburg].
Palcu, D.V., Golovina, L.A., Vernyhorova, Y.V., Popov, S.V., and Krijgsman, W. 2017. Middle Miocene paleoenvironmental crises in Central Eurasia caused by changes in marine gateway configuration. Global and Planetary Change, 158:57–71.
https://doi.org/10.1016/j.gloplacha.2017.09.013
Palcu, D.V., Popov, S.V., Golovina, L.A., Kuiper, K.F., Liu, S., and Krijgsman, W. 2019. The shutdown of an anoxic giant: Magnetostratigraphic dating of the end of the Maikop Sea. Gondwana Research, 67:82–100.
https://doi.org/10.1016/j.gr.2018.09.011
Paruch-Kulczycka, J. 1992. Małżoraczki środkowego miocenu (badenu) z otworu Broniszowice (SW Polska). Kwartalnik Geologiczny, 36:259–280.
Permanyer, A. and Esteban, M. 1973. El arrecife mioceno de Sant Pau d'Ordal (provincia de Barcelona). Barcelona, Institut d'Investigacions Geolegiques, 28:45–72.
Peryt, T.M. 2006. The beginning, development and termination of the Middle Miocene Badenian salinity crisis in Central Paratethys. Sedimentary Geology, 188-189:379–396.
Peryt, D., Garecka, M., and Peryt, T.M. 2021. Foraminiferal and calcareous nannoplankton biostratigraphy of the upper Badenian-lower Sarmatian in the SE Polish Carpathian Foredeep. Geological Quarterly, 65:18.
https://doi.org/10.7306/gq.1584
Pisera, A. 1985. Paleoecology and lithogenesis of the Middle Miocene (Badenian) algal-vermetid reefs from the Roztocze Hills, south-eastern Poland. Acta Geologica Polonica, 35:89–155.
Pollard, D.A. 1984. A review of ecological studies on seagrass-fish communities, with particular reference to recent studies in Australia. Aquatic Botany, 18:3–42.
https://doi.org/10.1016/0304-3770(84)90079-2
Popov, S.V., Rögl, F., Rozanov, A.Y., Steininger, F.F., Shcherba, I.G., and Kovać, M. (eds.). 2004. Lithological-paleogeographic maps of Paratethys. 10 maps late Eocene to Pliocene. Courier Forschungs-Institut Senckenberg, 250:1–46.
Procházka, V.J. 1900. Das ostböhmische Miocaen. Archiv für die naturwissenschaftliche Durchforschung Böhmens, 10:77–86
Radwańska, U. 1992. Fish otoliths in the middle Miocene (Badenian) deposits of southern Poland. Acta Geologica Polonica, 42:141–328.
Rafinesque, C.S. 1810. Caratteri di alcuni nuovi generi e nuove specie di animali e piante della sicilia, con varie osservazioni sopra i medisimi. Per le stampe di Sanfilippo, Palermo.
Regan, C.T. 1909. Descriptions of new marine fishes from Australia and the Pacific. Annals and Magazine of Natural History, 4:438–440.
Regan, C.T. 1910. The origin and evolution of the teleostean fishes of the order Heterosomata. Annals and Magazine of Natural History, 6:484–496.
Regan, C.T. 1912. The classification of the blennioid fishes. Annals and Magazine of Natural History, Series 8, 10:265–280.
Reichenbacher, B. 1993. Mikrofaunen, Paläogeographie und Biostratigraphie der miozänen Brack- und Süßwassermolasse in der westlichen Paratethys unter besonderer Berücksichtigung der Fisch-Otolithen. Senckenbergiana lethaea, 73: 277–374.
Reichenbacher, B. 1996. Continental European fish faunas (otoliths) at the Oligocene/Miocene boundary. Giornale di Geologia, 58:171–175.
Reichenbacher, B. and Weidmann, M. 1992. Fisch-Otolithen aus der oligo-/miozänen Molasse der West-Schweiz und der haute-Savoie (Frankreich). Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie), 184:1–83.
Reichenbacher, B. and Gaudant, J. 2003. On Prolebias meyeri (Agassiz) (Teleostei, Cyprinodontiformes) from the Oligo-Miocene of the Upper Rhinegraben area, with the establishment of a new genus and a new species. Eclogae Geologicae Helvetiae, 96:509–520.
https://doi.org/10.1007/s00015-003-1098-x
Reichenbacher, B. and Prieto, J. 2006. Lacustrine fish faunas (Teleostei) from the Karpatian of the northern Alpine Molasse Basin, with a description of two new species of Prolebias Sauvage. Palaeontographica Abteilung A, 278:87–95.
https://doi.org/10.1127/pala/278/2006/87
Reichenbacher, B., Sienknecht, U., Küchenhoff, H., and Fenske, N. 2007. Combined Otolith Morphology and Morphometry for Assessing Taxonomy and Diversity in Fossil and Extant Killifish (Aphanius, † Prolebias). Journal of Morphology, 268:898–915.
https://doi.org/10.1002/jmor.10561
Reichenbacher, B. and Kowalke, T. 2009. Neogene and present-day zoogeography of killifishes (Aphanius and Aphanolebias) in the Mediterranean and Paratethys areas. Palaeogeography, Palaeoclimatology, Palaeoecology, 281:43–56.
https://doi.org/10.1016/j.palaeo.2009.07.008
Reichenbacher, B., Feulner, G.R., and Schulz-Mirbach, T. 2009. Geographic variation in otolith morphology among freshwater populations of Aphanius dispar (Teleostei, Cyprinodontiformes) from the southeastern Arabian peninsula. Journal of Morphology, 270:469–484.
https://doi.org/10.1002/jmor.10702
Reichenbacher, B., Filipescu, S., and Miclea, A. 2019. A unique middle Miocene (Sarmatian) fish fauna from coastal deposits in the eastern Pannonian Basin (Romania). Palaeobiodiversity and Palaeoenvironments, 99:177–194.
https://doi.org/10.1007/s12549-018-0334-3
Reichenbacher, B. and Bannikov, A.F. 2022. Diversity of gobioid fishes in the late middle Miocene of northern Moldova, Eastern Paratethys - Part II: description of † Moldavigobius helenae gen. et sp. nov. PalZ, 97:365–381.
https://doi.org/10.1007/s12542-022-00639-1
Rivaton, J. and Bourret, P. 1999. Les otolithes des poissons de l’Indo-Pacifique. IRD. Doc. Sci. Tech.
Rögl, F. 1998. Palaeogeographic Considerations for Mediterranean and Paratethys Seaways (Oligocene to Miocene). Annalen des Naturhistorischen Museums in Wien, A, 99:279–310.
Rüppell, W.P.E.S. 1835. Neue Wirbelthiere zu der Fauna von Abyssinien gehörig. Fische des Rothen Meeres. Frankfurt-am-Main.
Sant, K., Palcu, D.V., Turco, E., Di Stefano, A., Baldassini, N., Kouwenhoven, T., Kuiper, K.F., and Krijgsman W. 2019. The mid-Langhian flooding in the eastern Central Paratethys: integrated stratigraphic data from the Transylvanian Basin and SE Carpathian Foredeep. International Journal of Earth Science, 108:2209–2232.
https://doi.org/10.1007/s00531-019-01757-z
Sant, K., Kuiper, K.F., Rybár, S., Grunert, P., Harzhauser, M., Mandic, O., Jamrich, M., Šarinová, K., Hudáčková, N., and Krijgsman, W. 2020. 40Ar/39Ar geochronology using high sensitivity mass spectromety: examples from middle Miocene horizons of the Central Paratethys. Geologica Carpathica, 71:166–182.
https://doi.org/10.31577/GeolCarp.71.2.5
Sauvage, H.E. 1874. Notice sur les poissons tertiaires d’Auvergne. Bulletin de la Société d’Histoire naturelle de Toulouse, 8:171–198.
Schneider, G.F. 1953. Fauna of ostracods from Miocene deposits of the western part of Ukraine. Geologicheski Sbornik VNIGRI, 2:108–109. (In Russian)
Schneider, G.F. 1959. Characteristics of the Middle Miocene ostracod fauna, p. 107–145. In Zhizhchenko, B.P. (ed.), Atlas of the Middle Miocene fauna of the North Caucasus and Crimea. Gosgeoltoptekhizdat, Moscow. (In Russian)
Schubert, R.J. 1902. Die Fischotolithen des österreichisch-ungarischen Tertiärs. I. Die Sciaeniden. Jahrbuch der Geologischen Reichanstalt, 51:301–316.
Schubert, R.J. 1905. Die Fischotolithen des österreichisch-ungarischen Tertiärs. II. Macruriden und Beryciden. Jahrbuch der kaiserlich-königlichen Geologischen Reichsanstalt, 55:613–638.
Schubert, R.J. 1906. Die Fischotolithen des österreichisch-ungarischen Tertiärs. III. Jahrbuch der kaiserlich-königlichen Geologischen Reichsanstalt, 56:623–706.
Schultz, O. 1979. Supplementary notes on elasmobranch and teleost fish remains from the Korytnica Clays (Middle Miocene; Holy Cross Mountains, Central Poland). Acta Geologica Polonica, 29:287–293.
Schultz, O. 1998. Tertiärfossilien Österreichs. Wirbellose, niedere Wirbeltiere und marine Säugetiere. Schöne, interessante, häufige und wichtige Makrofossilien aus dem Naturhistorischen Museum und Privatsammlungen. Goldschneck-Verlag, Korb.
Schultz, O. 2013. Pisces, p. 1–576. In Piller, W. (ed.), Catalogus Fossilium Austriae, 3. Verlag der Österreichischen Akademie der Wissenschaften, Wien.
Schultz, O. and Bellwood, D.R. 2004. Trigonodon oweni and Asima jugleri are different parts of the same species Trigonodon jugleri, a Chiseltooth Wrasse from the Lower and Middle Miocene in Central Europe (Osteichthyes, Labridae, Trigonodontinae). Annalen des Naturhistorischen Museums in Wien, 105A:287–305.
Schultz, O., Brzobohatý, R., and Kroupa, O. 2010. Fish teeth from the Middle Miocene of Kienberg at Mikulov, Czech Republic, Vienna Basin. Annalen des Naturhistorischen Museums in Wien A, 112:489–506.
Schwarzhans, W. 1978. Otolith-morphology and its usage for higher systematical units with special reference to the Myctophiformes s.l. Mededelingen van de Werkgroep voor Tertiaire en Kwartaire Geologie, 15:167–185.
Schwarzhans, W. 2010. The otoliths from the Miocene of the North Sea Basin. Backhuys Publishers, Leiden & Margraf Publishers, Leiden.
Schwarzhans, W. 2014. Otoliths from the middle Miocene (Serravallian) of the Karaman Basin, Turkey. Cainozoic Research, 14:35–69.
Schwarzhans W. 2017. A review of otoliths collected by W. Weiler from the Badenian of Romania and by B. Strashimirov from Badenian equivalents of Bulgaria. Cainozoic Research, 17:167–191.
Schwarzhans, W., Ahnelt, H., Carnevale, G., Japundžić, S., Bradić, K., and Bratishko, A. 2017a. Otoliths in situ from Sarmatian (middle Miocene) fishes of the Paratethys. Part III: tales from the cradle of the Ponto-Caspian gobies. Swiss Journal of Palaeontology, 136:45–92.
https://doi.org/10.1007/s13358-016-0120-7
Schwarzhans, W., Carnevale, G., Japundžić, S., and Bradić-Milinović, K. 2017b. Otoliths in situ from Sarmatian (middle Miocene) fishes of the Paratethys. Part IV: Scorpaenidae, Labridae, and Gobiesocidae. Swiss Journal of Palaeontology, 136:93–108.
https://doi.org/10.1007/s13358-017-0124-y
Schwarzhans, W., Carnevale, G., Japundžić, S., and Bradić-Milinović, K. 2017c. Otoliths in situ from Sarmatian (middle Miocene) fishes of the Paratethys. Part V: Bothidae and Soleidae. Swiss Journal of Palaeontology, 136:109–128.
https://doi.org/10.1007/s13358-017-0128-7
Schwarzhans, W., Brzobohatý, R., and Radwańska, U. 2020a. Goby otoliths from the Badenian (middle Miocene) of the Central Paratethys from the Czech Republic, Slovakia and Poland: A baseline for the evolution of the European Gobiidae (Gobiiformes; Teleostei). Bollettino della Società Palaeontologica Italiana, 59:125–173.
https://doi.org/10.4435/BSPI.2020.10
Schwarzhans, W., Agiadi, K., and Carnevale, G. 2020b. Late Miocene-early Pliocene evolution of Mediterranean gobies and their environmental and biogeographic significance. Rivista Italiana di Paleontologia e Stratigrafia, 126:657–724.
Schwarzhans, W. and Kovalchuk, O. 2022. New data on fish otoliths from the late Badenian (Langhian, Middle Miocene) back reef environment in the Carpathian Foredeep (Horodok, western Ukraine). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 303:317–326.
https://doi.org/10.1127/njgpa/2022/1051
Schwarzhans, W., Klots, O., Ryabokon, T., and Kovalchuk, O. 2022. A rare window into a back-reef fish community from the middle Miocene (late Badenian) Medobory Hills barrier reef in western Ukraine, reconstructed mostly by means of otoliths. Swiss Journal of Palaeontology, 141:18.
https://doi.org/10.1186/s13358-022-00261-3
Schwarzhans, W. and Radwańska, U. 2022. A review of lanternfish otoliths (Myctophidae, Teleostei) of the early Badenian (Langhian, middle Miocene) from Bęczyn, southern Poland. Cainozoic Research, 22:9–24.
Schwarzhans, W., Bratishko, A., and Vernyhorova, Y.V. 2023. Approaching the Khersonian Crisis: Fish otoliths from the upper Bessarabian (middle Sarmatian s.l.; Late Miocene) of Jurkine (Kerch Peninsula, Crimea). Palaeontologia Electronica, 26:a31.
https://doi.org/10.26879/1300
Schwarzhans, W. and Carnevale, G. 2024. Fish otoliths from the upper Oligocene and Lower Miocene of the Monferrato and Turin Hills, northern Italy. Rivista Italiana di Paleontologia e Stratigrafia.
https://doi.org/10.13130/2039-4942/14185
Scopoli, G.A. 1777. Introductio ad historiam naturalem, sistens genera lapidum, plantarum et animalium hactenus detecta, caracteribus essentialibus donata, in tribus divisa, subinde ad leges naturae. Wolfgang Gerle, Pragae.
Šegvić, B., Lukács, R., Mandic, O., Strauss, P., Badurina, L., Guillong, M., and Harzhauser, M. 2023. U-Pb zircon age and mineralogy of St. Georgen halloysite tuff shed light on the timing of the middle Badenian transgression, ash dispersal and paleoenvironmental conditions in the central Vienna Basin, Austria. Journal of the Geological Society, 180:jgs2022-106.
https://doi.org/10.1144/jgs2022-106
Smale, M.J., Watson, G., and Hecht, T. 1995. Otolith atlas of southern African marine fishes. J.L.B. Smith Institute of Ichthyology, Ichthyological Monographs, 1:1–402.
Śmigielska, T. 1966. Otoliths of fishes from the Tortonian of Southern Poland. Annales de la Société Géologique de Pologne, 36:205–275.
Stancheva, M. 1962. Ostracodna fauna ot neogena v severozapadna Bulgaria. I. tortonskii ostracodi. Travaux sur la Géologie de Bulgaria, Paléontologie, 4:5–75.
Stancheva, M. 1963. Ostracodna fauna ot neogena v severozapadna Bulgaria. II. sarmatskii ostracodi. Travaux sur la Géologie de Bulgaria, Paléontologie, 5:1–75.
Stanley, S.M. 2014. Evolutionary radiation of shallow-water Lucinidae (Bivalvia with endosymbionts) as a result of the rise of seagrasses and mangroves. Geology, 42:803–806.
https://doi.org/10.1130/G35942.1
Strashimirov, B. 1972. Otolity ot tarkhana na Severoistochna Bulgaria. Annuaire de l’Ecole Supérieure des Mines et de Geologie, Sofia, 18:301–313.
Swainson, W. 1839. On the natural history and classification of fishes, amphibians, & reptiles, or monocardian animals. Spottiswoode & Co., London.
Szabó, M. and Kocsis, L. 2020. Supplementary data on the Middle Miocene (Badenian) fish assemblage of Nyirád (Hungary): revision and new results on faunal composition and paleoenvironment. Palaeontographica, Abteilung A: Palaeozoology - Stratigraphy, 315:121–191.
https://doi.org/10.1127/pala/2020/0094
Szabó, M., Kocsis, L., Bosnakoff, M., and Sebe, K. 2021. A diverse Miocene fish assemblage (Chondrichthyes and Osteichthyes) from the Pécs-Danitzpuszta sand pit (Mecsek Mts, Hungary). Földtani Közlöny, 33:31–64.
https://doi.org/10.17111/FragmPalHung.2016.33.31
Szczechura, J. 2006. Middle Miocene (Badenian) ostracods and green algae (Chlorophyta) from Kamienica Nawojowska, Nowy Sacz Basin (Western Carpathians, Poland). Geologica Carpathica, 57:103–122.
Szuromi-Korecz, A. and Szegő, É. 2001. Data for knowledge of foraminifera and ostracoda microfauna of Kovácsszénája (SW-Hungary). Folia Komloensis, 10:51–74.
Tanaka, S. 1908. Notes on some Japanese fishes, with descriptions of fourteen new species. Journal of the College of Science. Imperial University Tokyo, 23:1–54.
Thacker, C.E. 2009. Phylogeny of Gobioidei and placement within Acanthomorpha, with a new classification and investigation of diversification and character evolution. Copeia, 2009:93–104.
https://doi.org/10.1643/ci-08-004
Thornton, H.G. 1953. Sergei Nicholaevitch Winogradsky, 1856-1953. Obituary Notices of Fellows of the Royal Society, 8:635–626.
https://doi.org/10.1098/rsbm.1953.0022
Tóth, E. 2008. Sarmatian (Middle Miocene) ostracod fauna from the Zsámbék Basin, Hungary. Geologica Pannonica, 36:101–151.
Tunoğlu, C. and Bilen, C. 2001. Burdigalian-Langhian (Miocene) ostracod biostratigraphy and chronostratigraphy of the Kasaba Basin (Kaş/Antalya), SW Turkey. Geologica Carpathica, 52:247–258.
Van der Heide, T., Govers, L.L., de Fouw, J., Olff, H., van der Geest, M., van Katwijk, M.M., Piersma, T., van de Koppel, J., Silliman, B.R., Smolders, A.J.P., and van Gils, J.A. 2012. A three-stage symbiosis forms the foundation of seagrass ecosystems. Science, 336:1432–1434.
https://doi.org/10.1126/science.1219973
Van Hinsbergh, V. and Hoedemakers, K. 2022. Zanclean and Piacnezian otolith-based fish faunas of Estepona (Málaga, Spain). Cainozoic Research, 22:241–352.
Vernyhorova, Y. 2018. Biostratigraphy of the Konkian (Middle Miocene of the Eastern Paratethys) deposits of Southern Ukraine based on foraminifera. Geologia Croatica, 71:135–146.
Vernyhorova, Y.V., Holcová, K., Doláková, N., Reichenbacher, B., Scheiner, F., Ackerman, L., Rejšek, J., De Bortoli, L., Trubač, J., and Utescher, T. 2023. The Miocene Climatic Optimum at the interface of epicontinental sea and large continent: A case study from the Middle Miocene of the Eastern Paratethys. Marine Micropaleontology, 181:102231.
https://doi.org/10.1016/j.marmicro.2023.102231
Vinciguerra, D. 1883. Risultati ittiologici delle crociere del Violante. Annali del Museo Civico di Storia Naturale di Genova, 18:465–590.
Weiler, W. 1942. Die Otolithen des rheinischen und nordwestdeutschen Tertiärs. Abhandlungen des Reichsamts für Bodenforschung, Neu Folge, 206:9–140.
Weiler W. 1943. Die Otolithen aus dem Jungtertiär Süd-Rumäniens, I. Buglow und Sarmat. Senckenbergiana Lethaea, 26:87–115.
Weinfurter, E. 1950. Die oberpannonische Fischfauna vom Eichkogel bei Mödling Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften mathematisch naturwissenschaftlichen Klasse, Abteilung 1, 159:37–50.
Whitley, G.P. 1950. New fish names. Proceedings of the Royal Zoological Society of New South Wales, volume for 1948-49:44.
Zorn, I. 1998. Ostracoda aus dem Karpat (Unter-Miozän) des Korneuburger Beckens (Niederösterreich). Beiträge zur Paläontologie, 23:175–271.
Zorn, I. 2003. Ostracods of the Karpatian, p. 236-240. In Brzobohatý, R., Cicha, I., Kovač, M., and Rögl, F. (eds.), The Karpatian. A Lower Miocene stage of the Central Paratethys. Masaryk University, Brno.
Zorn, I. 2004. Ostracoda from the Lower Badenian (Middle Miocene) Grund Formation (Molasse Bazin, Lower Austria). Geologica Carpathica, 55:179–189.
