

 Two exceptional Balaenomorpha (Cetacea: Mysticeti) from the Biemenhorst Subformation (middle/late Miocene) of Bocholt (W Münsterland, Germany) with a critical appraisal on the anatomy of the periotic bone
Two exceptional Balaenomorpha (Cetacea: Mysticeti) from the Biemenhorst Subformation (middle/late Miocene) of Bocholt (W Münsterland, Germany) with a critical appraisal on the anatomy of the periotic bone
Article number: 26.3.a37
https://doi.org/10.26879/1268
Copyright Society of Vertebrate Paleontology, September 2023
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 13 January 2023. Acceptance: 8 September 2023.
ABSTRACT
Here we present two new Miocene basal balaenomorph mysticetes recognized by periotic bones with a unique combination of characters. The fossil periotics originated from former shallow marine sediments and were discovered in a suburban quarry of Bocholt in the German Münsterland holding a number of remains of a multispecies community of mysticetes. A detailed accompanying analysis of the internal and external structures of mysticete periotics resulted in a critical review and revision of two important characteristics of the pars cochlearis. The partition between the openings for the facial nerve and the vestibulocochlear nerve, named here ‘bony septum’ for baleen whales, has been developed so prominently during mysticete evolution that the crista transversa is no longer recognizable in derived forms, as it lies deep inside the inner ear canal and within the fundus of the internal acoustic meatus where it separates the cochlear nerve from the inferior vestibular nerve. We show that the designation of the opening for the superior vestibular nerve as ‘foramen singulare’ is misleading since this term describes the opening for the nerve to the posterior semicircular canal within the fundus of the internal acoustic meatus.
Indira S. Ritsche. Bezirksamt Lichtenberg von Berlin, Rathaus Lichtenberg, Möllendorfstraße 6, D-10367 Berlin, Germany. puddelruss@gmx.de
Oliver Hampe. Museum für Naturkunde, Leibniz-Institut für Evolutions- und Biodiversitätsforschung, Invalidenstraße 43, D-10115 Berlin, and Institut für Geologische Wissenschaften, Fachrichtung Paläontologie, Freie Universität Berlin, Malteserstr. 74-100, 12249 Berlin, Germany. Corresponding author. oliver.hampe@mfn.berlin
Keywords: Diorocetidae; Breda Formation; IAM; bony septum; crista transversa
Final citation: Ritsche, Indira S., and Hampe, Oliver. 2023. Two exceptional Balaenomorpha (Cetacea: Mysticeti) from the Biemenhorst Subformation (middle/late Miocene) of Bocholt (W Münsterland, Germany) with a critical appraisal on the anatomy of the periotic bone. Palaeontologia Electronica, 26(3):a37.
https://doi.org/10.26879/1268
palaeo-electronica.org/content/2023/3958-two-exceptional-balaenomorpha
Copyright: September 2023 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
It is broadly accepted that the periotics has been an important source for significant valid characters in taxonomic and phylogenetic studies of Mysticeti (Steeman, 2007; 2010; Hampe and Ritsche, 2011; Ekdale et al., 2011; Marx and Fordyce, 2015). Many periotic characters are used for phylogenetic analysis in baleen whales, but most important is the character selection because of the dramatic morphological change of the periotic bone during ontogeny, for example, recognized in the living blue whale (Balaenoptera musculus) and minke whale (B. acutorostrata). In consequence, one should expect this in fossil mysticetes, too, and for some species it is already documented, for example, Herpetocetus morrowi (El Adli et al., 2014) and Waharoa ruwhenua (Boessenecker and Fordyce, 2015a). It must keep in mind that there are often only one or two specimens per species existing and therefore lacking a complete impression of intraspecific variability. However, the morphology of the periotic bone is highly diagnostic for individual species of baleen whales and in some cases also for mysticete clades, like Eomysticetidae possessing a discontinuous and low superior process of the periotic with anterior and posterior apices (Boessenecker and Fordyce, 2015a, b) or Cetotheriidae possessing a high ‘crista transversa’ reaching the cerebral surface of the pars cochlearis, internal opening for the vestibulocochlear nerve subequal to the internal opening for the facial nerve, and a posterior process well exposed on the lateral wall of the skull (El Adli et al., 2014). In consideration of all these aspects, the periotic bone is an important diagnostic tool for taxonomic analysis of fossil and modern baleen whales but limited for a phylogenetic debate. We would, therefore, like to utilise this work, above all, to revisit and clarify the internal anatomy of the periotic in the details of the fundus of the internal acoustic meatus.
 In December 1924, fossil baleen whale remains were donated to the Museum für Naturkunde in Berlin to the attention of Josef Felix Pompeckj (1867-1930), Professor and Director of the Geological-Palaeontological Institute and Museum of the Friedrich-Wilhelms-Universität (today: Humboldt-Universität zu Berlin) for scientific research. The scattered findings from a locality near Bocholt in the German Münsterland comprise at least 13 isolated periotic bones and about 20 tympanic bullae or bullar fragments, more than 30 vertebrae including atlases and axes, one scapula associated with a lower jaw, and several phalanges. Furthermore, some postcranial bones of a toothed whale, possibly a physeteroid, were documented in this bundle. Until 1942 (according to the remained correspondence), the owner, Mr. Paul Mauersberger, has repeatedly left remains of cetaceans from outcrops of the locality Bocholt-Biemenhorst to the Museum für Naturkunde Berlin for scientific research. The complete material was archived as ‘Collection Paul Mauersberger (Halberstadt)’. The town of Bocholt belongs politically and culturally to western Münsterland, but in terms of landscape it is already part of the Lower Rhine Lowlands (Figure 1).
In December 1924, fossil baleen whale remains were donated to the Museum für Naturkunde in Berlin to the attention of Josef Felix Pompeckj (1867-1930), Professor and Director of the Geological-Palaeontological Institute and Museum of the Friedrich-Wilhelms-Universität (today: Humboldt-Universität zu Berlin) for scientific research. The scattered findings from a locality near Bocholt in the German Münsterland comprise at least 13 isolated periotic bones and about 20 tympanic bullae or bullar fragments, more than 30 vertebrae including atlases and axes, one scapula associated with a lower jaw, and several phalanges. Furthermore, some postcranial bones of a toothed whale, possibly a physeteroid, were documented in this bundle. Until 1942 (according to the remained correspondence), the owner, Mr. Paul Mauersberger, has repeatedly left remains of cetaceans from outcrops of the locality Bocholt-Biemenhorst to the Museum für Naturkunde Berlin for scientific research. The complete material was archived as ‘Collection Paul Mauersberger (Halberstadt)’. The town of Bocholt belongs politically and culturally to western Münsterland, but in terms of landscape it is already part of the Lower Rhine Lowlands (Figure 1).
We present two new basal Balaenomorpha recognized by periotic bones found in the collection just introduced showing characters or character combinations never seen before in known mysticetes. The exact points where the fossil periotics were found could not be determined through the obtained correspondence but a stratigraphic correlation with the Biemenhorst Subformation was possible regarding preservational condition and colour of the fossils. Since we have only single periotic bones to be described we avoid defining new taxa and leave them in open taxonomy. Consequently, another phylogenetic analysis based on these new fossils does not seem appropriate at this stage.
STRATIGRAPHIC BACKGROUND
The Biemenhorst Subformation (von der Hocht, 1988: Biemenhorst-Schichten) belongs to the upper part of the Breda Formation underlying the Uedem Subformation. The Biemenhorst member consists essentially of illite, silt, and intercalated fine sands. It is usually free of limestone with dark grey to dark olive grey colour (Jansen, 2004). The upper part of the Breda Formation is especially common in the Lower Rhine region (Hiss, 2013). For the Breda Formation, the Groote Heide borehole near Venlo was additionally defined to be the type of profile in addition to the reference borehole from the eponymous town Breda (Doppert et al., 1975).
 The present biostratigraphical age datings are based on calcareous nannoplankton, dinoflagellates, bolboforms, planktonic foraminifers, molluscs, and elasmobranch teeth. The Biemenhorst Subformation is correlated with the regional Neogene North Sea basin stage Langenfeldian (late Serravallian to early Tortonian; Figure 2), which correlates roughly with the global late middle Miocene (Laursen et al., 1998; King, 2016). The deposits describe a shallow marine environment for this former area. After a regression phase at the Palaeogene/Neogene boundary, oceanic sedimentation restarted during the Hemmorian (Burdigalian, early Miocene) in the Lower Rhine area. The fact that the Biemenhorst Subformation contains fossil marine mammals has been mentioned before. Cetaceans (e.g., Heinrichs, 1956, 1957; see also Jansen, 2004) or even remains of monachid pinnipeds (Springhorn, 1978) have been documented here from several brickwork pits.
The present biostratigraphical age datings are based on calcareous nannoplankton, dinoflagellates, bolboforms, planktonic foraminifers, molluscs, and elasmobranch teeth. The Biemenhorst Subformation is correlated with the regional Neogene North Sea basin stage Langenfeldian (late Serravallian to early Tortonian; Figure 2), which correlates roughly with the global late middle Miocene (Laursen et al., 1998; King, 2016). The deposits describe a shallow marine environment for this former area. After a regression phase at the Palaeogene/Neogene boundary, oceanic sedimentation restarted during the Hemmorian (Burdigalian, early Miocene) in the Lower Rhine area. The fact that the Biemenhorst Subformation contains fossil marine mammals has been mentioned before. Cetaceans (e.g., Heinrichs, 1956, 1957; see also Jansen, 2004) or even remains of monachid pinnipeds (Springhorn, 1978) have been documented here from several brickwork pits.
MATERIAL AND METHODS
 For external examination and documentation, a Canon PowerShot SX50 HS camera and a portable Somikon NC1727 professional photo studio box with two photo lamps (power 19.8 W, 1200 lm) was utilised providing high-resolution photographs of the periotics. Dimensions such as length, width, and distances were made using a dial vernier caliper (measuring section definitions, see Figure 3).
For external examination and documentation, a Canon PowerShot SX50 HS camera and a portable Somikon NC1727 professional photo studio box with two photo lamps (power 19.8 W, 1200 lm) was utilised providing high-resolution photographs of the periotics. Dimensions such as length, width, and distances were made using a dial vernier caliper (measuring section definitions, see Figure 3).
The tomography was done in collaboration with the Helmholtz-Zentrum Berlin für Materialien und Energie (HZB) to provide a well-designed setup and an accurate data analysis. The X-ray tomography facility at HZB is an industrial computed tomography scanner that was developed and constructed on site. Cone beam voltage is continuously adjustable from 40 to 150 kV and spatial resolution was in the range between 10 and 50 µm. A flat panel detector with a 2316 x 2316 pixel density (resolution) was used. To utilise the entire gray scale range of the detector, the exposure time was adjusted. Appropriate filter (copper or aluminium), accelerating voltage and emission current of the X-ray source were selected accordingly (see also Arlt et al., 2018). 3D image reconstruction of the CT scans was done using Octopus 8 (Inside Matters). For visualisation and analysis of the compiled data the following programs were utilised: Fiji Is Just ImageJ 1.51d (open source) for image processing and analysis of reconstructed CT data and VGStudio MAX 2.2 (Volume Graphics) for visualisation of reconstructed CT data.
Institutional abbreviations
ChM - The Charleston Museum, Charleston, South Carolina
FMNH - Field Museum of Natural History, Chicago
IRSNB - Institut royal des Sciences naturelles de Belgique, Brussels
MB - Museum für Naturkunde Berlin, Palaeontological Collection
USNM - United States National Museum, Smithsonian Institution, Washington, D.C.
ZMB - Museum für Naturkunde Berlin, Zoological Collection
SYSTEMATIC PALAEONTOLOGY
MAMMALIA Linnaeus, 1758
CETACEA Brisson, 1762
MYSTICETI Flower, 1864
CHAEOMYSTICETI Mitchell, 1989
BALAENOMORPHA Geisler and Sanders, 2003
BALAENOMORPHA ‘TYPE BOCHOLT A’
Figure 4
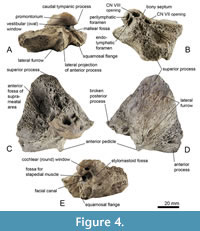 Material. MB.Ma. 51619, a right periotic bone.
Material. MB.Ma. 51619, a right periotic bone.
Occurrence. Biemenhorst Subformation, upper part of Breda Formation, Langenfeldian, late Serravallian, late middle Miocene of Bocholt-Biemenhorst, W Münsterland, North Rhine-Westphalia State, Germany.
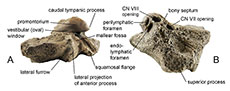
Description. The periotic of Balaenomorpha ‘type Bocholt A’ (Figure 4) is well preserved but the posterior process is broken away. In lateral view, the outline of the periotic equates almost an acute triangle (Figure 4D). Also, the anterior process is triangular in lateral view with a slightly concave anterodorsal margin and extends ventrally. The elongate anterior pedicle is prominent and far ventrally located (Figure 4C, D). The lateral projection of the anterior process is moderately developed and anterolaterally pointed (Figure 4A). There is no distinct lateral fissure between periotic body and anterior process and the depression immediately anterior to the lateral projection of the anterior process is only slightly deepened. The superior process is extraordinary elongated and pointed (Figure 4B, C), its dorsolateral edge is very thick and laterally rounded and the lateral surface is furrowed and almost flat (Figure 4D). In posterior view, the flattened and tilted pars cochlearis is dorsoventrally elongated and somewhat pointed dorsomedially (Figure 4E), especially its anterior part, and it possesses a ventrally distinct inflated promontorium (Figure 4A). The caudal tympanic process is weakly developed or even reduced. The vestibular window is partially covered by the bulged promontorium (Figure 4A). The cochlear window is slightly elongated dorsoventrally but small, and it is well separated from the perilymphatic foramen by a weakly depressed area (Figure 4B, E). Perilymphatic and endolymphatic foramina are almost equal in size and positioned parallel with a slightly shifted perilymphatic foramen (Figure 4B). Both are more slit-like, and the endolymphatic foramen is slightly elongated laterally. The openings for vestibulocochlear nerve (CN VIII) and facial nerve (CN VII) are almost equal in size and separated by a very thick and protruding bony septum (Figure 4B; for further explanations see section “Review on the internal acoustic meatus anatomy”). The CN VIII-opening is more medially oriented and circular compared to the teardrop-shaped and rather anteriorly oriented CN VII-opening. The opening for the greater petrosal nerve is located within the latter. The suprameatal area is rather small, cragged, and tapered dorsolaterally, and a distinct ridge separates suprameatal area and anterior fossa of the suprameatal area (Figure 4C). The latter is deep, broad, and extends together with the superior process dorsolaterally. The broad stylomastoid fossa is shallow and dorsolaterally expanded onto the periotic body (Figure 4E). The fossa for the stapedial muscle is broad with a well-defined outline (Figure 4E). The ventral opening for the facial nerve is large and passes on to an elongated and distinct sulcus for facial nerve, and a shallow mallear fossa is located anterolateral to the ventral opening for the facial nerve (Figure 4E). The fossa incudis is inconspicuous. The ventrolateral ridge is narrow and weakly developed and adjacent to the latter there is a small and ridge-like squamosal flange, which is separated from posterior process by a deep furrow (Figure 4A).
The dimensions are as follows (measurement ID in brackets refers to Figure 3):
(1) Maximum anteroposterior extent from stylomastoid fossa to the tip of anterior process 62 mm
(2) Anteroposterior distance of pars cochlearis excluding caudal tympanic process 30 mm
(3) Dorsoventral distance of pars cochlearis from the tip of the internal acoustic meatus (IAM) to most bulged point of promontorium 34 mm
(4) Dorsoventral distance of periotic body from ventrolateral ridge to the tip of superior process 59 mm
(5) Thickness of periotic body: maximum medial-lateral extent 42 mm
(6) Anteroposterior distance of anterior process beginning from ventral opening of facial nerve 49 mm
(7) Anteroposterior distance of anterior process beginning from the anterior edge of pars cochlearis 37 mm
(8) Dorsoventral distance of anterior process: measured at the fissure between periotic body and anterior process and included lateral projection of anterior process 64 mm
(9) Thickness of base of anterior process at the connection to periotic body 32 mm
(10) Length of internal acoustic meatus (IAM) from anterior to posterior edge 23 mm
(11) Width of IAM superior to inferior edge 9 mm
(12) Width of fossa for stapedial muscle 7 mm
(13) Distance between cochlear window and vestibular window 8 mm
(14) Distance between cochlear window and perilymphatic foramen 8 mm
(15) Cross-section of cochlear window: dorsal to ventral 4 mm
(16) Cross-section of cochlear window: medial to lateral 2 mm
BALAENOMORPHA ‘TYPE BOCHOLT B’
Figure 5, Figure 6
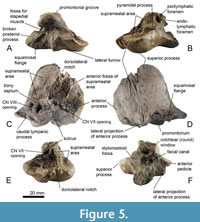 Material. MB.Ma. 51618, a left periotic bone and MB.Ma. 51621, a right periotic bone.
Material. MB.Ma. 51618, a left periotic bone and MB.Ma. 51621, a right periotic bone.
Occurrence. Biemenhorst Subformation, upper part of Breda Formation, Langenfeldian, late Serravallian, late middle Miocene of Bocholt-Biemenhorst, W Münsterland, North Rhine-Westphalia State, Germany.
Description. The first periotic of Balaenomorpha ‘type Bocholt B’ (MB.Ma. 51618; Figure 5) is well preserved but without the broken posterior process. The referred specimen, a right periotic (MB.Ma. 51621; Figure 6), lacks both the anterior and posterior processes. In consideration of the broken anterior process, both periotic bones may belong to one individual due to its almost identical morphology and size. In lateral view, the outline of the left periotic bone is roughly triangular even though the anterodorsal margin of the anterior process is well rounded (Figure 5D). The ventrally extended anterior process possesses a prominent, elongate anterior pedicle. The lateral projection of anterior process is well developed and tapered laterally (Figure 5D, Figure 6D). There is no distinct lateral fissure between periotic body and anterior process, thus the latter is almost completely fused with the periotic body. The lateral surface of the periotic body is smooth with few slight furrows (Figure 5D). The caudal tympanic process is weakly developed or even reduced (Figure 5C). In posterior view, the flattened and tilted pars cochlearis occurs elevated and squared with a ventrally inflated promontorium (Figure 5F, Figure 6D).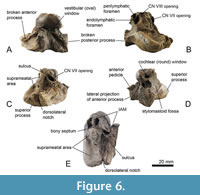 The cochlear window is dorsoventrally elongated and diamond-shaped but small and well separated from the perilymphatic foramen by an uneven area (Figure 5F, Figure 6D). Perilymphatic and endolymphatic foramina are positioned almost parallel to each other with a slightly shifted perilymphatic foramen (Figure 5B, Figure 6B). The latter is small and oval compared to a more slit-like and laterally elongated endolymphatic foramen. The openings for the vestibulocochlear nerve (CN VIII) and facial nerve (CN VII) are almost circular and separated by a thick and protruding bony septum, which passes on to a distinct sulcus (Figure 5C, Figure 6B). The CN VIII-opening is large with a sharp edge dorsolaterally, and it is more medially oriented. The CN VII-opening is smaller than the CN VIII-opening and more anteriorly oriented (Figure 6B). The opening for the greater petrosal nerve is located within the opening for the facial nerve. Anterolateral to the endolymphatic foramen, a prominent pyramidal process is developed (Figure 5B). The superior process is elongated and tongue-shaped with a slightly concave and fine-pored suprameatal area. The latter is flanked anteriorly by a porous, uneven, and shallow anterior fossa of suprameatal area (Figure 5E, F, Figure 6C, D). The cragged dorsolateral edge of the superior process is well developed and laterally rounded with a distinct dorsolateral notch (Figure 5E, Figure 6E). The stylomastoid fossa is shallow and dorsolaterally expanded onto the periotic body, and it is separated to perilymphatic and endolymphatic foramina by a thin and fragile brink (Figure 5F). The fossa for the stapedial muscle is here also broad with a well-defined outline (Figure 5A). The vestibular window is small; the right periotic still contains the largely obscured stapes in situ (Figure 6A). The ventral opening for the facial nerve is large and passes on to an elongated and distinct sulcus for the facial nerve (Figure 5F). Fossa incudis and mallear fossa are inconspicuous. The ventrolateral ridge is narrow and moderately developed and adjacent to the latter there is a distinct and bulging squamosal flange, which is separated from posterior process by a shallow furrow (Figure 5A).
The cochlear window is dorsoventrally elongated and diamond-shaped but small and well separated from the perilymphatic foramen by an uneven area (Figure 5F, Figure 6D). Perilymphatic and endolymphatic foramina are positioned almost parallel to each other with a slightly shifted perilymphatic foramen (Figure 5B, Figure 6B). The latter is small and oval compared to a more slit-like and laterally elongated endolymphatic foramen. The openings for the vestibulocochlear nerve (CN VIII) and facial nerve (CN VII) are almost circular and separated by a thick and protruding bony septum, which passes on to a distinct sulcus (Figure 5C, Figure 6B). The CN VIII-opening is large with a sharp edge dorsolaterally, and it is more medially oriented. The CN VII-opening is smaller than the CN VIII-opening and more anteriorly oriented (Figure 6B). The opening for the greater petrosal nerve is located within the opening for the facial nerve. Anterolateral to the endolymphatic foramen, a prominent pyramidal process is developed (Figure 5B). The superior process is elongated and tongue-shaped with a slightly concave and fine-pored suprameatal area. The latter is flanked anteriorly by a porous, uneven, and shallow anterior fossa of suprameatal area (Figure 5E, F, Figure 6C, D). The cragged dorsolateral edge of the superior process is well developed and laterally rounded with a distinct dorsolateral notch (Figure 5E, Figure 6E). The stylomastoid fossa is shallow and dorsolaterally expanded onto the periotic body, and it is separated to perilymphatic and endolymphatic foramina by a thin and fragile brink (Figure 5F). The fossa for the stapedial muscle is here also broad with a well-defined outline (Figure 5A). The vestibular window is small; the right periotic still contains the largely obscured stapes in situ (Figure 6A). The ventral opening for the facial nerve is large and passes on to an elongated and distinct sulcus for the facial nerve (Figure 5F). Fossa incudis and mallear fossa are inconspicuous. The ventrolateral ridge is narrow and moderately developed and adjacent to the latter there is a distinct and bulging squamosal flange, which is separated from posterior process by a shallow furrow (Figure 5A).
The dimensions are as follows:
Left periotic, MB, Ma. 51618
(1) Maximum anteroposterior extent from stylomastoid fossa to the tip of anterior process 53 mm
(2) Anteroposterior distance of pars cochlearis excluding caudal tympanic process 27 mm
(3) Dorsoventral distance of pars cochlearis from the tip of the IAM to most bulged point of promontorium 29 mm
(4) Dorsoventral distance of periotic body from ventrolateral ridge to the tip of superior process 52 mm
(5) Thickness of periotic body: maximum medial-lateral extent 43 mm
(6) Anteroposterior distance of anterior process beginning from ventral opening of facial nerve 41 mm
(7) Anteroposterior distance of anterior process beginning from the anterior edge of pars cochlearis 24 mm
(8) Dorsoventral distance of anterior process: measured at the fissure between periotic body and anterior process and included lateral projection of anterior process 51 mm
(9) Thickness of base of anterior process at the connection to periotic body 24 mm
(10) Length of IAM from anterior to posterior edge 23 mm
(11) Width of IAM superior to inferior edge 9 mm
(12) Width of fossa for stapedial muscle 6 mm
(13) Distance between cochlear window and vestibular window 7 mm
(14) Distance between cochlear window and perilymphatic foramen 8 mm
(15) Cross-section of cochlear window: dorsal to ventral 4 mm
(16) Cross-section of cochlear window: medial to lateral 2 mm
Right periotic, MB, Ma. 51621
(1) Maximum anteroposterior extent from stylomastoid fossa to the tip of anterior process not applicable
(2) Anteroposterior distance of pars cochlearis excluding caudal tympanic process 27 mm
(3) Dorsoventral distance of pars cochlearis from the tip of the IAM to most bulged point of promontorium 27 mm
(4) Dorsoventral distance of periotic body from ventrolateral ridge to the tip of superior process 50 mm
(5) Thickness of periotic body: maximum medial-lateral extent 41 mm
(6) Anteroposterior distance of anterior process beginning from ventral opening of facial nerve not applicable
(7) Anteroposterior distance of anterior process beginning from the anterior edge of pars cochlearis not applicable
(8) Dorsoventral distance of anterior process: measured at the fissure between periotic body and anterior process and included lateral projection of anterior process 49 mm
(9) Thickness of base of anterior process at the connection to periotic body 23 mm
(10) Length of IAM from anterior to posterior edge 23 mm
(11) Width of IAM superior to inferior edge 8 mm
(12) Width of fossa for stapedial muscle 6 mm
(13) Distance between cochlear window and vestibular window 8 mm
(14) Distance between cochlear window and perilymphatic foramen 8 mm
(15) Cross-section of cochlear window: dorsal to ventral 4 mm
(16) Cross-section of cochlear window: medial to lateral 2 mm
DISCUSSION
Balaenomorpha ‘type Bocholt A’ shares with the Diorocetidae (for purposes of clarity in the following comparisons, here strictly after diagnosis of Steeman, 2007: p. 886, 2010) a flattened and tilted pars cochlearis with a ventrally bulged promontorium but distinctly differs from all of them by an extraordinary elongated and pointed superior process. Also, in Balaenomorpha ‘type Bocholt A’, the pars cochlearis, and especially its anterior part, is more dorsoventrally elongated and somewhat pointed dorsomedially in comparison to all other Diorocetidae. Balaenomorpha ‘type Bocholt A’ further differs from Diorocetidae except Amphicetus later from the lower Miocene of Belgium and Balaenomorpha ‘type Bocholt B’ in possessing a more medially oriented opening for the vestibulocochlear nerve, and it features as well as Balaenomorpha ‘type Bocholt B’ a more anteriorly oriented opening for the facial nerve. These significant morphological differences, especially a dorsoventrally elongated and dorsomedially pointed pars cochlearis, an extraordinary elongated and pointed superior process, and a small and cragged suprameatal area with an adjacent deep and broad anterior fossa of suprameatal area, which extends together with the superior process dorsolaterally, make the discovered periotic of Balaenomorpha ‘type Bocholt A’ unique.
It must be mentioned at this point that the family Diorocetidae, erected by Steeman (2007), is doubted by many authors, among others, by Marx (2011) and Bisconti et al. (2013), and shows at least no support in younger comprehensive phylogenetic analyses (e.g., Marx and Fordyce, 2015; Bisconti et al., 2022; Boessenecker et al., 2023) and are considered as a paraphyletic group within the basal balaenomorphs respectively thalassotherians.
Balaenomorpha ‘type Bocholt B’ shares with the Diorocetidae the flattened, tilted, and ventrally bulged pars cochlearis but differs from all of them by a tongue-shaped, elongated superior process with a distinct dorsolateral notch. Additionally, the pars cochlearis occurs somewhat elevated and squared in contrast to Diorocetidae. The periotics of Balaenomorpha ‘type Bocholt B’ further differs from the Diorocetidae except Amphicetus later and Balaenomorpha ‘type Bocholt A’ in possessing a more medially oriented opening for the vestibulocochlear nerve, and Balaenomorpha ‘type Bocholt B’ has a more anteriorly oriented opening for facial nerve. Both openings are well separated by a distinct sulcus in the Balaenomorpha ‘type Bocholt B’ periotics. The significant tongue-shaped superior process with a distinct dorsolateral notch and a somewhat elevated and squared pars cochlearis with distinctly separated openings for vestibulocochlear nerve (CVIII) and facial nerve (CVII) are new and could not be seen in any other mysticete periotic.
Diorocetus hiatus from the Langhian Calvert Formation of Maryland differs from Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’ in having a regular shaped edge which encircles the large, ovoidal and deepened internal acoustic meatus. The openings for CVII and CVIII are equal in size and separated by a slightly thickened and recessed bony septum (Kellogg, 1968a). Diorocetus sp. ‘type Freetz’ from the Serravallian of Lower Saxony, Germany, shares with Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’ an elongated superior process, which is less prominent and has a more rounded dorsolateral margin in dorsal view (Hampe and Ritsche, 2011). Because of the abraded endocranial aspect of the pars cochlearis, no valid evaluation is possible with the Bocholt specimens.
Thinocetus arthritus from the middle Miocene Choptank Formation of the Atlantic east coast (Virginia) differs from Balaenomorpha ‘type Bocholt B’ by following characters: the morphology of dorsal surface, especially the IAM with the openings for CNVII and CNVIII separated by a thick bony septum, and a suprameatal area, which is pitted and ornamented with short bony spicules (Kellogg, 1969). But it has the presence of a thick bony septum with Balaenomorpha ‘type Bocholt A’ and Amphicetus in common.
Amphicetus later (IRSNB 575) differs from Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’ in having a more massive periotic body and a rather slightly compressed pars cochlearis (Steeman, 2010).
Parietobalaena (including here P. palmeri from the middle Miocene of Maryland and P. campiniana from the middle Miocene of Belgium and the Netherlands) differs from Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’ in the triangle-shaped pars cochlearis in dorsal view with a concave dorsal surface, a distinctly reduced anterior process, and a convex lateral surface of periotic body (Kellogg, 1924, 1968b; Steeman, 2007, 2010; Bisconti et al., 2013).
The Cetotheriidae (here considered Brandtocetus chongulek from the lower Tortonian of Crimea, Piscobalaena nana from the upper Tortonian to Messinian of Peru, Metopocetus durinasus probably from the upper Miocene St. Marys Formation of Virginia, Nannocetus eremus from the upper Miocene of California, Herpetocetus transatlanticus from the lower Pliocene of North Carolina, and H. morrowi from the upper Pliocene of California) differ from Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’ in having a flat suprametal and a reduced superior process of the periotic body (see e.g., Bouetel and Muizon, 2006; Steeman, 2007; Whitmore and Barnes, 2008; El Adli et al., 2014; Gol’din and Startsev, 2014). Also, Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’ do not have an alignment of CNVII and CNVIII openings and perilymphatic foramen in comparison with the Cetetheriidae.
Halicetus ignotus from the middle Miocene Choptank Formation of Maryland show many differences to Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’: a distinct posterior pointed caudal tympanic process, a strongly reduced anterior process, the lateral projection of anterior process which is ventrally positioned and wing-like, and a convex indentation of the lateral surface of periotic body (Kellogg, 1969).
Uranocetus gramensis from the upper Serravallian to Tortonian of South Jutland, Denmark, a cetothere sensu lato after cladistic analyses of Boessenecker and Fordyce (2015a), differs from Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’ by distinct coronoid processes anterior on the anterior process. With respect to the state of preservation of the periotic and its pars cochlearis, the latter does not seem to be tilted and flattened and lacks a ventrally inflated promontorium (Steeman, 2009).
The periotic of Pelocetus calvertensis, also from the middle Miocene section of the Calvert Formation of Maryland, possesses a distinct and triangular caudal tympanic process with a curved ventral margin or outline, a short anterior process, and a lateral projection of anterior process or even anterior process which is extended far ventrally (Kellogg, 1965; Steeman, 2007; Marx and Fordyce, 2015), characters that are different from Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’. The recently presented Persufflatius renefraaijeni from the upper Miocene of Liessel/Netherlands has obvious similarities to Pelocetus calvertensis and Atlanticetus patulus (after Bosselaers and Munstermann, 2022), especially the massive periotic body, the comparatively small pars cochlearis without an inflated promontorium.
Besides Diorocetidae, Steeman (2007) introduced the new family Aglaocetidae, also by its ear bone characters (here: tympanic bulla and periotic). Some authors (among others, Deméré et al., 2008; Hampe and Ritsche, 2011; Marx, 2011) doubted about the validity of the “Aglaocetidae”. Furthermore, unclarified remained the situation of Aglaocetus analysed by Bisconti et al. (2013). Recently, Bisconti et al. (2021) separated species existing besides the type species Aglaocetus moreni. A. moreni was described first by Lydekker (1894) based on a relatively well-preserved skull. It was initially designated as Cetotherium moreni but later recognized as generically distinct from Cetotherium. Kellogg (1934) has established the new genus Aglaocetus. He described a new discovered cranium from Chubut and referred this specimen to A. moreni. “Aglaocetus” patulus was then erected by Kellogg (1968c) and considered to be congeneric with A. moreni, also by Steeman (2010). Later phylogenetic analyses questioned the position of “A.” patulus within the genus (e.g., Marx, 2011; Boessenecker and Fordyce, 2015a, b; Bisconti and Bosselaers, 2016). Unfortunately, A. moreni, stored in the Museo de Ciencias Naturales de La Plata, was unavailable or without well-preserved and isolated periotic bones (Chicago specimen, FMNH 13407). Bisconti et al. (2021) erected a new species and assigned A. patulus and the new one to the genus Atlanticetus.
Atlanticetus patulus (USNM 23049 and USNM 23690) differ from both Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’ in having a thick periotic body with a strongly convex lateral side and encompassed by a small anterior process. The lateral surface of the periotic bone is characterised by furrows extending dorsally from the lateral projection of anterior process. The caudal tympanic process of Atlanticetus contrasts with Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’ well developed and forms a broad triangular shelf. The stylomastoid fossa is broad and deepened and not shallow as in Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’.
The modern Balaenopteridae differ clearly from Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’ with the presence of an elongated pars cochlearis and endocranially extended, reduced, or even recessed superior process, and - in alternate definition - a large caudal tympanic process (Steeman, 2007), or developed as a robust, ventrally directed projection (Marx and Fordyce, 2015), or large and posteriorly extended or with proximally expanded base (our observation).
Very distinct from Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’ are the periotics of the Balaenidae. These ear bones are characterised by a short anterior process with a hypertrophied lateral projection of anterior process and a significantly massive periotic body (Bisconti, 2003, 2005; Ekdale et al., 2011; Marx et al., 2016). This characterisation is already documented in the primitive Morenocetus parvus from the Burdigalian of Argentina (Buono et al., 2017).
REVIEW ON THE INTERNAL ACOUSTIC MEATUS ANATOMY
In cetaceans, the internal acoustic meatus (IAM) is a prominent feature of the endocranial side of the pars cochlearis. Due to the excessive growth of the bony tissue endocranially, the anatomy of the internal acoustic meatus is dramatically modified in Mysticeti and may lead to misinterpretation of anatomical structures within the IAM and its fundus. The basic anatomy of the fundus was described by using the human ear with focus on crista transversa, Bill’s bar, foramen singulare, and courses of vestibular nerve parts (Braus, 1960; Schünke et al., 2009). In several mysticete ears, the origin of the nerves and its egression through the fundus was shown by mapping of all the nerves deep within the pars cochlearis up to the fundus of the internal acoustic meatus (FIAM).
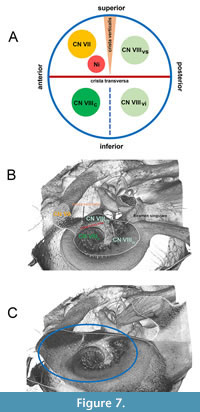 Principally, the internal acoustic meatus provides a passage through which the vestibulocochlear nerve (CN VIII) and the facial nerve (CN VII) can pass from inside the skull to the inner ear and face. Within the FIAM the crista transversa separates the superior part from the inferior part whereas the crista verticalis or Bill’s bar separates the anterosuperior part (which transmits the facial nerve) from the posterosuperior part (which transmits the superior vestibular nerve; Figure 7A). During the evolution of the Mysticeti, the facial nerve was separated distinctly from the coinjoined vestibulocochlear nerve by a prominent bony septum. This structure was recognized by Sanders and Barnes (2002: p. 323). The bony septum could probably be considered as homologous to crista verticalis or Bill’s bar since it separates primarily the facial nerve from the superior vestibular nerve as could be traced back in Eomysticetus whitmorei from the late Oligocene. Geisler and Luo (1996: p. 1055) used the term ‘transverse septum’ for the bony septum, which separates the foramen for the facial nerve from the internal acoustic meatus, and confusingly also as a synonym for the term ‘crista transversa’ (Geisler and Luo, 1996: p. 1049). Several authors (e.g., Mead and Fordyce, 2009; Marx et al., 2015; Boessenecker and Fordyce, 2017) also used the term ‘crista transversa’ for the bony septum between CN VII and CN VIII openings. For evolved Mysticeti it is strongly recommended to simply use the term “bony septum” as a designation of the crest between CN VII and CN VIII openings avoiding confusion with the crista transversa.
Principally, the internal acoustic meatus provides a passage through which the vestibulocochlear nerve (CN VIII) and the facial nerve (CN VII) can pass from inside the skull to the inner ear and face. Within the FIAM the crista transversa separates the superior part from the inferior part whereas the crista verticalis or Bill’s bar separates the anterosuperior part (which transmits the facial nerve) from the posterosuperior part (which transmits the superior vestibular nerve; Figure 7A). During the evolution of the Mysticeti, the facial nerve was separated distinctly from the coinjoined vestibulocochlear nerve by a prominent bony septum. This structure was recognized by Sanders and Barnes (2002: p. 323). The bony septum could probably be considered as homologous to crista verticalis or Bill’s bar since it separates primarily the facial nerve from the superior vestibular nerve as could be traced back in Eomysticetus whitmorei from the late Oligocene. Geisler and Luo (1996: p. 1055) used the term ‘transverse septum’ for the bony septum, which separates the foramen for the facial nerve from the internal acoustic meatus, and confusingly also as a synonym for the term ‘crista transversa’ (Geisler and Luo, 1996: p. 1049). Several authors (e.g., Mead and Fordyce, 2009; Marx et al., 2015; Boessenecker and Fordyce, 2017) also used the term ‘crista transversa’ for the bony septum between CN VII and CN VIII openings. For evolved Mysticeti it is strongly recommended to simply use the term “bony septum” as a designation of the crest between CN VII and CN VIII openings avoiding confusion with the crista transversa.
The foramen singulare is the most posteriorly situated foramen within the fundus of the IAM. This very small foramen is found on the posterior wall, inferior to the opening of the inferior vestibular nerve and transmits the posterior ampullar nerve of the posterior semicircular duct (Braus, 1960; Schünke et al., 2009). Several authors (e.g., Sanders and Barnes, 2002; Mead and Fordyce, 2009; Marx et al., 2015; Boessenecker and Fordyce, 2017) used the term ‘foramen singulare’ for the opening of the vestibular nerve. It is strongly recommended not to use the term ‘foramen singulare’ as designation of the opening of the vestibular nerve.
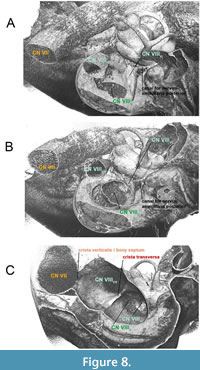 To resolve the confusion about distinct structures within the IAM, the diverse conditions were described with corresponding figures as outlined below. The fundus of IAM in Tursiops truncatus resembles approximately the morphology of fundus of IAM seen in humans (Figure 7B, C). In E. whitmorei, the opening of the superior vestibular nerve has evolved as a single distinct opening within a broad and elongated IAM (Figure 8A-C). It is separated from CN VII by a prominent crista verticalis/bony septum besides a well-developed crista transversa, which separates the superior vestibular nerve from the inferior vestibular nerve and the cochlear nerve (Figure 8C).
To resolve the confusion about distinct structures within the IAM, the diverse conditions were described with corresponding figures as outlined below. The fundus of IAM in Tursiops truncatus resembles approximately the morphology of fundus of IAM seen in humans (Figure 7B, C). In E. whitmorei, the opening of the superior vestibular nerve has evolved as a single distinct opening within a broad and elongated IAM (Figure 8A-C). It is separated from CN VII by a prominent crista verticalis/bony septum besides a well-developed crista transversa, which separates the superior vestibular nerve from the inferior vestibular nerve and the cochlear nerve (Figure 8C).
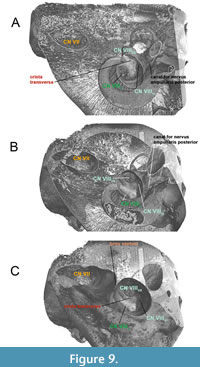 In crown Mysticeti (Balaenomorpha), the bony septum is very prominent and separates the facial nerve and the coinjoined vestibulocochlear nerve in two distinct parts within the IAM as seen in Parieobalaena palmeri (Figure 9A-C). The crista transversa is no longer recognizable in derived forms, as it lies deep inside the IAM and within the FIAM. The occurrence of a recessed bony septum as seen in the living Balaenoptera borealis (Figure 10A-C) is caused by the excessive growth of the pars cochlearis endocranially. The separation between CN VII and CN VIII by a very thick bony septum or sulcus (e.g.,
In crown Mysticeti (Balaenomorpha), the bony septum is very prominent and separates the facial nerve and the coinjoined vestibulocochlear nerve in two distinct parts within the IAM as seen in Parieobalaena palmeri (Figure 9A-C). The crista transversa is no longer recognizable in derived forms, as it lies deep inside the IAM and within the FIAM. The occurrence of a recessed bony septum as seen in the living Balaenoptera borealis (Figure 10A-C) is caused by the excessive growth of the pars cochlearis endocranially. The separation between CN VII and CN VIII by a very thick bony septum or sulcus (e.g., 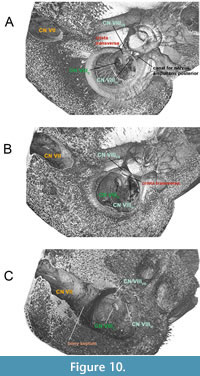 Balaenomorpha ‘type Bocholt B’ or Amphicetus later) is probably depending on the cerebral elongation of the pars cochlearis and on the ontogenetic stage of the specimen (see also Bisconti, 2001). It is also important to consider the preservation status of the pars cochlearis and its delicate endocranial structures.
Balaenomorpha ‘type Bocholt B’ or Amphicetus later) is probably depending on the cerebral elongation of the pars cochlearis and on the ontogenetic stage of the specimen (see also Bisconti, 2001). It is also important to consider the preservation status of the pars cochlearis and its delicate endocranial structures.
Definition and Recommended Use of Terminology (in alphabetical order)
Bony septum. Separates Nervus facialis (CN VII) from Nervus vestibulocochlearis (CN VIII) within the internal acoustic meatus. An independent structure from the fundus of the internal acoustic meatus in evolved mysticetes.
Crista transversa. Separates the superior part from the inferior part within the fundus of the internal acoustic meatus and has an almost anteroposterior orientation.
Crista verticalis = Bill’s bar. Separates the anterosuperior part (which transmits the Nervus facialis, CN VII) from the posterosuperior part (which transmits the Nervus utriculoampullaris, CN VIIIVS) within the fundus of the internal acoustic meatus.
Foramen singulare. Most posteriorly situated foramen within the fundus of the internal acoustic meatus. It is positioned on the posterior wall, inferior to the opening of the Nervus vestibularis inferior and transmits the Nervus ampullaris posterior of the posterior semicircular duct.
Fundus of the internal acoustic meatus. Lateral end of the internal acoustic meatus formed by a thin cribriform plate of bone that separates the cochlea and vestibule which are in close proximity.
Internal acoustic meatus. Opening that always includes both the passages for the Nervus vestibulocochlearis (CN VIII) and Nervus facialis (CN VII).
CONCLUSIONS
Whale remains from the upper Serravallian of Bocholt-Biemenhorst in the Münsterland, Germany, yielded two forms with hitherto unknown combination of periotic characters. Balaenomorpha ‘type Bocholt A’ is characterised by an anterior portion of pars cochlearis, which is more elongated dorsoventrally and somewhat pointed dorsomedially and a significantly elongated and pointed superior process. Also, the opening for the vestibulocochlear nerve (CN VIII) orients more medially and the opening for the facial nerve (CN VII) orients more anteriorly. Balaenomorpha ‘type Bocholt B’ is characterised by an elevated and squared pars cochlearis, and a tongue-shaped, elongated superior process with a distinct dorsolateral notch. This second Bocholt form has also an opening for the vestibulocochlear nerve (CN VIII) that orients more medially and an opening for the facial nerve (CN VII) that orients more anteriorly. Both openings are separated here by a distinct sulcus.
The presented periotics, Balaenomorpha ‘type Bocholt A’ and ‘type Bocholt B’ share distinct characters with members of the Diorocetidae. A validity of the Diorocetidae regarding periotic morphology is supported by well-established taxa such as Diorocetus (Kellogg, 1968a; Gottfried et al., 1994; Steeman, 2007, 2010; Hampe and Ritsche, 2011) and Amphicetus (Van Beneden, 1880; Steeman, 2010; Hampe and Ritsche, 2011), including Thinocetus (Kellogg, 1969; Tanaka et al., 2018) and are primarily characterised by having a tilted, flattened pars cochlearis with a ventrally inflated promontorium and a broad, shallow stylomastoid fossa laterally expanded onto the periotic body.
Detailed analyses of the internal and external structures of whale periotics resulted in a critical review and revision of two important characteristics of the pars cochlearis. First, the partition between the openings for the facial nerve (CN VII) and the superior vestibular nerve superior, which is the crista verticalis in the human fundus of the internal acoustic meatus, should be named “bony septum” in evolved baleen whales. This bony septum has been developed so prominently during mysticete evolution and does not belong anymore to the fundus of the internal acoustic meatus. The crista transversa is a structure of the fundus of the internal acoustic meatus and no longer recognizable in derived forms, as it lies deep inside the inner ear canal where it separates the cochlear nerve from the inferior vestibular nerve.
Secondly, the designation of the opening for the superior vestibular nerve as ‘foramen singulare’ is misleading since this term describes the opening for the nerve to the posterior semicircular canal at the bottom of the internal acoustic meatus. ‘Foramen singulare’ was used as the opening for the superior vestibular nerve in various taxa (e.g., Sanders and Barnes, 2002 for Zygorhiza; Mead and Fordyce, 2009 for Tursiops truncatus; Boessenecker and Fordyce, 2015a for Waharoa ruwhenua). The term ‘foramen singulare’ should only be used for the opening of the superior vestibular nerve.
The high diversification and the continuing discoveries of new mysticete taxa especially in European waters is not astonishing at all. Transgression events during the Late Palaeogene and Early Neogene based on orogenic-tectonic movements produced several local marine evolutionary centres around the European continent which certainly allowed constantly the generation of new populations (Hampe and Fahlke, 2022).
ACKNOWLEDGEMENTS
This work could not have been realised without the kind assistance of the following colleagues, curators, collection managers and preparators to whom we are much obliged for providing access to the collections and/or exhibitions: D. Willborn (Berlin), E. Horstrup (Bremervörde, Germany), A. Folie and O. Lambert (Brussels), M. Bosselaers (Middelburg, The Netherlands), C. de Muizon (Paris), M.T. Olsen (Copenhagen), E. Fordyce (Dunedin, New Zealand), N. Pyenson and D. Bohaska (Washington, D.C.), and J. McCormick (Charleston, SC).
We are deeply thankful to A. Hilger, N. Kardjilov, and F. Wieder (all Berlin) for the fruitful collaboration with CT scanning and 3D image reconstruction of the ear bones.
We are indebted to J. Fahlke (Kiel, Germany) for intensive and inspiring discussions and to C. Hartkopf-Fröder (Krefeld and Cologne) for additional information about the local geology. Finally, we want to thank two anonymous reviewers for their constructive and detailed comments and suggestions for improvement.
This research on fossil and extant whales was funded by grants from the German Scientific Foundation (DFG) HA 1776/13-1, FA 889/2-1, and BA 1170/25-1).
REFERENCES
Arlt, T., Wieder, F., Ritsche, I., Hilger, A., Kardjilov, N., Fahlke, J.M., Hampe, O., and Manke, I. 2018. Röntgen- und Neutronentomographie am knöchernen Innenohr der Bartenwale. Materials Testing, 60(2):173-178.
https://doi.org/10.3139/120.111129
Bisconti, M. 2001. Morphology and postnatal growth trajectory of rorqual petrosal. Italian Journal of Zoology, 68:87-93.
https://doi.org/10.1080/11250000109356390
Bisconti, M. 2003. Evolutionary history of Balaenidae. Cranium, 20:9-49.
Bisconti, M. 2005. Skull morphology and phylogenetic relationships of a new diminutive balaenid from the lower Pliocene of Belgium. Palaeontology, 48:793-816.
https://doi.org/10.1111/j.1475-4983.2005.00488.x
Bisconti, M. and Bosselaers, M. 2016. Fragilicetus velponi: a new mysticete genus and species and its implications for the origin of Balaenopteridae (Mammalia, Cetacea, Mysticeti). Zoological Journal of the Linnean Society, 177:450-474.
https://doi.org/10.1111/zoj.12370
Bisconti, M., Damarco, P., Mao, S., Pavia, M., and Carnevale, G. 2021. The earliest baleen whale from the Mediterranean: large‐scale implications of an early Miocene thalassotherian mysticete from Piedmont, Italy. Papers in Palaeontology, 7(2):1147-1166.
https://doi.org/10.1002/spp2.1336
Bisconti, M., Lambert, O. and Bosselaers, M. 2013. Taxonomic revision of Isocetus depauwi (Mammalia, Cetacea, Mysticeti) and the phylogenetic relationships of archaic ‘cetothere’ mysticetes. Palaeontology, 56:95-127.
https://doi.org/10.1111/j.1475-4983.2012.01168.x
Bisconti, M., Raineri, G., Tartarelli, G., Monegatti, P., and Carnevale, G. 2022. The periotic of a basal balaenopterid from the Tortonian of the Stirone River, northern Italy (Cetacea, Mysticeti, Balaenopteridae). Palaeobiodiversity and Palaeoenvironments.
https://doi.org/10.1007/s12549-022-00550-2
Bosselaers, M. and Munsterman, D.K. 2022. The discovery of a Balaenomorpha (Persufflatius renefraaijeni n. gen., n. sp.) from the upper Miocene of the Netherlands sheds new light on the cranial anatomy of archaic rorqual relatives. Geodiversitas, 44(30):933-973.
https://doi.org/10.5252/geodiversitas2022v44a30
Boessenecker, R.W. and Fordyce, R.E. 2015a. Anatomy, feeding ecology, and ontogeny of a transitional baleen whale: a new genus and species of Eomysticetidae (Mammalia: Cetacea) from the Oligocene of New Zealand. PeerJ, 3:e1129.
https://doi.org/10.7717/peerj.1129
Boessenecker, R.W. and Fordyce, R.E. 2015b. A new genus and species of eomysticetid (Cetacea: Mysticeti) and a reinterpretation of ‘ Mauicetus’ lophocephalus Marples, 1956: transitional baleen whales from the upper Oligocene of New Zealand. Zoological Journal of the Linnean Society, 175:607-660.
https://doi.org/10.1111/zoj.12297
Boessenecker, R.W. and Fordyce, R.E. 2017. A new eomysticetid from the Oligocene Kokoamu Greensand of New Zealand and a review of the Eomysticetidae (Mammalia, Cetacea). Journal of Systematic Palaeontology, 15(6):429-469.
https://doi.org/10.1080/14772019.2016.1191045
Boessenecker, R.W., Beatty, B.L., and Geisler, J.H. 2023. New specimens and species of the Oligocene toothed baleen whale Coronodon from South Carolina and the origin of Neoceti. PeerJ, 11:e14795.
https://doi.org/10.7717/peerj.14795
Bouetel, V. and Muizon, C. de 2006. The anatomy and relationships of Piscobalaena nana (Cetacea, Mysticeti), a Cetotheriidae s.s. from the early Pliocene of Peru. Geodiversitas, 28:319-395.
Braus, H. 1960. Anatomie des Menschen, Ein Lehrbuch für Studierende und Ärzte. Band 3: Periphere Leitungsbahnen II. Centrales Nervensystem. Sinnesorgane. 2nd Edition. Springer, Berlin.
Brisson, A.D. 1762. Regnum animale in Classes IX. distributum, sive Synopsis methodica sistens generalem Animalium distributionem in Classes IX, & duarum primarum Classicum, Quadrupedum scilicet & Cetaceorum, particularem divisionem in Ordines, Sectiones, Genera & Species. Haak, Leiden.
Buono, M.R., Fernández, M.S., Cozzuol, M.A., Cuitiño, J.I., and Fitzgerald, E.M.G. 2017. The early Miocene balaenid Morenocetus parvus from Patagonia (Argentina) and the evolution of right whales. PeerJ, 5: e4148.
https://doi.org/10.7717/peerj.4148
Deméré, T.A., McGowen, M.R., Berta, A., and Gatesy, J. 2008. Morphological and molecular evidence for a stepwise evolutionary transition from teeth to baleen in mysticete whales. Systematic Biology, 57(1):15-37.
https://doi.org/10.1080/10635150701884632
Doppert, J.W.C., Ruegg, G.H.J., van Staalduinen, C.J., Zagwijn, W.H., and Zandstra, J.G. 1975. Formaties van het Kwartair en Boven-Tertiair in Nederland, p. 11-56. In Zagwijn, W.H. and van Staalduinen, C.J. (eds.), Toelichting bij geologische overzichtskaarten van Nederland. Rijks Geologische Dienst, Haarlem.
Ekdale, E.G., Berta, A., and Deméré, T.A. 2011. The comparative osteology of the petrotympanic complex (ear region) of extant baleen whales (Cetacea: Mysticeti). PLoS One, 6:e21311.
https://doi.org/10.1371/journal.pone.0021311
El Adli, J.E., Deméré, T.A., and Boessenecker, R.W. 2014. Herpetocetus morrowi (Cetacea: Mysticeti), a new species of diminutive baleen whale from the upper Pliocene (Piacenzian) of California, USA, with observations on the evolution and relationships of the Cetotheriidae. Zoological Journal of the Linnean Society, 170:400-466.
https://doi.org/10.1111/zoj.12108
Flower, W.H. 1864. Notes on the skeletons of whales in the principal museums of Holland and Belgium, with descriptions of two species apparently new to science. Proceedings of the Zoological Society of London, 1864:1-38.
Gaillard, F. 2022. Nerves of the internal acoustic meatus (diagram), Case study, Radiopaedia.org.
https://doi.org/10.53347/rID-36049
Geisler, J.H. and Luo, Z. 1996. The petrosal and inner ear of Herpetocetus sp. (Mammalia: Cetacea) and their implications for the phylogeny and hearing of archaic mysticetes. Journal of Paleontology, 70(6):1045-1066.
https://doi.org/10.1017/s0022336000038749
Geisler, J.H. and Sanders, A.E. 2003. Morphological evidence for the phylogeny of Cetacea. Journal of Mammalian Evolution, 10:23-129.
https://doi.org/10.1023/a:1025552007291
Gol’din, P. and Startsev, D. 2014. Brandtocetus, a new genus of baleen whales (Cetacea, Cetotheriidae) from the late Miocene of Crimea, Ukraine. Journal of Vertebrate Paleontology, 34:419-433.
https://doi.org/10.1080/02724634.2013.799482
Gottfried, M.D., Bohaska, D.J., and Whitmore, F.C. Jr. 1994. Miocene cetaceans of the Chesapeake Group. In Berta, A. and Deméré, T.A. (eds.), Contributions in Marine Mammal Paleontology Honoring Frank C. Whitmore Jr. Proceedings of the San Diego Society of Natural History, 29:229-238.
Hampe, O. and Fahlke, J.M. 2022. A large chaeomysticete (Mammalia: Cetacea) from the middle/late Miocene mica-clay of Groß Pampau (North Sea Basin, North Germany). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 305(1):11-38.
https://doi.org/10.1127/njgpa/2022/1075
Hampe, O. and Ritsche, I. 2011. Die Bartenwalfauna (Cetacea: Mysticeti: Balaenomorpha) aus dem Lüneburgium (Serravallium, Mittelmiozän) von Freetz bei Sittensen (Niedersachsen, Deutschland). Zeitschrift für geologische Wissenschaften, 39(2):83-110.
Heinrichs, P. 1956. Paläontologische Funde in und um Bocholt. Unser Bocholt, 1956(2):30-34.
Heinrichs, P. 1957. Bericht über die Ausgrabung von zwei Urwalen in Biemenhorst. Unser Bocholt, 1957(1):32-36.
Hiss, M. 2013. Breda-Formation, LithoLex, BGR Hannover, Record No. 2008128.
http://www.bgr.bund.de/litholex
Jansen, F. 2004. Erläuterungen zu Blatt 4205 Hamminkeln. Geologische Karte von Nordrhein-Westfalen 1:25000. Geologischer Dienst NRW, Krefeld.
Kellogg, R. 1924. Description of a new genus and species of whalebone whale from the Calvert Cliffs, Maryland. Proceedings of the United States National Museum, 63:1-14.
https://doi.org/10.5479/si.00963801.63-2483.1
Kellogg, R. 1934. The Patagonian fossil whalebone whale, Cetotherium moreni (Lydekker). Carnegie Institution of Washington Publication, 447:64-81.
Kellogg, R. 1965. Fossil marine mammals from the Miocene Calvert Formation of Maryland and Virginia. Part 1. A new whalebone whale from the Miocene Calvert Formation. United States National Museum Bulletin, 247:1-45.
Kellogg, R. 1968a. Fossil marine mammals from the Miocene Calvert Formation of Maryland and Virginia. Part 6. A hitherto unrecognized Calvert cetothere. United States National Museum Bulletin, 247:133-161.
Kellogg, R. 1968b. Fossil marine mammals from the Miocene Calvert Formation of Maryland and Virginia. Part 8. Supplement to description of Parietobalaena palmeri. United States National Museum Bulletin, 247:175-201.
Kellogg, R. 1968c. Fossil marine mammals from the Miocene Calvert Formation of Maryland and Virginia. Part 7. A sharp-nosed cetothere from the Miocene Calvert. United States National Museum Bulletin, 247:163-173.
Kellogg, R. 1969. Cetothere skeletons from the Miocene Choptank Formation of Maryland and Virginia. United States National Museum Bulletin, 294:1-40.
https://doi.org/10.5479/si.03629236.294.1
King, C. 2016. The North Sea basin: mid-Miocene to early Pleistocene, p. 269-327. In King, C., Gale, A.S., and Barry, T.L. (eds.), A revised correlation of Tertiary rocks in the British Isles and adjacent areas of NW Europe. Geological Society Special Reports, 27.
https://doi.org/10.1144/sr27.13
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. Laurentii Salvii, Stockholm.
Lydekker, R. 1894. Contributions to a knowledge of the fossil vertebrates of Argentina 2. Cetacean skulls from Patagonia. Anales del Museo de La Plata, Paleontología Argentina, 2:1-13.
Marx, F.G. 2011. The more the merrier? A large cladistic analysis of mysticetes, and comments on the transition from teeth to baleen. Journal of Mammalian Evolution, 18:77-100.
https://doi.org/10.1007/s10914-010-9148-4
Marx, F.G. and Fordyce, R.E. 2015. Baleen boom and bust: a synthesis of mysticete phylogeny, diversity and disparity. Royal Society Open Science, 2:140434.
https://doi.org/10.1098/rsos.140434
Marx, F.G., Lambert, O., and Uhen, M.D. 2016. Cetacean Paleobiology. Wiley Blackwell, Chichester, UK.
https://doi.org/10.1002/9781118561546
Marx, F.G., Tsai, C.-H., and Fordyce, R.E. 2015. A new early Oligocene toothed ‘baleen’ whale (Mysticeti: Aetiocetidae) from western North America: one of the oldest and the smallest. Royal Society Open Science, 2:150476.
https://doi.org/10.1098/rsos.150476
Mead, J.G. and Fordyce, R.E. 2009. The therian skull: A lexicon with emphasis on the odontocetes. Smithsonian Contributions to Zoology, 627:1-248.
https://doi.org/10.5479/si.00810282.627
Mitchell, E.D. 1989. A new cetacean from the Late Eocene La Meseta Formation, Seymour Island, Antarctic Peninsula. Canadian Journal of Fisheries and Aquatic Sciences, 46:2219-2235.
https://doi.org/10.1139/f89-273
Sanders, A.E. and Barnes, L.G. 2002. Paleontology of the late Oligocene Ashley and Chandler Bridge Formations of South Carolina, 3: Eomysticetidae, a new family of primitive mysticetes (Mammalia: Cetacea), p. 313-356. In Emry, R.J. (ed.), Cenozoic mammals of land and sea: Tributes to the career of Clayton E. Ray. Smithsonian Contributions to Paleobiology, 93.
Schünke, M., Schulte, E., and Schumacher, U. 2009. Prometheus LernAtlas der Anatomie. Kopf, Hals und Neuroanatomie. 2nd Edition. Georg Thieme Verlag, Stuttgart.
Springhorn, R. 1978. Extremitätenreste einer Mönchsrobbe (Phocidae, Carnivora, Mammalia) aus dem Miozän von Bocholt (Niederrhein). Paläontologische Zeitschrift, 52(1-2):19-27.
https://doi.org/10.1007/bf03006727
Steeman, M.E. 2007. Cladistic analysis and a revised classification of fossil and recent mysticetes. Zoological Journal of the Linnean Society, 150:875-894.
https://doi.org/10.1111/j.1096-3642.2007.00313.x
Steeman, M.E. 2009. A new baleen whale from the Late Miocene of Denmark and early mysticete hearing. Palaeontology, 52:1169-1190.
https://doi.org/10.1111/j.1475-4983.2009.00893.x
Steeman, M.E. 2010. The extinct baleen whale fauna from the Miocene-Pliocene of Belgium and the diagnostic cetacean ear bones. Journal of Systematic Palaeontology, 8:63-80.
https://doi.org/10.1080/14772011003594961
Tanaka, Y., Ando, T., and Sawamura, H. 2018. A new species of middle Miocene baleen whale from the Nupinai Group, Hikatagawa Formation of Hokkaido, Japan. PeerJ, 6:e4934.
https://doi.org/10.7717/peerj.4934
Van Beneden, P.J. 1880. Les Mysticètes à courts fanons des sables des environs d'Anvers. Bulletins de l'Académie royale des sciences, des lettres et des beaux-arts de Belgique, 2me Série, 50:11-25.
von der Hocht, F. 1988. Die Knorpelfischfauna der Walfundstelle von Kervenheim. Natur am Niederrhein Neue Folge, 3:13-24.
von Laursen, G., Poulsen, N.E., and Rasmussen, L.B. 1998. Correlation of Northwest European Miocene stages with the international stages - preliminary results. Newsletters on Stratigraphy, 36(1):55-61.
https://doi.org/10.1127/nos/36/1998/55
Whitmore, F.C. Jr. and Barnes, L.G. 2008. The Herpetocetinae, a new subfamily of extinct naleen whales (Mammalia, Cetacea, Cetotheriidae), p. 141-180. In Ray, C.E., Bohaska, D.J., Koretsky, I.A., Ward, L.W., and Barnes, L.G. (eds.), Geology and Paleontology of the Lee Creek Mine, North Carolina, IV. Virginia Museum of Natural History Special Publication, 14.
