

 Late middle Eocene caviomorph rodents from Contamana, Peruvian Amazonia
Late middle Eocene caviomorph rodents from Contamana, Peruvian Amazonia
Article number: 20.1.19A
https://doi.org/10.26879/742
Copyright Palaeontological Association, April 2017
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 3 November 2016. Acceptance: 5 April 2017
{flike id=1822}
ABSTRACT
Caviomorph rodents represent one of the most successful groups of placental mammals from South America. Despite their modern, Neogene and late Paleogene high diversity, their early evolutionary history has long remained obscure. Recent field expeditions in Peruvian Amazonia have yielded among the earliest representatives of that group, in deposits dating from the late middle Eocene (Contamana, CTA-27 locality, ~41 Ma). Here, we provide an exhaustive analysis of the rodent material from CTA-27 and from new sub-coeval localities discovered in the same area and geological formation (Pozo Formation): CTA-47, CTA-51, CTA-73, CTA-66, and CTA-29. A total of 20 rodent taxa are identified in these localities, among which one from CTA-29 (Pozomys ucayaliensis gen. et sp. nov.) remains with uncertain suprafamilial affinities. Additionally, the material of CTA-27 previously attributed to Eobranisamys sp. is assigned here to the new species Eobranisamys javierpradoi. In terms of taxonomic composition, Eocene localities from Contamana area have many taxa in common (Cachiyacuy, Canaanimys, Eobranisamys, and Eoespina, or very close relatives). These Eocene assemblages are clearly distinct from Oligocene ones, mostly documented at mid and high latitudes. In contrast, they share some affinities with the late Eocene-earliest Oligocene Santa Rosa locality (Peruvian Amazonia), from which the two genera Eobranisamys and Eoespina were originally described. This faunal closeness might more reflect biogeographic affinities than contemporaneity. In addition, the occlusal pattern of some upper molars of Eosallamys from Santa Rosa recalls that of Cachiyacuy and Canaanimys. These low-latitude caviomorph assemblages provide new insights into the early evolutionary history, biogeography, and paleodiversity of that group.
Myriam Boivin. Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR 5554, CNRS/UM/IRD/EPHE), c.c. 064, Université de Montpellier, place Eugène Bataillon, F-34095 Montpellier Cedex 05, France. myriam.boivin@umontpellier.fr
Laurent Marivaux. Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR 5554, CNRS/UM/IRD/EPHE), c.c. 064, Université de Montpellier, place Eugène Bataillon, F-34095 Montpellier Cedex 05, France. laurent.marivaux@umontpellier.fr
Maëva J. Orliac. Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR 5554, CNRS/UM/IRD/EPHE), c.c. 064, Université de Montpellier, place Eugène Bataillon, F-34095 Montpellier Cedex 05, France. maeva.orliac@umontpellier.fr
Francois Pujos. Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales (IANIGLA), CCT-CONICET-Mendoza, Av. Ruiz Leal s/n, Parque Gral. San Martín, 5500 Mendoza, Argentina. fpujos@mendoza-conicet.gob.ar
Rodolfo Salas-Gismondi. Departamento de Paleontología de Vertebrados, Museo de Historia Natural - Universidad Nacional Mayor San Marcos (MUSM), Av. Arenales 1256, Lima 11, Peru. rsalasgismondi@gmail.com
Julia V. Tejada-Lara. Departamento de Paleontología de Vertebrados, Museo de Historia Natural - Universidad Nacional Mayor San Marcos (MUSM), Av. Arenales 1256, Lima 11, Peru, and Department of Earth and Environmental Sciences, Columbia University in the City of New York and Division of Paleontology, American Museum of Natural History, Central Park West at 79th Street, New York, NY 10024, USA. julia.tejada@columbia.edu
Pierre-Olivier Antoine. Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR 5554, CNRS/UM/IRD/EPHE), c.c. 064, Université de Montpellier, place Eugène Bataillon, F-34095 Montpellier Cedex 05, France. pierre-olivier.antoine@umontpellier.fr
Keywords: Barrancan; South America; Systematics; Biostratigraphy; new genus; new species
Final citation: Boivin, Myriam, Marivaux, Laurent, Orliac, Maëva J., Pujos, Francois, Salas-Gismondi, Rodolfo, Tejada-Lara, Julia V., and Antoine, Pierre-Olivier. 2017. Late middle Eocene caviomorph rodents from Contamana, Peruvian Amazonia. Palaeontologia Electronica 20.1.19A: 1-50. https://doi.org/10.26879/742
palaeo-electronica.org/content/2017/1822-eocene-amazonian-caviomorphs
http://zoobank.org/540D23AA-F705-4A05-8E10-FADAD3356D9C
INTRODUCTION
The caviomorph rodents (Caviomorpha Wood, 1955) constitute one of the main placental mammal groups of South America. They are represented nowadays by four superfamilies (Erethizontoidea Bonaparte, 1845 [New World porcupines], Octodontoidea Waterhouse, 1839 [spiny rats and their kin], Chinchilloidea Bennett, 1833 [chinchillas and their kin], and Cavioidea Fischer de Waldheim, 1817 [guinea pig and their kin]), including a total of 13 families (e.g., Wilson and Reeder, 2005). Their modern specific richness amounts to ~11% of the rodent diversity (255/2277; Wilson and Reeder, 2005). Moreover, caviomorphs show a wide morphological disparity, many dietary and locomotory adaptations, and they occupy diverse environments (e.g., Mares and Ojeda, 1982; Patton et al., 2015). This rodent group, documented in the fossil record of South America since the late middle Eocene (Antoine et al., 2012), also shows a great past diversity (e.g., Woods, 1984; Candela and Picasso, 2008; Rinderknecht and Blanco, 2008).
 Until recently, the oldest fossil occurrence of caviomorphs in South America was recorded from one locality of Peruvian Amazonia: CTA-27 from Contamana (Loreto Department; Antoine et al., 2012). This locality was dated at ~41 Ma (late middle Eocene, Barrancan South American Land Mammal Age [SALMA]) by radiometric Ar/Ar analyses (Antoine et al., 2012). On the basis of dental remains, five taxa were described from this locality, three were new to science: Cachiyacuy contamanensis Antoine et al., 2012, Cachiyacuy kummeli Antoine et al., 2012, and Canaanimys maquiensis Antoine et al., 2012. These taxa are particularly interesting because they display a suite of primitive dental characters (for caviomorphs), which are otherwise found only in coeval African phiomorphs (Antoine et al., 2012; Barbière and Marivaux, 2015). This striking morphological affinity strengthens support for the hypothesis of the African origin of the caviomorph clade. Another locality from Peruvian Amazonia as well, Santa Rosa (Ucayali Department), originally considered as ?late Eocene in age (Frailey and Campbell, 2004), has yielded a rich rodent assemblage (17 taxa), among which the earliest representatives of three caviomorph superfamilies: Erethizontoidea, Octodontoidea, and Cavioidea. However, the late Eocene age of the Santa Rosa locality proposed by Campbell (2004) is highly questioned, with some other authors advocating the possibility of a younger age (i.e., earliest Oligocene or later; Shockey et al., 2004; Croft et al., 2009; Antoine et al., 2012; Kay, 2015; Antoine et al., 2017).
Until recently, the oldest fossil occurrence of caviomorphs in South America was recorded from one locality of Peruvian Amazonia: CTA-27 from Contamana (Loreto Department; Antoine et al., 2012). This locality was dated at ~41 Ma (late middle Eocene, Barrancan South American Land Mammal Age [SALMA]) by radiometric Ar/Ar analyses (Antoine et al., 2012). On the basis of dental remains, five taxa were described from this locality, three were new to science: Cachiyacuy contamanensis Antoine et al., 2012, Cachiyacuy kummeli Antoine et al., 2012, and Canaanimys maquiensis Antoine et al., 2012. These taxa are particularly interesting because they display a suite of primitive dental characters (for caviomorphs), which are otherwise found only in coeval African phiomorphs (Antoine et al., 2012; Barbière and Marivaux, 2015). This striking morphological affinity strengthens support for the hypothesis of the African origin of the caviomorph clade. Another locality from Peruvian Amazonia as well, Santa Rosa (Ucayali Department), originally considered as ?late Eocene in age (Frailey and Campbell, 2004), has yielded a rich rodent assemblage (17 taxa), among which the earliest representatives of three caviomorph superfamilies: Erethizontoidea, Octodontoidea, and Cavioidea. However, the late Eocene age of the Santa Rosa locality proposed by Campbell (2004) is highly questioned, with some other authors advocating the possibility of a younger age (i.e., earliest Oligocene or later; Shockey et al., 2004; Croft et al., 2009; Antoine et al., 2012; Kay, 2015; Antoine et al., 2017).
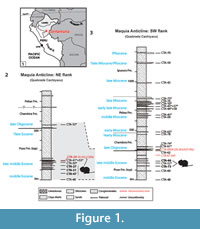
Recently, Antoine et al. (2016) published a noteworthy paleontological and geological survey of the Cenozoic deposits along the Quebrada Cachiyacu near Contamana in Peruvian Amazonia. Of the 19 caviomorph-bearing localities identified by these authors in a single section, 10 are Eocene in age (middle Eocene-late middle Eocene; including CTA-27). Three of these localities (CTA-47, CTA-51, and CTA-73; Figure 1.2-3) are stratigraphically below CTA-27, while the others (CTA-66, CTA-52, CTA-50, CTA-41, CTA-53, and CTA-29; Figure 1.2-3) are located above CTA-27 in the same section. In this paper, we 1) provide an exhaustive analysis (description and comparison) and a revision of the rodent material from CTA-27; and 2) describe the material from other late middle Eocene localities of Contamana (CTA-47, CTA-51, CTA-73, CTA-66, and CTA-29). The study of these caviomorph assemblages, which are the oldest known at a South American scale, thus allows furthering our understanding of the early evolutionary history and paleodiversity of that group during its earliest adaptive radiation.
MATERIAL AND METHODS
 The rodent fossils described in the present work come from six Eocene localities of Contamana (Loreto department, Peru; Figure 1.1): CTA-47, CTA-51, CTA-73, CTA-27, CTA-66, and CTA-29. The four other Eocene caviomorph-bearing localities from the same section (CTA-52, CTA-50, CTA-41, and CTA-53; Antoine et al., 2016) have been excluded because they only yielded fragments of rodent incisors, unidentifiable at a family level or below. CTA-47 (7°19’52.1”S, 74°57’4.6”W), CTA-51 (7°19’46.1”S, 74°56’57.1”W), CTA-27 (7°19’48.4”S, 74°56’50.4”W), and CTA-29 (7°19’42.9”S, 74°56’44”W) are situated on the northeastern flank of the Maquía Anticline, whereas CTA-73 (7°20’29.8”S, 74°57’8.7”W) and CTA-66 (7°20’29.8”S, 74°57’15.5”W) are located on the southwestern flank of that anticline (Antoine et al., 2016, figure 2.A). Following Antoine et al. (2016, figure 3), all these localities are referred to the lower member of the Pozo Formation (“Pozo Sands”; Figure 1.2-3) and correspond to scattered inframetric channelized sandstone lenses (Antoine et al., 2016, figure 5.A-B). At CTA-51, CTA-27, CTA-29, and CTA-66, these sandstones are massive, gray to yellowish, and unconsolidated. They include millimetric calcareous nodules of diagenetic origin (Antoine et al., 2016, figure 5.D), locally encrusting fossil elements (charophyte oogonia, crab claws, fish scales, and vertebrate teeth and bones). CTA-29 consists in a red-brown tuffaceous silty sand dated at 43.44 ± 2.5 Ma by Ar/Ar on biotites (late middle Eocene, Barrancan SALMA; Antoine et al., 2012, 2016; Figure 1.2). The Eocene localities of Contamana have yielded a wide array of aquatic and terrestrial organisms (plants, crabs, molluscs, chondrichthyans, osteichthyans, frogs, turtles, snakes, and crocodylomorphs), including numerous terrestrial mammals (allotherian, metatherians, xenarthrans, South American native ungulates [i.e., astrapothere, litoptern, notungulates, and pyrotheres], and rodents). In addition, CTA-27 and CTA-66 have also yielded bat teeth (Antoine et al., 2016). In the Contamana section, fossil material of each locality was collected by screen-washing (1 mm mesh): ~205 kg of sediment for CTA-47 (in 2010 and 2011), 97 kg of sediment for CTA-51 (in 2010 and 2011), 11 kg of sediment for CTA-73 (2014), 665 kg of sediment for CTA-27 (from 2008 to 2010), 31 kg of sediment for CTA-66 (in 2013 and 2014), and 270 kg of sediment for CTA-29 (from 2011 to 2013). During the last field expedition in the Contamana area (August 2016), our team regrettably noted the irremediable loss of several concerned localities (CTA-51, CTA-27, CTA-73, and CTA-29), either due to landslides, erosion, and silting caused by flooding events that have occurred since 2014 (Appendix 1).
The rodent fossils described in the present work come from six Eocene localities of Contamana (Loreto department, Peru; Figure 1.1): CTA-47, CTA-51, CTA-73, CTA-27, CTA-66, and CTA-29. The four other Eocene caviomorph-bearing localities from the same section (CTA-52, CTA-50, CTA-41, and CTA-53; Antoine et al., 2016) have been excluded because they only yielded fragments of rodent incisors, unidentifiable at a family level or below. CTA-47 (7°19’52.1”S, 74°57’4.6”W), CTA-51 (7°19’46.1”S, 74°56’57.1”W), CTA-27 (7°19’48.4”S, 74°56’50.4”W), and CTA-29 (7°19’42.9”S, 74°56’44”W) are situated on the northeastern flank of the Maquía Anticline, whereas CTA-73 (7°20’29.8”S, 74°57’8.7”W) and CTA-66 (7°20’29.8”S, 74°57’15.5”W) are located on the southwestern flank of that anticline (Antoine et al., 2016, figure 2.A). Following Antoine et al. (2016, figure 3), all these localities are referred to the lower member of the Pozo Formation (“Pozo Sands”; Figure 1.2-3) and correspond to scattered inframetric channelized sandstone lenses (Antoine et al., 2016, figure 5.A-B). At CTA-51, CTA-27, CTA-29, and CTA-66, these sandstones are massive, gray to yellowish, and unconsolidated. They include millimetric calcareous nodules of diagenetic origin (Antoine et al., 2016, figure 5.D), locally encrusting fossil elements (charophyte oogonia, crab claws, fish scales, and vertebrate teeth and bones). CTA-29 consists in a red-brown tuffaceous silty sand dated at 43.44 ± 2.5 Ma by Ar/Ar on biotites (late middle Eocene, Barrancan SALMA; Antoine et al., 2012, 2016; Figure 1.2). The Eocene localities of Contamana have yielded a wide array of aquatic and terrestrial organisms (plants, crabs, molluscs, chondrichthyans, osteichthyans, frogs, turtles, snakes, and crocodylomorphs), including numerous terrestrial mammals (allotherian, metatherians, xenarthrans, South American native ungulates [i.e., astrapothere, litoptern, notungulates, and pyrotheres], and rodents). In addition, CTA-27 and CTA-66 have also yielded bat teeth (Antoine et al., 2016). In the Contamana section, fossil material of each locality was collected by screen-washing (1 mm mesh): ~205 kg of sediment for CTA-47 (in 2010 and 2011), 97 kg of sediment for CTA-51 (in 2010 and 2011), 11 kg of sediment for CTA-73 (2014), 665 kg of sediment for CTA-27 (from 2008 to 2010), 31 kg of sediment for CTA-66 (in 2013 and 2014), and 270 kg of sediment for CTA-29 (from 2011 to 2013). During the last field expedition in the Contamana area (August 2016), our team regrettably noted the irremediable loss of several concerned localities (CTA-51, CTA-27, CTA-73, and CTA-29), either due to landslides, erosion, and silting caused by flooding events that have occurred since 2014 (Appendix 1).
 All field missions were performed as part of an International Specific Agreement elaborated between the “Museo de Historia Natural, Universidad Nacional Mayor de San Marcos,” Lima, Peru (MUSM) and the “Université de Montpellier” (Institut des Sciences de Montpellier [ISE-M]). The fossil material described in this paper is permanently housed in the paleontological collections of the MUSM.
All field missions were performed as part of an International Specific Agreement elaborated between the “Museo de Historia Natural, Universidad Nacional Mayor de San Marcos,” Lima, Peru (MUSM) and the “Université de Montpellier” (Institut des Sciences de Montpellier [ISE-M]). The fossil material described in this paper is permanently housed in the paleontological collections of the MUSM.
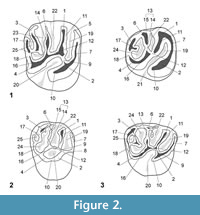
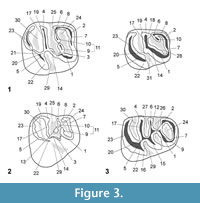
The studied material was identified preliminarily in Antoine et al. (2016). As some determinations have evolved since then, synonymies are specified. The terminology for rodent dentition (Figure 2, Figure 3) follows the nomenclature of Boivin et al. (2017) based on Wood and Wilson (1936), Fields (1957), Marivaux et al. (2004, 2017), and Antoine et al. (2012). Lower case letters are used for the lower dentition (i.e., dp, for decidual premolar; p, for premolar; m, for molar) and upper case letters for the upper dentition (i.e., dP, for decidual premolar; P, for premolar; M, for molar). The caviomorph taxa used for comparison in this study are listed in Appendix 2. When fossils from the Eocene localities of Contamana are compared with several taxa, the latter are primarily listed according to their chronostratigraphic order (from the oldest to the latest) and then alphabetically if they are coeval. All measurements are given in mm (Appendix 3). Two ratios, HIg and Hlb, deriving from the hypsodonty indice of Janis (1986), have been calculated from the maximum lingual crown height (Hg) and the labial height (Hb) of the less worn teeth available, respectively. As the hypsodonty indices of measured teeth are inferior to 1, most specimens can be considered as brachydont. Depending on specimens, photographs were taken with two scanning electron microscopes (SEM): HITACHI S 4000 and HITACHI S 4800.
 Institutional Abbreviations
Institutional Abbreviations
LACM, Los Angeles County Museum, Los Angeles, USA; MACN, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina; MLP, Museo de Ciencias Naturales de La Plata, La Plata, Argentina; MNHN, Musée national d'Histoire naturelle, Paris, France; MUSM, Museo de Historia Natural de la Universidad Nacional Mayor San Marcos, Lima, Peru; UM, Université de Montpellier, Montpellier, France.
Other Abbreviations
Hb, maximum labial crown height; Hg, maximum lingual crown height; HIb, hypsodonty index calculated from Hb; HIg, hypsodonty index calculated from Hg; ML, maximum anteroposterior length; MW, maximum transversal width; S, south; SALMA, South American Land Mammal Age; W, west.
SYSTEMATIC PALEONTOLOGY
Nomenclatural Remark
The new species and genera described below must be referred to Boivin, 2017, following the article 50.1 and the “recommendation 50A concerning multiple authors” of the International Code of Zoological Nomenclature (ICZN, 1999: 52, 182).
Order RODENTIA Bowdich, 1821
Infraorder HYSTRICOGNATHI Tullberg, 1899
Parvorder CAVIOMORPHA Wood, 1955
Superfamily CAVIOIDEA Fischer de Waldheim, 1817
Genus EOBRANISAMYS Frailey and Campbell, 2004
Type species.Eobranisamys romeropittmanae Frailey and Campbell, 2004.
Species content. The type species mentioned above and Eobranisamys riverai Frailey and Campbell, 2004; Eobranisamys javierpradoi nov. sp. (this work).
Geographic and stratigraphic distribution. Contamana (CTA-27 and CTA-66 localities), Pozo Formation, lower member (late middle Eocene), Loreto department, Peru; Santa Rosa (LACM 6289 locality), “Yahuarango Formation” (?late Eocene/early Oligocene), Ucayali department, Peru.
Emended generic diagnosis. Occlusal pattern similar to that of Branisamys by its tetralophodont and non-taeniodont lower molars; pentalophodont and taeniodont dP4s and upper molars with a strong third transverse crest and metaloph, which is lingually connected to the posteroloph. Eobranisamys differs from Branisamys in having a lower crown, thin loph(-id)s, and identifiable cusp(-id)s (modified after Frailey and Campbell, 2004, p. 79).
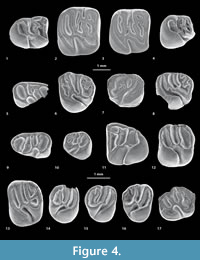
Eobranisamys javierpradoi sp. nov.
Figure 4.7-8, Appendix 3
zoobank.org/54337122-7076-48E5-81CC-83D8A1435825
 2012 Eobranisamys sp. Antoine et al., p. 1321-1322.
2012 Eobranisamys sp. Antoine et al., p. 1321-1322.
2016 Eobranisamys sp. Antoine et al., Supplementary data, p. 7.
2017 Eobranisamys sp. Antoine et al., Supplementary data, p. 9.
Etymology. In honour of Javier Prado, who founded the “Museo de Historia Natural, Universidad Mayor de San Marcos” in Lima, Peru, in 1918.
Holotype. MUSM 1897, left M1 (in Antoine et al., 2012, figure 2b’). Deposited in the "Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos”, Lima, Peru.
Referred material. In addition to the holotype (MUSM 1897) ̶̶ MUSM 2795, left m1 or m2; MUSM 1898, right fragmentary m2 (in Antoine et al., 2012, figure 2c’); MUSM 1899, left m3 (in Antoine et al., 2012, figure 2d’); MUSM 2796, right m3; MUSM 2797, right dP4 (Figure 4.7); MUSM 1896 (in Antoine et al., 2012, figure 2a’) and 2798, left P4s; MUSM 2799, right M1 or M2; MUSM 2800, left M1; MUSM 2801, left M3 (Figure 4.8).
Type Locality. Contamana CTA-27, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2012, 2016).
Diagnosis. Smallest species of Eobranisamys (about half the size of E. romeropittmanae and 20% smaller than E. riverai), showing sharper crests, more salient cusps, and longer M3 with a less rounded occlusal outline than other referred species.
Description. The lower molars are tetralophodont (in Antoine et al., 2012, figure 2c’-d’). Mesially, there is no trace of anterocingulid at the base of the crowns. On MUSM 1899 (m3; in Antoine et al., 2012, figure 2d’), the protoconid is the largest cuspid of the tooth. On MUSM 1899 and 2795 (m1 or 2; in Antoine et al., 2012, figure 2c’), the metaconid is situated more mesially than the protoconid, with a strong and high posterior arm. The protoconid and metaconid are connected by a strong and complete metalophulid I. On m3, the second transverse cristid is strong and complete. Labially, this cristid starts from a well-defined and oblique (distolingually oriented) posterior arm of the protoconid, then it makes an angle and runs lingually to reach the distal extremity of a long posterior arm of the metaconid. The same is true on m1-2s. On MUSM 1898 (in Antoine et al., 2012, figure 2c’), a noticeable angulation in the middle of this cristid could correspond to the junction of two cristids. Furthermore, on this tooth, the lingual connection of this second cristid could be a mesostylid. So, the question remains as to whether this second cristid is a complete metalophulid II or a combination of two cristids, i.e., a short posterior arm of the protoconid and a lingual neomesolophid. The metalophulid I and second transverse cristid are well-separated, and isolate a broad and oval anterofossettid. The ectolophid is very short, medial (at the center of the tooth), and fully or roughly longitudinal. There is no mesoconid. The hypoconid and entoconid are labiolingually opposed. The hypolophid, strong and oblique (slightly forwardly oriented), connects to both a thin, oblique and moderately long consists of two cristulids. The hypoconid is mesiodistally compressed. The posterolophid on m2 and m3 is massive but short. It is clearly separate from the entoconid on MUSM 1898 and 1899; it reaches the base of that cuspid on MUSM 2796. In any case, the posteroflexid remains open lingually.
Many upper teeth are referable to this taxon: one dP4, two P4s, and nine molars.
The dP4 (MUSM 2797; Figure 4.7) is damaged and highly worn. It is subrectangular, with a mesiodistal long axis. Only the mesiodistally elongated mesostyle is clearly distinct on the labial margin. The mesostyle is mesially separate from the paracone but distally connected to the metacone. The dP4 is pentalophodont and taeniodont (absence of lingual protoloph). The anteroloph is markedly curved, strongly connected to the protocone, and runs labially to reach and connect to the paracone mesiolabially. The mure, oblique and aligned with the anterior arm of the hypocone, reaches mesially the short and transverse protoloph. The third transverse crest, interrupted labially and not reaching the mesostyle, is identified as a mesolophule. There are two accessory enamel crestules: one on the mesolophule and one on the anterior arm of the hypocone. The metacone is crestiform (mesiodistally elongated) and connected to the posteroloph distolabially. The metaloph, connected labially to the metacone, runs lingually and bends distally near the midline of the crown to link the posteroloph.
The P4s are oval in occlusal view, with a tetralophodont pattern (MUSM 1896 and 2798; in Antoine et al., 2012, figure 2a’). The paracone and metacone are twinned on MUSM 1896, whereas they are less connected on MUSM 2798. The protocone is mesiodistally developed and lingually opposed to the labial paracone-metacone complex. There is a minute hypocone (distinct on MUSM 2798; broken on MUSM 1896), located distolabially to the protocone. These two lingual cusps, also twinned, form a protocone-hypocone complex. The paracone-metacone and protocone-hypocone complexes, associated with long and strongly developed posteroloph and anteroloph, design a circular enamel wall on the entire crown margin. On MUSM 1896 (in Antoine et al., 2012, figure 2a’), there is only a small depression on this wall, located mesially, at the level of the anteroloph-paracone junction. This depression is absent on MUSM 2798. The protoloph is short and limited to its labial part. There is no lingual protoloph: the tooth is taeniodont. The hypocone has an oblique anterior arm ending near the center of the tooth. This hypocone arm extends labially via a straight and long third transverse crest (mesolophule on MUSM 1896 and mesolophule/mesoloph? on MUSM 2798). There is a tiny enamel spur mesiolingually directed at the junction between the hypocone arm and the third transverse crest.
The M1s referred here to Eobranisamys (MUSM 1897 and 2800; in Antoine et al., 2012, figure 2b’) are subquadrate, while the M3 (MUSM 2801; Figure 4.8) is smaller and shows a more rounded occlusal outline. On all upper molars, the protocone appears mesiodistally opposed to the hypocone. However, the hypocone is more reduced with respect to the protocone on M3 than on M1. There is no significant difference of crown height with upper molars of other rodent species from CTA-27. All upper molars are fully pentalophodont. The main cusps are sometimes faintly visible, but still distinct. These upper molars are fully taeniodont. The paraflexus is merged with the hypoflexus to form a continuous labiolingual groove, without any vestige of lingual protoloph. The protocone is slightly labiolingually pinched and connected to a strong anteroloph. This mesial transverse crest runs labially and connects to a well-defined parastyle. There is no connection between the parastyle and the paracone, even if they are very close on MUSM 2799 (where the paraflexus is almost completely labially closed). The hypocone is slightly labiolingually pinched. Its anterior arm is strong and frequently runs labiomesially to join the lingual extremity of the interrupted protoloph. Thus, the mure is present but rather indistinct except on MUSM 2800 where it is long and mesiolabially oriented. The third transverse crest is strongly connected to a well-defined and isolated mesostyle. This crest is a composite of either equally developed mesolophule and mesoloph (MUSM 2800), or of a long mesoloph with a short spur-like mesolophule (MUSM 1897 and 2801; in Antoine et al., 2012, figure 2b’). In addition, the mesostyle is separate from the paracone and metacone by two notches (the posterior arm of the paracone and anterior arm of the metacone are absent or weakly developed). On all upper molars, the metacone is labiolingually compressed and oblique. The metaloph is strong but short and backwardly directed in its lingual end, and it is variably connected to the medial part of the posteroloph (i.e., from slightly [MUSM 2800 and 2801] to strongly [MUSM 1897] connected). With such a configuration, the lingual part of the posteroflexus is confluent with the metaflexus. The metaloph and posteroloph isolate a small and oval posterofossette (the remaining labial part of the posteroflexus). The posteroloph is low compared to other transverse crests, and it links the distal aspects of the hypocone and of the metacone. In addition, the posteroloph bears a labial secondary crestule on M3 (Figure 4.8).
Comparisons. This new species of Eobranisamys is roughly similar in size to the sympatric Cachiyacuy contamanensis. The transverse crests and cristids are well-marked and slightly higher than in molars of species referred to Cachiyacuy or Canaanimys. Teeth studied here display features reminiscent of those of Eobranisamys and Branisamys Hoffstetter and Lavocat, 1970 (especially upper molars; Lavocat, 1976; Frailey and Campbell, 2004). This is particularly shown in the development of a taeniodont and pentalophodont pattern, characterized by the presence of a strong third transverse crest, and a strong and well-defined metaloph, which is transverse and connected to the posteroloph. As in Eobranisamys romeropittmanae and E.riverai, the teeth of E. javierpradoi nov. sp. are much lower crowned than teeth of Branisamys. However, E. javierpradoi differs substantially from E. romeropittmanae in being about half the size, and from E. riverai in being 20% smaller, and in showing sharper crests and more salient cusps. Besides, E. romeropittmanae and E. riverai tend to have M3 shorter and with a more rounded occlusal outline than E. javierpradoi.
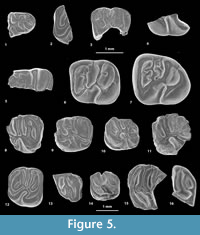
Eobranisamys sp.
Figure 5.15-16, Appendix 3
2016 Caviomorpha indet. Antoine et al., Supplementary data, p. 8.
 2017 Caviomorpha indet. Antoine et al., Supplementary data, p. 9.
2017 Caviomorpha indet. Antoine et al., Supplementary data, p. 9.
Referred material. MUSM 2841, fragmentary left upper molar? (Figure 5.15); MUSM 2842, fragmentary left upper molar (Figure 5.16).
Locality. Contamana CTA-66, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Description. MUSM 2841 (Figure 5.15) could be a fragment of an upper molar, upon which it can be recognized a part of the anteroloph, the protoloph, the mure, the third transverse crest, and a mesial part of the anterior arm of the hypocone. The crests are very thick. The lingual protoloph is reduced to a short crestule, stemming from the labial branch of the protoloph and forwardly directed, but without connection with the protocone (pseudo-taeniodont pattern). The paracone displays short anterior and posterior arms connecting to the labial end of the anteroloph and the posterior arm of the mesostyle, respectively, thereby involving the labial closure of both the paraflexus and mesoflexus. The protoloph runs from the paracone to an oblique mure. The third transverse crest, labially linked to a strong mesostyle, is thin at the level of its lingual connection with the anterior arm of the hypocone. Such a pattern suggests that the third transverse crest corresponds to a long mesoloph, and possibly to a very short mesolophule lingually. The protoloph and third transverse crest are roughly parallel and slightly oblique.
MUSM 2842 (Figure 5.16) is a labiodistal fragment of a medium-sized upper molar, upon which it can be easily recognized labially, the mesostyle and metacone, and distolingually, the hypocone. The mesostyle is particularly strong, appearing as large as the metacone and hypocone. The mesostyle is separate from the metacone by a narrow but deep notch, while the metacone is strongly connected to the posteroloph. The third transverse crest is strong, straight, and well-connected to the mesostyle, but it is distinct from the mesial end of the anterior arm of the hypocone. This pattern suggests that it is only formed by a mesoloph. The metaloph is well-defined, long, and well-curved. It runs lingually from the metacone, then turns backwardly and connects to an accessory cusp of the posteroloph mediolingually (just below the middle line of the crown). The metaloph, mesoloph, and posteroloph remain well-separated. The hypocone displays a very long and oblique anterior arm. The posteroloph is strong and gently curved. The flexi are deep. The most posterior flexus (labial part of the posteroflexus) is entirely closed and forms a deep and oval posterofossette.
Comparisons. On MUSM 2842 (Figure 5.16), the presence of a strong and non-oblique third transverse crest, as well as a well-defined, long, and curved backwardly metaloph, which is lingually connected to the posteroloph, in addition to the brachydonty, are characters found in three Paleogene genera: Cachiyacuy, Eosallamys Frailey and Campbell, 2004, and Eobranisamys. In Cachiyacuy and Eosallamys, the development and connections of the metaloph are highly variable. The metaloph can be long, reduced, or almost absent. It is totally absent on two upper molars of Cachiyacuy : on MUSM 2744 of C. contamanensis and on MUSM 2785 of C. kummeli (Figure 4.12). When it is present in Cachiyacuy and Eosallamys, it can be connected either to the anterior arm of the hypocone, to the third transverse crest, or to the posteroloph. In Eobranisamys, the metaloph is present, well-defined, well-separated from its adjacent mesial and distal transverse crests (third transverse crest and posteroloph, respectively), and it is always connected to the posteroloph. As for MUSM 2842, the mesostyle is always labially separate from the metacone in Eosallamys and Eobranisamys, while in Cachiyacuy, these two cusps can be linked. On MUSM 2842, the development and configuration of the metaloph generate a large, deep, and rounded posterofossette (labial part of the posteroflexus), a feature characteristic of Eobranisamys. In C. contamanensis and Eosallamys, this posterofossette is smaller and shallower. This fragmentary tooth from CTA-66 is therefore likely to be the distal part of an upper molar of an Eobranisamys -like taxon. The size of the specimens is closer to the species of Eobranisamys from CTA-27 (E. javierpradoi) and Eobranisamys riverai than to Eobranisamys romeropittmanae. On MUSM 2842, as for E. romeropittmanae and E. riverai, the posterofossette is larger than in E. javierpradoi. As for E. javierpradoi, MUSM 2842 differs from E. romeropittmanae and E. riverai in showing sharper crests and more salient cusps.
Superfamily OCTODONTOIDEA Waterhouse, 1839
Genus EOESPINA Frailey and Campbell, 2004
Type species.Eoespina woodi Frailey and Campbell, 2004
Species content. Only the type species.
Geographic and stratigraphic distribution. Contamana (CTA-51 locality), Pozo Formation, lower member (late middle Eocene), Loreto department, Peru; Santa Rosa (LACM 6289 locality), “Yahuarango Formation” (?late Eocene/early Oligocene), Ucayali department, Peru.
Eoespina sp.
Figure 5.12, Appendix 3
2016 Cachiyacuy aff. kummeli Antoine et al., Supplementary data, p. 5.
2017 Cachiyacuy aff. kummeli Antoine et al., Supplementary data, p. 9.
Referred material. MUSM 2655, right M2 (Figure 5.12).
Locality. Contamana CTA-51, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Description. The M2 (MUSM 2655; Figure 5.12) is worn but all its occlusal structure is well-recognizable, with a subquadrate outline. The hypocone is slightly displaced labially and smaller than the protocone, thereby indicating that this tetralophodont tooth is a M2. The transverse crests (anteroloph, protoloph, third transverse crest, and posteroloph) are subparallel, with a slightly oblique protoloph. Both the anteroloph and posteroloph are massive and strongly connected to the lingual and labial cusps. The tooth is not taeniodont. The mure is short and almost longitudinal. Labially, there is a large mesostyle, twinned with the metacone and strongly connected to the lingually thinning third transverse crest (therefore interpreted as a mesoloph). A deep and narrow notch separates the mesostyle from the paracone, and thus the mesoflexus remains open labially. The mesoflexus is particularly narrow due to the close proximity of the mesoloph with the protoloph. Labially, the metacone is virtually indistinct and entirely incorporated within the labial posteroloph. The mesial enamel edge of the posteroloph expends mesially, and forms an enamel platform-like surface. This structure might correspond to a relic of the metaloph. The paraflexus and the posteriormost flexus (fused meta- and posteroflexus) are close lingually. The hypoflexus is mesiodistally constricted but remains open.
Comparisons. This specimen has a size comparable to that of teeth of Cachiyacuy kummeli,Canaanimys maquiensis, Eoespina woodi Frailey and Campbell, 2004, and Eosachacui lavocati Frailey and Campbell, 2004. Except for Canaanimys, the concerned upper molars have a pattern close to that of MUSM 2655: non-taeniodont and with a metaloph reduced or absent. In Eosallamys, upper molars are non-taeniodont but their metaloph is still present and long. The subquadrate occlusal outline of MUSM 2655 and the strong connections of the anteroloph and posteroloph to the labial cusps, respectively, better match the conditions found in Eoespina. Given the scarcity the available material, we provisionally assign this tooth to Eoespina sp.
Remarks. Following Frailey and Campbell (2004), Eoespina woodi and Eosachacui lavocati, both found at Santa Rosa, are two close taxa sharing many similarities. This is particularly shown in their brachydonty, non-taeniodonty, subquadrate upper molars with rounded corners, and pentalophodont/tetralophodont upper molars with a very reduced or absent metaloph. Frailey and Campbell (2004, p. 88-91) described seven characters distinguishing both taxa. However, these characters are not found in all specimens referred to each taxon and can result from an intraspecific variation. A taxonomic revision of both taxa would hence be required, notably in considering their possible synonymy. According to Frailey and Campbell (2004, p. 88-91):
1. Eoespina is slightly smaller than Eosachacui. However, the size range of its dental variation matches that of Eosachacui;
2. Contrary to Eoespina, upper molars of Eosachacui often display additional spurs or cuspules in their metaflexus. However, this addition of enamel structures is only limited to three specimens (LACM 143292, 143387, and 143401; Frailey and Campbell, 2004, p. 128);
3. In Eosachacui, the deflections of the protoloph are less severe than those observed in Eoespina. These deflections of the protoloph in Eosachacui probably correspond to the obliquity of the protoloph, which is more pronounced (strongly oblique) and tends to be in line with the mure. However, some upper molars of Eosachacui have a transverse protoloph (LACM 143387 and 143388; Frailey and Campbell, 2004, p. 128), while this crest can be slightly oblique in Eoespina (e.g., LACM 143286 and 149436; Frailey and Campbell, 2004, p. 124-125);
4. Contrary to Eosachacui, lower molars of Eoespina display accessory cristulids and cuspids in their anteroflexid. Nevertheless, one lower molar of Eosachacui shows these secondary structures (LACM 143325; Frailey and Campbell, 2004, p. 129). Interestingly, the presence or absence of these structures is also observed in Cachiyacuy contamanensis, but that remains very variable;
5. In lower molars of Eoespina, the second transverse cristid (named “metalophid” by Frailey and Campbell, 2004, p.91) is not uniform either in height or thickness. Indeed, the second transverse cristid appears as a combination of two cristids (i.e., neomesolophid and posterior arm of the protoconid), more or less developed depending on the specimens. From our personal observation of the Santa Rosa specimens figured in Frailey and Campbell (2004, p. 124-130), this apparent composite second cristid in Eoespina is also found in lower molars of Eosachacui ;
6. The second transverse cristid and hypolophid (named “protolophid” in Frailey and Campbell, 2004, p. 91) are mesiodistally closer in Eosachacui than in Eoespina. However, in both genera, these two cristids are variably spaced, which implies a variation of the shape and size of the flexids (anteroflexid and mesoflexid); and
7. Finally, in lower molars of Eosachacui, the hypolophid is transverse or slightly backwardly directed, while it is transverse or slightly forwardly directed in Eoespina. However, as noticed by Frailey and Campbell (2004, p. 91), this character is highly variable.
cf. Eoespina sp.
Figure 4.17, Appendix 3
Referred material. MUSM 1913 (in Antoine et al., 2012, figure 2f’) and 2802 (Figure 4.17), left M2s; MUSM 1912, right M2 (in Antoine et al., 2012, figure 2e’).
Locality. Contamana CTA-27, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2012, 2016).
Description. Only three minute upper molars can be referred to cf. Eoespina sp. (MUSM 1912, 1913, and 2802; in Antoine et al., 2012, figure 2e’-f’; Figure 4.17). They have a rounded crown outline in occlusal view. These teeth are primarily tetralophodont with strong and long third transverse crest, antero-, proto-, and posteroloph (no metaloph). The cusps are still well-defined. There is no parastyle. Mesially, a strong anteroloph runs from the protocone to the mesiolabial aspect of the paracone. The teeth are not taeniodont and display a short and longitudinal mure, which is slightly situated lingually to the medial axis of the crown. There is a strong but short anterior arm of the hypocone that connects the mure. The hypocone is more labial and reduced with respect to the protocone. The metacone is mesiodistally elongated and slightly more lingual than the paracone, and it is merged with a strong mesostyle. The paracone and mesostyle are only separate by a narrow but deep notch, and the mesoflexus remains open labially as a result. A strong and continuous third transverse crest runs from the mesial extremity of the anterior arm of the hypocone to the mesostyle. The protoloph and third transverse crest are nearly parallel. On the distolabial region of MUSM 1913 (in Antoine et al., 2012, figure 2f’), it may occur a vestigial metaloph, very low, short, backwardly directed, and practically subsumed within the posteroloph. On all teeth, the hypoflexus is narrow and somewhat constricted lingually.
Comparison. Teeth of this taxon are slightly smaller than those of Canaanimys maquiensis. These three diminutive upper molars from CTA-27 exhibit a tetralophodont/non-taeniodont pattern, with strong and long third transverse crest, antero-, proto-, and posteroloph (no metaloph or very reduced), as it can be observed in “?Vallehermosomys merlinae Vucetich et al., 2010," Sallamys Hoffstetter and Lavocat, 1970 or Eoespina/Eosachacui. The general morphology of these upper teeth from CTA-27, with notably a strong and long posterior arm of the paracone and a rounded crown outline, recall upper molars of E. woodi/E. lavocati. However, these specimens from CTA-27 differ from E. woodi / E.lavocati in having slightly more transverse upper molars, usually characterized by less inflated labial cusps. These upper molars have a longer and more mesially positioned third transverse crest contrary to the condition observed on the M3 of ?Vallehermoso mysmerlinae. Indeed, the latter displays a wider mesoflexus, and displays a third transverse crest that is clearly more displaced distally and close to the posteroloph. These upper molars from CTA-27 also differ from those of Sallamys in being lower-crowned, and in having a stronger third transverse crest, which always connects to the anterior arm of the hypocone.
CAVIOIDEA Fischer de Waldheim, 1817 or CHINCHILLOIDEA Bennett, 1833
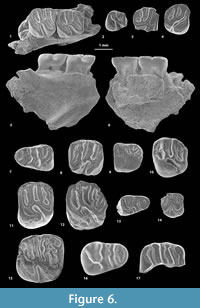
Gen. et sp. indet.
Figure 6.15-17, Appendix 3
2016 Eobranisamys sp. Antoine et al., Supplementary data, p. 9.
 2016 Cachiyacuy cf. contamanensis Antoine et al., Supplementary data, p. 9.
2016 Cachiyacuy cf. contamanensis Antoine et al., Supplementary data, p. 9.
2016 Caviomorpha indet., sp. 1 Antoine et al., Supplementary data, p. 9.
2017 Eobranisamys sp. Antoine et al., Supplementary data, p. 9.
2017 Cachiyacuy cf. contamanensis Antoine et al., Supplementary data, p. 9.
2017 Caviomorpha indet. sp. 1, Antoine et al., Supplementary data, p. 9.
Referred material. MUSM 2834, right dp4 (Figure 6.16); MUSM 2835, right m2 (Figure 6.15); MUSM 2836-2837, fragmentary right upper molars (Figure 6.17).
Locality. Contamana CTA-29, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Description. The dp4 (MUSM 2834; Figure 6.16) is relatively larger compared to other dp4s found at CTA-29. This tooth is pristine and brachydont, with slender transverse cristids, and small but well-defined and acute mesostylid, entoconid, and hypoconid. The metaconid and protoconid are much more crestiform. This deciduous lower premolar is pentalophodont. Mesially, the metalophulid I, well-curved, connects to the protoconid labially and ends and the base of the metaconid lingually. The metaconid, mesiodistally pinched, is faintly linked to the crestiform protoconid via its posterior arm, which forms a quasi-complete metalophulid II. There is a minute enamel cuspid on the anteroflexid at the base of the distal wall of the metalophulid I. The metaconid does not develop a posterior arm. In the middle part of the tooth, the lingual margin bears a well-defined and isolated mesostylid. The mesial ectolophid is almost transverse and appears in continuity with a mesolophid, which extends lingually to the mesostylid. Labially, the mesial and sub-transverse ectolophid displays an enamel swelling, which could be interpreted as a neomesoconid-like cuspid. The distal ectolophid is small and very low. As such, the hypoflexid and distal mesoflexid are virtually confluent. In this context, the third transverse cristid has no link with the surrounding cristids, and the hypoflexid-distal mesoflexid and mesial mesoflexid are transversely open. The hypoconid is the largest cuspid of the tooth, labially opposed to the entoconid. Both cuspids are linked by a thin and sinuous hypolophid. The second transverse cristid, mesolophid, and hypolophid are parallel and slightly curved. The anterior arm of the hypoconid is faintly marked, lower and grooved, and it is weakly connected to the hypolophid (i.e., pseudo-taeniodont). Distolabially, the posterolophid faintly links the hypoconid, distolingually ending its course far from the entoconid. Thus, the broad metaflexid remains open lingually.
The m2 (MUSM 2835; Figure 6.15) is corroded and damaged, with a dental structure still visible. This tetralophodont tooth is particularly large with a trigonid as wide as the talonid. Two enamel wrinkles are directed backwardly to the metalophulid I, and one directed forwardly to the second transverse cristid. Due to wear, it is difficult to tell if this second cristid was connected or not to the mesostylid. This cristid could be a complete metalophulid II or a composite cristid made by a labial short posterior arm of the protoconid associated with a short neomesolophid. The ectolophid is oblique and distally connected both to a short but strong anterior arm of the hypoconid (i.e., non-taeniodont pattern) and to the transverse and strong hypolophid. The posterolophid is lingually separate from the entoconid. The mesoflexid and posteroflexid are lingually open.
Both labial fragments (MUSM 2836 and 2837; Figure 6.17) document large upper molars. The labial cusps are well-defined: parastyle, paracone, mesostyle, and metacone. MUSM 2836 is tetralophodont (Figure 6.17): there is no trace of metaloph between the mesolophule and the posteroloph. The metacone is the largest cusp, at the labial end of the posteroloph. On MUSM 2836, all transverse flexi are labially open, whereas the metaflexus is labially closed on MUSM 2837 (more worn).
Comparisons. The morphology of MUSM 2834 (Figure 6.16) is reminiscent of that found in dp4s of Eobranisamys from Santa Rosa (Peru, ?late Eocene/early Oligocene; Frailey and Campbell, 2004) and Branisamys from Salla (Bolivia, late Oligocene; Hoffstetter and Lavocat, 1970; Lavocat, 1976), notably in lacking the connections between the second transverse cristid (metalophulid II) and the mesolophid (or with their associated cuspids). However, in Eobranisamys and Branisamys, there is a distal ectolophid between the mesolophid and the hypolophid, and a labial connection between the metalophulid I and the metalophulid II. MUSM 2834 is pentalophodont whereas the dp4s of Eobranisamys and Branisamys are hexalophodont, in showing the addition of a neolophid between the metalophulid I and the metalophulid II. MUSM 2835 (Figure 6.15) is similar in dental size to Eobranisamys , while Branisamys is significantly larger. That m2 has a pattern close to that of lower molars of Eobranisamys, which can have accessory cristids on the metalophulid I and second transverse cristid. Although the size of the fragmentary upper molars (MUSM 2836 and 2837; Figure 6.17) is compatible with that of Eobranisamys, the former are tetralophodont while Eobranisamys and Branisamys have pentalophodont upper molars, with a strong metaloph. To sum up, this taxon might show close affinities to Eobranisamys, and to a lesser extent to Branisamys. However, the particular characteristics of the dp4 and the tetralophodont pattern of upper molars do not allow for a formal generic assignment. Branisamys was initially attributed to the Dasyproctidae (i.e., Cavioidae) by Lavocat (1976), but this genus may be more closely related to the Chinchilloidea following Kramarz et al. (2013) and Vucetich et al. (2015). Eobranisamys was originally assigned to the Cavioidea (Frailey and Campbell, 2004), but its alleged close relationship with Branisamys would question, in turn, the suprafamilial attribution of Eobranisamys. In the present state, we provisionally refer the material of CTA-29 to as “Cavioidea or Chinchilloidea indet.,” although a critical revision of Eobranisamys and Branisamys would be necessary in that purpose.
Superfamily indet.
Genus POZOMYS gen. nov.
zoobank.org/903578CB-4B98-4430-AF8B-5C30839265D5
Type species. Pozomys ucayaliensis, sp. nov.
Species content. Only the type species.
Etymology. From the Pozo Formation, which includes the CTA-29 locality, and mŷs, Greek for mouse.
Generic diagnosis. As for the type and only species.
Pozomys ucayaliensis sp. nov.
Figure 6.1-6, Appendix 3
zoobank.org/E38E9E2A-6303-4278-888F-0F8FC05C5010
2016 Canaanimys sp. Antoine et al., Supplementary data, p. 9.
2016 Cachiyacuy cf. kummeli Antoine et al., Supplementary data, p. 9.
2016 Chinchilloidea indet. Antoine et al., Supplementary data, p. 9.
2017 Canaanimys sp. Antoine et al., Supplementary data, p. 9.
2017 Cachiyacuy cf. kummeli Antoine et al., Supplementary data, p. 9.
2017 Chinchilloidea indet. Antoine et al., Supplementary data, p. 9.
Etymology. Refers to the Ucayali River, which is a major tributary of the Amazon River flowing near Contamana, Peru.
Holotype. MUSM 2833, right M2. Deposited in the Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos, Lima, Peru.
Referred material. In addition to the holotype (MUSM 2833) ̶ MUSM 2821, left p4 (Figure 6.2); MUSM 2820, fragmentary right p4; MUSM 2822, right mandibular fragment bearing m1 and m2 (Figure 6.1, 6.5-6); MUSM 2833, right M2 (Figure 6.4); MUSM 2819, fragmentary right M3 (Figure 6.3).
Type Locality. Contamana CTA-29, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Diagnosis. Tiny rodent characterized by tetralophodont p4s and lower molars. Pozomys differs from Eoincamys in having no posterior arm of the protoconid (or very reduced one) on p4, and less oblique transverse cristids on the lower molars. Differs from Cachiyacuy and Canaanimys in having a residual metaloph, which can be merged with the metacone-posteroloph complex on upper molars. Differs from Cachiyacuy, Eoespina/Eosachacui and Incamys in having a metalophulid I more or less disconnected to the protoconid on lower molars. Its teeth tend to be taeniodont, contrary to Eoespina/Eosachacui and most teeth of Cachiyacuy. Differs from Incamys in being lower-crowned and in showing a similar thickness of the enamel layer on the mesial and distal flanks of the hypoflexus. Differs from Platypittamys and Deseadomys in having a p4 with a hypolophid. Differs from Galileomys and Paulacoutomys in having a very thin lateral crest, which does not reaches the masseteric crest below the m1 on mandible.
Description. The nearly complete p4 (MUSM 2821; Figure 6.2) is moderately eroded, and shows rounded corners. The shape of this premolar is characterized by a talonid, which is nearly twice wider than the trigonid. Despite of the wear, the four main cuspids (metaconid, entoconid, protoconid, and hypoconid) are well-recognizable, as they are well-defined. Although the tooth is slightly broken mesially (it lacks the enamel), the metaconid and protoconid appear well-separated and connected mesially by a transverse and straight metalophulid I. The rounded protoconid seems to display a short posterior arm, which is in connection and in line with a strongly oblique ectolophid. The hypolophid is also oblique and in line with the ectolophid. These two cristids form a distolingual-mesiolabial directed central and diagonal cristid, which links the entoconid and protoconid (its posterior arm?). Lingually, the mesostylid is faintly visible to indistinct and linked to a short and low posterior arm of the metaconid. The second transverse cristid is short, straight, and strikingly labiodistally directed. This second transverse cristid is lingually linked to the posterior arm of the metaconid-mesostylid complex, and labially to the ectolophid, and as such it could represent a neomesolophid rather than a metalophulid II. Labiodistally, the hypoconid is massive but crestiform (mesiodistally pinched), and bears a noticeably long labial outgrowth. Distolabially, the hypoconid is entirely merged with the strong posterolophid. This latter cristid runs lingually and connects to a short, well-marked, and high posterior arm of the entoconid. A very short, low, and thin anterior arm of the hypoconid is faintly linked to the hypolophid (the tooth is almost taeniodont). Therefore, the distal metafossettid is almost confluent with the hypoflexid, which exhibits a wide labial aperture. The fragment of p4 (MUSM 2820) is less eroded than the former one described above. This tooth fragment does not present any second transverse cristid. A thin and deep furrow separates the mesostylid from the metaconid.
MUSM 2822 is a mandibular fragment preserving m1 and m2 (Figure 6.1, 6.5-6). Labially, the broken masseteric crest is posteroventrally directed. It is widely prominent and reduced at its anterior tip, which probably ends below the premolar (p4). The part of the ascending ramus, which runs toward the coronoid process, begins below the m2. That mandibular fragment shows neither horizontal crest nor lateral crest nor notch for the insertion of the tendon of the zygomatico-mandibularis pars infraorbitalis. The m1 has a trigonid narrower than the talonid, whereas the trigonid is roughly as wide as the talonid on m2. Although the occlusal surface of the molars is worn, their cuspids are still well-recognizable, notably on m2. Both teeth are tetralophodont. The metalophulid I is very thin in its labial part and it is clearly separate from the protoconid by a wide notch on m2. On both molars, the posterior arm of the metaconid is long and reaches a well-defined mesostylid. The latter is well-separated from the entoconid. The short second transverse cristid is straight and complete but constricted in its middle part, which suggests that it could result from the coalescence of two cristids (lingually, a neomesolophid, and labially, a posterior arm of the protoconid). On m1, the pinching of the second cristid is labially located, which would indicate that the neomesolophid is dominant, whereas the two cristids are equal in length on m2. The hypolophid is strong and straight transversely. The second transverse cristid and hypolophid are parallel and mesiodistally close in position. Both isolate a labiolingually long and mesiodistally narrow furrow-like mesoflexid. The ectolophid, longitudinal, is short due to the advanced stage of wear. The anterior arm of the hypoconid, poorly developed (faintly visible and very low), separates the metaflexid from the hypoflexid. Yet, given the weak development and low elevation of the anterior arm of the hypoconid, we can describe this dental pattern as “pseudo-taeniodont.” The hypoconid is crestiform (mesiodistally pinched), with a long and labially oriented outgrowth. This outgrowth and the distal flank of the protoconid isolate a narrow and nearly transverse hypoflexid (notably on m2). Lingually, on both teeth, the entoconid does not develop an anterior arm, and thus the furrow-like mesoflexid remains open lingually. The metaflexid is lingually open on m1 but closed on m2.
MUSM 2833 (Figure 6.4) is slightly wider than long. The hypocone is reduced compared to the protocone, and displaced labially like on some M3s and M2s. This specimen has interstitial facets on its mesial and distal margins, which allows us to identify it as a M2. The protocone, massive and slightly oblique, displays a short but thick posterior outgrowth. Connected to the protocone, the anteroloph runs labially. It ends its course at the mesial base of the paracone, without connecting to it, thereby letting the paraflexus faintly open labially. The paracone is large and rounded, and much more differentiated than the metacone (nearly indistinct and subsumed within the posteroloph). From the paracone, the oblique labial protoloph runs distolingually and joins the mure-anterior arm of the hypocone complex, subparallel to the protoloph. The lingual protoloph is lacking, and thus the hypoflexus connects the paraflexus, thereby illustrating a full taeniodont pattern. Labially, at mid-distance between the paracone and metacone-posteroloph, the mesostyle is strong and well-defined, but only connected to the metacone-posteroloph complex by a strong longitudinal crest (posterior arm of the mesostyle and/or anterior arm of the metacone). The central transverse crest between the mesostyle and the mesial extremity of the anterior arm of the hypocone is sinuous, indicating that this crest could be a composite crest, including a short mesolophular spur and a long mesoloph. Distolabially, the posteroloph shows a distinct mesiodistal enlargement, thus suggesting the presence of a residual metaloph, backwardly oriented and merged with the posteroloph. The metaflexus is the unique flexus entirely closed labially.
The fragmentary MUSM 2819 (Figure 6.3) is subpentalophodont and non-taeniodont, with a rounded labial margin and a hypocone more labial than the protocone, which indicates that the specimen is a M3. The protoloph is slightly oblique, close and parallel to the anteroloph (narrow paraflexus). Although broken labiomesially, these two mesial crests seem not to be linked labially. The metacone is small, slightly more lingual than the paracone, and merged to the posteroloph. The mesostyle is almost as large as the paracone and isolated on the labial margin. Neither a posterior arm of the paracone nor an anterior arm of the metacone is developed, and the mesoflexus and metaflexus remain open labially. The hypocone displays a strong and sagittal anterior arm. It connects to the protoloph via a strong mure, although almost undifferentiated. The central transverse crest consists of two disjoint crests: lingually, a short mesolophule and labially, a long mesoloph stemming from the mesostyle. A short metaloph, stemming from the metacone, runs mesiolingually toward the labial extremity of the mesolophule, without being connected to it. Distolingually, the posteroloph is faintly connected to the hypocone, and a shallow notch separates both structures.
Comparisons. The size of these specimens is comparable to that of Cachiyacuy kummeli, Canaanimys maquiensis, Eoespina woodi/Eosachacui lavocati, and Eoincamys ameghinoi Frailey and Campbell, 2004. On the lateral view of the MUSM 2822 mandible (Figure 6.5), the general disposition and strong development of the masseteric crest on the dentary is found in all Paleogene caviomorphs for which the mandible is known (e.g., Eobranisamys, Andemys Bertrand et al., 2012, Platypittamys Wood, 1949, Scotamys Loomis, 1914, Cephalomys Ameghino, 1897, Branisamys, Incamys Hoffstetter and Lavocat, 1970, Sallamys, Migraveramus Patterson and Wood, 1982, Paulacoutomys Vucetich et al., 1993, Galileomys Vucetich and Kramarz, 2003, Acarechimys Patterson, 1965 (in Patterson and Wood, 1982), Leucokephalos Vucetich et al., 2015, Loncolicu Vucetich et al., 2015, and Llitun Vucetich et al., 2015). Contrary to MUSM 2822, Galileomys and Paulacoutomys develop a very thin lateral crest, stemming from the ascending ramus, and almost reaching the masseteric crest below the m1. The lower molars of Pozomys ucayaliensis nov. gen. et sp. display an association of characters that can be found in Canaanimys and/or in Eoincamys (Frailey and Campbell, 2004). The molars of the MUSM 2822 mandible show a metalophulid I that tends to be disconnected to the protoconid, a configuration which can be observed in lower molars of Eoincamys and Canaanimys. As on the m2 of MUSM 2822, the m1s and m2s of Eoincamys pascuali clearly show an interrupted metalophulid I, which is separate from the protoconid by a large notch. In Eoincamys ameghinoi and Canaanimys maquiensis , this feature is more variable: the metalophulid I can show the same configuration than the m2 of MUSM 2822 and Eoincamys pascuali (on LACM 149435 [in Frailey and Campbell, 2004, p. 114] and MUSM 1893, 2788-2789 [in Antoine et al., 2012, figure 2x], respectively), but also it can be complete in linking the metaconid to the protoconid (on LACM 143435 [in Frailey and Campbell, 2004, p. 114] and MUSM 2787, 2790-2791, respectively). In Eoincamys, the second transverse cristid is almost limited to its lingual part (neomesolophid), which tends to be disconnected to a very short, spur-like posterior arm of the protoconid. In Canaanimys, the second transverse cristid is either complete or discontinuous (split into two or three parts). The molars of the MUSM 2822 mandible are characterized by a pseudo-taeniodont pattern, like in some lower molars of Canaanimys (MUSM 1892 and 1893; in Antoine et al., 2012, figure 2w-x). This is also observed on a tooth of Eoincamys ameghinoi (LACM 149442; in Frailey and Campbell, 2004, p. 114). In contrast, Eoincamys pascuali has more taeniodont lower molars. Contrary to the m1-2 of P. ucayaliensis and Canaanimys, the transverse cristids of the lower molars are more oblique in Eoincamys: the hypolophid is slightly oblique and tends to be aligned with an oblique ectolophid along with the posterior arm of the protoconid. The ectolophid is oblique in Eoincamys and Canaanimys, contrary to P. ucayaliensis. The p4 of P. ucayaliensis also resembles that of Eoincamys in showing a rounded crown outline and a reduced second cristid; yet, the posterior arm of the protoconid is more developed in Eoincamys than in P. ucayaliensis (small or absent). A reduced second cristid is also found in Draconomys Vucetich et al., 2015, Leucokephalos, Platypittamys, Deseadomys Wood and Patterson, 1959, and Incamys. As for Eoincamys , the p4 of P. ucayaliensis is relatively shorter with respect to the p4 of Draconomys, Leucokephalos, Platypittamys, Deseadomys, and Incamys (Wood, 1949; Wood and Patterson, 1959; Hoffstetter and Lavocat, 1970; Lavocat, 1976; Vucetich et al., 2010, 2015). Besides, the MUSM 2821 p4 (Figure 6.2) differs from Platypittamys and Deseadomys in the presence of a hypolophid. Although being taeniodont, the M2 of P. ucayaliensis is very distinct from that of Eoincamys or Canaanimys. Indeed, there is a prominent outgrowth of the protocone on M1-2 in C. maquiensis, which is less developed in P. ucayaliensis. On MUSM 2833 (and MUSM 2819) of P. ucayaliensis, the mesoloph is still linked to the mesolophular spur of the anterior arm of the hypocone, while the mesoloph tends to have no lingual connection with the anterior arm of the hypocone in E. ameghinoi (i.e., absence of the mesolophular spur) and the mesoloph is backwardly directed and connected with the posteroloph in E. pascuali. Finally, the MUSM 2833 M2 (Figure 6.4) of P. ucayaliensis shows a posteroloph with a distinct mesiodistal enlargement, which probably corresponds to a residual metaloph fused with the posteroloph. On the contrary, there is no metaloph in Eoincamys. In Canaanimys, there is a metaloph, but the latter is longer than in MUSM 2833 and lingually connected to the third transverse crest. However, Canaanimys can have a reduced metaloph but only on M3. A reduced metaloph on MUSM 2833 can be documented in Eoespina/Eosachacui and Incamys. Contrary to MUSM 2833 and Incamys, Eoespina/Eosachacui displays no taeniodont or pseudo-taeniodont molars. MUSM 2833 shows a slight oblique (diagonal) alignment of the protoloph with the mure and the anterior arm of the hypocone, as in Incamys (and Eoincamys). Contrary to MUSM 2833, upper molars of Incamys and Eoincamys can have a third transverse crest lingually free or connected to the posteroloph. MUSM 2833 (P. ucayaliensis) is lower-crowned than in Incamys, with equally thick enamel layer on the mesial and distal flanks of the hypoflexus (characteristic of most chinchilloids; Kramarz et al., 2013). Indeed, the leading edges are thicker than the trailing edges in Incamys and most chinchilloids (Kramarz et al., 2013; Vucetich et al., 2015). MUSM 2833 differs from all aforementioned taxa in having a shorter protoloph and third transverse crest. In sum, this material from CTA-29 is attributed to a new genus and species following this particular association of dental traits. Despite its non-taeniodont pattern, the MUSM 2819 M3 (Figure 6.3) is tentatively attributed to this new taxon.
Genus CACHIYACUY Antoine et al., 2012
Type species. Cachiyacuy contamanensis Antoine et al., 2012.
Species content. The type species and Cachiyacuy kummeli Antoine et al., 2012; Cachiyacuy cf. contamanensis, morph 1 (this work); Cachiyacuy cf. contamanensis, morph 2 (this work); Cachiyacuy cf. kummeli (this work).
Geographic and stratigraphic distribution. Contamana CTA-51, CTA-27, and CTA-29; Pozo Formation, lower member (late middle Eocene); Loreto Department, Peru.
Diagnosis sensu Antoine et al. (2012, p. 1320-1321). Rodents characterized by brachydont and bunolophodont teeth. Upper molars are pentalophodont with strong mesolophule and distinct and long metaloph. Differs from Eobranisamys, Branisamys, Eosallamys, Sallamys, Eoespina, and Draconomys in showing a long metaloph not backwardly directed and connected to the posteroloph, but transverse and without lingual connection. Accessory, thin and short enamel crests may connect the metaloph either to the posteroloph or/and to the mesolophule. Differs from Eobranisamys, Branisamys, and Canaanimys in the absence of taeniodont pattern on upper teeth due to the presence of a strong lingual protoloph. Differs from Eoincamys and Incamys in having brachydont instead of hypsodont teeth, pentalophodont instead of tetralophodont upper molars, thinner and transverse instead of strong and oblique crests, and in the absence of a taeniodont pattern on both upper and lower teeth (i.e., lingual protoloph and anterior arm of hypoconid lacking).
Cachiyacuy contamanensis Antoine et al., 2012
Figure 4.1-6, Figure 7, Figure 8, Appendix 3
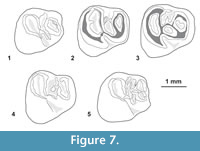 Referred material. In addition to the holotype (MUSM 1871, right M2; in Antoine et al., 2012, figure 2b) - MUSM 2661-2667, left dp4s; MUSM 1880 (in Antoine et al., 2012, figure 2k) and MUSM 2668-2673 (Figure 4.5), right dp4s; MUSM 2674-2675 (Figure 7.1), left p4s; MUSM 1879 (in Antoine et al., 2012, figure 2j, Figure 7.4) and 2676-2678 (Figure 4.4, Figure 7.2-3, 7.5), right p4s; MUSM 2679-2682, left m1s; MUSM 1878 (in Antoine et al., 2012, figure 2i, Figure 8.1) and 2683-2694 (Figure 8.3, 8.5, 8.8), right m1s; MUSM 2695-2703 (Figure 8.6), left m2s; MUSM 1877 (in Antoine et al., 2012, figure 2h), 1914-1915 (Figure 4.2-3, Figure 8.9-10) and 2704-2710 (Figure 8.2, 8.4), right m2s; MUSM 2711, left m3; MUSM 1876 (in Antoine et al., 2012, figure 2g) and 2712-2714, right m3s (Figure 4.1, Figure 8.7); MUSM 1875, dP3 or P3 (in Antoine et al., 2012, figure 2f); MUSM 1874 (in Antoine et al., 2012, figure 2e) and 2715-2717, left dP4s; MUSM 2718-2723, right dP4s; MUSM 2724, left P4; MUSM 1873 (in Antoine et al., 2012, figure 2d) and 2725-2729, right P4s; MUSM 1872 (in Antoine et al., 2012, figure 2c) and 2730-2738, left M1s; MUSM 2739-2746, right M1s; MUSM 2747-2750, left M2s; MUSM 2751-2757, right M2s; MUSM 2758-2761, left M3s (Figure 4.6); MUSM 1870, right M3 (in Antoine et al., 2012, figure 2a).
Referred material. In addition to the holotype (MUSM 1871, right M2; in Antoine et al., 2012, figure 2b) - MUSM 2661-2667, left dp4s; MUSM 1880 (in Antoine et al., 2012, figure 2k) and MUSM 2668-2673 (Figure 4.5), right dp4s; MUSM 2674-2675 (Figure 7.1), left p4s; MUSM 1879 (in Antoine et al., 2012, figure 2j, Figure 7.4) and 2676-2678 (Figure 4.4, Figure 7.2-3, 7.5), right p4s; MUSM 2679-2682, left m1s; MUSM 1878 (in Antoine et al., 2012, figure 2i, Figure 8.1) and 2683-2694 (Figure 8.3, 8.5, 8.8), right m1s; MUSM 2695-2703 (Figure 8.6), left m2s; MUSM 1877 (in Antoine et al., 2012, figure 2h), 1914-1915 (Figure 4.2-3, Figure 8.9-10) and 2704-2710 (Figure 8.2, 8.4), right m2s; MUSM 2711, left m3; MUSM 1876 (in Antoine et al., 2012, figure 2g) and 2712-2714, right m3s (Figure 4.1, Figure 8.7); MUSM 1875, dP3 or P3 (in Antoine et al., 2012, figure 2f); MUSM 1874 (in Antoine et al., 2012, figure 2e) and 2715-2717, left dP4s; MUSM 2718-2723, right dP4s; MUSM 2724, left P4; MUSM 1873 (in Antoine et al., 2012, figure 2d) and 2725-2729, right P4s; MUSM 1872 (in Antoine et al., 2012, figure 2c) and 2730-2738, left M1s; MUSM 2739-2746, right M1s; MUSM 2747-2750, left M2s; MUSM 2751-2757, right M2s; MUSM 2758-2761, left M3s (Figure 4.6); MUSM 1870, right M3 (in Antoine et al., 2012, figure 2a).
Type locality. Contamana CTA-27, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member late middle Eocene (Antoine et al., 2012, 2016).
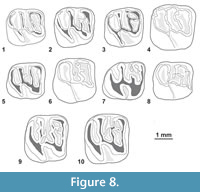 Diagnosis sensu Antoine et al. (2012, p. 1321). Cachiyacuy contamanensis (body mass estimated at 80-120 g) is ~30% larger than C. kummeli. Differs from C. kummeli in having upper molars with labial cusps and styles generally more marked and lower molars sometimes developing accessory enamel crests.
Diagnosis sensu Antoine et al. (2012, p. 1321). Cachiyacuy contamanensis (body mass estimated at 80-120 g) is ~30% larger than C. kummeli. Differs from C. kummeli in having upper molars with labial cusps and styles generally more marked and lower molars sometimes developing accessory enamel crests.
Description. The dp4 is about twice as long as wide, and it has a talonid wider than the trigonid (Antoine et al., 2012, figure 2k; Figure 4.5). The main cuspids are easily recognizable, but the protoconid and metaconid are slightly smaller than the hypoconid and entoconid. There is neither distinct anteroconid on the metalophulid I nor hypoconulid on the posterolophid. The mesial margin of the crown is rounded and formed by a curved metalophulid I. This cristid links the metaconid to the protoconid. The protoconid develops a short posterior arm, which extends lingually. The latter can reach the labial base of the metaconid (= metalophulid II) or not (MUSM 2663, 2665, 2670 [Figure 4.5], and 2673). Another cristid, stemming from the metaconid or its posterior arm, can participate to the formation of the second transverse cristid (MUSM 2663, 2665, and 2673). The mesostylid is well-developed and nearly as large as the protoconid. The mesostylid can be lingually isolated between the entoconid and metaconid or connected to the metaconid notably at a late wear stage (MUSM 2663, 2670, and 2669). Centrally, there is a mesolophid situated between the mesostylid and the distal ectolophid. The mesolophid is not strongly connected to these structures and can also be reduced (MUSM 2670; Figure 4.5). The majority of dp4s shows a long mesial ectolophid, mesially longitudinally oriented, and distally linguodistally directed. It connects the protoconid to the mesolophid and the distal ectolophid. However, on some dp4s (MUSM 1880, 2464, and 2671), it appears composed of different structures. On MUSM 2464 and 2671 for instance, there are two cristulids: one is linguodistally directed, stemming from the mesolophid-distal ectolophid junction, and another one is longitudinal, separate from the first and the protoconid by tiny notches. On MUSM 1880, the second cristulid (longitudinal one) is also found while the first (linguodistally directed) corresponds here to a large cuspid displaying anterior and posterior arms. That cuspid may be interpreted as a neomesoconid-like cuspid. Accessory enamel wrinkles occur between the second transverse cristid and the mesolophid on most dp4s. The distal ectolophid is very short and links the hypolophid to the mesolophid labially. The hypoconid and entoconid are labiolingually opposed and linked together by a long transverse hypolophid, which connects a short and thin anterior arm of the hypoconid. On two dp4s (MUSM 2670 [Figure 4.5] and 2671), the anterior arm of the hypoconid is low in its middle part and thus appears composed of two cristulids: a mesial one from the ectolophid and a distal one from the hypoconid. The distal margin of the crown is circular and formed by a strong posterolophid, which can be connected to the posterior arm of the entoconid. The posterolophid and hypolophid isolate a broad metaflexid/metafossettid.
The p4s differs substantially from dp4 in having a talonid significantly wider than the trigonid, and in being as long as large (in Antoine et al., 2012, figure 2j; Figure 4.4). The main cuspids are equally sized and salient. Mesially, the metalophulid I is well-elevated and labiolingually straight. The second transverse cristid is complete and continuous on the most specimens (but most are worn). It extends from the protoconid to the mesostylid, which is faintly distinct, being merged with the posterior arm of the metaconid. Two p4s (MUSM 1879 [in Antoine et al., 2012, figure 2j; Figure 7.4] and 2678 [Figure 4.4, Figure 7.5]) show a complex pattern of the second transverse cristid. On these p4s, this cristid is divided into three parts: two labial and a lingual one. The more mesial and labial part, stemming from the protoconid, corresponds to the posterior arm of the protoconid. The second labial part is more reduced, notably on MUSM 1879, and may be a residual branch of the mesolophid (labial part) because it is connected to a distinct but low mesoconid (on MUSM 2678), which occupies a more central position than on dp4. The homology of the lingual part stemming from an isolated mesostylid is more obscure (probably a neomesolophid). From its cuspid connections, its position and shape, the continuous second transverse cristid of other p4s (e.g., MUSM 2674, Figure 7.1) is probably a fusion of the first labial part with the lingual one found in MUSM 1879 (Figure 7.4) and 2678 (Figure 4.4, Figure 7.5). MUSM 2678 is very particular because it shows a very low cristulid (a neostructure?) between the hypolophid and the second cristid. It appears divided with a lingual part lingually connected to a small cuspid (neostructure too?) and a labial part that reaches the base of the ectolophid. Two other p4s (MUSM 2676 [Figure 7.2] and 2677 [Figure 7.3]) display fragments of that cristulid. On all p4s, the hypolophid is complete and joins the short and thin anterior arm of the hypoconid. The posterolophid is strongly developed and connects a strong and short posterior arm of the entoconid. There is no trace of hypoconulid.
The lower molars (m1-3) share the same dental structure (in Antoine et al., 2012, figure 2g-i; Figure 4.1-3). The m1s are clearly smaller than m2s, and m3s are of the same size or smaller than m2s. The m3s are characteristic in having a small entoconid, which is more labial in position to the metaconid and in showing a distal expansion of the posterolophid (Figure 4.1). The occlusal pattern is similar from m1 to m3 and strongly reminiscent of that of p4. Molars differ from p4s in being much more quadrate, and in showing a strong development of the posterior arm of the metaconid (metastylar fold), which is directed to a small but well distinct mesostylid. The posterior arm of the metaconid and mesostylid are connected (twinned) or separated by a tiny and shallow notch. In contrast, a deep and narrow furrow usually separates the mesostylid from the entoconid. Mesially, at the base of the crown, there is no trace of anterocingulid. The metaconid and protoconid are labiolingually opposed and linked by a straight and well-trenchant metalophulid I. The second transverse cristid is somewhat difficult to describe inasmuch as it appears highly variable (Figure 8). On some teeth, this cristid is formed by a posterior arm of the protoconid, which is connected to the mesostylid or the posterior arm of the metaconid (= metalophulid II; Figure 8.5). However, the posterior arm of the protoconid can also be short in some cases (Figure 8.2, 8.7-8). On some other molars (Figure 8.1, 8.3, 8.9-10), like on p4s, it may appear as a composite of two cristids that merge together: one being a short posterior arm of the protoconid (lingually directed), and the other being a short cristid stemming from the mesostylid (labially directed, = neomesolophid). These two cristids can be often disjoined (Figure 8.7-8). On some molars, there is a neomesolophid and no posterior arm of the protoconid or a very short one (e.g., MUSM 2708, Figure 8.4). In many cases, a longitudinal accessory enamel cristulid may occur, joining the second transverse cristid to the metalophulid I (Figure 8.3, 8.9-10). Like on p4s, some lower molars (MUSM 1914, 1915, 2689, 2692, 2701, 2708, and 2714; Figure 4.2-3, Figure 8.5-10) display an accessory “third” transverse cristulid and/or a small lingual cuspid, (neostructures?). In MUSM 1914 (Figure 4.2, Figure 8.10), that “third” cristulid is long but interrupted. Its lingual part is connected to the small lingual cuspid and remains labially free. Its shorter labial part is labially linked to the hypolophid. Also on this tooth occurs a short longitudinal enamel wrinkle that bridges the second and third transverse cristids. On two lower molars (MUSM 1915 [Figure 4.3, Figure 8.9] and 2689 [Figure 8.8]), there is a fragment of cristulid that could be the labial part of a mesolophid. On MUSM 1915, there are two labial cristulids situated between the second transverse cristid and the hypolophid, one stemming from the ectolophid (but without mesoconid) and the other from the hypolophid. The first might be a fragment of the mesolophid. On MUSM 2689, there is a short cristulid distolabially directed, linked to the lingual part of the second transverse cristid and labially free. It might be a fragment of the mesolophid or the “third” transverse cristulid. On all molars, the hypoconid and entoconid are labiolingually opposed and linked by a long and straight hypolophid, which connects labially a short, but strong anterior arm of the hypoconid (absence of taeniodont pattern). Many lower molars have a minute mesoconid, located directly mesial to the labial extremity of the hypolophid. On these lower molars, there is no trace of mesolophid stemming from the mesoconid. The entoconid can display anterior and posterior arms. When it is present, the anterior arm of the entoconid is thin and faintly visible, contrary to the posterior arm (usually stronger, except on MUSM 2708 [Figure 8.4]). The hypoconid is slightly mesiodistally compressed. A strong and curved posterolophid connects the hypoconulid but not the posterior arm of the entoconid, which is separate from the posterolophid by a narrow and shallow notch. However, in some teeth showing an advanced degree of wear, the posterolophid can be connected to the posterior arm of the entoconid. The anteroflexid and metaflexid are wide.
As for the lower dentition, all the dental loci are documented for the upper dentition.
Only one specimen of DP3 is available (MUSM 1875; in Antoine et al., 2012, figure 2f). It is a single-rooted, peg-like tooth, with a circular outline. On the occlusal surface, there are two crests and two cusps. The higher crest connects both cusps.
We identify as dP4s trapezoidal teeth that show their labial crown margin longer than their lingual one (in Antoine et al., 2012, figure 2e). These teeth are brachydont and exhibit a pentalophodont pattern with transverse crests that are moderately elevated. The main cusps (protocone, hypocone, paracone, and metacone) remain well-defined. The anteroloph is oblique, directed mesiolabially. Lingually, it connects the mesial border of the protocone, and labially, a distinct parastyle. The parastyle is slightly mesiolingual to the paracone and separate from it by a narrow but deep notch (paraflexus labially open). The labial protoloph is long, oblique, and parallel to the anteroloph, and it connects a short lingual protoloph (= posterior arm of the protocone). As such, the internal sinus (hypoflexus) remains separate from the paraflexus (absence of taeniodont pattern). The protocone is slightly pinched labiolingually and somewhat oblique in position. On some dP4s (MUSM 2720 and 2721), a secondary cusp is present on the distal wall of the paracone. On MUSM 2717, that accessory cusp is neither on the wall of the paracone nor on that of the mesostyle but it is located on the opening of the mesoflexus. On all dP4s, the hypocone is slightly labial to the protocone. Both cusps are equally sized. The hypocone displays a strong, long, and oblique anterior arm, which extends mesially (via a short but complete mure) to join the confluence between the lingual protoloph and the lingual extremity of the labial protoloph. This entire complex delimits the hypoflexus, which appears narrow and deep. The third transverse crest usually connects the mesial extremity of the anterior arm of the hypocone to a strong mesostyle, but this crest often appears as a composite crest including a mesolophule and a mesoloph. The mesolophule is always long, while the mesoloph (when present) is short and either connected or not to the mesolophule. Otherwise, in most specimens, the mesolophule displays in its lingual part a small but well-defined enamel swelling (accessory neocusp). On MUSM 1874 (in Antoine et al., 2012, figure 2e), a tiny accessory cusp is present and twinned to the mesostyle. This accessory cusp displays a short crestule that runs lingually between the protoloph and the central transverse crest, and ends without connection. On all dP4s, the mesostyle is mesiodistally enlarged and often twinned with the metacone. The metacone is distally positioned with respect to the hypocone. These two distal cusps are linked by a strong and complete posteroloph. The fourth transverse crest is a metaloph, which shows a variable development. It is either long, stemming from the metacone (MUSM 1874; in Antoine et al., 2012, figure 2e), or disconnected to that cusp and usually very reduced or even absent (MUSM 2716 and 2718). Generally, the lingual extremity of the metaloph does not reach the anterior arm of the hypocone. It remains free lingually or linked to the mesolophule and/or the posteroloph via thin, short, and longitudinal accessory enamel crestules. On MUSM 2719, there are three structures (crestules and cusps) on the most distal flexus (confluence of the metaflexus and posteroflexus), which may correspond at least in part to the metaloph. One of these structures is labially and lingually connected to the posteroloph, which forms a small and rounded fossette.
In occlusal view, the crown outline of the P4s is oval- to heart-shaped (mesiodistal compression), and as such, these teeth are clearly distinguishable from the dP4s (in Antoine et al., 2012, figure 2d). The anteroloph shows the same connexions (labially with the parastyle and lingually with the protocone) as it does in the dP4s. It is continuous but can be divided in two parts. Its labial part is slightly less elevated than the other transverse crests. Its lingual one is more elevated and corresponds to a long anterior arm of the protocone. The protocone is mesially canted and labiolingually pinched. It develops a long and lingual posterior outgrowth, which is backwardly directed. This outgrowth connects a minute and crestiform hypocone, and entirely closes the hypoflexus lingually as a result. As on dP4s, P4s exhibit the same arrangement of the anterior arm of the hypocone, which connects the protoloph via a complete longitudinal mure. The metacone is crestiform and mesiodistally elongated. Labially, there is a minute and isolated mesostyle, which is separate from the paracone to the metacone. The third transverse crest can be complete: it connects the mesostyle to the mesial extremity of the anterior arm of the hypocone. On MUSM 1873 (in Antoine et al., 2012, figure 2d), the third transverse crest shows a break in its middle part and thus, corresponds to the combination of a mesoloph and a mesolophule. On two specimens (MUSM 2724 and 2725), the third transverse crest is reduced: it lacks its lingual connection with the anterior arm of the hypocone (= mesoloph; MUSM 2724), or its labial one with the mesostyle (= mesolophule; MUSM 2725). On MUSM 2726, the third transverse crest is entirely lacking. When it is present (MUSM 1873 and 2727), a short metaloph, stemming from the metacone, runs lingually. On MUSM 2727, the metaloph is lingually free, while it is connected to the posteroloph via a very short and longitudinal accessory enamel crestule on MUSM 1873 (in Antoine et al., 2012, figure 2d).
The upper molar proportions are similar to those observed for the lower molars (in Antoine et al., 2012, figure 2a-c; Figure 4.6). The upper molars (M1-3) share the same dental structure. M1s are significantly smaller than M2s, and M3s appear roughly equal in size to M2s. However, M3s differ substantially from M2s in having a hypocone strongly displaced labially. As for the dP4s, the upper molars are brachydont, fully pentalophodont, and non-taeniodont. On pristine teeth, the five transverse crests (anteroloph, protoloph, mesolophule/mesoloph, metaloph, and posteroloph) are thin and moderately elevated, and the main cusps and styles are well-defined. There is no conule. Lingually, the protocone is massive and slightly labiolingually compressed. It displays a short posterior arm (= lingual protoloph), which connects the long and transverse labial protoloph (absence of taeniodont pattern). The lingual branch of the protoloph is often grooved in its middle part and thus, appears composed of two crestules: a mesial one from the hypocone, and a distal one from the protoloph-mure junction. On M1s, the hypocone is distal and as large as the protocone. On M2s, the hypocone is slightly smaller and also slightly displaced labially. Except on some M3s, the upper molars bear a strong and longitudinal mure, which connects the anterior arm of the hypocone to the protoloph lingually. On the three molar loci, the third transverse crest, strongly developed, links the anterior arm of the hypocone to a strong and well-defined mesostyle, situated in the labialmost margin of the crown. As for dP4s and P4s, this crest seems to be formed by the union of a mesoloph and a mesolophule. The mesoloph is often long, while the mesolophule can be either long, reduced (spur), or absent. The mesostyle is usually remote from the metacone except on five molars (among less worn upper molars; MUSM 2739, 2740, 2744, 2750, and 2759) where it is close but still separated by a narrow and shallow notch. The metaloph is usually present and well-developed, but it can also be reduced, interrupted, or absent on M1-3. It is usually strong on M1s and M2s and reduced on M3s. On M1-2s, the metaloph runs lingually (slightly oblique on M2s) but remains without lingual connection. Accessory, thin and short enamel crestules may lingually connect the metaloph either to the posteroloph or/and to the third transverse crest. On M3s, the metaloph is associated with other crestules stemming from accessory cusps on the posteroloph (Figure 4.6). The hypoflexus is narrow, and its lingual aperture is somewhat mesiodistally constricted. The paraflexus and mesoflexus remain open labially. When the mesostyle and metacone are very close, the metaflexus appears almost labially closed. When the metaloph is lingually connected to the posteroloph, the posteroflexus is divided into two parts: a labial one, which is delimited by the metaloph and posteroloph, and a lingual one, which is delimited by the metaloph, the anterior arm of the hypoconid and lingual end of the posteroloph.
Cachiyacuy kummeli Antoine et al., 2012
Figure 4.9-13, Appendix 3
Referred material. In addition to the holotype (MUSM 1882, left M1; in Antoine et al., 2012, figure 2m) - MUSM 1888 (in Antoine et al., 2012, figure 2s) and 2762-2763 (Figure 4.9), left dp4s; MUSM 2764-2765, right dp4s; MUSM 2766, left p4 (Figure 4.10); MUSM 2767-2768, left m1s; MUSM 1887 (in Antoine et al., 2012, figure 2r) and 2769-2771, right m1s; MUSM 2772, left m2; MUSM 1886 (in Antoine et al., 2012, figure 2q) and 2773-2775, right m2s; MUSM 2776, right m2?; MUSM 2777-2779, left m3s; MUSM 1885 (in Antoine et al., 2012, figure 2p) and 2780-2781 (Figure 4.11), right m3s; MUSM 2783, left P4; MUSM 1881, right P4 (in Antoine et al., 2012, figure 2l); MUSM 2784, right M1 or M2; MUSM 2785, left M1 (Figure 4.12); MUSM 1883 (in Antoine et al., 2012, figure 2n) and 2786 (Figure 4.13), left M2s; MUSM 1884, left M3 (in Antoine et al., 2012, figure 2o).
Type locality. Contamana CTA-27, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2012, 2016).
Diagnosis sensu Antoine et al. (2012, p. 1321).Cachiyacuy kummeli (body mass estimated at 30-40 g) is ~30% smaller than C. contamanensis. The molars have slightly thinner transverse crests and the cusp(id)s are more salient than in C. contamanensis.
Description. The occlusal morphology of the upper and lower teeth referred to Cachiyacuy kummeli is basically similar to that of the teeth of C.contamanensis. C. kummeli differs substantially from C. contamanensis in being ~30% smaller, and in having molars with transverse crests slightly thinner and cusp(id)s more salient. The dp4s of C. kummeli also differ from those referred to C. contamanensis, notably in the distal region of the talonid, which shows a posterolophid less curved (Figure 4.9). The unique p4 of C. kummeli (MUSM 2766; Figure 4.10) has neither mesoconid, nor mesolophid nor neocristulid(s) between the second transverse cristid and the hypolophid, contrary to some specimens of C. contamanensis (Figure 7). On the lower molars, especially on m1s, the mesoconid is more frequently absent and the mesostylid is often less indistinct, being entirely subsumed in the strong posterior arm of the metaconid (Figure 4.11). The second transverse cristid, which could appear as a long and complete metalophulid II, also seems to be a composite of two short cristids (posterior arm of the protoconid and neomesolophid) that merge together medially. The accessory enamel cristids are rarer and less long. Like in C.contamanensis, the metaloph on upper molars can be absent (MUSM 2785; Figure 4.12). On M2s of C. kummeli (MUSM 1883 and 2786; Figure 4.13), the metaloph is less developed than that of M2s of C.contamanensis. On MUSM 1883, the metaloph corresponds to a tiny low crestule on the mesial slope of the posteroloph without any connection with the third transverse crest, the anterior arm of the hypocone or the posteroloph. MUSM 2786 (Figure 4.13) shows an unusual crest, which is mesiolingually directed, and located between the third transverse crest and the posteroloph. That crest is composed of two merged crestules, one stemming from the posteroloph to the other from the third transverse crest. These crestules may be remanent connections of the metaloph to the third transverse crest and posteroloph.
Comparisons.Cachiyacuy differs from Eoincamys from Santa Rosa (Peru, ?late Eocene/early Oligocene; Frailey and Campbell, 2004) and Incamys from Salla (Bolivia, late Oligocene; Hoffstetter and Lavocat, 1970; Lavocat, 1976) in having brachydont instead of hypsodont teeth, pentalophodont instead of tetralophodont upper molars, and in developing thinner and transverse instead of strong and oblique crests, and in the absence of a taeniodont pattern on both upper and lower teeth (i.e., lingual protoloph and anterior arm of hypoconid lacking). Contrary to Eobranisamys, Branisamys, and Canaanimys, Cachiyacuy does not exhibit a taeniodont pattern of the upper molars due to the presence of a strong lingual protoloph (Hoffstetter and Lavocat, 1970; Lavocat, 1976; Frailey and Campbell, 2004). Cachiyacuy differs from Eobranisamys, Branisamys, Eosallamys, Sallamys, Eoespina, Eosachacui, Vallehermosomys Vucetich et al., 2010 and Draconomys in showing long metalophs neither backwardly directed nor connected to the posteroloph, but transverse (lingually directed) and without lingual connection instead. Accessory, thin and short enamel crests may connect the metaloph to the posteroloph and/or to the central transverse crest. However, on some upper molars of Eosallamys (LACM 143422, 143363, 143376, 143380; in Frailey and Campbell, 2004, p. 120-122), the metaloph can be connected to the central transverse crest via a secondary crestule. However, in Cachiyacuy, the crests and cristids are slenderer, and they show a thinner enamel thickness than in Eosallamys. In Cachiyacuy (especially in C. contamanensis), lower molars have a complete second transverse cristid (even if it is composed of two parts) and accessory cristulids within the anterofossettid. In contrast, in Draconomys there is no accessory cristulid within the anterofossettid, and the second transverse cristid is always interrupted before to reach the mesostylid (i.e., absence of neomesolophid in this taxon), being only composed of a long posterior arm of the protoconid. Besides, Cachiyacuy differs from Draconomys in having transverse crests on its upper molars, which are labiolingually shorter. Finally, Cachiyacuy differs from Sallamys in having a second composite transverse cristid more complete on lower molars.
Cachiyacuy cf. contamanensis, morph 1
Figure 5.5-7, Appendix 3
2016 Cachiyacuy cf. contamanensis Antoine et al., Supplementary data, p. 5.
2017 Cachiyacuy cf. contamanensis Antoine et al., Supplementary data, p. 9.
Referred material. MUSM 2651, fragmentary right dp4 (Figure 5.5); MUSM 2652, left m1 (Figure 5.6); MUSM 2653, left m3 (Figure 5.7).
Locality. Contamana CTA-51, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Description. The dp4 (MUSM 2651; Figure 5.5) is worn and broken labiomesially. Due to the fragmentary state of the tooth, only the lingual cuspids are preserved: a part of the metaconid, the mesostylid and the entoconid. The posterior arm of the metaconid is linked to a crestiform mesostylid. The second transverse cristid is divided into two independent parts: the lingual one is connected to the metaconid, and the labial one was probably linked to the protoconid (posterior arm of the protoconid). Its lingual part is oriented labiodistally. In the mesiocentral part of the tooth, the mesolophid appears discontinuous and limited to two parts: an accessory cuspid and a tiny spur of ectolophid. The accessory cuspid, situated between the second transverse cristid and the hypolophid, connects to the lingual part of the second transverse cristid. A tiny spur of the ectolophid runs labially and ends directly distolabially to the accessory cuspid. The entoconid is the highest and largest cuspid. The hypolophid is straight and transverse. The anterior arm of the hypoconid may be present. The labial part of the posterolophid is curved and does not reach the distal aspect of the entoconid, ending its course far away from this main lingual cuspid.
The m1 (MUSM 2652; Figure 5.6) is nearly pristine. This tooth is subrectangular in occlusal outline, with a talonid slightly larger than the trigonid. It exhibits a tetralophodont pattern, characterized by strong and elevated cuspids and cristids. The metaconid, massive, crestiform and mesiolabially canted, is prominent over other cuspids. The protoconid, hypoconid, and entoconid are low, virtually equally sized, but the entoconid is the most cuspate cuspid, the others being more crestiform. The protoconid and hypoconid are mesiodistally opposed, but the hypoconid displays a long, narrow, and mesiolabially oriented outgrowth that widens the talonid. There is no anterocingulid. A strong and straight metalophulid I forms the mesial margin of the crown. This cristid strongly connects the metaconid to the protoconid, this latter being slightly more distal in position to the metaconid. The metaconid displays a long and strong posterior arm, which runs distally to reach a faintly visible mesostylid. This stylid is in fact entirely incorporated within the posterior arm of the metaconid. The second transverse cristid is the lowest cristid of the crown surface. It is straight and transversely continuous, and could be identified as a complete metalophulid II (but see description of the m3, hereafter). With this longitudinal cristid configuration, as well as the connection between the posterior arm of the metaconid and the mesostylid, the anteroflexid is lingually closed and forms an anterofossettid. A longitudinal and short enamel spur (accessory cristulid) occurs in the middle part of the second transverse cristid. It runs toward the metalophulid I, but does not connect to it. The ectolophid is oblique and links the distal aspect of the protoconid to the hypolophid. Distolingually, the entoconid displays short anterior and posterior arms. The anterior arm runs mesially but does not reach the mesostylid, thereby generating a well-marked and deep furrow that makes the mesoflexid open lingually. The entoconid and the hypoconid are labiolingually opposed, and linked by a strong and curved posterolophid that closes the metaflexid lingually (= metafossettid). Distolabially, the anterior arm of the hypoconid is complete but low in its middle part (grooved), and thus it appears composed of two cristulids: a mesial one stemming from the ectolophid and a distal one from the hypoconid). Within the metafossettid, there is a small accessory cuspid, located on the distal wall of the hypolophid. All flexids/fossettids are wide and deep.
The m3 (MUSM 2653; Figure 5.7), despite having the distal part of its talonid labiolingually pinched, basically exhibits a similar dental pattern to that of m1. The mesostylid is better defined and separate from the posterior arm of the metaconid by a thin and shallow notch. On this tooth, the second transverse cristid appears continuous, but there is a disruption on its middle, which indicates that this transverse cristid is a combination of two cristids. Its lingual part seems to be a neomesolophid that runs labially from the mesostylid. The labial part of the second transverse cristid corresponds to the posterior arm of the protoconid. As on m1, there is an accessory longitudinal enamel cristulid, which is situated at the labial extremity of the short neomesolophid that runs toward the metalophulid I without connecting it. The labial part of the posterior arm of the protoconid also bears a short accessory cristulid, forwardly directed. Lingually, an accessory cuspid is also present on the mesoflexid, close to the mesostylid. However, there is no secondary cuspid on the metafossettid as it does on m1. Otherwise, this tooth also differs from m1 in showing a weaker development of the anterior arm of the hypoconid, which appears composed of two cristulids, as on m1, but the latters are disconnected, thereby involving an incipient taeniodonty.
Comparisons. The three teeth have trenchant cristids but the latter are clearly thinner compared with Deseadan or Miocene caviomorph species. Basically, the occlusal pattern of the lower molars from CTA-51 is simply constructed and similar to that found in most of pre-Deseadan caviomorphs (i.e., brachydonty, tetralophodonty, and non-taeniodonty without oblique cristids). The specimens recall Cachiyacuy contamanensis by their size, although they are slightly smaller, and by some morphological characters. On the MUSM 2651 dp4 (Figure 5.5), the second transverse cristid is discontinuous with its lingual part distally directed, and the mesolophid is limited to a very short cuspid-like cristid. On the dp4 of Cachiyacuy kummeli, Eobranisamys, Eoespina, Eosachacui, Eosallamys simpsoni, Sallamys pascuali, Sallamys cf. pascuali, and Branisamys (Hoffstetter and Lavocat, 1970; Lavocat, 1976; Frailey and Campbell, 2004; Shockey et al., 2009), the mesolophid is well-developed and labiolingually complete. On LACM 143451 (Eosallamys paulacoutoi ; in Frailey and Campbell, 2004, p. 122), the mesolophid is reduced but appears as a well-defined cristulid still connected to the mesostylid. In Sallamys quispea Shockey et al., 2009, the mesolophid is reduced and includes two cristulids: one stemming from the mesostylid (neomesolophid) and the other from the ectolophid (true mesolophid). In Canaanimys maquiensis, the mesolophid on dp4 is reduced to a short cuspid-like cristid, like in MUSM 2651. However, on MUSM 2651, there is a lingual part of the second transverse cristid, while this structure is absent on the dp4 of Canaanimys maquiensis (MUSM 1888; in Antoine et al., 2012, figure 2k) . On the dp4 of C. contamanensis, the cristid arrangement in the trigonid is highly variable. In this species, the second transverse cristid and mesolophid of dp4s can be transversely complete but with a tendency to be divided and reduced (MUSM 2665, 2670, 2671, 2672, and 2673). One dp4 of C. contamanensis (MUSM 2670; Figure 4.5) has a similar configuration to that found in MUSM 2651 (i.e., discontinuous second transverse cristid with its lingual part distally directed, and a mesolophid reduced to a cuspid-like cristid). In addition, in having a posterior arm of the metaconid high and strongly connected to the mesostylid, the MUSM 2651 dp4 is somewhat peculiar among the Paleogene taxa known, since this character is otherwise found only in C achiyacuy, Sallamys quispea and Sallamys cf. pascuali (Shockey et al., 2009). In C achiyacuy, this character is rather variable and only found on some dp4s (MUSM 2663, 2670 [Figure 4.5], 2669, and 2763-2765). Finally, on MUSM 2652 and 2653, the addition of accessory cristulids connected to the second transverse cristid can also be observed in some lower molars of C. contamanensis,Eobranisamys and Eosachacui. Contrary to the MUSM 2652 and 2653 molars (Figure 5.6-7), or the molars of C. contamanensis, in Eobranisamys and Eosachacui, these cristulids are shorter and absent on m3s. Lower molars from CTA-51 differ from those attributed to C. maquiensis (CTA-27) in having a high posterior arm of the metaconid and a continuous metalophulid I. In sum, due to the strong dental affinities, an attribution of these lower teeth from CTA-51 to Cachiyacuy contamanensis could by reasonable. However, given the small difference of size noticed, we attribute MUSM 2651, 2652, and 2653 to Cachiyacuy cf. contamanensis, morph 1.
Cachiyacuy cf. contamanensis, morph 2
Figure 6.7-12, Appendix 3
2016 Cachiyacuy cf. contamanensis Antoine et al., p. 13 and Supplementary data, p. 9.
2017 Cachiyacuy cf. contamanensis Antoine et al., Supplementary data, p. 9.
Referred material. MUSM 2823, fragment of a right dp4 or p4; MUSM 2825, left dp4 (Figure 6.7); MUSM 2826, fragmentary right m1; MUSM 2827, right m1 (Figure 6.8); MUSM 2828, right dP4 (Figure 6.9); MUSM 2829-2830, fragmentary right upper molars; MUSM 2831, left M1 (Figure 6.10); MUSM 2832, left M2 (Figure 6.12); MUSM 2563, right M2 (Figure 6.11).
Locality. Contamana CTA-29, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Description. The MUSM 2825 (Figure 6.7) is a complete but moderately worn dp4. It is pentalophodont and non-taeniodont. The curved metalophulid I and second transverse cristid are both lingually and labially linked to the metaconid and protoconid, respectively, these two cuspids being linguolabially opposed. The second transverse cristid corresponds to a long posterior arm of the protoconid, which connects to the metaconid (= metalophulid II). A well-developed and transverse mesolophid is lingually connected to a well-defined mesostylid and labially to the mesial ectolophid. The mesial ectolophid is slightly linguodistally extended, and it is not linked to the protoconid. There is a short and low distal ectolophid. The hypolophid is well-developed and slightly oblique due to the more mesial position of the entoconid with respect to the hypoconid. The lingual end of the posterolophid reaches the posterior arm of the entoconid. Only the anteroflexid and metaflexid are entirely closed, thereby forming an anterofossettid and a metafossettid, respectively.
Both specimens (MUSM 2826 and 2827; Figure 6.8) are m1s and are characterized by a trigonid slightly narrower than the talonid. They have a typical morphology of most pre-Deseadan caviomorphs, in being tetralophodont, non-taeniodont, and with a long second transverse cristid (unique or composite). On these two specimens, the second transverse cristid is complete and continuous, seemingly being only composed of a long posterior arm of the protoconid (= metalophulid II). MUSM 2826 differs from MUSM 2827 (Figure 6.8) in having a tiny cristulid extending between the metalophulid I and metalophulid II, and in lacking the connection between the metaconid and the mesostylid.
The dP4 (MUSM 2828; Figure 6.9) is markedly eroded, notably in its lingual part, and its dental structure has been virtually erased. This tooth is trapezoidal with a lingual edge shorter than the labial one. The paracone is mesiolabially canted and strongly connected to the anteroloph. The protoloph is distolingually connected to the paracone, and is lingually complete, being connected distolabially to the protocone (absence of taeniodonty). Lingually, the hypocone displays a markedly oblique anterior arm that connects the protoloph mediolabially via a strong and oblique mure. Distolabially, the metacone is rather indistinct, being subsumed within the posteroloph. On the labial margin of the tooth, the mesostyle is faintly visible, but it displays a thin posterior arm, which reaches the anterior arm of the metacone. The third transverse crest and metaloph are short but well-defined, and they are lingually linked, thereby forming a V-shaped structure. The lingual base of this structure is connected to the mesial extremity of the anterior arm of the hypocone via a small crestule, which is interpreted here as a very short mesolophule. The labial branch of the third crest is in contrast interpreted as a mesoloph. Another tiny accessory crestule occurs distally, and links the lingual part of the posteroloph to the distal aspect of the anterior arm of the hypocone. All the flexi are entirely closed and they form fossettes except for the mesoflexus.
The upper molars (Figure 6.10-12) are brachydont, primarily tetralophodont in appearance (anteroloph, protoloph, third transverse crest, and posteroloph; but see below concerning the presence of a metaloph), non-taeniodont, and somewhat transverse in occlusal outline (i.e., wider than long). The paracone is the highest and largest cusp of the crowns. The metacone is slightly more lingual in position and rather indistinct, being entirely subsumed within the posteroloph. In contrast, there is a strong and well-defined mesostyle between the paracone and the metacone. While a deep furrow separates the paracone from the mesostyle (mesoflexus open labially), the metacone is connected to the mesostyle via a well-marked anterior arm. However, tiny and shallow labial notches, absent on MUSM 2563 (Figure 6.11), tend to separate the metacone from both the mesostyle and posteroloph on MUSM 2831 (Figure 6.10). Lingually, the protocone is massive. It develops a strong and long posterior outgrowth, which runs distally and nearly closes the hypoflexus lingually (mesiodistal constriction of the hypoflexus). Contrary to MUSM 2832 (Figure 6.12), the lingual protoloph is composed of two crestules on MUSM 2831 (Figure 6.10): one stemming from the protocone and the other from the protoloph. The hypocone is smaller and slightly more labial in position. It displays a strong and oblique anterior arm, which continues its course mesially via a well-defined longitudinal or slightly oblique mure that links the protoloph in its lingual part. A trenchant third transverse crest extends linguolabially between the mesial extremity of the anterior arm of the hypocone and the mesostyle. It is parallel and close to the protoloph. On MUSM 2832 (Figure 6.12), the third transverse crest is slightly curved. It is strongly linked to the mesostyle. In contrast, it shows a lingual thinning on MUSM 2831, and is almost disconnected to the anterior arm of the hypocone on MUSM 2563 and 2832 (i.e., = mesoloph). On MUSM 2832, a secondary minute crestule, backwardly directed, is present on the distolingual wall of the mesoloph. The two least worn upper molars (MUSM 2831 and 2832) are tetra to pentalophodont because of a variable development of the metaloph-like crest, whereas on the more worn upper molar (MUSM 2563; Figure 6.11), there is no metaloph but a tiny spur on the posteroloph, which is probably a relic of that loph. When present, the metaloph is lingually free. On all teeth, the labial end of the anteroloph reaches the base of the paracone and generates, with the protoloph, an oval-shaped, narrow and labiolingually developed anterofossette. On MUSM 2831 (Figure 6.10), there are two secondary longitudinal crestules within the anterofossette: one situated on the distal wall of the anteroloph and one situated on the mesial one of the protoloph.
Comparisons. The size and morphology of these teeth correspond to those of Cachiyacuy contamanensis. The dP4 is morphologically similar with the dP4s referred to C. contamanensis, notably in having the metaloph that reaches the third transverse crest lingually. Moreover, as for MUSM 2828 (Figure 6.9), many dP4s of C. contamanensis have a secondary longitudinal crestule stemming from the posteroloph. Like in Cachiyacuy, Canaanimys, and Eosallamys, the upper molars described here are subrectangular rather than subquadrate as in Eoespina/Eosachacui and ?Vallehermosomys merlinae. On these CTA-29 specimens, the development of the metaloph is highly variable (long, small, or almost absent), a variation which is found on the M1-2s of Cachiyacuy and Eosallamys. Like in Cachiyacuy, these upper molars from CTA-29 differ from those of Eosallamys in having more slender crests with a thinner enamel thickness. They also display a long posterior outgrowth of the protocone, which remains shorter than that observed in Canaanimys. The non-taeniodont pattern of these upper molars excludes affinities with Vallehermosomys mazzoni, Canaanimys, Eobranisamys, Branisamys, Eoincamys and Incamys (Hoffstetter and Lavocat, 1970; Lavocat, 1976; Frailey and Campbell, 2004; Vucetich et al., 2010). Their brachydonty also contrasts with the higher crowned teeth characterizing Branisamys and Incamys. The pentalophodonty of the MUSM 2825 dp4 (Figure 6.7) excludes Eobranisamys and Branisamys. Compared with the dp4 of Canaanimys maquiensis (MUSM 1895; in Antoine et al., 2012, figure 2z), MUSM 2825 from CTA-29 does not have a lingual connection between the metaconid and mesostylid. This character is variable in dp4s of Cachiyacuy (i.e., presence or absence of that connection). In addition, on MUSM 2825, the protoconid is clearly separate from the mesial ectolophid like in Eosallamys simpsoni (as well as in Eobranisamys and Branisamys). However, this character is not found in Cachiyacuy and Eosallamys paulacoutoi , while it is variable in Eoespina/Eosachacui. Given the very similar dimensions and morphological features of this dental material from CTA-29 with C. contamanensis, we provisionally assigned the latter to Cachiyacuy cf. contamanensis, morph 2. Because of the apparent larger size of this dental material form CTA-29 with respect to MUSM 2651-2653 from CTA-51 (attributed to Cachiyacuy cf. contamanensis, morph 1, see above) we refer these isolated teeth to as two different taxa.
Cachiyacuy cf. kummeli
Figure 5.11, Appendix 3
2016 Canaanimys cf. maquiensis Antoine et al., Supplementary data, p. 5.
2017 Canaanimys cf. maquiensis Antoine et al., Supplementary data, p. 9.
Referred material. MUSM 2654, fragmentary left M1 (Figure 5.11).
Locality. Contamana CTA-51, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Description. The MUSM 2654 M1 (Figure 5.11) has an occlusal surface and a crown outline heavily worn, and lacks its posterolingual part. Although broken, the hypocone does not appear to have occupied a labial position, which indicates that this tooth is likely a M1. The posterior outgrowth of the protocone is short, which involves a very small mesiodistal constriction of the hypoflexus. The protoloph is transversely complete, its long labial part is slightly oblique (distolingually oriented), and its lingual part (posterior arm of the protocone) is short but strong (i.e., non-taeniodont pattern). The third transverse crest is strong, complete and links the mesial extremity of the oblique anterior arm of the hypocone (AAH) to a strong mesostyle labially. A strong, short, and longitudinal mure connects the third transverse crest-AAH junction to the lingual extremity of the labial protoloph, and closes the hypoflexus labially. Distolabially, there is a small metaloph, posterolabially directed, labially connected to the metacone/posteroloph and lingually to the mesolophule. The lingual connection of the metaloph with the third transverse crest is extended and includes a tiny fossette, which can be a part of the metaflexus. Its separation from the rest of the metaflexus is probably due to the strong occlusal wear. Mediodistally, there is a longitudinal accessory crestule linked to the posteroloph and almost to the metaloph. Because of the advanced state of wear, all flexi are entirely closed, except the hypoflexus which remains open lingually.
Comparisons. This specimen is comparable in size with Cachiyacuy kummeli, Canaanimys maquiensis, Eoespina woodi, and Eosachacui lavocati (Frailey and Campbell, 2004). The absence of taeniodont pattern of MUSM 2654 (Figure 5.11) excludes affinities with Eoincamys, Eobranisamys, and Vallehermosomys mazzonii (Frailey and Campbell, 2004; Vucetich et al., 2010) . Although the M3s of C. maquiensis tend to have a lingual connection between the protocone and the protoloph (pseudo-taeniodont pattern), its M1-2s show a clear taeniodont pattern. The presence of a well-developed metaloph on MUSM 2654 is in sharp contrast with the condition found in E. woodi, E. lavocati, ?Vallehermosomys merlinae, and Draconomys verai (Frailey and Campbell, 2004; Vucetich et al., 2010), in which this crest is absent, incipient or corresponding to a thickened posteroloph. In Cachiyacuy and Eosallamys, the metaloph tends to be more developed, like on MUSM 2654. In Cachiyacuy, the metaloph is transverse, and its lingual extremity can be connected to the third transverse crest and posteroloph via accessory crestules. In Eosallamys, the metaloph is usually only linked lingually to the posteroloph but on some upper molars (LACM 143422, 143363, 143376, 143380; in Frailey and Campbell, 2004, p. 120-122), it can be connected to the third transverse crest via an accessory crestule. Nevertheless, MUSM 2654 shows a smaller size, more slender crests with thinner enamel than upper molars of Eosallamys, and as such it appears closer in morphology and proportions to the upper molars of Cachiyacuy.
?Cachiyacuy kummeli
Figure 5.3, Appendix 3
2016 ?Canaanimys sp. Antoine et al., Supplementary data, p. 5.
2017 ?Canaanimys sp. Antoine et al., Supplementary data, p. 9.
Referred material. MUSM 2648, fragmentary left dP4 (Figure 5.3).
Locality. Contamana CTA-47, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016). This is the lowermost locality having yielded rodents in the Quebrada Cachiyacu section of Contamana (Antoine et al., 2016).
Description. MUSM 2648 (Figure 5.3) is a labial fragment of a brachydont upper tooth. Due to the trapezoidal shape of the occlusal outline, this is identified as a dP4. The paracone is clearly visible because it is the largest cusp of the tooth. In its mediolabial part, the anteroloph bears a tiny and backwardly directed spur that reaches a second spur linked to the protoloph. There is a lingual connection between the anteroloph and the protoloph due to wear. Because of the important thickness of that link, it may be expected that this dP4 did not exhibit a taeniodont pattern when it was pristine. There is a very thin and low crestule, distally placed to the protoloph, and labially linked to a minute cusp. These structures are too small and anteriorly placed to be identified as the main central transverse crest and mesostyle, given that these latters are well-defined and situated just posteriorly. As such, they appear to be accessory dental traits. With the advanced degree of wear, it is difficult to know if the lingual end of that secondary crestule is free or linked to the central transverse crest. The latter, running labiodistally from the junction between the mure and the mesial extremity of the anterior arm of the hypocone to the mesostyle, corresponds to a mesolophule. Due to wear, the enamel of the distal flank of the mesolophule reaches one of the last distal crests, a configuration which forms a wide enamel plate rather than a metaflexus. The dentine part of the apparent fourth crest is wide and slightly excavated, which suggests that it might include both a metaloph and a posteroloph. The metaflexus is strongly reduced and limited to a very small and faintly visible fossette.
Comparisons. MUSM 2648 (Figure 5.3) has a size close to that of Cachiyacuy kummeli, Canaanimys maquiensis, Eoespina woodi, and Eosachacui lavocati (Frailey and Campbell, 2004). In Antoine et al. (2016, 2017), this tooth was attributed to ?Canaanimys on the basis of a different interpretation of dental homologies from that proposed here (see description). Topographically, the third apparent transverse crest was originally interpreted as a metaloph (with a configuration as that observed on upper molars of Canaanimys [i.e., oriented anteriorly]), while here it is interpreted as a mesoloph. Likewise, the posteriormost crest, which was originally interpreted as a posteroloph, might be a fusion of the metaloph with the posteroloph instead. Finally, the central tiny crestule is reinterpreted here as a neostructure instead of a mesoloph. If our current interpretations regarding the homologies are correct, this pattern characterized by a metaloph associated with the posteroloph is reminiscent of that found in Cachiyacuy contamanensis. Moreover, Cachiyacuy is the only genus that often displays a central crestule (more or less long) and/or cusp between the protoloph and the third transverse crest on dP4 (MUSM 1874, 2717, 2720, and 2721; in Antoine et al., 2012, figure 2e) and on upper molars (MUSM 1871 and 2742; in Antoine et al., 2012, figure 2b). On these teeth, the accessory crestule and/or cusp is on the distal wall of the paracone and protoloph, except on two teeth: on MUSM 2717, the accessory cusp is neither on the wall of the paracone nor on that of the mesostyle but is located on the opening of the mesoflexus, while on MUSM 1874, the accessory crestule and cusp are closer to the central transverse crest, like on MUSM 2648. To summarise, MUSM 2648 has more affinities with the upper molars of C. kummeli.
Genus CANAANIMYS Antoine et al., 2012
Type species.Canaanimys maquiensis Antoine et al., 2012.
Species content. Only the type species.
Geographic and stratigraphic distribution. Contamana CTA-27; Pozo Formation, lower member (late middle Eocene); Loreto Department, Peru.
Canaanimys maquiensis, Antoine et al., 2012
Figure 4.14-16, Appendix 3
Referred material. In addition to the holotype (MUSM 1890, right M2; in Antoine et al., 2012, figure 2u) - MUSM 1895, right dp4 (in Antoine et al., 2012, figure 2z); MUSM 1894 (in Antoine et al., 2012, figure 2y) and 2787-2788, left m1s; MUSM 2789-2790, right m1s; MUSM 1893, left m2 (in Antoine et al., 2012, figure 2x); MUSM 1892 (in Antoine et al., 2012, figure 2w) and 2791, right m3s; MUSM 1891, left M1 (in Antoine et al., 2012, figure 2v); MUSM 2792, left M3 (Figure 4.16); MUSM 1889 (in Antoine et al., 2012, figure 2t) and 2793-2794 (Figure 4.14-15), right M3s.
Type Locality. Contamana CTA-27, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2012, 2016).
Emended diagnosis (modified after Antoine et al., 2012, p. 1321). Tiny rodent (body mass estimated at approx. 40 g) characterized by brachydont and bunolophodont teeth. It differs from all other caviomorphs in having teeth with moderately low and sharp transverse crest(id)s, lower molars having generally incomplete second transverse cristid, and pentalophodont upper molars with a well-developed metaloph that turns anteriorly (not posteriorly) and connects either to the third transverse crest lingually or to the mesial extremity of the anterior arm of the hypocone, a primitive condition that is reminiscent of that found in stem Hystricognathi (e.g., Baluchimys, Protophiomys, Waslamys, Phiomys, Hodsahibia, Bugtimys and Ottomania). The lingual protoloph is either slightly developed or lacking (i.e., taeniodont pattern).
Description. The lower dentition is documented by several molars and one deciduous premolar. There is no permanent premolar identified in the recovered material.
The dp4 is damaged in its mesial part, and as such, it is impossible to describe the protoconid region (in Antoine et al., 2012, figure 2z). This tooth appears somewhat slender, with thin and low transverse and longitudinal cristids. The main cuspids are salient and well-defined. The metaconid is labiolingually compressed and bears a posterior arm that extends backwardly and connects a strong mesostylid. The latter is as large as the metaconid. There is a supernumerary cuspid (nearly reaching the size of the mesostylid) occupying a central position within the mesoflexid and that could be related to a local enlargement of a mesolophid. Thin enamel cristulids radiate from this central cuspid, directed either to the mesostylid, to the ectolophid, or forward. The latter, the longest one, may correspond to the posterior arm of the protoconid. The ectolophid is complete and does not bear any mesoconid. On the talonid, the hypoconid and entoconid are labiolingually opposed, and linked together by a long hypolophid that connects a thin and short anterior arm of the hypoconid. The latter, grooved in its middle part, is formed by two cristulids. Distally, the hypoconid and entoconid are linked by a massive and curved posterolophid. This latter plus the hypolophid isolate a broad metaflexid.
Lower molars (m1-3s) share the same dental structure (in Antoine et al., 2012, figure 2w-y). On m1s, the protoconid and metaconid, as well as the hypoconid and entoconid, are labiolingually opposed. On the unique m2 recognized, the same is true for the mesial cuspids. In contrast, the m2 talonid cuspids differ in showing the entoconid noticeably displaced mesially with respect to the hypoconid. On pristine teeth, cuspids are salient and clearly dominate the cristids, which remain relatively low (moderately elevated). Mesially, there is no trace of an anterocingulid at the neck. Lingually, between the metaconid and entoconid, generally there is a prominent and isolated mesostylid. There is no strong or elevated posterior arm of the metaconid. On the only unworn lower molar (MUSM 1893; in Antoine et al., 2012, figure 2x), the metaconid-mesostylid connection is absent. On other molars, the metaconid is connected to the mesostylid but the latter remains distinct, albeit minute. Like in Cachiyacuy, the second transverse cristid is highly variable and always composite. It can be continuous labiolingually (i.e., long posterior arm of the protoconid + long neomesolophid; MUSM 1892 and 2790; in Antoine et al., 2012, figure 2w), but it is usually interrupted and divided into two (i.e., long posterior arm of the protoconid + short neomesolophid MUSM 2787) or three parts (i.e., long posterior arm of the protoconid short neomesolophid + accessory cuspid; MUSM 1893, 1894, and 2789; in Antoine et al., 2012, figure 2x-y). There is no mesoconid on the complete and obliquely oriented ectolophid. The hypolophid is strong, trenchant, and slightly oblique. Labially, it connects a short anterior arm of the hypoconid, sometimes grooved in its middle part as for the dp4. The posterolophid is massive and strongly connected to the hypoconid, without a hypoconulid. The posterolophid runs lingually and slopes gently to finish its course distolingually at the base of the entoconid.
There is neither deciduous nor permanent upper premolar recovered for this taxon.
The upper molars are brachydont and bunolophodont (in Antoine et al., 2012, figure 2t-v; Figure 4.14-16). The transverse crests are thin and low; the main cusps are well-marked and salient as a result. There is no conule. We identify as M1 a rectangular tooth that is slightly wider than long. While M1 has a hypocone distally opposed to the protocone, the M2 is recognizable by its hypocone, slightly smaller and more labial. M3s are rectangular with a labiolingual long axis (MUSM 1889 [in Antoine et al., 2012, figure 2t] and 2794 [Figure 4.14]) or mesiodistally expanded on the labial part (MUSM 2792 and 2793; Figure 4.15-16). The hypocone of the M3s occupies a more labial position than it does on M2. On upper molars, the protocone, mesially canted, usually develops a strong and long posterior outgrowth, which nearly closes the hypoflexus lingually. Mesially, the protocone is connected to an anteroloph running labially to join the base of the paracone mesially, thereby closing the paraflexus labially. There is no trace of parastyle. The protoloph, long and oblique (slightly backwardly directed), does not reach the protocone. There is no lingual protoloph, and as such, the hypoflexus is confluent with the paraflexus, thereby involving a taeniodont pattern. However, some M3s (MUSM 1889, 2792, and 2794) may have a residual lingual protoloph: a thin, very low, and short spur extends from the protoloph lingually to the protocone, or another crestule stemming from the protocone. On M1-3, the anterior arm of the hypocone is thin and moderately long, running mesiolabially. A thin, short, and longitudinal mure links the lingual extremity of the protoloph to the anterior arm of the hypocone. The third transverse crest is a thin and long mesolophule running labially to reach the lingual base of a small and isolated mesostyle. Unlike MUSM 1889 and 1890 (in Antoine et al., 2012, figure 2t-u), the linguolabial course of the mesolophule is sinuous on all other upper molars. On MUSM 2793 (Figure 4.15), the mesolophule is lower and more oblique (i.e., labiomesially oriented), and it displays a weaker connection with the anterior arm of the hypocone. On the unique M1 specimen (MUSM 1891; in Antoine et al., 2012, figure 2v), the mesolophule, shorter, does not reach the labial margin. On this tooth, the mesostyle is practically indistinct, as it is entirely merged with the metacone. Interestingly, some upper molars of Canaanimys maquiensis (MUSM 1889, 1890, and 1891) exhibit a very unusual arrangement of the metaloph among caviomorphs. Indeed, the metaloph is slightly oblique, forwardly directed, and connected either to the mesolophule lingually or to the mesial extremity of the anterior arm of the hypocone. On some M3s (MUSM 2792 and 2793; Figure 4.15-16), a small crestule stems from a cusp on the posteroloph, likely the relic of a metaloph or a secondary crestule. This crestule may be absent (on MUSM 2794; Figure 4.14). Distally, the teeth show a well-developed posteroloph that connects the hypocone to the base of the metacone. The posteroflexus is generally closed labially (= posterofossette), but in some cases (e.g., MUSM 1890 and 1891), a narrow and very shallow furrow persists between the posteroloph and the metacone.
Comparisons. Canaanimys differs from all other caviomorphs in having teeth with moderately low and sharp transverse crests and cristids, a long outgrowth of the protocone on upper molars, and pentalophodont M1-2 with a well-developed metaloph that turns anteriorly (not posteriorly) and connects either to the mesolophule lingually or to the mesial extremity of the anterior arm of the hypocone ̶ ̶ a plesiomorphic condition reminiscent of that found in stem Hystricognathi (e.g., Baluchimys Flynn et al., 1986, Protophiomys Jaeger et al., 1985, “Waslamys ” Sallam et al., 2009, Phiomys Osborn, 1908, Hodsahibia Flynn et al., 1986, Bugtimys Marivaux et al., 2002, or Ottomania de Bruijn et al., 2003). The lingual protoloph is either slightly developed or lacking (i.e., taeniodont pattern) like in Eobranisamys, Branisamys, Eoincamys and Incamys (Hoffstetter and Lavocat, 1970; Lavocat, 1976; Frailey and Campbell, 2004). As for Cachiyacuy, Canaanimys differs from Eoincamys and Incamys in having brachydont instead of hypsodont teeth, pentalophodont instead of tetralophodont upper molars, and thinner and transverse instead of strong and oblique crests. Canaanimys has transverse crests labiolingually longer on its upper molars than in Draconomys from La Cantera (Argentina, late early Oligocene; Vucetich et al., 2010), and a composite second cristid including a neomesolophid and a posterior arm of the protoconid on lower molars, whereas lower molars of Draconomys lack a neomesolophid.
?Canaanimys sp.
Figure 5.1-2, Appendix 3
2016 ?Canaanimys sp. Antoine et al., Supplementary data, p. 5.
2017 ?Canaanimys sp. Antoine et al., Supplementary data, p. 9.
Referred material. MUSM 2645, fragmentary right dp4 (Figure 5.1); MUSM 2646, fragmentary left lower molar (Figure 5.2).
Locality. Contamana CTA-47, Loreto Department, Peru. This is the lowermost locality having yielded rodents in the Quebrada Cachiyacu section of Contamana (Antoine et al., 2016).
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Description. The brachydont dp4 (MUSM 2645; Figure 5.1) is broken in its posterior part, lingually at the level of the hypolophid and labially at the level of the labial end of the posterolophid. Despite the wear, the protoconid, metaconid, and mesostylid are well-recognizable. The metalophulid I is curved and connected to the protoconid and metaconid. This cristid is thick except in its labial end, which tapers before connecting the protoconid. The posterior arm of the protoconid does not reach the metaconid. This cristid ends at mid-line of the tooth width. The mesial ectolophid is separate from the protoconid by a wide labial opening of the mesial mesoflexid. The posterior arm of the metaconid runs distally and reaches the mesostylid. Labially, at the intersection of the mesial ectolophid and distal one, there is a tiny spur, directed toward the mesostylid, probably corresponding to a very short mesolophid. There is a labial connection between the hypolophid and the posterolophid (no taeniodont). Given the short posterior arm of the protoconid and mesolophid, the anteroflexid, mesial mesoflexid, and distal mesoflexid are merged. That mixed flexid is lingually open between the mesostylid and entoconid, as well as labially between the protoconid and the mesial ectolophid.
MUSM 2646 (Figure 5.2) is a mesial fragment of a lower molar. Only the metalophulid I and a part of the second transverse crestid are visible. The latter is reduced and composed of different structures (a posterior arm of the protoconid and probably a neocristid).
Comparisons. These teeth have a size close to that of Cachiyacuy kummeli, Canaanimys maquiensis, and Eoespina/Eosachacui (Frailey and Campbell, 2004). In MUSM 2645 (Figure 5.1), the presence of a short posterior arm of the protoconid (not connected to the metaconid) and a strong posterior arm of the metaconid recall the conditions found in C. maquiensis. However, the discontinuity of the second transverse cristid and its reduction (although its labial part remains well-defined) is also found in Draconomys verai (Vucetich et al., 2010). Even though the general morphology of MUSM 2645 appears quite primitive by some characters (e.g., brachydonty and absence of taeniodonty), this tooth differs from the dp4s characterizing pre-Deseadan species for which this dental locus is known in having a mesolophid virtually undeveloped. A reduced mesolophid is also observed in dp4s of Cachiyacuy contamanensis and Canaanimys maquiensis but, in these taxa, this cristid is short and still well-recognizable. Given the scarcity and damaged condition of the available material, we tentatively and provisionally assign these teeth to ?Canaanimys sp.
Gen. et sp. indet. 1
Figure 5.4, Appendix 3
2016 ?Canaanimys sp. Antoine et al., Supplementary data, p. 5.
2017 ?Canaanimys sp. Antoine et al., Supplementary data, p. 9.
Referred material. MUSM 2647, fragmentary right lower molar (Figure 5.4).
Locality. Contamana CTA-47, Loreto Department, Peru. This is the lowermost locality having yielded rodents in the Quebrada Cachiyacu section of Contamana (Antoine et al., 2016).
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Description. MUSM 2647 (Figure 5.4) is the labial part of a right lower molar. Mesially, the tooth is broken along the posterior arm of the protoconid. Distally, only the labial extremity of the hypoconid apex is present. As the mesiolabial outgrowth of the hypoconid is not curved, the hypoflexid is straight and transverse, which clearly indicates that this specimen is a lower molar.
The size of MUSM 2647 is comparable to that of Cachiyacuy kummeli and Canaanimys maquiensis but, due to its fragmentary state, it is impossible to provide a formal taxonomic assignment of this specimen.
Gen. et sp. indet. 2
Figure 5.8-10, Appendix 3
2016 Cachiyacuy aff. kummeli Antoine et al., Supplementary data, p. 5.
2016 Canaanimys cf. maquiensis Antoine et al., Supplementary data, p. 5.
2017 Cachiyacuy aff. kummeli Antoine et al., Supplementary data, p. 9.
2017 Canaanimys cf. maquiensis Antoine et al., Supplementary data, p. 9.
Referred material. MUSM 2656, fragmentary right lower molar (Figure 5.8); MUSM 2657 left dP4 (Figure 5.9); MUSM 2658, fragmentary left upper tooth (Figure 5.10).
Locality. Contamana CTA-51, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Description. The lower molar (MUSM 2656; Figure 5.8) is heavily worn and broken posteriorly. It is tetralophodont and non-taeniodont. The second transverse cristid appears continuous but it is not rectilinear, its lingual part being distally displaced. This configuration suggests that this cristid is an association of two cristids: labially, a long, straight, and lingually oriented posterior arm of the protoconid, and labially, a short and labially oriented neomesolophid. The latter cristid originates from a faintly visible mesostylid, situated in the distal most part of a strong and long posterior arm of the metaconid.
The dP4 (MUSM 2657; Figure 5.9) is damaged and eroded. This tooth is longer than wide, tetralophodont and non-taeniodont. A fractured long lingual root avoids any confusion with lower molars. The protoloph is slightly oblique (a few linguodistally directed) and parallel to the transverse and adjacent third transverse crest. Although complete, the third transverse crest is very thin in its lingual tip (where it connects the anterior arm of the hypocone) and forms a short spur forwardly oriented. This arrangement indicates that this crest is composed of a very short lingual mesolophule and a long labial mesoloph. The mure is mesiolabially directed and continuing the oblique anterior arm of the hypocone. The posteroloph is a little thickened, but neither the metaloph nor the metacone are distinct. Given the degree of wear of the tooth, it is difficult to tell if the paraflexus and mesoflexus remain open or if they are closed labially. The most posterior flexus (confluence of the metaflexus with the posteroflexus?) is labially closed, thus forming a fossette. The paraflexus and mesoflexus are roughly equal in surface whereas the most posterior flexus is more extensive, notably in width.
MUSM 2658 (Figure 5.10) shows a similar general pattern to that of the dP4 (MUSM 2657; Figure 5.9), except for the mure configuration, which is longitudinal, and for the paraflexus and the most posterior flexus, which are equally sized. Moreover, on this tooth, the posteroloph displays a tiny spur that may correspond to a residual metaloph.
Comparisons. Although eroded, these teeth have a size compatible to that of the molars of Cachiyacuy kummeli, Canaanimys maquiensis, and Eoespina woodi/Eosachacui lavocati (Frailey and Campbell, 2004). Concerning these specimens from CTA-51, the morphology and evolutionary stage of their dental pattern is characteristic of most early caviomorphs, notably the pre-Deseadan ones. The MUSM 2657 dP4 is tetralophodont, while in Cachiyacuy contamanensis, this tooth is pentalophodont. It is worth noting that the tetralophodonty and non-taeniodonty of MUSM 2657 (Figure 5.9) are reminiscent to the conditions found on the upper molars of E. woodi/E. lavocati or even Cachiyacuy kummeli. Hence, we prefer to assign MUSM 2658 to “Caviomorpha gen. et sp. indet. 2,” pending new findings.
Gen. et sp. indet. 3
Figure 5.14, Appendix 3
2016 Caviomorpha indet., sp. 1 Antoine et al., Supplementary data, p. 6.
2017 Caviomorpha indet., sp. 1 Antoine et al., Supplementary data, p. 9.
Referred material. MUSM 2660, left upper molar (Figure 5.14).
Locality. Contamana CTA-73, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Description. This single tooth (MUSM 2660; Figure 5.14) is heavily worn and damaged. It is small, brachydont, and non-taeniodont. The protoloph is slightly oblique, well-separated, and non-parallel to the third transverse crest (mesolophule or mesoloph?). A short and faintly visible metaloph might be present, but it is entirely merged with the posteroloph. The paraflexus and the most posterior flexus (confluence of the metaflexus with the posteroflexus) are labially closed, thus forming a fossette.
Comparisons. The size of this specimen is comparable to that of Cachiyacuy kummeli, Canaanimys maquiensis, and Eoespina woodi/Eosachacui lavocati (Frailey and Campbell, 2004). The shape of the occlusal outline of upper molars is similar to that of E. woodi/E. lavocati, which is less transverse than that characterizing upper molars of C. kummeli and C. maquiensis. Given the lack of diagnostic characters, we provisionally assign this tooth to a Caviomorpha gen. et sp. indet. 3.
Gen. et sp. indet. 4
Figure 5.13, Appendix 3
2016 Caviomorpha indet. sp. 2, Antoine et al., Supplementary data, p. 6.
2017 Caviomorpha indet. sp. 2, Antoine et al., Supplementary data, p. 9.
Referred material. MUSM 2659, fragmentary right lower molar (Figure 5.13).
Locality. Contamana CTA-73, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Description. The specimen MUSM 2659 (Figure 5.13) is a small fragment of the anterior part of a right lower molar. The metalophulid I, second transverse cristid, and hypolophid are recognizable. The second transverse cristid is complete but not straight, and seems to be a mixed structure including a part of the posterior arm of the protoconid labially and, lingually, a neomesolophid. Both cristids are similar in length. The branch of the posterior arm of the protoconid bears a tiny spur on its posterior edge. The ectolophid is oblique. The anteroflexid and mesoflexid appear closed lingually. This lingual closure is due to the strong development of a long posterior arm of the metaconid, which incorporates a faintly visible mesostylid, and also due to the connection between the posterior arm of the mesostylid and the anterior arm of the entoconid.
Comparisons. The morphology of this anterior fragment of lower molar is comparable to that found in most of Eocene/early Oligocene caviomorphs (all taxa from CTA-27, 29, 51 and Santa Rosa [except Eoincamys and Eopululo Frailey and Campbell, 2004]), notably by the configuration of the second transverse cristid (a neomesolophid + a long posterior arm of the protoconid). Its size is close to that of lower molars of C. cf. contamanensis from CTA-51, which is slightly smaller than that of C. contamanensis from CTA-27. We provisionally refer this material of CTA-73 to Caviomorpha gen. et sp. indet. 4.
Gen. et sp. indet. 5
Figure 6.13, Appendix 3
2016 Cachiyacuy cf. kummeli Antoine et al., (Supplementary data, p. 9).
2017 Cachiyacuy cf. kummeli Antoine et al., (Supplementary data, p. 9).
Referred material. MUSM 2838, right dp4 (Figure 6.13).
Locality. Contamana CTA-29, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Description. The dp4 (MUSM 2838; Figure 6.13) is worn and shows a well-marked distal contact facet. The talonid is almost twice as wide as the pinched labiolingually trigonid. It is pentalophodont with large, complete cristids and cuspids still well-defined and bulbous (except the metaconid), and it is non-taeniodont. Mesially, the protoconid is labially opposed to the metaconid, and these two cuspids are linked mesially by a curved metalophulid I, and distally by a straight and complete metalophulid II. Mediolabially, the mesial branch of the ectolophid is linguolabially oriented and not connected to the protoconid mesially. With this cristid arrangement, the hypoflexid is narrow and not mesially extensive. A strong and slightly oblique mesolophid runs mesiolingually from the ectolophid, to reach the lingual margin, where it is connected to a strong mesostylid. This latter is entirely merged with the short but elevated posterior arm of the metaconid. With the mesolophid connection and the mesial ectolophid arrangement, the mesial mesoflexid is closed lingually but open labially. The hypoconid and entoconid are labiolingually opposed. The hypoconid displays a strong and long anterior arm that connects to a short hypolophid stemming from the entoconid. The distal ectolophid is strong, almost longitudinal, and links the mesolophid and anterior arm of the hypoconid/hypolophid junction. Lingually, neither the mesostylid nor the lingual end of the posterolophid is linked to the entoconid distally and mesially, respectively. The anteroflexid is the only flexid closed lingually (= anterofossettid).
Comparisons. MUSM 2838 (Figure 6.13) is tetralophodont like the dp4s of most pre-Deseadan caviomorphs, except Eobranisamys and Branisamys (Hoffstetter and Lavocat, 1970; Lavocat, 1976; Frailey and Campbell, 2004), which have hexalophodont dp4s. MUSM 2838 shares with the dp4 referred to C. maquiensis (MUSM 1895) and some dp4s of C. contamanensis (MUSM 2663, 2670, and 2669), a lingual connection between the metaconid and the mesostylid, contrary to all other pre-Deseadan caviomorph species documented by dp4s. Yet, on MUSM 2838, the second transverse cristid does not form a longitudinal cristid, connected to the mesolophid, contrary to what is observed in dp4s of C. maquiensis. On MUSM 2838, the protoconid is clearly separate from the mesial ectolophid, as in Eosallamys simpsoni and some dp4s of Eoespina/Eosachacui (as well as in Eobranisamys and Branisamys; Hoffstetter and Lavocat, 1970; Lavocat, 1976; Frailey and Campbell, 2004). This character is not found in Cachiyacuy and Eosallamys paulacoutoi. Thus, this specimen from CTA-61 is provisionally identified as Caviomorpha gen. et sp. indet. 5.
Gen. et sp. indet. 6
Figure 6.14, Appendix 3
2016 Caviomorpha indet., sp. 2 Antoine et al., Supplementary data, p. 9.
2017 Caviomorpha indet., sp. 2 Antoine et al., Supplementary data, p. 9.
Referred material. MUSM 2839, right M1 or 2 (Figure 6.14).
Locality. Contamana CTA-29, Loreto Department, Peru.
Formation and age. Pozo Formation, lower member, late middle Eocene (Antoine et al., 2016).
Description. The upper molar (MUSM 2839; Figure 6.14) is particularly worn and eroded, and also distolabially broken. Its crown outline is quadrate in occlusal view. There are four crests but the posteroloph exhibits a short enamel spur on its mesial edge (remnant metaloph?). The anteroloph and protoloph are linked labially and broadly merged lingually due to wear. This tooth is non-taeniodont, and the mure is longitudinal and well-differentiated with respect to the oblique anterior arm of the hypocone and the transverse protoloph. Centrally, there is a strong but moderately long mesolophule, transverse and parallel to the protoloph. Labially, the mesolophule is connected to a strong mesostyle. Although the upper molar is broken distolabially (at the place where a metacone would have occurred), a small enamel ridge indicates that the mesostyle was connected to the metacone-posteroloph complex. In contrast, mesially, the mesostyle remains well-separate from the paracone (mesoflexus labially open). The parafossette is small, narrow, and labially limited, while the most posterior fossette (confluence between the metaflexus and posteroflexus) is broader and more lingually extended.
Comparisons. This upper molar is the smallest specimen discovered from CTA-29. The small size of this tooth, its quadrate shape and its tetralophodonty (with a possible relic of a metaloph), recall the dental pattern of Eoespina/Eosachacui . However, as for MUSM 2660 from CTA-73, due to wear, it is impossible to propose a formal taxonomic assignment.
DISCUSSION
Eocene Localities of Contamana: A Homogeneous Set of Assemblages?
A total of 20 caviomorph taxa are identified from the Contamana localities (including six taxa referred to Caviomorpha indet.): three in CTA-47, four in CTA-51, two in CTA-73, five in CTA-27, one in CTA-66, and five in CTA-29 (Table 1). In addition to be geographically close (within a perimeter of ~5 km2), the Eocene localities of Contamana yield very similar mammal assemblages (Antoine et al., 2016), which is true for the rodent faunas as well. It is worth noting that many rodent taxa (12/20) are attributed to Cachiyacuy contamanensis, Cachiyacuy kummeli, Canaanimys, Eobranisamys, and Eoespina or closely related forms (six taxa left in open nomenclature). CTA-29 may appear somewhat distinct from the other Eocene localities of Contamana in recording a new genus and species: Pozomys ucayaliensis. Startlingly, all these recorded taxa share similar dental characters: small size, brachydonty, bunolophodonty, thin and transverse loph(-id)s, and tetralophodont lower molars (except few molars attributed to Cachiyacuy contamanensis, which tend to be pentalophodont). All these features, together with the pentalophodonty of M1-2s observed in most of taxa, are also found in the earliest Afro-Asian hystricognaths (“phiomyids” and “baluchimyines”; Antoine et al., 2012). As the result of their close morphological and phylogenetic proximity with some Afro-Asian “phiomorphs” (Antoine et al., 2012; Marivaux et al., 2014; Barbière and Marivaux, 2015), Cachiyacuy and Canaanimys are considered as stem Caviomorpha (Antoine et al., 2012). Other taxa mentioned in this study, the cavioid Eobranisamys, the octodontoid Eoespina , and the caviomorph incertae sedisPozomys, display a combination of characters, which appear somewhat more derived, such as: no connection of the metaloph with the third transverse crest or the anterior arm of the hypocone, taeniodont upper teeth (in Eobranisamys and at least in M2s of Pozomys), a reduction of crests on P4 (in Eobranisamys ), a reduction or even the absence of the metaloph and metacone on upper molars (notably in Eoespina and Pozomys), and pseudo-taeniodont lower molars (in Pozomys). Interestingly, the metaloph can be also reduced or absent on some upper molars of Cachiyacuy and Canaanimys.
All Eocene localities of Contamana consist of sandstone lenses deposited within a low-energy small stream (Antoine et al., 2016). All the concerned palaeontological assemblages show both aquatic and terrestrial affinities, with consistent mollusc, fish, and mammalian assemblages (e.g., Antoine et al., 2017). Furthermore, the palynoflora found at CTA-47 and CTA-27 points to the persistence of a tropical rainforest habitat during the concerned interval (Antoine et al., 2012, 2016), even if CTA-47 differs from other Eocene assemblages in recording dinoflagellates, thereby clearly suggesting a marine influence.
To sum up, the Eocene localities of Contamana constitute a homogeneous set from a geographical, taxonomic, and environmental perspective, contrary to the Deseadan (CTA-32 and CTA-61; Boivin et al., 2017) and Neogene ones (Antoine et al., 2016). The latter are characterized by distinct environmental/depositional conditions, leading to different taxonomic assemblages in terms of composition and diversity.
Comparison with Other Paleogene Caviomorph-bearing Localities and Biostratigraphic Implications
Among the pre-Deseadan caviomorph-bearing localities formerly described (Termas del Flaco, Chile [Wyss et al., 1993; Bertrand et al., 2012]; Santa Rosa, Peru [Frailey and Campbell, 2004]; and La Cantera, Argentina [Vucetich et al., 2010]), the rodent assemblages of the Eocene localities from Contamana are more similar to that of Santa Rosa, which might more primarily reflect biogeographic affinities within a tropical-equatorial region (further known to yield long-lasting lineages) than contemporaneity (see Antoine et al., 2017). Indeed, three localities of Contamana (CTA-51, CTA-27, and CTA-66) record at least one genus in common, which was originally described in Santa Rosa (Eobranisamys and/or Eoespina). The CTA-27 locality yields representatives of both genera, including a new species of Eobranisamys, E. javierpradoi that is smaller than its counterparts from Santa Rosa. The localities CTA-51 and CTA-27 testify to the earliest occurrence of Eoespina and Eobranisamys, respectively. Contrastingly, the Eocene localities of Contamana do not share any genus/species with Termas del Flaco, Chile (~35°S) or La Cantera, Patagonia Argentina (~45°S). Additionally, there are only brachydont rodents at Santa Rosa, in La Cantera, and in the Eocene localities of Contamana, contrary to Termas del Flaco (where is recorded the hypsodont taxon of chinchilloid affinities Eoviscaccia frassinettii Bertrand et al., 2012). Finally, Eosallamys from Santa Rosa (E. paulacoutoi and E. simpsoni) is morphologically close to Canaanimys and Cachiyacuy. Indeed, Eosallamys is the only other caviomorph taxon developing a lingual connection between the metaloph and the third transverse crest on M1-2s, as in Canaanimys and Cachiyacuy, a plesiomorphic character otherwise found only in the earliest Afro-Asian hystricognaths. Thanks to the present work, the occurrences of Cachiyacuy range now from CTA-51 to CTA-29, while Canaanimys is reliably documented only at CTA-27.
The Eocene rodents from Contamana do not display any affinity with Deseadan taxa known in South America, such as those from Cabeza Blanca in Argentina or Salla in Bolivia (i.e., no genera/species in common). Besides, any Eocene rodent assemblage of Contamana yields a low taxonomic richness (at suprafamilial and specific levels) associated with a low morphological disparity (only brachydont taxa with thin loph[-id]s), notably compared with that of Santa Rosa (Frailey and Campbell, 2004) and well-documented Deseadan localities (CTA-32, CTA-61, Salla, and Cabeza Blanca). Put together, with the presence of caviomorphs showing very primitive features (i.e., Cachiyacuy and Canaanimys ), these low diversity and disparity suggest that a short time range of caviomorph evolutionary history had undergone in South America prior this early Contamanan rodent record, as already proposed by Antoine et al. (2012). Although these localities potentially document the first representatives of octodontoids ( Eoespina) and cavioids (Eobranisamys), they probably predate the early adaptive radiation(s) of Oligocene-Neogene caviomorph communities (including obligate representatives of extant superfamilies), which is consistent with the results of Arnal and Vucetich (2015) and Vucetich et al. (2015).
CONCLUSIONS
The late middle Eocene caviomorph assemblages from Contamana, Peruvian Amazonia, encompassing at least 20 taxa, highlight the early diversity and great antiquity of that South American mammal group at low latitudes. Three caviomorph taxa have been identified in CTA-47 locality, four in CTA-51, two in CTA-73, five in CTA-27, one in CTA-66, and five in CTA-29. In the latter locality, a new taxon of uncertain suprafamilial affinities is described: Pozomys ucayaliensis gen. et sp. nov. Besides, the material from CTA-27 previously attributed to Eobranisamys sp. is assigned here to a new species, Eobranisamys javierpradoi. In terms of taxonomic composition, all Eocene localities of Contamana have many taxa in common (Cachiyacuy, Canaanimys, Eobranisamys, and Eoespina, or very close relatives), which is consistent with roughly similar depositional environments and accumulating factors, and with a short stratigraphic range between these localities. Cachiyacuy is the most common taxon, being recorded in CTA-51, CTA-27, and CTA-29. Compared with other pre-Deseadan caviomorph faunas, the assemblages studied here only share two genera with the Santa Rosa fauna ( Eobranisamys and Eoespina). In addition, the pattern of some upper molars of Eosallamys found at Santa Rosa is reminiscent of that of Cachiyacuy or Canaanimys from Contamana, thus likely to testify to the presence of a Paleogene lineage of stem caviomorphs restricted to tropical South America. As a consequence, these rodent assemblages provide a better understanding of the early evolutionary history of caviomorphs for at least two reasons: 1) they contribute to complete the pre-Deseadan fossil record of this group, which is known by few localities; and 2) they document the oldest undisputable representatives of caviomorphs and among the earliest stages of evolution of that group.
A cladistic assessment of the dental evidence available must now be undertaken in order to formalize the phylogenetic position of these early taxa within a high level caviomorph phylogenetic framework (work in progress). Given the recent groundbreaking paleontological potential of Peruvian Amazonia for highlighting the early evolutionary history of many mammalian clades (e.g., Bond et al., 2015; Antoine et al., 2016, 2017; Marivaux et al., 2016), paleontological field efforts must be sustained, especially in these regions, in order to better characterize the early adaptive radiation of caviomorphs, and to further our understanding of the emergence of the four extant super-families and their historical biogeography in South America.
ACKNOWLEDGMENTS
We especially thank the Canaan Shipibo Native Community and Maple Gas Peru S.A. for granting access to the field and the IRD-Peru-Petro Convention Programme for logistic support. We are particularly grateful to our guides Manuel and Manuel, for their long standing help in the forest during the field seasons. Many thanks to S. Adnet (ISEM, Montpellier, France), A.J. Altamirano-Sierra (MUSM, Lima, Peru), G. Billet (MNHN, Paris, France), A. Boada-Saña (Spain), F. Duranthon (Muséum de Toulouse, France), and whoever helped us in the field and in the lab. We are much indebted to M.E. Pérez (MPEF, Trelew, Argentina), M.G. Vucetich, and A. Candela (MLP, La Plata, Argentina), and A.G. Kramarz (MACN, Buenos Aires, Argentina) for discussing on rodent affinities, and providing crucial bibliographic references. C. Argot (MNHN), A.G. Kramarz, and M. Reguero (MLP) kindly granted access to the collections under their care. We gratefully thank M.G. Vucetich, who kindly provided casts of fossil rodents from Argentinean Patagonia. We warmly thank C. Cazevieille (Montpellier RIO Imaging, Institut des Neurosciences de Montpellier, France) and D. Cot (Institut Européen des Membranes [IEM], Montpellier, France) for access to a scanning electron microscope facility. We thank J. Louys, K. Claeson, and C. Grohé (members of the Palaeontologia Electronica Editorial Board), as well as the anonymous reviewers, who provided formal reviews of this manuscript that enhanced the final version. This work was supported by CNRS “Eclipse 2”, CNRS “Paleo2”, Toulouse University “SPAM” programs, by the L. S. B. Leakey Foundation, the ANR PALASIAFRICA (ANR-08-JCJC-0017, ANR-ERC), the National Geographic Society (grant 9679-15), an “Investissements d’Avenir” grant managed by the “Agence Nationale de la Recherche” (CEBA, ANR-10-LABX-0025-01), and by the COOPINTEER CNRS/CONICET and the ECOS-SUD/FONCyT international collaboration programs. This is ISEM publication 2017-013-Sud.
REFERENCES
Ameghino, F. 1897. Mammifères Crétacés de l’Argentine. Deuxième contribution à la connaissance de la faune mammalogique des couches à Pyrotherium. Boletín del Instituto Geográfico Argentino, 18:406-429, 431-521.
Antoine, P.-O., Abello, M., Adnet, S., Altamirano Sierra, A.J., Baby, P., Billet, G., Boivin, M., Calderóne, Y., Candela, A., Chabain, J., Corfu, F., Croft, D.A., Ganerød, M., Jaramillo, C., Klaus, S., Marivaux, L., Navarrete, R.E., Orliac, M.J., Parra, F., Perez, M.E., Pujos, F., Rage, J.-C., Ravel, A., Robinet, C., Roddaz, M., Tejada-Lara, J.V., Vélez-Juarbe, J., Wesselingh, F.P., and Salas-Gismondi, R. 2016. A 60-million-year Cenozoic history of western Amazonian ecosystems in Contamana, eastern Peru. Gondwana Research, 31:30-59.
Antoine, P.-O., Marivaux, L., Croft, D.A., Billet, G., Ganerød, M., Jaramillo, C., Martin, T., Orliac, M.J., Tejada-Lara, J., Altamirano, A.J., Duranthon, F., Fanjat, G., Rousse, S., and Salas-Gismondi, R.S. 2012. Middle Eocene rodents from Peruvian Amazonia reveal the pattern and timing of caviomorph origins and biogeography. Proceedings of the Royal Society B, 279:1319-1326.
Antoine, P.-O., Salas-Gismondi, R., Pujos, F., Ganerød, M., and Marivaux, L. 2017. Western Amazonia as a hotspot of mammalian biodiversity throughout the Cenozoic. Journal of Mammalian Evolution. 24: 5–17.
Arnal, M. and Vucetich, M.G. 2015. Main radiation events in Pan-Octodontoidea (Rodentia, Caviomorpha). Zoological Journal of the Linnean Society , 175:587-606.
Barbière, F. and Marivaux, L. 2015. Phylogeny and evolutionary history of hystricognathous rodents from the Old World during the Tertiary: new insights into the emergence of modern “phiomorph” families, p. 87-1120. In Cox, P. and Hautier, L. (eds.), Evolution of the Rodents- Advances in Phylogeny, Functional Morphology and Development. Cambridge University Press, Cambridge.
Bennett, E.T. 1833. On the Chinchillidae, a family of herbivorous Rodentia, and on a new genus referrible to it. Transactions of the Zoological Society of London, 1:35-64.
Bertrand, O.C., Flynn, J.J., Croft, D.A., and Wyss, A.R. 2012. Two new taxa (Caviomorpha, Rodentia) from the Early Oligocene Tinguiririca fauna (Chile). American Museum Novitates, 3750:1-36.
Boivin, M., Marivaux, L., Candela, A.M., Orliac, M.J., Pujos, F., Salas-Gismondi, R., Tejadalara, J.V., and Antoine, P-O. 2017. Late Oligocene caviomorph rodents from Contamana, Peruvian Amazonia. Papers in Palaeontology. 3: 69–109.
Bonaparte, C.L.J.L. 1845. Catalogo methodico dei mammiferi Europei. Luigi di Giacomo Pirola, Milano.
Bond, M., Tejedor, M.F., Campbell, K.E., Chornogubsky, L., Novo, N., and Goin, F.J. 2015. Eocene primates of South America and the African origins of New World monkeys. Nature, 520:538-541.
Bowdich, T.E. 1821. An analysis of the natural classifications of Mammalia: for the use of students and travellers. Printed by Smith, J., Paris.
Campbell, K. E. (ed.) 2004. The Paleogene Mammalian Fauna of Santa Rosa, Amazonian Peru. Natural History Museum of Los Angeles County, Science Series 40.
Candela, A.M. and Picasso, M.B.J. 2008. Functional anatomy of the limbs of Erethizontidae (Rodentia, Caviomorpha): indicators of locomotor behavior in Miocene porcupines. Journal of Morphology, 269:552-593.
Croft, D.A., Anaya, F., Auerbach, D., Garzione, C., and Macfadden, B.J. 2009. New data on Miocene neotropical provinciality from Cerdas, Bolivia. Journal of Mammalian Evolution, 16:175-198.
de Bruijn, H., Ünay, E., Saraç, G., and Yïlmaz, A. 2003. A rodent assemblage from the Eo/Oligocene boundary interval near Süngülü, Lesser Caucasus, Turkey. Coloquios de Paleontología, Volumen Extraordinario, 1:47-76.
Fields, R.W. 1957. Hystricomorph rodents from the late Miocene of Colombia, South America, University of California Publications in Geological Sciences , 32:273-404.
Fischer de Waldheim, G. 1817. Adversaria zoologica. Mémoires de la Société Impériale des Naturalistes de Moscou, 5:357-428.
Flynn, L.J., Jacobs, L.L., and Cheema, I.U. 1986. Baluchimyinae, a new Ctenodactyloid rodent subfamily from the Miocene of Baluchistan. American Museum Novitates, 2841:1-58.
Frailey, C.D. and Campbell, K.E. 2004. Paleogene rodents from Amazonian Peru: the Santa Rosa local fauna, p. 71-130. In Campbell, K.E. (ed.), The Paleogene Mammalian Fauna of Santa Rosa, Amazonian Peru. Natural History Museum of Los Angeles County, Science Series 40.
Hartenberger, J.-L. 1975. Nouvelles découvertes de rongeurs dans le Déseadien (Oligocène inférieur) de Salla Luribay (Bolivie). Comptes Rendus des Séances, Académie de Science, Paris, 280:427-430.
Hoffstetter, R. and Lavocat, R. 1970. Découverte dans le Déséadien de Bolivie des genres pentalophodontes appuyant les affinités africaines des rongeurs caviomorphes. Comptes-rendus de l’Académie des Sciences, Série D, 271:172-175
Jaeger, J.-J., Denys, C., and Coiffart, B. 1985. New Phiomorpha and Anomaluridae from the late Eocene of north-west Africa: phylogenetic implications. p. 567-588. In Luckett, W.P. and Hartenberger, J.-L. (eds.), Evolutionary Relationships among Rodents, a Multidisciplaplinary Analysis. Springer US, New York.
Janis, C.M. 1986. An estimation of tooth volume and hypsodonty indices in ungulate mammals, and the correlation of these factors with dietary preference, p. 367-387. In Russell, D.E., Santoro, J.P., and Sigogneau-Russell, D. (eds.), Teeth Revisited. Proceedings of the 7th International Symposium on Dental Morphology (Paris), 53. Mémoires du Muséum national d'Histoire Naturelle, Série C.
Kay, B.R.F. 2015. New World monkey origins. Science, 347:1068-1069.
Kramarz, A.G., Vucetich, M.G., and Arnal, M. 2013. A new early Miocene chinchilloid hystricognath rodent; an approach to the understanding of the early chinchillid dental evolution. Journal of Mammalian Evolution, 20:249-261.
Lavocat, R. 1976. Rongeurs caviomorphes de l’Oligocène de Bolivie. II. Rongeurs du bassin déséadien de Salla. Palaeovertebrata, 7:15-90.
Loomis, F.B. 1914. The Deseado formation of Patagonia. Runiford Press, Concord.
Mares, M.A. and Ojeda, R.A. 1982. Patterns of diversity and adaptation of South American hystricognathous rodents, p. 393-432. In Mares, M.A. and Genoways, H. (eds.), Mammalian Biology in South America. Special Publication, Pymatuning Laboratory of Ecology, University of Pittsburg, Pittsburg, USA.
Marivaux, L., Adnet, S., Altamirano-Sierra, A.J., Boivin, M., Pujos, F., Ramdarshan, A., Salas-Gismondi, R., Tejada-Lara, J.V., and Antoine P.-O. 2016. Neotropics provide insights into the emergence of New World monkeys: new dental evidence from the late Oligocene of Peruvian Amazonia. Journal of Human Evolution , 97:159-175.
Marivaux, L., Adnet, S., Benammi, M., Tabuce, R., and Benammi, M. 2017. Anomaluroid rodents from the earliest Oligocene of Dakhla, Morocco, reveal the long-lived and morphologically conservative pattern of the Anomaluridae and Nonanomaluridae during the Tertiary in Africa. Journal of Systematic Palaeontology . doi:10.1080/14772019.2016.1206977.
Marivaux, L., Essid, E.M., Marzougui, W., Ammar, H.K., Adnet, S., Marandat, B., Merzeraud, G., Tabuce, R., and Vianey-Liaud, M. 2014. A new and primitive species of Protophiomys (Rodentia, Hystricognathi) from the late middle Eocene of Djebel el Kébar, Central Tunisia. Palaeovertebrata, 38:e2.
Marivaux, L., Vianey-Liaud, M., and Jaeger, J-J. 2004. High-level phylogeny of early Tertiary rodents: dental evidence. Zoological Journal of the Linnean Society, 142:105-134.
Marivaux, L., Welcomme, J.L., Vianey-Liaud, M., and Jaeger, J.-J. 2002. The role of Asia in the origin and diversification of hystricognathous rodents. Zoologica Scripta, 31: 225-239.
Osborn, H.F. 1908. New fossil mammals from the Fayûm Oligocene, Egypt. Bulletin of the American Museum of Natural History, 24:265-272.
Patterson, B. and Wood, A.E. 1982. Rodents from the Deseadan Oligocene of Bolivia and the relationships of the Caviomorpha. Bulletin of the Museum of Comparative Zoology, 149:371-543.
Patton, J.L., Pardiñas, U.F., and d’Elía, G. (eds.). 2015. Mammals of South America, Volume 2: Rodents. University of Chicago Press, Chicago and London.
Rinderknecht, A. and Blanco, R.E. 2008. The largest fossil rodent. Proceedings of the Royal Society B, 275:923-928.
Sallam, H.M., Seiffert, E.R., Steiper, M.E., and Simons, E.L. 2009. Fossil and molecular evidence constrain scenarios for the early evolutionary and biogeographic history of hystricognathous rodents. Proceedings of the National Academy of Sciences, 106:16722-16727.
Shockey, B.J., Hitz, R., and Bond, M. 2004. Paleogene notoungulates from the Amazon Basin of Peru, p. 61-70. In Campbell, K.E. (ed.), The Paleogene Mammalian Fauna of Santa Rosa, Amazonian Peru. Natural History Museum of Los Angeles County, Science Series 40.
Shockey, B.J., Salas-Gismondi, R., Gans, P.B., Jeong, A., and Flynn, J.J. 2009. Paleontology and geochronology of the Deseadan (late Oligocene) of Moquegua, Peru. American Museum Novitates, 3668:1-24.
Tullberg, T. 1899. Ueber das system der Nagethiere: eine phylogenetische studie. Nova Acta Regiae Societatis Scientarium Upsaliensis, 18:1-514.
Vucetich, M.G., de Souza Cunha, F.L., and Alvarenga, H.M.F. de 1993. Un roedor Caviomorpha de la Formación Tremembé (Cuenca de Taubaté), Estado de São Paulo, Brasil. Anais da Academia Brasileira de Ciências, 65:247-251.
Vucetich, M.G., Dozo, M.T., Arnal, M., and Pérez, M.E. 2015. New rodents (Mammalia) from the late Oligocene of Cabeza Blanca (Chubut) and the first rodent radiation in Patagonia. Historical Biology, 27:236-257.
Vucetich, M.G. and Kramarz, A. G. 2003. New Miocene rodents of Patagonia (Argentina) and their bearing in the early radiation of the octodontoids (Hystricognathi). Journal ofVertebrate Paleontology, 23:435-444.
Vucetich, M.G., Vieytes, E.C., Perez, M.E., and Carlini, A.A. 2010. The rodents from La Cantera and the early evolution of caviomorphs in South America, p. 189-201. In Madden, R.H., Carlini, A.A., Vucetich, M.G., and Kay, R.F. (eds.), The Paleontology of Gran Barranca, Evolution and Environmental Change through the Middle Cenozoic of Patagonia. Cambridge University Press, Cambridge.
Waterhouse, G.R. 1839. Observations on the Rodentia, with a view to point out groups as indicated by
Wilson, D.E. and Reeder, D.M. (eds.). 2005. Mammal Species of the World: A Taxonomic and Geographic Reference (3rd edition). Johns Hopkins University Press, Baltimore.
Wood, A.E. 1949. A new Oligocene rodent genus from Patagonia. American Museum Novitates, 1435:1-54.
Wood, A.E. 1955. A revised classification of the rodents. Journal of Mammalogy, 36:165-187.
Wood, A.E. and Patterson, B. 1959. The rodents of the Deseadan Oligocene of Patagonia and the beginnings of South American rodent evolution. Bulletin of the Museum of Comparative Zoology, 120:281-428.
Wood, A.E. and Wilson, R.W. 1936. A suggested nomenclature for the cusps of the cheek teeth of rodents. Journal of Paleontology, 10:388-391.
Woods, C.A. 1984. Hystricognath rodents, p. 389-446. In Anderson, S. and Jones, J.K. Jr. (eds.), Orders and Families of Mammals of the World. Wiley, New York.
Wyss, A.R., Flynn, J.J., Norell, M.A., Swisher, C.C. III, Charrier, R., Novacek, M.J., and McKenna, M.C. 1993. South America's earliest rodent and recognition of a new interval of mammalian evolution. Nature, 365:434-437.
zoobank.org/E38E9E2A-6303-4278-888F-0F8FC05C5010
