

 Mammals from the earliest Uintan (middle Eocene) Turtle Bluff Member, Bridger Formation, southwestern Wyoming, USA, Part 3: Marsupialia and a reevaluation of the Bridgerian-Uintan North American Land Mammal Age transition
Mammals from the earliest Uintan (middle Eocene) Turtle Bluff Member, Bridger Formation, southwestern Wyoming, USA, Part 3: Marsupialia and a reevaluation of the Bridgerian-Uintan North American Land Mammal Age transition
Article number: 21.2.25A
https://doi.org/10.26879/804
Copyright Society for Vertebrate Paleontology, July 2018
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 13 July 2017. Acceptance: 7 June 2018
{flike id=2240}
ABSTRACT
This is the third and last of a series of reports that provide detailed descriptions and taxonomic revisions of the fauna from the Turtle Bluff Member (TBM) of the middle Eocene Bridger Formation of southwestern Wyoming. The TBM has been designated as the stratotype section for biochron Ui1a (earliest Uintan) of the Uintan North American Land Mammal age and here we document new faunal elements along with new U-Pb geochronologic and paleomagnetic data for the TBM. Prior to these reports, detailed systematic accounts of the taxa from the TBM were unavailable with the exception of one primate (Hemiacodon engardae). Here we document the occurrence of the following didelphimorphian marsupials from the TBM: Herpetotherium knighti, Herpetotherium marsupium, Peradectes chesteri, and Peradectes californicus. New U-Pb dates of 47.31 ± 0.06 Ma and 46.94 ± 0.14 Ma from the TBM provide precise constraints on the age of the fauna. These dates plus new paleomagnetic data further support the existing evidence that the TBM Fauna and the boundary between the Bridgerian and Uintan North American Land Mammal ages occurs within the lower part of Chron C21n of the Geomagnetic Polarity Time Scale. The only other fauna from North America that can confidently be assigned to biochron Ui1a is the Basal Tertiary Local Fauna from the Devil's Graveyard Formation of Texas. Revisions of the faunal characterizations of biochrons Ui1a (earliest Uintan) and Ui1b (early Uintan) of the Uintan North America Land Mammal age are proposed to further clarify their differences.
Paul C. Murphey. Research Associate, Department of Paleontology, San Diego Museum of Natural History, 1788 El Prado, San Diego, California 92101, USA. pmurphey@sdnhm.org
Thomas S. Kelly. Research Associate, Vertebrate Paleontology Department, Natural History Museum of Los Angeles County, 900 Exposition Blvd., Los Angeles, California 90007, USA. tom@tskelly.gardnerville.nv.us
Kevin R. Chamberlain. Research Professor, Department of Geology and Geophysics, University of Wyoming, 1000 E. University Ave., Laramie, Wyoming 82071, USA and Faculty of Geology and Geography, Tomsk State University, Tomsk 634050, Russia. KChamber@uwyo.edu
Kaori Tsukui. Postdoctoral Associate, Department of Earth Atmospheric and Planetary Sciences, Massachusetts Institute of Technology, 77 Massachusetts Avenue, Cambridge, Massachusetts 02139, USA. ktsukui@mit.edu
William C. Clyde. Professor, Department of Earth Sciences, University of New Hampshire, 56 College Road, Durham, New Hampshire, 03824, USA. will.clyde@unh.edu
Keywords: biostratigraphy; Eocene; Marsupialia; Uintan; magnetostratigraphy; U-Pb dating
Final citation: Murphey, Paul C., Kelly, Thomas S., Chamberlain, Kevin R., Tsukui, Kaori, and Clyde, William C. 2018. Mammals from the earliest Uintan (middle Eocene) Turtle Bluff Member, Bridger Formation, southwestern Wyoming, USA, Part 3: Marsupialia and a reevaluation of the Bridgerian-Uintan North American Land Mammal Age transition. Palaeontologia Electronica 21.2.25A 1-52. https://doi.org/10.26879/804
palaeo-electronica.org/content/2018/2240-tbm-mammals-pmag-geochron
Copyright: July 2018 Society of Vertebrate Paleontology
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0/
INTRODUCTION
 The Turtle Bluff Member (TBM) of the Bridger Formation in the Bridger Basin of southwestern Wyoming, USA, has been designated as the stratotype section for the earliest Uintan biochron Ui1a of the Uintan North American Land Mammal age (Evanoff et al., 1994; Murphey and Evanoff, 2007; Murphey and Dunn, 2009; Gunnell et al., 2009). In the Bridger Basin, the TBM overlies the Blacks Fork and Twin Buttes members of the Bridger Formation, which have been designated as the stratotype sections for the middle Bridgerian biochron Br2 and late Bridgerian biochron Br3 of the Bridgerian North American Land Mammal age, respectively (Gunnell et al., 2009). As such, greater knowledge of the taxa comprising the TBM Fauna is important to better characterize the beginning of the Uintan and further our understanding of the Bridgerian-Uintan faunal transition. The TBM, formerly referred to as the Bridger E, was previously considered only sparsely fossiliferous (Matthew, 1909; West and Hutchinson, 1981). However, owing to many years of field work by one of us (PCM) and crews from the University of Colorado Natural History Museum and San Diego Natural History Museum, numerous fossil mammals have been recovered from seven superposed stratigraphic levels in the TBM (Figure 1). This is the third and last report in a series (Kelly and Murphey, 2016a; Murphey and Kelly, 2017) that provides a comprehensive taxonomic analysis and revision of the mammals from the TBM and includes the marsupials along with a reevaluation of the Bridgerian-Uintan North American Land Mammal age transition based on new paleomagnetic and radioisotopic data.
The Turtle Bluff Member (TBM) of the Bridger Formation in the Bridger Basin of southwestern Wyoming, USA, has been designated as the stratotype section for the earliest Uintan biochron Ui1a of the Uintan North American Land Mammal age (Evanoff et al., 1994; Murphey and Evanoff, 2007; Murphey and Dunn, 2009; Gunnell et al., 2009). In the Bridger Basin, the TBM overlies the Blacks Fork and Twin Buttes members of the Bridger Formation, which have been designated as the stratotype sections for the middle Bridgerian biochron Br2 and late Bridgerian biochron Br3 of the Bridgerian North American Land Mammal age, respectively (Gunnell et al., 2009). As such, greater knowledge of the taxa comprising the TBM Fauna is important to better characterize the beginning of the Uintan and further our understanding of the Bridgerian-Uintan faunal transition. The TBM, formerly referred to as the Bridger E, was previously considered only sparsely fossiliferous (Matthew, 1909; West and Hutchinson, 1981). However, owing to many years of field work by one of us (PCM) and crews from the University of Colorado Natural History Museum and San Diego Natural History Museum, numerous fossil mammals have been recovered from seven superposed stratigraphic levels in the TBM (Figure 1). This is the third and last report in a series (Kelly and Murphey, 2016a; Murphey and Kelly, 2017) that provides a comprehensive taxonomic analysis and revision of the mammals from the TBM and includes the marsupials along with a reevaluation of the Bridgerian-Uintan North American Land Mammal age transition based on new paleomagnetic and radioisotopic data.
METHODS AND MATERIALS
U-Pb Geochronology
Zircon grains were separated from two tuffaceous rock samples (PCM3Aug14-04 and PCM3Aug14-07) collected from the TBM (Bridger E) on Cedar Mountain in Uinta County, Wyoming. Sample PCM3Aug14-04 came from the base of the tuffaceous white sandstone bed, which occurs at about 74-75 m above the base of the TBM (Figure 1). Sample PCM3Aug14-07 came from Locality SDSNH 5844 (Roll the Bones) at 105 m above the base of the TBM on Cedar Mountain (Figure 1). The rock samples were mechanically crushed and zircons were concentrated by a combination of ultrasonic defloculation of slurries and Wilfley shaker table separations. Zircons were further purified by magnetic separation and heavy liquid flotation of lighter minerals.
 Each sample yielded a range of zircon morphologies (Figure 2-Figure 3), including euhedral subpopulations with characteristics typical of volcanic origins and ash-fall deposition, such as acicular grains with elongate tips, axial melt trails and transverse channels (Corfu et al., 2003; Hoskin and Schaltegger, 2003; Machlus et al., 2015). More equant, euhedral grains that did not exhibit these volcanic characteristics were also abundant, and each sample also contained rounded, detrital zircons that were likely entrained during deposition. Zircons from the volcanic/ash-fall and euhedral subpopulations were preferentially chosen for dissolution and U-Pb analysis.
Each sample yielded a range of zircon morphologies (Figure 2-Figure 3), including euhedral subpopulations with characteristics typical of volcanic origins and ash-fall deposition, such as acicular grains with elongate tips, axial melt trails and transverse channels (Corfu et al., 2003; Hoskin and Schaltegger, 2003; Machlus et al., 2015). More equant, euhedral grains that did not exhibit these volcanic characteristics were also abundant, and each sample also contained rounded, detrital zircons that were likely entrained during deposition. Zircons from the volcanic/ash-fall and euhedral subpopulations were preferentially chosen for dissolution and U-Pb analysis.
Selected single grain zircons were annealed at 850 °C for 50 hours, then dissolved in two steps in a chemical abrasion, thermal ionization mass spectrometric U-Pb dating method (CA-TIMS) modified from Mattinson (2005). The first dissolution step was in hydrofluoric acid (HF) and nitric acid (HNO3) at 180 °C for 12 hours. This removed the most metamict zircon domains in the annealed crystals. Individual grains were then spiked with a mixed 205Pb-233U-235U tracer (ET535), completely dissolved in HF and HNO3 at 240 °C for 30 hours, and then converted to chlorides. The dissolutions were loaded onto rhenium filaments with phosphoric acid and silica gel without any further chemical processing. Pb and UO2 isotopic compositions were determined in single Daly-photomultiplier mode on a Micromass Sector 54 mass spectrometer. Data were reduced, and ages calculated using PbMacDat and ISOPLOT/EX after Ludwig (1988, 1991, 1998). Total common Pb varied from 0.5 to 4 picograms and was all assigned to blank. All analyses were conducted at the Radiogenic Isotope Laboratory at the University of Wyoming.
Magnetostratigraphy
 A set of pilot paleomagnetic samples was collected to determine the potential of creating a reliable magnetostratigraphy for the uppermost Bridger Formation and to spot test the polarity of key stratigraphic levels. Two separately oriented blocks were collected from three different stratigraphic levels, including: 1) the tuffaceous white sandstone bed; 2) the level of the Roll the Bones Locality in the stratotype section of the TBM (Bridger E) on the southwest flank of Cedar Mountain; and 3) from a tuff (= the Basal E tuff of Murphey and Evanoff, 2007) in the uppermost Twin Buttes Member (Bridger D) on Sage Creek Mountain. The name (Basal E tuff) that was used by Murphey and Evanoff (2007) to refer to this stratigraphic unit is confusing considering that it does not occur in the TBM (formerly the Bridger E), so it is here renamed the uppermost Twin Buttes Member tuff. PCM6Sept15-02 and PCM6Sept15-03 were collected from the base of a tuffaceous white sandstone, 74 m above base of the TBM, and PCM6Sept15-04 and PCM6Sept15-05 were collected from Locality SDSNH 5844 (Roll the Bones), 105 m above base of the TBM (Figure 1). PCM7Sept15-01 and PCM7Sept15-02 were collected from the uppermost Twin Buttes Member tuff, about 8 m below the base of the TBM. Each oriented block was cut down to form three separate 8 cm 3 samples. Samples were analyzed in the Paleomagnetism Laboratory at the University of New Hampshire with an HSM2 SQUID cryogenic magnetometer, a Molspin tumbling alternating-field demagnetizer, and an ASC Model TD48 SC thermal demagnetizer. Sample demagnetization protocol included: 1) step-wise alternating field demagnetization in 2 mT steps up to 15 mT, then 5 mT steps up to 40 mT, and then 10 mT steps thereafter (to a maximum of 100 mT); and 2) step-wise (up to 18 steps) thermal demagnetization. After demagnetization, samples with Natural Remanent Magnetization (NRM) directions exhibiting a relatively linear decay to the origin were characterized by least squares analysis (Kirschvink, 1980). Samples that exhibited non-linear unstable demagnetization behavior were not considered further. Oriented blocks that produced three samples with well-defined Characteristic Remanent Magnetizations (ChRM) that cluster together and passed the Watson test for randomness (Watson, 1956) were considered reliable and are reported here using standard Fisher statistics (Fisher, 1953).
A set of pilot paleomagnetic samples was collected to determine the potential of creating a reliable magnetostratigraphy for the uppermost Bridger Formation and to spot test the polarity of key stratigraphic levels. Two separately oriented blocks were collected from three different stratigraphic levels, including: 1) the tuffaceous white sandstone bed; 2) the level of the Roll the Bones Locality in the stratotype section of the TBM (Bridger E) on the southwest flank of Cedar Mountain; and 3) from a tuff (= the Basal E tuff of Murphey and Evanoff, 2007) in the uppermost Twin Buttes Member (Bridger D) on Sage Creek Mountain. The name (Basal E tuff) that was used by Murphey and Evanoff (2007) to refer to this stratigraphic unit is confusing considering that it does not occur in the TBM (formerly the Bridger E), so it is here renamed the uppermost Twin Buttes Member tuff. PCM6Sept15-02 and PCM6Sept15-03 were collected from the base of a tuffaceous white sandstone, 74 m above base of the TBM, and PCM6Sept15-04 and PCM6Sept15-05 were collected from Locality SDSNH 5844 (Roll the Bones), 105 m above base of the TBM (Figure 1). PCM7Sept15-01 and PCM7Sept15-02 were collected from the uppermost Twin Buttes Member tuff, about 8 m below the base of the TBM. Each oriented block was cut down to form three separate 8 cm 3 samples. Samples were analyzed in the Paleomagnetism Laboratory at the University of New Hampshire with an HSM2 SQUID cryogenic magnetometer, a Molspin tumbling alternating-field demagnetizer, and an ASC Model TD48 SC thermal demagnetizer. Sample demagnetization protocol included: 1) step-wise alternating field demagnetization in 2 mT steps up to 15 mT, then 5 mT steps up to 40 mT, and then 10 mT steps thereafter (to a maximum of 100 mT); and 2) step-wise (up to 18 steps) thermal demagnetization. After demagnetization, samples with Natural Remanent Magnetization (NRM) directions exhibiting a relatively linear decay to the origin were characterized by least squares analysis (Kirschvink, 1980). Samples that exhibited non-linear unstable demagnetization behavior were not considered further. Oriented blocks that produced three samples with well-defined Characteristic Remanent Magnetizations (ChRM) that cluster together and passed the Watson test for randomness (Watson, 1956) were considered reliable and are reported here using standard Fisher statistics (Fisher, 1953).
Paleontology
Teeth were measured with an optical micrometer to the nearest 0.01 mm. Upper molars were measured following the methods used by Rothecker and Storer (1996), Eberle and Storer (1995), and Kihm and Schumaker (2015), which results in slightly shorter anteroposterior lengths and slightly wider transverse widths than the method used by Korth (1994). Dental terminology follows Marshall et al. (1990). Upper and lower teeth are designated by uppercase and lowercase letters, respectively. All specimens described here are curated in the research collections of three different institutions: the Department of Paleontology at the San Diego Natural History Museum, the Paleontology Section of the University of Colorado Museum of Natural History, and the Department of Earth Sciences, Denver Museum of Nature and Science. Detailed locality data are available at these institutions.
Subzones or subbiozones of the Bridgerian and Uintan North American Land Mammal ages (e.g., Br2, Br3, Ui1a, Uilb, Ui2, and Ui3) follow Gunnell et al. (2009).
Abbreviations
ap, greatest anteroposterior length; L, left; GPTS, Geomagnetic Polarity Time Scale; kyr, one thousand years in the radioisotopic time scale; Ma, megannum (one million years in the radioisotopic time scale); R, right; tr, greatest transverse width; tra, anterior transverse width; trp, posterior transverse width.
Institutional Acronyms
AMNH, American Museum of Natural History; DMNH, Denver Museum of Nature and Science; SDNHM, San Diego Natural History Museum; LACM (CIT), California Institute of Technology, specimens now housed at the Natural History Museum of Los Angeles County; SDSNH, San Diego Society of Natural History; TMM, Texas Memorial Museum; UCM, University of Colorado Museum of Natural History; UCMP V-, University of California, Museum of Paleontology, Berkeley, vertebrate fossil locality; USNM, National Museum of Natural History, Smithsonian Institution; YPM, Yale Peabody Museum of Natural History.
U-Pb ZIRCON GEOCHRONOLOGY
Results
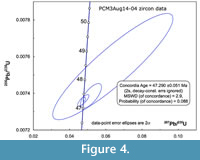 Four single-grain analyses from PCM3Aug14-04 produced concordant data that overlap within error (Figure 4, Table 1). Weighted mean analytical 206Pb/238U date is 47.313 ± 0.059 Ma (Figure 5, 95% confidence, mean square weighted deviation [MSWD] 0.97) with correction for 230Th disequilibrium (after Schärer 1984) assuming a magma Th/U of 2.2.
Four single-grain analyses from PCM3Aug14-04 produced concordant data that overlap within error (Figure 4, Table 1). Weighted mean analytical 206Pb/238U date is 47.313 ± 0.059 Ma (Figure 5, 95% confidence, mean square weighted deviation [MSWD] 0.97) with correction for 230Th disequilibrium (after Schärer 1984) assuming a magma Th/U of 2.2.  The 230Th correction increased the date by about 60 kyr. The Concordia Age (Ludwig 1998) from these data, which incorporates both the 206Pb/238U and 207Pb/235U data, is 47.290 ± 0.051 Ma (2 sigma, MSWD 2.9, Figure 4) without propagating uncertainties in the two U decay constants. These dates can be compared internally to other U-Pb dates to evaluate age differences, as the decay constant errors will bias the U-Pb dates systematically. When tracer errors and decay constant errors are included following Schoene et al. (2006), the weighted mean 206Pb/238U date and Concordia Age for this sample become 47.313 ± 0.081 Ma and 47.290 ± 0.075 Ma, respectively. These dates include all external sources of U-Pb errors and can be directly compared to 40Ar/39Ar dates as long as the 40Ar/39Ar dates have similarly included all external errors and have been recalculated to reflect the new age for the Fish Canyon 40Ar/39Ar standard (e.g., Kuiper et al., 2008; Smith et al., 2010). All older 40Ar/39Ar dates reported here have been recalculated relative to the astronomically calibrated age of 28.201 Ma for the Fish Canyon sanidine standard (Kuiper et al., 2008) and include ± 2σ fully propagated uncertainty.
The 230Th correction increased the date by about 60 kyr. The Concordia Age (Ludwig 1998) from these data, which incorporates both the 206Pb/238U and 207Pb/235U data, is 47.290 ± 0.051 Ma (2 sigma, MSWD 2.9, Figure 4) without propagating uncertainties in the two U decay constants. These dates can be compared internally to other U-Pb dates to evaluate age differences, as the decay constant errors will bias the U-Pb dates systematically. When tracer errors and decay constant errors are included following Schoene et al. (2006), the weighted mean 206Pb/238U date and Concordia Age for this sample become 47.313 ± 0.081 Ma and 47.290 ± 0.075 Ma, respectively. These dates include all external sources of U-Pb errors and can be directly compared to 40Ar/39Ar dates as long as the 40Ar/39Ar dates have similarly included all external errors and have been recalculated to reflect the new age for the Fish Canyon 40Ar/39Ar standard (e.g., Kuiper et al., 2008; Smith et al., 2010). All older 40Ar/39Ar dates reported here have been recalculated relative to the astronomically calibrated age of 28.201 Ma for the Fish Canyon sanidine standard (Kuiper et al., 2008) and include ± 2σ fully propagated uncertainty.
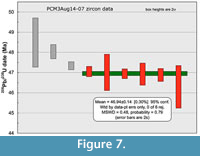
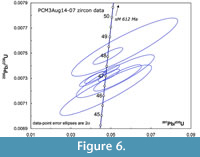 Ten single grain analyses from PCM3Aug14-07 yielded nine concordant analyses with 206Pb/238U dates that range from 48.5 to 46.3 Ma (Figure 6, Table 1) and a single concordant analysis at 612 ± 1.3 Ma (sample 07 ash s (single grain) M in Table 1). The range of ca. 47 Ma dates is interpreted to reflect minor inherited zircon components, minor differences in magma chamber residence times and possibly some detrital zircon gains, although less than half of the recovered zircons were demonstrably rounded, detrital grains (Figure 3), and these grains were avoided during dating. The weighted mean 206Pb/238U date from the six youngest and overlapping analyses is 46.94 ± 0.14 Ma (Figure 7, 95% confidence, MSWD 0.48, corrected for 230Th disequilibrium) and is interpreted as the best estimate of the eruption age of this ash. The date with fully propagated uncertainties, including those from tracer and decay constant is 46.94 ± 0.15 Ma.
Ten single grain analyses from PCM3Aug14-07 yielded nine concordant analyses with 206Pb/238U dates that range from 48.5 to 46.3 Ma (Figure 6, Table 1) and a single concordant analysis at 612 ± 1.3 Ma (sample 07 ash s (single grain) M in Table 1). The range of ca. 47 Ma dates is interpreted to reflect minor inherited zircon components, minor differences in magma chamber residence times and possibly some detrital zircon gains, although less than half of the recovered zircons were demonstrably rounded, detrital grains (Figure 3), and these grains were avoided during dating. The weighted mean 206Pb/238U date from the six youngest and overlapping analyses is 46.94 ± 0.14 Ma (Figure 7, 95% confidence, MSWD 0.48, corrected for 230Th disequilibrium) and is interpreted as the best estimate of the eruption age of this ash. The date with fully propagated uncertainties, including those from tracer and decay constant is 46.94 ± 0.15 Ma.
 Sample PCM3Aug14-04 is 30 m stratigraphically below sample PCM3Aug14-07 and the internal 206Pb/238U dates do not overlap (Figure 8). The differences in these ages require a minimum of 174 kyr between deposition of these two strata and a maximum of 572 kyr.
Sample PCM3Aug14-04 is 30 m stratigraphically below sample PCM3Aug14-07 and the internal 206Pb/238U dates do not overlap (Figure 8). The differences in these ages require a minimum of 174 kyr between deposition of these two strata and a maximum of 572 kyr.
Remarks
The new radioisotopic data provide important constraints for the age of the TBM as well as its Ui1a fauna. The new U-Pb dates are generally consistent with the U-Pb and 40Ar/39Ar ages of ash beds from the Bridger Formation in Smith et al. (2008, 2010) and Tsukui (2016). Of the two dated samples in this study, the date of sample PCM3Aug14-04 from the tuffaceous white sandstone bed is in close agreement with the U-Pb date of the Sage Creek Mountain tuff by Tsukui (2016), and the two ages overlap within analytical error. These two U-Pb dates are also in agreement with the 40Ar/39Ar date of Smith et 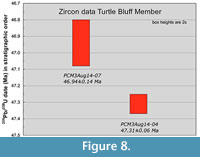 al. (2008, 2010) for the Sage Creek Mountain tuff when the astronomically calibrated age of the Fish Canyon sanidine is used, although the 40Ar/39Ar date is comparatively older. Based on the age as well as the description of the bed from which the sample was collected, it is likely that the Sage Creek Mountain tuff of Smith et al. (2008, 2010) and Tsukui (2016) is the same bed as the bed from which PCM3Aug14-04 was collected. The best age estimate for the Ui1a fauna in the TBM is thus provided by the radioisotopic (U-Pb and 40Ar/39Ar) dates from the level of Locality SDSNH 5844 (Roll the Bones) as well as the stratigraphically lower tuffaceous white sandstone bed and the apparently laterally equivalent Sage Creek Mountain tuff.
al. (2008, 2010) for the Sage Creek Mountain tuff when the astronomically calibrated age of the Fish Canyon sanidine is used, although the 40Ar/39Ar date is comparatively older. Based on the age as well as the description of the bed from which the sample was collected, it is likely that the Sage Creek Mountain tuff of Smith et al. (2008, 2010) and Tsukui (2016) is the same bed as the bed from which PCM3Aug14-04 was collected. The best age estimate for the Ui1a fauna in the TBM is thus provided by the radioisotopic (U-Pb and 40Ar/39Ar) dates from the level of Locality SDSNH 5844 (Roll the Bones) as well as the stratigraphically lower tuffaceous white sandstone bed and the apparently laterally equivalent Sage Creek Mountain tuff.
The Bridger Formation is now calibrated in absolute time by 40Ar/39Ar and U-Pb dates of four ash beds including the Church Butte tuff that forms the contact between the lower and middle B, the Henrys Fork tuff that forms the contact between the middle C and upper C, as well as the tuffaceous white sandstone bed and Roll the Bones level in the TBM (Smith et al., 2008, 2010; Tsukui, 2016; this paper). Taken together, deposition of the Bridger Formation at minimum spanned from ~48.9 Ma to ~46.9 Ma, and a simple calculation indicates that sedimentation rate between the Church Butte tuff and the tuffaceous white sandstone bed (0.314 mm/yr) took place at a rate ~4 times faster than the succeeding interval between the tuffaceous sandstone bed and the Roll the Bones Level (0.081 mm/yr).
PALEOMAGNETIC ANALYSIS
 Results
Results
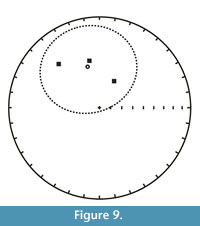
Paleomagnetic samples collected from three sites at two stratigraphic levels produced reliable, well-clustered paleomagnetic directions (samples PCM6Sept15-02, PCM6Sept15-03, PCM7Sept15-01). These samples were characterized by ChRM directions that (a) were defined by a line fit through more than 10 contiguous demagnetization points and a MAD (Maximum Angular Deviation) of less than 20° (Appendix 1) and (b) significantly clustered together based on the Watson test for randomness (Watson, 1956). The mean site directions indicate normal polarity for these stratigraphic levels (Table 2, Figure 9). Paleomagnetic samples PCM6Sept15-04 and PCM6Sept15-05 from Locality SDSNH 5844 (Roll the Bones) were very unstable and did not yield reliable results.
Remarks
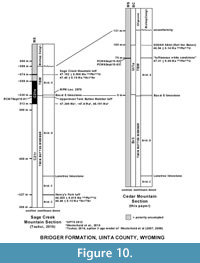 By correlation to the GPTS (Gradstein et al., 2012), the U-Pb age of 47.313 ± 0.059 Ma from the same level (tuffaceous white sandstone bed) as the paleomagnetic samples, PCM6Sept15-02 and PCM6Sept15-03, indicates that these samples with normal polarity determination are likely to correlate to Chron C21n (Figure 10). The magnetostratigraphy of Tsukui (2016) from the Bridger Formation does not extend up to the Sage Creek Mountain tuff. However, the top of her magnetostratigraphic section, which is ~30 m below the Sage Creek Mountain tuff, was assigned to Chron C21n. Thus, correlating the two samples PCM6Sept15-02 and PCM6Sept15-03 to Chron C21n is consistent with the magnetostratigraphy of Tsukui (2016).
By correlation to the GPTS (Gradstein et al., 2012), the U-Pb age of 47.313 ± 0.059 Ma from the same level (tuffaceous white sandstone bed) as the paleomagnetic samples, PCM6Sept15-02 and PCM6Sept15-03, indicates that these samples with normal polarity determination are likely to correlate to Chron C21n (Figure 10). The magnetostratigraphy of Tsukui (2016) from the Bridger Formation does not extend up to the Sage Creek Mountain tuff. However, the top of her magnetostratigraphic section, which is ~30 m below the Sage Creek Mountain tuff, was assigned to Chron C21n. Thus, correlating the two samples PCM6Sept15-02 and PCM6Sept15-03 to Chron C21n is consistent with the magnetostratigraphy of Tsukui (2016).
The stratigraphic level of sample PCM7Sept15-01 from the uppermost Twin Buttes Member tuff was magnetostratigraphically correlated to Chron C21n by Tsukui (2106), and thus, it is consistent with our determination that the sample has normal polarity, and PCM7Sept15-01 is likely to also correlate to Chron C21n. The new U-Pb and paleomagnetic polarity data provide further support for the magnetostratigraphic chron assignment of the Bridgerian-Uintan boundary to Chron C21n (Tsukui, 2016).
SYSTEMATIC PALEONTOLOGY
Cohort MARSUPIALIA Illiger, 1811
Order DIDELPHIMORPHIA Gill, 1872
Family HERPETOTHERIIDAE Trouessart, 1879
Genus HERPETOTHERIUM Cope, 1873a
Type species. Herpetotherium fuzax Cope, 1873a by original designation, emended to Herpetotherium fugax Cope 1873c.
Other included species. H. comstocki (Cope, 1884); H. valens (Lambe, 1908); H. merriami (Stock and Furlong, 1922); H. marsupium Troxell, 1923a; H. youngi (McGrew, 1937); H. edwardi (Gazin, 1952); and H. knighti (McGrew, 1959).
Herpetotherium knighti (McGrew, 1959) in McGrew et al. (1959)
Figure 11.1-9, Figure 12.1-10, Table 3
1959 Peratherium knighti; McGrew, in McGrew et al., p. 147, figure 3.
 1962 Peratherium morrisi; Gazin, p. 21, pl. 1, figure 1.
1962 Peratherium morrisi; Gazin, p. 21, pl. 1, figure 1.
1970 Peratherium sp.; McGrew and Sullivan, p.74.
1973 Peratherium knighti; West and Dawson, p. 35.
1975 Peratherium cf. P. knighti; Setoguchi, p. 267, figures 3-6.
1976 Peratherium knighti; Gazin, p. 2, 7.
1976 Peratherium sp., cf. P. knighti, in part; Lillegraven, p. 86, pl. 1, figures 1a-c, pl. 2, figures 1a-c, 2a-c, 3a-c, pl. 3, figures 1a-c, 2a-c, 3a-c, pl. 4, figures 1a-c, 2a-c, 3a-c, 4a-c, pl. 5, figures 1a-c.
1982 Peratherium knighti; Bown, p. A43.
1983b Peratherium knighti; Krishtalka and Stucky, p. 235, figure 2.
1984 Peratherium knighti; Storer, p. 17.
1985 Peratherium sp., cf. P. knighti; Eaton, p.347, figure 3a.
1996 Herpetotherium sp., cf. H. knighti; Rotheker and Storer, p. 772.
1996 Herpetotherium knighti; Storer, p. 245, 247.
1996 Herpetotherium knighti; Stucky et al., p. 44.
1998 Peratherium knighti; Gunnell, p. 88.
2008 Herpetotherium knighti; Korth, p. 42.
Referred specimens. From locality SDSNH 5841: Lm2 or 3, SDSNH 110339. From locality DMNH 4672: LM1, DMNH 75286; Ldp3, DMNH 75324. From locality SDSNH 5844: LM1, SDSNH 110429; partial LM1 or 2, SDSNH 110430; partial RM1 or 2, SDSNH 110431; partial Rm2 or 3, SDSNH 110427. From locality UCM 92189: LM1, UCM 68793; RM2s, UCM 70636, 70639, 95783; partial RM2 or 3, UCM 68622; RM3s, UCM 68794, 70770; partial RM3, UCM 69973; RM4, UCM 95784; partial dentary with Lp2, UCM 95797; Rdp3, UCM 68902; Lp3, UCM 68908; Rp3, 68937; Rm1, UCM 95792; Lm2 or 3s, UCM 70977, 95790; Rm2 or 3s, UCM 78454, 95785; partial Rm2 or 3, UCM 68901; partial dentary with Rm2-3, UCM 68574; partial dentary with Lm3-4, UCM 95780; Rm4s, UCM 67896, 95793.
Description. In the TBM sample, all the upper molars are isolated teeth. In Paleogene and early Neogene herpetotheriids, the M1 can usually be distinguished from M2 by having a larger ap/tr ratio, that is the ap dimension is usually greater than the tr dimension, whereas in M2 the ap/tr ratio is smaller (more transverse) with the ap dimension usually smaller than the tr dimension (Korth, 1994; Kihm and Schumaker, 2015). The M3 is more easily distinguished from the first two upper molars by having a more V-shaped occlusal outline and its ap/tr ratio is usually smaller than that of M2 (Korth, 1994; Kihm and Schumaker, 2015). Thus, the ap/tr ratio progressively decreases from M1 to M3. The above criteria were used in this study to identify the tooth positions of the first three upper molars, but it should be noted that without intact dentitions our identifications for M1 and M2 are provisional. In the TBM sample, two M1s (Figure 11.1-11.2) are identified with ap/tr ratios of 1.03 and 1.11, whereas the four M2s (Figure 11.3-11.6) are identified with a mean ap/tr ratio of 0.92 (range = 0.88-0.96).
Besides the M2s being more transverse relative to the M1s, their occlusal morphologies are very similar. They have a dilambdodont centrocrista and a very shallow ectoflexus. The paracone is smaller and lower in height than the metacone. The protocone is robust and anteriorly positioned. The preprotocrista and postprotocrista extend labially from the protocone to terminate at a well-developed protoconule and metaconule, respectively. All stylar cusps are positioned along the labial edge of the stylar shelf. Stylar cusp A is absent in five specimens, only represented by an anterolabial spur that is connected to the labial terminus of the anterior cingulum, and on one specimen it is vestigial (a very small, low cusp on the spur). Stylar cusp B is robust, connected lingually to the preparacrista and positioned opposite of the paracone apex. Stylar cusp C is moderately developed and positioned just posterior of the ectoflexus. Stylar cusp D is moderately developed, slightly smaller than C in five specimens and about equal in size to C in one specimen and positioned very close to C in four specimens and joined to C in two specimens. A stylar cusp E is lacking on all specimens.
The occlusal morphology of the M3 (Figure 11.7-11.8) is quite similar to that of the M1-2 (Figure 11.1-11.6), but minor differences can be discerned. As noted above, the M3 has a more V-shaped occlusal outline and is slightly more transversely broad than M1-2, which is due to a relatively shorter metastylar wing on the stylar shelf. The ectoflexus of M3 is slightly deeper than that of M1-2, but still quite shallow. The stylar cusp C is positioned slightly more anteriorly, close to the center of the ectoflexus. Stylar cusps C and D are slightly better developed and slightly more separated.
The M4 (Figure 11.9) has a very transversely elongated occlusal outline. The paracone is about equal in size to the metacone. The protocone is robust with its apex nearly in line with that of the paracone. The paraconule and metaconule are small and poorly developed. A metastylar shelf is lacking posteriorly, whereas the stylar shelf extends anterolabially from near the labial base of the metacone to join the anterolabial terminus of the elongated preprotocrista. The anterior cingulum is distinct, extending from the protoconule to the anterolabial projection of the stylar shelf. A single stylar cusp is present that is positioned just slightly posterior of the paracone apex as a flattened, oval bulge on labial edge of the stylar shelf, and probably represents cusp C. The ectoflexus is nearly straight.
The p2 and p3 have a simple morphology, typical of those of Herpetotherium (Lillegraven, 1976; Korth, 1994), with a relatively tall protoconid and weakly-developed anterior and posterior cuspulids that are nearly in line with the protoconid apex. The p2 differs from the p3 by being smaller, narrower, and more elongated anteroposteriorly with the protoconid apex positioned more anteriorly, resulting in a longer, more steeply inclined central ridge from the protoconid apex to the posterior cuspulid and by having a slightly better developed posterior cuspulid.
 Two lower teeth agree well in proportions and overall structure to those previously identified as a deciduous p3 of Herpetotherium (Lillegraven, 1976; Rothecker and Storer, 1996; Kihm and Schumaker, 2015). They exhibit an occlusal morphology that is similar to that of the lower molars, including a well-developed entoconid that is taller, larger, and separated from the shelf-like, posteriorly projecting hypoconulid by a distinct notch (Figure 12.1). It differs from the lower molars by the following: 1) smaller size; 2) a lower, relatively smaller and slightly more anteriorly projecting paraconid, resulting in the trigonid being longer than wide; 3) the protoconid larger than the metaconid with their apices positioned relatively closer to each other; and 4) the trigonid and talonid narrower relative to the length.
Two lower teeth agree well in proportions and overall structure to those previously identified as a deciduous p3 of Herpetotherium (Lillegraven, 1976; Rothecker and Storer, 1996; Kihm and Schumaker, 2015). They exhibit an occlusal morphology that is similar to that of the lower molars, including a well-developed entoconid that is taller, larger, and separated from the shelf-like, posteriorly projecting hypoconulid by a distinct notch (Figure 12.1). It differs from the lower molars by the following: 1) smaller size; 2) a lower, relatively smaller and slightly more anteriorly projecting paraconid, resulting in the trigonid being longer than wide; 3) the protoconid larger than the metaconid with their apices positioned relatively closer to each other; and 4) the trigonid and talonid narrower relative to the length.
One tooth is identified as an m1 (Figure 12.3). It differs from m2-3 (Figure 12.2, 12.4-12.8) by being slightly smaller in size and by having the paraconid projecting slightly more anteriorly and positioned slightly more labially. Otherwise, the m1-3 are very similar in occlusal morphology. The protoconid is the tallest primary cusp and is about equal in size to the metaconid. The paraconid is anteroposteriorly compressed, projects anterolabially, and is lower in height than the protoconid and metaconid. The width of the talonid is usually equal to or slightly wider than the trigonid. The hypoconid is the largest and tallest talonid cusp. The entoconid is robust, conical, and taller than the hypoconulid, and separated from it by a distinct notch. The hypoconulid is shelf-like, projecting well posterior of the entoconid, and is connected to the hypoconid by a distinct, relatively tall postcristid. The cristid obliqua extends anterolabially from the hypoconid apex to terminate on the posterior wall of the talonid, about halfway between the protolophid notch and protoconid apex. The anterior and posterior cingulids are moderately robust.
The m4 (Figure 11.9-11.10) only differs from the m1-3 by having a considerably narrower talonid that results in it appearing slightly more anteroposteriorly elongated and a slightly more labially positioned hypoconulid.
Remarks. The familial and subfamilial status of Herpetotherium and other closely related North American and Eurasian genera vary among investigators, with some allocating them to the subfamily Herpetotheriinae Trouessart, 1879, within the Didelphidae Gray, 1921 (e.g., Korth, 1994, 2008; Hayes, 2005), whereas others have elevated the subfamily to familial rank as Herpetotheriidae (e.g., Case et al., 2005; Kelly, 2010; Ladevèze et al., 2012; Williamson et al., 2012). Regardless of which rank is preferred, most investigators agree on their generic composition. Here, we follow Willamson et al. (2012) and allocate Herpetotherium to Herpetotheriidae.
The Paleogene and early Neogene herpetotheriid marsupials of North America have a rather complicated taxonomic history. Krishtalka and Stucky (1983b) and Korth (1994, 2008) documented detailed historic accounts of their taxonomy, from which a brief updated summary is provided here.
Cope (1873a) described a new genus and species, Herpetotherium fuzax, from what is now known as the late Eocene and early Oligocene White River Formation of Colorado, which he regarded as an insectivore. Shortly afterwards, Cope (1873c) corrected the specific name for the genotype to Herpetotherium fugax and described five additional species of Herpetotherium, which he also regarded as insectivores. Eleven years later, Cope (1884) recognized that Herpetotherium fugax actually represented a marsupial and transferred all six of his species to the European genus Peratherium Aymard, 1850. Most subsequent investigators followed this generic allocation and referred many Paleogene and early Neogene North America marsupials to Peratherium (e.g., Matthew, 1903; Stock and Furlong, 1922; McGrew, 1937; Stock, 1936; Gazin, 1952; Galbreath, 1953; Guthrie, 1971; West and Dawson, 1974; Setoguchi, 1974; Lillegraven, 1976; Krishtalka and Stucky, 1983a, 1983b, 1984; Russell, 1984; Storer, 1984). However, of Cope's (1873a, 1873c) original six species, one (Herpetotherium marginale) was later transferred to the geolabidid eulipotyphlan genus Centetodon Marsh, 1872a (McKenna, 1960; Lillegraven et al., 1981), and two (Herpetotherium hunti and Herpetotherium stevesonii) were transferred to other marsupial genera (Crochet, 1980; Scott, 1941; Krishtalka and Stucky, 1983a, 1983b; Korth, 1994).
Lavocat (1951) and Hough (1961) first proposed that the type species, Herpetotherium fugax, should be regarded as generically distinct from Peratherium based on the incorrect assumption that an inflected mandibular angle was lacking in the former, but continued to include most of the remaining North American species in Peratherium. Crochet (1977) went further with the resurrection of Herpetotherium, referring many of the North American species to the genus based on certain differences in the cheek teeth, but his proposal was not widely accepted. Stock (1936) described Peratherium californicum from the Uintan of California and Gazin (1952) described Peratherium chesteri from the late Wasatchian of Wyoming. Setoguchi (1973) proposed that Peradectes protinnominatus Simpson, 1928, is a junior synonym of Peratherium chesteri, which he considered as probably assignable to Peradectes within a peradectid lineage that included Peradectes Matthew and Granger, 1921, and Nanodelphys McGrew, 1937. Setoguchi (1973) also suggested that Nanodelphys may be a junior synonym of Peradectes, whereas Lillegraven (1976) retained Nanodelphys as a valid genus and transferred Peradectes californicum to Nanodelphys as N. californicus. Bown (1979) followed Setoguchi's (1973) proposal and formally synonymized Peratherium protinnominatus with Peratherium chesteri, and assigned Peratherium chesteri to Peradectes. Crochet (1978, 1979, 1980) provided a different taxonomic scenario as follows: 1) erected the subgenus Peradectes for the type species, Peradectes elegans Matthew and Granger, 1921; 2) reduced Nanodelphys to a subgenus of Peradectes, which resulted in Peradectes californicus along with Nanodelphys minutus McGrew, 1937, and Peradectes protinnominatus being assigned to Peradectes (Nanodelphys) and 3) transferred Peradectes chesteri to Herpetotherium (a mistaken assignment that Krishtalka and Stucky [1983b] later corrected). Subsequently, Krishtalka and Stucky (1983b) demonstrated that Peradectes protinnominatus is specifically distinct from Peradectes chesteri and, following Setoguchi (1973), regarded Nanodelphys as a synonym of Peradectes. However, Korth (1994) provided convincing evidence that Nanodelphys minutus is a junior synonym of Herpetotherium hunti Cope, 1873c, then recombined the nomenclature for the genotype to Nanodelphys hunti and demonstrated that it is generically distinct from Peradectes, making the genus monotypic.
Fox (1983) recognized certain differences in the antemolar dentition between Herpetotherium fugax and European species of Peratherium, supporting Crochet's (1977, 1980) original proposal. Korth (1994) provided further evidence that most Chadronian through Hemingfordian herpetotheriid species previously referred to Peratherium should be allocated to Herpetotherium, but retained all Duchesnean and older species in Peratherium. Based on size and certain dental differences, Korth (1994) also erected a new genus, Copedelphys, wherein he transferred two Chadronian species previously assigned to Peratherium (Peratherium titanelix Matthew, 1903, and Peratherium stevensonii Cope, 1873c) to his new genus. Rothecker and Storer (1996) concluded that Peratherium innominatum Simpson, 1928, actually represents a primitive species of Copedelphys and reassigned it to C. innominata. Rothecker and Storer (1996) also recognized additional differences between the cheek teeth of Herpetotherium and Peratherium, and referred almost all Wasatchian through Hemingfordian North American herpetotheriids to Herpetotherium, which was also followed by McKenna and Bell (1997) and Korth (2008). However, Beard and Dawson (2001) referred material from the early Eocene (Wasatchian) Tuscahoma Formation of Mississippi to the European species, Peratherium constans. Thus, except for Peradectes constans, Peradectes californicus, Peradectes chesteri, Nanodelphys hunti, the three species now assigned Copedelphys (C. titanelix, C. stevensonii, C. innominata) and the one species transferred to Centetodon (C. marginale), the current consensus is that all the other Paleogene and early Neogene North American herpetotheriids originally assigned or transferred to Peratherium are now allocated to Herpetotherium. Two additional herpetotheriid genera have also been recognized from the Paleogene of North America, Swaindelphys Johanson, 1996, from the middle Paleocene of Montana and Estelestes Novacek et al., 1991, from the middle Eocene of Baja California, Mexico. Horovitz et al. (2009) referred two partial skulls to Herpetotherium sp., cf. H. fugax that exhibit procumbent lower incisors and a large stylar cusp C on M3, characters typical of the genus (Fox, 1983; Korth, 1994).
Copedelphys innominata, Peradectes chesteri, and two species of Herpetotherium, H. knighti (McGrew, 1959), and H. marsupium (Troxell, 1923a), are currently recognized from the Bridger Formation (Gazin, 1976; West and Hutchinson, 1981; Krishtalka and Stucky, 1983b; Gunnell, 1998; Rothecker and Storer, 1996; Korth, 2008). The TBM specimens can be confidently assigned to Herpetotherium because they exhibit the following diagnostic characters (Setoguchi, 1975; Krishtalka and Stucky, 1983a, 1983b; Rothecker and Storer, 1996; Korth, 2008): 1) a dilambdodont centrocrista on M1-2; 2) a reduced paracone on M1-2 that is smaller and lower in height than the metacone; 3) a large, conical, relatively tall entoconid on m1-3; and 4) a posteriorly projecting hypoconulid on m1-3 that is shelf-like, smaller, and notably lower in height than the entoconid and separated from it by a distinct notch.
As with other Paleogene and early Neogene marsupials, species of Herpetotherium are differentiated primarily by size, dental proportions, and differences in the occurrence and positions of the upper molar stylar cusps (Korth, 1994, 2008). Although many early studies dealt with very small samples, more recent investigations utilizing larger sample sizes for North American species of Herpetotherium have demonstrated that a fair degree of individual variation in size and in the occurrence and positions of the upper molar stylar cusps can be discerned (e.g., Eberle and Storer, 1995; Rothecker and Storer, 1996; Hayes, 2005; Kihm and Schumaker, 2015; Korth, 2015). Ladevèze et al. (2005) reported similar examples of individual variation in the herpetotheriid marsupials of Europe. These studies indicate that the frequency and morphological trends for these character states should be utilized to differentiate species rather than definitive diagnostic statements regarding these character states.
Based on a partial dentary with p3-m2 (YPM 13518) from the Bridger Formation of the Bridger Basin, Troxell (1923a) described Herpetotherium marsupium and differentiated it from Entomacodon minutus Marsh, 1872a (= Herpetotherium knighti see McKenna, 1960 and Robinson, 1968; see also Krishtalka and Stucky, 1983b for nomenclatural priority) by having a presumably more diagonally transverse trigonid wall on the lower molars. The holotype of Herpetotherium knighti, a partial maxilla with M1-3 (AMNH 55684), came from the Bridger Formation at Tabernacle Butte and was initially characterized by McGrew (1959) by having the following: 1) length of M1-3 equals 5.9 mm; 2) three principal stylar cusps on M1-3 that are low and positioned on extreme labial border of the stylar shelf; and 3) a well-developed paraconule and metaconule on M2-3. Lillegraven (1976) assigned specimens from the Bridger Formation and Teepee Trail Formation of Wyoming to Herpetotherium knighti and specimens from the Uintan Friars and Mission Valley formations of California to Herpetotherium sp., cf. H. knighti. However, Krishtalka and Stucky (1983b) considered Lillegraven's sample from California to represent a mixed sample of the larger Herpetotherium knighti and the smaller Copedelphys. innominata. Subsequently, samples ranging from the Wasatchian through the Duchesnean have been referred to Herpetotherium knighti and Herpetotherium marsupium, with H. knighti being distinguished from the latter by having the following (Setoguchi, 1975; West and Dawson, 1975; Bown, 1982; Krishtalka and Stucky, 1983a, 1983b, 1984; Russell, 1984; Storer, 1984; Rothecker and Storer, 1996): 1) smaller size; 2) significantly shallower M2-3 ectoflexi, especially on M3; 3) M1-3 stylar cusps positioned along extreme labial border of the stylar shelf; 4) stylar shelf usually lacking crenulation along its labial border; 5) less tendency toward reduction of cusp C on M1-3 and less separation of cusps C and D on M1-2; 6) lack of a tendency towards twinning of stylar cusp C on M1-3; and 7) m1-3 cristid obliqua with tendency to terminate slightly more labially on the posterior wall of trigonid. The TBM specimens are intermediate in size between typotypic samples of Herpetotherium knighti and Herpetotherium marsupium, but within the observed ranges of the larger specimens of Lillegraven's (1976) sample of Herpetotherium sp., cf. H. knighti from the Uintan of California, which Krishtalka and Stucky (1983b) referred to H. knighti. In all other dental characters, especially the shallow ectoflexi on M1-3, labially positioned stylar cusps and the close positioning of cusps C and D on M1-2 with the tendency to share a common base, the TBM specimens are indistinguishable from those of Herpetotherium knighti. Considering the individual variation in size noted above for large samples Herpetotherium, we refer the TBM specimens to Herpetotherium knighti.
Herpetotherium marsupium Troxell, 1923a
1923 Herpetotherium marsupium; Troxell, 1923a, p. 508, figures 1-2.
1928 Peratherium marsupium; Simpson, p. 5, figure 4.
1975 Peratherium marsupium; West and Dawson, p. 237.
1970 Peratherium cf. marsupium; McGrew and Sullivan, p. 74.
1973 Peratherium cf. P. marsupium; West, p. 80.
1975 Peratherium cf. P. marsupium; Setoguchi, p. 264, figures 1-4.
1976 Peratherium marsupium; Gazin, p. 2.
1981 Peratherium marsupium; West and Hutchinson, p. 61.
1982 Peratherium cf. marsupium; Bown, p. A43.
1982 Peratherium marsupium; West, p. 2, figures 2a-b.
1982 Herpetotherium sp., cf. H. marsupium; Eaton, p. 162.
1983a Peratherium marsupium; Krishtalka and Stucky, p. 214, figure 3.
1983b Peratherium marsupium; Krishtalka and Stucky, p. 232.
1984 Peratherium marsupium; Krishtalka and Stucky, p. 33, figure 1.
1984 Peratherium marsupium; Storer, p. 16, figures 1c-f.
1996 Herpetotherium sp., cf. H. marsupium; Rothecker and Storer, p.772.
1996 Herpetotherium marsupium; Storer, p. 245, 247.
1998 Peratherium marsupium; Gunnell, p. 85, figure 2a.
2008 Herpetotherium marsupium; Korth, p. 42.
Remarks. The holotype of Herpetotherium marsupium, a partial right dentary with p3-m3 (YPM 13518), came from an undetermined stratigraphic level (either Br2 or Br3) of the Bridger Formation in the Bridger Basin (Troxell, 1923a). Subsequently, numerous specimens of Herpetotherium marsupium have been recorded from the Blacks Fork and Twin Buttes members (biochrons Br2 and Br3) in the Bridger Basin (Gazin, 1976; Krishtalka and Stucky, 1983a, 1983b). Gunnell (1998) referred to Herpetotherium marsupium three specimens collected from exposures of the lowermost Bridger Formation (early Bridgerian, biochron Br1a = Bridger A of Matthew [1909]) in the northern portion of the Bridger Basin near the town of Opal. West and Hutchinson (1981) described a single upper molar (MPM 5888) of Herpetotherium marsupium from locality MPM 2970, which occurs low in the TBM on the southwest flank of Sage Creek Mountain. Herpetotherium marsupium has also been recorded from the early Eocene (late Wasatchian, biochron Wa6) through the middle Eocene (Duchesnean, biochron Du) of North America (Krishtalka and Stucky, 1983b; Korth, 2008), resulting in a geochronologic range of about 12.4 Ma. No additional specimens of this species were recovered from our study area of the TBM exposed on southwest flank of Cedar Mountain.
Family PERADECTIDAE Crochet, 1979
Genus PERADECTES Matthew and Granger, 1921
Type species. Peradectes elegans Matthew and Granger, 1921, by original designation.
Other included species. P. californicus (Stock, 1936); P. chesteri Gazin, 1952; P. pauli Gazin, 1956; P. protinnominatus McKenna, 1960; P. austrinum Sigé, 1971; P. louisi Crochet, 1979; P. minor Clemens, 2006; P. gulottai Rose, 2010; P. coprexeches Willamson and Taylor, 2011.
Peradectes chesteri (Gazin, 1952)
Figure 13.1-4, Table 3
1952 Peratherium chesteri; Gazin, p. 18, pl. 1, figure 1.
1962 Peratherium chesteri; Gazin, p. 21.
1973 Peratherium innominatum, in part; West, p. 79.
1980 Peradectes chesteri: Bown, p. 131.
1982 Peratherium sp., cf. P. innominatum; Bown, p. A43.
1983a Peradectes chesteri; Krishtalka and Stucky, p. 219.
1983b Peradectes chesteri; Krishtalka and Stucky, p. 249, figure 5.
1984 Peradectes chesteri; Krishtalka and Stucky, p. 37, figure 3.
1994 P eradectes chesteri; Bown et al., p. 4, tab. 1.
1998 Peradectes chesteri; Gunnell, p. 88.
2008 Peradectes chesteri; Korth, p. 43.
Referred specimens. From locality UCM 92189: LM1, UCM 68427; partial maxilla with LM2-3, UCM 95781; Lm2 or 3, UCM 78458. From locality SDSNH 5842: partial RM1, SDSNH 110418. From locality DMNH 4672: RdP3, DMNH 75275.
Description. DMNH 75275 is identified as a dP3 (Figure 13.1) because of its nearly equilateral dimensions (just slightly longer than wide), very elongated posterior metastylar wing, and reduced anterior parastylar shelf. It is significantly smaller than the M2-3. The paracone and metacone are equal in size and height. The protocone and talon basin are reduced as compared to those of the M2-3. The paraconule is weakly developed with very short, vestigial pre- and post paraconulecristae, whereas a metaconule is lacking. Stylar cusp C is represented by an elongated, weak bulge along the labial rim of the stylar shelf approximately opposite of the metacone apex, whereas stylar cusps A, B, D, and E are lacking. The ectoflexus is very shallow. The posterior cingulum (postcingulum) is weakly developed and an anterior cingulum (precingulum) is lacking.
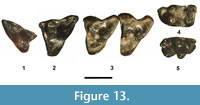 An isolated M1 (UCM 68427, Figure 13.2) is compatible in size and very similar in occlusal morphology to the associated M2-3 (UCM 95781, Figure 13.3) described below, both of which were recovered from UCM Locality 92189. It differs from M2-3 by the following: 1) slightly longer relative to its width (slightly less transversely broad); 2) a weakly-developed stylar cusp (probably cusp D) present on the stylar shelf posterior of the metacone apex and anterior to the labial terminus of the metastylar wing; 3) a shallower ectoflexus; and 4) a metacone slightly larger than the paracone. All of these differences are typically used to separate the M1 from the M2-3 of Peradectes (McGrew, 1939; Lillegraven, 1976; Krishtalka and Stucky, 1983b). Like the M2-3, UCM 68427 is lacking stylar cusps A, C, and E, and a protoconule and metaconule, and has a non-dilambdodont centrocrista with the metacone slightly taller than the paracone.
An isolated M1 (UCM 68427, Figure 13.2) is compatible in size and very similar in occlusal morphology to the associated M2-3 (UCM 95781, Figure 13.3) described below, both of which were recovered from UCM Locality 92189. It differs from M2-3 by the following: 1) slightly longer relative to its width (slightly less transversely broad); 2) a weakly-developed stylar cusp (probably cusp D) present on the stylar shelf posterior of the metacone apex and anterior to the labial terminus of the metastylar wing; 3) a shallower ectoflexus; and 4) a metacone slightly larger than the paracone. All of these differences are typically used to separate the M1 from the M2-3 of Peradectes (McGrew, 1939; Lillegraven, 1976; Krishtalka and Stucky, 1983b). Like the M2-3, UCM 68427 is lacking stylar cusps A, C, and E, and a protoconule and metaconule, and has a non-dilambdodont centrocrista with the metacone slightly taller than the paracone.
UCM 95781 (Figure 13.3) consists of a partial right maxilla with fragments of the M1 due to its crown being broken off, a nearly complete M2 with only the apex of the paracone broken off, and a complete M3. Although the occlusal morphologies of the M2 and M3 are quite similar, the M3 differs by being more transverse and by having a slightly deeper, more defined ectoflexus and a significantly more anteroposteriorly compressed protocone. The M2-3 have the paracone and metacone subequal in size, with the former slightly lower in height. A protoconule and metaconule are lacking. Stylar cusp B is moderately developed and connected lingually to the preparacrista on M2, but on the M3 it is isolated from the labial terminus of the preparacrista by a very shallow, narrower valley. Stylar cusps A, C, D, and E are lacking. The stylar shelf is relatively wide and the ectoflexus relatively deep, especially on M3. Anterior and posterior cingula are lacking.
An isolated left lower molar, either m2 or 3 (Figure 13.4), was recovered from the same locality as the partial maxilla with M2-3. Its trigonid and talonid are equal in width. The primary cusps of the trigonid (protoconid, metaconid, and paraconid) are robust, with the paraconid about one-half the height of the protoconid and the metaconid just slightly lower in height than the protoconid. The paracristid is straight, connecting the paraconid and metaconid. The entoconid is about equal in size to the hypoconulid, closely positioned to it, and separated from it by a shallow notch. The cristid obliqua extends from the hypoconid apex to terminate on the posterior wall of the trigonid, just below the protocristid notch. The anterior and posterior cingula are moderately distinct. The labial border between the trigonid and talonid is almost straight with very little emargination.
Remarks. Crochet (1979) erected the tribe Peradectini, based primarily on North American species of Peradectes. Subsequent investigators have either elevated its rank to the subfamily level as Peradectinae (e.g., Korth, 2008) or family level as Peradectidae (e.g., Reig et al., 1985; Johanson, 1996; Rothecker and Storer, 1996; Case et al., 2005; Horovitz et al., 2009; Rose, 2010) or used both ranks (e.g., Korth, 1994). Williamson et al. (2012) regarded "Peradectidae" sensu lato as an unranked clade composed a basal polytomy of species, which also included certain Cretaceous Eurasian and Paleocene South American taxa. Following Horovitz et al. (2009), we regard Peradectidae as a familial rank.
Peradectes is characterized by the following (Krishtalka and Stucky, 1983b; Korth, 2008): 1) the M1-3 paracone and metacone subequal in size and not dilambdodont; 2) the M1-3 stylar cusps, protoconule and metaconule weakly developed and in some species all stylar cusps, except B, vestigial or absent; 3) the M1-3 posterolingual base of the protocone unexpanded; 4) the m1-3 talonid relatively short with a labially positioned cristid obliqua and a low entoconid; and 5) the m1-3 hypoconulid and entoconid subequal in size, positioned relatively close to each other, and separated by a weak notch. The TBM specimens exhibit the above diagnostic characters of Peradectes and can be confidently assigned to the genus.
The holotype of Peradectes chesteri, a partial right dentary with m3 (USNM 19199), came from the late Wasatchian (biochron Wa7) Wasatch Formation near La Barge, Wyoming (Gazin, 1952). Subsequently, Peradectes chesteri has also been recorded from earliest Bridgerian (biochron Br1a) through the early Uintan (biochron Ui1b), resulting in a geochronologic range of about 7 Ma (West, 1973; Krishtalka and Stucky, 1983a, 1983b; Gazin, 1976; Walsh, 1996; Gunnell, 1998; Gunnell et al., 2009).
Peradectes chesteri is characterized by having the following (Krishtalka and Stucky, 1983a, 1983b, 1984): 1) small size; 2) M1-3 with a moderately reduced stylar cusp B and other stylar cusps vestigial or absent; 3) M1-3 paracone taller than protocone and stylar cusp B; 4) M1-3 paraconule and metaconule greatly reduced (vestigial); 5) M2-3 ectoflexus well defined and moderately deep; 6) M3 with a highly compressed protocone and more transversely broadened than M2; 7) p3 with short talonid that is lower in height than m1; 8) m1-3 narrow in proportion to length with little to no labial emargination between the trigonid and talonid; 9) m1 smaller than m2-3; and 10) m4 talonid shorter than trigonid. The specimens from the TBM are indistinguishable in size and occlusal morphology from those of Peradectes chesteri and are referred to that species.
Peradectes californicus (Stock, 1936)
Figure 13.5, Table 3
1936 Peratherium californicum; Stock, p. 123, figures 2, 2a.
1975 Nanodelphys cf. N. minutus, in part; Setoguchi, p. 269.
1976 Nanodelphys californicus; Lillegraven, p. 91, pl. 5, figure 3, pl. 6, figures 1a-c, pl. 7, figures 1a-c, 2a-c, pl. 8, figures 1a-c, pl. 9, figures 1a-c, 2a-c, 3a-c, pl. 10, figures 1a-c, 2a-c, 3a-c, 4a-c.
1983a Peradectes californicus; Krishtalka and Stucky, p. 219.
1983b Peradectes californicus; Krishtalka and Stucky, p. 247.
1984 Peradectes californicus; Storer, p. 21, figures 1k.
1990 Peradectes californicus; Kelly, p. 8.
1994 Peradectes californicus; Kelly and Whistler, p. 2, figure 1.
1996 Peradectes californicus; Rothecker and Storer, p. 773, figure 1o-q.
2008 Peradectes californicus; Korth, p. 43.
2010 Peradectes californicus; Kelly, p. 160, figure 1b-f.
2013 Peradectes californicus; Kelly, p. 58, figure 2c.
Referred specimen. From locality SDSNH 5844: Rm2 or 3, SDSNH 110432.
Description. The lower molar trigonid is slightly wider than the talonid. The primary cusps of the trigonid are robust, with the protoconid larger and slightly taller than the metaconid. The paraconid and metaconid are subequal in size, with the paraconid about one-half the height of the protoconid. The paracristid extends anterolabially from the protoconid apex and then turns more directly labially to join the paraconid apex, giving it a curved appearance in occlusal view. The entoconid and hypoconulid are subequal in size, closely positioned to one another and separated by a shallow notch. The cristid obliqua extends anterolabially from the hypoconid apex to terminate on the posterior wall of the trigonid, between the paracristid notch and the protoconid apex. The labial margin between the trigonid and talonid is distinctly emarginated. The anterior cingulid is robust, whereas the posterior cingulid is slightly less developed.
Remarks. One lower molar, either m2 or 3 (SDSNH 110432, Figure 13.5), that came from the highest fossil yielding level in the TBM, can be eliminated as belonging to Herpetotherium or Copedelphys by having the following combination of characters: 1) small size; 2) the entoconid and hypoconulid about equal in size and height, positioned close to each other and separated by a weak notch; and 3) the hypoconulid cuspate (not shelf-like) and not projecting well posterior of the entoconid. This talonid morphology is typical of members of the Peradectes - Nanodelphys lineage (Krishtalka and Stucky, 1983b; Korth, 1994).
As discussed above, the taxonomic history of Peradectes californicus is complicated. The holotype, a partial right dentary with p3-m2 (LACM [CIT] 1943), came from the late Uintan (biochron Ui3) part of the Sespe Formation, Ventura County, California (Stock, 1936). Peradectes californicus has also been recorded from the late Uintan Mission Valley and Santiago Formations of San Diego County, California, the late Duchesnean (biochron Du) part of the Sespe Formation, California, the medial Uintan to early Duchesnean (biochrons Ui2-Du) Wagon Bed Formation of the Wind River Basin, Wyoming, the Duchesnean (biochron Du) Tepee Trail Formation of Wyoming, and late Uintan to Duchesnean (biochrons Ui3- Du) Cypress Hills Formation of Saskatchewan (Setoguchi, 1975; Lillegraven, 1976; Krishtalka and Stucky, 1983b; Storer, 1984; Rothecker and Storer, 1996; Korth, 2008; Kelly, 2010, 2013).
Peradectes californicus is similar in size to Peradectes chesteri, but differs by the following (Lillegraven, 1976; Krishtalka and Stucky, 1983a, 1983b, 1984): 1) M1-3 length shorter relative to width (ap and tr nearly equal); 2) M1-3 paraconule, metaconule usually slightly better developed, but still weak; 3) M1-3 with very small stylar cusps C and D commonly present; 3) M3 less transverse with less compression of protocone; 4) p3 talonid absent; and 5) m1-3 wider relative to length and with a distinct labial emargination between the trigonid and talonid. As in Peradectes californicus, SDSNH 110432 exhibits a very distinct emargination between the trigonid and talonid and its width relative to its length is wider (tra/ap = 0.62) than that of the m2 or 3 referred above to Peradectes chesteri from lower in the TBM (tra/ap = 0.51).
Setoguchi (1973) originally assigned certain specimens from the Uintan and Duchesnean Tepee Trail Formation at Badwater Creek to two informal species of Nanodelphys; Nanodelphys sp., cf. N. minutus and Nanodelphys sp. nov. Later, Setoguchi (1975) subsumed the sample of Nanodelphys sp. nov. into his Nanodelphys sp., cf. N. minutus. Krishtalka and Stucky (1983b) determined that Setoguchi's (1975) sample of Nanodelphys sp., cf. N. minutus actually consisted of three species; Peradectes californicus, Peratherium innominatum (= Copedelphys innominata, see Rothecker and Storer [1996]), and Nanodelphys sp., cf. N. minutus (= Nanodelphys sp. nov. of Setoguchi [1973]). Korth (1994) noted that the upper molars that Krishtalka and Stucky (1983b) retained in Nanodelphys sp., cf. N. minutus may represent a new species of Peradectes because their length to width ratios are intermediate between those of Nanodelphys and Peradectes, and they have less reduction of the stylar cusps than that of Nanodelphys. Korth (1994) also noted that the lower molars of Nanodelphys differ from those of Peradectes by having relatively narrower and more elongated trigonids and straighter paracristids. In SDSNH 110432, the paracristid is slightly curved posteriorly and the ratio for its trigonid length/ap is 0.54, whereas that for the mean m2-3 trigonid length/ap of Nanodelphys hunti is 0.64 (Korth, 1994).
Based on the above comparisons, SDSNH 110432 can be eliminated as representing either Peradectes chesteri or Nanodelphys. sp., cf. N. minutus, but is indistinguishable from the lower molars of Peradectes californicus and is assigned to that species.
A REEVALUATION OF EARLIEST UINTAN FAUNAS
Correlations of Ui1a and Ui1b Faunas
Flynn (1986) proposed the "Shoshonian" subage for what he regarded as earliest Uintan faunas whose compositions appeared transitional between those of the Bridgerian and Uintan. He based his new subage primarily on the faunas from the Bone Bed A (Horizon D) of the type Tepee Trail Formation, Wyoming, and what he considered as correlative faunas from the Friars and Mission Valley Formations in the greater San Diego area, California. Based on his magnetostratigraphic correlations alone, Flynn (1986) also proposed that a "Shoshonian" fauna should be present within the middle to upper part of the lower subunit of the Adobe Town Member of the Washakie Formation, Wyoming. After Flynn's (1986) proposal, many investigators debated the validity of the "Shoshonian" subage and it was not generally accepted (e.g., Krishtalka et al., 1987; Prothero and Emry, 1996; Walsh, 1996a; Janis, 1998). Robinson et al. (2004) questionably proposed a new biochron named Ui1 for what they regarded as earliest Uintan faunas, which included most of the faunas that Flynn (1986) referred to his "Shoshonian" subage. For many years, investigators referred to earliest Uintan faunas informally as "Shoshonian" in age and subsequent to Robinson et al. (2004), also as Ui1 faunas. Most recently, Gunnell et al. (2009) divided Ui1 into two biochrons, Ui1a and Ui1b, with Ui1a based on the oldest transitional faunas and Ui1b based on younger transitional faunas with both biochrons characterized by the following: 1) the presence of index taxa; and 2) the lowest range datum's (LRD = FAD, first appearance datum of Berggren and Van Couvering, 1974, and Woodburne, 2004a) and highest range datum's (HRD = LAD, last appearance datum of Woodburne, 2004a) of certain key taxa. The changing nomenclature and definition of what the term "earliest Uintan" refers to has led to some confusion in the literature, wherein investigators have sometimes included both Ui1a and Ui1b faunas in the earliest Uintan (e.g., Tsukui, 2016). For this reason, we present a reevaluation of the correlations of the Ui1a and Ui1b faunas, which will hopefully clarify the faunal distinctions between these biochrons.
Gunnell et al. (2009) assigned the type section of the TBM as the stratotype section for Ui1a, and also referred the Tertiary Basal Local Fauna (Walton, 1992) of the Devil's Graveyard Formation, Texas, to Ui1a along with possibly all of informal unit A of the Uinta Formation in the eastern Uinta Basin and the lower part of unit B of the Uinta Formation in the western Uinta Basin, Utah. Gunnell et al. (2009) did not recognize a stratotype section for the Ui1b, but referred the following sections to it: 1) the Washakie Formation of the Sand Wash Basin of Colorado; 2) the middle unit of the Adobe Town Member of the Washakie Formation of Wyoming; 3) the lowermost Tepee Trail Formation of the East Fork Basin of Wyoming; 4) the Friars Formation and the upper part of Member B of the Santiago Formation of southern California; 5) the portion of the lower member of the Devil's Graveyard Formation yielding the Whistler Squat Local Fauna of Texas; and 6) tentatively all or a portion of the informal units A and B1 of the Uinta Formation of the eastern part of the Uinta Basin and the lowermost part of unit B in the western part of the Uinta Basin of Utah.
Tsukui (2016) provided the most complete record of the magnetostratigraphy of the Bridger Formation along with precise U-Pb geochronology, including the placement of the Uintan-Bridgerian boundary within the lower part of Chron C21n. Our new U-Pb dates from the stratotype section of the TBM (Figure 1, Figure 10) provide accurate constraints on the ages of the Ui1a localities in the stratigraphic interval from the tuffaceous white sandstone to the highest localities (Localities SDSNH 5843 and 5844, and Locality DMNH 4673). However, a precise date for the lowermost Ui1a localities (Localities UCM 92189 and SDSNH 5841), which occur at about 1 to 2 m above the base of the TBM (= Basal E Limestone) or about 72-73 m below the tuffaceous white sandstone (Figure 1), has not been determined. Murphey et al. (1999) reported an 40Ar/39Ar date of 47.13 ± 0.45 Ma (corrected) for the uppermost Twin Buttes Member tuff (= Basal E tuff of Murphey and Evanoff, 2007) that occurs about 8 m below the base of the TBM in the uppermost Twin Buttes Member, but this date has a large uncertainty so that it is of little value in constraining the age of the beginning of the Ui1a in the TBM. The base of the TBM begins at the Basal E Limestone at about 27 m above the base of Chron C21n (= the boundary between chrons C21r and C21n) in the Sage Creek Mountain sections of Tsukui (2016). The C21r/C21n boundary has been calibrated to 47.349 Ma according to the GPTS 2012 (Ogg, 2012). However, a number of studies using different methods have suggested varying calibrations for the Paleogene segment of the GPTS (e.g., Tsukui and Clyde, 2012; Westerhold and Rӧhl, 2009; Westerhold et al., 2015; Tsukui, 2016). If a consensus on the age for the base of Chron C21n emerges, the age for the beginning of the Ui1a in the TBM will be better constrained. The date for the beginning of the Ui1a in the TBM could be further clarified by re-dating the uppermost Twin Buttes Member tuff using the more precise U-Pb method.
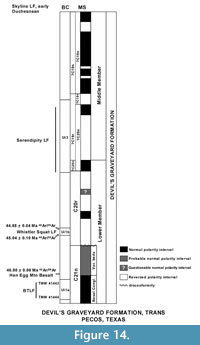
The magnetostratigraphy of the Devil's Graveyard Formation (DGF) of the Agua Fria area, Texas, is complicated with a number of possible interpretations for Chron assignments in the middle member (Walton, 1992; Prothero, 1996b). The polarity of the samples from the variegated beds (maximum thickness of about 33 m) overlying the Basal Conglomerate was difficult to assess, but two samples showing normal polarity suggested that this interval may represent Chron C21n (Walton, 1992). Furthermore, the contact between the variegated beds and the overlying strata containing the Whistler Squat Quarry is erosional (Stevens et al., 1984), indicating that some portion of the strata may be missing. So, the Basal Tertiary Local Fauna (BTLF, Walton, 1992, = localities TMM 41443 [locality name Junction] and TMM 41444 [locality name.6 miles east of Junction]) and the Hen Egg Mountain localities in the lowermost part of the lower member of the DGF appear not to occur at the top of Chron C21n, but somewhere within the chron (Figure 14). The BTLF and the Hen Egg Mountain localities are Ui1a in age (Gunnell et al., 2009; Kelly and Murphey, 2016a; Murphey and Kelly, 2017), whereas the stratigraphically higher Whistler Squat Quarry in the lower member of the DGF is Ui1b in age and occurs in Chron C20r (Gunnell et al., 2009; Campisano et al. 2014; Kelly and Murphey, 2016a; Murphey and Kelly, 2017). Murphey and Kelly (2017) retained the BTLF in the Ui1a, but noted it may be slightly younger than the Ui1a fauna from the TBM of the Bridger Formation based on certain faunal differences (e.g., the larger proportion of Bridgerian hold over taxa present in the TBM, the lack of selenodont artiodactyls in the TBM and the fact that the rodent cf. Pareumys sp. of the TBM is slightly less derived than Pareumys boskeyi Wood, 1973, of the BTLF), along with an 40Ar/39Ar date of 46.80 ± 0.08 Ma (corrected) for a basalt overlying the Hen Egg Mountain localities (Miggins, 2009). The proposal that the TBM is older than the BTLF is further supported by our new U-Pb dates from the TBM, which provide precise dates for the fauna (46.94 Ma for locality Roll the Bones and 47.31 Ma for the tuffaceous white sandstone). Thus, what can be confidently stated is that the BTLF occurs within Chron C21n and the Whistler Squat Quarry occurs somewhere within the lower part Chron C20r (Figure 14).
 The history of the biostratigraphic/lithostratigraphic divisions of the Uinta Formation of the Uinta Basin is somewhat complicated (Prothero, 1996a). Peterson (in Osborn, 1895) recognized three horizons, in ascending stratigraphic order, Uinta A, B, and C, and considered the lithostratigraphic boundary between his Uinta B and C to occur at the change from gray mudstone beds to red and orange claystone beds in the Devil's Playground area of the eastern Uinta Basin. However, Osborn and Matthew (1909) and Osborn (1929) redefined the Uinta B-C boundary as the Amynodon sandstone of Riggs (1912), a locally restricted unit that occurs stratigraphically lower in the section (Cashion, 1986; Townsend et al., 2006). Moreover, Osborn (1929) further modified Peterson's (in Osborn, 1895) divisions of the Uinta Formation as follows: 1) Peterson's Uinta A was divided with the lower half retained as Uinta A and the upper half named Uinta B1; and 2) Peterson's original Uinta B was renamed Uinta B2. Many of the early collections of fossil mammals from the Uinta Basin lacked detailed locality data and stratigraphic control, but recent investigations indicate that the Ui2-Ui3 faunal transition actually occurs at about 53-63 m above the Amynodon sandstone or at about 10-20 m below the revised lithologic boundary of the Uinta B-C in the eastern Uinta Basin (Rasmussen et al., 1999; Townsend et al., 2006).
The history of the biostratigraphic/lithostratigraphic divisions of the Uinta Formation of the Uinta Basin is somewhat complicated (Prothero, 1996a). Peterson (in Osborn, 1895) recognized three horizons, in ascending stratigraphic order, Uinta A, B, and C, and considered the lithostratigraphic boundary between his Uinta B and C to occur at the change from gray mudstone beds to red and orange claystone beds in the Devil's Playground area of the eastern Uinta Basin. However, Osborn and Matthew (1909) and Osborn (1929) redefined the Uinta B-C boundary as the Amynodon sandstone of Riggs (1912), a locally restricted unit that occurs stratigraphically lower in the section (Cashion, 1986; Townsend et al., 2006). Moreover, Osborn (1929) further modified Peterson's (in Osborn, 1895) divisions of the Uinta Formation as follows: 1) Peterson's Uinta A was divided with the lower half retained as Uinta A and the upper half named Uinta B1; and 2) Peterson's original Uinta B was renamed Uinta B2. Many of the early collections of fossil mammals from the Uinta Basin lacked detailed locality data and stratigraphic control, but recent investigations indicate that the Ui2-Ui3 faunal transition actually occurs at about 53-63 m above the Amynodon sandstone or at about 10-20 m below the revised lithologic boundary of the Uinta B-C in the eastern Uinta Basin (Rasmussen et al., 1999; Townsend et al., 2006).
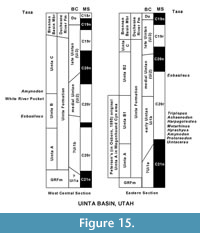 Prothero (1996a) documented the magnetostratigraphy of the Uinta Formation in the Uinta Basin. Assuming Prothero's (1996a) magnetostratigraphy is correct, what is recognized as Uinta A in the eastern section is older than the Uinta A in the west-central section. There are no documented mammal fossils with reliable locality data from the Uinta A (in the modern sense) in the eastern section or west-central section of the Uinta Formation. Kelly et al. (2012) reevaluated the correlation of late Uintan and Duchesnean faunas of the Uinta Basin to the GPTS and determined that late Uintan faunas (Ui3) occur from somewhere within Chron C20n to the lower third of Chron C19n, and that the Uintan-Duchesnean boundary occurs in about the lower third of Chron C19n. Considering that certain taxa whose LRDs within the eastern section (e.g., the rhino Uintaceras Holbrook and Lucas, 1997, and the agriochoerid Protoreodon Scott and Osborn, 1887) are low or near the base of the Uinta B1 or are recorded within the Uinta B1 (e.g., the brontothere Metarhinus Osborn, 1908, the rhino Amynodon Marsh, 1877, and the entelodont Achaenodon Cope, 1873b), then by definition (Gunnell et al., 2009), this interval represents the Ui1b (Figure 15). The speculation of Gunnell et al. (2009) as to what portions of the Uinta Formation represent the Ui1a is still uncertain until the discovery of age diagnostic taxa. It is speculative to use the paleomagnetic data to assign with any certainty biochrons to stratigraphic intervals within the Uinta Formation without diagnostic mammal fossil assemblages to support the correlations. Thus, what can be reasonably stated regarding the correlations of faunas to the magnetostratigraphy of the Uinta and Duchesne River Formations in the Uinta Basin are as follows (Figure 15): 1) no Ui1a faunas are currently recognized; 2) Ui1b faunas appear to occur at or near the top of Chron C21n to somewhere within Chron C20r; 3) Ui2 faunas occur from somewhere within Chron C20r to somewhere within Chron C20n; and 4) Ui3 faunas occur from somewhere within Chron C20n through about the lower third of Chron C19n.
Prothero (1996a) documented the magnetostratigraphy of the Uinta Formation in the Uinta Basin. Assuming Prothero's (1996a) magnetostratigraphy is correct, what is recognized as Uinta A in the eastern section is older than the Uinta A in the west-central section. There are no documented mammal fossils with reliable locality data from the Uinta A (in the modern sense) in the eastern section or west-central section of the Uinta Formation. Kelly et al. (2012) reevaluated the correlation of late Uintan and Duchesnean faunas of the Uinta Basin to the GPTS and determined that late Uintan faunas (Ui3) occur from somewhere within Chron C20n to the lower third of Chron C19n, and that the Uintan-Duchesnean boundary occurs in about the lower third of Chron C19n. Considering that certain taxa whose LRDs within the eastern section (e.g., the rhino Uintaceras Holbrook and Lucas, 1997, and the agriochoerid Protoreodon Scott and Osborn, 1887) are low or near the base of the Uinta B1 or are recorded within the Uinta B1 (e.g., the brontothere Metarhinus Osborn, 1908, the rhino Amynodon Marsh, 1877, and the entelodont Achaenodon Cope, 1873b), then by definition (Gunnell et al., 2009), this interval represents the Ui1b (Figure 15). The speculation of Gunnell et al. (2009) as to what portions of the Uinta Formation represent the Ui1a is still uncertain until the discovery of age diagnostic taxa. It is speculative to use the paleomagnetic data to assign with any certainty biochrons to stratigraphic intervals within the Uinta Formation without diagnostic mammal fossil assemblages to support the correlations. Thus, what can be reasonably stated regarding the correlations of faunas to the magnetostratigraphy of the Uinta and Duchesne River Formations in the Uinta Basin are as follows (Figure 15): 1) no Ui1a faunas are currently recognized; 2) Ui1b faunas appear to occur at or near the top of Chron C21n to somewhere within Chron C20r; 3) Ui2 faunas occur from somewhere within Chron C20r to somewhere within Chron C20n; and 4) Ui3 faunas occur from somewhere within Chron C20n through about the lower third of Chron C19n.
 The Washakie Formation in the main Washakie Basin, Wyoming, is subdivided into the Kinney Rim Member and the much thicker and overlying Adobe Town Member. The Adobe Town Member has been subdivided into lower, middle and upper informal subunits (McCarroll et al., 1996a). The lowest stratigraphic datum (LSD) of Amynodon is low in the Adobe Town middle subunit (bed 630 of Roehler, 1973), which by definition equates this level to Ui1b (Gunnell et al., 2009). The Adobe Town middle subunit has also yielded the two Ui1b index species Achaenodon robustus Osborn, 1883, and Achaenodon insolens Cope, 1873b (McCarroll et al., 1996a; Gunnell et al., 2009), along with Uintaceras, Metarhinus, and the camelids Protylopus Wortman, 1898 and Poebrodon Gazin, 1955, of which the first two are typical Ui1b taxa (Figure 16). Tsukui (2016: figure 2.11) correlated the lower part of the Adobe Town middle subunit to the Ui1a, but it appears that her Ui1a interval actually represents the Ui1b, at least in part, because of the LSD of Amynodon (Walsh, 1996a; Gunnell et al., 2009). The upper boundary of the Ui1b is undetermined for the Washakie Formation, but could extend upward into Chron C20r. The faunas from the Kinney Rim Member and the Adobe Town lower subunit consist of typical late Bridgerian (Br3) taxa with no Ui1a or Ui1b index taxa (McCarroll et al., 1996a, 1996b). Most of the mammal fossils from the lower subunit came from the middle red beds (McCarroll et al., 1996a). The upper part of the lower subunit is sparsely fossiliferous with no Ui1a age diagnostic taxa, and the lowermost part of the middle subunit is unfossiliferous (McCarroll et al., 1996a). It is possible that some interval between the upper part of the lower subunit and the lowermost part of the middle subunit is Ui1a in age, but the lack of fossils makes this highly speculative and unprovable. The lower 90 m of the Skull Creek section in the Washakie Basin is unsampled (polarity undetermined). However, Stucky et al. (1996) stated that it contains a "Shoshonian" fauna. This fauna has not been documented in detail, so whether it represents a Ui1a or Ui1b fauna is undetermined. In summary, a Ui1b fauna occurs within Chron C21n and appears to extend upward into Chron C20r and there are no confidently recognized Ui1a faunas from Washakie Formation of the main Washakie Basin (Figure 16).
The Washakie Formation in the main Washakie Basin, Wyoming, is subdivided into the Kinney Rim Member and the much thicker and overlying Adobe Town Member. The Adobe Town Member has been subdivided into lower, middle and upper informal subunits (McCarroll et al., 1996a). The lowest stratigraphic datum (LSD) of Amynodon is low in the Adobe Town middle subunit (bed 630 of Roehler, 1973), which by definition equates this level to Ui1b (Gunnell et al., 2009). The Adobe Town middle subunit has also yielded the two Ui1b index species Achaenodon robustus Osborn, 1883, and Achaenodon insolens Cope, 1873b (McCarroll et al., 1996a; Gunnell et al., 2009), along with Uintaceras, Metarhinus, and the camelids Protylopus Wortman, 1898 and Poebrodon Gazin, 1955, of which the first two are typical Ui1b taxa (Figure 16). Tsukui (2016: figure 2.11) correlated the lower part of the Adobe Town middle subunit to the Ui1a, but it appears that her Ui1a interval actually represents the Ui1b, at least in part, because of the LSD of Amynodon (Walsh, 1996a; Gunnell et al., 2009). The upper boundary of the Ui1b is undetermined for the Washakie Formation, but could extend upward into Chron C20r. The faunas from the Kinney Rim Member and the Adobe Town lower subunit consist of typical late Bridgerian (Br3) taxa with no Ui1a or Ui1b index taxa (McCarroll et al., 1996a, 1996b). Most of the mammal fossils from the lower subunit came from the middle red beds (McCarroll et al., 1996a). The upper part of the lower subunit is sparsely fossiliferous with no Ui1a age diagnostic taxa, and the lowermost part of the middle subunit is unfossiliferous (McCarroll et al., 1996a). It is possible that some interval between the upper part of the lower subunit and the lowermost part of the middle subunit is Ui1a in age, but the lack of fossils makes this highly speculative and unprovable. The lower 90 m of the Skull Creek section in the Washakie Basin is unsampled (polarity undetermined). However, Stucky et al. (1996) stated that it contains a "Shoshonian" fauna. This fauna has not been documented in detail, so whether it represents a Ui1a or Ui1b fauna is undetermined. In summary, a Ui1b fauna occurs within Chron C21n and appears to extend upward into Chron C20r and there are no confidently recognized Ui1a faunas from Washakie Formation of the main Washakie Basin (Figure 16).
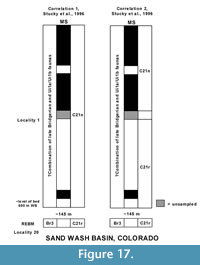 In the Sand Wash Basin, the putative "Shoshonian" localities have yielded a mixture of typical Bridgerian and Uintan taxa. As noted above, "Shoshonian" or "earliest Uintan" faunas of Flynn (1986) include both Ui1a and Ui1b faunas (Gunnell et al., 2009), so there is much confusion in the literature regarding the faunal separation of these biochrons. Of the two main Sand Wash Basin sections, Locality 1 (DMNH 296) of West and Dawson (1975) is reported to occur at the top of a reversed polarity interval within the lower section, which Stucky et al. (1996) regarded as "Shoshonian" in age, whereas the upper section (upper Vaughn Draw section) consists of a long normal interval with a short reversed interval occurring near its center (Figure 17). Except for Localities 1 and 20, Stucky et al. (1996: Figure 4) did not provide the relative stratigraphic positions of any of the other Sand Wash Basin localities relative to their two main sections, only a label reading "earliest Uintan" that was positioned in the lower part of the normal polarity interval in the upper Vaughn Draw section. Stucky et al. (1996) provided two possible correlations of the Sand Wash Basin sections to the magnetostratigraphic sections of McCarroll et al. (1996a) in the main Washakie Basin (Figure 17). In correlation 1, both of the main Sand Wash sections above the Robins Egg Blue Marker Bed (REBM) sample are correlated to Chron C21n. Correlation 1 assumes that the 90 m unsampled portion of the lower Skull Creek section of the main Washakie Basin will be found to be of reversed polarity in the future. In correlation 2, the lower reversed interval is correlated with the uppermost Chron C21r. The base of the lower Sand Wash Basin section was stated to be stratigraphically positioned at the Tbl/Tbu contact, which was previously mapped by McKay (1975) as the lower and upper parts of the Bridger Formation, respectively. Stucky et al. (1996) stated that Locality 20 is the only locality that occurs below the REBM (= bed 579 of Roehler, 1973) in the Sand Wash Basin. Stucky et al. (1996) sampled the polarity of the REBM from an isolated outcrop that occurs about 14.5 km north of the other two main Sand Wash Basin sections and found it to be reversed. Based on the stratigraphy in the main Washakie Basin to the north, Stucky et al. (1996) estimated that about 143 m of section separated the REBM from the base of their lower Sand Wash Basin section, placing Locality 1 at about 205 m above the REBM, which they also estimated to be about equal to the level of bed 600 of Roehler (1973) in the Washakie Formation of the Washakie Basin (McCarroll et al., 1996a). Assuming the REBM in the Sand Wash Basin is temporally equivalent to the REBM in the main Washakie Basin, which occurs in Chron C21r, a Br3 (late Bridgerian) age for Locality 20 is supported.
In the Sand Wash Basin, the putative "Shoshonian" localities have yielded a mixture of typical Bridgerian and Uintan taxa. As noted above, "Shoshonian" or "earliest Uintan" faunas of Flynn (1986) include both Ui1a and Ui1b faunas (Gunnell et al., 2009), so there is much confusion in the literature regarding the faunal separation of these biochrons. Of the two main Sand Wash Basin sections, Locality 1 (DMNH 296) of West and Dawson (1975) is reported to occur at the top of a reversed polarity interval within the lower section, which Stucky et al. (1996) regarded as "Shoshonian" in age, whereas the upper section (upper Vaughn Draw section) consists of a long normal interval with a short reversed interval occurring near its center (Figure 17). Except for Localities 1 and 20, Stucky et al. (1996: Figure 4) did not provide the relative stratigraphic positions of any of the other Sand Wash Basin localities relative to their two main sections, only a label reading "earliest Uintan" that was positioned in the lower part of the normal polarity interval in the upper Vaughn Draw section. Stucky et al. (1996) provided two possible correlations of the Sand Wash Basin sections to the magnetostratigraphic sections of McCarroll et al. (1996a) in the main Washakie Basin (Figure 17). In correlation 1, both of the main Sand Wash sections above the Robins Egg Blue Marker Bed (REBM) sample are correlated to Chron C21n. Correlation 1 assumes that the 90 m unsampled portion of the lower Skull Creek section of the main Washakie Basin will be found to be of reversed polarity in the future. In correlation 2, the lower reversed interval is correlated with the uppermost Chron C21r. The base of the lower Sand Wash Basin section was stated to be stratigraphically positioned at the Tbl/Tbu contact, which was previously mapped by McKay (1975) as the lower and upper parts of the Bridger Formation, respectively. Stucky et al. (1996) stated that Locality 20 is the only locality that occurs below the REBM (= bed 579 of Roehler, 1973) in the Sand Wash Basin. Stucky et al. (1996) sampled the polarity of the REBM from an isolated outcrop that occurs about 14.5 km north of the other two main Sand Wash Basin sections and found it to be reversed. Based on the stratigraphy in the main Washakie Basin to the north, Stucky et al. (1996) estimated that about 143 m of section separated the REBM from the base of their lower Sand Wash Basin section, placing Locality 1 at about 205 m above the REBM, which they also estimated to be about equal to the level of bed 600 of Roehler (1973) in the Washakie Formation of the Washakie Basin (McCarroll et al., 1996a). Assuming the REBM in the Sand Wash Basin is temporally equivalent to the REBM in the main Washakie Basin, which occurs in Chron C21r, a Br3 (late Bridgerian) age for Locality 20 is supported.
Locality 1 has yielded the following taxa (Stucky et al., 1996): Herpetotherium knighti (= Peratherium marsupium of West and Dawson, 1975, see Krishtalka and Stucky, 1983b); the pantolestid Pantolestes natans Matthew, 1909; the hyracodont Triplopus sp. (= Hyrachyus small sp. of West and Dawson, 1975, see Stucky and Snyder, 1992, and Stucky et al., 1996); and an indeterminate uintathere. Although the holotype of Herpetotherium knighti is from the Bridger Formation (Br3) at Tabernacle Butte (McGrew, 1959, in McGrew et al., 1959), it is a long ranging species (Wasatchian through Duchesnean), so its occurrence at Locality 1 is of little biostratigraphic significance (Krishtalka and Stucky, 1983). Pantolestes natans occurs in the late Bridgerian (Br3) of the Bridger Formation (Matthew, 1909; Gazin, 1976) and is also known from the Adobe Town lower member (Br3) of the Washakie Formation in the main Washakie Basin (McCarroll et al., 1996a). Westgate and Emry (1985) tentatively assigned a partial dentary lacking teeth to cf. Pantolestes natans. This specimen came from the Crow Creek Quarry of the Jackson Group, Arkansas, which is probably no older than late Uintan (Ui3). Although the Crow Creek Quarry edentulous dentary appears to represent a pantolestid, its specific identification is dubious and cannot be regarded as a reliable record for the species. Triplopus sp. was also reported to occur at Locality 3 in the Sand Wash Basin (Stucky et al., 1996). Triplopus Cope, 1880, is a common Uintan genus that extends into the Duchesnean (Prothero, 1998) and its LRD is otherwise in the Turtle Bluff Member Fauna (Ui1a) of the Bridger Formation (Murphey and Kelly, 2017).
West and Dawson (1975) reported the occurrence of the equid Orohippus sylvaticus (Leidy, 1870) from Localities 3, 4, and 20 of the Washakie Formation in the Sand Wash Basin. West and Dawson (1975) only compared the Sand Wash Basin equid specimens to Orohippus agilis Marsh, 1873, and Orohippus sylvaticus and assigned them to O. sylvaticus based on the subequal widths of the trigonids and talonids in p4 and a broad m3 hypoconulid. The Sand Wash Basin specimens exhibit relatively wide p4 trigonids that are nearly equal in width to the talonids, as seen in O. sylvaticus, whereas in O. agilis the p4 talonid is much wider relative to the p4 trigonid. Also, in O. agilis, the p4 is the widest of all the lower teeth (Kitts, 1957). The character state of a broad m3 hypoconulid is not present in O. sylvaticus, instead in this species the m3 hypoconulid is relatively long, narrow, and unconstricted anteriorly, whereas in O. agilis the m3 hypoconulid is relatively short and wide (Kitts, 1957).
The equids Orohippus Marsh, 1872b, and Epihippus Marsh, 1878, are distinguished from each other by the progressive molarization of the upper and lower premolars in the latter (Granger, 1908; Kitts, 1957; MacFadden, 1980; Kelly and Murphey, 2016b). In both Orohippus and Epihippus, the p4 is molariform. The main difference in the lower premolars of Orohippus and Epihippus is the greater degree of molarization of p2-3 in Epihippus (Granger, 1908; Kitts, 1957; MacFadden, 1980; Kelly and Murphey, 2016b). Separating these genera based on the morphology of p4 is difficult since both exhibit a molariform p4. Moreover, in both Orohippus sylvaticus and Epihippus, the p4 trigonid and talonids widths are subequal, so this character cannot be used to differentiate them. In examining the p4s of O. sylvaticus from the type Bridger Formation, one difference may be a slightly better developed p4 metastylid in Epihippus, which is seen in the unworn p4 of UCM 24302 from the Sand Wash Basin. West and Dawson (1975) did not compare the Sand Wash equid specimens to Epihippus and they did not report any specimens preserving the p2 or p3. This is problematic because a definitive generic diagnosis requires knowledge of the morphologies of p2 and p3. Interestingly, Stucky et al. (1996) listed the occurrence of Epihippus parvus Granger, 1908 (= E. gracilis [Marsh, 1871], see MacFadden, 1980), from West and Dawson's (1975) Locality 16. This specimen was not described or illustrated by Stucky et al. (1996), so confirmation of their assignment cannot be made. The specimens illustrated by West and Dawson (1975, figures 5-7) are not complete enough to confirm their generic assignments and could represent either Orohippus or Epihippus. Based on the fact that the RBEM is well within Chron C21r in the Washakie Formation of the main Washakie Basin and that interval has yielded only late Bridgerian mammals, the equid specimen from Locality 20 may very well represent Orohippus. In addition, the other taxa recorded from Locality 20 (the viverravid Viverravus sp., the primate Hemiacodon sp (= Washakius sp. of West and Dawson, 1975, see Stucky et al., 1996), and the rodent Tillomys sp., cf. T. senex Marsh, 1872b, are typical Bridgerian taxa. The equid specimens from Localities 3 and 4 should be reexamined to see whether there are any specimens that can be confidently assigned to Orohippus. For now, all that can be said of the Sand Wash Basin equids is that their generic assignments are uncertain and the specimens are in need of a more detailed systematic analysis.
Of further interest is the fact that Stucky et al. (1996) provided additional revisions to certain other taxa in the Sand Wash Basin faunal list of West and Dawson (1975). The specimen from Locality 3 originally referred to the rhinocerotoid Hyrachyus modestus Leidy, 1870, represents either Hyrachyus eximus Leidy, 1871, or Uintaceras, with Hyrachyus eximus known to have its HRD in the lower part of the Foggy Day beds of the Tepee Trail Formation (Ui1b, Chron C20r) and Uintaceras to have its LRD near the base of the Holy City beds (Ui1b) of the Tepee Trail Formation and the Uinta B1 (Ui1b) of the Uinta Formation in the eastern Uinta Basin. The specimen from Locality 3 originally referred to the dichobunid Homacodon sp., cf. H. vagans Marsh, 1872a, actually represents the homacodont Hylomeryx sp., a genus with its LRD in the Whistler Squat Local Fauna (Ui1b, Chron C20r) of the Devil's Graveyard Formation, but also known from as late as the Ui3 in the Uinta Formation (Myton Member). There is also a new record of Protoreodon sp. from Locality 3. The LRD for this genus is otherwise in the Poway Fauna of the Friars Formation (Ui1b) and the Uinta B1 (Ui1b) of the Uinta Formation in the eastern Uinta Basin. New records of oromerycid Oromeryx Marsh, 1894, were recorded from Localities 9 and 12. The LRD for this genus is otherwise in the Uinta B (Ui2, Chron C20r) of the Uinta Formation. Although Stucky et al. (1996) used the faunal list of West and Dawson (1975) for the brontotheres, Mihlbachler (2008) revised the brontothere taxa from the Sand Wash Basin as follows: 1) Tanyorhinus blairi Cook, 1926, and Tanyorhinus bridgeri Cook, 1926, = Sphenocoelus uintensis Osborn, 1895; 2) Telmatherium accola Cook, 1926, and Telmatherium advocata Cook, 1926 = Metarhinus sp.; 3) a nomen dubium assignment for Tanyorhinus harundivorax Cook, 1926, Manteoceras foris Cook, 1926, and Manteoceras pratensis Cook, 1926 (all generically and specifically indeterminate); and 4) Tanyorhinus sp. = indeterminate brontothere. Mihlbachler (2008) also recorded the occurrence of a species closely related to, but slightly larger than, Mesatirhinus junius (Leidy, 1872) from the Sand Wash Basin. The LRD of Sphenocoelus Osborn, 1895, is otherwise in the Adobe Town middle subunit of the Washakie Formation (Ui1b) in the Washakie Basin. The LRD of Metarhinus is otherwise in the Adobe Town middle subunit (Ui1b) of the Washakie Basin. The LRD of Mesatirhinus junius is undetermined because the holotype specimen's provenance is unknown, but it has been confidently recorded from the upper Bridger Formation (Br3) in the Bridger Basin and the Kinney Rim Member and lower subunit of the Adobe Town Member of the Washakie Formation (McCarroll et al., 1996; Mihlbachler, 2008). Mihlbachler (2008) also described a new brontothere, Wickia brevirhinus, based on a partial skull from the Sand Wash Basin and a referred specimen from above bed 17 of Granger (1909) in the Adobe Town middle subunit (Ui1b) of the main Washakie Basin. The LRD for Wickia is otherwise from the Roll the Bones Locality (Ui1a) of the TBM, Bridger Formation (Murphey and Kelly, 2017). Thus, only Sphenocoelus uintensis, Wickia brevirhinus, Metarhinus sp., and Mesatirhinus sp. are confidently known from the Sand Wash Basin. The dinocerate Eobasileus cornutus (Cope, 1872) is reported from Localities 11 and 13 in the Sand Wash Basin (West and Dawson, 1975). The LRD of Eobasileus Cope, 1872, is otherwise in the Adobe Town middle subunit of the Washakie Formation (?Ui1b) in the Washakie Basin, but is also known from the Uinta B (Ui2) in the western Uinta Basin and the Uinta B2 of the eastern Uinta Basin (Ui2). The primate Notharctus robustior Leidy, 1872, was reported to occur at Locality 3 in the Sand Wash Basin (West and Dawson, 1975), a Bridgerian hold over species that is also known from Adobe Town middle subunit (Ui1b) of the Washakie Formation in the main Washakie Basin and the Turtle Bluff Member (Ui1a) of the Bridger Formation (McCarroll et al., 1996a; Kelly and Murphey, 2016a). West and Dawson (1975) referred specimens from Localities 2, 3, and 12 in the Sand Wash Basin to the hyposodontid Hyopsodus despicians Matthew, 1909, whereas McCarroll et al. (1996) referred these specimens to Hyopsodus markmani Abel and Cook, 1925, without explanation. Previously, Gazin (1968) regarded Hyopsodus markmani as a synonym of Hyopsodus despicians, whereas West (1979) regarded H. markmani as a synonym of Hyopsodus paulus Leidy, 1870. Regardless of which final species diagnosis is accepted for the Sand Wash Basin Hyopsodus, the chronologic range of Hyopsodus Leidy, 1870, extends from the Wasatchian (early Eocene) to the Chadronian (late Eocene), so the occurrence of the genus cannot be used to separate Bridgerian, Ui1a, and Ui1b faunas (Gazin, 1968; Archibald, 1998). West and Dawson (1975) reported the occurrence of Viverravus minutus Wortman, 1901, from Locality 3, Viverravus sp. from Locality 20, and the miacid Uintacyon vorax Leidy, 1873, from Locality 4. Viverravus minutus is also known from the early Bridgerian (Br1b through Br2) and the genus has a chronologic range of the Clarkforkian through the late Uintan, Ui3 (Flynn, 1998). Uintacyon vorax is also known from the early Bridgerian Powder Wash Fauna of the Green River Formation and the Black's Fork Member of the Bridger Formation (Br2). Uintacyon has a long chronologic range in the western interior, being reported from the Clarkforkian (Ck2) through the Duchesnean (Flynn, 1998).
The above faunal evidence suggests that the fauna from Locality 20 is late Bridgerian in age, and the localities that occur well above Locality 20, or at least part of the fauna from these localities in the Sand Wash Basin, may be Ui1b in age. However, the question arises whether there could be a combination of Ui1a and Ui1b faunas from the Sand Wash Basin. Before assigning the entire or any part of the main Sand Wash fauna from these localities to a biochron or subage, the relative stratigraphic positions of all of the Sand Wash Basin localities are sorely needed along with a detailed account of the systematic paleontology justifying the taxonomic assignments. Without this information, any faunal correlations or interpretations of the magnetostratigraphy of the Sand Wash Basin are highly speculative.
 In the Tepee Trail Formation of the Absaroka Range, Wyoming (= East Fork Basin of Flynn, 1986, and Tsukui, 2016), the "Holy City beds" (HCB) yielded a fauna with Amynodon (LSD near middle of HCB), Epihippus, Hyrachyus eximus, Uintaceras (LSDs near base of HCB), and Hyopsodus sp., cf. H. paulus (Eaton, 1985; Walsh,1996a), indicating that this interval represents a Ui1b fauna (Gunnell et al., 2009). The occurrence of Achaenodon at locality UCMP V-78003, Metarhinus at locality UCMP V-77022, Hyrachyus eximus at locality UCMP V-77022 and Hyopsodus sp., cf. H. paulus at localities UCMP V-78008 and V-78012, also indicates a Ui1b age for about the lower three-quarters of the "Foggy Day beds." Therefore, the Ui1b faunal interval in the Tepee Trail Formation appears to span from about the middle of Chron C21n to somewhere within Chron C20r (Figure 18). There are no known mammal fossils from the "lower" part of the Tepee Trail Formation (= about lower half of Chron C21n), so its age is undetermined. An unconformity separates the Blue Point Marker bed (BPM) from the overlying "lower" part of the Tepee Trail Formation (Sundell et al., 1984; Eaton, 1985). The BPM has yielded a Bridgerian fauna (?Br3) from Locality UCMP V-78001 (Eaton, 1982; Sundell et al., 1984; Walsh, 1996a). Thus, no Ui1a faunas can be confidently recognized from the Tepee Trail Formation (Figure 18).
In the Tepee Trail Formation of the Absaroka Range, Wyoming (= East Fork Basin of Flynn, 1986, and Tsukui, 2016), the "Holy City beds" (HCB) yielded a fauna with Amynodon (LSD near middle of HCB), Epihippus, Hyrachyus eximus, Uintaceras (LSDs near base of HCB), and Hyopsodus sp., cf. H. paulus (Eaton, 1985; Walsh,1996a), indicating that this interval represents a Ui1b fauna (Gunnell et al., 2009). The occurrence of Achaenodon at locality UCMP V-78003, Metarhinus at locality UCMP V-77022, Hyrachyus eximus at locality UCMP V-77022 and Hyopsodus sp., cf. H. paulus at localities UCMP V-78008 and V-78012, also indicates a Ui1b age for about the lower three-quarters of the "Foggy Day beds." Therefore, the Ui1b faunal interval in the Tepee Trail Formation appears to span from about the middle of Chron C21n to somewhere within Chron C20r (Figure 18). There are no known mammal fossils from the "lower" part of the Tepee Trail Formation (= about lower half of Chron C21n), so its age is undetermined. An unconformity separates the Blue Point Marker bed (BPM) from the overlying "lower" part of the Tepee Trail Formation (Sundell et al., 1984; Eaton, 1985). The BPM has yielded a Bridgerian fauna (?Br3) from Locality UCMP V-78001 (Eaton, 1982; Sundell et al., 1984; Walsh, 1996a). Thus, no Ui1a faunas can be confidently recognized from the Tepee Trail Formation (Figure 18).
The Poway Fauna from the Friars Formation of the greater San Diego area, California, includes the LSD for Amynodon near the base of the formation, and the fauna is regarded as Ui1b based on this fact plus the other taxa in the fauna (Figure 19). The Mesa Drive Local Fauna of member B of the Santiago Formation is Ui1b in age, and the Murray Canyon Local Fauna of the Stadium Conglomerate is probably Ui1b in age (Walsh, 1996b). These faunas span the upper part of Chron C21n through a portion of Chron C20r (Walsh, 1996a, 1996b; Walsh et al., 1996). The Black's Beach Local Fauna consists of an indeterminate uintathere and an indeterminate brontothere, which are not age diagnostic and the fauna could represent either a Ui1a or late Bridgerian age. The faunas from the Santiago Formation of San Diego and Orange Counties span the Ui1b through the Duchesnean (Walsh, 1996b; Kelly, 2015; Kelly and Murphey, 2016b). The only confirmed Bridgerian fauna from the greater San Diego area is the Swami's Point Local Fauna (Walsh, 1996a, Miyata and Deméré, 2016) from the Delmar Formation, which occurs in Chron C21r. Thus, no Ui1a faunas can be confidently recognized from the greater San Diego area.
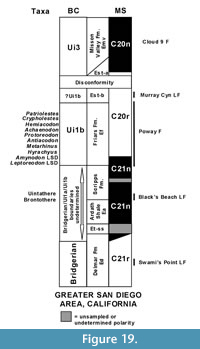 To summarize, Tsukui's (2016) magnetostratigraphy of the Bridger Formation is the most reliable for placement of the Bridgerian/Uintan boundary, which is in the lower part of Chron C21n (Figure 10). The Ui1a TBM Fauna occurs in the lower part of Chron C21n with two 206Pb/238U dates, 47.31 ± 0.06 Ma and 46.94 ± 0.14 Ma, directly associated with the fauna in the stratotype section. The Ui1a Basal Tertiary Local Fauna of Devil's Graveyard Formation also occurs in Chron C21n (Walton, 1992; Prothero, 1996b). Based on faunal correlations, no Ui1a faunas can be confidently recognized from the Uinta Formation, the Washakie Formation of the main Washakie Basin, the Tepee Trail Formation, and the greater San Diego area. The fauna from Locality 20 in the Washakie Formation of the Sand Wash Basin appears to be late Bridgerian in age. An age assignment for the fauna (or any part of the fauna) from all the other localities that occur stratigraphically much higher in the Washakie Formation of the Sand Wash Basin cannot be determined with confidence until the relative stratigraphic levels of the localities are determined and a detailed account of the systematic paleontology of the fauna is documented. The Ui1b Whistler Squat Quarry (local fauna) of the Devil's Graveyard Formation occurs within the lower part of Chron C20r and is 40Ar/39Ar dated at 45.04 ± 0.10 Ma and 44.88 ± 0.04 Ma for the lower tuff and upper tuff, respectively (Walton, 1992; Prothero, 1996; Campisano et al., 2014). In most other correlated sections, Ui1b faunas appear to occur from somewhere near the middle to the upper part of Chron C21n to somewhere between about the middle to the upper part of Chron C20r. The "Shoshonian" of Flynn (1986) includes Ui1a and Ui1b faunas as now understood (Gunnell et al., 2009), and investigators need to differentiate these two biochrons based on faunal compositions in order to make accurate faunal correlations. Investigators also need to recognize that biochrons do not necessarily have to be isochronous throughout North America with some possibly being time transgressive (Clemens, 2010; Kelly, 2014).
To summarize, Tsukui's (2016) magnetostratigraphy of the Bridger Formation is the most reliable for placement of the Bridgerian/Uintan boundary, which is in the lower part of Chron C21n (Figure 10). The Ui1a TBM Fauna occurs in the lower part of Chron C21n with two 206Pb/238U dates, 47.31 ± 0.06 Ma and 46.94 ± 0.14 Ma, directly associated with the fauna in the stratotype section. The Ui1a Basal Tertiary Local Fauna of Devil's Graveyard Formation also occurs in Chron C21n (Walton, 1992; Prothero, 1996b). Based on faunal correlations, no Ui1a faunas can be confidently recognized from the Uinta Formation, the Washakie Formation of the main Washakie Basin, the Tepee Trail Formation, and the greater San Diego area. The fauna from Locality 20 in the Washakie Formation of the Sand Wash Basin appears to be late Bridgerian in age. An age assignment for the fauna (or any part of the fauna) from all the other localities that occur stratigraphically much higher in the Washakie Formation of the Sand Wash Basin cannot be determined with confidence until the relative stratigraphic levels of the localities are determined and a detailed account of the systematic paleontology of the fauna is documented. The Ui1b Whistler Squat Quarry (local fauna) of the Devil's Graveyard Formation occurs within the lower part of Chron C20r and is 40Ar/39Ar dated at 45.04 ± 0.10 Ma and 44.88 ± 0.04 Ma for the lower tuff and upper tuff, respectively (Walton, 1992; Prothero, 1996; Campisano et al., 2014). In most other correlated sections, Ui1b faunas appear to occur from somewhere near the middle to the upper part of Chron C21n to somewhere between about the middle to the upper part of Chron C20r. The "Shoshonian" of Flynn (1986) includes Ui1a and Ui1b faunas as now understood (Gunnell et al., 2009), and investigators need to differentiate these two biochrons based on faunal compositions in order to make accurate faunal correlations. Investigators also need to recognize that biochrons do not necessarily have to be isochronous throughout North America with some possibly being time transgressive (Clemens, 2010; Kelly, 2014).
Revision of Age Qualifiers for Uintan Biochrons
The North American Land Mammal ages were first established by the Wood Committee (Wood et al., 1941). Subsequently, investigators provided further refinements and subdivisions of the North American Land Mammal ages (e.g., Rensberger, 1971; Fisher and Rensberger, 1972; Lindsay, 1972; Krishtalka et al., 1987; Woodburne, 1987). As noted above, Flynn (1986) proposed the "Shoshonian Subage" of the Uintan for certain older faunas that appeared transitional between typical Bridgerian and Uintan faunas. Krishtalka et al. (1987) divided the Uintan faunally into early and late segments. Woodburne (2004b) and Robinson et al. (2004) recognized two Uintan biochrons, the Ui2 and Ui3, and provisionally recognized a third earliest Uintan biochron, the Ui1, which they questionably equated with the "Shoshonian" of Flynn (1986). Gunnell et al. (2009) proposed the division of Ui1 into two biochrons, the Ui1a and Ui1b, with faunas from the former regarded as earliest Uintan in age. After Robinson et al. (2004) but prior to Gunnell et al. (2009), Ui1, Ui2, and Ui3 faunas were generally referred to as earliest, early and late Uintan faunas, respectively. However, with the recognition that Ui1a faunas are older and faunally distinctive from Ui1b faunas, the problem of what informal age qualifier is used with each biochron needs revision. To clarify the age qualifiers and attempt to avoid future confusion in the literature, we propose the following descriptive age terms for each Uintan biochron: 1) Ui1a as earliest Uintan; 2) Ui1b as early Uintan; 3) Ui2 as medial (or middle) Uintan; and 4) Ui3 as late Uintan.
Faunal Characterization of Biochrons Ui1a and Ui1b
Many investigators have discussed in detail the concerns and problems of using fossil datum events in defining biochrons and biostratigraphic zones (e.g., Berggren and Van Couvering, 1974; Opdyke et al., 1977; Lindsay et al., 1987; Lindsay and Tedford, 1990; Aubry, 1995; Berggren et al., 1995; Walsh, 1998; Woodburne, 2004a, 2004b). Here we follow Gunnell et al. (2009) and Campisano et al. (2014) in the use of the following temporal and biostratigraphic terms: 1) LRD (lowest range datum) defined as the first appearance (= oldest record) of a taxon among its fossil occurrences in a large geographic region, either by origin or immigration; 2) HRD (highest range datum) defined as the last occurrence (= youngest record) of a taxon among its fossil occurrences in a large geographic region, either by emigration or extinction; 3) LSD (lowest stratigraphic datum) defined as the lowest stratigraphic occurrence of a taxon within a given section; and 4) HSD (highest stratigraphic datum) defined as the highest stratigraphic occurrence of a taxon within a given section. Lindsay et al. (1987), Walsh (1998), and Woodburne (2004a) recognized that LRD and HRD are not necessarily synchronous with LSD and HSD, respectively. However, if a taxon's LSD or HSD within a given section can be determined by paleomagnetic data and/or radioisotopic dates to be the earliest or latest record of that taxon, then that LSD or HSD becomes equivalent (synchronous) to its LRD or HRD, respectively (Woodburne, 2004a). Some investigators have advocated the use of a single taxon LRD to define the beginning of a mammalian biochron (e.g., Woodburne, 1977, 1987; Flynn, 1986; Walsh, 1996a). Flynn (1986) and Walsh (1996a) used the LRD of Amynodon to define the beginning of the Uintan. However, there can be inherent problems with using a single taxon LRD due to paleoenvironmental, taphonomic, and sampling biases (Lindsay et al., 1987; Walsh, 1998). Nevertheless, Amynodon is a widespread taxon, known from both the western interior, the Gulf Coast and the Pacific Coast of North America, so its LSD within a given section can still be useful if that section has been placed in a temporal context by paleomagnetic data and/or radioisotopic dates. The use of LRDs of multiple taxa to define a biochron or land mammal age boundary can help to alleviate the problem of knowing whether a single taxon's LSD and LRD are actually isochronous over a wide geographical area. Index taxa are also useful in defining a biochron because they are restricted to a single biochron. However, often index taxa are at the species level and are not widespread geographically, but endemic to a particular formation or small geographic area. Even considering these concerns, the most practical method to assign a fauna to a biochron age is to use the LRDs of multiple taxa and the occurrence of index taxa (Robinson et al., 2004; Gunnell et al., 2009; Campisano et al., 2014).
Our proposals for the faunal characterizations of biochrons Ui1a and Ui1b are presented in Table 4 and Table 5. Table 4 is a revised version of table 5 of Murphey and Kelly (2017), and Table 5 is a revised version of table 7 of Gunnell et al. (2009).
CONCLUSIONS
T  he Turtle Bluff Member of the Bridger Formation exposed on the southwest flank of Cedar Mountain, Uinta County, Wyoming, is the stratotype section for the earliest Uintan, biochron Ui1a, of the Uintan North American Land Mammal age (Gunnell et al., 2009). With this third report along with those of Kelly and Murphey (2016a) and Murphey and Kelly (2017), detailed systematic accounts of all known taxa comprising the Ui1a TBM Fauna are now complete, and their stratigraphic distributions within the TBM are well defined (Figure 20). These studies provide a much greater understanding of the faunal transition between the Bridgerian and Uintan North American Land Mammal ages. The Ui1a TBM Fauna is characterized by the following (Table 4): 1) the occurrence of at least five index species, and probably more; 2) the first appearances of at least 16 species; 3) the first appearances of at least five genera (the rodent Metanoiamys, the brontothere Wickia, the hyracodont Triplopus, the equid Epihippus, and the oromerycid Merycobunodon) and 4) the occurrence of 29 Bridgerian (Br2 and/or Br3) hold over taxa. With the exception of Hemiacodon engardae, all other Primates from the TBM are Bridgerian hold over taxa, whereas a moderate diversification occurred in the rodents and lipotyphlans from the TBM (Kelly and Murphey, 2016a; Murphey and Kelly, 2017). Of the four marsupials documented here from the TBM, three are also known from the Bridgerian, but one (Peradectes californicus), whose previous LRD was in the Ui1b Poway Fauna from the Friars Formation of the greater San Diego area, has a new record for its LRD at Locality SDSNH 5844 in the TBM. Based on our reevaluation and revised characterizations of Ui1a and Ui1b faunas, currently only two Ui1a faunas can be confidently recognized from North America, the TBM Fauna and the Basal Tertiary Local Fauna from the Devil's Graveyard Formation of Texas. Because of the prior confusion in the literature regarding the term "earliest Uintan," we propose that it only be applied to faunas that can defined faunally as Ui1a in age. We also propose that the informal age qualifiers of early Uintan, medial Uintan, and late Uintan be applied to biochrons Ui1b, Ui2, and Ui3, respectively.
he Turtle Bluff Member of the Bridger Formation exposed on the southwest flank of Cedar Mountain, Uinta County, Wyoming, is the stratotype section for the earliest Uintan, biochron Ui1a, of the Uintan North American Land Mammal age (Gunnell et al., 2009). With this third report along with those of Kelly and Murphey (2016a) and Murphey and Kelly (2017), detailed systematic accounts of all known taxa comprising the Ui1a TBM Fauna are now complete, and their stratigraphic distributions within the TBM are well defined (Figure 20). These studies provide a much greater understanding of the faunal transition between the Bridgerian and Uintan North American Land Mammal ages. The Ui1a TBM Fauna is characterized by the following (Table 4): 1) the occurrence of at least five index species, and probably more; 2) the first appearances of at least 16 species; 3) the first appearances of at least five genera (the rodent Metanoiamys, the brontothere Wickia, the hyracodont Triplopus, the equid Epihippus, and the oromerycid Merycobunodon) and 4) the occurrence of 29 Bridgerian (Br2 and/or Br3) hold over taxa. With the exception of Hemiacodon engardae, all other Primates from the TBM are Bridgerian hold over taxa, whereas a moderate diversification occurred in the rodents and lipotyphlans from the TBM (Kelly and Murphey, 2016a; Murphey and Kelly, 2017). Of the four marsupials documented here from the TBM, three are also known from the Bridgerian, but one (Peradectes californicus), whose previous LRD was in the Ui1b Poway Fauna from the Friars Formation of the greater San Diego area, has a new record for its LRD at Locality SDSNH 5844 in the TBM. Based on our reevaluation and revised characterizations of Ui1a and Ui1b faunas, currently only two Ui1a faunas can be confidently recognized from North America, the TBM Fauna and the Basal Tertiary Local Fauna from the Devil's Graveyard Formation of Texas. Because of the prior confusion in the literature regarding the term "earliest Uintan," we propose that it only be applied to faunas that can defined faunally as Ui1a in age. We also propose that the informal age qualifiers of early Uintan, medial Uintan, and late Uintan be applied to biochrons Ui1b, Ui2, and Ui3, respectively.
The new 206 Pb/ 238 U dates from the TBM, 47.31 ± 0.06 Ma from the tuffaceous white sandstone and 46.94 ± 0.14 Ma from the Roll the Bones locality (Locality SDSNH 5844), provide precise age constraints for the TBM Fauna and, along with the new paleomagnetic data, further support the proposal of Tsukui (2016) that the Uintan-Bridgerian boundary occurs in the lower part of Chron C21n of the GPTS.
ACKNOWLEDGEMENTS
We thank the U.S. Bureau of Land Management for supporting our research by providing permits to complete our field work over the years. We are grateful to B. Breithaupt of the Wyoming BLM for facilitating the curation of TBM specimens at the DMNS. We are greatly indebted to T.A. Deméré and K.A. Randall of the SDNHM, L. Ivy and J. Sertich of the DMNS, T. Culver and J. Eberle of the UCM, and J. Ming, R. O'Leary, and A. Gishlick of the AMNH, for their considerate assistance in procuring loans of specimens for this study and assistance with locality and specimen numbers. Special thanks are extended to: P. Robinson, E. Evanoff, P. Monaco, the late D. Engard, and all of the UCM students and volunteers who assisted with the recovery of the fossils from UCM Locality 92189; P. Sena, M. Madsen, S. Madsen, H. Finalyson, and T. Temme, who assisted with quarrying and hauling and wet screening of matrix samples of the more recently discovered TBM localities; and F. Alshamsi for help with paleomagnetic data collection. K. Chamberlain was partially supported by Mega-Grant 14.Y26.31.0012 of the government of the Russian Federation. Our greatest gratitude goes to our late friend and colleague S.L. Walsh of the SDNMH for his participation in the TBM field work, wet screening, heavy liquid separation, and fossil identifications; as well as his many contributions to middle Eocene mammalian paleontology, biostratigraphy, and biochronology.
REFERENCES
Abel, O. and Cook, H.J. 1925. A preliminary study of early mammals in a new fauna from Colorado. Proceedings of the Colorado Museum of Natural History, 5:33-36.
Archibald, J.D. 1998. Archaic ungulates ("Condylarthra"), p. 292-329. In Janis, C.M., Scott, K.M., and Jacobs, L.L. (eds.), Evolution of Tertiary Mammals of North America, Volume 1: Terrestrial Carnivores, Ungulates and Ungulatelike Mammals. Cambridge University Press, New York.
Asher, R.J., McKenna, M.C., Emry, R.J., Tabrum, A.R., and Kron, D.G. 2002. Morphology and relationships of Apternodus and other extinct, zalambdodont, placental mammals. Bulletin of the American Museum of Natural History, 273:1-117. https://doi.org/10.1206/0003-0090(2002)273%3C0001:MAROAA%3E2.0.CO;2
Aubry, M.-P. 1995. From chronology to stratigraphy: interpreting the lower and middle Eocene stratigraphic record in the Atlantic Ocean, p. 213-274. In Berggren, W.A., Kent, D.V., and Hardenbol, J. (eds.), Geochronology, Time Scales and Global Stratigraphic Correlations: A United Temporal Framework for an Historical Geology. Society of Economic Paleontologists and Mineralogists Special Publication 54. https://doi.org/10.2110/pec.95.04.0213
Aymard, A. 1850. Compte rendu de la séance du 12 Avril 1849, réponse à M. Robert sur les mammifères fossiles des calcaires du Puy. Annales de la Société d'Agriculture, Sciences, Arts et Commerce du Puy, 14:80-86. (In French)
Beard, K.C. and Dawson, M.R. 2001. Early Wasatchian mammals from the Gulf Coastal Plain of Mississippi: biostratigraphic and paleobiogeographic implications, p. 75-94. In Gunnell, G.F. (ed.), Eocene Biodiversity: Unusual Occurrences and Rarely Sampled Habitats. Kluwer Academic/Plenum Press, New York. https://doi.org/10.1007/978-1-4615-1271-4
Berggren, W.A., Kent, D.V., Swisher III, C.C., and Aubry, M-P. 1995. A revised Cenozoic geochronology and chronostratigraphy, p. 129-212. In Berggren, W.A., Kent, D.V., and Hardenbol, J. (eds.), Geochronology, Time Scales and Global Stratigraphic Correlations: a Unified Temporal Framework for an Historical Geology. Society of Economic Paleontologist and Mineralogists, Special Publication 54. https://doi.org/10.2110/pec.95.04.0129
Berggren, W.A. and Van Couvering, J.A. 1974. The late Neogene: biostratigraphy, geochronology and paleoclimatology of the last 15 million years in marine and continental sequences. Palaeogeography, Palaeoclimatology, Palaeoecology, 16:1-216. https://doi.org/10.1016/0031-0182(74)90032-7
Bown, T.M. 1979. Geology and mammalian paleontology of the Sand Creek facies, lower Willwood Formation (lower Eocene), Washakie County, Wyoming. Geological Survey of Wyoming Memoir, 2:1-151.
Bown, T.M. 1980. The Willwood Formation (lower Eocene) of the southern Bighorn Basin, Wyoming, and its mammalian faunas, p. 127-133. In Gingerich, P.D. (ed.), Early Cenozoic Paleontology and Stratigraphy of the Big Horn Basin, Wyoming. University of Michigan Papers on Paleontology, 24.
Bown, T.M. 1982. Geology, paleontology, and correlation of Eocene volcaniclastic rocks, southeast Absaroka Range, Hot Springs County, Wyoming. USGS Professional Paper, 1201-A:1-76.
Bown, T.M., Rose, K.D., Simons, E.L., and Wing, L.S. 1994. Distribution and stratigraphic correlation of upper Paleocene and lower Eocene fossil mammal and plant localities of the Fort Union, Willwood, and Tatman Formations, southern Bighorn Basin, Wyoming. USGS Professional Paper, 1540:1-103.
Campisano, C.J., Kirk, E.C., Townsend, K.E.B., and Deino, A.L. 2014. Geochronological and taxonomic revisions of the middle Eocene Whistler Squat Quarry (Devil's Graveyard Formation, Texas) and implications for the early Uintan in Trans-Pecos Texas. https://doi.org/10.1371/journal.pone.0101516
Case, J.A., Goin, F.J., and Woodburne, M.O. 2005. "South American" marsupials from the late Cretaceous of North America and the origin of marsupial cohorts. Journal of Mammalian Evolution, 12:461-494. https://doi.org/10.1023/b:jomm.0000047339.39630.82
Cashion, W.D. 1982. Geologic map of the Bonanza Quadrangle, Uintah County, Utah. USGS Miscellaneous Field Studies Map, MF-1895, scale 1:24,000.
Clemens, W.A. 2006. Early Paleocene (Puercan) peradectid marsupials from northeastern Montana, North American western interior. Palaeontographica Abteilung A, Palaozoologie-Stratigraphie, 277:19-31.
Clemens, W.A. 2010. Were immigrants a significant part of the earliest Paleocene mammalian fauna of the North American interior. Vertebrata PalAsiatica, 48:285-307.
Cook, H.J. 1926. New Eocene titanotheres from Moffat County, Colorado. Proceedings of the Colorado Museum of Natural History, 7:12-18.
Cope, E.D. 1872. Notices of new vertebrata from the upper waters of Bitter Creek, Wyoming Territory. Proceedings of the American Philosophical Society, 12:483-486.
Cope, E.D. 1873a. Third notice of extinct Vertebrata from the Tertiary of the Plains. Palaeontological Bulletin, 16:1-8.
Cope, E.D. 1873b. Fourth notice of extinct Vertebrata from the Bridger and Green River Territories. Palaeontological Bulletin, 17:1-4.
Cope, E.D. 1873c. Synopsis of New Vertebrata from the Tertiary of Colorado, Obtained during the Summer of 1873. U.S. Government Printing Office, Washington, D.C.
Cope, E.D. 1880. A new genus of tapiroids. American Naturalist, 14:382-383.
Cope, E.D. 1884. The Vertebrata of the Tertiary formations of the West. Report of the U.S. Geological Survey of the Territories, F.V. Hayden, Geologist in Charge. U.S. Government Printing Office, Washington, D.C.
Corfu, F., Hanchar, J.M, Hoskin, P.W.O, and Kinny, P. 2003. Chapter 16, atlas of zircon morphologies, p. 469-500. In Hanchar, J.M. and Hoskin, P.W.O (eds.), Zircon. Reviews in Mineralogy and Geochemistry, Volume 53.
Crochet, J.-Y. 1977. Les Didelphidae (Marsupicarnivora, Marsupialia) holarctiques tertiaires. Comptes Rendus Hebdomadaires des Séances de L'Académie des Sciences, Paris, Séries D, 24:357-360. (In French)
Crochet, J.-Y. 1979. Diversité systématique des Didelphidae (Marsupiala) européens tertiaires. Geobios, 12:365-378. (In French) https://dx.doi.org/10.1016/S0016-6995(79)80117-5
Crochet, J.-Y. 1980. Les marsupiaux du Tertiaire d'Europe. Éditions de la Fondation. Singer-Polignac, Paris, France. (In French)
Eaton, J.G. 1982. Paleontology and correlation of Eocene volcanic rocks in the Carter Mountain area, Park County, southeastern Absaroka Range, Wyoming. Contributions to Geology, University of Wyoming, 21:153-194.
Eaton, J.G. 1985. Paleontology and correlation of the Eocene Tepee Trail and Wiggins formations in the north fork of Owl Creek area, southeastern Absaroka Range, Hot Springs County, Wyoming. Journal of Vertebrate Paleontology, 5:345-370. https://doi.org/10.1080/02724634.1985.10011871
Eberle, J. and Storer, J.E. 1995. Herpetotherium valens (Lambe), a didelphid marsupial from the Calf Creek Local Fauna (Chadronian), Saskatchewan. Journal of Vertebrate Paleontology, 15:785-794. https://doi.org/10.1080/02724634.1995.10011261
Evanoff, E., Robinson, P., Murphey, P.C., Kron, D.G., Engard, E., and Monaco, P. 1994. An early Uintan fauna from Bridger E. Society of Vertebrate Paleontology, Abstracts with Programs Supplement, 14:24A.
Fisher, R.A., 1953, Dispersion on a sphere. Proceedings of the Royal Society (London), A217:295-305. https://doi.org/10.1098/rspa.1953.0064
Fisher, R.V. and Rensberger, J.M. 1972. Physical stratigraphy of the John Day Formation, Oregon. University of California, Publications in the Geological Sciences, 101:1-95.
Flynn, J.J. 1986. Correlation and geochronology of middle Eocene strata from the western United States. Palaeogeography, Palaeoclimatology, Palaeoecology, 55:335-406. https://doi.org/10.1016/0031-0182(86)90155-0
Flynn, J.J. 1998. Early Cenozoic Carnivora ("Miacoidea"), p. 110-123. In Janis, C.M., Scott, K.M., and Jacobs, L.L. (eds.), Evolution of Tertiary mammals of North America, Volume 1: Terrestrial Carnivores, Ungulates and Ungulatelike Mammals. Cambridge University Press, New York.
Fox, R.C. 1983. Notes on the North American Tertiary marsupials Herpetotherium and Peradectes. Canadian Journal of Earth Sciences, 20:1565-1578. https://doi.org/10.1139/e83-146
Galbreath, E.C. 1953. A contribution to the Tertiary geology and paleontology of northeastern Colorado. University of Kansas Paleontological Contributions, Vertebrata, 4:1-120.
Gazin, C.L. 1952. The lower Eocene Knight Formation of western Wyoming and its mammalian faunas. Smithsonian Miscellaneous Collections, 117:1-82.
Gazin, C.L. 1955. A review of the Upper Eocene Artiodactyla of North America. Smithsonian Miscellaneous Collections, 281:1.
Gazin, C.L. 1956. Paleocene mammalian faunas of the Bison Basin in south-central Wyoming. Smithsonian Miscellaneous Collections, 131:1-57.
Gazin, C.L. 1962. A further study of the lower Eocene mammalian faunas of southwestern Wyoming. Smithsonian Miscellaneous Collections, 144:1-98.
Gazin, C.L. 1968. A study of the Eocene condylarthran mammal Hyopsodus. Smithsonian Miscellaneous Collections, 153:1-90.
Gazin, C.L. 1976. Mammalian faunal zones of the Bridger middle Eocene. Smithsonian Contributions to Paleobiology, 26:1-25. https://dx.doi.org/10.5479/si.00810266.26.1
Gill, T. 1872. Arrangement of the families of mammals with analytical tables. Smithsonian Miscellaneous Collections, 11:1-98. https://doi.org/10.5962/bhl.title.14607
Golz, D.J. 1976. Eocene Artiodactyla of southern California. Natural History Museum of Los Angeles County Science Bulletin, 26:1-85.
Granger, W.A. 1908. A revision of the American Eocene horses. Bulletin of the American Museum of Natural History, 24:221-264.
Gray, J.E. 1821. On the natural arrangement of vertebrose animals. London Medical Repository, 15:269-310.
Gunnell, G.F. 1998. Mammalian fauna from the lower Bridger Formation (Bridger A, early middle Eocene) of the southern Green River Basin, Wyoming. Contributions from the Museum of Paleontology, University of Michigan, 30:83-130.
Gunnell, G.F., Bown, T.M., Bloch, J.I., and Boyer, D.M. 2008. "Proteutheria," p. 63-81. In Janis, C.M, Gunnell, G.F., and Uhen, M.E. (eds.), Evolution of Tertiary Mammals of North America, Volume 2, Small Mammals, Xenarthrans, and Marine Mammals. Cambridge, Cambridge University Press. https://doi.org/10.1017/CBO9780511541438
Gunnell, G.F., Murphey, P.C., Stucky, R.K., Townsend, K.E. Beth, Robinson, P., Zonneveld, J.-P., and Bartels, W.S. 2009. Biostratigraphy and biochronology of the latest Wasatchian, Bridgerian, and Uintan North American Land Mammals “Ages,” p. 279-330. In Albright, L. B. III (ed.), Papers on Geology, Vertebrate Paleontology, and Biostratigraphy in Honor of Michael O. Woodburne, Museum of Northern Arizona Bulletin 65, Flagstaff, Arizona.
Guthrie, D.A. 1971. The mammalian fauna of the Lost Cabin Member, Wind River Formation (lower Eocene) of Wyoming. Annals of Carnegie Museum, 43:47-113.
Hayes, F.G. 2005. Arikareean (Oligocene-Miocene) Herpetotherium (Marsupialia, Didelphidae) from Nebraska and Florida. Bulletin of the Florida Museum of Natural History, 45:341-360.
Holbrook, L.T. and Lucas, S.G. 1997. A new genus of rhinocerotoid from the Eocene of Utah and the status of North American "Forstercooperia." Journal of Vertebrate Paleontology, 17:384-396. https://doi.org/10.1080/02724634.1997.10010983
Horovitz, I., Martin, T., Bloch, J., Ladevèze, S., Kurz, C., and Sánchez-Villagra, M.R. 2009. Cranial anatomy of the earliest marsupials and the origin of opossums. PLOS ONE 4:e8278:1-9. https://doi.org/10.1371/journal.pone.0008278
Hoskin, P.W. and Schaltegger, U. 2003. The composition of zircon and igneous and metamorphic petrogenesis. Reviews in Mineralogy and Geochemistry, 53:27-62.
Hough, J.R. 1961. Review of Oligocene didelphid marsupials. Journal of Paleontology, 35:218-228.
Illiger, C. 1811. Prodromus systematis Mammalium et Avium additis terminis zoographicis utrudque classis. C. Salfeld, Berlin. (In German)
Janis, C.M. 1998. Introduction, p. 1-7. In Janis, C.M. and Jacobs, L.L. (eds.), Evolution of Tertiary Mammals of North America, Volume 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals. Cambridge, Cambridge University Press.
Johanson, Z. 1996. New marsupial from the Fort Union Formation, Swain Quarry, Wyoming. Journal of Paleontology, 70:1023-1031. https://doi.org/10.1017/S0022336000038725
Kelly, T.S. 1990. Biostratigraphy of Uintan and Duchesnean land mammal assemblages from the Middle Member of the Sespe Formation, Simi Valley, California. Contributions in Science, 419:1-42.
Kelly, T.S. 2010. New records of Marsupialia, Lipotyphla, and Primates from the Duchesnean (middle Eocene) Simi Valley Landfill Local Fauna, Sespe Formation, California. Paludicola, 7:158-169.
Kelly, T.S. 2013. New records of small mammals from the latest Uintan (middle Eocene) Strathearn Local Fauna, Sespe Formation, California. Paludicola, 9:56-69.
Kelly, T.S. 2014. Preliminary report on the mammals from Lane's Little Jaw Site Quarry: a latest Cretaceous (earliest Puercan?) local fauna, Hell Creek Formation, southeastern Montana. Paludicola, 10:50-91.
Kelly, T.S. 2015. Final report on the fossil mammals recovered during the paleontologic mitigation program for Rancho Mission Viejo planning area 2 (subarea 2.1 and 2.2), TT17561, Orange County, California. Proprietary report available from Archeological Resource Management Corporation, Rancho Palos Verdes, California, p. 24.
Kelly, T.S. and Murphey, P.C. 2016a. Mammals from the earliest Uintan (middle Eocene) Turtle Bluff Member, Bridger Formation, southwestern Wyoming, Part 1: Primates and Rodentia. Palaeontologia Electronica 19.2.24A:1-55. https://palaeo-electronica.org/content/2016/1518-earliest-uintan-mammals. https://doi.org/10.26879/586
Kelly, T.S. and Murphey, P.C. 2016b. New records of Epihippus (Equidae) and a rhinocerotoid perissodactyl from the middle Eocene Santiago Formation of southern California. Paludicola, 11:8-30.
Kelly, T.S., Murphey, P.C., and Walsh, S.L. 2012. New records of small mammals from the middle Eocene Duchesne River Formation, Utah, and their implications for the Uintan-Duchesnean North American Land Mammal Age transition. Paludicola, 8:194-237.
Kelly, T.S. and Whistler, D.P. 1994. Additional Uintan and Duchesnean (middle and late Eocene) mammals from the Sespe Formation, Simi Valley, California. Contributions in Science, 439:1-29.
Kihm, A.J. and Schumaker, K.K. 2015. Marsupials from the Chadronian (latest Eocene) Medicine Pole Hills Local Fauna, North Dakota. Paludicola, 10:93-112.
Kirschvink, J.L. 1980. The least-squares line and plane and the analysis of paleomagnetic data. Geophysical Journal of the Royal Astronomical Society, 62:699-718.
Kitts, D.B. 1957. A revision of the genus Orohippus (Perissodactyla, Equidae). American Museum Novitates, 1864:1-40.
Korth, W.W. 1994. Middle Tertiary marsupials (Mammalia) from North America. Journal of Paleontology, 68:376-397. https://doi.org/10.1017/S0022336000022952
Korth, W.W. 2008. Marsupialia, p. 39-47. In Janis, C.M, Gunnell, G.F., and Uhen, M.E. (eds.), Evolution of Tertiary Mammals of North America, Volume 2, Small Mammals, Xenarthrans, and Marine Mammals. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511541438
Korth, W.W. 2015. Mammals from the Blue Ash Local Fauna (late Oligocene), South Dakota. Additional specimens of Herpetotherium Cope (Marsupialia, Didelphidae). Paludicola, 10:113-118.
Krishtalka, L. 1976. North American Nyctitheriidae (Mammalia, Insectivora). Annals of Carnegie Museum, 46:7-28.
Krishtalka, L. and Stucky, R.K. 1983a. Revision of the Wind River faunas, early Eocene of central Wyoming. Part 3. Marsupialia. Annals of Carnegie Museum, 52:205-227.
Krishtalka, L. and Stucky, R.K. 1983b. Paleocene and Eocene marsupials of North America. Annals of Carnegie Museum, 52:229-263.
Krishtalka, L. and Stucky, R.K. 1984. Middle Eocene marsupials (Mammalia) from northeastern Utah and the mammalian fauna from Powder Wash. Annals of Carnegie Museum, 53:31-45.
Krishtalka, L., West, R.M., Black, C.C., Dawson, M.R., Flynn, J.J., Turnbull, W.D., Stucky, R.K., McKenna, M.C., Bown, T.M., Golz, D.J., and Lillegraven, J.A. 1987. Eocene (Wasatchian through Duchesnean) biochronology of North America, p.77-117. In Woodburne, M.O. (ed.), Cenozoic Mammals of North America. Berkeley, University of California Press.
Krogh, T.E. 1973. A low-contamination method for hydrothermal decomposition of zircon and extraction of U and Pb for isotopic age determinations. Geochimica et Cosmochimica Acta, 37:485-494. https://doi.org/10.1016/0016-7037(73)90213-5
Kuiper, K.F., Deino, A., Hilgen, F.J., Krijgsman, W., Renne, P.R., and Wijbrans, J.R. 2008. Synchronizing rock clocks of earth history. Science, 320:500-504. https://doi.org/10.1126/science.1154339
Ladevèze, S., Smith, R., and Smith, T. 2005. Reassessment of the morphology and taxonomic status of the earliest herpetotheriid marsupials of Europe. Journal of Mammalian Evolution, 12. https://doi.org/10.1007/s10914-012-9195-0
Lambe, L.M. 1908. The Vertebrata of the Oligocene of the Cypress Hills, Saskatchewan. Contributions to Canadian Palaeontology, 3:1-65. https://doi.org/10.5962/bhl.title.64818
Lavocat, A.R. 1951. Révision de la faune des mammifères oligocènes d'Auvergne et du Velay. Paris Éditions Sciences et Avenir, Paris, France. (In French)
Leidy, J. 1870. Remarks on a collection of fossils from the western Territories. Proceedings of the Academy of Natural Sciences, Philadelphia, 22:109-110.
Leidy, J. 1871. Remarks on fossil vertebrates from Wyoming. American Journal of Science, Series 3, 23:229-373.
Leidy, J. 1872. Remarks on fossils from Wyoming. Proceedings of the Academy of Natural Sciences, Philadelphia, 24:19-21.
Leidy, J. 1873. Contributions to the extinct vertebrate fauna of the western Territories, p.14-358. In Hayden, F.V. (ed.), Report of the United States Geological Survey of the Territories. Washington, DC, Department of the Interior. https://doi.org/10.5962/bhl.title.61805
Lillegraven, J.A. 1976. Didelphids (Marsupialia) and Uintasorex (?Primates) from later Eocene sediments of San Diego County, California. Transactions of the San Diego Society of Natural History, 18:85-112.
Lillegraven, J.A., McKenna, M.C., and Krishtalka, L. 1981. Evolutionary relationships of middle Eocene and younger species of Centetodon (Mammalia, Insectivora, Geolabididae) with a description of the dentition of Ankylodon (Adapisoricidae). University of Wyoming Publications, 45:1-115.
Lindsay, E.H. 1972. Small mammals from the Barstow Formation, California. University of California, Publications in the Geological Sciences, 93:1-104.
Lindsay, E.H., Opdyke, N.D., Johnson, N.M., and Butler, R.F. 1987. Mammalian chronology and the Magnetic Polarity Time Scale, p. 269-264. In Woodburne, M.O. (ed.), Cenozoic Mammals of North America. Berkeley, University of California Press.
Lindsay, E.H. and Tedford, R.H. 1990. Development and application of land mammal ages in North America and Europe, a comparison, p. 601-624. In Lindsay, E.H., Fahlbursch, V., and Mein, P. (eds.), European Neogene Mammal Chronology. NATO Advanced Science Institute Series 180. New York, Plenum Press. https://doi.org/10.1007/978-1-4899-2513-8_36
Ludwig, K.R. l988. PBDAT for MS-DOS, a computer program for IBM-PC compatibles for processing raw Pb-U-Th isotope data, version 1.24. U.S. Geological Survey, Open-File Report 88-542, 32 pp.
Ludwig, K.R. l991. ISOPLOT for MS-DOS, a plotting and regression program for radiogenic-isotope data, for IBM-PC compatible computers, version 2.75. U.S. Geological Survey, Open-File Report 91-445, 45 pp.
Ludwig, K. R. 1998. On the treatment of concordant uranium-lead ages. Geochimica et Cosmochimica Acta, 62:665-676. https://doi.org/10.1016/S0016-7037(98)00059-3
MacFadden, B. J. 1980. Eocene Perissodactyla from the type section of the Tepee Trail Formation of northwestern Wyoming. Contributions to Geology, University of Wyoming, 18:135-143.
Machlus, M.L., Ramezani, J., Bowring, S.A., Hemming, S.R., Tsukui, K., and Clyde, W.C. 2015. A strategy for cross-calibration U-Pb chronology and astrochronology of sedimentary sequences: an example from the Green River Formation, Wyoming, USA. Earth and Planetary Science Letters, 413:70-78.
Marsh, O. C. 1871. Notice of some new fossil mammals from the Tertiary formation. American Journal of Science, Series 3, 2:35-44. https://doi.org/10.2475/ajs.s3-2.7.35
Marsh, O.C. 1872a. Preliminary description of new Tertiary mammals. American Journal of Science, Series 3, 4:122-128. https://doi.org/10.2475/ajs.s3-4.20.122
Marsh, O.C. 1872b. Preliminary description of new Tertiary mammals. American Journal of Science, Series 3, 4:202-224. https://doi.org/10.2475/ajs.s3-4.21.202
Marsh, O.C. 1873. Notice of new Tertiary mammals. American Journal of Science, Series 3, 6:407-410. http://doi.org/10.2475/ajs.s3-5.29.407
Marsh, O.C. 1877. Notice of new vertebrate fossils. American Journal of Science, Series 3, 14:249-256. https://doi.org/10.2475/ajs.s3-14.81.249
Marsh, O. C. 1878. The introduction and succession of vertebrate life in America. Popular Science Monthly, 7:513-527, 672-697.
Marsh, O.C. 1894. Description of Tertiary artiodactyls. American Journal of Science, 48:259-274. https://doi.org/10.2475/ajs.s3-48.285.259
Marshall, L.G., Case, J.A., and Woodburne, M.O. 1990. Phylogenetic relationships of the families of marsupials, p. 433-505. In Genoways, H.H. (ed.), Current Mammalogy, Volume 2. Plenum Press, New York.
Matthew, W.D. 1903. The fauna of the Titanotherium beds at Pipestone Springs, Montana. Bulletin of the American Museum of Natural History, 19:1-19.
Matthew, W.D. 1909. The Carnivora and Insectivora of the Bridger Basin, middle Eocene. American Museum of Natural History Memoirs, 9:289-567.
Matthew, W.D. and Granger, W. 1921. New genera of Paleocene mammals. American Museum Novitates, 13:1-17.
Mattinson, J.M. 2005. Zircon U-Pb chemical abrasion (“CA-TIMS”) method: combined annealing and multi-step partial dissolution analysis for improved precision and accuracy of zircon ages. Chemical Geology, 220:47-66. https://doi.org/10.1016/j.chemgeo.2005.03.011
McCarroll, S.M., Flynn, J.J., and Turnbull, W.D. 1996a. Biostratigraphy and magnetostratigraphy of the Bridgerian-Uintan Washakie Formation, Washakie Basin, Wyoming, p. 25-39. In Prothero, D.R. and Emry, R.J. (eds.), The Terrestrial Eocene-Oligocene Transition in North America. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511665431.003
McCarroll, S.M., Flynn, J.J., and Turnbull, W.D. 1996b. The mammalian faunas of the Washakie Formation, Eocene age, of southern Wyoming, Part III. The perissodactyls. Fieldiana Geology New Series, 33:1-38. https://doi.org/10.5962/bhl.title.2705
McGrew, P.O. 1937. New marsupials from the Tertiary of Nebraska. Journal of Geology, 45:448-455. https://dx.doi.org/10.1086/624552
McGrew, P.O.1939. Nanodelphys, an Oligocene didelphine. Geologic Series of Field Museum of Natural History, 6:393-399. https://doi.org/10.5962/bhl.title.5208
McGrew, P.O. 1959. The geology and paleontology of the Elk Mountain and Tabernacle Butte area, Wyoming. Bulletin of the American Museum of Natural History, 117:121-176.
McGrew, P.O. and Sullivan, R. 1970. The stratigraphy and paleontology of Bridger A. Contributions to Geology, University of Wyoming, 9:66-85.
McKay, E.J. 1974. Geologic map of the Lone Mountain Quadrangle, Moffat County, Colorado. U.S. Geological Survey Map GQ-1144.
McKenna, M.C. 1960. Fossil Mammalia from the early Wasatchian Four Mile fauna, Eocene of northwest Colorado. University of California Publications of the Department of Geological Sciences, 37:1-130.
McKenna, M.C. and Bell, S.K. 1997. Classification of Mammals Above the Species Level. Columbia University Press, New York.
McKenna, M.C. and Simpson, G.G. 1959. A new insectivore from the middle Eocene of Tabernacle Butte, Wyoming. American Museum Novitates, 1952:1-12.
Miggins D.P. (2009) Temporal and geochemical insights related to volcanic and plutonic activity within Big Bend National Park, Texas. Unpublished Ph.D. dissertation, University of Texas at El Paso, Texas, USA.
Mihlbachler, M.C. 2008. Species taxonomy, phylogeny, and biogeography of the Brontotheriidae (Mammalia: Perissodactyla). Bulletin of the American Museum of Natural History, 311:1-475.
Miyata, K. and Deméré, T.A. 2016. New material of a "short-faced" Trogosus (Mammalia, Tillodontia) from the Delmar Formation (Bridgerian), San Diego County, California, U.S.A. Journal of Vertebrate Paleontology, 36:1-27. https://doi.org/10.1080/02724634.2016.1089878
Murphey, P.C. and Dunn, R.H. 2009. Hemiacodon engardae, a new species of omomyid primate from the earliest Uintan Turtle Bluff Member of the Bridger Formation, southwestern Wyoming, USA. Journal of Human Evolution, 57:123-130. https://doi.org/10.1016/j.jhevol.2009.05.006
Murphey, P.C. and Evanoff, E. 2007. Stratigraphy, fossil distribution, and depositional environments of the upper Bridger Formation (middle Eocene), southwestern Wyoming. Wyoming State Geological Survey, Report of Investigations, 57:1-107.
Murphey, P.C. and Kelly, T.S. 2017. Mammals from the earliest Uintan (middle Eocene) Turtle Bluff Member, Bridger Formation, southwestern Wyoming, USA, Part 2: Apatotheria, Lipotyphla, Carnivoramorpha, Condylarthra, Dinocerata, Perissodactyla and Artiodactyla. Palaeontologia Electronica 20.1.20a:1-50. https://doi.org/10.26879/720
https://palaeo-electronica.org/content/2017/1844-bridger-turtle-bluff-member-mammals.
Novacek, M.J., Ferrusquía-Villafranca, I., Flynn, J.J., Wyss, A.R., and Norell, M. 1991. Wasatchian (early Eocene) mammals and other vertebrates from Baja California, Mexico: the Lomas Las Tetas de Cabra Fauna. Bulletin of the American Museum of Natural History, 208:1-88.
Ogg, J.G. 2012. Geomagnetic Polarity Time Scale, p. 85-114. In Gradstein, F.M., Ogg, J.G., Schmitz, M.D., and Ogg, G.M. (eds.), The Geologic Time Scale 2012. Elsevier, Oxford, United Kingdom. https://doi.org/10.1016/B978-0-444-59425-9.00005-6
Opdyke, N.D., Lindsay, E.H., Johnson, N.M., and Downs, T. 1977. The paleomagnetism and magnetic polarity stratigraphy of the mammal-bearing section of Anza-Borrego State Park, California. Quaternary Research, 7:316-329. https://doi.org/10.1016/0033-5894(77)90024-2
Osborn, H.F. 1883. Achaenodon, an Eocene bunodont. Contributions from the E.M. Museum of Geology and Archaeology of Princeton College Bulletin, 3:23-45.
Osborn, H.F. 1895. Fossil mammals of the Uinta Basin. Expedition of 1894. Bulletin of the American Museum of Natural History, 7:71-105.
Osborn, H.F. 1908. New or little known titanotheres from the Eocene and Oligocene. Bulletin of the American Museum of Natural History, 24:599-617.
Osborn, H.F. 1929. The Titanotheres of ancient Wyoming, Dakota and Nebraska. USGS Monograph, 1:1-701. https://doi.org/10.5962/bhl.title.36431
Osborn, H.F. and Matthew, W.D. 1909. Cenozoic mammal horizons of western North America. USGS Bulletin, 361:1-138. https://doi.org/10.5962/bhl.title.86403
Parrish, R.R., Roddick, J.C., Loveridge, W.D., and Sullivan, R.D. l987. Uranium-lead analytical techniques at the geochronology laboratory, Geological Survey of Canada, p. 3-7. In Radiogenic age and isotopic studies, Report 1: Geological Survey of Canada Paper 87-2. https://doi.org/10.4095/122739
Prothero, D.R. 1996a. Magnetic stratigraphy and biostratigraphy of the middle Eocene Uinta Formation, Uinta Basin, Utah, p. 4-23. In Prothero, D.R. and R.J. Emry (eds.), The Terrestrial Eocene-Oligocene Transition in North America. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511665431.002
Prothero, D.R. 1996b. Magnetostratigraphy of the Eocene-Oligocene transition in Trans-Pecos Texas, p. 189-198. In Prothero, D.R. and Emry, R.J. (eds.), The Terrestrial Eocene-Oligocene Transition in North America. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511665431.010
Prothero, D.R. 1998. Hyracodontidae, p. 589-594. In Janis, C.M., Scott, K.M., and Jacobs, L.L. (eds.), Evolution of Tertiary Mammals of North America, Volume 1: Terrestrial Carnivores, Ungulates and Ungulatelike Mammals. Cambridge University Press, New York.
Prothero, D.R. and Emry, R.J. 1996. Summary, p. 664-683. In Prothero, D.R. and Emry, R.J. (eds.), The Terrestrial Eocene-Oligocene Transition in North America. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511665431.031
Rasmussen, D.T., Conroy, G.C., Friscia, A.R., Townsend, K.E., and Kinkle, M.D. 1999. Mammals of the middle Eocene Uinta Formation, p. 402-420. In Gillette, D.D. (ed.), Vertebrate Paleontology in Utah. Utah Geological Survey, Salt Lake City, Utah.
Reig, O.A., Kirsch, J.A., and Marshall, L.G. 1985. New conclusions on the relationships of the opossum-like marsupials, with an annotated classification of the Didelphimorphia. Ameghiniana, 21:335-343.
Rensberger, J.M. 1971. Entoptychine pocket gophers (Mammalia, Geomyoidea) of the early Miocene John Day Formation, Oregon. University of California, Publications in the Geological Sciences, 90:1-209.
Riggs, E.S. 1912. New or little known Titanotheres from the lower Uintah formations, with notes on the stratigraphy and distributions of fossils. Field Museum of Natural History Geological Series, 159:17-41. https://doi.org/10.5962/bhl.title.3381
Robinson, P. 1968. Nyctitheriidae (Mammalia, Insectivora) from the Bridger Formation of Wyoming. Contributions to Geology, University of Wyoming, 7:129-138.
Robinson, P., Gunnell, G.F., Walsh, S.L., Clyde, W.C., Storer, J.E., Stucky, R.K., Froehlich, D.J., Villafranca, I.F., and McKenna, M.C. 2004. Wasatchian through Duchesnean biochronology, p. 106-155. In Woodburne, M.O. (ed.), Late Cretaceous and Cenozoic Mammals of North America. Columbia University Press, New York.
Roehler, H.W. 1973. Stratigraphy of the Washakie Formation in the Washakie Basin, Wyoming. U.S. Geological Survey Bulletin, 1369:1-40.
Rose, K.D. 2010. New marsupial from the early Eocene of Virginia. Journal of Paleontology, 84:561-565. https://doi.org/10.1666/09-140.1
Rothecker, J. and Storer, J.E. 1996. The marsupials of the Lac Pelletier Lower Fauna, middle Eocene (Duchesnean) of Saskatchewan. Journal of Vertebrate Paleontology, 16:770-774. https://doi.org/10.1080/02724634.1996.10011365
Russell, L.S. 1984. Tertiary mammals of Saskatchewan Part VII: Oligocene marsupials. The Royal Ontario Museum, Life Sciences Contributions, 139:1-13. https://doi.org/10.5962/bhl.title.52239
Schärer, U., 1984. The effect of initial 230 Th equilibrium on young U-Pb ages: the Makula case, Himalaya. Earth and Planetary Science Letters, 67:191-204. https://doi.org/10.1016/0012-821X(84)90114-6
Schoene, B., Crowley, J.L., Condon, D.J., Schmitz, M.D., and Bowring, S.A. 2006. Reassessing the uranium decay constants for geochronology using ID-TIMS U-Pb data. Geochemica et Cosmochemica Acta, 70:426-445. https://doi.org/10.1016/j.gca.2005.09.007
Scott, W.B. 1941. The mammalian fauna of the White River Oligocene. Part V. Perissodactyla. Transactions of the American Philosophical Society, 28:747-980. https://doi.org/10.2307/1005518
Scott, W.B. and Osborn, H.F. 1877. Preliminary report on the vertebrate fossils of the Uinta Formation, collected by Princeton Expedition of 1886. Proceedings of the American Philosophical Society, 24:255-264.
Setoguchi, T. 1973. The late Eocene marsupials and insectivores from the Tepee Trail Formation, Badwater, Wyoming. Unpublished MS Thesis, Texas Tech University, USA.
Setoguchi, T. 1975. Paleontology and geology of the Badwater Creek area, central Wyoming Part II. Late Eocene marsupials. Annals of Carnegie Museum, 45:263-275.
Sigé, B. 1971. Les Didelphoidea de Laguna Umayo (Formation Vilquechico, Crétacé supérieur, Pérou), et le peuplement marsupial d'Amérique de Sud. Comptes Rendus des Séances de l'Académie des Sciences de Paris, 273:2479-2481.
Simpson, G.G. 1928. American Eocene didelphids. American Museum Novitates, 303:1-7.
Smith, M.E., Carroll, A.R., and Singer, B.S., 2008. Synoptic reconstruction of a major ancient lake system: Eocene Green River Formation, western United States. Geological Society of America Bulletin, 120:54-84. https://doi.org/10.1130/b26073.1
Smith, M.E., Chamberlain, K.R., Singer, B.S., Carroll, A.R. 2010. Eocene Clocks Agree: Coeval 40Ar/39Ar, U-Pb, and Astronomical Ages from the Green River Formation. Geology, 38:527-530. https://doi.org/10.1130/G30630.1
Stacey, J.S. and Kramers, J.D., 1975, Approximation of terrestrial lead isotope evolution by a two-stage model. Earth and Planetary Science Letters, 26:207-221. https://doi.org/10.1016/0012-821X(75)90088-6
Steiger, R.H. and Jäger, E. 1977. Subcommission on geochronology: convention on the use of decay constants in geo- and cosmochronology. Earth and Planetary Science Letters, 36:359-362. https://doi.org/10.1016/0012-821X(77)90060-7
Stevens J.B., Stevens, M.S., and Wilson, J.A. 1984. The Devil's Graveyard Formation (new) Eocene and Oligocene age Trans-Pecos Texas. Texas Memorial Museum Bulletin, 32:1-21.
Stock, C. 1936. Sespe Eocene didelphids. Proceedings of the National Academy of Sciences, Philadelphia, 22:122-124. https://doi.org/10.1073/pnas.22.2.122
Stock, C. and Furlong, 1922. A marsupial from the John Day Oligocene of Logan Butte, eastern Oregon. University of California Publications in Geological Sciences, 13:311-317.
Storer, J.E. 1984. Mammals of the Swift Current Creek Local Fauna (Eocene: Uintan), Saskatchewan. Natural History Contributions, Saskatchewan Museum of Natural History, 7:1-158.
Storer, J.E. 1996. Eocene-Oligocene faunas of the Cypress Hills Formation, Saskatchewan, p. 240-261. In Prothero, D.R. and Emry, R.J. (eds.), The Terrestrial Eocene-Oligocene Transition in North America. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511665431
Stucky, R.K, Prothero, D.R., Lohr, W.G., and Snyder, J.R. 1996. Magnetic stratigraphy, sedimentology, and mammalian faunas of the early Uintan Washakie Formation, Sand Wash Basin, northwestern Colorado, p. 40-51. In Prothero, D.R. and Emry, R.J. (eds.), The Terrestrial Eocene-Oligocene Transition in North America. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511665431
Stucky, R.K. and Snyder, J.R. 1992. Mammalian fauna of the Sand Wash Basin, Colorado (Washakie Formation, middle Eocene, earliest Uintan). Abstracts with Programs, Fifty-Second Annual Meeting, Society of Vertebrate Paleontology, 12:54A.
Sundell, K.A., Shive, P.N., and Eaton, J.A. 1984. Measured sections, magnetic polarity, and biostratigraphy of the Eocene Wiggins, Tepee Trail and Aycross formations with the southeastern Absaroka Range, Wyoming. Earth and Science Bulletin of the Wyoming Geological Association, 17:1-48.
Szalay, F.S. 1969. Mixodectidae, Microsyopidae, and the insectivore-primate transition. Bulletin of the American Museum of Natural History, 140:195-330.
Szalay, F.S. 1976. The systematics of the Omomyidae (Tarsiiformes, Primates): taxonomy, phylogeny and adaptations. Bulletin of the American Museum of Natural History, 156:157-450.
Townsend, K.E., Friscia, A.R. and Rasmussen, D.T. 2006. Stratigraphic distribution of upper middle Eocene fossil vertebrate localities in the eastern Uinta Basin, Utah, with comments on Uintan biostratigraphy. The Mountain Geologist, 43:115-134.
Troxell, E.L. 1923a. A new marsupial. American Journal of Science, 5:507-510. https://doi.org/10.2475/ajs.s5-5.30.507
Troxell, E.L. 1923b. The Eocene rodents Sciuravus and Tillomys. American Journal of Science, 5:383-396. https://doi.org/10.2475/ajs.s5-5.29.383
Tsukui, K. 2016. Chronology and faunal evolution of the middle Eocene Bridgerian North American Land Mammal "Age": achieving high precision geochronology. Unpublished PhD Thesis, Columbia University, New York, USA.
Tsukui, K. and Clyde, W.C. 2012. Fine-tuning the calibration of the early to middle Eocene geomagnetic polarity time scale: paleomagnetism of radioisotopically dated tuffs from Laramide foreland basins. Geological Society of America Bulletin, 124:870-885. https://doi.org/10.1130/B30545.1
Turnbull, W.D. 2002. The mammalian faunas of the Washakie Formation, Eocene age, of southern Wyoming. Part IV. The Uintatheres. Fieldiana Geology, 47:1-189. https://doi.org/10.5962/bhl.title.3567
Walsh, S.L. 1996a. Theoretical biochronology, the Bridgerian/Uintan boundary, and the "Shoshonian subage" of the Uintan, p. 52-74. In Prothero, D.R. and Emry, R.J. (eds.), The Terrestrial Eocene-Oligocene Transition in North America. Cambridge University Press, New York. https://doi.org/10.1017/CBO9780511665431
Walsh, S.L. 1996b. Middle Eocene mammalian faunas of San Diego County, California, p. 75-119. In Prothero, D.R. and Emry, R.J. (eds.), The Terrestrial Eocene-Oligocene Transition in North America. Cambridge University Press, New York. https://doi.org/10.1017/CBO9780511665431
Walsh, S.L. 1998. Fossil datum and paleobiological event terms, paleontostratigraphy, chronostratigraphy, and the definition of land mammal "age" boundaries. Journal of Vertebrate Paleontology, 18:150-179. https://doi.org/10.1080/02724634.1998.10011041
Walsh, S.L., Prothero, D.R., and Lundquist, D.J., 1996. Stratigraphy and paleomagnetism of the middle Eocene Friars Formation and Poway Group, southwestern San Diego County, California, p. 120-151. In Prothero, D.R. and Emry, R.J. (eds.), The Terrestrial Eocene-Oligocene Transition in North America. Cambridge University Press, New York. https://doi.org/10.1017/CBO9780511665431
Walton, A.H. 1992. Magnetostratigraphy and the ages of Bridgerian and Uintan faunas in the lower and middle members of the Devil's Graveyard Formation, Trans-Pecos Texas, p. 74-87. In Prothero, D.R. and Berggren, W.A. (eds.), Eocene-Oligocene Climatic and Biotic Evolution. Princeton University Press, Princeton, New Jersey. https://doi.org/10.1515/9781400862924
Watson, G.S. 1956. A test for randomness of directions. Monthly Notices of the Royal Astronomical Society Geophysical Supplement, 7:160-161. https://doi.org/10.1111/j.1365-246X.1956.tb05561.x
West, R.M. 1973. Geology and mammalian paleontology of the New Fork-Big Sandy area, Sublette County, Wyoming. Fieldiana, 29:1-193. https://doi.org/10.5962/bhl.title.3573
West, R.M. 1974. New North American middle Eocene nyctithere (Mammalia, Insectivora). Journal of Paleontology, 48:983-987.
West, R.M. 1979. Paleontology and geology of the Bridger Formation, southern Green River Basin, southwestern Wyoming. Part 3. Notes on Hyopsodus. Contributions in Biology and Geology, Milwaukee Public Museum, 25:1-52.
West, R.M. 1982. Fossil mammals from the lower Buck Hill Group, Eocene of Trans-Pecos Texas. Pearce-Sellards Series, 35:1-20.
West, R.M and Dawson, M.R. 1973. Fossil mammals from the upper part of the Cathedral Bluffs tongue of the Wasatch Formation (early Bridgerian), northern Green River Basin, Wyoming. Contributions to Geology, University of Wyoming, 12:33-41.
West, R.M. and Dawson, M.R. 1975. Eocene fossil Mammalia from the Sand Wash Basin, northwestern Moffat County, Colorado. Annals of Carnegie Museum, 45:231-253.
West, R.M. and Hutchison, J.H. 1981. Paleontology and geology of the Bridger Formation, southern Green River Basin, southwestern Wyoming, Part 6, the fauna and correlation of Bridger E. Contributions in Biology and Geology, Milwaukee Public Museum, 46:1-8.
Westerhold, T., and Rӧhl, U. 2009. High resolution cyclostratigraphy of the early Eocene - new insights into the origin of the Cenozoic cooling trend. Climate of the Past, 5:309-327. https://doi.org/10.5194/cp-5-309-2009
Westerhold, T., Röhl, U., Frederichs, T., Bohaty, S.M., and Zachos, J.C. 2015. Astronomical calibration of the geological timescale: closing the middle Eocene gap. Climate of the Past, 11:1665-1699. https://doi.org/10.5194/cp-11-1181-2015
Westerhold, T., Röhl, U., Laskar, J., Raffi, I., Bowles, J., Lourens, L.J., and Zachos, J.C., 2007. On the duration of magnetochrons C24r and C25n and the timing of early Eocene global warming events: Implications from the Ocean Drilling Program Leg 208 Walvis Ridge depth transect. Paleoceanography, 22:PA2201. https://doi.org/10.1029/2006PA001322
Westerhold, T., Rӧhl, U., Raffi, I., Fornaciari, E., Monechi, S., Reale, V., Bowles, J., and Evans, H.F. 2008. Astronomical calibration of the Paleocene time. Palaeogeography, Palaeoclimatology, Palaeoecology, 257:377-403. https://doi.org/10.1016/j.palaeo.2007.09.016
Westgate, J. and Emry, R.J. 1985. Land mammals of the Crow Creek Local Fauna, late Eocene, Jackson Group, St. Francis County, Arkansas. Journal of Paleontology, 59:242-248.
Williams, B.A. and Kirk, E.C., 2008. New Uintan primates from Texas and their implications for North American patterns of species richness during the Eocene. Journal of Human Evolution, 55:927-941. https://doi.org/10.1016/j.jhevol.2008.07.007
Williamson, T.E., Brusatte, S.L., Carr, T.D., Weil, A., and Standhardt, B.R. 2012. The phylogeny and evolution of Cretaceous-Palaeogene metatherians: cladistic analysis and description of new early Palaeocene specimens from the Nacimiento Formation, New Mexico. Journal of Systematic Palaeontology, 10:625-651. https://doi.org/10.1080/14772019.2011.631592
Williamson, T.E. and Taylor, L.H. 2011. New species of Peradectes and Swaindelphys (Mammalia: Metatheria) from the early Paleocene (Torrejonian) Nacimiento Formation, San Juan Basin, New Mexico. Palaeontologia Electronica, 14.3.23A:1-16.
Wilson, J.A. 1984. Vertebrate faunas 49 to 36 million years ago and additions to the species of Leptoreodon (Mammalia: Artiodactyla) found in Texas. Journal of Vertebrate Paleontology, 4:199-207. https://doi.org/10.1080/02724634.1984.10012003
Wilson, J.A. 1986. Stratigraphic occurrence and correlation of early Tertiary vertebrate faunas, Trans-Pecos Texas: Agua Fria-Green Valley areas. Journal of Vertebrate Paleontology, 6:350-373. https://doi.org/10.1080/02724634.1986.10011630
Wilson, J.A. and Schiebout, J.A. 1984. Early Tertiary vertebrate faunas, Trans-Pecos Texas: Ceratomorpha less Amynodontidae. Pearce-Sellards Series, 39:1-47.
Wilson, R.W. 1937. Two new Eocene rodents from the Green River Basin, Wyoming. American Journal of Science, 34:447-456. https://doi.org/10.2475/ajs.s5-34.204.447
Wood, A.E. 1959. The geology and paleontology of the Elk Mountain and Tabernacle Butte area, Wyoming, Rodentia. Bulletin of the American Museum of Natural History, 117:157-169.
Wood, A.E. 1973. Eocene rodents, Pruett Formation, southwest Texas: their pertinence to the origin of the south American Caviomorpha. Pearce-Sellards Series, 20:1-41.
Wood, A.E., Chaney, R.W., Clark, J., Colbert, E.H., Jepsen, G.L., Reeside, Jr., J.B., and Stock, C. 1941. Nomenclature and correlation of the North American continental Tertiary. Bulletin of the Geological Society of America, 52:1-48. https://doi.org/10.1130/GSAB-52-1
Woodburne, M.O. 1977. Definition and characterization in mammalian chronostratigraphy. Journal of Paleontology, 51:220-234.
Woodburne, M.O. 1987. Introduction, p. 1-8. In Woodburne, M.O. (ed.), Cenozoic Mammals of North America. Berkeley, University of California Press.
Woodburne, M.O. 2004a. Principles and procedures, p. 1-20. In Woodburne, M.O. (ed.), Late Cretaceous and Cenozoic Mammals of North America. Berkeley, University of California Press. https://doi.org/10.7312/wood13040-003
Woodburne, M.O. 2004b. Global events and the North American Mammalian biochronology, p. 315-343. In Woodburne, M.O. (ed.), Late Cretaceous and Cenozoic Mammals of North America. Berkeley, University of California Press. https://doi.org/10.7312/wood13040-010
Wortman, J.L. 1898. The extinct Camelidae of North America and some associated forms. Bulletin of the American Museum of Natural History, 10:93-142.
Wortman, J.L. 1901. Studies of Eocene mammals in the Marsh Collection, Peabody Museum, Part 1: Carnivora. American Journal of Science, Series 4, 7:143-154. https://doi.org/10.2475/ajs.s4-12.68.143
