

 New records of Bennettitales and associated flora from the Jurassic of the Cualac Formation, Mexico
New records of Bennettitales and associated flora from the Jurassic of the Cualac Formation, Mexico
Article number: 27.1.a14
https://doi.org/10.26879/1293
Copyright Paleontological Society, February 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 14 April 2023. Acceptance: 17 January 2024.
ABSTRACT
The present work contributes to the palaeofloristic knowledge of the Jurassic in the Cualac Formation of Mexico with the first records of the genera Anomozamites, Cycadolepis, Czekanowskia, Mexiglossa, and Williamsoniella. A new species of bisexual structure, Williamsonia rosarensis, is described; the species Zamites diquiyui is emended and the cuticular characters of the dominant species Mexiglossa varia are described for the first time. These data substantially increase the plant diversity of this stratigraphic unit and provide information about the environment in which they were preserved. These new reports come from a new locality of the Cualac Formation, in the state of Oaxaca, Mexico, called Rosario. After considering some sedimentological and taphonomic aspects, it is proposed that this locality represented an alluvial fan depositional environment that evolved into a flood plain, which favored the state of preservation of the specimens, and even preserved cuticular characters that allows an updated and more detailed taxonomy of three species. A notable find is Williamsoniella rosarensis nov. sp., a genus which has been previously reported from two Middle Jurassic localities in the UK and one in Mexico. A possible organic connection between this structure and leaves of Zamites diquiyui is also proposed. The findings mentioned above indicate that Bennettitales is the most abundant order (61.4%) with eight genera and 15 species. The present contribution confirms that paleofloristic lists are still incomplete, and thus it is necessary to continue the work in the Jurassic formations described for Mexico.
María Patricia Velasco de León. Colección de Paleontología, Facultad de Estudios Superiores, Zaragoza, Universidad Nacional Autónoma de México, Guelatao avenue, Ejercito de Oriente, Mexico City, 09230, Mexico. pativel@unam.mx
Erika Lourdes Ortiz Martínez. Colección de Paleontología, Facultad de Estudios Superiores, Zaragoza, Universidad Nacional Autónoma de México, Guelatao avenue, Ejercito de Oriente, Mexico City, 09230, Mexico. erikaortizmartinez01@gmail.com
Miguel Angel Flores Barragan. Colección de Paleontología, Facultad de Estudios Superiores, Zaragoza, Universidad Nacional Autónoma de México, Guelatao avenue, Ejercito de Oriente, Mexico City, 09230, Mexico (corresponding author). 08mikeangel@gmail.com
Diana Silvia Guzmán Madrid. Colección de Paleontología, Facultad de Estudios Superiores, Zaragoza, Universidad Nacional Autónoma de México, Guelatao avenue, Ejercito de Oriente, Mexico City, 09230, Mexico. dianaguz_8@yahoo.com.mx
Pedro Christian Martínez Martinez. Colección de Paleontología, Facultad de Estudios Superiores, Zaragoza, Universidad Nacional Autónoma de México, Guelatao avenue, Ejercito de Oriente, Mexico City, 09230, Mexico. pitpedrito@yahoo.com.mx
Key words: New species; Mesozoic; Paleoflora; Zamites; Williamsoniella; Mexiglossa
Final citation: Velasco de León, Maria Patricia, Ortiz Martinez, Erika Lourdes, Flores Barragan, Miguel Angel, Guzmán Madrid, Diana Silvia, and Martinez Martinez, Pedro Christian. 2024. New records of Bennettitales and associated flora from the Jurassic of the Cualac Formation, Mexico. Palaeontologia Electronica, 27(1):a14.
https://doi.org/10.26879/1357
palaeo-electronica.org/content/2024/5138-bennettitales-from-mexico
Copyright: February 2024 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
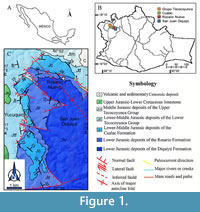 The Jurassic floras of Tezoatlán de Segura y Luna, in the municipality of Oaxaca, Mexico, have been studied for more than a century. Here, the sedimentary Tezoatlan paleobasin is located (Figure 1). The exposed stratigraphic sequence of this basin comprises the Diquiyui, Rosario, and Cualac formations and the Lower Tecocoyunca Group (Erben, 1956; Moran-Zenteno et al., 1993; Zepeda-Martínez et al., 2018). The sequence has been dated to 180-188 m.y. by the U-Pb zircon method (Martini et al., 2016; Zepeda-Martínez et al., 2018). Its lithology includes volcanic rocks and a rhythmic superposition of continental to marine deposits, composed of coarsening-upward successions of conglomerate, siltstone-sandstone, and mudstone.
The Jurassic floras of Tezoatlán de Segura y Luna, in the municipality of Oaxaca, Mexico, have been studied for more than a century. Here, the sedimentary Tezoatlan paleobasin is located (Figure 1). The exposed stratigraphic sequence of this basin comprises the Diquiyui, Rosario, and Cualac formations and the Lower Tecocoyunca Group (Erben, 1956; Moran-Zenteno et al., 1993; Zepeda-Martínez et al., 2018). The sequence has been dated to 180-188 m.y. by the U-Pb zircon method (Martini et al., 2016; Zepeda-Martínez et al., 2018). Its lithology includes volcanic rocks and a rhythmic superposition of continental to marine deposits, composed of coarsening-upward successions of conglomerate, siltstone-sandstone, and mudstone.
Fossil plants, e.g., permineralizations, impressions, and carbonaceous compressions, have been described from the stratigraphic units; Formation Rosario, Cualac and Grupo Tecocoyunca Inferior (Wieland, 1914; Delevoryas and Pearson, 1975; Pearson and Delevoryas, 1982; Silva-Pineda, 1984; Jiménez-Rentería, 2004; Ortega-Cháves et al., 2017; Carrasco-Ramírez et al., 2016; Lozano-Carmona and Velasco-de León, 2016). However, of the formations mentioned above, the Cualac Formation is the least studied with only a single report of poorly preserved material (Silva-Pineda and González-Gallardo, 1988).
The classic paleobotanical work on the Jurassic flora of Mexico in the region of Consuelo, Oaxaca, was carried out by Wieland (1914), who reported 50 species and six varieties (Table 1), from the Rosario and Tecocoyunca Group stratigraphic units. Silva-Pineda (1970) studied material from the region of Tezoatlán and identified 23 species of fossil flora, including three from the Cualac Formation: Otozamites graphicus Schimper, O. aguilerai Wieland, and Zamites sp cf. Z. feneonis Brongniart (Table 1).
Person and Delevoryas (1982) later collected material from the middle Jurassic of Oaxaca in the region of Tezoatlán. However, due to the state of preservation of some specimens examined by Wieland (1914) and the lack of a detailed description of the species mentioned, only 26 plant species are considered valid for the Rosario and Zorrillo formations (Table 1); this table includes the original reports of each author.
The Cualac Formation was reported for the first time in the state of Guerrero by Guzmán (1950). However, no plant fossils were reported until those by Silva-Pineda and González-Gallardo (1988) from siltstone. The authors attributed these to Otozamites hespera Wieland, Ptillophyllum sp. and Brachyphyllum (Brongniart) Harris. However, the described material was scarce and poorly preserved (Silva-Pineda, 1970; Silva-Pineda and González- Gallardo, 1988). The fossil-bearing outcrops there are composed of thick layers of white to yellowish quartz matrix conglomerate composed of milky quartz mica schist pebbles, gneiss, and ignimbrite occasionally interspersed with siltstone and shale (Corona-Esquivel, 1981).
There are few reports of fossil plants in this formation (Table 1), which is described as not very diverse and sparse, as well as fragmented and poorly preserved (Silva-Pineda, 1970; Silva-Pineda and González-Gallardo, 1988). Thus, the species descriptions mentioned above are mostly insufficient and lack of epidermal details cuticular data. Cuticular characters are important in the taxonomic assignment of different plant groups when macroscopic characters do not allow a sound systematic placement (Pott et al., 2010). In the Jurassic of Oaxaca cuticles of Bennettitales, Coniferales, and groups of uncertain affinity have been described previously (Silva-Pineda, 1969; Ortiz-Martínez, 2014; Guzmán-Madrid and Velasco- de León, 2015, 2017, 2021; Guzmán-Madrid et al., 2019).
Here, we describe well-preserved plant fossils from a recently discovered new locality in the Cualac Formation, near the town of Rosario Nuevo, in the municipality of Tezoatlán de Segura y Luna, Oaxaca.
STUDY AREA AND METHODS
The material studied was collected in the municipality of Tezoatlán de Segura y Luna, around the town of Rosario Nuevo, Oaxaca, Mexico. The fossil-bearing strata belong to the Cualac Formation. In the study area this unit has a thickness of around 100 m. This sequence of rocks is found above the fluvial deposits belonging to the Rosario Formation and is located below the strata of the Tecocoyunca Group. It has been interpreted that this unit was deposited in an alluvial plain environment that was occasionally invaded by marine waters (Morán-Zenteno et al., 1993).
The Cualac Formation is composed of conglomerates, fine-conglomeratic sandstone, and thin shale strata (Zepeda-Martínez, et al., 2021). Ten lithofacies have been recognized that are organized into two different architectural elements, Gravel Bars and Bedforms (GB) and Floodplain fines (FF), which according to Zepeda-Martínez et al. (2021) was part of a coalescent alluvial fan system.
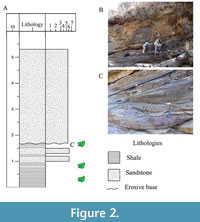 Specifically, the collection area of this work is located in the upper part of the stratigraphic column (meters 73-78) proposed for the Tezoatlan area (fig. 5A, Zepeda Martines et al., 2021), which is characterized by an alternation of coarse-grained shale and sandstone strata (Figure 2).
Specifically, the collection area of this work is located in the upper part of the stratigraphic column (meters 73-78) proposed for the Tezoatlan area (fig. 5A, Zepeda Martines et al., 2021), which is characterized by an alternation of coarse-grained shale and sandstone strata (Figure 2).
The fossil material recovered in the studied area corresponds to well-preserved plant fragments, whose stratigraphic position, state of preservation, and type of preserved structure were recorded, their degree of fragmentation (they must retain two-thirds of their size to be considered as complete leaves), the size of the organs, and their abundance (Greenwood, 1991; Mancuso and Marsicano, 2008) to support the degree of material transport. These made it possible to analyze the transport to which they were subjected (Fergunson, 2005). The specimens were deposited in the Paleontological Collection of the Facultad de Estudios Superiores Zaragoza of the Universidad Nacional Autónoma de México under the acronym CFZCr. For the description and subsequent taxonomic identification of the fossil material collected, observations were made under an Olympus SZ51 stereo microscope and photographs were obtained with a Nikon 5100 camera. The photographs were used to measure each specimen in ImageJ version 1.38x.
Cuticular extraction was performed using the following techniques: acetate peel, dispersed cuticles with 10% hydrofluoric acid (HF), and mechanical extraction. The extracted cuticles were treated with a 3:1 Schultze solution (HNO3 and KClO3), 40% nitric acid (HNO3), and 5% ammonium hydroxide (NH4OH), bleached with 6% sodium hypochlorite (NaClO) and 30% hydrogen peroxide (H2O2), washed with distilled water, and mounted in 5% glycerin jelly (Kerp, 1990; Knowenberg et al., 2007; Guzmán-Madrid, 2016; Guzmán-Madrid and Velasco-de León, 2017). In the case of CFZCr68, cellulose acetate was placed in the basal part of the flower where there is a layer of carbon to try to recover organic material. The cuticular samples were observed under an Olympus BX41 optical microscope and photographed with an Olympus E-620 digital camera. Specialized literature was used for the taxonomic determination of the specimens (Thomas, 1915; Harris, 1953; Harris and Miller, 1974; Pott and McLoughlin, 2009).
Registration
The new name is registered with a unique PFN number in the Plant Fossil Names Registry, hosted and operated by the National Museum, Prague for the International Organisation of Palaeobotany.
RESULTS
Thus far, 97% of the 517 fossils collected have been identified, of which 90% correspond to complete leaves and belong to three orders: Bennettitales, Czekanowskiales, and Filicales, representing 61.4%, 0.6%, and 1% in this locality, respectively. The rest correspond to Mexiglossa Person and Delevoryas. Of these orders, the Bennettitales are the most diverse group with nine genera: Otozamites Wieland, Ptilophyllum Morris, Zamites Brongniart, Anomozamites Schimper, Mexiglossa, Cycadolepis Saporta, Pterophyllum Brongniart, Williamsonia Carruthers, and Williamsoniella Thomas. The latter are reported for the first time in this stratigraphic unit. In the case of the Filicales, the specimens collected thus far correspond to small fragments of poorly preserved sterile fronds that are not possible to identify, and the specimens of Williamsonia are rare and not very diverse. Table 2 shows a list of the species present in this formation.
It is important to note that the species described for the Rosario locality result in a three-fold increase in diversity compared to previous reports from the Cualac Formation (Table 2). This is due to the good state of preservation of the material in carbonaceous shale, which allowed identification of four genera of Bennettitales and one of Czekanowskiales (Czekanowskia Heer) that had not been previously described for the Cualac Formation (Table 2). Previously, Czekanowskia had been recorded only for the Zorrillo Formation of the Middle Jurassic and with few specimens. The new collection also allows expanding the description of the taxa Mexiglossa varia and Zamites diquiyui (Wieland) Person and Delevoryas. In the case of the latter, the characters described in the present work allowed emending the original description by Person and Delevoryas (1982). Finally, a new species of Williamsoniella is proposed. The new and best-preserved taxa are described below.
SYSTEMATIC PALEONTOLOGY
Order BENNETTITALES Engler, 1892
Family WILLIAMSONIACEAE (Carruthers, 1870) Nathorst, 1913
Genus ANOMOZAMITES Schimper (1870) emend. Pott et McLoughlin, 2009
Type species. Anomozamites nilssonii (Phillips, 1829) Harris, 1969, Jurassic of Yorkshire, England
Anomozamites sp.
Figure 3A-B
Material examined. CFZ-Cr 3, Cr 4, Cr 7, Cr 27, Cr 53.
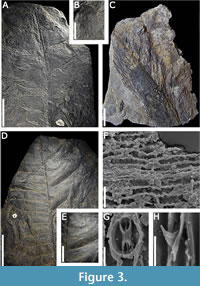 Description. Incomplete compound leaf, regularly segmented, with a length of 9.7-8.9 cm and a width of 4.4 cm. Rachis traversed longitudinally by three distinct, about 0.2 cm wide, striae (Figure 3A). Leaflets oppositely positioned, oblong and tapering towards the base, 2.5 cm long and 1.1 cm wide, length/width ratio is 2:1, laterally attached to the stem along their entire base, which is recurrent, apex rounded to obtuse (Figure 3B). The macroscopic margin is straight (90˚), while the basiscopic margin is decurrent. Leaflet longitudinally traversed by six-11 distinct veins per leaflet that run perpendicular to the rachis, bifurcating once near the base (Figure 3B).
Description. Incomplete compound leaf, regularly segmented, with a length of 9.7-8.9 cm and a width of 4.4 cm. Rachis traversed longitudinally by three distinct, about 0.2 cm wide, striae (Figure 3A). Leaflets oppositely positioned, oblong and tapering towards the base, 2.5 cm long and 1.1 cm wide, length/width ratio is 2:1, laterally attached to the stem along their entire base, which is recurrent, apex rounded to obtuse (Figure 3B). The macroscopic margin is straight (90˚), while the basiscopic margin is decurrent. Leaflet longitudinally traversed by six-11 distinct veins per leaflet that run perpendicular to the rachis, bifurcating once near the base (Figure 3B).
Discussion. The fossil taxon Anomozamites comprises segmented leaves with leaflets as broad as long or twice as long as broad, with a broad base generally inserted laterally to the rachis, with numerous and parallel veins, simple or once-forked (Figure 3A). In recent decades, the assignment of fossil leaves based on the classic definitions made by Harris (1932) for Anomozamites and Pterophyllum have been questioned (Pott and McLoughlin, 2009), This has led to the revision and in many cases the taxonomic relocation of leaves. The macroscopic characteristics of the specimens from the Terreno Mixteco agree with the description amended by Pott and McLoughlin (2009) for Anomozamites, since they are leaves up to twice as long as they are wide. This is an essential character for their assignment, since according to Pott and McLoughlin (2009) the species of the genus Pterophyllum have leaves with leaflets that are more than twice as long as they are wide.
Other characters that corroborate the assignment to Anomozamites are the uniform shape of the leaflets along the length of the leaf, numerous strong parallel veins, simple or once bifurcated near the leaflet base. At the species level, the specimens were compared with A. angustifolium Pott et McLoughlin, specimens with affinity to the species A. triangularis and with A. sp. described by Lozano-Carmona et al. (2019) for the Zorrillo-Taberna Formation, since, although Person and Delevoryas (1982) mention for the first time the presence of the genus for the Rosario (Early Jurassic) and Zorrillo (Middle Jurassic) formations, the conservation status of the specimens did not allow assigning them to a species. On the other hand, the bibliographic review of the morphological characteristics of 30 species of Anomozamites described for Laurasia by Xu et al. (2019) and the characteristics described for the species from the Terreno Mixteco indicate that the specimens analyzed here differ from the species with which they were compared in the characteristics of the rachis and the size, shape, and number of veins per leaflet. It is therefore impossible to assign them to a previously described species of the genus.
Anomozamites is one of the most important genera of the extinct order Bennettitales and also one of the most diverse in Laurasia. Analysis of its distribution during the Mesozoic in Asia shows that Anomozamites appeared in China during the Late Triassic, where it reached its maximum diversity. At the end of this period, the number of its species decreased significantly but its distribution extended. It was in Mexico in the region of Tezoatlán (Oaxaca) during the Lower Jurassic and up to the Middle Jurassic, but finally disappeared from the planet during the Cretaceous (Xu et al., 2019).
Genus CYCADOLEPIS Saporta 1875 emend. Harris, 1953
Type species. Cycadolepis villosa Saporta, 1875
Cycadolepis aff. C. mexicana Wieland, 1914
Figure 3C
Material examined. CFZ-Cr 137
Description. Lanceolate bract impression, with four longitudinal veins, margin entire, 6.1 cm long and 1.8 cm wide at maximum, hairs 10.4-21.0 mm long and 0.4-1.4 mm wide on the margin, without apparent arrangement.
Discussion. Cycadolepis is used to name the set of bracts that likely surrounded the reproductive structures of some Bennettitales and detached with floral maturity (McLoughlin, 2017). The species C. mexicana Wieland has been reported from the Jurassic of Mexico, and has the same shape, size, and presence of hairs as the specimen described in the present work (Figure 3C). However, the fragmentation of the specimen only allows proposing an affinity with this species. This genus has a cosmopolitan distribution throughout the Mesozoic (Beheresmeyer and Turner, 2022).
Genus ZAMITES Brongniart, 1828 emend. Harris, 1969
Type species. Zamites gigas Lindley and Hutton, 1831
Zamites diquiyui (Wieland) Person et Delevoryas, 1982
Figure 3D-H
1914 Otozamites or Zamites Wieland, pl.19, figs. 6, 7.
1914 Otozamites (Otopteris) sp. Wieland, pl. 44, fig. 2.
1982 Zamites diquiyui Person and Delevoryas, pl. 6, fig. 39.
Holotype. IGPB 420. U. of Texas P. C. No. 164
Epitype. CCr10-1
Type locality. Rosario, Cualac Formation
Age. Lower-Middle Jurassic
Deposited. Facultad de Estudios Superiores Zaragoza, Universidad Nacional Autónoma de México.
Material examined. CFZ Cr. 1-3, Cr4-9, Cr9-4, Cr10-1, Cr20-1, Cr24-4, Cr28-6, Cr28-11, Cr31-1, 3, and 4, Cr40-2, Cr42-10, Cr45-1, Cr46-1, Cr51-8, Cr55-1, Cr58-3, Cr62-1 and 2, Cr64-1, Cr66-2, Cr73-2, Cr76-3, Cr78-3, Cr82-5, Cr84-2, Cr94-1 and 2, Cr95-1, Cr97-5, 6, 7, and 8, Cr98-1, Cr114-3, and Cr120-1, Cr123-1,
Emended diagnosis. Compound leaves, leaflet entire-margined, base cordate even quadrate, acute apex, inserted to the rachis by the central part of the base, and sub-oppositely arranged. Fine and robust occasionally bifurcating veins. Cuticle with syndetocheilic stomata, with rectangular epidermal cells and hook-shaped trichomes.
Description. Elliptic leaf 26.5 cm long and 19.6 cm wide, rachis longitudinally striated and tapering towards the leaf apex with a maximum width of 0.4 cm. Leaflet sup-oppositely arranged, close together, 4.9-10.6 cm long and 1.7-3.5 cm wide (Figure 3D). Acute apex, base cordate, quadrate in some cases (Figure 3D). Pinnae laterally inserted to the rachis in angles of 80°-85°, robust veins at the base, widely spaced, at a density of seven-10 per leaflet, occasionally bifurcating once near the base. Two finer veins between the larger veins (Figure 3E).
Cuticle with rectangular epidermal cells 6.4-18.9 μm wide and 31.1-80.7 μm long, short, and elongated cells are interspersed, elongating, and thinning towards the rachis with a width of 4.2-6.7 μm and a length of 39.6-72.4 μm. Undulate to sinuous cell walls 1.4-6.2 μm wide, which are straight in the rachis and 0.5-1 μm wide (Figure 3F). Dispersed simple, unicellular, hook-shaped trichomes 3-7 μm long with a rounded base 1.7-5.6 μm wide between the stomata, size increases in the veins to 30.1 μm in diameter at the base and up to 47.4 μm in length. Oval-rounded syndetocheilic stomata 14.6-20.9 μm wide and 19.9-25.2 μm long at the base, elongating towards the center to up to 38.7 μm. Open ostioles are 3-4.5 μm wide and 7.6-8.1 μm long and closed ostioles are 1-2.9 μm wide and 10.7-24.9 μm long. Guard cells are 1.5-2 μm wide and 5.1-9.8 μm long, subsidiary cells are 4.7-6.8 μm wide and 7.1-22.6 μm long (Figure 3G).
Discussion. Given that the material used to establish Z. diquiyui is based only on poorly preserved isolated pinnae (Person and Delevoryas, 1982, pl. 6, fig. 39), its description is ambiguous, and thus it was necessary to assign a new type with a higher number of morphological and cuticular characters, which were used for the emendation of the taxon.
Epitype CCr10-1 has compound leaves, pinnae with a symmetric base, and parallel veins that never anastomose, which are characters that allow corroborating its assignment to the genus Zamites (Harris, 1969). The species Z. oaxacensis, Z. lucerensis, and Z. tribulosus recorded in the Rosario locality (Table 2) differ in leaf and pinna size and have a higher vein density, a variable vein arrangement, and lack trichomes.
Genus WILLIAMSONIELLA Thomas, 1915
Type species. Williamsoniella coronata (Thomas) Harris,1944
Williamsoniella rosarensis sp. nov.
Figure 4A-G
PFN003250
Holotype. CFZCr115
Paratype. CFZCr11, 45-5, 68, 82, 112, 128
 Type locality. Rosario, Cualac Formation
Type locality. Rosario, Cualac Formation
Age. Lower-Middle Jurassic
Deposited. Facultad de Estudios Superiores Zaragoza, Universidad Nacional Autónoma de México.
Etymology. The specific epithet is dedicated to the village of Rosario
Diagnosis. Pedunculate, flower-like hermaphrodite structure; obovate gynoecium with obtuse crown, rhomboidal interseminal scales. The androcecium consists of at least four free microsporophylls surrounding the gynoecium. The peduncle is robust, longitudinally striated and has bracteoles on two levels. Elongated epidermal cells, thin-walled and straight, simple single-celled hooked trichomes. Oval syndetochelic stomata.
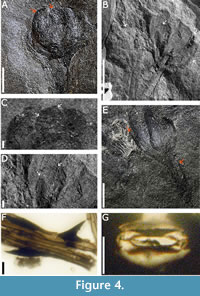
Description. Impression of pedunculated, flower-shaped, hermaphrodite structure, 9.8 mm high and 11.2 mm wide, gynoecium is 8.34 mm long, 3.5 mm at its widest part, and 2.0 mm at the base. Rhomboidal structures of 0.6 mm x 0.6 mm on the distal right area corresponded an interseminal scales. There are observed four free microsporophylls surrounding the gynoecium, up to 6.9-8.9 mm long and 1.1- 1.4 mm wide, forming the androecium (Figure 4A-B). Peduncle is 11 mm long and 5.2 mm wide with two bracts arranged in two different levels at 5.33 mm and 8.7 mm, respectively. It is possible to observe hairs 2.25 mm long on the apex of some of these bracts. The peduncle of paratype CFZCr68 (Figure 4B) is longitudinally striated and 2.6 mm wide and 9 mm long. In Figure 4B, another specimen of W. rosarensis is observed on the left side, which is observed from an abaxial view. Elongated epidermal cells (3.6-6.7 µm wide and 22.4-106.1 µm long) with thin and straight walls and dispersed simple, unicellular, sting-shaped trichomes 8-37 μm long with a rounded base 7-25 μm wide (Figure 4F). Oval syndetocheilic stomata 15-20 μm wide and 20-25 μm long. Open ostioles are 3-5 μm wide and 12-15 μm long. Guard cells are 6-8 μm wide and 20-22 μm long (Figure 4G).
Williamsoniella is characterised by male and female organs united in a small flower-like structure (Thomas, 1915). Three species have been described worldwide, all from the Jurassic of the UK. The first description was of Williamsoniella coronata Thomas from the UK, whose diagnostic characters are the high number (12-16) of microsporophylls; bearing 4-6 synangia with abundant pollen (Thomas, 1915). The gynoecium is pear-shaped, ending in a well-differentiated truncated apex (a corona sensu Thomas, 1915). The number of microsporophylls, as well as the shape of the gynoecium and corona differ from the species described here; in Figure 4B four microsporophylls are observed, in addition to the obovate shape of the gynoecium, which ends in an obtuse apex (Figure 4D) that hardly resembles the structure called corona. Another recorded species is Williamsoniella papillosa Cridland (Harris, 1957), described on disarticulated specimens, characterised by the presence of microsporophylls and large gynoecium, which are twice the size of the specimens from Oaxaca. The name W. papillosa refers to the presence of papillae on the micropyle, but no such structures have been observed in the cuticles obtained so far from the receptaculum, so it is not assigned to this species.
The third species, Williamsoniella lignieri Nathorst, from the Bajocian of Yorkshire, was proposed based on its pollen, which is very similar to that of W. coronata (Harris, 1974). In the specimens reviewed, no pollen was obtained, so it is not assigned to this species either. Another report for this genus is by Flores-Barragan et al. (2017) from the Middle Jurassic Zorrillo Taberna Formation of Oaxaca, Mexico. This structure is like W. rosarensis in the number and size of the microsporophylls, however, it lacks a gynoecium; therefore, it is suggested to refer to this specimen as Williamsonia cf. W. rosarensis. In the Cualac Formation seven specimens have been found that correspond to Williamsonia rosarensis, the flowers have variations in the opening of their microsporophylls, e.g., Figure 4A shows closed flowers, unlike the fossil CFZC68 (Figure 4B), the microsporophylls are separated in size, which allows to observe the gynoecium (arrow); these differences were observed in W. coronata by Thomas (1915), who proposed that they represent up to six stages of maturity; in addition, he commented that from stage 4 both sexes can be differentiated. Finally, the distribution of Williamsoniella rosarensis is restricted to the Cualac Formation, where leaves of Zamites diquiyui Delevoryas and Person, restricted to this locality, are abundant. In previous paragraphs the cuticle of Zamites diquiyui was described, and cuticle from the upper part of the peduncle of Williamsonia rosarensis was also obtained; the presence of sting-shaped trichomes and similar stomata in both taxa was observed, which suggests a possible biological association between both organs.
In the case of W. papillosa, it is associated with Nilssoniopteris major Lindley and Hutton, and the abundant W. coronata Thomas is associated with Nilssoniopteris vittata (Crane and Herendeen, 2009); The absence of these leaf fossil taxa in the flora of Mexico supports the presence of a new species of reproductive structure.
Order INCERTAE SEDIS
Genus MEXIGLOSSA
Delevoryas and Person, 1975
Type species. Mexiglossa varia Delevoryas and Person, 1975
Mexiglossa varia Delevoryas and Person, 1975
Figure 5A-G
 Material examined. CFZCr2-1, Cr4-2, 3, 6, 8, 10, Cr5-1, 2, 3, Cr7-1, 2, 5-8, Cr8-1, 2, 5-8, Cr9-1-3, Cr10-2-10, 13-15, Cr11-1, 2, 5-8, Cr12-1, 3, Cr13-1,2, Cr14-1-3, Cr15-1, 2, Cr17-1, Cr18-2-4, Cr19-3, Cr21-2, 3, 5-9, Cr22-1, 6, 8, Cr24-1, 3, Cr25-1, Cr28-1, 5, 7, 8, 10, 12-14, Cr30-2, 4, 9, 12, Cr38-1, 2, Cr39-1-4, 6, Cr40-1, Cr41-1, Cr42-2-6, 9, 11, 12, Cr43-1-7, Cr48-1-4, Cr49-1, 4, Cr50-2, 4, Cr51-1, 5, 7, 9, 11, 12, 15, 17, Cr52-1, 3, 5, 8, 10, 12, 13, Cr53-2, 5-7, 9, 17, 18, Cr54-1, 2, Cr55-6, Cr56-5, 8, 10,12, Cr55-6, Cr56-5, 8, 10, 11, Cr65-1, Cr66-1, Cr67-1, Cr69-1, Cr70-1, 2, Cr71-1, 2, Cr72-1, Cr73-1, Cr74-1, Cr75-2, 3, Cr76-1, Cr77-1, Cr79-1, Cr80-1, Cr81-1-3, Cr82-1, 2, 9-12, Cr84-1,3, 6, 7, Cr88-1, 2, Cr89-2, 3, Cr90-2, Cr97-3, Cr104-2, Cr105-1, Cr118-1, 2, 4, 6, Cr121-1, 2, Cr123-2, Cr124-2, Cr129-1
Material examined. CFZCr2-1, Cr4-2, 3, 6, 8, 10, Cr5-1, 2, 3, Cr7-1, 2, 5-8, Cr8-1, 2, 5-8, Cr9-1-3, Cr10-2-10, 13-15, Cr11-1, 2, 5-8, Cr12-1, 3, Cr13-1,2, Cr14-1-3, Cr15-1, 2, Cr17-1, Cr18-2-4, Cr19-3, Cr21-2, 3, 5-9, Cr22-1, 6, 8, Cr24-1, 3, Cr25-1, Cr28-1, 5, 7, 8, 10, 12-14, Cr30-2, 4, 9, 12, Cr38-1, 2, Cr39-1-4, 6, Cr40-1, Cr41-1, Cr42-2-6, 9, 11, 12, Cr43-1-7, Cr48-1-4, Cr49-1, 4, Cr50-2, 4, Cr51-1, 5, 7, 9, 11, 12, 15, 17, Cr52-1, 3, 5, 8, 10, 12, 13, Cr53-2, 5-7, 9, 17, 18, Cr54-1, 2, Cr55-6, Cr56-5, 8, 10,12, Cr55-6, Cr56-5, 8, 10, 11, Cr65-1, Cr66-1, Cr67-1, Cr69-1, Cr70-1, 2, Cr71-1, 2, Cr72-1, Cr73-1, Cr74-1, Cr75-2, 3, Cr76-1, Cr77-1, Cr79-1, Cr80-1, Cr81-1-3, Cr82-1, 2, 9-12, Cr84-1,3, 6, 7, Cr88-1, 2, Cr89-2, 3, Cr90-2, Cr97-3, Cr104-2, Cr105-1, Cr118-1, 2, 4, 6, Cr121-1, 2, Cr123-2, Cr124-2, Cr129-1
Description. Impressions and compressions of oblanceolate to lanceolate, even elliptic, leaflets 7-26 cm long and 2-8 cm wide, margin entire, thick midvein tapering towards the apex measuring 3-0.7 mm between eight to 11 anastomosing secondary veins. Mucronate, acuminate, acute, or rounded apices (Figure 5A-B). The cuticle has quadrangular to rectangular epidermal cells 3-5 μm wide and 8-15 μm long, the largest are 9-25 μm wide and 23.2-69.4 μm long. Slightly undulate cell walls with a thickness of 0.5-1.5 μm, and the thickest of up to 9 μm (Figure 5C-E). Sunken stomata without apparent arrangement, stomatal opening is 0.5-3 μm wide and 2-5 μm long, from paracytic to anisocytic. Rounded to oval stomata 10-13 μm wide and 10-20 μm long in some specimens. Thin, 1-2 μm wide and 1.5-3 μm long, and thickened, 4-7 μm wide and 6-10 μm long, guard cells. Anisocytic subsidiary cells 2.5-4.2 μm wide and 5-8 μm long (Figure 5F). Simple trichomes in the midvein 0.4 μm wide and 0.3-0.5 long (Figure 5G).
Discussion. When analyzing the morphological characters of this species, differences are mainly observed in the venation patterns, and there are also anatomical variants in specimens Cr4-6, Cr8-6, Cr11-1, Cr30-13, and Cr70-2 with smaller epidermal cells and very thin cell walls, larger paracytic stomata, and thickened guard cells without trichomes. These data allow the identification two fossil taxa of this genus that probably correspond to independent species. However, this point is beyond the scope of the present work and should be reevaluated in future studies.
Order CZEKANOWSKIALES Pant, 1957
Family LEPTOSTROBASTACEAE Meyen, 1984
Genus CZEKANOWSKIA Heer, 1897
Type species. Czekanowskia setacea Heer, 1876
Czekanowskia sp.
Figure 5H-I
Material examined. CFZCr105, Cr135
Description. Incomplete laciniate leaf impressions, stiff and firm in appearance; bundle of up to nine laciniae united at base, without short outgrowths, largest leaves up to 71.8 mm long and 0.61-2 mm wide, at base the specimen is dichotomous up to twice (Figure 5H). Each lacinia bears a distinct median vein, the apex is unknown. The laciniae diverge at angles of 15°-22°.
Discussion. Leaves of the Czekanowskiales are borne on short shoots, are deciduous and are surrounded by squamiform leaves; the laminae are elongate, simple, or dichotomously segmented, with a single vein at the base of the leaf, usually forked several times, not anastomosing, so that at least one vein remains per leaf segment resulting. There are three genera; the first, Solenites Lindley and Hutton is characterized by a leaf without segmentation; Phoenicopsis Heer with leaves that are narrow wedge-shaped with numerous veins and Czekanowskia Heer with linear, segmented leaves 1 mm wide and a vein (Harris and Miller 1974; Watson et al., 2001). The material studied here has a single vein and leaves with a width of 1.2 mm and bifurcation is therefore assigned to Czekanowskia.
Czekanowskia is distributed in the Northern Hemisphere from the upper Triassic to the Cretaceous (Taylor et al., 2009). Harris and Miller (1974) recorded four species in Yorkshire, UK: C. furcula Harris and Miller, 1 mm wide over most of the leaf, tapering towards the apex and over 120 mm long; it also has eight-leaved bundles; the latter character distinguishes it from the Oaxacan species as seen in the specimen, CFZCr 105-135, Figure 5 H-I. Czekanowskia thomasi Harris and Miller has few, delicate, unevenly sized leaves; its foliage with leaf bundles of seven to eight leaves, and up to 80 mm in length. Most leaf segments fork once and are mostly 0-5 mm wide. The fossil studied here does not have any of the diagnostic characters of C. thomasi the third species, Czekanowskia blacki Harris and Miller, has twice forked leaves with unequal size and about seven leaves per bundle, over 140 mm in length; again, the specimens from Mexico are characterized by leaves of constant size and are therefore not assignable to this species. Finally, Czekanowskia microphylla Harris and Miller has leaves forked three times, in bundles of five leaves. Both of these characters differentiate it from the Cualac Formation specimen. Therefore, the material presented here differs from that of the UK. Other species have been proposed from Russia, many of them from the Cretaceous, and others from the Wealden flora by Watson et al. (2001) where cuticle is used as the main character. As no cuticle could be obtained so far from our specimens the assignment to any species is impossible, and we leave the record at genus level only.
DISCUSSION
Taphocenosis
The flora included in the paleofloristic list of the Cualac Formation comes from two lithofacies. In the Olinalá area, the paleobotanical record is preserved in siltstone interstratified with conglomerate deposited in a high-energy environment that caused wear, transport, and fragmentation of the plants. This is reflected in their poor state of preservation and low diversity and abundance. The characteristics mentioned for the flora of the Cualac Formation in the outcrops of the state of Guerrero suggest that its elements are allochthonous (Silva-Pineda and Gallardo, 1988).
In the region of Tezoatlán, fossil plants were preserved in shale and laminated siltstone, which correspond to the Fl lithofacies reported by Zepeda-Martínez (2021). Miall (2000) reported that these rocks are characteristic of alluvial plains, that is, low-energy depositional environments. This would indicate little transport of the elements that comprise the taphocenosis described for the Rosario locality. This idea is reinforced by the abundance of complete large leaflets of Bennetittales (61.4%) and almost complete well-preserved leaves of Mexiglossa (37 %), which allows defining the flora as paraautochthonous in the sense of Gastaldo (1988).
In the case of Czkanowskiales and Filicales, they have a low abundance (0.6 % and 1 %, respectively), and a high fragmentation level. In the case of Czekanowskia, only united leaves have been found but without dwarf shoots and isolated pinnae, which may be interpreted as uncommon taxa in the taphocenosis of the Rosario locality or as possible allochthonous elements.
Current Knowledge of the Paleoflora
As observed in Table 2, Mexiglossa is the most abundant taxon in this formation. However, its distribution is restricted to the Rosario and Cualac Formations (Tezoatlán paleobasin), which has complicated its taxonomic assignment thus far despite its morphological similarity to Glossopteris Brongniart and Sagenopteris Presl (reticulate veins and compound leaves, respectively), and thus is still considered Incertae sedis until a greater number of characters can be found to elucidate their nature, such as an association with some reproductive structure.
Williamsoniella has a distribution that, in Mexico, is thus far restricted to Oaxaca and, specifically, to the Cualac and Zorrillo-Taberna Formations undifferentiated. Worldwide, it has only been reported from the United Kingdom, in the Grisethorpe and Whitby localities, with two species. In both countries, this genus has low abundance and diversity, unlike Williamsonia and Weltrichia (Brongniart) Harris, which are present in all the Jurassic formations of Mexico. In the localities where these rocks crop out, the diversity of these genera includes more than two species in many cases (Delevoryas 1991; Silva-Pineda et al., 2011; Velasco- de León et al., 2019; Lozano-Carmona, 2021). Even though there are no analyses of reproductive systems of gymnosperms, the discovery of new genera will allow future comparisons of seed number, as reported by Crane and Herenden (2009) (higher in bisexual structures), which is interpreted as an evolutionary strategy of plants to colonize new environments. This is a long-term goal of the present work.
The results allow proposing a floristic assemblage of Zamites diquiyui and Williamsoniella-Mexiglossa as a biofacies indicator in the Cualac Formation.
CONCLUSIONS
The discovery of a new locality and exhaustive sampling has increased the floristic diversity of the Cualac Formation, with the description of: Anomozamites, Cycadolepis, Williamsoniella, and Czekanowskia. It is confirmed that an alluvial or flood plain depositional environment favored quality preservation of the specimens, as well as of small and delicate structures (Williamsoniella). This allowed the emendation of Zamites diquiyui and the proposal of nov. sp. Williamsoniella rosarensis. A possible biological association between the species Zamites diquiyui and Williamsoniella rosarensis is proposed. The Bennettitales and Mexiglossa are established as the most abundant paleofloristic elements in this area. Finally, the cuticular analysis of Mexiglossa allows the identification of two possible independent fossil taxa.
ACKNOWLEDGMENTS
The results of this article are part of the PAPIIT funded project with number IN100721. We also appreciate D.E. Lozano-Carmona y E.Ortega-Chavez for their help in the development of the field work. We thank Dr. M. Zepeda-Martínez for her help on Figure 1. We also thank reviewers of this article, Dr. M. Pole and the anonymous reviewers for the comments and observations that contributed to the writing.
REFERENCES
Behrensmeyer, A.K. and Turner, A. 2023. Bennettitales occurrence data, records downloaded 10 January 2023. Paleobiology Database.
https://paleobiodb.org/#/
Brongniart, A. 1828a. Histoire des végétaux fossiles, ou, Recherches botaniques et géologiques sur les végétaux renfermés dans les diverses couches du globe. Imprimerie de Fain, París.
https://doi.org/10.5962/bhl.title.60992
Brongniart, A. 1828b. Prodrôme d’une histoire des végétaux fossiles. Dictionnaire des Sciences Naturelles, 57:16–212.
Carrasco-Ramírez, R.S., Ferrusquía-Villafranca, I., Buitrón-Sánchez, B.E., and Ruiz-González, J.E. 2016. Litoestratigrafía del grupo Tecocoyunca (jurásico medio) en el área del río Ñumi (cercanías de Tlaxiaco), Oaxaca y consideraciones sobre la distribución regional de su biota. Revista Geológica de América Central, 55:115–136.
Carruthers, W. 1870. On fossil cycadean stems from the secondary rocks of Britain. Transactions of the Linnean Society of London, 26:675–708.
Corona-Esquivel, R.J. 1981. Revista Mexicana de Ciencias Geológicas, 5:17–24.
Crane, P.R. and Herendeen, P.S. 2009. Bennettitales from the Gristhorpe Bed (Middle Jurassic) at Cayton Bay, Yorkshire, UK. American Journal of Botany, 96:284–295.
https://doi.org/10.3732/ajb.0800193
Delevoryas, T. 1991. Investigations of north American cycadeoids: Weltrichia and Williamsonia from the Jurassic of Oaxaca, Mexico. American Journal of Botany, 78:177–182.
https://doi.org/10.1002/j.1537-2197.1991.tb15744.x
Delevoryas, T. and Person, C.P. 1975. Mexiglossa varia gen. et sp. nov.; a new genus of glossopteroid leaves from the Middle Jurassic of Oaxaca, México. Paleontographica, Abteilung B, 154:114–120.
Engler, A. 1892. Anacardiaceae, p. 138–178. In Engler, A. and Prantl, K. (eds.), Die natürlichen Pflanzenfamilien, III. Engelmann, Leipzig.
Erben, H.K. 1956. Estratigrafía y Paleontología del Mesozoico de la Cuenca Sedimentaria de Oaxaca y Guerrero. Especialmente del Jurásico Inferior y Medio, p. 11-36. Conference Proceedings of the XX Congreso Geológico Internacional, México.
Ferguson, D.K. 2005. Plant Taphonomy: Ruminations of the Past, the Present and the Future. Palaios, 20:418–428.
Flores-Barragan, M.A., Velasco de León, M.P., and Corro, O.M.G. 2017. El género Williamsoniella (Thomas) y flora asociada de la Formación Zorrillo-Taberna Indiferenciadas (Jurásico Medio) Oaxaca, México. Paleontología Mexicana, 6:59–65.
Gastaldo, R.A. 1988. A conspectus of phytotaphonomy, p. 14–28. In Di Michele, W.A. and Wing, S.L. (eds.), Methods and applications of plant paleoecology: notes for a short course. Cambridge University Press, UK.
Greenwood, D.R. 1991. The taphonomy of plant macrofossils, p. 141–169. In Donovan, S.K. (ed.), The processes of fossilization. Columbia University Press, UK.
Guzmán, E.J. 1950. Geología del noreste de Guerrero. Boletín Asociación Mexicana de Geólogos Petroleros, 2:95–156.
Guzmán-Madrid, D.S. 2016. Estudio morfológico y cuticular de Mexiglossa varia del Jurásico Inferior de la formación Cuarcítica Cualac, localidad Rosario Nuevo, Oaxaca. Unpublished Bachelor’s Thesis, Facultad de Estudios Superiores Zaragoza, Universidad Nacional Autónoma de México, Mexico.
Guzmán-Madrid, D.S. and Velasco-de León, M.P. 2015. Morfología de Mexiglossa varia del Jurásico Inferior de la Formación Cuarcítica Cualac, Localidad Rosario Nuevo, Oaxaca, p. 63. Conference Proceedings of the XIV Congreso Mexicano de Paleontología. San Luis Potosi, México.
Guzmán-Madrid, D.S. and Velasco-de León, M.P. 2017. Presencia de Zamites diquiyui (Wieland) Person et Delevoryas, en el Jurásico Inferior de la Formación Cuarcítica Cualac, Localidad Rosario Nuevo, Oaxaca, México, p. 63. Conference Proceedings of the XIV Congreso Mexicano de Paleontología. Coahuila, México.
Guzmán-Madrid, D.S. and Velasco-de León, M.P. 2021. Weltrichia magna sp. nov., a new record for the Middle Jurassic of Oaxaca, México. Acta Palaeobotanica, 61:95–106.
https://doi.org/10.35535/acpa-2021-0005
Guzmán-Madrid, D.S., Velasco-De León, M.P., and Lozano-Carmona, D.E. 2019. Nuevo registro de Weltrichia Braun, 1847 emend. Harris, 1969 del Jurásico de Oaxaca (summary), p. 136. Conference Proceedings of the XVI Congreso Mexicano de Paleontología. Chihuahua, México.
Harris, T.M. 1944. A revision of Williamsoniella: Philosophical Transactions of the Royal Society of London B, 231:313–328.
Harris, T.M. 1953. Notes on the Jurassic flora of Yorkshire. Annals and Magazine of Natural History London, 12:33–52.
Harris, T.M. 1969. The Yorkshire Jurassic Flora III. Bennettitales. Trustees of the British Museum (Natural History), London, UK.
Harris, T.M. 1974. Williamsoniella lignieri: its pollen and the compression of spherical pollen grains. Palaeontology, 17:48–125.
Harris, T.M. 1979. The Yorkshire Jurassic Flora. V. Coniferales. Trustees of the British Museum (Natural History), London, UK.
Harris, T.M., Millington, W., and Miller, J. 1974. The Yorkshire Jurassic Flora. IV. Ginkgoales and Czekanowskiales. Trustees of the British Museum (Natural History), London, UK.
Heer, O. 1876. Beiträge zur Jura-Flora Ostsibiriens und des Amurlandes. Mémoires de l’Academie Imperiale des Sciences de Saint-Petersbourg, 22:1–222.
Jímenez-Rentería, J. 2004. Estudio Palinoestratigráfico de los grupos Consuelo y Tecocoyunca (Triásico-Jurásico), en la Barranca Rosario Nuevo, Región de Tezoatlán, al Noroeste del estado de Oaxaca. Unpublished Bachelor’s Thesis, Facultad de Ingeniería, Universidad Nacional Autónoma de México, Mexico.
Kerp, H. 1990. The study of fossil gymnosperms by means of cuticular analysis. Palaios, 5:548–569.
https://doi.org/10.2307/3514861
Kouwenberg, L.L.R., Hines, R.R., and McElwain, J.C. 2007. A new transfer technique to extract and process thin and fragmented fossil cuticle using polyester overlays. Review of Palaeobotany and Palynology, 145:243–248.
https://doi.org/10.1016/j.revpalbo.2006.11.002
Lozano-Carmona, D.E. and Velasco de León, M.P. 2016. Jurassic flora in Southeast Mexico: importance and prospects of recent findings in the Mixteco Terrane. Paleontología Mexicana, 5:87–101.
Lozano-Carmona, D.E. and Velasco-de León, M.P. 2021. Bennettitales from the Middle Jurassic of northwestern Oaxaca, Mexico: Diversity, sedimentary environments, and phytogeography. Journal of South American Earth Sciences, 110:103404.
https://doi.org/10.1016/j.jsames.2021.103404
Lozano-Carmona, D.E., Velasco-de León, M.P., and Flores-Barragan, M.A. 2019. Jurassic Flora in Southern Mexico: Anomozamites Schimper, 1870 emend. Pott et McLoughlin, 2009 from Mixteco Terrane, Phytogeographical Implications of Williamsoniaceae Family (Bennettitales). Open Journal of Geology, 9:142–156.
Mancuso, A.C. and Marsicano, C.A. 2008. Paleoenvironments and taphonomy of a Triassic lacustrine system (Los Rastros Formation, Central-Western Argentina). Palaios, 23:535–547.
Martini, M., Ramírez-Calderón, M., Solari, L., Villanueva-Amadoz, U., Zepeda Martínez, M., Ortega-Gutiérrez, F., and Elías-Herrera, M. 2016. Provenance analysis of Jurassic sandstones from the Otlaltepec Basin, southern Mexico: Implications for the reconstruction of Pangea breakup. Geosphere, 12:1842–1864.
McLoughlin, S., Pott, C., and Sobbe, I.H. 2017. The diversity of Australian Mesozoic bennettitopsid reproductive organs. Paleobiodiversity and Palaeoenviroments, 98:71–95.
https://doi.org/10.1007/s12549-017-0286-z
Meyen, S.V. 1987. Fundamentals of Paleobotany. Chapman and Hall, London, UK.
Miall, A.D. 2000. Principles of Sedimentary Basin Analysis. Springer-Verlag, Berlin.
Morán-Zenteno, D., Caballero-Miranda, J., Silva-Romo, C., Ortega-Gutiérrez, G., and González, T.G. 1993. Jurassic-Cretaceous paleogeographic evolution of the northern Mixteca terrane southern México. Geofísica Internacional, 32:453–473.
Morris, J. 1840. Memoir to illustrate a geological map of Cutch (C.W. Grant). Transactions of Geological Society, 5:289–329.
Nathorst, A.G. 1913. How are the names Williamsonia and Wielandiella to be used? A question of nomenclature. Geologiska Föreningen i Stockholm Förhandlingar, 35:361–366.
https://doi.org/10.1080/11035891309443477
Ortega-Chávez, E., Velasco-de León, M.P., and Jiménez-Rentería, J. 2017. Agathoxylon sp. del Jurásico Inferior, Rosario Nuevo, Oaxaca, México. Paleontología Mexicana, 6:73–77.
Ortiz-Martínez, E.L. 2014. Estudio paleoecológico del jurásico inferior y medio de la zona norte del Estado de Oaxaca. Unpublished Doctoral Thesis, Facultad de Estudios Superiores Zaragoza, Universidad Autónoma de México, Mexico.
Pant, D.D. 1957. The classification of gymnospermous plants. Palaeobotanist, 6:65–70.
Person, C.P. and Delevoryas, T. 1982. The middle Jurassic flora of Oaxaca México. Palaeontographica Abteilung B, 180:82–119.
Phillips, 1829. Illustrations of the geology of Yorkshire, or a description of the strata and organic remains of the Yorkshire Coast: accompanied by a geological map, sections, and plates of the fossil plants and animals. Thomas and Sons, Hogh-Ousegate, York, UK.
Pott, C. and McLoughlin, S. 2009. Bennettitalean foliage from the Rhaetian-Bajocian (latest Triassic-Middle Jurassic) floras of Scania, southern Sweden. Review of Palaeobotany and Palynology, 158:117–166.
https://doi.org/10.1016/j.revpalbo.2009.08.004
Pott, C., McLoughlin, S., and Lindström, A. 2010. Late Palaeozoic foliage from China displays affinities to Cycadales rather than to Bennettitales necessitating a reevaluation of the Palaeozoic Pterophyllum species. Acta Palaeontologica Polonica, 55:157–168.
https://doi.org/10.4202/app.2009.0070
Saporta, G. 1875. Plantes jurassiques, 2. Cycadées. Dans: Paleontologie Franpaise ou Description des Fossiles de la France. Végétaux. Masson, Paris, France.
Schimper, W.P. 1870. Traite de Paléontologie Végétale ou la flore de monde primitif. J.B. Bailliére et Fils, Paris, France.
Silva-Pineda, A. 1969. Plantas fósiles del Jurásico Medio de Tecomatlan, Estado de Puebla. Paleontología Mexicana, 27:1–77.
Silva-Pineda, A. 1970. Plantas fósiles del Jurásico Medio de la región de Tezoatlán, Oaxaca, Sociedad Geológica Mexicana, 1:129–153.
Silva-Pineda, A. 1984. Revisión taxonómica y tipificación de las plantas jurásicas colectadas y estudiadas por Wieland (1914) en la región de El Consuelo, Oaxaca. Paleontología Mexicana, 49:1–103.
Silva-Pineda, A. and González-Gallardo, S. 1988. Algunas Bennettitales (Cycadophyta) y Coniferales (Coniferophyta) del Jurásico Medio del Área de Cualac, Guerrero. Revista Mexicana de Ciencias Geológicas, 7:244–248.
Silva-Pineda, A., Velasco-De León, M.P., Gil, J.A., and Grimaldo, J.R. 2011. Una nueva especie de Weltrichia (Bennettitales) del Jurásico Medio de la Formación Tecomazuchil (Oaxaca, México). Geobios, 44:519–525.
https://doi.org/10.1016/j.geobios.2011.01.004
Taylor, T.N., Taylor, E.L., and Krings, M. 2009. Paleobotany. The biology and evolution of fossil plants. Academic Press, Amsterdam, The Netherlands.
Thomas, H.H. 1915. III. On Williamsoniella, a new type of bennettitalean flower: Philosophical Transactions of the Royal Society of London, B, 207:113–148.
Velasco-de León, M.P., Ortiz-Martínez, E.L., Lozano-Carmona, D.E., and Flores-Barragán, M.A. 2019. Paleofloristic comparison of the Ayuquila and Otlaltepec basins, Middle Jurassic, Oaxaca, México. Journal of South American Earth Sciences, 93:1–13.
https://doi.org/10.1016/j.jsames.2019.04.008
Watson, J., Lydon, S.H., and Harrison, N.A. 2001. A revision of the English Wealden flora, III: Czekanowskiales, Ginkgoales and allied Coniferales. Bulletin Natural History Museum London (Geology), 57:29–82.
Wieland, G.R. 1914. La flora Liásica de la Mixteca Alta: México, Secretaría de Fomento, Instituto Geológico de México, 31:1–165.
Xu, X., Gou, X., Wan, S., Liu, H., Hai-Bo, W., Liu, J., Ding, J., Yang, B., Shu, Y., Hua, Y., and Feng, Z. 2019. Anomozamites (Bennettitales) in China: species diversity and temporo-spatial distribution. Palaeontographica Abteilung B Stuttgart, 300:21-46.
Zepeda-Martínez, M., Martini, M., and Solari, L. 2018. A major provenance change in sandstones from the Tezoatlán basin, southern México, controlled by Jurassic, sinistral normal motion along the Salado River fault: implications for the reconstruction of Pangea. Journal of South American Earth Science, 86:447–460.
https://doi.org/10.1016/j.jsames.2018.07.008
Zepeda-Martínez, M., Martini, M., Solari, L., and Mendoza-Rosales, C.C. 2021. Reconstructing the tectono-sedimentary evolution of the Early–Middle Jurassic Tlaxiaco Basin in southern Mexico: New insights into the crustal attenuation history of southern North America during Pangea breakup. Geosphere, 17:1294–1317.
https://doi.org/10.1130/GES02309.1
