

 Early Pliocene Deer from the Gray Fossil Site, Appalachian Highlands, Tennessee, USA
Early Pliocene Deer from the Gray Fossil Site, Appalachian Highlands, Tennessee, USA
Article number: 28.2.a36
https://doi.org/10.26879/1560
Copyright Society of Vertebrate Paleontology, August 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 9 April 2025. Acceptance: 27 July 2025.
ABSTRACT
The Early Pliocene age Gray Fossil Site (GFS) of northeastern Tennessee is well-known for its diverse and abundant fauna and flora. Perissodactyls, including tapirs and rhinos, are by far the most common large mammals at the site. The only artiodactyls noted from GFS so far are rare remains of several peccary species and a few camelid specimens. This paper describes the first remains of deer from the site, which include a partial juvenile maxilla, an isolated molar, and postcranial elements including humerus, tibiae, astragalus, calcaneum, and phalanges. Comparisons were made to a wide range of modern and fossil cervids, utilizing both qualitative comparisons and quantitative analyses based on measurements from the upper dentition and postcranial elements. The GFS cervid specimens are remarkably similar in both morphology and size to contemporaneous records of the monotypic genus Eocoileus (E. gentryorum) from the Early Pliocene of Florida, which supports taxonomic assignment to that species here. Eocoileus from the Early Pliocene of Tennessee and Florida are generally smaller than extant and fossil cervids in North and Central America studied here, with the exception of Key deer (Odocoileus virginianus clavium), Yucatan brown brocket deer (O. pandora), and Central American red brocket deer (Mazama temama). Dated to 4.9-4.5 Ma, the GFS deer are among the earliest records of the family in North America and combined with similar age occurrences from Florida and Washington these indicate deer dispersed rapidly across the continent in the latest Miocene or Early Pliocene. Overall similarity of the Eocoileus specimens to modern deer (Odocoileus spp.) suggest the former occupied comparable niches, being versatile browsers well-suited to occupy a broad range of habitats. The GFS deer records suggest deer have filled a similar role in the forests of the Appalachian region for nearly 5 million years, persisting through dramatic changes in climate and biota over time.
Joshua X. Samuels. Center of Excellence in Paleontology, East Tennessee State University, Johnson City, Tennessee, U.S.A. and Department of Geosciences, East Tennessee State University, Johnson City, Tennessee, U.S.A. samuelsjx@etsu.edu
Olivia R. Williams. Department of Geosciences, East Tennessee State University, Johnson City, Tennessee, U.S.A. williamsor@etsu.edu
Shay Maden. Center of Excellence in Paleontology, East Tennessee State University, Johnson City, Tennessee, U.S.A. madens@etsu.edu
Blaine W. Schubert. Center of Excellence in Paleontology, East Tennessee State University, Johnson City, Tennessee, U.S.A. and Department of Geosciences, East Tennessee State University, Johnson City, Tennessee, U.S.A. schubert@etsu.edu
Key Words: Cervidae; Eocoileus; ruminant
Final citation: Samuels, Joshua X., Williams, Olivia R., Maden, Shay, and Schubert, Blaine W. 2025. Early Pliocene Deer from the Gray Fossil Site, Appalachian Highlands, Tennessee, USA. Palaeontologia Electronica, 28(2):a36.
https://doi.org/10.26879/1560
palaeo-electronica.org/content/2025/5616-early-pliocene-deer-from-tennessee
Copyright: August 2025 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
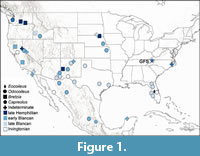 Deer (Cervidae) are the most abundant and species rich group of ruminants in the New World, and represent key components of nearly every terrestrial ecosystem in the Americas and Eurasia (Hanley, 1996; Geist, 1998; Webb, 2000; Rooney, 2001; Côté et al., 2004). The family dispersed to North America from Asia in the late Miocene or earliest Pliocene, with the earliest records likely being Bretzia pseudalces from Washington (Gustafson, 2015; Emery-Wetherell and Schilter, 2020) and Eocoileus gentryorum from Florida (Webb, 2000; Webb et al., 2008). In addition to Bretzia and Eocoileus, Pliocene age occurrences of cervids are known from across the continent (Figure 1; Supplemental Table 1), including many records of Odocoileus (Hibbard, 1941; Robertson, 1976; Webb, 1998; Wheatley and Ruez, 2006; Palma-Ramirez et al., 2023) and one record of Capreolus (Jiménez-Hidalgo and Bravo-Cuevas, 2015). Age and distribution of these records suggest cervids dispersed and diverged rapidly after their arrival in the Americas (Webb, 2000; Croitor, 2022).
Deer (Cervidae) are the most abundant and species rich group of ruminants in the New World, and represent key components of nearly every terrestrial ecosystem in the Americas and Eurasia (Hanley, 1996; Geist, 1998; Webb, 2000; Rooney, 2001; Côté et al., 2004). The family dispersed to North America from Asia in the late Miocene or earliest Pliocene, with the earliest records likely being Bretzia pseudalces from Washington (Gustafson, 2015; Emery-Wetherell and Schilter, 2020) and Eocoileus gentryorum from Florida (Webb, 2000; Webb et al., 2008). In addition to Bretzia and Eocoileus, Pliocene age occurrences of cervids are known from across the continent (Figure 1; Supplemental Table 1), including many records of Odocoileus (Hibbard, 1941; Robertson, 1976; Webb, 1998; Wheatley and Ruez, 2006; Palma-Ramirez et al., 2023) and one record of Capreolus (Jiménez-Hidalgo and Bravo-Cuevas, 2015). Age and distribution of these records suggest cervids dispersed and diverged rapidly after their arrival in the Americas (Webb, 2000; Croitor, 2022).
Today, there are seven cervid species that occur in North America, including two that historically occur in the southern Appalachian Highlands region (Odocoileus virginianus and Cervus canadensis). Deer, particularly O. virginianus, are the most abundant large herbivores in modern communities of the southeastern United States, and represent a large proportion of herbivore biomass within ecosystems (VerCauteren et al., 2011; Greenspoon et al., 2023). While currently abundant and similarly well-represented in Pleistocene age sites from eastern North America, there are few records of the family in the region prior to the Pleistocene. The only occurrences described thus far are Early Pliocene (late Hemphillian or early Blancan) records of Eocoileus gentryorum from the Palmetto Fauna of Florida (Webb, 2000; Webb et al., 2008) and early records of Odocoileus from a number of late Blancan and early Irvingtonian sites in Florida (Robertson, 1976).
Here, we describe the first cervid fossils from the Early Pliocene age Gray Fossil Site of Tennessee, which represent the first ruminants known from the site. The specimens described here are among the oldest records of the family in North America and are the only pre-Pleistocene records of deer from the Appalachian Highlands. These early deer help reveal the origin of a ubiquitous component of Appalachian forests and improve understanding of Neogene faunal evolution in the southern Appalachians.
MATERIALS AND METHODS
Fossil cervid specimens are typically identified and diagnosed based on morphology of the antlers (Gustafson, 2015), which poses a challenge to providing taxonomic identifications of other skeletal material. There are characteristics of the dentition that vary across cervid taxa (Webb, 1998; Webb, 2000; Gustafson, 2015; Emery-Wetherell and Schilter, 2020), but identification of a specimen to species level may be impossible when the type specimen is an antler (Gustafson, 2015). Dental nomenclature used here follows Bärmann and Rössner (2011) and Gustafson (2015); we use dP3 and dP4 to refer to the upper deciduous third and fourth premolars, and M1 and M2 for the upper first and second molars. We have gathered eight measurements from the upper dentition and 18 from postcranial elements of cervids (measurement definitions and references are provided in Supplemental Table 2). Measurements of the teeth and postcranial elements, to the nearest 0.01 mm, were made using Mitutoyo Absolute digital calipers. Measurements of upper teeth include maximum anteroposterior length and transverse breadth taken at occlusal level. Specimens were photographed using a Nikon DSLR D7000 with a Micro Nikkor 60 mm lens.
Comparative specimens of both extant and fossil cervid species were examined at the following institutions: American Museum of Natural History (AMNH); Florida Museum of Natural History, University of Florida (UF); Gray Fossil Site & Museum, Zoology collection (ETMNH Z); National Museum of Natural History, Smithsonian Institution (USNM); University of Michigan Museum of Paleontology (UMMP). Since publication of Webb (2000), additional cervid material has been discovered in the Palmetto Fauna localities in the Central Florida Phosphate Mining District in Polk County, Florida, including elements not represented in the original type and hypodigm series of Eocoileus gentryorum (appendix 4.1 in Webb, 2000). These unpublished UF specimens include a range of dental and postcranial specimens directly comparable to material in the original Palmetto Fauna sample described by Webb (2000).
Some comparative specimens of Pliocene Odocoileus from the Rexroad Fauna of Kansas and Hagerman Local Fauna of Idaho are referred to here as O. brachyodontus, following their original designation; Wheatley and Ruez (2006) consider that taxon to be a nomen dubium, but they only studied the dentition of several specimens and indicated further study was warranted. GFS cervid material was also compared to specimens and measurements from a wide range of publications (including Vislobokova et al., 1995; Gustafson, 2015; Jiménez-Hidalgo and Bravo-Cuevas, 2015; Emery-Whetherell and Schilter, 2020; Palma-Ramirez et al., 2023). A complete listing of studied specimens and measurement data are included in Supplemental Table 3. All specimens described here are housed at the Gray Fossil Site & Museum (cataloged under the acronym ETMNH), Gray, Tennessee.
GEOLOGICAL AND PALEONTOLOGICAL SETTING
Studies of the geology of the Early Pliocene age Gray Fossil Site suggest it formed as an ancient sinkhole with a relatively small and deep lake that filled with sediments over the course of ~4,500 to 11,000 years (Shunk et al., 2006, 2009). Cores from the site reveal rhythmite sediments in the upper lacustrine strata, with alternating layers of fine-grained silty clay and coarse-grained, organic rich sediments (Shunk et al., 2006, 2009). Both fauna and flora from the site are diverse and well-preserved (e.g., Parmalee et al., 2002; Wallace and Wang, 2004; Zobaa et al., 2011; Mead et al., 2012; Ochoa et al., 2012, 2016; Worobiec et al., 2013; Oberg and Samuels, 2022). Both macro- and microfossils of plants reveal a forest dominated by oak (Quercus), hickory (Carya), and pine (Pinus), accompanied by a variety of herbaceous taxa (Ochoa et al., 2016; and references therein). Ochoa et al. (2016) interpreted the environment at the site as a woodland or woodland savanna with frequent disturbance. Isotopic analyses of ungulate and proboscidean teeth from GFS support the presence of relatively dense forest, but suggest more open grass-dominated habitats occurred within the dispersal range of large herbivores (DeSantis and Wallace, 2008). Both isotopic (DeSantis and Wallace, 2008) and ecometric analyses (Schap et al., 2021) suggest the climate had little seasonal variation in temperature and precipitation. Among the fauna are multiple organisms that are indicative of aquatic environments, specifically fish, neotenic salamanders, aquatic turtles, Alligator, a desman mole, and beavers (Parmalee et al., 2002; Boardman and Schubert, 2011; Mead et al., 2012; Bourque and Schubert, 2015; Jasinski, 2018; Oberg and Samuels, 2022; Maden, 2023). Presence of reptiles intolerant of extended freezing conditions (e.g., Alligator and Heloderma) (Parmalee et al., 2002; Mead et al., 2012) and mammals characteristic of forested habitats (e.g., tree squirrels, flying squirrels, tapirs, and the ailurid Pristinailurus) (Wallace and Wang, 2004; DeSantis and MacFadden, 2007; Hulbert et al., 2009; Crowe, 2017; Samuels et al., 2018; Grau-Camats et al., 2025) further support interpretation of the site as a relatively warm and closed forest. A range of large and small mammals from GFS represent immigrant taxa from Eurasia, including ailurids, meline badgers, several moles, and a giant flying squirrel (Wallace and Wang, 2004; Oberg and Samuels, 2022; Grau-Camats et al., 2025). In total, the site’s flora and fauna represents a unique combination among biotas of North America.
There has been some controversy over age estimates for the Gray Fossil Site, based primarily on occurrence of Teleoceras (Short et al., 2019). The extinction of rhinoceroses in North America has been traditionally considered to have occurred at the end of the Hemphilllian NALMA (Wood et al., 1941; Lundelius et al., 1987), and the absence of rhinoceroses has been interpreted as characterizing Blancan faunas. However, Teleoceras is now known from multiple sites that include Blancan taxa (Bell et al., 2004; Farlow et al., 2001, 2010; Samuels et al., 2018; Martin, 2021), and Gustafson (2012) asserted that the presence of Teleoceras at a site is insufficient justification for assigning a Hemphillian NALMA age. Improved sampling and understanding of microfauna at both Pipe Creek Sinkhole in Indiana and GFS have led to revised interpretations of their ages. Martin (2021) and Martin and Kelly (2023) place Pipe Creek Sinkhole as a basal Blancan age site based on occurrences of Ogmodontomys and Pliophanacomys at the site, as occurrences of arvicolines south of 55°N latitude are used to define the Blancan NALMA (Bell et al., 2004). As a result of those arvicoline occurrences, Martin (2021) and Martin and Kelly (2023) interpret Teleoceras as a holdover taxon at Pipe Creek Sinkhole. Arvicolines have not been recovered from the Gray Fossil Site, but several other taxa characteristic of early Blancan faunas do occur, including the leporids Alilepus vagus and Notolagus lepusculus, cricetids Neotoma, Peromyscus, and Symmetrodontomys, and the mephitid Buisnictis breviramus (Samuels et al., 2018; Samuels and Schap, 2021; Xu, 2023).
Based on biochronology of mammal taxa with good fossil records and limited chronologic ranges, the age of GFS is currently estimated to be Early Pliocene, between 4.9 and 4.5 Ma, near the Hemphillian - Blancan transition (Samuels et al., 2018; Samuels and Schap, 2021; Xu, 2023). We elect for a conservative interpretation of the age of GFS (Early Pliocene, either in the latest Hemphillian or early Blancan NALMA), but note that none of the mammal taxa from the site are restricted to the Miocene or the Hemphillian NALMA, while multiple taxa are restricted to the Pliocene and characteristic of Blancan faunas. It is possible that GFS dates to the end of the Hemphillian, and the site’s records of Symmetrodontomys (also known from Pipe Creek), Peromyscus, and Notolagus lepusculus represent first appearances of those taxa. Alternatively, the records of Teleoceras at the site may represent the last known rhinoceroses in North America (Short et al., 2019), having survived into the early Blancan where they co-occurred with a number of novel rodent and lagomorph taxa. Both GFS and Pipe Creek Sinkhole are geographically distant from most other records of Teleoceras, and the forested habitats of eastern North America may have represented a refugium where those rhinoceroses survived later than most other parts of the continent.
SYSTEMATIC PALEONTOLOGY
Class MAMMALIA Linnaeus, 1758
Order ARTIODACTYLA Owen, 1848
Family CERVIDAE Goldfuss, 1820
Subfamily CAPREOLINAE Brookes, 1828
Genus EOCOILEUS Webb, 2000
Eocoileus gentryorum Webb, 2000
(Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Table 1, Table 2)
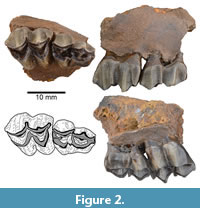 Referred Specimens. ETMNH 6166 partial right maxilla with dP3 and dP4; ETMNH 40000 left M2; ETMNH 10559 distal right humerus; ETMNH 8030, ETMNH 14923 right tibia; ETMNH 17218 left astragalus; ETMNH 12448 partial right calcaneum; ETMNH 19528 metapodial distal condyle; ETMNH 10546, ETMNH 13918 proximal phalanx
Referred Specimens. ETMNH 6166 partial right maxilla with dP3 and dP4; ETMNH 40000 left M2; ETMNH 10559 distal right humerus; ETMNH 8030, ETMNH 14923 right tibia; ETMNH 17218 left astragalus; ETMNH 12448 partial right calcaneum; ETMNH 19528 metapodial distal condyle; ETMNH 10546, ETMNH 13918 proximal phalanx
Locality. Gray Fossil Site, Washington County, Tennessee.
Age. Early Pliocene (late Hemphillian or earliest Blancan).
Diagnosis. In the GFS cervid the upper molar protocone is fused to the metaconule, there is a small entostyle between the lingual crescents, and the posterior crest extending from the upper molar protocone is bifurcated, which are traits common among capreoline cervids. In contrast to the GFS cervid, Eocoileus gentryorum, and extant Odocoileus, the upper molar protocone and metaconule are unfused in Cervus, Bretzia, and dromomerycids. The GFS cervid upper molar displays rugose anterior and posterior cingula bearing small cuspules, similar to some specimens of E. gentryorum from Florida. These cingula are distinct, but not rugose in Bretzia and are weak or absent in studied specimens of Odocoileus, Capreolus, and Mazama. The GFS cervid astragalus has a straight proximal edge of the sustentacular facet, as is typical of capreolines. In the GFS cervid and E. gentryorum the sustentacular facet of the astragalus is more rectangular in shape (anteroposteriorly elongate) than in Bretzia and Odocoileus. Morphological features of both the teeth and postcranial elements, along with size of the specimens, are within the range of variation of known specimens of E. gentryorum and support assignment to that species.
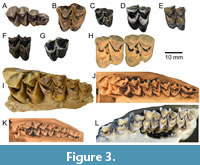 Description–upper dentition. ETMNH 6166 represents a fragmentary portion of the right maxilla with the dP3 and dP4 preserved (Figure 2 and Figure 3), and ETMNH 40000 is an isolated left M2 (Figure 3). The upper dentition is selenodont and brachydont, with crescentic protocone and metaconule lingually and rhomboidal paracone and metacone labially. Crests extend from the anterior and posterior margins of each of those cusps. Enamel on the cusps is not distinctly crenulate, and there are no cingula surrounding the lingual cusps. The dP3 of ETMNH 6166 is relatively asymmetric and narrower anteriorly than posteriorly. The paracone and protocone of the dP3 are distinctly smaller than the metacone and metaconule. The parastyle, mesostyle, and metastyle of the dP3 are all distinct, and the paracone rib is more prominent than the indistinct metacone rib. A small spur extends labially from the posterior crest of the metaconule. In contrast to the relatively asymmetric dP3, the dP4 of ETMNH 6166 displays a trapezoidal shape with protocone and metaconule of similar size and paracone and metacone of similar size. The dP4 parastyle and mesostyle are prominent, and both are larger than the metastyle. The paracone rib and metacone rib of the dP4 are also distinct, though the former is more pronounced. The dP4 has a distinct entostyle present between the paracone and metaconule. The posterior crest of the protocone of the dP4 shows no evidence of bifurcation or presence of a protoconal fold, and there is no metaconule spur extending from the posterior crest of the metaconule. Though the dP4 is only slightly worn, the posterior crest of the protocone is fused to the anterior crest of the metaconule. The dP4 lacks both anterior and posterior cingula.
Description–upper dentition. ETMNH 6166 represents a fragmentary portion of the right maxilla with the dP3 and dP4 preserved (Figure 2 and Figure 3), and ETMNH 40000 is an isolated left M2 (Figure 3). The upper dentition is selenodont and brachydont, with crescentic protocone and metaconule lingually and rhomboidal paracone and metacone labially. Crests extend from the anterior and posterior margins of each of those cusps. Enamel on the cusps is not distinctly crenulate, and there are no cingula surrounding the lingual cusps. The dP3 of ETMNH 6166 is relatively asymmetric and narrower anteriorly than posteriorly. The paracone and protocone of the dP3 are distinctly smaller than the metacone and metaconule. The parastyle, mesostyle, and metastyle of the dP3 are all distinct, and the paracone rib is more prominent than the indistinct metacone rib. A small spur extends labially from the posterior crest of the metaconule. In contrast to the relatively asymmetric dP3, the dP4 of ETMNH 6166 displays a trapezoidal shape with protocone and metaconule of similar size and paracone and metacone of similar size. The dP4 parastyle and mesostyle are prominent, and both are larger than the metastyle. The paracone rib and metacone rib of the dP4 are also distinct, though the former is more pronounced. The dP4 has a distinct entostyle present between the paracone and metaconule. The posterior crest of the protocone of the dP4 shows no evidence of bifurcation or presence of a protoconal fold, and there is no metaconule spur extending from the posterior crest of the metaconule. Though the dP4 is only slightly worn, the posterior crest of the protocone is fused to the anterior crest of the metaconule. The dP4 lacks both anterior and posterior cingula.
The GFS M2 (ETMNH 40000) is heavily worn, and it displays a trapezoidal shape with protocone and metaconule of similar size and paracone and metacone of similar size. A distinct entostyle is present between the paracone and metaconule of the M2. The M2 posterior crest of the protocone is bifurcated in ETMNH 40000, forming a small enamel island (protoconal fold) where it fuses to the anterior crest of the metaconule. There is a small metaconule spur in ETMNH 40000 that extends labially from the posterior crest of the metaconule. The M2 (ETMNH 40000) has a clear anterior cingulum with a transversely elongate cingular cuspule along the anterior margin of the protocone, as well as a rugose posterior cingulum on the lingual and posterior margins of the metaconule bearing small cuspules.
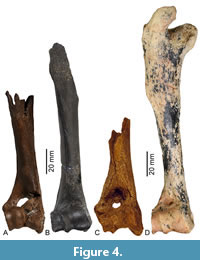 Postcranial Material. A partial right humerus recovered from the site (ETMNH 10559, Figure 4) preserves the distal end and most of the diaphysis. The proximal end of the bone is missing, there is some weathering and damage to the medial portion of the trochlea, and the thin bone of the olecranon and coronoid fossa has eroded away to form an oval-shaped hole just proximal to the distal articulation. The distal articulation of ETMNH 10559 shows morphology typical of ruminants, with a capitulum characterized by a raised spline (carina of Gustafson, 2015) and distinct grooves in the capitulum and trochlea, which restrict mediolateral movements of the elbow joint. The lateral epicondyle and supracondylar ridge are fairly prominent, as are the medial projection associated with the origin of the flexor muscle complex and medial caudal protuberence.
Postcranial Material. A partial right humerus recovered from the site (ETMNH 10559, Figure 4) preserves the distal end and most of the diaphysis. The proximal end of the bone is missing, there is some weathering and damage to the medial portion of the trochlea, and the thin bone of the olecranon and coronoid fossa has eroded away to form an oval-shaped hole just proximal to the distal articulation. The distal articulation of ETMNH 10559 shows morphology typical of ruminants, with a capitulum characterized by a raised spline (carina of Gustafson, 2015) and distinct grooves in the capitulum and trochlea, which restrict mediolateral movements of the elbow joint. The lateral epicondyle and supracondylar ridge are fairly prominent, as are the medial projection associated with the origin of the flexor muscle complex and medial caudal protuberence.
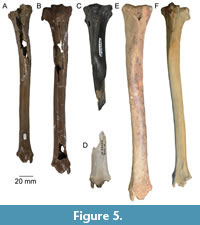 Two nearly complete right tibiae have been recovered (ETMNH 8030 and ETMNH 14923, Figure 5). The proximal epiphysis and portions of the proximal portion of the diaphysis are missing in ETMNH 8030, while some portions of the proximal diaphysis and distal diaphysis are missing in ETMNH 14923. Each tibia is particularly gracile, and the tibial spine extends from the proximal epiphysis to nearly halfway down the diaphysis. The proximal end of ETMNH 14923 has a fairly broad and low intercondylar eminence, and the lateral surface has a prominent facet for articulation with the head of the fibula. The distal end has a prominent medial malleolus and deep fossae for articulation with the condyles of the astragalus.
Two nearly complete right tibiae have been recovered (ETMNH 8030 and ETMNH 14923, Figure 5). The proximal epiphysis and portions of the proximal portion of the diaphysis are missing in ETMNH 8030, while some portions of the proximal diaphysis and distal diaphysis are missing in ETMNH 14923. Each tibia is particularly gracile, and the tibial spine extends from the proximal epiphysis to nearly halfway down the diaphysis. The proximal end of ETMNH 14923 has a fairly broad and low intercondylar eminence, and the lateral surface has a prominent facet for articulation with the head of the fibula. The distal end has a prominent medial malleolus and deep fossae for articulation with the condyles of the astragalus.
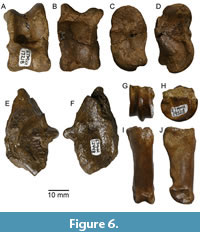 The left astragalus (ETMNH 17218, Figure 6A-D) has some erosion evident across its surface, but the morphology of the bone is preserved with the exception of the medial tibial shelf, which is partially lost. The lateral and medial proximal condyles are prominent, and the proximal trochlear groove is relatively broad. There is a deep and mediolaterally broad interarticular fossa, as well as a prominent tibial stop (Gustafson, 2015). The lateral cubonavicular condyle has a distinct distal keel, which is relatively medially placed. The distal trochlea, between the lateral and medial cubonavicular condyles, is shallow. The sustentacular facet is roughly rectangular in shape (anteroposteriorly elongate) and has a relatively small lateral process on its proximal corner, though this feature is somewhat worn. The proximal edge of the sustentacular facet is relatively straight and at a right angle to the long axis of the bone.
The left astragalus (ETMNH 17218, Figure 6A-D) has some erosion evident across its surface, but the morphology of the bone is preserved with the exception of the medial tibial shelf, which is partially lost. The lateral and medial proximal condyles are prominent, and the proximal trochlear groove is relatively broad. There is a deep and mediolaterally broad interarticular fossa, as well as a prominent tibial stop (Gustafson, 2015). The lateral cubonavicular condyle has a distinct distal keel, which is relatively medially placed. The distal trochlea, between the lateral and medial cubonavicular condyles, is shallow. The sustentacular facet is roughly rectangular in shape (anteroposteriorly elongate) and has a relatively small lateral process on its proximal corner, though this feature is somewhat worn. The proximal edge of the sustentacular facet is relatively straight and at a right angle to the long axis of the bone.
The partial right calcaneum (ETMNH 12448) has the articular end preserved well, but most of the body and tuber calcanei are missing (Figure 6E-F). The preserved portion of the calcaneal body suggests the bone would have tapered toward the tuber calcanei. The sustentaculum is broken but appears relatively small and obliquely oriented to the long axis of the bone. The sulcus for the deep flexor tendon is well-developed. The fibular condyle is particularly prominent and S-shaped, and there is a small shelf between the fibular facet and the interosseus fossa. The astragalar facet is broad and slightly convex. The cubonavicular facet faces relatively distally and extends onto a prominent triangular distal projection of the calcaneum.
The only metapodial element is ETMNH 19528, which represents a metapodial distal condyle (Figure 6G-H). The inner and outer trochlear surfaces are similarly broad and separated by a prominent and steeply inclined ridge. There are small, but distinct pits at the posterior (palmar) end of the trochlear surfaces. Though fragmentary, the shape of ETMNH 19528 indicates the presence of a pronounced intertrochlear notch.
There are two proximal phalanx specimens (ETMNH 10546, ETMNH 13918), and both taper gently distally (Figure 6I-J). The proximal articular surfaces are concave with a deep midsagittal groove, and the lateral glenoid is cavity shallower than the medial one. Small articular facets are present at the proximal posterior (palmar) surface. The distal articular surface has distinct medial and lateral condyles separated by a broad groove. Just anterior to the distal articulation on the posterior (palmar surface) is a distinct projection for insertion of digital flexors.
Comparisons. Characteristics of the GFS cervid specimens are consistent with those of other early cervids in North America, and details of the dentition and postcrania facilitate comparisons to both fossil and extant cervid taxa. Upper dentition of Eocoileus has not been previously figured nor have any of the deciduous teeth, specimens from Tennessee and Florida as well as a range of comparative taxa are figured here (Figure 2 and Figure 3). The dP3 of ETMNH 6166 is narrower anteriorly (smaller protocone and paracone) than in other cervids studied (Table 1, Figure 3, but note that dP3 was only present in specimens of Odocoileus virginianus, O. hemionus, and Mazama temama). The relatively small dP3 protocone of ETMNH 6166 is similar to the dP3 protocone of other pecorans like the Dromomerycidae, possibly reflecting retention of a plesiomorphic state in the deciduous dentition of this early cervid. In both GFS cervid specimens, the posterior crest of the upper molar protocone is fused with anterior crest of the metaconule early in wear. Fusion of the protocone to metaconule is typical of worn upper molars in cervids, but does show variability in studied specimens of Eocoileus, Odocoileus, Capreolus, and Mazama (illustrated in Figure 3). According to Gustafson (2015), the protocone and metaconule are always unfused in Bretzia, and those cusps are similarly unfused in the upper molars of dromomerycids. In both upper molar specimens from GFS, there is a small entostyle present between lingual crescents, as is typical of cervids. The upper molar entostyle is always present in Bretzia (Gustafson 2015), but occurrence is variable in some other taxa: Eocoileus gentryorum (present in 3 of 5), Mazama temama (present in 2 of 4), and the small subspecies of white-tailed deer from the Yucatan of Mexico (O. virginianus yucatanensis) (present in 1 of 4). The posterior crest extending from the M2 protocone is bifurcated in ETMNH 40000, which is typical of cervids. Note that this bifurcation does variably occur in studied specimens of Eocoileus gentryorum (bifurcated in 4 of 5) and Capreolus capreolus (bifurcated in 3 of 4). The occurrence of anterior and posterior cingula varies in specimens of Eocoileus gentryorum from Florida (Figure 3). In ETMNH 40000 (Figure 3B) there is a rugose posterior cingulum on the lingual and posterior margins of the metaconule bearing small cuspules, and similar features are present in UF 27518 from Florida (Figure 3G). While anterior and posterior cingula are present in Bretzia (Gustafson, 2015), similarly rugose cingula were not seen among studied samples of any other cervid taxa from North America. More pronounced cingula are present in some earlier fossil cervids from Eurasia (Vislobokova, 2012; Croitor, 2014, 2018). It is important to note that the unusual features of ETMNH 6166 (small dP3 protocone, lack of bifurcation of posterior crest of dP4 protocone) led the lead author to initially identify that specimen as a dromomerycid (noted in the biostratigraphic list of Samuels et al., 2018), but broader examination of those features has documented their occurrence in other cervid samples.
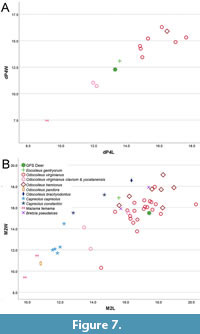 The sizes of upper teeth from GFS are interesting, as ETMNH 6166 is similar in size to a dP4 specimen (UF 27479) in the sample of Eocoileus gentryorum from Florida (Table 1, Figure 7A) and smaller than Pliocene Bretzia pseudalces and Capreolus constantini, and modern Odocoileus other than O. pandora, O. virginianus clavum, and O. virginianus yucatanensis. In contrast, ETMNH 40000 is longer but narrower than M2 specimens of E. gentryorum from Florida (Figure 7B), comparable in size to modern and Pleistocene records of Odocoileus virginianus. The disparate sizes of these specimens may reflect sexual dimorphism or other forms of intraspecific variation in the sample of deer from GFS. Among the sample of O. virginianus studied here there is large variation in tooth size (Figure 7, Supplemental Table 3), greater than 25% among individuals (even when excluding O. virginianus clavium and O. virginianus yucatanensis). While ETMNH 40000 is relatively large, nearly as large as Pliocene Odocoileus brachyodontus (UMMP 28140) and early Pleistocene O. virginianus (UF 276885), the morphological features of the specimen are within the range of variation of known specimens of E. gentryorum and support assignment to that species rather than some other cervid taxon.
The sizes of upper teeth from GFS are interesting, as ETMNH 6166 is similar in size to a dP4 specimen (UF 27479) in the sample of Eocoileus gentryorum from Florida (Table 1, Figure 7A) and smaller than Pliocene Bretzia pseudalces and Capreolus constantini, and modern Odocoileus other than O. pandora, O. virginianus clavum, and O. virginianus yucatanensis. In contrast, ETMNH 40000 is longer but narrower than M2 specimens of E. gentryorum from Florida (Figure 7B), comparable in size to modern and Pleistocene records of Odocoileus virginianus. The disparate sizes of these specimens may reflect sexual dimorphism or other forms of intraspecific variation in the sample of deer from GFS. Among the sample of O. virginianus studied here there is large variation in tooth size (Figure 7, Supplemental Table 3), greater than 25% among individuals (even when excluding O. virginianus clavium and O. virginianus yucatanensis). While ETMNH 40000 is relatively large, nearly as large as Pliocene Odocoileus brachyodontus (UMMP 28140) and early Pleistocene O. virginianus (UF 276885), the morphological features of the specimen are within the range of variation of known specimens of E. gentryorum and support assignment to that species rather than some other cervid taxon.
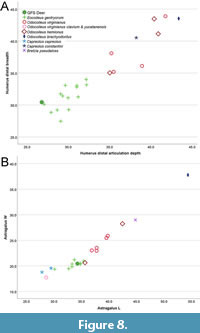 The right humerus from GFS, ETMNH 10559, is very similar in size to specimens of Eocoileus gentryorum from Florida (Table 2, Figure 8), but far smaller than Pliocene records of Odocoileus brachyodontus, Capreolus constantini, and Bretzia pseudalces and modern O. virginianus and O. hemionus. The structure of the distal articulation of ETMNH 10559 is typical of cervids (Figure 4), with the medial trochlea substantially mediolaterally broader than the lateral capitulum. The lateral epicondyle and supracondylar ridge of ETMNH 10559 are relatively prominent in comparison to extant Odocoileus, which is also true of late Pliocene O. brachyodontus (USNM 299483) from the Hagerman local fauna of Idaho (Figure 4). Similarly, the medial projection associated with the origin of the flexor muscle complex is prominent in ETMNH 10559, O. brachyodontus USNM 299483, and some extant O. virginianus, much more so than in studied specimens of Eocoileus gentryorum from Florida or B. pseudalces (Gustafson, 2015). The medial caudal protuberance is large in ETMNH 10559, similar to that of O. virginianus (Jacobson, 2003) and larger than in O. hemionus.
The right humerus from GFS, ETMNH 10559, is very similar in size to specimens of Eocoileus gentryorum from Florida (Table 2, Figure 8), but far smaller than Pliocene records of Odocoileus brachyodontus, Capreolus constantini, and Bretzia pseudalces and modern O. virginianus and O. hemionus. The structure of the distal articulation of ETMNH 10559 is typical of cervids (Figure 4), with the medial trochlea substantially mediolaterally broader than the lateral capitulum. The lateral epicondyle and supracondylar ridge of ETMNH 10559 are relatively prominent in comparison to extant Odocoileus, which is also true of late Pliocene O. brachyodontus (USNM 299483) from the Hagerman local fauna of Idaho (Figure 4). Similarly, the medial projection associated with the origin of the flexor muscle complex is prominent in ETMNH 10559, O. brachyodontus USNM 299483, and some extant O. virginianus, much more so than in studied specimens of Eocoileus gentryorum from Florida or B. pseudalces (Gustafson, 2015). The medial caudal protuberance is large in ETMNH 10559, similar to that of O. virginianus (Jacobson, 2003) and larger than in O. hemionus.
The cervid tibiae recovered from GFS (ETMNH 8030, 14923) are both from the right side, indicating they come from two different individuals. These tibiae are relatively gracile (Figure 5, Table 2), like studied specimens of Eocoileus gentryorum from Florida and extant Capreolus capreolus, and slenderer than those of Odocoileus virginianus, C. constantini, and Bretzia pseudalces. As in E. gentryorum from Florida and O. virginianus, the tibial spine (=anterior tibial crest of Gustafson, 2015) extends from the proximal epiphysis to nearly halfway down the diaphysis, which contrasts with the relatively shorter tibial crest of B. pseudalces, C. capreolus, and C. constantini (Gustafson, 2015; Jiménez-Hidalgo and Bravo-Cuevas, 2015). The relatively broad and low intercondylar eminence in the proximal end of ETMNH 14923 (Figure 5B) is similar to specimens of E. gentryorum from Florida, B. pseudalces, and extant Odocoileus, as is the structure of the distal end, which has a prominent medial malleolus and deep fossae for articulation with the astragalus.
Morphology and size of the GFS astragalus (ETMNH 17218, Figure 6A-D) falls within the range of variation of specimens of Eocoileus gentryorum from Florida and is smaller than the studied specimens of Odocoileus hemionus and O. virginianus from multiple subspecies, with the exception of the Key Deer O. virginianus clavium (Table 2, Figure 8B). The astragali of E. gentryorum and O. virginianus are relatively anteroposteriorly elongate compared to O. brachyodontus, Bretzia, and Capreolus. As is the case for specimens of Eocoileus gentryorum from Florida, the sustentacular facet of ETMNH 17218 is more rectangular in shape (anteroposteriorly elongate) than in Bretzia and Odocoileus (O. brachyodontus and O. virginianus). The straight proximal edge of the sustentacular facet in ETMNH 17218 is also seen in Odocoileus, Mazama, Capreolus, and Bretzia (Morejohn and Dailey, 2004; Gustafson, 2015; Palma-Ramírez et al., 2023). Jacobson (2003) noted a well-defined ridge running at an angle medially to laterally along the distal edge of the lateral proximal condyle in O. virginianus, but not O. hemionus. ETMNH 17218 has erosion to its surface in that area, precluding confident assessment of that feature, but that ridge is present in E. gentryorum (UF 27516) and early Pleistocene O. virginianus (UF 276402) from Florida. The incomplete preservation of the right calcaneum from GFS (ETMNH 12448, Figure 6E-F) precludes detailed comparisons to other cervid taxa. Morphological features and size of ETMNH 12448 fall within the range of variation observed among studied specimens of Eocoileus gentryorum from Florida (UF 27530, 125058, 135604); the GFS calcaneum is also similar in morphology to an incomplete calcaneum of Odocoileus brachyodontus (USNM 418959) from the Pliocene of Idaho, but the GFS specimen is substantially smaller.
ETMNH 19528 (Figure 6G-H), a metapodial distal condyle, is similar in morphology to several metatarsal specimens of Eocoileus gentryorum from Florida (UF 134798, 134199). Those specimens all have distal condyles with similarly broad inner and outer trochlear surfaces separated by a steeply inclined ridge, as well as small pits at the posterior ends of the trochlear surfaces. Both metatarsals of Eocoileus also display a deep intertrochlear notch, as is suggested by the morphology of ETMNH 19528. The GFS and Florida Eocoileus specimens have distal condyles similar in size, and these are all substantially smaller than in studied specimens of Odocoileus (O. virginianus, O. hemionus, and O. brachyodontus) and dimensions reported for Bretzia (Gustafson, 2015) and Capreolus (Vislobokova et al., 1995; Jiménez-Hidalgo and Bravo-Cuevas, 2015). The two proximal phalanges from GFS (ETMNH 10546, ETMNH 13918) are similar in length and general shape to a phalanx identified as cf. Eocoileus sp. from Fort Green Mine in Florida (UF 216983). The morphology of both the GFS and Florida specimens referred to Eocoileus is similar to studied specimens of extant and fossil Odocoileus (O. virginianus, O. hemionus, and O. brachyodontus). The GFS phalanges are more mediolaterally robust than UF 216983 (Table 2), showing similar robustness to that of Capreolus constantini from Mexico (Jiménez-Hidalgo and Bravo-Cuevas, 2015). With the exception of Key deer (O. virginianus clavium), the GFS and Florida specimens are also smaller than all studied specimens of Odocoileus (Table 2), those noted by Palma-Ramírez et al. (2023), and proximal phalanges reported for Capreolus (Jiménez-Hidalgo and Bravo-Cuevas, 2015). The insertion for the distal flexor on the posterior surface of the phalanx is more prominent in the GFS specimens than in the Florida phalanx, similarly to that of O. brachyodontus from Idaho (USNM 418959). It is also worth noting that the morphology of the proximal articular surfaces of ETMNH 10546 are compatible for articulation with the distal metapodial condyle of ETMNH 19528 (Figure 6I-J).
DISCUSSION
The rare records of a cervid from the Gray Fossil Site, referred here to Eocoileus gentryorum, are the first ruminants recovered from this well-known site. Much has been written about the GFS tapirs (Hulbert et al., 2009; McConnell and Zavada, 2013; Maclaren et al., 2018; DeSantis et al., 2020; Schap and Samuels, 2020) and rhinos (Short et al., 2019), and those large perissodactyls are clearly the predominant members of the large herbivore guild. Tapirs at GFS have an MNI of >160 and rhinos 11 (Inabinett, personal commun.). Equids, which are particularly abundant at most fossil sites from the Early Pliocene, are only currently represented by 15 total specimens from GFS, which have been referred to Cormohipparion emsliei by Wallace et al., 2011 (in Schubert and Mead, eds. 2011). Far less is known about the artiodactyls from GFS, the best known of which are tayassuids (Doughty et al., 2018), represented by at least four individuals. Rare remains of camel have also been noted from GFS (Wallace and Wang, 2004), currently being described as representing two taxa (Maden and Samuels, In Preparation).
Morphology and size of the GFS deer specimens are consistent with the range of variation of contemporaneous Eocoileus gentryorum from Florida. Unfortunately, the type specimen and figured specimens in the original description of E. gentryorum (Webb, 2000) do not overlap with the GFS materials, but elements referred to E. gentryorum in the original publication (appendix 4.1 of Webb, 2000) and additional Palmetto fauna specimens referred to Eocoileus in the UF collection match GFS dental and postcranial specimens well. While an antler or cranium from GFS might someday indicate that this represents a distinct species, at this time the most parsimonious conclusion is that the Tennessee and Florida samples are one species, thus we refer to the GFS cervid as Eocoileus gentryorum. These Early Pliocene deer from Tennessee and Florida are smaller than extant and fossil cervids in North and Central America, with the exception of Key deer (O. virginianus clavium), Yucatan brown brocket deer (Odocoileus pandora), and Central American red brocket deer (Mazama temama).
The Eocoileus specimens described here from Tennessee are among the oldest records of Cervidae in North America and the only pre-Pleistocene records of deer from the Appalachian region. The earliest records of the family Cervidae in North America date to the late Miocene or earliest Pliocene, including occurrences of Bretzia pseudalces in Washington (Gustafson, 2015; Emery-Wetherell and Schilter, 2020) and Eocoileus gentryorum in Florida (Webb, 2000; Webb et al., 2008). Along with the early occurrence of Eocoileus at GFS are early records of Bretzia from California, New Mexico, and Nebraska. In addition to Bretzia and Eocoileus, other Pliocene age occurrences of cervids include Odocoileus (Hibbard, 1941; Robertson, 1976; Webb, 1998; Wheatley and Ruez, 2006; Palma-Ramirez et al., 2023) and Capreolus (Jiménez-Hidalgo and Bravo-Cuevas, 2015). In sum, the North American fossil record of cervids indicates rapid dispersal across the continent in the late Miocene or Early Pliocene (Figure 1, Supplemental Table 1) and early divergence of modern taxa (Webb, 2000; Croitor, 2022).
Deer (Cervidae) are the most abundant and species-rich group of ruminants in the New World, and represent key components of nearly every terrestrial ecosystem in the Americas and Eurasia (Hanley, 1996; Geist, 1998; Webb, 2000; Rooney, 2001; Côté et al., 2004). Similarity of the GFS deer to Eocoileus and extant deer (Odocoileus spp.) suggest they occupied similar niches, providing an important point of continuity between Early Pliocene and modern mammal communities in the Appalachian Highlands. These deer are versatile browsers capable of occupying a wide range of habitats, and these new fossils may indicate they have survived in the Appalachian forests through dramatic climate changes of the Pliocene and Pleistocene. Though fragmentary, these early deer fossils help reveal the origin of a ubiquitous component of Appalachian forests and improve understanding of Neogene faunal evolution in the southern Appalachians.
ACKNOWLEDGEMENTS
Specimen collection at the Gray Fossil Site in Tennessee was partially funded through a National Science Foundation Grant (NSF Grant #0958985) to SC Wallace and BW Schubert. The remainder of the funding for the project was provided by internal funding from the Center of Excellence in Paleontology at East Tennessee State University and ETSU RDC Major Grant (#23-011M) to JX Samuels. The following collection managers kindly assisted with access to specimens in their care and provided valuable information: M. Inabinett and B. Compton (ETMNH), A. Woodruff and R. Narducci (UF), J. Galkin (AMNH), D. Lunde, and J. Ostosky (USNM), and A. Rountrey (UMMP). These specimens from GFS were collected through the important efforts of ETSU volunteers, students, and staff at GFS. We also thank L. Emmert and S. Haugrud for their outstanding preparation of specimens described here. Finally, we thank V. Bravo-Cuevas and an anonymous reviewer for their helpful feedback, which has greatly improved the quality of this manuscript.
REFERENCES
Bärmann, E.V. and Rössner, G.E. 2011. Dental nomenclature in Ruminantia: Towards a standard terminological framework. Mammalian Biology, 76:762-768.
https://doi.org/10.1016/j.mambio.2011.07.002
Bell, C.J., Lundelius, E.L., Jr., Graham, R.W., Barnosky, A.D., Lindsay, E.H., Ruez, D.R., Jr., Semken, H.A., Jr., Webb, S.D., Zakrzewski, R.J., and Woodburne, M.O. 2004. The Blancan, Irvingtonian, and Rancholabrean mammal ages. In Woodburne, M.O. (ed.), Late Cretaceous and Cenozoic mammals of North America; biostratigraphy and geochronology. New York: Columbia University Press, 232-314.
Boardman, G.S. and Schubert, B.W. 2011. First Mio-Pliocene salamander fossil assemblage from the southern Appalachians. Palaeontologia Electronica, 14(2):16A.
https://palaeo-electronica.org/2011_2/257/index.html
Bourque, J.R. and Schubert, B.W. 2015. Fossil musk turtles (Kinosternidae, Sternotherus) from the late Miocene-early Pliocene (Hemphillian) of Tennessee and Florida. Journal of Vertebrate Paleontology, 35(1):e885441.
https://doi.org/10.1080/02724634.2014.885441
Brookes, J. 1828. A catalogue of the anatomical & zoological museum of Joshua Brookes, Esq., F.R.S. F.L.S. &c. London.
Carrasco, M.A., Barnosky, A.D., Kraatz, B.P., and Davis, E.B. 2007. The Miocene mammal mapping project (MIOMAP): an online database of Arikareean through Hemphillian fossil mammals. Bulletin of the Carnegie Museum of Natural History 39:183-188.
https://doi.org/10.2992/0145-9058(2007)39[183:TMMMPM]2.0.CO;2
Côté, S.D., Rooney, T.P., Tremblay, J.P., Dussault, C., and Waller, D.M., 2004. Ecological impacts of deer overabundance. Annual Review of Ecology, Evolution, and Systematics, 35(1):113-147.
https://doi.org/10.1146/annurev.ecolsys.35.021103.105725
Croitor R. 2014. Deer from Late Miocene to Pleistocene of Western Palearctic: matching fossil record and molecular phylogeny data. Zitteliana B, 32:115-153.
Croitor, R. 2018. Plio-Pleistocene deer of western Palearctic: Taxonomy, Systematics, Phylogeny. Institute of Zoology of the Academy of Sciences of Moldova. 140 pp.
Croitor, R. 2022. Paleobiogeography of crown deer. Earth, 3(4):1138-1160.
https://doi.org/10.3390/earth3040066
Crowe, C. 2017. Sciurids (Rodentia: Sciuridae) of the Late Mio-Pliocene Gray Fossil Site and the Late Miocene Tyner Farm: implications on ecology and expansion of the sciurid record. Master’s thesis, East Tennessee State University.
DeSantis, L.R. and MacFadden, B. 2007. Identifying forested environments in deep time using fossil tapirs: evidence from evolutionary morphology and stable isotopes. Courier Forschungsinstitut Senckenberg, 258:147-157.
DeSantis, L.R. and Wallace, S.C. 2008. Neogene forests from the Appalachians of Tennessee, USA: geochemical evidence from fossil mammal teeth. Palaeogeography, Palaeoclimatology, Palaeoecology, 266(1):59-68.
https://doi.org/10.1016/j.palaeo.2008.03.032
DeSantis, L.R., Sharp, A.C., Schubert, B.W., Colbert, M.W., Wallace, S.C., and Grine, F.E. 2020. Clarifying relationships between cranial form and function in tapirs, with implications for the dietary ecology of early hominins. Scientific Reports, 10(1):8809.
https://doi.org/10.1038/s41598-020-65586-w
Doughty, E.M., Wallace, S.C., Schubert, B.W., and Lyon, L.M. 2018. First occurrence of the enigmatic peccaries Mylohyus elmorei and Prosthennops serus from the Appalachians: latest Hemphillian to Early Blancan of Gray Fossil Site, Tennessee. PeerJ, 6, p.e5926.
https://doi.org/10.7717/peerj.5926
Emery-Wetherell, M.M. and Schilter, J.F. 2020. A new early occurrence of Cervidae in North America from the Miocene-Pliocene Ellensburg Formation in Washington, USA. Palaeontologia Electronica, 23(1):a08.
https://doi.org/10.26879/946
Farlow, J.O., Sunderman, J.A., Havens, J.J., Swinehart, A.L., Holman, J.A., Richards, R.L., Miller, N.G., Martin, R., Hunt, R.M., Jr., Storrs, G.W., Curry, B.B., Fluegeman, R.H., Dawson, M.R., and Flint, M.E.T. 2001. The Pipe Creek Sinkhole biota, a diverse late Tertiary continental fossil assemblage from Grant County, Indiana. American Midland Naturalist, 145:367-378.
https://doi.org/10.1674/0003-0031(2001)145[0367:TPCSBA]2.0.CO;2
Farlow, J.O., Richards, R.L., Garniewicz, R.C., Greenan, M., Wepler, W.R., Shunk, A.J., Ludvigson, G.A., Shaffer, N.R., and Wenning, A.L. 2010. Occurrence and features of fossiliferous sediments of the Pipe Creek Sinkhole (late Neogene, Grant County, Indiana). Geology of the Late Neogene Pipe Creek Sinkhole (Grant County, Indiana), 812:1-60.
Geist, V. 1998. Deer of the world: Their evolution, behaviour, and ecology. Mechanicsburg: Stackpole Books.
Goldfuss, GA. 1820. Handbuch der Zoologie, vol. 2. Nürnberg.
Graham, R.W. and Lundelius, E.L., Jr. 2010. FAUNMAP II: New Data for North America with a Temporal Extension for the Blancan, Irvingtonian and early Rancholabrean. FAUNMAP II Database, version 1.0.
http://www.ucmp.berkeley.edu/faunmap (Accessed via Neotoma Paleoecology Database 20 August 2024).
Grau-Camats, M., Casanovas-Vilar, I., Crowe, C.J., and Samuels, J.X. 2025. Gliding between continents: a review of the North American record of the giant flying squirrel Miopetaurista (Rodentia, Sciuridae) with the description of new material from the Gray Fossil Site (Tennessee). Journal of Mammalian Evolution, 32, 8.
https://doi.org/10.1007/s10914-025-09751-w
Greenspoon, L., Krieger, E., Sender, R., Rosenberg, Y., Bar-On, Y.M., Moran, U., Antman, T., Meiri, S., Roll, U., Noor, E., and Milo, R., 2023. The global biomass of wild mammals. Proceedings of the National Academy of Sciences, 120(10), p.e2204892120.
https://doi.org/10.1073/pnas.2204892120
Gustafson, E.P. 2012. New records of rhinoceroses from the Ringold Formation of central Washington and the Hemphillian-Blancan boundary. Journal of Vertebrate Paleontology, 32(3), pp. 727-731. https://doi.org/10.1080/02724634.2012.658481
Gustafson, E.P. 2015. An Early Pliocene North American Deer: Bretzia pseudalces, Its Osteology, Biology, and Place in Cervid History. Bulletin of the Museum of Natural History, University of Oregon, (25).
https://hdl.handle.net/1794/29271
Hanley, T.A., 1996. Potential role of deer (Cervidae) as ecological indicators of forest management. Forest Ecology and Management, 88(1-2), pp.199-204.
https://doi.org/10.1016/S0378-1127(96)03803-0
Hibbard, C.W. 1941. Mammals of the Rexroad fauna from the upper Pliocene of southwestern Kansas. Transactions of the Kansas Academy of Science, 44:265-313.
Hulbert, R.C., Wallace, S.C., Klippel, W.E., and Parmalee, P.W. 2009. Cranial morphology and systematics of an extraordinary sample of the late Neogene dwarf tapir, Tapirus polkensis (Olsen). Journal of Paleontology, 83(2):238-262.
https://doi.org/10.1666/08-062.1
Jacobson, J.A. 2003. Identification of mule deer (Odocoileus hemionus) and white-tailed deer (Odocoileus virginianus) postcranial remains as a means of determining human subsistence strategies. Plains Anthropologist, 48(187):287-297.
Jasinski, S.E. 2018. A new slider turtle (Testudines: Emydidae: Deirochelyinae: Trachemys) from the late Hemphillian (late Miocene/early Pliocene) of eastern Tennessee and the evolution of the deirochelyines. PeerJ, 6:e4338.
https://doi.org/10.7717/peerj.4338
Jiménez-Hidalgo, E. and Bravo-Cuevas, V.M. 2015. A roe deer from the Pliocene of Hidalgo, central Mexico. Acta Paleontologia Polonica, 60(4):807-813.
https://doi.org/10.4202/app.00044.2013
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Vol. 1: Regnum animale. Editio decima, 1758. Stockholm: Societatis Zoologicae Germanicae.
Lundelius, E.L., Jr., Churcher, C.S., Downs, T., Harington, C.R., Lindsay, E.H., Schultz, G.E., Semken, H.A., Webb, S.D., and Zakrzewski, R.J. 1987. The North American Quaternary sequence. In Cenozoic mammals of North America: Geochronology and Biostratigraphy, ed. Woodburne MO. Berkeley: University of California Press, pp. 211-235.
Maclaren, J.A., Hulbert, R.C., Jr., Wallace, S.C., and Nauwelaerts, S. 2018. A morphometric analysis of the forelimb in the genus Tapirus (Perissodactyla: Tapiridae) reveals influences of habitat, phylogeny and size through time and across geographical space. Zoological Journal of the Linnean Society, 184(2):499-515.
https://doi.org/10.1093/zoolinnean/zly019
Maden, S. 2023. An Early Pliocene Fish Assemblage from the Southern Appalachians: Ichthyofauna of the Gray Fossil Site. Master’s thesis, East Tennessee State University.
Martin, R.A. 2021. Correlation of Pliocene and Pleistocene fossil assemblages from the central and eastern United States: toward a continental rodent biochronology. Historical Biology, 33(6):880-896.
https://doi.org/10.1080/08912963.2019.1666118
Martin, R.A. and Kelly, T.S. 2023. Biostratigraphy and biochronology of late Cenozoic North American rodent assemblages. Palaeontologia Electronica, 26(2):1-29.
https://doi.org/10.26879/1303
McConnell, S.M. and Zavada, M.S. 2013. The occurrence of an abdominal fauna in an articulated tapir (Tapirus polkensis) from the Late Miocene Gray Fossil Site, northeastern Tennessee. Integrative Zoology, 8(1):74-83.
https://doi.org/10.1111/j.1749-4877.2012.00320.x
Mead, J.I., Schubert, B.W., Wallace, S.C., and Swift, S.L. 2012. Helodermatid lizard from the Mio-Pliocene oak-hickory forest of Tennessee, eastern USA, and a review of monstersaurian osteoderms. Acta Palaeontologica Polonica, 57(1), pp. 111-121.
https://doi.org/10.4202/app.2010.0083
Morejohn, G.V. and Dailey, D.C. 2004. The identity and postcranial osteology of Odocoileus lucasi (Hay) 1927: a Plio-Pleistocene deer from California and Idaho. Sierra College Natural History Museum Bulletin, 1:1-54.
Oberg, D.E. and Samuels, J.X. 2022. Fossil moles from the Gray Fossil Site (Tennessee): Implications for diversification and evolution of North American Talpidae. Palaeontologia Electronica, 25.3.a33.
https://doi.org/10.26879/1150
Ochoa, D., Whitelaw, M., Liu, Y.S., and Zavada, M. 2012. Palynology from Neogene sediments at the Gray Fossil Site, Tennessee, USA: Floristic implications. Review of Palaeobotany and Palynology, 184:36-48.
https://doi.org/10.1016/j.revpalbo.2012.03.006
Ochoa, D., Zavada, M.S., Liu, Y., and Farlow, J.O. 2016. Floristic implications of two contemporaneous inland upper Neogene sites in the eastern US: Pipe Creek Sinkhole, Indiana, and the Gray Fossil Site, Tennessee (USA). Palaeobiodiversity and Palaeoenvironments, 96(2):239-254.
https://doi.org/10.1007/s12549-016-0233-4
Owen, R. 1848. Description of teeth and portions of jaws of two extinct Anthracotherioid quadrupeds (Hyopotamus vectianus and Hyop. bovinus) discovered by the Marchioness of Hastings in the Eocene deposits on the NW coast of the Isle of Wight: with an attempt to develope Cuvier’s idea of the Classification of Pachyderms by the number of their toes. Quarterly Journal of the Geological Society of London, 4:103-141
Palma-Ramírez, A., Bravo-Cuevas, V.M., and Villanueva Amadoz, U. 2023. First record of Odocoileus virginianus (Artiodactyla: Cervidae) from the Blancan of Sonora, Northwestern Mexico. Historical Biology, 36(10):2209. https://doi.org/10.1080/08912963.2023.2248163
Parmalee, P.W., Klippel, W.E., Meylan, P.A., and Holman, J.A. 2002. A late Miocene-early Pliocene population of Trachemys (Testudines: Emydidae) from east Tennessee. Annals Carnegie Museum, 71:233-239.
https://doi.org/10.5962/p.329869
Robertson, J.S. 1976. Latest Pliocene mammals from Haile XV A, Alachua County, Florida. Bulletin of the Florida State Museum Biological Sciences, 20(3):111-186.
https://doi.org/10.58782/flmnh.fzbw5044
Rooney, T.P. 2001. Deer impacts on forest ecosystems: a North American perspective. Forestry, 74(3):201-208.
https://doi.org/10.1093/forestry/74.3.201
Samuels, J.X., Bredehoeft, K.E., and Wallace, S.C. 2018. A new species of Gulo from the Early Pliocene Gray Fossil Site (Eastern United States); rethinking the evolution of wolverines. PeerJ, 6:e4648.
https://doi.org/10.7717/peerj.4648
Samuels, J.X. and Schap, J. 2021. Early Pliocene Leporids from the Gray Fossil Site of Tennessee. Eastern Paleontologist, 8:1-23.
https://eaglehill.us/epal-pdfs-regular/EPAL-008-Samuels.pdf
Schap, J.A. and Samuels, J.X. 2020. Mesowear analysis of the Tapirus polkensis population from the Gray Fossil Site, Tennessee, USA. Palaeontologia Electronica, 23(2):a26.
https://doi.org/10.26879/875
Schap, J.A., Samuels, J.X., and Joyner, T.A., 2021. Ecometric estimation of present and past climate of North America using crown heights of rodents and lagomorphs. Palaeogeography, Palaeoclimatology, Palaeoecology, 562, p.110144.
https://doi.org/10.1016/j.palaeo.2020.110144
Schubert, B.W. and Mead, J.I. 2011. Gray Fossil Site: 10 Years of Research. Johnson City, TN: Don Sundquist Center of Excellence in Paleontology, East Tennessee State University.
Short, R., Wallace, S., and Emmert, L. 2019. A new species of Teleoceras (Mammalia, Rhinocerotidae) from the late Hemphillian of Tennessee. Bulletin of the Florida Museum of Natural History, 56:183-260.
https://doi.org/10.58782/flmnh.kpcf8483
Shunk, A.J., Driese, S.G., and Clark, G.M. 2006. Latest Miocene to earliest Pliocene sedimentation and climate record derived from paleosinkhole fill deposits, Gray Fossil Site, northeastern Tennessee, U.S.A. Palaeogeography, Palaeoclimatology, Palaeoecology, 231:265-278.
https://doi.org/10.1016/j.palaeo.2005.08.001
Shunk, A.J., Driese, S.G., and Dunbar, J.A. 2009. Late Tertiary paleoclimatic interpretation from lacustrine rhythmites in the Gray Fossil Site, northeastern Tennessee, USA. Journal of Paleolimnology, 42:11-24.
https://doi.org/10.1007/s10933-008-9244-0
The NOW Community. 2024. New and Old Worlds Database of Fossil Mammals (NOW). Licensed under CC BY 4.0. Retrieved 20 August 2024 from
https://nowdatabase.org/now/database/
Vercauteren, K.C., Anderson, C.W., Van Deelen, T.R., Drake, D., Walter, W.D., Vantassel, S.M., and Hygnstrom, S.E. 2011. Regulated commercial harvest to manage overabundant white‐tailed deer: An idea to consider? Wildlife Society Bulletin, 35(3):185-194.
https://doi.org/10.1002/wsb.36
Vislobokova, I.A. 2012. Giant deer: origin, evolution, role in the biosphere. Paleontological Journal, 46:643-775.
https://doi.org/10.1134/S0031030112070027
Vislobokova, I., Dmitrieva, E., and Kalmykov, N. 1995. Artiodactyls from the Late Pliocene of Udunga, Western Trans-Baikal, Russia. Journal of Vertebrate Paleontology, 15:146-159.
https://doi.org/10.1080/02724634.1995.10011214
von Den Driesch, A. 1976. A guide to the measurement of animal bones from archaeological sites. Peabody Museum of Archaeology and Ethology, 1:1-137.
Wallace, S.C. and Wang, X. 2004. Two new carnivores from an unusual late Tertiary forest biota in eastern North America. Nature, 431:556-559.
https://doi.org/10.1038/nature02819
Webb, S.D. 1998. Cervidae and Bovidae. In Janis, C.M., Scott, K.M., and Jacobs, L.L. (eds.), Evolution of Tertiary mammals of North America, vol. 1: Terrestrial carnivores, ungulates, and ungulatelike mammals. Cambridge: Cambridge University Press, 691 pp.
Webb, S. 2000. Evolutionary history of new world Cervidae. In Vrba, E., Schaller, G. (eds.), Antelopes, Deer, and Relatives: Fossil Record, Behavioral Ecology, Systematics, and Conservation. New Haven: Yale University Press. 38-64.
Webb, S.D., Hulbert, R.C., Jr., Morgan, G.S., and Evans, H.F. 2008. Terrestrial mammals of the Palmetto Fauna (early Pliocene, latest Hemphillian) from the central Florida phosphate district. Natural History Museum Los Angeles County Science Series, 41:293-312.
Wheatley, P.V. and Ruez, Jr., D.R. 2006. Pliocene Odocoileus from Hagerman Fossil Beds National Monument, Idaho, and comments on the taxonomic status of Odocoileus brachyodontus. Journal of Vertebrate Paleontology, 26(2), pp.462-465.
https://doi.org/10.1671/0272-4634(2006)26[462:POFHFB]2.0.CO;2
Wood, H.E., II, Chaney, R.W., Clark, J., Colbert, E.H., Jepsen, G.L., Reeside, J.B., Jr., and Stock, C. 1941. Nomenclature and correlation of the North American continental Tertiary. Bulletin of the Geological Society of America, 52:1-48, Plate 1.
Worobiec, E., Liu, Y., and Zavada, M.S. 2013. Palaeoenvironment of late Neogene lacustrine sediments at the Gray Fossil Site, Tennessee, U.S.A. Annales Societatis Geologorum Poloniae, 83:51-63.
Xu, Z. 2023. Early Pliocene Mice and Rats from the Gray Fossil Site of Eastern Tennessee: Implications for the Evolution of Cricetidae and Understanding of the Past Ecosystem Master’s thesis, East Tennessee State University.
Zobaa, M.K., Zavada, M.S., Whitelaw, M., Shunk, A.J., and Oboh-Ikuenobe, F.E. 2011. Palynology and palynofacies analyses of the Gray Fossil Site, eastern Tennessee: their role in understanding the basin-fill history. Palaeogeography, Palaeoclimatology, Palaeoecology, 308(3-4):433-444.
https://doi.org/10.1016/j.palaeo.2011.05.051
