

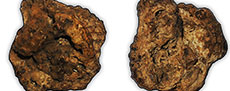
 First fossil snake from McFaddin Beach, Texas, USA
First fossil snake from McFaddin Beach, Texas, USA
Article number: 27.2.a37
https://doi.org/10.26879/1304
Copyright Paleontological Society, July 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 8 May 2023. Acceptance: 11 July 2024.
ABSTRACT
McFaddin Beach is an archeological and paleontological locality extending approximately 32 km along the coast of Jefferson County, Texas, USA. Vertebrate fossils recovered from McFaddin Beach include Mammuthus, Equus, Mammut, and Holmesina, which indicates that at least some of the faunal material washed onto the beach is Pleistocene in age. A fossil snake was found on McFaddin Beach. It is a sample of partly cemented matrix containing more than 29 associated snake vertebrae, many in articulated sections. The fossil was CT-scanned and individual vertebrae were digitally segmented. Through this digital visualization, the vertebrae were identified to be from the precloacal trunk region. The snake was then qualitatively described and compared to the genera Lampropeltis (Kingsnakes), Pantherophis (Ratsnakes, Cornsnakes, and Foxsnakes), Rhinocheilus (Long-nosed snakes), and Cemophora (Scarlet snakes), which are all limited geographically to species currently present in southeastern North America. The fossil most resembled the genus Lampropeltis. The nearest fossils attributed to this genus are from Bee County, Texas, approximately 92 km from the modern Texas coast. This record expands prehistoric ranges of Lampropeltis to the continental shelf in times of low sea level.
Deanna Flores. Department of Earth Sciences, 100 Cascade Hall, 1272 University of Oregon Eugene, OR 97403, USA. dflores9@uoregon.edu
Antonio Meza. School of Life Sciences, Arizona State University, 427 East Tyler Mall, Tempe, AZ 85287, USA. ameza36@asu.edu
Christopher J. Bell. Jackson School of Geosciences, The University of Texas at Austin, Austin, Texas 78712, USA. cjbell@jsg.utexas.edu
Stacie Skwarcan. Jackson School of Geosciences, The University of Texas at Austin, Austin, Texas 78712, USA. sskwarca@utexas.edu
William Godwin. Sam Houston State Natural History Collections, Sam Houston State University, Huntsville, Texas 77341, USA. wbg004@shsu.edu
Jesse Fremont. Sam Houston State Natural History Collections, Sam Houston State University, Huntsville, Texas 77341, USA. jessefremont@msn.com
Patrick J. Lewis. Sam Houston State Natural History Collections, Sam Houston State University, Huntsville, Texas 77341, USA and Department of Biological Sciences, Sam Houston State University, Huntsville, Texas 77341, USA. pjl001@shsu.edu
Keywords: snake; McFaddin Beach; Texas; Pleistocene; Gulf of Mexico
Final citation: Flores, Deanna, Meza, Antonio, Bell, Christopher J., Skwarcan, Stacie, Godwin, William, Fremont, Jesse, and Lewis, Patrick J. 2024. First fossil snake from McFaddin Beach, Texas, USA. Palaeontologia Electronica, 27(2):a37.
https://doi.org/10.26879/1304
palaeo-electronica.org/content/2024/5271-mcfaddin-beach-fossil-snake
Copyright: July 2024 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
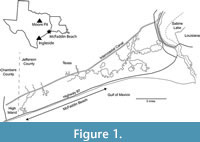 During the examination of a private collection of McFaddin Beach (MB) material, a specimen was found consisting of partly cemented matrix that contains more than 29 associated snake vertebrae. McFaddin Beach extends approximately 32 km along the Gulf Coast of Jefferson County, Texas, USA (Figure 1). This beach is a secondary deposit for Paleoindian artifacts and fossils, most likely Pleistocene in age (Russell, 1975; Long, 1977; Turner and Tanner, 1994). The fauna described by this study and by Russell (1975) represent taxa that are characteristic of the Texas Coastal Plains as well as the greater Gulf Coast area. The identified megafauna can be divided into three groups, “extinct”, “extant”, and “local”, as described by Russell (1975). The extinct group includes taxa such as Mammut, Equus, and Smilodon, that were extinct by the end of the Pleistocene. The extant group includes extant taxa that no longer inhabit the Coastal Plains of Texas such as Bison and Cynomys. The local group includes taxa that are still found in the area at the present time such as Didelphis, Procyon lotor, Odocoileus virginianus, and Canis latrans. The source deposit (or deposits) for the artifacts and fossils is hypothesized to be somewhere on the continental shelf, as far as ~80 km out from the current shoreline near the ancient river valleys of what are today the Trinity, Sabine, Calcasieu, and Neches rivers (Russell, 1975; Long, 1977; Turner and Tanner, 1994). It is unknown if the fossils represent local populations that inhabited the area of the source location or if they represent vagrant groups that historically may have moved through the area.
During the examination of a private collection of McFaddin Beach (MB) material, a specimen was found consisting of partly cemented matrix that contains more than 29 associated snake vertebrae. McFaddin Beach extends approximately 32 km along the Gulf Coast of Jefferson County, Texas, USA (Figure 1). This beach is a secondary deposit for Paleoindian artifacts and fossils, most likely Pleistocene in age (Russell, 1975; Long, 1977; Turner and Tanner, 1994). The fauna described by this study and by Russell (1975) represent taxa that are characteristic of the Texas Coastal Plains as well as the greater Gulf Coast area. The identified megafauna can be divided into three groups, “extinct”, “extant”, and “local”, as described by Russell (1975). The extinct group includes taxa such as Mammut, Equus, and Smilodon, that were extinct by the end of the Pleistocene. The extant group includes extant taxa that no longer inhabit the Coastal Plains of Texas such as Bison and Cynomys. The local group includes taxa that are still found in the area at the present time such as Didelphis, Procyon lotor, Odocoileus virginianus, and Canis latrans. The source deposit (or deposits) for the artifacts and fossils is hypothesized to be somewhere on the continental shelf, as far as ~80 km out from the current shoreline near the ancient river valleys of what are today the Trinity, Sabine, Calcasieu, and Neches rivers (Russell, 1975; Long, 1977; Turner and Tanner, 1994). It is unknown if the fossils represent local populations that inhabited the area of the source location or if they represent vagrant groups that historically may have moved through the area.
A persistent challenge in the fossil record of snakes is that the majority of fossils attributable to snakes are isolated vertebrae, or short series of articulated (or associated) vertebral elements (e.g., Auffenberg, 1963; Rodriguez-Robles and De Jesus-Escobar, 1999; Holman, 2000; Caldwell et al., 2015; Caldwell, 2020; Head et al., 2022). A problem in the interpretation of those fossils is the degree to which isolated vertebrae, or short series of vertebrae, can reliably be diagnosed to finer levels in a taxonomic hierarchy (Head et al., 2022; Smith and Georgalis, 2022). Many herpetofaunal species lineages have poor documentation of evolutionary morphology due to many Quaternary paleontologists using a traditional comparative approach instead of an apomorphy based approach to identifications (Holman, 1999; Bell et al., 2010). Recent work has shown that the apomorphy based approach to identification results in higher level taxonomic resolution instead of species level resolution (Bell et al., 2004). Much of the traditional approach does have a tendency to ignore some level of variation (sexual dimorphism, ontogeny, etc.) which can result in some members being mistaken for a different species (Bell et al., 2010). Both approaches also suffer from previously published characteristics that were restricted to a specific geographic context instead of a wider phylogenetic context (Bell et al., 2010). Utilization of geographic data can help refine taxonomic resolution, such as North American natricine snakes are able to be distinguished from North American elapids but not all natricine snakes can be separated from all elapid snakes (Bell et al., 2004). Here, we qualitatively describe the MB fossil through CT scanning and digital visualization methods. We use some apomorphies (e.g., presence or absence of hypapophyses or of distinct subcentral ridges) along with geographic data to narrow down higher-level taxonomy of the specimen. To further narrow the specimen down to a genus level identification, we utilize the traditional approach to compare the specimen to known genera for phenetic similarity.
MATERIAL AND METHODS
The MB specimen (SHSU-1-311) was donated to the Sam Houston State Natural History Collections by Jesse Fremont, curator of the Orangefield Cormier Museum. The specimen currently is on loan to be displayed at the Orangefield Cormier Museum in Orangefield, Texas.
Some vertebral sections are visible on the surface of the MB specimen SHSU-1-311, but much of the material is obscured by matrix. To view the specimen in greater detail, it was CT-scanned at the University of Texas at Austin High-Resolution X-ray Computed Tomography Facility (UT-CT) using the NSI scanner. The machine at UT-CT had Hamamatsu X-ray focus high power source operating at 160 kV and 0.2mA with an aluminum filter and a Perkin Elmer detector. The CT scan was continuous with two frames averaged, zero skipped frames, 3000 projections, four gain calibrations, 5 mm calibration phantom, data range of -20 to 360 adjusted grayscale values, and beam-hardening correction of 0.25. Post-reconstruction ring correction applied by Dr. Jessie Maisano used parameters oversample of two, radial bin width of 21, sectors of 32, minimum arc length of two, angular bin width of nine, and an angular screening factor of four. Voxel size is 32.9 μm, based on distance of source to object 240.838 mm and distance of source to detector 1465.675 mm. The total number of slices is 1958. The CT scans and a surface scan are catalogued and digitally stored with the natural history collections at Sam Houston State University. The scan was imported in Avizo (version 9.7), and three articulated sections of vertebral column were recognized, including a total of 29 individual vertebrae that could be digitally isolated. Additional vertebrae present in the specimen could not be isolated due to their similarity in density with the surrounding matrix.
To identify the specimen down to the genus level, the vertebral morphology was compared to that of specimens from the genera Lampropeltis (51 specimens consisting of L. getula, L. triangulum, L. calligaster, and L. mexicana), Pantherophis (26 specimens consisting of P. obsoletus, P. vulpinus, and P. guttatus), Rhinocheilus (27 specimens consisting of R. lecontei), and Cemophora (10 specimens consisting of C. coccinea) at Sam Houston State University and the Vertebrate Paleontology Lab at The University of Texas at Austin. Comparative specimens were limited geographically to southeastern North America. Specimen availability also limited the geographic range of comparative specimens. Descriptions and images from the literature were also used to assist in identification on the basis of vertebral morphology, commonly used with other extinct snake specimens (Auffenberg, 1963; Holman, 1995, 2000; Krysko, 2001; McCartney et al., 2014; Szyndlar and Georgios, 2023). Other taxa (e.g., natricine snakes or tribe Sonorini) were excluded based on size, general dimensions of the vertebrae (e.g., short stubby neural spine, concave neural arches), and presence of a hypophysis.
Abbreviations
SHSU-Sam Houston State University, Natural History Collections
MB-McFaddin Beach, Texas
TxVP-Vertebrate Paleontology Lab, University of Texas at Austin
SYSTEMATIC PALEONTOLOGY
Class Reptilia Laurenti, 1768
Order Squamata Oppel, 1811
Family Colubridae Oppel, 1811
Genus LAMPROPELTIS Fitzinger, 1843
Lampropeltis sp. Fitzinger, 1843
(Figure 2, Figure 3)
SHSU-1-311. A conglomeration of matrix with articulated segments of precloacal trunk vertebrae and ribs imbedded.
Description and Diagnosis
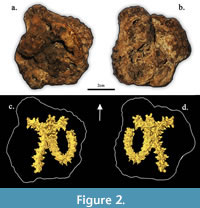 The specimen identification was narrowed down to belonging to the family Colubridae due to the lack of hypapophyses on the trunk vertebrae, distinct subcentral ridges/grooves, and prominent well-projected prezygapophyseal accessory processes (Holman, 2000). From the family, possible identifications were narrowed down based on size and similarity. Because the vertebrae have distinct synapophyses (rib attachment points) and various unsegmented ribs are preserved within the material encasing the specimen, the vertebrae are interpreted to be from the precloacal
The specimen identification was narrowed down to belonging to the family Colubridae due to the lack of hypapophyses on the trunk vertebrae, distinct subcentral ridges/grooves, and prominent well-projected prezygapophyseal accessory processes (Holman, 2000). From the family, possible identifications were narrowed down based on size and similarity. Because the vertebrae have distinct synapophyses (rib attachment points) and various unsegmented ribs are preserved within the material encasing the specimen, the vertebrae are interpreted to be from the precloacal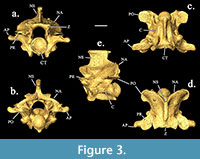 trunk region of the vertebral column. The vertebrae have an average centrum length of 5.62 mm and an average neural arch width of 5.01 mm. The ratios of neural spine length to neural spine height, and cotyle width to cotyle height of SHSU-1-311 are 1.39 and 0.99, respectively. Neural spine heights are greater than lengths, and they also have a posterior overhang. The neural arches are moderately vaulted with convex laminae. The prezygapophyseal accessory processes exhibit lengths less than the greatest lengths of the prezygapophyseal processes and are thick with rounded/blunted ends.
trunk region of the vertebral column. The vertebrae have an average centrum length of 5.62 mm and an average neural arch width of 5.01 mm. The ratios of neural spine length to neural spine height, and cotyle width to cotyle height of SHSU-1-311 are 1.39 and 0.99, respectively. Neural spine heights are greater than lengths, and they also have a posterior overhang. The neural arches are moderately vaulted with convex laminae. The prezygapophyseal accessory processes exhibit lengths less than the greatest lengths of the prezygapophyseal processes and are thick with rounded/blunted ends.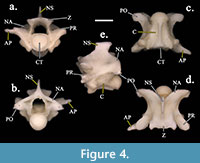 The zygapophyses are pronounced laterally, and the zygosphene lacks an anteriorly flattened roof. The subcentral ridges are strongly developed. These characters are all consistent with referral to the specimen of Lampropeltis (Figure 3, Figure 4).
The zygapophyses are pronounced laterally, and the zygosphene lacks an anteriorly flattened roof. The subcentral ridges are strongly developed. These characters are all consistent with referral to the specimen of Lampropeltis (Figure 3, Figure 4).
 Comparisons
Comparisons
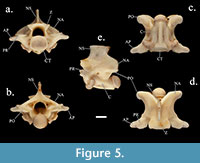
Members of Pantherophis have characteristics similar to Lampropeltis, but Pantherophis has less depressed neural arches, higher and thinner neural spines, and longer, less rounded accessory processes than seen in SHSU-1-311 (Auffenberg, 1963; Parmley, 1986a; Holman, 1995, 2000). Specimens of Pantherophis (Figure 5) have a similar general shape to SHSU-1-311, especially with the neural spine and zygapophyses. The vertebrae differ in the lack of lateral distinctions of the zygosphene, in wider neural arches, and in how the accessory processes do not have as much of an upward curve seen in cranial and caudal views (Figure 5).
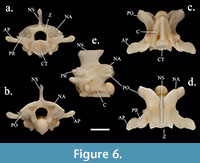 The neural spine length is greater than the height in Rhinocheilus compared to that of SHSU-1-311 (Figure 3, Figure 6). Both specimens have a posterior overhang. The neural spine has a less rounded ventral portion anteriorly and a more rounded ventral portion posteriorly in comparison to SHSU-1-311. The neural arches appear to be more rounded and wider in the Rhinocheilus specimen (Figure 6). The prezygapophysis and postzygapophysis are similar in shape to SHSU-1-311.
The neural spine length is greater than the height in Rhinocheilus compared to that of SHSU-1-311 (Figure 3, Figure 6). Both specimens have a posterior overhang. The neural spine has a less rounded ventral portion anteriorly and a more rounded ventral portion posteriorly in comparison to SHSU-1-311. The neural arches appear to be more rounded and wider in the Rhinocheilus specimen (Figure 6). The prezygapophysis and postzygapophysis are similar in shape to SHSU-1-311.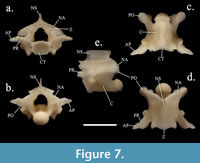 The accessory process is rounded but extends more laterally. The zygosphene is similar in shape with a more rounded medial portion.
The accessory process is rounded but extends more laterally. The zygosphene is similar in shape with a more rounded medial portion.
Specimens of Cemophora have a greater neural spine length than height, with a slight posterior overhang (Figure 7). The neural arches are not as depressed as those of SHSU-1-311. The prezygapophysis and postzygapophysis are both similar in shape to SHSU-1-311. The accessory process is not rounded (Figure 7). The zygosphene is similar in the lateral distinctions but is not as flat medially as seen in SHSU-1-311 (Figure 3, Figure 7). This specimen has a projection on the portion below the neural spine in lateral view, not seen in SHSU-1-311.
TAPHONOMY
A detailed taphonomic study of the specimen has not been completed. This fossilized aggregation of snake vertebrae is well preserved and is currently the only articulated specimen from MB. The current hypothesis is that cement deposited around the bones during fossilization yielded tightly articulated vertebrae and created a protective layer against wave action for the specimen as a whole (Figure 2). An articulated, multi-element skeleton suggests that the specimen may have been rapidly buried, was in a burrow, was fossorial, was trapped in a rapid influx of sediment, or a combination of these possibilities (Lopes et al., 2008). Small crystalline structures are found scattered around the surface of the fossil.
DISCUSSION
The specimen, SHSU-1-311, is the only snake fossil known from MB and is one of the smallest-bodied, most well-preserved fossils collected from the locality. The fossil was difficult to identify using externally visible morphology as much of it was obscured (Figure 2A-B). The use of CT was vital to adequately identify the specimen. The use of X-ray computed tomography and digital segmentation software allows for the study of and visualization of microfossils in three dimensions for specimens that are not easily extracted using traditional preparation methods. The use of CT scanning is mainstream due to its practicality (Bell and Mead, 2014; Gignac et al., 2016; Georgalis and Scheyer, 2022). For the study of microvertebrates, the use of CT has allowed many researchers to ask and explore new and exciting questions. The implementation of CT provides an avenue to access new fossil snakes and gather quantitative data, in the form of geometric morphometrics, to help diagnose and identify specimens.
Fossils assigned to Lampropeltis first appeared during the Miocene (Rodriguez-Robles and De Jesus-Escobar, 1999; Armstrong et al., 2001; Krysko, 2001; Bryson, et al., 2007; Pyron and Burbrink, 2009; Krysko et al., 2017) and are reported from sites throughout the United States and Mexico (Holman, 2000). In Texas, fossils assigned to Lampropeltis were found in locations dated throughout the Pleistocene (Holman, 1964; Parmley, 1986b; Holman and Winkler, 1987; Parmley, 1988a, 1988b, 1990; Holman, 2000) and were previously assigned to the species Lampropeltis triangulum, Lampropeltis calligaster, Lampropeltis mexicana, and Lampropeltis getula. Several species, including Lampropeltis alterna, L. annulata, L. triangulum, L. calligaster, and L. getula, are also found in the modern fauna of Texas, but only three species (Lampropeltis triangulum, Lampropeltis calligaster, and Lampropeltis getula) have modern ranges which include the coast (Werler and Dixon, 2010; Dixon et al., 2020). Fossil material of Lampropeltis has not been reported from any coastal localities in Texas. At least 13 localities in Texas were previously reported to include fossils of Lampropeltis; the closest locality to McFaddin Beach is Berclair Terrace Site 1 in Bee County, Texas, which is approximately 96 km from the coast (Conkin and Conkin, 1962).
The current range of Lampropeltis does include the Texas Coast, but no fossil specimens of Lampropeltis are known from the modern coast. Although the original site of deposition of the fossil is unknown, it may have been close to the shoreline at that time; during the Pleistocene (the hypothesized age of the specimen based on associated fauna) the coast was as much as 129 km south of its modern position (Anderson, 2007). This fossil material supports the presence of Lampropeltis on the coast in the Pleistocene regardless of how far the unknown source location is from the modern shoreline.
Although few fossils from McFaddin Beach have been formally published, this site and the fossils found there provide valuable insight into the Gulf Coast region during the Pleistocene. Faunal similarities with deposits in Florida suggest a potential faunal region specific to the Gulf Coast. Fossil deposits are uncommon on continental shelves, and only a few are known from North America (e.g., Whitmore et al., 1967; Hoyle et al., 2004), the North Sea (Erdbrink and Van Bree, 1995; Van Kolfschoten and Laban, 1995; Van Essen and Mol, 1996; Mol et al., 2006), Argentina (Tonni and Cione, 1999; Cione et al., 2005), Uruguay (Rinderknecht, 2006), and Brazil (Lopes et al., 2008; Lopes and Buchmann, 2011). McFaddin Beach was for many decades a popular site for private collectors, and many of the known fossils and artifacts are not curated in public institutions. There are three main institutionalized collections from McFaddin Beach: the natural history collections at Sam Houston State University (Huntsville, Texas), the Texas Vertebrate Paleontology Laboratory (Austin, Texas), and the Museum of the Gulf Coast (Port Arthur, Texas). The collections manager and students at Sam Houston State University (SHSU) continue to work with local private collectors that collected, or still collect, materials from McFaddin Beach, and have entered into collaborations that result in transfer of private collections to the museum. As the McFaddin Beach faunal material is studied, it will provide an excellent opportunity to evaluate how taphonomic processes of shelf deposits differ from inland deposits that preserve vertebrate fossils.
CONCLUSIONS
The vertebrae are, to our knowledge, the first articulated fossils found from MB. Articulated specimens, especially of this size, are highly unusual at MB because the beach recovery site is a secondary deposit where elements are redeposited by waves from unknown primary offshore deposits. The discovery of the Lampropeltis specimen provides a rare glimpse into the make-up of the small vertebrate community of MB as much of the site’s fossil record is biased toward larger-bodied animals.
The Jesse Fremont collection includes other fossils that yield insight into the McFaddin Beach fauna. Collections staff at the natural history collections at SHSU have been successful in working with private collectors to archive four other formerly private collections into formal collections held in the public trust (e.g., Bell et al., 2020; Flores, 2022). Those collectors have provided valuable fossils and shared knowledge about the history of deposits, collections, and research on fossil material from McFaddin Beach that greatly enhances not only the understanding of the fossils themselves, but also their historical context and scientific relevance.
ACKNOWLEDGEMENTS
We thank W.I. Lutterschmidt and C. Sagebiel for access to modern specimens in their care. We thank J. Maisano and M. Colbert at The University of Texas High-Resolution X-ray CT Facility for their efforts on our behalf.
REFERENCES
Anderson, J.B. 2007. The Formation and Future of the Upper Texas coast: A Geologist Answers Questions about Sand, Storms, and Living by the Sea. Texas A&M University Press, College Station, Texas.
Armstrong, M.P., Frymire, D., and Zimmerer, E.J. 2001. Analysis of sympatric populations of Lampropeltis triangulum syspila and Lampropeltis triangulum elapsoides, in western Kentucky and adjacent Tennessee with relation to the taxonomic status of the scarlet kingsnake. Journal of Herpetology, 35:688–693.
https://doi.org/10.2307/1565915
Auffenberg, W. 1963. The fossil snakes of Florida. Tulane Studies in Zoology, 10:131–216.
https://doi.org/10.5962/bhl.part.4641
Bell, C.J., Head, J.J., and Mead, J.I. 2004. Synopsis of the herpetofauna from Porcupine Cave. In Barnosky, A.D. (ed.), Biodiversity Response to Climate Change in the Middle Pleistocene: the Porcupine Cave Fauna from Colorado. University of California Press, Berkeley, California, pp. 117–126.
Bell, C.J., Gauthier, J.A., and Bever, G.S. 2010. Covert biases, circularity, and apomorphies: a critical look at the North American Quaternary Herpetofaunal Stability Hypothesis. Quaternary International, 217: 30–36.
https://doi.org/10.1016/j.quaint.2009.08.009
Bell, C.J. and Mead, J. 2014. Not enough skeletons in the closet: collections-based anatomical research in an age of conservation conscience. The Anatomical Record, 297:344–348.
https://doi.org/10.1002/ar.22852
Bell, C.J., Jenkins, K., and Lewis, P. J. 2020. First fossil manatees in Texas: Trichechus manatus bakerorum in the Pleistocene fauna from beach deposits along the Texas Coast of the Gulf of Mexico. Palaeontologia Electronica, 23:a47.
https://doi.org/10.26879/1006
Bryson, R.W. Jr., Pastorini, J., Burbrink, F.T., and Forstner M.R. 2007. A phylogeny of the Lampropeltis mexicana complex (Serpentes: Colubridae) based on mitochondrial DNA sequences suggests evidence for species-level polyphyly within Lampropeltis. Molecular Phylogenetics and Evolution, 43:674–684.
https://doi.org/10.1016/j.ympev.2006.11.025
Caldwell, M.W. 2020. The Origin of Snakes: Morphology and the Fossil Record. CRC Press, Boca Raton, Florida.
https://doi.org/10.1201/9781315118819
Caldwell, M.W., Nydam, R.L., Palci, A., and Apesteguía, S. 2015. The oldest known snakes from the Middle Jurassic-Lower Cretaceous provide insights on snake evolution. Nature Communications, 6:1–11.
https://doi.org/10.1038/ncomms6996
Cione, A.L., Tonni, E.P., and Dondas, A.. 2005. A mastodont (Mammalia, Gomphotheriidae) from the Argentinian continental shelf. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 10:614–630.
https://doi.org/10.1127/njgpm/2005/2005/614
Conkin, J.E. and Conkin, B.M. 1962. Pleistocene Berclair terrace of Medio Creek, Bee County, Texas. AAPG Bulletin, 46:344–353.
https://doi.org/10.1306/bc743825-16be-11d7-8645000102c1865d
Dixon, J.R., Werler, J.E., and Forstner, M. 2020. Texas Snakes: a Field Guide. University of Texas Press, Austin, Texas.
https://doi.org/10.7560/706750
Erdbrink, D.B. and Van Bree, P. 1995. Hyeana brevirostris, a curious fragment from the bottom of the North Sea. Beaufortia, 45:105–114.
Fitzinger, L. 1843. Systema Reptilium. Fasciculus Primus. Ambyglossae. Braumüller et Seidel Bibliopolas, Vindobonae [= Wien]. 106 pp. + [vi] pp. + [3] pp.
https://doi.org/10.5962/bhl.title.4694
Flores, M.D. 2022. Identification of Pleistocene fauna from McFaddin Beach, Texas. Unpublished Master’s Thesis, Sam Houston State University, Huntsville, Texas.
Georgalis, G.L. and Scheyer, T.M. 2022. Crushed but not lost: a colubriform snake (Serpentes) from the Miocene Swiss Molasse, identified through the use of micro-CT scanning technology. Swiss Journal of Geosicences, 115:1–9.
https://doi.org/10.1186/s00015-022-00417-w
Gignac, P.M., Kley, N.J., Clarke, J.A., Colbert, M.W., Morhardt, A.C., Cerio, D., Cost, I.N., Cox, P.G., Daza, J.D., Early, C.M., Echols, M.S., Henkelman, R.M., Herdina, A.N., Holliday, C.M., Zhiheng Li, Mahlow, K., Merchant, S., Müller, J., Orsbon, C.P., Paluh, D.J., Thies, M.L., Tsai, H.P., and Witmer, L.M.. 2016. Diffusible iodine-based contrast-enhanced computed tomography (diceCT): an emerging tool for rapid, high-resolution, 3-D imaging of metazoan soft tissues. Journal of Anatomy, 228:889–909.
https://doi.org/10.1111/joa.12449
Head, J.J., Howard, A.F.C., and Müller, J. 2022. The Mesozoic fossil record of snakes and its implications for origin hypotheses, biogeography, and mass extinction, p. 26–54. In Gower, D.J. and Zaher, H. (ed.), The Origin and Early Evolutionary History of Snakes. Cambridge University Press, Cambridge, United Kingdom.
Holman, J.A. 1964. Pleistocene amphibians and reptiles from Texas. Herpetologica, 20:73–83.
Holman, J.A. 1995. Pleistocene Amphibians and Reptiles in North America. Oxford University Press, New York, New York.
Holman, J.A. 1999. Comments on Holarctic Pleistocene herpetofaunas. Bulletin of the Chicago Herpetological Society 34: 245–249.
Holman, J.A. 2000. Fossil Snakes of North America: Origin, Evolution, Distribution, Paleoecology. Indiana University Press, Bloomington, Indiana.
Holman, J.A. and Winkler, A.J. 1987. A mid-Pleistocene (Irvingtonian) herpetofauna from a cave in south central Texas. Pearce-Sellards Series, 44:1–17.
Hoyle, B.G., Fisher, D.C., Borns Jr., H.W., Churchill-Dickson, L.L., Dorion, C.C., and Weddle, T.K. 2004. Late Pleistocene mammoth remains from coastal Maine, USA. Quaternary Research, 61:277–288.
https://doi.org/10.1016/j.yqres.2004.02.006
Krysko, K.L. 2001. Ecology, conservation, and morphological and molecular systematics of the kingsnake, Lampropeltis getula (Serpentes: Colubridae). Unpublished PhD Dissertation, University of Florida, Gainsville, Florida.
Krysko, K.L., Nuñez, L.P., Newman, C.E., and Bowen, B.W. 2017. Phylogenetics of kingsnakes, Lampropeltis getula complex (Serpentes: Colubridae), in eastern North America. Journal of Heredity, 108:226–238.
https://doi.org/10.1093/jhered/esw086
Laurenti, J.N. 1768. Specimen Medicum, Exhibens Synopsin Reptilium Emendatam cum Experimentis circa Venena et Antidota Reptilium Austriacorum. Typis Joan. Thomæ Nob. de Trattnern, Cæs. Reg. Aulæ Typographi, et Bibliop., Viennæ [Vienna]. [2] + 214 + [1] pp., 5 plates.
https://doi.org/10.5962/bhl.title.5108
Long, R.J. 1977. McFaddin Beach. The Pattillo Higgins Series of Natural History and Anthropology, 1.
Lopes, R.P. and Buchmann, F.S. 2011. Pleistocene mammals from the southern Brazilian continental shelf. Journal of South American Earth Sciences, 31:17–27.
https://doi.org/10.1016/j.jsames.2010.11.003
Lopes, R.P., Buchmann, F.S., and Caron, F. 2008. Taphonomic analysis on fossils of Pleistocene mammals from deposits submerged along Southern Rio Grande do Sul coastal plain, Brazil. Arquivos do Museu Nacional, Rio de Janeiro, Brazil 66:213–229.
McCartney, J.A., Stevens, N.J., and O’Connor, P.M. 2014. The earliest Colubroid-dominated snake fauna from Africa: perspectives from the Late Oligocene Nsungwe Formation of southwestern Tanzania. PLoS One, 9: e90415.
https://doi.org/10.1371/journal.pone.0090415
Mol, D., Post, K., Reumer, J.W., van der Plicht, J., de Vos, J., van Geel, B., van Reenen, G., Pals, J.P., and Glimmerveen, J. 2006. The Eurogeul–first report of the palaeontological, palynological and archaeological investigations of this part of the North Sea. Quaternary International, 142:178–185.
https://doi.org/10.1016/j.quaint.2005.03.015
Oppel, M. 1811. Die Ordnungen, Familien und Gattungen der Reptilien als Prodrom einer Naturgeschichte derselben. Joseph Lindauer, München. xii + 86 pp.
https://doi.org/10.5962/bhl.title.4911
Parmley, D. 1986a. An annotated key to isolated trunk vertebrae of Pantherophis (Colubridae) species occurring in Texas. Texas Journal of Science, 38:41–44.
Parmley, D. 1986b. Herpetofauna of the Rancholabrean Schulze Cave local fauna of Texas. Journal of Herpetology, 20:1–10.
Parmley, D. 1988a. Middle Holocene herpetofauna of Klein Cave, Kerr County, Texas. The Southwestern Naturalist, 33:378–382.
https://doi.org/10.2307/3671775
Parmley, D. 1988b. Additional Pleistocene amphibians and reptiles from the Seymour Formation, Texas. Journal of Herpetology, 22:82–87.
https://doi.org/10.2307/1564359
Parmley, D. 1990. Late Pleistocene snakes from Fowlkes Cave, Culberson County, Texas. Journal of Herpetology, 24:266–274.
https://doi.org/10.2307/1564393
Pyron, R.A. and Burbrink, F.T. 2009. Neogene diversification and taxonomic stability in the snake tribe Lampropeltini (Serpentes: Colubridae). Molecular Phylogenetics and Evolution, 52:524–529.
https://doi.org/10.1016/j.ympev.2009.02.008
Rinderknecht, A. 2006. Vertebrados fósiles de la costa uruguaya, p. 343–349. In Menafra, R., Rodríguez-Gallego, L., Scarabino, F., and Conde, D. (eds.), Bases para la conservación y el manejo de la costa uruguaya. Vida Silvestre Uruguay, Montevideo.
Rodriguez-Robles, J.A. and De Jesus-Escobar, J.M. 1999. Molecular systematics of New World lampropeltinine snakes (Colubridae): implications for biogeography and evolution of food habits. Biological Journal of the Linnean Society, 68:355–385.
https://doi.org/10.1111/j.1095-8312.1999.tb01176.x
Russell, J.D. 1975. Identification of Pleistocene fossils from McFaddin Beach, Texas. Unpublished Master's Thesis, Lamar University, Beaumont, Texas.
Smith, K.T. and Georgalis, G.L. 2022. The diversity and distribution of Palaeogene snakes. A review with comments on vertebral sufficiency, p. 55–84. In Gower, D.J., and Zaher, H. (eds.), The Origin and Early Evolutionary History of Snakes. Cambridge University Press, Cambridge, United Kingdom.
https://doi.org/10.1017/9781108938891.006
Stright, M., Lear, E.M., and Bennett, J.F. 1999. Spatial data analysis of artifacts redeposited by coastal erosion: A case study of McFadden Beach, Texas. US Department of the Interior, Minerals Management Service, 1:1–321.
Szyndlar, Z. and Georgalis, G.L. 2023. An illustrated atlas of the vertebral morphology of extant non-caenophidian snakes, with special emphasis on the cloacal and caudal portions of the column. Vertebrate Zoology, 73:717–886.
https://doi.org/10.3897/vz.73.e101372
Tonni, E. and Cione, A. 1999. Pleistocene continental vertebrates from the present marine shelf of Argentina. Current Research in the Pleistocene, 16:134–136.
Turner, E.S. and Tanner, P. 1994. The McFaddin Beach site on the upper Texas coast. Bulletin of the Texas Archaeological Society, 65:319–336.
Van Essen, H. and Mol, D. 1996. Plio-Pleistocene proboscideans from the southern bight of the North Sea and the Eastern Scheldt, The Netherlands, p. 214–224. In Shoshani, J. and Tassy, P. (eds.), The Proboscidea: Trends in Evolution and Palaeocology. Oxford University Press, Oxford, United Kingdom.
Van Kolfschoten, T. and Laban. C. 1995. Pleistocene terrestrial mammal faunas from the North Sea. Mededelingen Rijks Geologische Dienst, 52:135–151.
Werler, J.E. and Dixon, J.R. 2010. Texas Snakes: Identification, Distribution, and Natural History. University of Texas Press, Austin, Texas.
Whitmore, F.C., Emery, K.O., Cooke, H., and Swift, D.J. 1967. Elephant teeth from the Atlantic continental shelf. Science, 156:1477–1481.
https://doi.org/10.1126/science.156.3781.1477
