

 A new skull of Decennatherium rex Ríos, Sánchez and Morales, 2017 from Batallones-4 (upper Vallesian, MN10, Madrid, Spain)
A new skull of Decennatherium rex Ríos, Sánchez and Morales, 2017 from Batallones-4 (upper Vallesian, MN10, Madrid, Spain)
Article number: 22.2.pvc_1
https://doi.org/10.26879/965
Copyright Paleontological Society, August 2019
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 5 February 2019. Acceptance: 28 June 2019.
{flike id=2653}
ABSTRACT
Giraffids are pecoran ruminants with bi-lobed canines and a special type of cranial appendage called ossicones, which are epiphyseal in origin and don’t fuse completely to the skull until later in life. Although the family Giraffidae has only two extant living African genera, in the past this family was much more diverse and widespread. Today we can recognize more than 40 fossil species, from multiple sites in Eurasia and Africa, which range from the early Miocene (20 m.y.) until today. Here we describe the exceptionally well-preserved remains of the giraffid found in the late Miocene deposits of Batallones-4 (upper Vallesian, MN10, Torrejón de Velasco, Madrid, Spain). Batallones-4 is part of the Cerro de los Batallones fossil site complex, and although it was excavated during the year 2000 there is no published result. The specimen from Batallones-4 has been identified as an old male of Decennatherium rex first identified from Batallones-10 as they share key morphological cranial similarities. This new skull helps us to better understand this species cranial anatomy, as well as its morphological development and variability.
María Ríos. Departamento de Paleobiología, Museo Nacional de Ciencias Naturales- Consejo Superior de Investigaciones Científicas, Madrid, Spain. maria.rios.iba@gmail.com
Jorge Morales. Departamento de Paleobiología, Museo Nacional de Ciencias Naturales- Consejo Superior de Investigaciones Científicas, Madrid, Spain. mcnm166@mncn.csic.es
Keywords: Giraffidae; Artiodactyla; Ruminantia; Miocene; Cerro de los Batallones
Ríos, María and Morales, Jorge. 2019. A new skull of Decennatherium rex Ríos, Sánchez and Morales, 2017 from Batallones-4 (upper Vallesian, MN10, Madrid, Spain). Palaeontologia Electronica 22.2.pvc_1 1-16. https://doi.org/10.26879/965
palaeo-electronica.org/content/pvc_1
Copyright: August 2019 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
Giraffids are crown-pecoran ruminants, living relics of a once large and diverse clade called the Giraffomorpha (Sánchez et al., 2015). The Giraffidae is characterized by possessing bi-lobed canines and a type of epiphyseal cranial appendage called ossicone (Hamilton, 1978). There are only two extant genera, both of them African, Okapia and Giraffa, although more than 40 fossil taxa have been described from multiple sites in Eurasia and Africa (Solounias, 2007). Giraffids range from the early Miocene (20 m.y.) until today (Ríos et al., 2017).
Three late Miocene giraffid species have been described from the Iberian Peninsula: Decennatherium pachecoi Crusafont, 1952 (early Vallesian, MN9), present in the Calatayud-Teruel, Duero and Tagus basins, Decennatherium rex Rios et al., 2017 (late Vallesian, MN10), present in the Madrid basin, and Birgerbohlinia schaubi Crusafont, 1952 (early Turolian, MN11) from Piera and Crevillente-2 (Crusafont, 1952; Montoya and Morales, 1991; Montoya, 1994).
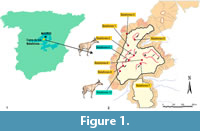 The description of the late Miocene (MN10) Decennatherium rex from the fossil site of Batallones-10 (Madrid Basin, Madrid province, Spain; Figure 1), represented by a complete sample of cranial, dental and postcranial remains shed light on the complex subject of giraffid evolution (Ríos et al., 2017). Decennatherium rex was recovered as a basal offshoot of a sivathere-clade that links together both samotheres and sivatheres, being the earliest evidence of the Sivatherium -ossicone plan (Ríos et al., 2017). In this work we describe the cranial remains of Decennatherium rex from Batallones-4 (BAT4 hereafter) with the aim of comparing them with the type skulls from Batallones-10, describing the intraspecific variability of the species regarding cranial features and specifically that of the cranial appendages.
The description of the late Miocene (MN10) Decennatherium rex from the fossil site of Batallones-10 (Madrid Basin, Madrid province, Spain; Figure 1), represented by a complete sample of cranial, dental and postcranial remains shed light on the complex subject of giraffid evolution (Ríos et al., 2017). Decennatherium rex was recovered as a basal offshoot of a sivathere-clade that links together both samotheres and sivatheres, being the earliest evidence of the Sivatherium -ossicone plan (Ríos et al., 2017). In this work we describe the cranial remains of Decennatherium rex from Batallones-4 (BAT4 hereafter) with the aim of comparing them with the type skulls from Batallones-10, describing the intraspecific variability of the species regarding cranial features and specifically that of the cranial appendages.
Locality
The fossil site of BAT4 (Figure 1) is one of the late Miocene (Vallesian age) localities of the fossil site-complex of Cerro de los Batallones (Batallones Butte). A complete description of the geological setting of the fossil sites in the butte is given by Pozo et al. (2004) and Calvo et al. (2013). Since the discovery of BAT1, the nine researched sites in the Butte have yielded some of the richest and best-preserved Neogene mammal assemblages worldwide (Morales, 2008; Domingo et al., 2011; Valenciano et al., 2015). BAT4, found in the year 2000, was the first fossil locality of Cerro de los Batallones in yielding giraffid remains (Figure 1). During that year some isolated large-size Hipparion remains were also found. The deeper levels of the site were excavated during 2009, yielding a rich micromammal fauna together with other macromammal remains identified as Bovidae and Suidae. When comparing it with the fauna from other sites of the Cerro de los Batallones complex, the BAT4 fauna is that of the BAT10-type sites, composed mainly by large herbivores. However, the fossil association of BAT4 is scarce when comparing it with the astonishingly rich BAT10.
MATERIALS AND METHODS
Fossil Material
We have studied the specimen BAT-04'00-37 from BAT4 curated by the MNCN-CSIC (Madrid, Spain) which consists in a complete skull with both hemimandibles.
Measurements
We follow the set of measurements proposed by Ríos et al., (2016a, 2016b) (S1, figures S1.1-S1.2). All measurements were taken with digital calipers (Table 1, Table 2, Table 3, Table 4, Table 5, Table 6, Table 7).
Nomenclature
We use the terminology of Barone (Barone 1999) for anatomic nomenclature of the cranial l skeleton, and that published by Azanza (2000) for nomenclature of the dentition.
Institutional / technical abbreviations
MNCN-CSIC, Museo Nacional de Ciencias Naturales-CSIC, Madrid, Spain; Batallones-4, BAT4; Batallones-10, BAT10.
SYSTEMATIC PALAEONTOLOGY
Class MAMMALIA Linnaeus, 1758
Order CETARTIODACTYLA Montgelard, Catzeflis and Douzery, 1997
Suborder RUMINANTIA Scopoli, 1777
Infraorder PECORA sensu Webb and Taylor, 1980
Family GIRAFFIDAE Gray, 1821
Genus Decennatherium Crusafont, 1952
Type species. Decennatherium pachecoi Crusafont, 1952, by original designation.
Decennatherium rex (Ríos, Sánchez and Morales, 2017)
Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8
 Holotype
Holotype
BAT-10’13-E2-69, skull with ossicones and mandible (Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8).
Locality, age and horizon
BAT4, BAT10, Cerro de los Batallones, late Vallesian, MN 10, local zone J, Madrid province, Spain. (ca 9 m.y.) (Figure 1).
Emended Diagnosis
Large giraffid with two pairs of ossicones. Smaller anterior pair located above the orbits and anteriorly oriented. Larger posterior pair located posterior to the orbits and posteriorly oriented. Posterior ossicones curved, showing a high number of ridges on their surface. Posterior ossicones exhibit secondary bone growth deposition in adult individuals in the form of dorsal, ventral and dorsoventral round bumps. The location of these bumps has a genetic component. Middle indentation of the hard palate slightly posterior to the M3. Broad occipital. Long premaxillae. Elongated diastema. p3 lacking mesolingual conid. Slightly molarized p4. Cervical vertebrae of medium length. Scapula with acromion. Robust postcranial skeleton. Metacarpals of medium length showing a medium depth palmar trough. Differs from Decennatherium pachecoi in: p3 with an isolated wall-like mesolingual conid structure; anterior stylid and conid of the p3 always present; and more robust metapodials (Ríos et al., 2017).
Material
BAT-04'00-37 (skull and mandible).
ANATOMICAL DESCRIPTION
Ossicones
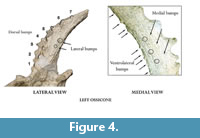
 The skull has two pairs of ossicones (Figure 2, Figure 3, Figure 4, Figure 5, Table 1, Table 2). The smaller anterior pair is located above the orbits on the frontals and are anteriorly oriented forming an angle with the skull roof of 53º. There is a slight degree of torsion on the skull so that the angle represents an approximation of the real condition. Its surface has no ridges. The larger posterior pair (ca. 40 cm) is located posterior to the orbits over the parietals and are oriented posteriorly and slightly outwards, with and angle of insertion to the skull roof of approximately 50º in the right ossicone and 40º in the left one. The posterior ossicones are curved, showing a high number of deep ridges on their surface that run longitudinally from the base to the tip. They also show several (mainly anterior) bumps (Figure 4, Table 2). The right posterior ossicone shows eight large dorsal bumps and a larger ventrolateral one, six small ventrolateral bumps, six small medial bumps and six small ventral bumps. The left posterior ossicone shows seven large dorsal bumps, seven small ventromedial bumps, three small medial bumps, nine small ventrolateral bumps and two small lateral bumps (Figure 4). The tips are blunt.
The skull has two pairs of ossicones (Figure 2, Figure 3, Figure 4, Figure 5, Table 1, Table 2). The smaller anterior pair is located above the orbits on the frontals and are anteriorly oriented forming an angle with the skull roof of 53º. There is a slight degree of torsion on the skull so that the angle represents an approximation of the real condition. Its surface has no ridges. The larger posterior pair (ca. 40 cm) is located posterior to the orbits over the parietals and are oriented posteriorly and slightly outwards, with and angle of insertion to the skull roof of approximately 50º in the right ossicone and 40º in the left one. The posterior ossicones are curved, showing a high number of deep ridges on their surface that run longitudinally from the base to the tip. They also show several (mainly anterior) bumps (Figure 4, Table 2). The right posterior ossicone shows eight large dorsal bumps and a larger ventrolateral one, six small ventrolateral bumps, six small medial bumps and six small ventral bumps. The left posterior ossicone shows seven large dorsal bumps, seven small ventromedial bumps, three small medial bumps, nine small ventrolateral bumps and two small lateral bumps (Figure 4). The tips are blunt.
The surface of the ossicones is very porous, as in extant Giraffa. This may indicate that even with their large size they were covered in integument, irrigated by the cornual artery (O’Brien et al., 2016).
Skull
 There is an almost complete skull in the BAT4 sample (Figure 3, Figure 4, Figure 5, Table 1, Table 2, Table 3, Table 4, Table 5, Table 6, Table 7), only lacking a portion of the premaxillae. Contrary to most of BAT10 skulls, it has no diagenetic dorsoventral deformation, although there is a longitudinal distortion that affects the position of the ossicones (Figure 3).
There is an almost complete skull in the BAT4 sample (Figure 3, Figure 4, Figure 5, Table 1, Table 2, Table 3, Table 4, Table 5, Table 6, Table 7), only lacking a portion of the premaxillae. Contrary to most of BAT10 skulls, it has no diagenetic dorsoventral deformation, although there is a longitudinal distortion that affects the position of the ossicones (Figure 3).
In dorsal view the two supraorbital foramina are placed behind the anterior ossicones on the frontals. The occipital is wide and rugose indicating strong neck musculature and the superior occipital edge extends backwards and upwards. There is no development of the frontal sinus to hollow the ossicones. The nasal bone is prominent and the rostral process of the nasal is developed (Figure 5.1).
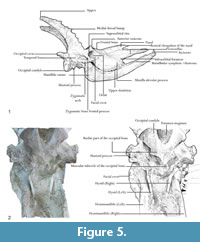 In lateral view the lacrimal canal foramen is closed and the caudal lacrimal process is barely developed. The supraorbital canal orifices are rounded and wide. The ethmoidal fissure is open. The temporal fossa is deep and subrectangular. The orbit is circular, and the supraorbital margin is massive and has a very rugose surface (Figure 5.1). The zygomatic process of the frontal bone is also very developed and rugose, and the zygomatic arch is prominent under the orbit. The occipital condyles are massive and are oriented backwards and slightly upwards. The masseteric is large, with an α value of 8.94 (n=1) and a β value of 87.85º (Figure 6), which fall within the range of the specimens from BAT10 Figure 6. These values indicate a relatively large masseteric fossa in comparison to other giraffids, which may indicate a mixing feeding with slightly more grazing habits (Solounias, 2007). This is also supported by the presence of an enlarged maxilla and a large masseter profundus (Figure 5.1) (Ríos et al., 2017). The rostral elongation of the nasal bone points downwards and forms an angle with the premaxillae of approximately 20º. The premaxilla portion that is preserved is thick proximally, narrows distally, and is oriented downwards. The mandibular symphysis and lower incisors are horizontal in relation to the basipalatal axis. The maxillar tuberosity is very developed.
In lateral view the lacrimal canal foramen is closed and the caudal lacrimal process is barely developed. The supraorbital canal orifices are rounded and wide. The ethmoidal fissure is open. The temporal fossa is deep and subrectangular. The orbit is circular, and the supraorbital margin is massive and has a very rugose surface (Figure 5.1). The zygomatic process of the frontal bone is also very developed and rugose, and the zygomatic arch is prominent under the orbit. The occipital condyles are massive and are oriented backwards and slightly upwards. The masseteric is large, with an α value of 8.94 (n=1) and a β value of 87.85º (Figure 6), which fall within the range of the specimens from BAT10 Figure 6. These values indicate a relatively large masseteric fossa in comparison to other giraffids, which may indicate a mixing feeding with slightly more grazing habits (Solounias, 2007). This is also supported by the presence of an enlarged maxilla and a large masseter profundus (Figure 5.1) (Ríos et al., 2017). The rostral elongation of the nasal bone points downwards and forms an angle with the premaxillae of approximately 20º. The premaxilla portion that is preserved is thick proximally, narrows distally, and is oriented downwards. The mandibular symphysis and lower incisors are horizontal in relation to the basipalatal axis. The maxillar tuberosity is very developed.
 In ventral view (Figure 5.2) the anterolateral edges of the occipital are rugose and prominent and anteriorly oriented. The foramen magum is rounded and deep, and the intercondylar groove is wide. The jugular process is very developed and rugose, but not as much as in Giraffa. The expansion of the retroarticular process is not fused to the bullae. The bullae are not very developed, being similar to those of the specimens from BAT10 and Giraffa rather than the large bullae of Okapia. The position of the sheath of the eardrum varies little relative to the bullae in comparison to other giraffids. The auditory meatus is circular and laterally oriented. The bassioccipital muscular tubercles of the occipital are very prominent and rounded as in the specimens from BAT10 and are much more developed than in Giraffa and Okapia. The mastoid is reduced. The middle indentation of the hard palate is slightly posterior to the M3. The stylohial part of the hyoid bone is long and thin. These features are also present on the skull of the type series from BAT10.
In ventral view (Figure 5.2) the anterolateral edges of the occipital are rugose and prominent and anteriorly oriented. The foramen magum is rounded and deep, and the intercondylar groove is wide. The jugular process is very developed and rugose, but not as much as in Giraffa. The expansion of the retroarticular process is not fused to the bullae. The bullae are not very developed, being similar to those of the specimens from BAT10 and Giraffa rather than the large bullae of Okapia. The position of the sheath of the eardrum varies little relative to the bullae in comparison to other giraffids. The auditory meatus is circular and laterally oriented. The bassioccipital muscular tubercles of the occipital are very prominent and rounded as in the specimens from BAT10 and are much more developed than in Giraffa and Okapia. The mastoid is reduced. The middle indentation of the hard palate is slightly posterior to the M3. The stylohial part of the hyoid bone is long and thin. These features are also present on the skull of the type series from BAT10.
Mandible
The two hemimandibles of BAT-04'00-37 are preserved. The coronoid process is developed, and its uppermost part is oriented laterally. The condylar process is oval and extends medially. The mandibular foramen is large and stands in the caudoventral side of a very shallow pterygoidian fossa. The masseteric fossa is ventrocaudally slightly expanded, and its surface is only slightly rugose except at the vascular incision where there is a more wrinkled and prominent area. This configuration indicates that the deepest part of the masseteric muscle is more developed than the superficial part suggesting grazing masticatory features (König and Liebich, 2005). The diastema is very elongated. The mentonial foramen is large and is located in the middle of the distal part of the mandibular symphysis, right before the incisors start.
Dentition
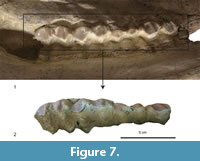 The mouth of BAT-04'00-37 has been preserved in an anatomic closed position, with the upper teeth resting outside the lower teeth. Due to this only part of the upper teeth is visible, and only the length of the lower teeth can be measured. The lower teeth are not visible. The molars show a high degree of wear, to the extent that there is barely any enamel left (Figure 7).
The mouth of BAT-04'00-37 has been preserved in an anatomic closed position, with the upper teeth resting outside the lower teeth. Due to this only part of the upper teeth is visible, and only the length of the lower teeth can be measured. The lower teeth are not visible. The molars show a high degree of wear, to the extent that there is barely any enamel left (Figure 7).
The P4 (Figure 7, Table 6) is wider than long. The labial cone is moderate and close to the anterior style. The anterior style is moderately developed, thicker than the posterior style.
The M1 (Figure 7, Table 6) has a labial wall with strong parastyle and mesostyle and a much weaker metastyle. The enamel is very rugose. The M2 (Figure 7) is similar to the M1 but larger. The protocone and metaconule are lingually V-shaped and are well separated, with the posterior lobe slightly wider than the anterior one. The paracone is very strong. There is a metaconule-fold. The M3 is very similar to the M1 and M2, but it shows a very pointy metaconule and the metacone is rounder than the paracone.
RESULTS
 The giraffid skull from BAT4 is morphologically and metrically almost identical the skulls of the type series of Decennatherium rex, although due taphonomical processes some features are different. For instance, the holotype of Decennatherium rex from BAT10, the skull of a large male (BAT10’13.E2-69) shows dorsoventral deformation, while the specimen from this study only shows torsion on the sagittal plane altering the location and orientation of the ossicones, but preserving almost in perfect conditions otherwise. They also differ somewhat in the ossicone morphology, as the anterior ossicones in the BAT4 specimen are shorter than those found in other male specimens found in BAT10. We also found an extreme case of secondary bone growth in form of bumps in the BAT4 specimen, which has larger and more numerous bumps than the specimens found in BAT10. We also find the specimen with the higher degree of erosion In the BAT4 specimen. Despite these differences, both the specimens from BAT10 and the specimen from BAT4 share key features that separate them from any other giraffid. Besides sharing many cranial features, as the same bassicranial morphology, the morphology of the dentition and mandible, and the development of the occipital region, the specimens from BAT10, and the specimen of BAT10 share the same ossicone bauplan. The location, development and orientation of their ossicones are almost identical. Their ossicones also share the same curvature and ornamentation based on deep longitudinal ridges and secondary bone growth in the form of bumps. Even further, the disposition of the bumps in the posterior ossicones of BAT-04'00-37 is very similar to the bumps present in BAT-10'07-G4-127 (Figure 8). BAT-10'13-E2-69 (Figure 2) has only a large bump, located in the middle of the ossicone shaft dorsally, in the same place as BAT-04'00-37 and BAT-10'07-G4-127 show their larger dorsal bump.
The giraffid skull from BAT4 is morphologically and metrically almost identical the skulls of the type series of Decennatherium rex, although due taphonomical processes some features are different. For instance, the holotype of Decennatherium rex from BAT10, the skull of a large male (BAT10’13.E2-69) shows dorsoventral deformation, while the specimen from this study only shows torsion on the sagittal plane altering the location and orientation of the ossicones, but preserving almost in perfect conditions otherwise. They also differ somewhat in the ossicone morphology, as the anterior ossicones in the BAT4 specimen are shorter than those found in other male specimens found in BAT10. We also found an extreme case of secondary bone growth in form of bumps in the BAT4 specimen, which has larger and more numerous bumps than the specimens found in BAT10. We also find the specimen with the higher degree of erosion In the BAT4 specimen. Despite these differences, both the specimens from BAT10 and the specimen from BAT4 share key features that separate them from any other giraffid. Besides sharing many cranial features, as the same bassicranial morphology, the morphology of the dentition and mandible, and the development of the occipital region, the specimens from BAT10, and the specimen of BAT10 share the same ossicone bauplan. The location, development and orientation of their ossicones are almost identical. Their ossicones also share the same curvature and ornamentation based on deep longitudinal ridges and secondary bone growth in the form of bumps. Even further, the disposition of the bumps in the posterior ossicones of BAT-04'00-37 is very similar to the bumps present in BAT-10'07-G4-127 (Figure 8). BAT-10'13-E2-69 (Figure 2) has only a large bump, located in the middle of the ossicone shaft dorsally, in the same place as BAT-04'00-37 and BAT-10'07-G4-127 show their larger dorsal bump.
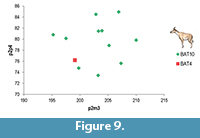
The ossicone index (Transverse Diameter at the Ossicone Base /Length of the Ossicone (Dorsal) of BAT-04'00-37 lies within the range of the BAT 10 specimens (Table 7). The same happens with the maxillae/mandible indices (Figure 9).
DISCUSSION
Sexual Dimorphism in Decennatherium Rex
 The Decennatherium rex skull from BAT4 corresponds to the Morphotype II of Rios et al. (2017). Morphotype I (e.g. BAT-10'08-G3-91) (Figure 2.3) is characterized by the presence of reduced ossicones, with frontal ossicones reduced to unattached little circular disks with a pointy projection in the middle, and slender and shorter fronto-parietal ossicones. Morphotype II (e.g., BAT-10'13-E2-69, BAT-10'07-G4-127) (Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8) is characterized by more robust and developed ossicones (both frontal and fronto-parietal) than those of Morphotype I, showing very deep ridges on their surface and a larger size. The fronto-parietal ossicones found in BAT4 are the largest (410 mm) in the entire Batallones sample, including those from the largest individuals from the type locality of BAT10, although not by much (the largest specimen in the BAT10 sample reaches 382 mm in length).
The Decennatherium rex skull from BAT4 corresponds to the Morphotype II of Rios et al. (2017). Morphotype I (e.g. BAT-10'08-G3-91) (Figure 2.3) is characterized by the presence of reduced ossicones, with frontal ossicones reduced to unattached little circular disks with a pointy projection in the middle, and slender and shorter fronto-parietal ossicones. Morphotype II (e.g., BAT-10'13-E2-69, BAT-10'07-G4-127) (Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8) is characterized by more robust and developed ossicones (both frontal and fronto-parietal) than those of Morphotype I, showing very deep ridges on their surface and a larger size. The fronto-parietal ossicones found in BAT4 are the largest (410 mm) in the entire Batallones sample, including those from the largest individuals from the type locality of BAT10, although not by much (the largest specimen in the BAT10 sample reaches 382 mm in length).
Ríos et al. (2017) linked Morphotype-I and II with female and male individuals, respectively. Hence the specimen BAT-04'00-37, is the most extreme example of a Decennatherium rex Morphotype-II found so far, and we considered it should be regarded as a male. Decennatherium rex males could have used their larger and more robust headgear as a display structure, and also for fighting between bulls in a Tetracerus fashion, with the paired ossicones of the opponents interlocking and applying pressure at the base of the cranial appendages (Bubenik, 1990). The hollow-based ossicones of Decennatherium rex, full of sinuses, is well constructed for use in sparring. Also, their cervical vertebrae are massive (Ríos et al., 2017), as is the occipital region of their skull. These features can be interpreted as reflecting a way to resist cranial wrestling, pushing or even butting (Churcher, 1978; Churcher, 1990).
Secondary Bone Apposition in Decennatherium Rex
The bumps present in the Decennatherium rex specimens are very similar to those of Sivatherium maurusium/olduvaiense Pomel, 1892, Schansitherium tafeli Killgus, 1922, or Giraffa camelopardalis Linneo, 1758 (Solounias, 2007; Guan, 1997) as they all represent examples represent secondary bone apposition in giraffid ossicones and even have a similar rounded and protruding morphology. In the sample from Cerro de los Batallones (BAT10 and BAT4) these bumps are mainly located over the dorsal surface, though they can also appear laterally (Figure 5, Figure 6, Figure 7). The largest bump appears always in the middle of the dorsal surface, more or less at the same height both in the right and left ossicones of each individual. The distribution of the bumps in the fronto-parietal ossicones of Decennatherium rex agrees with the observations of Solounias et al. (1988) who pointed out that these small irregular protuberances have a somewhat fixed position, with the first and largest bump always located dorsally and in the middle of the ossicone shaft, suggesting a genetic basis for the location of these secondary bone growth. Although these do not seem so easy to appreciate in Giraffa, the ossicones pictured by Singer and Boné (1960) show at least one large bump on the proximal part of the posterior ossicones of Sivatherium maurusium/olduvaiense that seems present in most of the pictured specimens (Singer and Boné, 1960: plates I-IV), also supporting Solounias observations (Solounias, 2007). There is, however, intraspecific variability, with some specimens showing either a higher number of bumps or larger-sized bumps, and also bumps in different locations. According to Harris (1974) the disparity in size and development of the bumps may represent individual variation. It is also possible that the amount of secondary bone apposition, in the form of ornamentation, may have an ontogenetic component and be correlated to the age of the individual. This is the case of the developmental process occurring in older males of Giraffa camelopardalis (Solounias and Tang, 1990) which accumulate secondary bone growth from fighting. This scenario could also be true for Decennatherium rex, Sivatherium maurusium/olduvaiense and Schansitherium tafeli, as we find specimens with bumps within the three taxa; and in the case of Decennatherium rex, we can also correlate them with the age thanks to the preservation of the dentition. Another important similarity is that as we do with Giraffa camelopardalis, we also find male specimens without the bumps within all three taxa (Singer and Boné, 1960; Guan, 1997; Geraads, 2009; Ríos et al., 2017) indicating these may represent younger adult individuals.
As for sexual dimorphism in the presence of secondary bone apposition, while in Giraffa camelopardalis older males the secondary bone covers the entire calvaria, the females show little or no such secondary bone growth. The same could be the case for Decennatherium rex females (morphotype I), as we have not found any evidence yet of morphotype I ossicones with any kind of secondary bone apposition in the form of bumps.
The morphology of the fronto-parietal ossicones of BAT-04'00-37, which has more numerous and larger bumps than the BAT10 specimens, fits in the hypothesis of the secondary bone apposition being related to both: age and sex. BAT-04'00-37 skull shows the largest bumps, but also the most worn-out molars (with almost no enamel left), indicating an advanced age for this specimen. This hypothesis must be checked through a) direct histologic analysis of the ossicones and b) microwear analysis to discard the possibility of a more abrasive diet as the agent of the advanced molar wearing (Danowitz et al., 2016), but it appears that these three variables (age, sex and secondary bone apposition) could be intrinsically related not only in the extant Giraffa camelopardalis, but also in fossil giraffids dated as far back as the late Miocene, and it may be also indicating a common ancestor for these taxa.
Feeding Habits in Decennatherium Rex
Following Solounias (2007) we have analyzed the shape and size of the masseteric fossa as well as other cranial and masticatory features. BAT-04'00-37 results very similar to BAT10 specimens, with α values of 8.94 (n=1) and β values of 87.85º (Figure 4). According to Ríos et al., (2017) these angles fall within the same category as Schansitherium tafeli, Palaeotragus microdon Bohlin, 1926, and Samotherium boissieri Forsyth Major, 1888 (Solounias, 2007). Only Samotherium major Bohlin, 1926, shows α values over 10º while Canthumeryx sirtensis Hamilton, 1973, Okapia johnstoni Sclater, 1901, and Giraffa camelopardalis show α values below 5º. This confirms that Decennatherium rex has a relatively larger masseteric fossa than most giraffids and may indicate more grazing habits than in the extant taxa (Ríos et al., 2017). Decennatherium rex also shows other grazer masticatory features as an enlarged maxilla and a large masseter profundus. As in other grazers the enlarged maxilla of Decennatherium rex tends to move the orbit posteriorly starting above the M3 or further back (Solounias and Dawson-Saunders, 1988).
CONCLUSIONS
We describe here a new complete skull of Decennatherium rex from the fossil site of Batallones-4 (MN10, Madrid basin, Spain). The complete skull BAT-04'00-37 corresponds to the Morphotype-II described for this species (identified as males) and offers an insight into the intraspecific variability of Decennatherium rex relative to the sexual dimorphism of the cranial appendages. The characteristic features of Morphotype-II are more developed in BAT-04'00-37 than in all the Morphotype-II specimens from the type locality of Batallones-10, with fronto-parietal ossicones sporting larger and more numerous bumps than the BAT10 specimens. The heavily worn-out molars of BAT-04'00-37 very possibly indicate an advanced age for this specimen, and thus the more developed structures of the fronto-parietal ossicones could be related to secondary bone apposition as in extant male giraffes. However this hypothesis will be checked out through the direct histologic analysis of the ossicones.
ACKNOWLEDGEMENTS
MR acknowledges a FPI predoctoral grant (Spanish Government MINECO) as well as the EEBB-FPI grant program 2013, 2014 and 2015 The Spanish Research Project CGL2015-68333-P (MINECO/FEDER-UE) and the Research Groups CSIC 641538 and CAM-UCM 910607 funded aspects of this research. We also want to thoughtfully thank P. Perez and S. Fraile (MNCN-CSIC, Madrid, Spain), L. Celia and D. DeMiguel (ICP, Barcelona, Spain), P. Montoya (MGUV, Valencia, Spain), J. Galkin, J. Meng and E.Westwig (AMNH; New York, USA), P. Brewer and S. Pappa (NHM, London, UK), C. Argot and S. Peigne (MNHN, Paris, France) and their respective home institutions for giving us access to their giraffid fossils. We thank E. Cantero, B. Gómez and P. Gutierrez for the preparation of the BAT4 and BAT10 remains. We thank O. Sanisidro and R. Guerrero Torralba for the Decennatherium rex illustrations.
REFERENCES
Azanza, B. 2000. Los Cervidae (Artiodactyla, Mammalia) del Mioceno de las cuencas del Duero, Tajo, Calatayud-Teruel y Levante. Memorias del Museo de Paleontología de la Universidad de Zaragoza, 8:1-376.
Barone, R. 1999. Anatomie Comparée des Mammifères Domestiques: Ostéologie. Vigot Frères, Paris, 761 pp.
Bubenik, A.B. 1990. Epigenetical, morphological, physiological, and behavioral aspects of evolution of horns, pronghorns, and antlers. p. 3-113. In Bubenik, G.A. and Bubenik, A.B. (eds.), Horns, Pronghorns, and Antlers. Springer, New York.
Calvo, J.P., Pozo, M., Silva, P.G., and Morales, J. 2013. Pattern of sedimentary infilling of fossil mammal traps formed in pseudokarst at Cerro de los Batallones, Madrid Basin, central Spain. Sedimentology, 60:1681-1708. https://doi.org/10.1111/sed.12048
Churcher, C.S. 1978. Chapter 25. Giraffidae; p. 509-535. In Maglio, V.J. and Cooke, H.B.S. (eds.), Evolution of African Mammals. Harvard University Press, Cambridge.
Churcher, C. 1990. Cranial Appendages of Giraffoidea, p. 180-94. In Bubenik, G. and Bubenik, A., (eds). Horns, Pronghorns, and Antlers. Springer, New York.
Crusafont, M. 1952. Los Jiráfidos Fósiles de España. Published PhD Thesis. Diputación Provincial de Barcelona. Memorias y comunicaciones del Instituto Geológico VIII: Premio extraordinario de Doctorado, Barcelona, Spain.
Danowitz, M., Hou, S., Mihlbachler, M., Hastings, V., and Solounias, N. 2016. A combined-mesowear analysis of late Miocene giraffids from North Chinese and Greek localities of the Pikermian Biome. Palaeogeography, Palaeoclimatology, Palaeoecology, 449:194-204. https://doi.org/10.1016/j.palaeo.2016.02.026.
Domingo, M.S., Domingo, L., Sánchez, I.M., Alberdi, M.T., Azanza, B., and Morales, J. 2011. New insights on the taphonomy of the exceptional mammalian sites of Cerro de los Batallones (Late Miocene, Spain) based on rare earth element geochemistry. Palaios, 26:55-65.https://doi.org/10.2110/palo.2010.p10-047r.
Geraads, D. 2009. Giraffidae (Mammalia) de la fin du Néogène de la République de Macédoine (ARYM). Geodiversitas, 31:893-909.https://doi.org/10.5252/g2009n4a893.
Gray, J.E. 1821. On the natural arrangement of vertebrate animals. London Medical Repository, 15:297-310.
Guan, J. 1997. The study on Schansitherium from Fugu, Shaanxi. Memoirs of Beijing Natural History Museum, 56:125-131.
Hamilton, W.R. 1978. Fossil giraffes from the Miocene of Africa and a revision of the phylogeny of the Giraffoidea. Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences, 283:165-229.
Harris, J.M. 1974. Orientation and variability in the ossicones of African Sivatheriinae (Mammalia: Giraffidae). Annals of the South African Museum, 65:189-198.
König, H.E. and Liebich, H.G. 2005. Anatomía de los Animales Domésticos: Texto y Altlas en Color. Ed. Médica Panamericana, Ciudad de México, 304 pp.
Linnaeus, C. 1758. Systema Naturae per Regna tria Naturae, secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Tenth edition. Laurentii Salvii, Stockholm, Sweden.
Montgelard, C., Catzeflis, F.M., and Douzery, E. 1997. Phylogenetic relationships of artiodactyls and cetaceans as deduced from the comparison of cytochrome b and 12S rRNA mitochondrial sequences. Molecular Biology and Evolution, 14:550-559.https://doi.org/10.1093/oxfordjournals.molbev.a025792.
Montoya P. 1994. Los Macromamíferos del Mioceno Uperior del Area de Crevillente (Alicante). PhD Thesis, Universitat de València, Facultat de Ciencies Biologiques, Departament de Geologia. 1994. http://roderic.uv.es/handle/10550/38910
Montoya, P. and Morales, J. 1991. Birgerbohlinia schaubi Crusafont, 1952 (Giraffidae, Mammalia) del Turoliense inferior de Crevillente-2 (Alicante, España). Filogenia e historia biogeográfica de la subfamilia Sivatheriinae. Bulletin du Muséum National d’Histoire Naturelle, Paris, 4º sér., 13:177-200.
Morales, J., Pozo, M., Silva, P.G., Domingo, M.S., López-Antoñanzas, R., Álvarez Sierra, M.A., Antón, M., Martín Escorza, C., Quiralte, V., Salesa, M.J., Sánchez, I.M., Azanza, B., Calvo, J.P., Carrasco, P., García-Paredes, I., Knoll, F., Hernández Fernández, M., van den Hoek Ostende, L., Merino, L., van der Meulen, A.J., Montoya, P., Peigné, S., Peláez-Campomanes, P., Sánchez-Marco, A., Turner, A., Abella, J., Alcalde, G.M., Andrés, M., DeMiguel, D., Cantalapiedra, J. L., Fraile, S., García Yelo, B.A., Gómez Cano, A.R., López Guerrero, P., Oliver Pérez, A., and Siliceo, G. 2008. El sistema de yacimientos de mamíferos miocenos del Cerro de los Batallones, Cuenca de Madrid: estado actual y perspectivas. Seminario Paleontología Zaragoza, 8:41-117.
O’Brien, H.D., Gignac, P.M., Hieronymus, T.L., and Witmer, L.M. 2016. A comparison of postnatal arterial patterns in a growth series of giraffe (Artiodactyla: Giraffa camelopardalis). PeerJ, 4:e1696. https://doi.org/10.7717/peerj.1696
Pozo, M., Calvo, J.P., Silva, P.G., Morales, J., and Peláez-Campomanes, P. 2004. Geologıa del sistema de yacimientos de mamıferos miocenos del Cerro de los Batallones, Cuenca de Madrid. Geogaceta, 35:143-159.
Ríos, M., Danowitz, M., and Solounias, N. 2016a. First comprehensive morphological analysis on the metapodials of Giraffidae. Palaeontologia Electronica, 19:1-39. https://doi.org/10.26879/653.
Ríos, M., Sánchez, I.M., and Morales, J. 2016b. Comparative anatomy, phylogeny, and systematics of the Miocene giraffid Decennatherium pachecoi Crusafont, 1952 (Mammalia, Ruminantia, Pecora): State of the art. Journal of Vertebrate Paleontology, 36:e1187624. https://doi.org/10.1080/02724634.2016.1187624.
Ríos, M., Sánchez, I.M., and Morales, J. 2017. A new giraffid (Mammalia, Ruminantia, Pecora) from the late Miocene of Spain, and the evolution of the sivathere-samothere lineage. PloS One, https://doi.org/10.1371/journal.pone.0185378
Sánchez, I.M., Cantalapiedra, J.L., Ríos, M., Quiralte, V., and Morales, J. 2015. Systematics and evolution of the Miocene three-horned palaeomerycid ruminants (Mammalia, Cetartiodactyla). PloS One, https://doi.org/10.1371/journal.pone.0143034
Scopoli, G.A. 1777. Introductio ad Historiam Naturalem, Sistens Genera Lapidium, Planatarum, et Animalium Hactenus Detecta, Caracteribus Essentialibus Donata, in Tribus Divisa, Subinde ad Leges Naturae. Prague, Gerle, 506 pp.
Singer, R. and Boné, E. 1960. Modern giraffes and the fossil giraffids of Africa. Annals of the South African Museum, 45:375-548.
Solounias, N. 2007. Family Giraffidae, p. 257-277. In Prothero, D.R.F. and Foss, S.E. (eds.), The Evolution of Artiodactyls. The Johns Hopkins University Press, Baltimore.
Solounias, N. and Dawson-Saunders, B. 1988. Dietary adaptations and paleoecology of the Late Miocene ruminants from Pikermi and Samos in Greece. Palaeogeography, Palaeoclimatology, Palaeoecology, 65:149-172. https://doi.org/10.1016/0031-0182(88)90021-1
Solounias, N. and Tang, N. 1990. The two types of cranial appendages in Giraffa camelopardalis (Mammalia, Artiodactyla). Journal of Zoology, 222:293-302.
Solounias, N., Teaford, M., and Walker, A. 1988. Interpreting the diet of extinct ruminants: The case of a non-browsing giraffid. Paleobiology, 69:845-848.
Valenciano, A., Abella, J., Sanisidro, O., Hartstone-Rose, A., Álvarez-Sierra, M.Á., and Morales, J. 2015. Complete description of the skull and mandible of the giant mustelid Eomellivora piveteaui Ozansoy, 1965 (Mammalia, Carnivora, Mustelidae), from Batallones (MN10), late Miocene (Madrid, Spain). Journal of Vertebrate Paleontology, 35:e934570. https://doi.org/10.1080/02724634.2014.934570
Webb, S.D. and Taylor, B.E. 1980. The phylogeny of hornless ruminants and a description of the cranium of Archaeomeryx. Bulletin of the American Museum of Natural History, 167:117-158.
