

 A taxonomic revision of the Cenomanian bothremydid turtle Algorachelus parva from Israel and morphological variation within its genus
A taxonomic revision of the Cenomanian bothremydid turtle Algorachelus parva from Israel and morphological variation within its genus
Article number: 28.1.a2
https://doi.org/10.26879/1398
Copyright Palaeontological Association, January 2025
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 12 April 2024. Acceptance: 20 December 2024.
ABSTRACT
The bothremydid turtle Algorachelus is identified as the oldest lineage of pelomedusoid turtles (Pleurodira) that dispersed from Gondwanan to Laurasian regions. It is recognized in the Cenomanian (early Late Cretaceous) of the Middle East (in Israel), Europe (in the Iberian Peninsula), and North America (in Utah). The European and North American species were identified and defined less than 10 years ago. However, two putative different species, originally attributable to the genus Podocnemis (i.e., a member of Podocnemididae now identified as exclusive to South America), were defined in the lower or middle Cenomanian levels of the ‘Ein Yabrud limestone quarry (Binyamin Region, West Bank, Israel). Over almost 50 years, no new data on the Cenomanian taxon or taxa of Bothremydidae from Israel have been published. A detailed first-hand review of the previously documented material is presented here. Several errors in the anatomical interpretation of these specimens are corrected. Unpublished articulated and well-preserved specimens from the type locality are analyzed. Characters not yet defined for the bothremydid or bothremydids from Israel, but essential for the comparison with those of Europe and North America, are studied. In addition, several anatomical regions hitherto not analyzed for the Israeli material, some of them being poorly known for Algorachelus (such as the skull) or never documented for this genus (such as the lower jaw), are analyzed. The presence of a single species in Israel is justified, being recognized as Algorachelus parva. Intraspecific variability is recognized. An emended diagnosis is proposed for this species, which belongs to a remarkably successful lineage of pleurodires that is characterized by its long persistence and multicontinental geographic range.
Adán Pérez-García. Grupo de Biología Evolutiva, Dpto. de Física Matemática y de Fluidos, Facultad de Ciencias, UNED, Avda. Esparta s/n, 28232, Las Rozas, Madrid, Spain. a.perez.garcia@ccia.uned.es
Keywords: Middle East; early Late Cretaceous; Pleurodira; Bothremydidae; Bothremydini.
Final citation: Pérez-García, Adán. 2025. A taxonomic revision of the Cenomanian bothremydid turtle Algorachelus parva from Israel and morphological variation within its genus. Palaeontologia Electronica, 28(1):a2.
https://doi.org/10.26879/1398
palaeo-electronica.org/content/?view=article&id=5424:a-bothremydid-turtle-in-israel&catid=707
Copyright: January 2025 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Bothremydidae is the most abundant and diverse group of pleurodiran turtles known from the Mesozoic fossil record (Lapparent de Broin, 2001; Gaffney et al., 2006; Pérez-García, 2017). Although most pleurodires are recognized as freshwater species (as in all current representatives), the adaptation of several of their lineages to transitional, coastal and even open marine environments, facilitated their dispersal from areas that had been part of Gondwana to Laurasian regions (Broin, 1988; Gaffney et al., 2011; Ferreira et al., 2015; Pérez-García et al., 2017a; Pérez-García, 2023). Thus, the oldest lineage of bothremydids recognized in Laurasia is that of Algorachelus Pérez-García, 2016, which reached Europe at least in the early Cenomanian, probably at the end of the Early Cretaceous; and North America at least in the late Cenomanian, thanks to its adaptation to coastal environments (Pérez-García, 2016; Pérez-García and Rubio, 2024). The description of European (Algorachelus peregrina Pérez-García, 2016) and North American species [ Algorachelus tibert (Joyce, Lyson and Kirkland, 2016)] was published in the last decade, from fossil specimens analyzed in detail (Pérez-García, 2016; Joyce et al., 2016). Algorachelus is also identified in areas that formerly belonged to Gondwana and is identified here as corresponding to finds from the lower or middle Cenomanian ‘Ein Yabrud quarry in Israel (Pérez-García, 2018), where a relatively diverse vertebrate fauna has been recognized, among which the reptiles are not only represented by this turtle but also by a mosasaur and some hind-limbed snakes (see Khalloufi et al., 2010; and references therein). In contrast to the North American and especially the European Algorachelus species, knowledge of the Israeli material (constrained to Haas, 1978a, b) does not taxonomically align with modern taxonomic understanding of Bothremydidae. In fact, the Israeli form has not been formally attributed to a bothremydid genus, but rather two species of the podocnemidid genus Podocnemis, which has modern representatives (Gaffney et al., 2011; Ferreira et al., 2018; Cadena and Vanegas, 2023).
In their review of the lineage of Bothremydidae, Gaffney et al. (2006) recognized that, following the ideas proposed by some previous authors (Antunes and Broin, 1988; Broin, 1988; Lapparent de Broin and Werner, 1998), both Israeli species (i.e., ‘Podocnemis’ parva Haas, 1978a, and ‘Podocnemis judaea’ Haas, 1978b) could form part of the variability of a single taxon. However, the material was not re-examined critically or adequately characterized and were attributed to Bothremydini incertae sedis (Gaffney et al., 2006). After the description of the oldest genus of Bothremydidae in Laurasia (i.e., Algorachelus), based on specimens from the uppermost middle or lowermost upper Cenomanian fossil site of Algora, in central Spain (Pérez-García, 2016), and its subsequent identification in other Iberian localities (i.e., the middle Cenomanian locality of Nazaré, in west central Portugal; Pérez-García et al., 2017b), both the North American species Paiutemys tibert Joyce, Lyson and Kirkland, 2016 and the aforementioned material from Israel presented by Haas (1978a, 1978b) were reassigned to Algorachelus. This decision was consistent with the view of Gaffney et al. (2006) wherein the Israeli material did not belong to a bothremydid genus known at the time, despite having been referred to Bothremys by other workers (Antunes and Broin, 1988; Lapparent de Broin and Werner, 1998). However, no new data on the turtles from Israel have been provided so far, although they were justified as attributable to a single species by Pérez-García (2016), i.e., Algorachelus parva (Haas, 1978a), exclusively based on the information published by Haas (1978a, 1978b).
There is currently no valid diagnosis for Algorachelus parva. Thus, Pérez-García (2018) only indicated some potential differences with other species of the genus. However, it is possible that at least some of them may be based on erroneous data or problematic interpretations provided in the works of Haas (1978a, 1978b), or on characters that may demonstrate intraspecific variability considering new specimens. The objective of this work is a detailed study of the pleurodiran specimens from ‘Ein Yabrud, based on the first-hand examination of all previously documented specimens and the study of unpublished ones. The current analysis expands our knowledge of the only known Gondwanan species of Algorachelus, the earliest dispersing turtle in Laurasia (Pérez-García, 2016, 2018).
MATERIAL AND METHODS
The detailed review of the pleurodiran specimens from ‘Ein Yabrud presented by Haas (1978a, 1978b) is performed here, based on the first-hand examination of all of them. Thus, it is intended to provide new anatomical data on the shell of the species from the lower or middle Cenomanian of Israel, which will help both its characterization and the recognition of intraspecific variability. Furthermore, almost five decades after the original identification of these remains, unpublished specimens of Algorachelus parva have been recognized in ‘Ein Yabrud, and some of them are very well-preserved. They are presented and analyzed here.
In addition to the shell, data on the skull of Algorachelus parva is provided here for the first time. Cranial information on the genus Algorachelus is currently limited to a partial skull of the Iberian Algorachelus peregrina (specimen ALG 64) which is lacking its anterior region (Pérez-García, 2016).
Institutional and Collection Abbreviations
ALG, Algora collection, deposited in the Museo de Paleontología de Castilla-La Mancha, Cuenca, Spain; EY, ‘Ein Yabrud Collection of the Paleontological Collection of The Hebrew University of Jerusalem (HUJ-PAL or HUJP), Jerusalem, Israel.
Anatomical Abbreviations
ab, abdominal; an, anal; ang, angular; c, costal; den, dentary; ep, epiplastron; ex, extragular; fe, femoral; fr, frontal; gu, gular; hp, hypoplastron; hu, humeral; hy, hyoplastron; ju, jugal; m, marginal; ms, mesoplastron; mx, maxilla; nu, nuchal; op, opisthotic; p, peripheral; pa, parietal; pc, pectoral; pf, prefrontal; pl, pleural; pm, premaxilla; po, postorbital; py, pygal; qu, quadrate; so, supraoccipital; sp, suprapygal; sq, squamosal; v, vertebral; xi, xiphiplastron.
SYSTEMATIC PALEONTOLOGY
Order TESTUDINES Batsch, 1788
Suborder PLEURODIRA Cope, 1864
Hyperfamily PELOMEDUSOIDES Cope, 1868
Family BOTHREMYDIDAE Baur, 1891
Tribe BOTHREMYDINI Gaffney, Tong and Meylan, 2006
Subtribe BOTHREMYDINA Gaffney, Tong and Meylan, 2006
Genus ALGORACHELUS Pérez-García, 2016
Type species. Algorachelus peregrina Pérez-García, 2016
Algorachelus parva (Haas, 1978a)
Figure 1, Figure 2, Figure 3
1978b Podocnemis parva; Haas, p. 169, 172, 175.
1978b Podocnemis judaea; Haas, p. 169-175, figures 1-4.
1988 Bothremys parva; Antunes and Broin, p. 163, 165, 174, 177, 179, 182, 184-185.
1988 Bothremys parva; Broin, p. 134.
1998 Bothremys parva; Lapparent de Broin and Werner, p. 164, 169.
2005 ‘Podocnemis’ parva; Zalmout, Mustafa, and Wilson, p. 158, 160, 171.
2005 ‘Podocnemis’ judaea; Zalmout, Mustafa, and Wilson, p. 158, 160, 171.
2006 ‘Podocnemis’ judea; Gaffney, Tong, and Meylan, p. 110.
2006 ‘Podocnemis’ parva; Gaffney, Tong, and Meylan, p. 110, 569, 683.
2012 ‘Podocnemis parva’; Rabi, Tong, and Botfalvai, p. 11.
2012 ‘Podocnemis judea’; Rabi, Tong, and Botfalvai, p. 11.
2013 ‘Podocnemis’ parva; Georgalis, Velitzelos, Velitzelos, and Kear, p. 284.
2013 ‘Podocnemis’ judea; Georgalis, Velitzelos, Velitzelos, and Kear, p. 284.
2016 ‘Podocnemis’ parva; Pérez-García, p. 710, 720-722, 726, 728.
2016 ‘Podocnemis’ parva; Joyce, Lyson, and Kirkland, p. 6, 15-16.
2017b Algorachelus parva; Pérez-García, Antunes, Barroso-Barcenilla, Callapez, Segura, Soares, and Torices, p. 65, 67.
2018 Algorachelus parvus; Pérez-García, p. 119, 132-134.
2020 Algorachelus parva; Pérez-García, p. 366.
2020 Algorachelus parva; Pérez-García, Bardet, Fregenal-Martínez, Martín-Jiménez, Mocho, Narváez, Torices, and Vullo, p. 2.
2024 Algorachelus parva; Pérez-García and Rubio, p. 2-4.
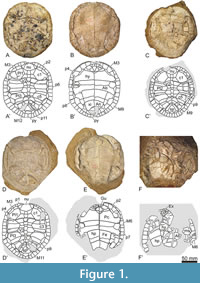 Type material. The shell EY795 (Figure 1A-B) corresponds to the holotype of ‘Podocnemis’ parva (referred as HUJP-Testudinata-3 by Haas 1978a), whose photographs, in dorsal and ventral views, were published by Haas (1978a, figures 3 and 4, respectively), as well as a schematic representation of the complete shell of the species, especially based on this individual (Haas, 1978a, figures 1 and 2). Three paratypes were established by Haas (1978a): the partial carapace EY790 (Figure 1F; referred as HUJP-Testudinata-4 by Haas 1978a), the partial skeleton EY797 (Figure 2A-B, Figure 3B-C; referred as HUJP-Testudinata-2 by Haas 1978a), and the shell EY792 (Figure 2D-E; referred as HUJP-Testudinata-1 by Haas 1978a). Photographs but no schematic representations of these paratypes were published (see figure 9 for EY790, figures 7-8 for EY797, and figures 5-6 for EY792 in Haas, 1978a).
Type material. The shell EY795 (Figure 1A-B) corresponds to the holotype of ‘Podocnemis’ parva (referred as HUJP-Testudinata-3 by Haas 1978a), whose photographs, in dorsal and ventral views, were published by Haas (1978a, figures 3 and 4, respectively), as well as a schematic representation of the complete shell of the species, especially based on this individual (Haas, 1978a, figures 1 and 2). Three paratypes were established by Haas (1978a): the partial carapace EY790 (Figure 1F; referred as HUJP-Testudinata-4 by Haas 1978a), the partial skeleton EY797 (Figure 2A-B, Figure 3B-C; referred as HUJP-Testudinata-2 by Haas 1978a), and the shell EY792 (Figure 2D-E; referred as HUJP-Testudinata-1 by Haas 1978a). Photographs but no schematic representations of these paratypes were published (see figure 9 for EY790, figures 7-8 for EY797, and figures 5-6 for EY792 in Haas, 1978a).
Other referred specimens. Four additional specimens from the type locality and horizon are here recognized as attributable to Algorachelus parva. The shell EY791 (Figure 1D-E) corresponds to the holotype of ‘Podocnemis judaea’ (referred as HUJP3664 by Haas 1978b), presented both through photographs (Haas, 1978b, figures 3 and 4) and schematic representations (Haas, 1978b, figures 1 and 2). The shell EY796 (Figure 1C), the partial skeleton EY793 (Figure 2C, Figure 3A), and the partial shell EY794 (Figure 2F) correspond to so far unpublished specimens.
Type locality and horizon. ‘Ein Yabrud limestone quarry, near the village of Beit Eil, Binyamin Region, West Bank, Israel, Middle East. Bet-Meir Formation or the slightly younger Amminadava Formation, middle part of the Judea Group, lower or middle Cenomanian (see Khalloufi et al., 2010; Cawley and Kriwet, 2018; and references therein).
Emended diagnosis. Member of Algorachelus differing from the other species of this genus by: a posterior carapace with evenly rounded, rather than nearly straight lateral sides; subtriangular to generally triangular first pair of marginals scutes, with short medial margins and a short overlap on the antero-lateral edges of the nuchal plate (covering less than half, and usually less than a third, of its length), and lacking a medial contact (i.e., allowing the first vertebral scute reaching the carapace margin) or showing a very short contact (i.e., the first vertebral occupying a position very close to that margin); wide vertebrals, the second being at least one and a half times wider than long; short anterior plastral lobe, not reaching the anterior carapace margin. It shares with Algorachelus peregrina, but not with Algorachelus tibert, the presence of a wider than long nuchal.
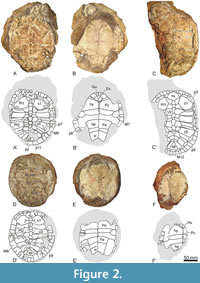 Description. All specimens referred here are represented by fully or partially articulated shells. The maximum carapace length reached approximately 22 cm in EY796 (Figure 2C). The carapace morphology varies from subrounded (Figure 1C, Figue 2D) to subelliptical (Figure 2C). The posterolateral margins of the carapace ranged from gently curved (Figure 1A, Figure 2A, C, D) to nearly straight (Figure 1C, D). A slight (Figure 1A, C; Figure 2D) to almost absent (Figure 2C) anterior notch, restricted to the nuchal plate and the medial half of the first pair of peripherals, is present on the anterior carapace margin.
Description. All specimens referred here are represented by fully or partially articulated shells. The maximum carapace length reached approximately 22 cm in EY796 (Figure 2C). The carapace morphology varies from subrounded (Figure 1C, Figue 2D) to subelliptical (Figure 2C). The posterolateral margins of the carapace ranged from gently curved (Figure 1A, Figure 2A, C, D) to nearly straight (Figure 1C, D). A slight (Figure 1A, C; Figure 2D) to almost absent (Figure 2C) anterior notch, restricted to the nuchal plate and the medial half of the first pair of peripherals, is present on the anterior carapace margin.
The nuchal plate is wider than long (Figure 1A, C, D; Figure 2A, C, D). Its anterior margin is equal or greater than half of the maximum width of the plate. Although the most common condition is the presence of six neurals (Figure 1A, C; Figure 2A, C, D), seven may also be present (see specimen EY791 in Figure 1D). The first neural is subrectangular (Figure 1A, C, D; Figure 2A, C, D), and the last one is pentagonal. The remainder are hexagonal and are generally longer than wide, with shorter antero-lateral than postero-lateral margins. Of the eight pairs of costals, a medial contact occurs between the sixth and the eighth in most specimens (Figure 1A, C; Figure 2A, C, D). However, contact occurs exclusively between the seventh and eighth pairs in EY791 (Figure 1D). The suprapygal is subtriangular and approximately as wide as long (Figure 1A, C; Figure 2C) or slightly wider than long (Figure 1D, Figure 2D). The pygal plate is approximately rectangular, and almost as wide as long.
The second vertebral scute is the widest and at least one and a half times wider than long (Figure 1A, C, D; Figure 2A, C). The first vertebral contacts the anterior carapace margin in EY795 (Figure 1A) or reaches a position very close to it in others (Figure 1C; Figure 2A, C, D). Laterally, this scute contacts the second pair of peripherals (Figure 1A, D) or a nearby position (Figure 1C, Figure 2C). The contact between the first and second, second and third, third and fourth, and fourth and fifth vertebrals is located on the first, third, fifth or sixth, and last pair of costals, respectively (Figure 1A, C, D; Figure 2A, C, D). The pleural scutes overlap the medial region of the peripheral plates. The first pair of marginals is generally subtriangular, showing a short medial margin. The first marginal scutes are wider than long. The overlap of these scutes on the antero-lateral edges of the nuchal are short, equivalent to less than half (Figure 2C) and even less than a third (Figure 1D; Figure 2D) of the length of the plate.
In ventral view, it is observed that the anterior plastral lobe is short, so that it does not reach the anterior edge of the carapace (Figure 1B, E). The anterior plastral lobe is visibly shorter than the posterior lobe (Figure 1B, E; Figure 2B). The anterior lobe is broadly rounded but the lateral margins of the posterior are nearly straight. The entoplastral morphology varies from subhexagonal, being a third wider than long (Figure 1B), to subrhomboidal, both dimensions being subequal (Figure 1E, Figure 2B). A pair of subrounded and laterally positioned mesoplastra are present (Figure 1B, E; Figure 2B, E). There is a well-developed anal notch that is more than twice as wide as long, and its profile is V-shaped (Figure 2B) or U-shaped (Figure 1B, E; Figure 2E).
The length of the gular scute is variable: in some specimens, it overlaps less than a quarter of the maximum entoplastral length (see specimen EY791 in Figure 1E) but can reach the pectoral scutes in others (see specimen EY795 in Figure 1A). When the latter condition is present, the humeral scutes lack a medial contact. The extragulars are triangular and show a relatively small size and can contact the anterior entoplastral margin (e.g., EY795; Figure 1B). The pectorals do not reach the epiplastra (Figure 1B, E, F; Figure 2B, F). However, they can overlap the anterior region of the mesoplastra (e.g., Figure 1B, E). A relatively long distance is observed between the anal scutes and the hypoplastra.
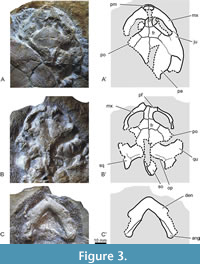 The skull is preserved in two individuals of Algorachelus parva studied herein (EY793 and EY797; Figure 2A-C; Figure 3). They are exposed from the matrixin dorsal view (Figure 3A, B). Both are dorsoventrally crushed. However, the skull is interpreted as a relatively wide element, its length being slightly greater than its width. It is wide along its entire length and has a short rostral region, with a well-defined anterior angle. The orbits are dorsolaterally facing. Nasals and an interorbital groove are absent. The prefrontals do not touch the parietals, due to the frontals contact along the midline (Figure 3A, B). The prefrontals are longer than wide. The frontals are slightly wider than long. The parietals do not contact the orbits, but the jugals do. No interparietal or other cranial scutes are recognized. A relatively deep temporal emargination is present, partially covering the otic chamber. There is a well-developed supraoccipital crest, and the squamosals show a well-developed posterior projection.
The skull is preserved in two individuals of Algorachelus parva studied herein (EY793 and EY797; Figure 2A-C; Figure 3). They are exposed from the matrixin dorsal view (Figure 3A, B). Both are dorsoventrally crushed. However, the skull is interpreted as a relatively wide element, its length being slightly greater than its width. It is wide along its entire length and has a short rostral region, with a well-defined anterior angle. The orbits are dorsolaterally facing. Nasals and an interorbital groove are absent. The prefrontals do not touch the parietals, due to the frontals contact along the midline (Figure 3A, B). The prefrontals are longer than wide. The frontals are slightly wider than long. The parietals do not contact the orbits, but the jugals do. No interparietal or other cranial scutes are recognized. A relatively deep temporal emargination is present, partially covering the otic chamber. There is a well-developed supraoccipital crest, and the squamosals show a well-developed posterior projection.
The lower jaw is identified for EY797, and is only accessible in ventral view (Figure 3C). It is robust. The sutures between the angular and the dentary are identified for both mandibular branches. The anterior angle defined by both branches is slightly obtuse, but the posterior angle between them is slightly acute.
DISCUSSION
Reinterpretation of Shell Characters for Algorachelus parva
Prior to the current study, knowledge of Algorachelus parva was based solely on Haas (1978a, b). The detailed first-hand study of all the specimens of this taxon published so far [i.e., the holotype of ‘Podocnemis’ parva, EY795 (Figure 1A-B); the three paratypes of the species, EY790 (Figure 1F), EY797 (Figure 2A-B, Figure 3B-C), and EY792 (Figure 2D-E); and the holotype of ‘Podocnemis judaea’, EY791 (Figure 1D-E)] allows me to identify several traits incorrectly interpreted by Haas (1978a, 1978b). This is essential for the characterization of the species, as well as for the diagnosis of other species of Algorachelus. Haas (1978a) misinterpreted the morphology and dimensions of the first neural of ‘Podocnemis’ parva considering that this region corresponded to two plates, so that the neural series was interpreted as composed of seven elements instead of six (compare Figure 1A and Figure 2A with figure 1 in Haas, 1978a). The same error was made for the holotype of ‘Podocnemis judaea’, where the neural series is here identified as containing seven, rather than eight plates (compare Figure 1D with figure 1 in Haas, 1978b). Therefore, other characters related to this, such as the number of neurals in contact with the first pair of costals, the position of the longest plate of the neural series, or the identification of the neurals on which the contacts between the first to fourth vertebral scutes were located, were also misinterpreted by Haas (1978a, 1978b). Haas (1978a) considered, in the diagnosis of ‘Podocnemis’ parva, the presence of a very short medial contact between the eighth costals (see figure 1 in Haas, 1978a), which he characterized as restricted to a point. First-hand study of both the holotype of the species (see specimen EY795 in Figure 1A) and the paratypes that preserve the posterior region of the carapace (see EY797 and EY792 in Figure 2A, B, D, E), allows confirmation that this interpretation was incorrect. Notable problems in the interpretation of the posterior costals of the holotype of ‘Podocnemis judaea’ were also identified in the work of Haas (1978b) (compare Figure 1D with figure 1 in Haas, 1978b, corresponding to the specimen EY791). Haas (1978b) incorrectly recognized a posterior extension of the medial region of the sixth pair of costals as not allowing contact between the plates of the seventh pair. Therefore, Haas (1978b) erroneously considered the medial contact of the costal series as only restricted to the long contact between the sixth costals. The current analysis of this specimen (EY791) allowed confirmation that medial contact between the sixth costals was absent, but present between the seventh and eighth costals (Figure 1D). Haas (1978a) also misrepresented the mesoplastra of EY795 as roughly triangular in the diagnosis of ‘Podocnemis’ parva (see figure 2 in Haas, 1978a). The mesoplastra are actually subrounded (Figure 1B), lacking the acute medial angle portrayed by Haas (1978a), especially for the left mesoplastron.
In addition to the erroneous interpretation of the medial region of the sixth and seventh costals of the holotype of ‘Podocnemis judaea’ discussed above, Haas (1978b) described several characters as putatively different between that individual (EY791) and the specimens of ‘Podocnemis’ parva, which allowed him to justify differentiation in the two species. Thus, Haas (1978b) interpreted the holotype of ‘Podocnemis judaea’ to have seven pairs of costals, instead of eight: he did not correctly locate the suture between the last pair of costals and the suprapygal plate. Rather, he characterized that taxon by the presence of a very large suprapygal, which contacted the anterior margin of the last pair of peripherals and more than the medial half of that of the next anterior pair (compare Figure 1D with figure 1 in Haas, 1978b). This led Haas (1978b) to erroneously recognize the medially disconnected scutes of the holotype of ‘Podocnemis judaea’ as unidentified "inguinal elements" (see figure 1 in Haas, 1978b), which were not found in the specimens attributed to ‘Podocnemis’ parva.
Haas (1978b) considered several additional differences between the two Podocnemis species he recognized, but examination of additional specimens from that type locality allows these differences to be considered part of the intraspecific variability of Algorachelus parva. Similar variation is known for the European species Algorachelus peregrina, which is represented by hundreds of shell remains. Thus, the neural series is variable in number for Algorachelus parva and can have a maximum of six (Figure 1A, C; Figure 2A, C, D) or seven plates (Figure 1D). The number of neurals is recognized as variable in the case of Algorachelus peregrina but ranges between five and seven neurals (Pérez-García, 2016, 2018). The medial contact of the posterior costal series in the shells of Algorachelus parva occurs from the sixth to the eighth pairs of costals in some specimens (Figure 1A, C; Figure 2A, C, D), and only between the last two pairs in EY791 (Figure 1D). This contact is also recognized as variable in Algorachelus peregrina, with both configurations recognized, as well as a longer medial contact of the costals, that also involves the fifth pair (Pérez-García, 2016, 2018). The variability in the overlap of the gular scute on the entoplastron in some Algorachelus parva is also consistent with the intraspecific variability documented for Algorachelus peregrina (Pérez-García, 2016, 2018). The remaining characters referred by Haas (1978b) as different between the originally described Podocnemis species from Israel (e.g., shape variation in the entoplastron, carapace, and anterior plastral lobe, along with vertebral scute and plastral lobe proportions) have also been recognized as variable at an intraspecific level within the genus Algorachelus (Pérez-García, 2016, 2018). In fact, other anatomical details considered by Haas (1978a) to be intraspecifically variable in ‘Podocnemis’ parva are similarly so in Algorachelus peregrina (see Pérez-García, 2016, 2018). For example, the length of the medial contact between the epiplastra can be less than three times the entoplastal length (see specimen EY795 in Figure 1B) or greater than one-third of the length of that plate in some specimens (see specimen EY797 in Figure 2B). The medial length of the hyoplastra of the taxon was recognized by Haas (1978a) as slightly lower than that of the hyoplastra, but approximately equal to that of the xiphiplastra. This condition is identified here as variable, not only at a intraspecific level but also for a specimen, comparing its right and left regions (e.g., the medial length of the right hypoplastron is similar to that of the right hyoplastron for the specimen in Figure 2B, but, on the left side, that of the hypoplastron is noticeably higher. That of the hyoplastra is not similar to that of the xiphiplastra in that specimen, neither on the right nor on the left side).
Some additional problems identified here in Haas (1978a, 1978b) can also be corrected. For instance, Haas (1978a) does not depict the contact between the pectoral scute and the left mesoplastron in the holotype of ‘Podocnemis’ parva (see figure 2 in Haas, 1978a), but that scute overlapped the anterior region of the mesoplastron, not only in that specimen (EY795, see Figure 1B) and in all other specimens from Israel in which this area is preserved (EY791, EY797, EY792; see Figure 1E; Figure 2B, E). The anterior edge of the nuchal plate, and the medial region of the first pair of marginal scutes, are reinterpreted as not preserved in the holotype of ‘Podocnemis judaea’ (EY791), despite the representation in Haas (1978b, Fig. 1). However, the anterior nuchal edge was probably located very close to the preserved margin (Figure 1D). The identification of the posterior margin of the fourth vertebral scute on the suprapygal plate in the holotype of ‘Podocnemis judaea’ was due to the aforementioned erroneous interpretation of the suprapygal (see figure 1 in Haas, 1978b). That sulcus is reinterpreted here as located on the last pair of costals (Figure 1D). Although only 10 pairs of peripherals were represented in the line drawing of the holotype of ‘Podocnemis judaea’ published by Haas (1978b; figure 1), he depicted 11 pairs, which is compatible with the condition recognized here for the specimen (Figure 1D). After accounting for numerous errors in the original morphological descriptions of Haas (1978a, b), the presence of a single bothremydid taxon is recognized for the Cenomanian record of ‘Ein Yabrud, with Algorachelus parva as the only representative of this lineage so far known from the fossil record of Israel.
Comparison of Shell of Algorachelus parva with Those of Other Congeners and Proposal of Emended Diagnosis
The first-hand review of all fossils of Algorachelus parva documented by Haas (1978a, 1978b), and the study of new specimens from its type locality, provide an important advance in the information of the shell of this taxon. For the first time, a rigorous comparison with both the type species of the genus (i.e., the Iberian Algorachelus peregrina) and the North American Algorachelus tibert can be made. Preliminarily, and exclusively considering the information published by Haas (1978a, 1978b), Pérez-García (2018) indicated three potential differences between Algorachelus parva and the type species. That relative to the putatively deeper anal notch, with rounded lateral margins (i.e., being U-shaped), for Algorachelus peregrina but not for Algorachelus parva cannot be supported here. The analysis of all material of the Iberian species published by Pérez-García (2016, 2018), including both juvenile and adult individuals, together with the examination of numerous unpublished specimens from its type locality (the Spanish site of Algora) allow the recognition of the relative depth of the anal notch as variable; as well as a more substraight (i.e., V-shaped) to more subrounded (i.e., U-shaped) morphology of its lateral margins, probably linked to both ontogeny and sexual dimorphism (Guerrero and Pérez-García, 2023). This intraspecific variability is compatible with that identified here for Algorachelus parva, some specimens showing substraight anal margins (Figure 2B) but being subrounded in others (Figure 1E), being relatively wider in some of those displaying the latter condition.
Two additional characters tentatively proposed by Pérez-García (2018) to differentiate between Algorachelus parva and Algorachelus peregrina can be confirmed. Based on the study of about 50 shells of Algorachelus peregrina from its type locality (i.e., its holotype, the seven articulated shells which preserve the anterior regions of both the carapace and the plastron analyzed by Pérez-García, 2018; and numerous unpublished shells found in the excavation campaigns carried out in Algora after that publication) permits confirmation that the anterior plastral margin reaches a level almost as far anteriorly as the carapace margin, and sometimes slightly exceeds it. This differs from the condition observed in Algorachelus parva (see Figure 1B, E), where the anterior margin of the plastron does not project as close to the extent of the carapace. The condition recognized in the holotype and only known specimen of the North American Algorachelus tibert is consistent with that in Algorachelus peregrina. The second character condition recognized here as different between Algorachelus parva and Algorachelus peregrina can also be confirmed based on the extensive review of the Algorachelus peregrina shell morphology. Thus, the carapaces of Algorachelus peregrina are subpentagonal, the posterolateral margins being nearly straight; those of Algorachelus parva being subrounded. Although the most posterior margin of the carapace is not preserved for the holotype and only known specimen of Algorachelus tibert, the peripheral series is preserved up to the antero-distal region of the left eleventh plate, allowing for the recognition that this carapace displayed nearly straight posterolateral margins (Joyce el al., 2016).
Additional characteristics are recognized here that further differentiate between species of Algorachelus. For instance, the posterior margin of the first pair of marginals was approximately perpendicular to the midline in Algorachelus peregrina (Pérez-García, 2016, 2018). Therefore, the length of the lateral and medial margins of these subrectangular scutes was subequal, a relatively wide distance being present between the first vertebral scute and the anterior carapace margin. This first pair of marginal scutes overlapped between one third and slightly more than half the anterolateral margin of the nuchal plate in Algorachelus peregrina. An even greater overlap of margins onto the nuchal is known for Algorachelus tibert, in which these marginal scutes were trapezoidal (Joyce et al., 2016). As in Algorachelus peregrina, a wide distance separated the first vertebral from the anterior carapace margin in the North American species. Therefore, the presence of generally subtriangular first pair of marginal scutes (with very short medial margins), with a short overlap onto the antero-lateral edges of the nuchal, and without medial contact more substantial than a single point (i.e., allowing the first vertebral scute to reach the nuchal notch), is exclusive to Algorachelus parva. The vertebral scutes of Algorachelus parva are proportionally wider than those of the other species and Algorachelus parva is the only species where the second vertebral is at least one and a half times wider than long. By comparison, the second vertebral is slightly wider than long in Algorachelus tibert, and between as wide as long to wider than long (generally not much wider than long, and always less than one and a half times it) in Algorachelus peregrina (Joyce et al., 2016). The presence of a nuchal with proportionally equal length and width in Algorachelus tibert differs from the wider nuchals of both Algorachelus parva and Algorachelus peregrina (Pérez-García, 2016, 2018).
A putative new North American bothremydid was recently described for the middle Cenomanian fossil record of Texas, having been attributed to a new genus and species, Pleurochayah appalachius Adrian, Smith, Noto, and Grossman, 2021. The shell elements attributed to it, all of them corresponding to isolated plates, are recognized here as compatible with those of the genus Algorachelus. The anatomical regions that allow the characterization of the different Algorachelus species currently recognized are unknown for the material attributed to the species from Texas.
Discussion of Cranial Characters of Algorachelus parva
Although one of the paratypes of Algorachelus parva preserved the skull (EY797; see Figure 2A-B, Figure 3B-C), Haas (1978a) refrained from describing it due to poor preservation, and the specimen was not figured in detail. No data on the skull of this species has been reported prior to the current study. A previously unpublished specimen of Algorachelus parva also preserving the skull also offers novel cranial insight (EY793; see Figure 2C, Figure 3A). This second skull is recognized as more robust than that of the paratype (EY797) and is associated with a shell that is 15% larger. The lack of knowledge about the morphology of the anal notch of the largest in relation to that of the minor (that is, about the region usually recognized as most relevant for the determination of sexual dimorphs in Pleurodira; see Páez et al., 2013; Sepúlveda-Seguro et al., 2020) does not allow to assess whether they might be two sexual dimorphs or whether the largest might represent a more advanced ontogenetic stage.
Algorachelus tibert is only known by a shell (Joyce et al., 2016). The skull of Algorachelus peregrina is known from a single skull belonging to a paratype (ALG 64; see figure 7 in Pérez-García, 2016). However, that specimen lacks the anterior cranial region, the posterior part of the supraoccipital crest, and the posterior margin of the skull roof, so that these regions can be characterized for the first time here for the genus Algorachelus. Significant differences between the material of Algorachelus peregrina and Algorachelus parva are not apparent in the scarce and incomplete preserved cranial material in both taxa. The new material presented here demonstrates a relatively long supraoccipital crest for the genus, a character that could not be evaluated until now. However, given damage to EY797, which is the only specimen preserving this region, termination of the crest at the posterior margin of the squamosals (typical for other Bothremydina) cannot be confirmed (Gaffney et al., 2006). The relatively wide overall morphology of the skull of Algorachelus parva is shared with most members of Bothremydina, except Araiochelys hirayamai (Figure 3A, B). The apparently relatively large size of the orbits of Algorachelus parva exceeds that of other forms of Bothremydina such as Araiochelys hirayamai, Bothremys spp., and Chedighaii spp., and is more similar to that of Rosasia soutoi (Antunes and Broin, 1988; Gaffney et al., 2006). However, considering the crushing of both of skulls of Algorachelus parva, this should be taken with caution. Slightly wider than long frontals and well-developed temporal emarginations, are shared with all other members of Bothremydina (Gaffney et al., 2006; Ferreira et al., 2018).
The holotype of the poorly known North American Cenomanian bothremydid Pleurochayah appalachius corresponds to a partial skull (see figure 2 in Adrian et al., 2021). This skull is not as wide as that documented for Algorachelus peregrina, but that condition is closer to that recognized in Algorachelus parva (the presence of intraspecific variability cannot be ruled out, as has been documented for some members of Pelomedusoides, including some bothremydids; see Martín-Jiménez and Pérez-García, 2022). The crushing of the only skull known for Pleurochayah appalachius does not allow an adequate comparison of the degree of opening of the incisura columella auris with that known for Algorachelus (being exclusively availaible for the paratype of Algorachelus peregrina), contrary to Adrian et al. (2021). This crushing, like those of the Algorachelus parva skulls, also do not allow to evaluate in which taxon the orbits are more dorsally directed. The skulls of Algorachelus parva show that the length of the contact between the postorbital and the frontal is not very different in this Israeli taxon from that recognized for Pleurochayah appalachius. The preservation of the skull of Pleurochayah appalachius does not allow us to confirm that, unlike what is documented here for the genus Algorachelus, its supraoccipital crest was short. The two additional cranial characters indicated by Adrian et al. (2021) as different between Pleurochayah appalachius and Algorachelus parva (i.e., the posterior elongation of the fossa orbitalis, considering the position of the septum orbitale; and the distance between the foramen stapediotemporale and the foramen nervi trigemini), cannot be adequately evaluated taking into account the relatively scarce and incomplete cranial material currently available for the genus Algorachelus, and the deformation of the holotype of the taxon from Texas, corresponding to the only skull attributable to it. Therefore, the potential identification of the material attributed to Pleurochayah appalachius as corresponding to the Algorachelus lineage should be evaluated through the future discovery and study of well-preserved Algorachelus skulls.
Algorachelus parva specimen EY797 has the only lower jaw known so far for this genus (Figure 3C). Although it is only exposed from the matrix in ventral view, it is notably more robust than that of Araiochelys hirayamai, displaying a condition similar to that in Bothremys spp. and Chedighaii spp.
CONCLUSIONS
Pelomedusoides is one of the two groups of pleurodiran turtles that are represented by modern species. The most successful of extinct lineages was Bothremydidae, especially during the Late Cretaceous and Eocene. Although Pelomedusoides had a Gondwanan origin, several representatives reached Laurasian regions through a series of diachronic dispersions. Among these dispersal events, those of Bothremydidae stands out for commencing in the earliest Late Cretaceous or, probably, at the end of the Early Cretaceous. The relatively recently discovered bothremydid genus Algorachelus has been documented in the Cenomanian of Israel (Algorachelus parva), the Iberian Peninsula (Algorachelus peregrina), and North America (Algorachelus tibert). The current study updates the taxonomic status of Algorachelus parva, which was erroneously described in the 1970s as belonging to two non-bothremydid species. It was originally attributed to the genus of Podocnemididae Podocnemis, which is now understood as exclusive to South America. The taxonomic revision conducted here was accompanied by descriptions that correct multiple errors in the original accounts and allows higher resolution comparison between Algorachelus spp. Contrary to interpretations made in the 1970s, all material of Bothremydidae from the ‘Ein Yabrud limestone quarry is attributed to the single species Algorachelus parva. Intraspecific variability is recognized and described in detail for several traits. The skull of Algorachelus parva is described for the first time and is represented by two specimens, expanding the knowledge of this poorly known region in the genus Algorachelus. The lower jaw of one skull is described as the only such element known for the genus. The current study corrects and supplements our understanding of Algorachelus parva and allows detailed comparisons with widely distributed congenerics, improving our understanding of bothremydid evolution and biostratigraphy.
ACKNOWLEDGMENTS
This research has been developed in the context of a SYNTHESYS project (IL-TAF-2568), under the EU FPVII framework. Author thanks the support of the Viceconsejería de Cultura of the Consejería de Educación, Cultura y Deportes of Castilla-La Mancha (Spain), which has provided funding for the discovery and preparation of specimens of Algorachelus peregrina from its type locality (SBPLY/22/180801/000020, SBPLY/23/180801/000046, and SBPLY/24/180801/000043); R. Rabinovich and G. Beiner (HUJ-PAL), for the access to the specimens from Israel studied here; A. Guerrero (UNED), for discussions on the variability of Algorachelus; and the editor M. Chikh Younes, and the reviewers E.A. Cadena and B. Adrian for comments and suggestions.
Jurisdiction Statement
The identification of the site of Ein Yabrud as part of Israel is purely geographical and has no intended political implications. This study is a scientific work and, as such, it is independent of the Israeli-Palestinian conflict, in which the editorial board of Palaeontologia Electronica takes a strictly neutral position.
REFERENCES
Adrian, B., Smith, H.F., Noto, C.R., and Grossman, A. 2021. An early bothremydid from the Arlington Archosaur Site of Texas. Scientific Reports, 11:9555.
https://doi.org/10.1038/s41598-021-88905-1
Antunes, M.T. and Broin, F. de. 1988. Le Crétacé terminal de Beira Litoral, Portugal: remarques stratigraphiques et écologiques; étude complémentaire de Rosasia soutoi (Chelonii, Bothremydidae). Ciências da Terra, 9:153–200.
Batsch, G.C. 1788. Versuch einer Anleitung, zur Kenntniss und Geschichte der Thiere und Mineralien. Jena, Akademische Buchhandlung, 1:1–528.
Baur, G. 1891. Notes on some little known American fossil tortoises. Proceedings of the Academy of Natural Sciences of Philadelphia, 43:411–430.
Broin, F. de 1988. Les tortues et le Gondwana. Examen des rapports entre le fractionnement du Gondawana et la dispersion géographique des tortues pleurodires à partir du Crétacé. Studia Geologica Salmanticensia, Studia Palaeocheloniologica, 2:103–142.
Cadena, E.-A. and Vanegas, R.D. 2023. A new fossil turtle ends the controversy on the occurrence of the extant genus Podocnemis Wagler, 1830 at the Miocene fauna of La Venta, Colombia. Geodiversitas, 45:127–138. https://doi.org/10.5252/geodiversitas2023v45a3
Cawley, J.J. and Kriwet, J. 2018. A new pycnodont fish, Scalacurvichthys naishi gen. et sp. nov., from the Late Cretaceous of Israel. Journal of Systematic Palaeontology, 16:659–673.
https://doi.org/10.1080/14772019.2017.1330772
Cope, E.D. 1864. On the limits and relations of the Raniformes. Publications of the Academy of Natural Sciences of Philadelphia, 16:181–183.
Cope, E.D. 1868. On the origin of genera. Publications of the Academy of Natural Sciences of Philadelphia, 20:242–300.
Ferreira, G.S., Bronzati, M., Langer, M.C., and Sterli, J. 2018. Phylogeny, biogeography and diversification patterns of side-necked turtles (Testudines: Pleurodira). Royal Society Open Science, 5:171773.
https://doi.org/10.1098/rsos.171773
Ferreira, G.S., Rincón, A.D., Solórzano, A., and Langer, M.C. 2015. The last marine pelomedusoids (Testudines: Pleurodira): a new species of Bairdemys and the paleoecology of Stereogenyina. PeerJ, 3:e1063.
https://doi.org/10.7717/peerj.1063
Gaffney, E.S., Tong, H., and Meylan, P.A. 2006. Evolution of the side-necked turtles: the families Bothremydidae, Euraxemydidae, and Araripemydidae. Bulletin of the American Museum of Natural History, 300:1–700.
http://hdl.handle.net/2246/5824
Gaffney, E.S., Meylan, P.A., Wood, R.G., Simons, E., and Almeida Campos, D. 2011. Evolution of the side-necked turtles: the family Podocnemididae. Bulletin of the American Museum of Natural History, 350:1–237.
https://doi.org/10.1206/350.1
Georgalis, G.L., Velitzelos, E., Velitzelos, D., and Kear, B.P. 2013. Nostimochelone lampra gen. et sp. nov., an enigmatic new podocnemidoidean turtle from the lower Miocene of northern Greece, p. 277–287. In Brinkman, D., Holroyd, P., and Gardner, J. (eds.), Morphology and evolution of turtles: papers in honor of Eugene S Gaffney. Springer, Dordrecht, The Netherlands.
Guerrero, A. and Pérez-García, A. 2023. A quantitative approach to the interindividual variability of the shell of the Spanish Cenomanian bothremydid turtle Algorachelus peregrina. Palaeontological Publications, 4:158.
Haas, G. 1978a. A Cretaceous pleurodire turtle from the surroundings of Jerusalem. Israel Journal of Zoology, 27:20–33.
Haas, G. 1978b. A new turtle of the genus Podocnemis from the lower Cenomanian of ‘Ein Yabrud’. Israel Journal of Zoology, 27:169–175.
Joyce, W.G., Lyson, T.R., and Kirkland, J.I. 2016. An early bothremydid (Testudines, Pleurodira) from the Late Cretaceous (Cenomanian) of Utah, North America. PeerJ, 4:e2502.
https://doi.org/10.7717/peerj.2502
Khalloufi, B., Zaragüeta-Bagils, R., and Leliévre, H. 2010. Rhombichthys intoccabilis, gen. et sp. nov. (Ellimmichthyiformes, Clupeomorpha, Teleostei), from the Cenomanian (Upper Cretaceous) of Ein Yabrud, Middle East: anatomical description and phylogenetic implications. Journal of Vertebrate Paleontology, 30:57–67.
https://doi.org/10.1080/02724630903409089
Lapparent de Broin, F. de. 2001. The European turtle fauna from the Triassic to the Present. Dumerilia, 4:155–216.
Lapparent de Broin, F. de. and Werner, C. 1998. New Late Cretaceous turtles from the Western Desert, Egypt. Annales de Paléontologie, 84:131–214.
Martín-Jiménez, M. and Pérez-García, A. 2022. The neuroanatomy of the bothremydid pleurodiran turtle Galianemys, from the Late Cretaceous (Cenomanian) of Morocco. The Anatomical Record, 306:1377–1395.
https://doi.org/10.1002/ar.25072
Páez, V.P., Restrepo, A., Vargas Ramírez, M., Bock, B.C., and Gallego García, N. 2013. Podocnemis lewyana (Duméril 1852). Tortuga del río Magdalena, tortuga de río. Asociación Colombiana de Herpetología, 1:1–5.
Pérez-García, A. 2016. A new turtle taxon (Podocnemidoidea, Bothremydidae) reveals the oldest known dispersal event of the crown Pleurodira from Gondwana to Laurasia. Journal of Systematic Palaeontology, 15:709–731.
https://doi.org/10.1080/14772019.2016.1228549
Pérez-García, A. 2017. The Iberian fossil record of turtles: an update. Journal of Iberian Geology, 43:155–191. https://doi.org/10.1007/s41513-017-0016-4
Pérez-García, A. 2018. New information on the Cenomanian bothremydid turtle Algorachelus based on new, well-preserved material from Spain. Fossil Record, 21:119–135.
https://doi.org/10.5194/fr-21-119-2018
Pérez-García, A. 2020. First evidence of a bothremydid turtle (crown Pleurodira) in the middle Cretaceous of Castile and Leon (Spain). Journal of Iberian Geology, 46:363–368.
https://doi.org/10.1007/s41513-020-00146-9
Pérez-García, A. 2023. The ornamented shell of a new bothremydid turtle from the Uppermost Cretaceous of Niger. Diversity, 15:375.
https://doi.org/10.3390/d15030375
Pérez-García, A. and Rubio, C. 2024. A turtle from northeastern Spain reveals that the dispersal of Pelomedusoides from Gondwana to Laurasia probably occurred in the Early Cretaceous. Cretaceous Research, 162:105938.
https://doi.org/10.1016/j.cretres.2024.105938
Pérez-García, A., Lapparent de Broin, F. de, and Murelaga, X. 2017a. The Erymnochelys group of turtles (Pleurodira, Podocnemididae) in the Eocene of Europe: new taxa and paleobiogeographical implications. Palaeontologia Electronica, 20.1.14A:1–28.
https://doi.org/10.26879/687
Pérez-García, A., Antunes, M.T., Barroso-Barcenilla, F., Callapez, P.M., Segura, M., Soares, A.F., and Torices, A., 2017b. A bothremydid from the middle Cenomanian of Portugal identified as one of the oldest pleurodiran turtles in Laurasia. Cretaceous Research, 78:61–70.
https://doi.org/10.1016/j.cretres.2017.05.031
Pérez-García, A., Bardet, N., Fregenal-Martínez, M.A., Martín-Jiménez, M., Mocho, P., Narváez, I., Torices, A., Vullo, R., and Ortega, F. 2020. Cenomanian vertebrates from Algora (central Spain): New data on the establishment of the European Upper Cretaceous continental faunas. Cretaceous Research, 115:104566.
https://doi.org/10.1016/j.cretres.2020.104566
Rabi, M., Tong, H., and Botfalvai, G. 2012. A new species of the side-necked turtle Foxemys (Pelomedusoides: Bothremydidae) from the Late Cretaceous of Hungary and the historical biogeography of the Bothremydini. Geological Magazine, 149:662–674.
https://doi.org/10.1017/S0016756811000756
Sepúlveda-Seguro, A.M., Páez, V.P., Gómez-Rincón, M.T., Morales-Betancourt, M.A., and Lasso, C.A. 2020. Evaluación de las características de las poblaciones y el uso de los hábitats de Podocnemis vogli (Müller 1935) durante una temporada reproductiva en la reserva natural privada de Bojonawi (Departamento de Vichada, Colombia). Revista Latinoamericana de Herpetología, 3:83–94.
https://doi.org/10.22201/fc.25942158e.2020.2.139
Zalmout, I.S., Mustafa, H.A., and Wilson, J.A. 2005. Karkaemys arabicus, a new side-necked turtle. (Pleurodira, Bothremydidae) from the Upper Cretaceous Wadi Umm Ghudran Formation of Karak, Jordan. Contributions from the Museum of Paleontology of the University of Michigan, 31:155–177.

 A Review of Handbook of Paleoichthyology Volume 8a: Actinopterygii I, Palaeoniscimorpha, Stem Neopterygii, Chondrostei
A Review of Handbook of Paleoichthyology Volume 8a: Actinopterygii I, Palaeoniscimorpha, Stem Neopterygii, Chondrostei Palaeontologia Electronica among the most influential palaeontological journals
Palaeontologia Electronica among the most influential palaeontological journals