

 New genus and species of Lamprosomatinae (Coleoptera: Chrysomelidae) from Eocene Baltic amber
New genus and species of Lamprosomatinae (Coleoptera: Chrysomelidae) from Eocene Baltic amber
Article number: 22.2.33
https://doi.org/10.26879/905
Copyright Paleontological Society, June 2019
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 11 July 2018. Acceptance: 15 May 2019.
{flike id=2584}
ABSTRACT
Fossil records of Lamprosomatinae (Coleoptera: Chrysomelidae) are scarce. Till now only three findings of this subfamily were reported and all originated from Eocene Baltic amber. Based on well-preserved specimen, Damzenius anitae gen. et sp. nov. is described and illustrated from this Lagerstätte. A key to the known extinct species of Lamprosomatinae is provided. Assumed minimum age of Lamprosomatinae is dated as mid-Eocene to Upper Eocene (47.8-33.9 Ma). The diverse fossil Lamprosomatinae representatives provide strong evidence for the presence of thermophilic, humid and mixed forest, and indicative of warm temperate, humid, equable climate with reduced thermal seasonality in the Eocene Europe.
Andris Bukejs. Institute of Life Sciences and Technologies, Daugavpils University, Vienības 13, Daugavpils, Latvia. carabidae@inbox.lv
Key words: leaf-beetles; Lamprosomatini; new taxa; Damzenius anitae; Paleogene; fossil resin
Bukejs, Andris. 2019. New genus and species of Lamprosomatinae (Coleoptera: Chrysomelidae) from Eocene Baltic amber. Palaeontologia Electronica 22.2.33A 1-10. https://doi.org/10.26879/905
palaeo-electronica.org/content/2019/2584-new-fossil-lamprosomatinae
Copyright: June 2019 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/C0985A7B-8189-44F2-9642-64B8CF1C5110
INTRODUCTION
Lamprosomatinae Lacordaire, 1848 (Coleoptera: Chrysomelidae) contains about 250 extant species belonging to 14 genera and four tribes: Cachiporrini Chamorro et Konstantinov, 2011 (1 genus), Lamprosomatini Larcordaire, 1848 (10 genera), Neochlamysini Monrós, 1958 (2 genera), and Sphaerocharini Clavareau, 1913 (1 genus) (Seeno and Wilcox, 1982; Chamorro and Konstantinov, 2011; Chamorro, 2014), and its distribution is mostly restricted to Neotropic (Jolivet et al., 2012). The Palaearctic fauna of this subfamily includes three genera and 29 species belonging to the tribe Lamprosomatini, most of them distributed in Asian part of the region (Konstantinov, 2010), while a single species, Oomorphus concolor (Sturm, 1807), is known from Recent Europe (Warchałowski, 2003).
Lamprosomatines mostly feed on woody plants. Documented hosts include Bombacaceae, Combretaceae, Fabaceae, Melastomataceae, and Myrtaceae (Jolivet, 1978; Caxambú and Almeida, 2003; Chamorro, 2014). Larvae build portable fecal cases and actively feed on the bark of live trees (Monrós, 1956; Erber, 1988; Caxambú and Almeida, 1999; Chamorro, 2014; Chaboo et al., 2016).
Fossils of Lamprosomatinae are scarce (Bukejs and Nadein, 2015). Till now only three fossils were reported and all belong to Lamprosomatini and originated from Eocene Baltic amber: Succinoomorphus warchalowskii Bukejs et Nadein, 2015, Archelamprosomius balticus Bukejs et Nadein, 2015, and A. kirejtshuki Bukejs et Nadein, 2015. In the present paper, a new extinct genus and species belonging to Lamprosomatinae is described from this fossil resin.
MATERIAL AND METHODS
The material examined is deposited in the collection of Andris Bukejs (Daugavpils, Latvia) [ACAB] maintained at Institute of Life Sciences and Technologies, Daugavpils University (Daugavpils, Latvia). The amber piece was polished by hand, allowing improved views of the included specimen, and was not subjected to any supplemental fixation.
Observations of this specimen were made using a Nikon SMZ 745T stereomicroscope. The photographs were taken using a Canon 70D camera with a macro lens (Canon MPE-65 mm). Extended depth of field at high magnifications was achieved by combining multiple images from a range of focal planes using Helicon Focus software. Measurements were taken using an ocular micrometer (expressed in millimeters). The following references were used for the comparison with extant taxa: Monrós (1956, 1958), Medvedev (1968), and Chamorro and Konstantinov (2011).
Baltic amber occurs in the “blau Erde” (“blue earth”), which can be found throughout northern Europe (Denmark, Sweden, Germany, Poland, and Lithuania), approximately 45 m below the surface, and in many places this unit is approximately 5 m below sea level (Engel, 2001). The greatest amber concentration, and from where most material originates, occurs on the Sambia peninsula (Samland), Kaliningrad Region, Russia. Exposed deposits can erode, and amber is washed up on shores not only of the Baltic countries, but as far away as the eastern coasts of England (Engel, 2001). Baltic amber is usually dated to the Upper Eocene (Priabonian, 37.2-33.9 Ma) (Aleksandrova and Zaporozhets, 2008; Perkovsky et al., 2007), although there are other estimates of the age of Baltic amber that extend as far back as the mid-Eocene (Lutetian, 47.8-41.3 Ma) (Ritzkowski, 1997; Szwedo and Sontag, 2013). According to Turkin (1997), Baltic amber was produced by Pinus succinifera (Conw.) Schub., which together with oak in the Eocene, dominated the humid mixed forest cover of Northern and Central Europe. More recent work on the chemical composition of Baltic amber has also suggested that trees within the family Araucariaceae or Sciadopityaceae might be candidates for the production of this amber deposit (Langenheim, 2003; Wolfe et al., 2009; Lambert et al., 2015).
SYSTEMATIC PALAEONTOLOGY
Order COLEOPTERA Linnaeus, 1758
Family CHRYSOMELIDAE Latreille, 1802
Subfamily LAMPROSOMATINAE Lacordaire, 1848
Tribe LAMPROSOMATINI Lacordaire, 1848
Remarks. The specimen considered here was assigned to the tribe Lamprosomatini on the basis of a combination of characters defined by Chamorro and Konstantinov (2011) and also by Monrós (1956, 1958): (1) abdominal ventrites with fine transverse folding on their borders; (2) abdominal ventrite 5 not excised in shape of arc; (3) pygidium completely covered by elytra; (4) scutellum acutely triangular; (5) elytral punctation arranged in regular striae; and (6) pronotum and elytra hairless.
DAMZENIUS gen. nov.
zoobank.org/04E84ADE-B752-4D09-B88F-F9209CAF9167
Type species. Damzenius anitae sp. nov., here designated
Included species. The new genus includes the type species only.
Differential diagnosis. Damzenius gen. nov. differs from extinct Succinoomorphus in possessing tarsal claws appendiculate with denticle basally; shape of antennomeres 9-10; epipleura with strong, convex projection in anterior one-third of elytral length; ventrolateral side of meso- and metatibiae without distinct longitudinal groove; pronotal lateral margins with bordering; body more convex and oval; and from Archelamprosomius in possessing tarsal claws with denticle basally; frons and vertex with longitudinal medial groove; head with groove at inner and upper margins of eye; compound eyes with inner margin slightly emarginated; shape of antennomeres 9-10; prohypomera distinctly concave on whole surface; metafemora extending to abdominal ventrite 2; femora and tibiae strongly flattened and widened. See also details provided in key (Appendix 1).
This new fossil genus resembles extant Oomorphoides Monrós, 1956, based on appendiculate tarsal claws, possession of a groove at inner and upper margins of eye, frons and vertex with longitudinal medial groove, strongly projecting basal third of the epipleura, but differs from it in slightly emarginate inner margin of eye (angularly notched in Oomorphoides), posterior pronotal angles not extending posterad beyond elytral base (extending posterad beyond elytral base in Oomorphoides), and elytral punctation very fine and inconspicuous, arranged in indistinct rows (small and distinct, forming distinct rows in Oomorphoides). Additionally Damzenius gen. nov differs from other extant genera of Lamprosomatini in the combination of the following characters: (1) tarsal claws appendiculate with denticle basally; (2) compound eyes with inner margin slightly emarginated; (3) head with groove at the inner and upper margins of eye; (4) frons and vertex with longitudinal medial groove; (5) tarsomere 4 projecting from tarsomere 3 to half of its length; (6) pronotal lateral margins slightly concave in lateral view, with narrow bordering; and (7) larger body size.
Etymology. The generic epithet is a patronym dedicated to Mr. Jonas Damzen (Vilnius, Lithuania), the collector of the amber piece. Gender masculine.
Note. The new genus is monotypic, represented by the type species only. Therefore, the generic description considerably overlaps that of the species.
Damzenius anitae sp. nov.
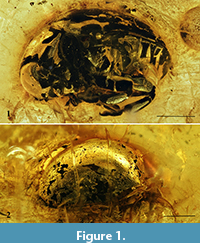
Figure 1.1-2, Figure 2.1-2
zoobank.org/6638E6E1-E8CD-4C90-AD67-965C07A75555
Type material. Holotype: collection number “048” [ACAB], adult, sex unknown. Rather complete beetle (apical antennomeres of left antenna missing) is included in small, transparent yellow amber piece with dimensions 21×10×4 mm. Syninclusions consist of one small Diptera specimen, one stellate Fagaceae trichome, and numerous small gas vesicles.
 Type strata. Baltic amber, mid-Eocene to Upper Eocene.
Type strata. Baltic amber, mid-Eocene to Upper Eocene.
Type locality. Baltic Sea coast, Yantarny settlement (formerly Palmnicken), Kaliningrad region, Russia.
Etymology. The specific epithet is a patronym dedicated to my mother, Anita Bukeja (Daugavpils, Latvia).
Description. Body length 4.1 mm, maximum width about 2.8 mm; widely oval, convex dorsally and flattened ventrally; unicolorous black; glabrous, prosternal process with moderately dense, fine, semierect setation, abdominal ventrites with very fine, inconspicuous pubescence (more distinct on ventrite 5).
Head slightly convex, evenly covered with fine, sparse punctation and very fine microreticulation (more distinct on frons). Frons and vertex with longitudinal medial groove. Compound eyes oval, small (vertical diameter about one-third as great as length of pronotal lateral margin), convex, with distinct facets, without interfacetal setae, inner margin apparently slightly roundly emarginated; with groove at inner and upper margins of eye; distance between eyes about 1.5× as great as vertical diameter of one eye. Clypeus transverse, convex, clearly delimited from frons. Antennae poorly visible because of beetle location in amber piece, filiform, moderately long, apparently slightly reach beyond posterior margin of prosternal process; scape large, 2.1× as long as wide, apparently with triangular projection dorsally, pedicel subcylindrical, shorter and narrower than scape, antennomeres 3-5 cylindrical, elongate, slender, antennomeres 6-8 nearly transverse, slightly dilated apically, antennomeres 9-10 distinctly dilated apically, asymmetrical (slightly serrate), antennomere 10 about 0.6× as long as ultimate antennomere, antennomere 11 elongated oval with slightly narrowed apex, about 1.6× as long as preceding antennomere, antennomeres 9-11 bearing few erect setae; antennal insertions widely separated.
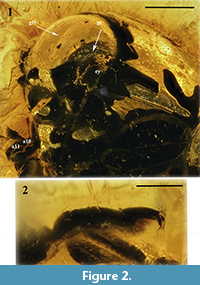 Pronotum strongly transverse, about 2× as wide as long, widest at base; anterior margin arcuate in frontal view; lateral margins slightly concave in lateral view, with narrow bordering, posterior margin bisinuate; anterior angles nearly right, bent ventrally and not visible from above, posterior angles right, not extending posterad beyond elytral base. Pronotal punctation fine and moderately dense. Prohypomera distinctly concave on whole surface, with small, shallow punctures and microreticulation. Intercoxal prosternal process large, distinctly longer than wide, lateral margins constricted medially, anterior margin almost straight, slightly swollen; densely covered with fine, deep punctures.
Pronotum strongly transverse, about 2× as wide as long, widest at base; anterior margin arcuate in frontal view; lateral margins slightly concave in lateral view, with narrow bordering, posterior margin bisinuate; anterior angles nearly right, bent ventrally and not visible from above, posterior angles right, not extending posterad beyond elytral base. Pronotal punctation fine and moderately dense. Prohypomera distinctly concave on whole surface, with small, shallow punctures and microreticulation. Intercoxal prosternal process large, distinctly longer than wide, lateral margins constricted medially, anterior margin almost straight, slightly swollen; densely covered with fine, deep punctures.
Scutellar shield minute, acutely triangular.
Elytra smooth, strongly convex, anterior margin slightly concave, elytral suture smooth; punctation very fine and inconspicuous, more distinct on disc in anterior portion, arranged in indistinct rows. Epipleura horizontal (not visible in lateral view), narrow, widest anteriorly and gradually narrowing posteriorly, with strong, convex projection in anterior one-third of elytral length. Metaventrite slightly elevated posteriorly and medially; with microteticulation and sparse, fine punctures (more distinct in anterior portion). Metacoxae transverse, elongate.
Legs robust, short; femora and tibiae strongly flattened, covered with fine, sparse punctures, microreticulation, and fine, semierect setae. Femora clavate, metafemora reaching abdominal ventrite 3. Tibiae with comb of erect setae apically, protibia nearly as long as profemora, meso- and metafemora about 0.8× as long as femora respectively, ventrolateral side of meso- and metatibiae without distinct longitudinal groove. Tarsomeres 1-3 of all legs distinctly dilated apically, transverse, tarsomere 3 deeply bilobed, tarsomere 4 subcylindrical, projecting from tarsomere 3 to about half of its length; protarsi: tarsomere 1 about 1.3× as wide as long, tarsomere 2 1.7× as wide as long, tarsomere 3 1.4× as wide as long, tarsomere 4 0.3× as wide as long. Tarsal claws free, appendiculate with denticle basally, narrowly separated.
Abdomen with five visible ventrites; covered with fine, sparse punctures and microreticulation; lateral borders of ventrites fine folding; ventrite 5 simple, without fovea or impression, not excised in shape of arc, with posterior margin widely rounded. Relative length ratios of ventrites 1-5 equal to 20:10:7:7:13 (medially). Pygidium completely covered by elytra.
DISCUSSION
Chrysomelidae are well presented in Baltic amber. Currently, 26 species belonging to seven subfamilies were described from this Lagerstätte: Galerucinae Latreille, 1802 (10 species in seven genera), Cassidinae Gyllenhal, 1813 (five species in five genera), Lamprosomatinae Lacordaire, 1848 (four species in three genera), Eumolpinae Hope, 1840 (four species in three genera), Bruchinae Latreille, 1802 (one species), Criocerinae Latreille, 1804 (one species), and Cryptocephalinae Gyllenhal, 1813 (one species) (Bukejs and Konstantinov, 2013; Bukejs and Nadein, 2013, 2014, 2015; Moseyko and Kirejtshuk, 2013; Biondi, 2014; Bukejs, 2014; Bukejs and Bezděk, 2014; Bukejs and Chamorro, 2015; Bukejs and Moseyko, 2015; Bukejs et al., 2015; Legalov, 2016; Bukejs and Schmitt, 2016; Nadein et al., 2016; and Bukejs and Alekseev, 2019). Apparently, lamprosomatines were diverse and abundant in the Eocene Baltic forest. Extant representatives of this subfamily occur mostly in subtropical and tropical regions, and associated with woody plants (Jolivet, 1978; Caxambú and Almeida, 2003; Jolivet et al., 2012; Chamorro, 2014; etc.). The diverse fossil Lamprosomatinae representatives provide strong evidence for the presence of thermophilic, humid and mixed forest (Alekseev and Alekseev, 2016), and indicative of warm temperate, humid, equable climate with reduced thermal seasonality (Alekseev, 2017) in the Eocene Europe.
All known lamprosomatine fossils (Succinoomorphus warchalowskii, Archelamprosomius balticus, A. kirejtshuki, and Damzenius anitae gen. et sp. nov.) are coming from Baltic amber. Based on this data, assumed minimum age of Lamprosomatinae is dated as mid-Eocene to Upper Eocene (47.8-33.9 Ma).
ACKNOWLEDGMENTS
The author is sincerely grateful to Mr. J. Damzen (Vilnius, Lithuania) for donation of interesting fossil material and permission to use his photographs, and to Dr. R. McKellar (Royal Saskatchewan Museum, Regina, Saskatchewan, Canada) for linguistic suggestions on an early version of the manuscript.
REFERENCES
Aleksandrova, G.N. and Zaporozhets, N.I. 2008. Palynological characteristic of the Upper Cretaceous and Paleogene sediments of the West of the Sambian peninsula (the Kaliningrad Region), Part 2. Stratigraphy and Geological Correlation, 16(5):75-86. https://doi.org/10.1134/S0869593808050067
Alekseev, V.I. 2017. Coleoptera from the middle-upper Eocene European ambers: Generic composition, zoogeography and climatic implications. Zootaxa, 4290:401-443. https://doi.org/10.11646/zootaxa.4290.3.1
Alekseev, V.I. and Alekseev, P.I. 2016. New approaches for reconstruction of the ecosystem of an Eocene amber forest. Biology Bulletin, 43(1):75-86. https://doi.org/10.1134/S1062359016010027
Biondi, M. 2014. Paolaltica eocenica new genus and new species of flea beetle (Coleoptera: Chrysomelidae: Galerucinae: Alticini) from Baltic amber. Zootaxa, 3852(4):496-500. https://doi.org/10.11646/zootaxa.3852.4.7
Bukejs, A. 2014. A new species of the genus Crepidodera Chevrolat (Coleoptera: Chrysomelidae) from Baltic amber. Zootaxa, 3815(2):286-290. https://doi.org/10.11646/zootaxa.3815.2.8
Bukejs, A. and Alekseev, V.I. 2019. Review of fossil Chrysomelidae (Insecta: Coleoptera) in Eocene Baltic amber, p. 16-18. In Book of Abstracts of “10th International Conference on Biodiversity Research”. Daugavpils University Academic Press “Saule”, Daugavpils.
Bukejs, A. and Bezděk, J. 2014. Calomicrus eocenicus sp. nov. (Coleoptera: Chrysomelidae: Galerucinae) from Baltic amber. Baltic Journal of Coleopterology, 14(1):73-75.
Bukejs, A., Biondi, M., and Alekseev, V.I. 2015. Sucinolivolia torpida—a new genus and species of flea-beetles (Coleoptera: Chrysomelidae: Galerucinae) from Baltic amber. Zootaxa, 4058(1):119-126. https://doi.org/10.11646/zootaxa.4058.1.7
Bukejs, A. and Chamorro, M.L. 2015. Two new fossil species of Cryptocephalus Geoffroy (Coleoptera: Chrysomelidae) from Baltic and Dominican amber. Proceedings of the Entomological Society of Washington, 117(2):116-125. https://doi.org/10.4289/0013-8797.117.2.116
Bukejs, A. and Konstantinov, A.S. 2013. New genus of flea beetle (Coleoptera: Chrysomelidae: Galerucinae: Alticini) from the Upper Eocene Baltic amber. Insecta Mundi, 0306:1-5.
Bukejs, A. and Moseyko, A.G. 2015. Two new species of the genus Taphioporus Moseyko et Kirejtshuk (Chrysomelidae: Eumolpinae) from Baltic amber. Zootaxa, 4032(4):395-406. https://doi.org/10.11646/zootaxa.4032.4.4
Bukejs, A. and Nadein, K. 2013. A second species of Psyllototus (Coleoptera: Chrysomelidae: Galerucinae: Alticini) from the Upper Eocene Baltic amber. Zootaxa, 3609(5):456-470. https://doi.org/10.11646/zootaxa.3609.5.2
Bukejs, A. and Nadein, K. 2014. Psyllototus groehni sp. nov. (Coleoptera: Chrysomelidae), a new species of the palaeoendemic genus from Baltic amber. Baltic Journal of Coleopterology, 14(2):163-166.
Bukejs, A. and Nadein, K. 2015. First fossil Lamprosomatinae leaf beetle (Coleoptera: Chrysomelidae) with descriptions of new genera and species from Baltic amber. Zootaxa, 3931(1):127-139. https://doi.org/10.11646/zootaxa.3931.1.9
Bukejs, A. and Schmitt, M. 2016. Lilioceris groehni sp. n.: the first authentic species of Criocerinae (Coleoptera, Chrysomelidae) from Baltic amber. ZooKeys, 618:67-77. https://doi.org/10.3897/zookeys.618.10085
Caxambú, M.G. and Almeida, L.M. 1999. Descrição dos estágios imaturos e redescrição de Lamprosoma azureum Germar (Chrysomelidae, Lamprosomatinae). Revista Brasileira de Zoologia, 16(Supl. 1):243-256. https://doi.org/10.1590/S0101-81751999000500017
Caxambú, M.G. and Almeida, L.M. 2003. Lamprosoma W. Kirby (Coleoptera, Chrysomelidae): descrição de nova espécie, redescrições e chave para algumas espécies sul americanas. Revista Brasileira de Zoologia, 20:329-337. https://doi.org/10.1590/S0101-81752003000200025
Chaboo, C.S., Chamorro, M.L., and Schöller, M. 2016. Catalogue of known immature stages of camptosomate leaf beetles (Coleoptera, Chrysomelidae, Cryptocephalinae and Lamprosomatinae). Proceedings of the Entomological Society of Washington, 118(2):150-217. https://doi.org/10.4289/0013-8797.118.1.150
Chamorro, M.L. 2014. 2.7.4 Lamprosomatinae Lacordaire, 1848, pp. 77-81. In Leschen, R.A.B. and Beutel, R.G. (eds.), Handbook of Zoology Arthropoda: Insecta: Coleoptera Volume 3: Morphology and Systematics (Phytophaga). De Gruyter, Berlin.
Chamorro, M.L. and Konstantinov, A.S. 2011. Cachiporrini, a remarkable new tribe of Lamprosomatinae (Coleoptera, Chrysomelidae) from South America. Zookeys, 78:43-59. https://doi.org/10.3897/zookeys.78.980
Clavareau, C.H. 1913. Pars 53: Chrysomelidae: Megascelinae, Megalopodinae, Clytrinae, Cryptocephalinae, Chlamydinae, Lamprosomatinae, p. 1-278. In Schenkling, S. (ed.), Coleopterorum Catalogus. W. Junk, Berlin.
Engel, M.S. 2001. A monograph of the Baltic amber bees and evolution of the Apoidea (Hymenoptera). Bulletin of the American Museum of Natural History, 259:1-192. https://doi.org/10.1206/0003-0090(2001)259<0001:AMOTBA>2.0.CO;2
Erber, D. 1988. Biology of Camptostomata Clytrinae-Cryptocephalinae-Chlamisinae-Lamprostomatinae, p. 513-552. In Jolivet, P., Petitpierre, E., and Hsiao, T.H. (eds.), Biology of Chrysomelidae. Kluwer Academic Publishers, London.
Gyllenhal, L. 1813. Insecta Suecica Descripta. Classis 1. Coleoptera sive Eleutherata. Tome 1. Pars III. Scaris, Leverentz.
Hope, F.W. 1840. The Coleopterist`s Manual, Part the Third, Containing Various Families, Genera and Species of Beetles, Recorded by Linnaeus and Fabricius. Also, Descriptions of Newly Discovered and Unpublished Insects. J.C. Bridgewater and Bowdery and Kerby, London.
Jolivet, P. 1978. Selection trophique chez les Clytrinae, Cryptocephalinae et Chlamisinae (Camptosoma) et les Lamprosomatinae (Cyclica) (Coleoptera, Chrysomelidae). Acta Zoologica et Pathologica Antwerpiensia, Antwerp, 70:167-200.
Jolivet, P., Petitpierre, E., and Hsiao, T.H. 2012. Biology of Chrysomelidae. Springer Science & Business Media, Dordrecht.
Konstantinov, A.S. 2010. Subfamily Lamprosomatinae Lacordaire, 1848, p. 563-564. In Löbl, I. and Smetana, A. (eds.), Catalogue of Palaearctic Coleoptera. Vol. 6. Chrysomeloidea. Apollo Books, Stenstrup.
Lacordaire, J.T. 1848. Monographie des coléoptères subpentamères de la famille des phytophages. Tome second. Mémoires de la Société Royale des Sciences de Liège, 5:vi + 890.
Lambert, J.B., Santiago-Blay, J.A., Wu, Y., and Levy, A.J. 2015. Examination of amber and related materials by NMR spectroscopy. Magnetic Resonance Chemistry, 53(1):2-8. https://doi.org/10.1002/mrc.4121
Langenheim, J.H. 2003. Plant Resins: Chemistry, Evolution, Ecology and Ethnobotany. Timber Press, Portland, Oregon.
Latreille, P.A. 1802. Histoire Naturelle, Générale et Particulière des Crustacés et des Insectes. Ouvrage Faisant Suite à L`histoire Naturelle Générale et Particulière, Composée par Leclerc de Buffon, et Rédigée par C. S. Sonnini, Membre de Plusieurs Sociétés Savantes. Tome Troisième. Familles Naturelles des Genres. F. Dufart, Paris.
Latreille, P.A. 1804. Histoire Naturelle, Générale et Particulière des Crustacés et des Insectes. Ouvrage Faisant Suite aux Oeuvres de Leclerc de Buffon, et Partie du Cours Complet D`histoire Maturelle Rédigée par C. S. Sonnini, Membre de Plusieurs Sociétés Savantes. Tome Douzième. F. Dufart, Paris.
Legalov, A.A. 2016. New genus of seed beetles (Coleoptera, Chrysomelidae, Bruchinae) in Baltic amber. Paleontological Journal, 50(1):73-77. https://doi.org/10.1134/S0031030116010056
Linnaeus, C. 1758. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, locis. Tomus I. Edition 10. Laurentii Salvii, Holmiae.
Medvedev, L.N. 1968. К познанию жуков-листоедов подсемейств Lamprosominае и Chlamysinae (Coleoptera, Chrysomelidae) Демократической Республики Вьетнам [On the knowledge of leaf-beetles of the subfamilies Lamprosominae and Chlamysinae (Coleoptera, Chrysomelidae) from Viet-Nam]. Entomological Review, 47(3):556-566. (In Russian)
Monrós, F. 1956. Revisión genérica de Lamprosomatinae con descripción de la algunos géneros y especies nuevas (Col., Chrysomelidae). Revista Agronómica del Noroeste Argentino [Tucuman], 2(1):25-77.
Monrós, F. 1958. Notes on Lamprosomatinae (Chrysomelidae). The Coleopterists Bulletin, 12:29-33. https://www.jstor.org/stable/3998928
Moseyko, A.G. and Kirejtshuk, A.G. 2013. On the taxonomic position of new taxa of the subfamily Eumolpinae (Coleoptera, Chrysomelidae) from Baltic amber. Terrestrial Arthropod Reviews, 6:163-172. https://doi.org/10.1163/18749836-06021063
Nadein, K.S., Perkovsky, E.E., and Moseyko, A.G. 2016. New Late Eocene Chrysomelidae (Insecta: Coleoptera) from Baltic, Rovno and Danish ambers. Papers in Palaeontology, 2(1):117-137.
Perkovsky, E.E., Rasnitsyn, A.P., Vlaskin, A.P., and Taraschuk, M.V. 2007. A comparative analysis of the Baltic and Rovno amber arthropod faunas: representative samples. African Invertebrates, 48(1):229-245.
Ritzkowski, S. 1997. K-Ar-Altersbestimmungen der bernsteinführenden Sedimente des Samlandes (Paläogen, Bezirk Kaliningrad). Metalla (Sonderheft), 66:19-23.
Seeno, T.N. and Wilcox, J.A. 1982. Leaf beetle genera (Coleoptera: Chrysomelidae). Entomography, 1:1-221.
Sturm, H. 1807. Deutschlands Fauna in Abbildungen nach der Natur mit Beschreibungen. V. Abtheilung. Die Insecten. Zweites Bändchen. Käfer. J. Sturm, Nürnberg.
Szwedo, J. and Sontag, E. 2013. The flies (Diptera) say that amber from the Gulf of Gdańsk, Bitterfeld and Rovno is the same Baltic amber. Polish Journal of Entomology, 82:379-388. https://doi.org/10.2478/pjen-2013-0001
Turkin, N.I. 1997. Preliminary results of microscopic research of tangential wood imprints in Baltic amber. Metalla (Sonderheft), 66:55-56.
Warchałowski, A. 2003, The Leaf-beetles (Chrysomelidae) of Europe and the Mediterranean Region. Natura Optima dux Foundation, Warszawa.
Wolfe, A.P., Tappert, R., Muehlenbachs, K., Boudreau, M., McKellar, R.C., Basinger, J.F., and Garrett, A. 2009. A new proposal concerning the botanical origin of Baltic amber. Proceedings of the Royal Society B: Biological Sciences, 276:3403-3412. https://doi.org/10.1098/rspb.2009.0806
