

 Catalogue and composition of fossil Anthicidae and Ischaliidae (Insecta: Coleoptera)
Catalogue and composition of fossil Anthicidae and Ischaliidae (Insecta: Coleoptera)
Article number: 22.1.18
https://doi.org/10.26879/885
Copyright Paleontological Society, April 2019
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 9 May 2018. Acceptance: 18 March 2019
{flike id=2484}
ABSTRACT
Despite the increasing rate of systematic research on extant tenebrionoid Coleoptera of the Anthicidae and Ischaliidae, their fossil records remained largely unrevised. In the current paper we review all hitherto named ant-like flower beetles and false fire-coloured beetles fossils. We suggest 16 fossil species can be reliably assigned to the Anthicidae and three species to the Ischaliidae. We proposed new placements for two fossil Anthicidae taxa: Petratypus nigri Kaddumi, 2005 moved from Anthicidae to Cucujiformia Familia incertae sedis and “Eurygenius” wickhami Cockerell, 1917 is re-described and moved from Eurygeniinae Anthicidae to Tenebrionoidea Familia and Genus incertae sedis. Additionally, three new species are described from Eocene Baltic amber, namely Nitorus succinius sp. nov., Steropes eleticinoides sp. nov. and Tomoderus saecularis sp. nov. An annotated catalogue of fossil Anthicidae and Ischaliidae is provided. We made a qualitative analysis of available data, evaluated the distribution of fossils in the light of current biogeography and geological time. The oldest hitherto known fossil record of the Anthicidae is 130.0-125.5 Ma (same for Macratriinae), of the Anthicinae - 37.2-33.9 Ma, of the Eurygeniinae - 55.8-48.6 Ma, of the Notoxinae, Steropinae and Tomoderinae - 37.2-33.9 Ma. The oldest hitherto know fossil record of the Ischaliidae is 37.2-33.9 Ma.
Dmitry Telnov. (ORCID: 0000-0003-3412-0089) Natural History Museum, Department of Life Sciences, Cromwell Road, SW7 5BD, London, United Kingdom. anthicus@gmail.com
and Institute of Biology, University of Latvia, Miera iela 3, LV-2169, Salaspils, Latvia
Andris Bukejs. Institute of Life Sciences and Technologies, Daugavpils University, Vienības iela 13, LV-5401, Daugavpils, Latvia. carabidae@inbox.lv
Keywords: fossils; review; new species; new placements; critical features
Final citation: Telnov, Dmitry and Bukejs, Andris. 2019. Catalogue and composition of fossil Anthicidae and Ischaliidae (Insecta: Coleoptera) Palaeontologia Electronica 22.1.18A 1-27. https://doi.org/10.26879/885
palaeo-electronica.org/content/2019/2484-fossil-anthicidae-ischaliidae
Copyright: April 2019 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/FDB6E319-C6D9-4861-89CE-E895C8C9D782
INTRODUCTION
Anthicidae (Insecta: Coleoptera), ant-like flower beetles, is a large group of Tenebrionoidea with a cosmopolitan distribution (Chandler, 2010) and over 3500 species (Telnov, 2008) in eight subfamilies (Chandler, 2010). Two groups of uncertain placements, until recently arranged to the Anthicidae, are the Afreminae and Lagrioidinae (Lawrence et al., 2010). There are various fossil records for ant-like flower beetles from around the world.
Ischaliidae (Insecta: Coleoptera), false fire-coloured beetles, is a small monotypic family of nearly 50 species almost exclusively of Northern Hemisphere distribution (Gusakov and Telnov, 2007; Young, 2011), with few fossil records from Eocene Baltic amber (Alekseev and Bukejs, 2017).
There has never been a phylogenetic analysis at the family level for either Anthicidae or Ischaliidae. The Anthicidae has not been formally established as a monophyletic group, and the family, particularly the Steropinae, is morphologically close to the Eleticinae (Meloidae) (Chandler, 2010) and Macratriinae appear closer to Ischaliidae than to Anthicinae (Batelka et al. 2016).Afreminae and Lagrioidinae may not be monophyletic with the Anthicidae (Lawrence and Ślipiński, 2013), and we follow Lawrence et al. (2010) who treated them as Tenebrionoidea incertae sedis (no fossils are known for these two groups). While knowledge of morphological evolution and biogeography of the Anthicidae has grown considerably over the last few decades, their fossils have remained insufficiently studied, with sparse records scattered over numerous publications with the first known records published already 180 years ago (Hope, 1836). In contrast, the first fossil representative of the Ischaliidae was only described a few years ago (Alekseev and Telnov, 2016).
Fossils represent an essential data source for resolving relationships, understanding morphological character evolution and assessing the tempo and mode of diversification (Tarasov et al., 2016). We argue that the minimal age for the Anthicidae should be aligned with the age of Camelomorpha longicervix Kirejtshuk, Azar et Telnov in Kirejtshuk and Azar, 2008 (Macratriinae), of which the oldest record is from the Early Cretaceous. The Eurygeniinae and the Lemodinae are known to appear first in the Late Cretaceous, while the Anthicinae - in the Palaeocene. For the Ischaliidae, all known fossil records are from the Eocene.
We plan to catalogue the currently known fossil records of the Anthicidae and Ischaliidae, describe new species and solve nomenclatural uncertainties, provide a preliminary analysis of minimal age of the main subgroups, briefly discuss their morphological features and compile references on the included taxa.
MATERIALS AND METHODS
The published data on fossil Anthicidae and Ischaliidae are summarized from previous reports and supplemented with three new species and additional records described in this paper. The catalogue comprises the following data: full valid taxon name, synonymy (if applicable), list of references and age of fossil records. The nomenclature of the Anthicidae generally follows Chandler (2010), of the Ischaliidae - Gusakov and Telnov (2007), Young (2011). All taxa are listed alphabetically since a phylogenetic arrangement is currently impossible. Authors’ comments are placed in square brackets. Specimens were studied using a Leica® S6D stereomicroscope and photographed using a Canon® EOS 450D DSLR camera attached to this stereomicroscope. Multiple photographs were taken at different focal planes and reassembled using CombineZP software.
Institutional Abbreviations
The material examined is deposited in the following collections:
- Center of Natural History (CeNak) (formerly Geologisch-Paläontologisches Institut und Museum, GPIH) of the University of Hamburg, GPIH (Hamburg, Germany);
- Kaliningrad Amber Museum, KAM (Kaliningrad, Russia);
- Latvian Natural History Museum, LDM (Rīga, Latvia);
- Museo de Ciencias Naturales de Álava, MCNA (Vitoria-Gasteiz, Spain);
- Muséum d’Histoire naturelle de Marseille, MHNM (Marseille, France);
- Muséum national d’Histoire naturelle, MNHN (Paris, France);
- Natural History Museum (British Museum, Natural History), BMNH (London, United Kingdom);
- Yale Peabody Museum of Natural History, YPM (New Haven, USA);
- collection of Christel and Hans Werner Hoffeins, CCHH (Hamburg, Germany) subsequently will be deposited in the collection of the Senckenberg Deutsches Entomologisches Institut, SDEI (Müncheberg, Germany); and
- collection of Francisco Molino-Olmedo, CMOM (Mancha Real (Jaén), Spain).
SYSTEMATIC PALAEONTOLOGY
Descriptions of New Taxa
Order Coleoptera Linnaeus, 1758
Suborder Polyphaga Emery, 1886
Superfamily Tenebrionoidea Latreille, 1802
Family Anthicidae Latreille, 1819
Subfamily Anthicinae Latreille, 1819
Tribus Anthicini Latreille, 1819
Genus Nitorus Telnov, 2007
Remarks. The specimen considered here was assigned to the genus Nitorus within the tribe Anthicini, based on the following morphological characters: (1) mesosternum simple, laterally curved to the mesocoxal cavities, (2) mesocoxal cavities not isolated from mesepisterna, (3) lateral margins of mesosternum nearly straight, (4) mesepisterna without fringe of setae and glossy, (5) elytra with distinct postbasal transverse impression and (6) dorsal body smooth, glossy and sparsely pubescent.
Nitorus succinius sp. nov.
Figure 1.1-2, Figure 2.1-4
zoobank.org/6104A725-3B18-405C-B036-7016A9B52484
Type material. Holotype: female ID “SDEI Coleoptera # 301526” [CCHH]: beetle inclusion in a small amber piece embedded in a block of GTS-polyester resin with dimensions 16×7×4 mm, adult female with exposed last visible ventrites. Tarsomeres 2-4 of right metatarsus are missing. No syninclusions present. Specimen provided with two additional printed labels: “HOLOTYPUS” [label red, black framed] and “NITORUS succinius sp. nov. det. D.Telnov and A.Bukejs, 2018”.
 Type stratum. Baltic amber, Mid-Eocene to Upper Eocene.
Type stratum. Baltic amber, Mid-Eocene to Upper Eocene.
Type locality. Yantarny settlement (formerly Palmnicken), Sambian (Samland) Peninsula, Kaliningrad Region, Russia.
Derivatio nominis. This first fossil representative of the genus Nitorus named from Latin “succinius” [amber].
Differential diagnosis. Since no other fossil Nitorus are presently known, this new species can be compared with extant taxa only. Few extant Nitorus share similar body colouration (uniformly pale forebody, uniformly dark brown elytra) with N. succinius sp. nov. Nitorus acuticollis (Krekich-Strassoldo, 1914) (E India) and N. latissimus (Pic, 1907) (“India” without exact locality) both have dorsal forebody densely punctured, opaque (sparsely punctured and smooth in N. succinius sp. nov.) and elytra are less strong punctured in basal half in these species (basal half of elytra covered with dense and deep punctures in N. succinius sp. nov.). Nitorus sensitivus (Krekich-Strassoldo, 1928) (N India, Nepal) has much longer (extending to mid-length of elytra) and slender antennae, tapered head base (broadly rounded in N. succinius sp. nov.), disc of pronotum with very large, irregular, coarse but shallow punctures in basal half (punctures are sparse, deep and not coarse in N. succinius sp. nov.). Other extant Nitorus species with pale forebody and dark coloured elytra all have pale elytral markings or transverse hair bands. Also in generally paler coloured species (like orange to pale castaneous Indian N. brevitarsis (Krekich-Strassoldo, 1931) or Oriental N. lictor (Fairmaire, 1896)) a paler (white or almost white) transverse elytral band is present in postbasal transverse impression.
Description. Measurements: Total body length about 2.8 mm [exposed last visible ventrites excluded], maximum combined width across middle of elytra 0.9 mm. Head about 0.6 mm long [not possible to take exact measurements due to the position of head], across eyes 1.03 mm wide, pronotum 0.5 mm long, maximum width 0.9 mm, minimum width in constriction area 0.65 mm, elytra 1.7 mm long, maximum width 0.9 mm.
 Colouration: Forebody pale orange with pale castaneous antennae. Elytra dark brown. Legs orange to pale castaneous, pro- and mesocoxae pale orange. Ventral side pale castaneous.
Colouration: Forebody pale orange with pale castaneous antennae. Elytra dark brown. Legs orange to pale castaneous, pro- and mesocoxae pale orange. Ventral side pale castaneous.
Head dorsally and ventrally smooth, with medium-sized, slightly prominent eyes. Head base very broadly rounded posterior to eyes. Dorsal surface deeply minutely punctate, intervening spaces smooth, about 3-4× as large as punctures. Pubescence sparse, very inconspicuous, appressed. Antennae extending to the postbasal transverse impression of elytra; antennomere 3 slender, 1.3× as long as antennomere 2; antennomeres 3-7 elongate, thickened distally; antennomeres 8-10 shortened, cylindrical and stronger thickened than preceding ones; terminal antennomere in female elongate, broad, tapered distally, about 1.8-2× as long as penultimate antennomere. Terminal maxillary palpomere fusiform, medially widened on inner margin.
Pronotum smooth dorsally, lateral and anterior margins evenly rounded, with strong lateral postmedian constriction. Prebasal area cylindrical, distinctly narrower than anterior portion of pronotum. Anterior rim narrow. Antebasal sulcus broad. Punctures on disc denser, larger (and of different sizes) and deeper than on head, intervening spaces smooth, about as large as to 2× as large as punctures. Punctures on lateral margins are minute, with wider intervening spaces. Antebasal sulcus area densely, largely, almost coarsely punctured. Pubescence longer and denser than on head, appressed, directed posteriorly. Scutellar shield small, rounded apically, covered with long and dense golden pubescence all along its apical and (partly) lateral margins.
Elytra elongate, smooth and shiny, slightly widened in the middle, with shallow but distinct postbasal transverse impression. Punctures very large, deep and dense in basal half, becoming significantly smaller and sparser in apical half. Intervening spaces smaller than to as large as punctures in basal half, but becoming 2-3× as large as those in apical half of elytra. Each puncture bearing long, appressed and curved seta. A few scattered rather short erect tactile setae in basal half and on sides of elytra. Sutural striae absent. Metathoracic wings apparently fully developed (humeri well developed).
Underside of the body smooth. Metasternum and abdominal ventrites evenly pubescent, setae appressed. Female morphological tergite VII and sternite VII both broadly rounded and densely setose apically.
Legs long and slender, smooth, finely and densely appressedly pubescent. Penultimate tarsomeres narrow, indistinctly bilobate. Claws simple, without appendages or teeth. Female basal metatarsomere as long as combined length of remaining tarsomeres.
Sexual dimorphism. Male is unknown.
Remarks. The present distribution of Nitorus is confined to southern and eastern Palaearctic (Central Asia, Far East of Russia, Japanese Archipelago, Korean Peninsula, China, and Himalaya), whole Afrotropical (inclusive Madagascar) and Oriental regions (inclusive insular systems of Greater Sundas and the Philippines). Nitorus is absent from the New World fauna. Its presence in Australia and on New Guinea is not yet confirmed. In Western Palaearctic Nitorus is known from southern boundaries, e.g., Egypt and Mauritania (N. unifasciatus unifasciatus (Desbrochers des Loges, 1875)). In Central and southern Asia several taxa present nowadays, namely N. apricans apricans (Krekich-Strassoldo, 1931) (Afghanistan, Pakistan), N. apricans lindbergi (Bonadona, 1964) (Afghanistan), N. petri (Pic, 1894) (Tian Shan Mountains, Kazakhstan, Turkmenistan), N. suasum Telnov, 2010 (Afghanistan), and N. unifasciatus unifasciatus (unconfirmed Iran record, see Telnov and Ghahari, 2018). The formerly wider distribution of Nitorus correlates with worldwide warmer and more uniform climate of Eocene.
Subfamily Steropinae Jacquelin du Val, 1863
Genus Steropes Steven, 1806
Remarks. The specimen considered here was assigned to the genus Steropes within the subfamily Steropinae on the basis of the following morphological features: (1) procoxal cavities open internally and (presumably, not visible in the holotype) externally, (2) mesoventrite triangular, (3) mesoventrite not completely fused with mesepisterna, suture present in-between, (4) pronotum dorsally without anterior rim, (5) pronotum with complete antebasal sulcus which ends in a pit at some distance from procoxa, (6) neck one-third of head width, (7) palpomeres 2-3 slightly widened and rounded distally, (8) terminal maxillary palpomere scalene-triangular, (9) procoxa elongate, truncate fusiform, (10) apical three antennomeres greatly lengthened, antennomeres 4-8 distally rounded, and (11) tarsal claw with spinose ventral appendage.
Steropes eleticinoides sp. nov.
Figure 3.1-2, Figure 4.1-2
zoobank.org/C6C19F5C-2F14-4121-85D9-E8700D34AA72
 Type material. Holotype: female ID “LDM G 404/78” [LDM]: beetle inclusion in a moderate large amber piece of irregular form with dimensions about 40×22×6 mm, adult female with exposed last visible ventrites. Syninclusions are represented by one fungus gnat (Insecta: Diptera: Sciaridae, Gen. sp.), one springtail (Collembola Fam. and Gen. sp.) and four fagacean stellate hairs. Specimen provided with two additional printed labels: “HOLOTYPUS” [red label with black frame] and “STEROPES eleticinoides sp. nov. det. D.Telnov and A.Bukejs, 2018”.
Type material. Holotype: female ID “LDM G 404/78” [LDM]: beetle inclusion in a moderate large amber piece of irregular form with dimensions about 40×22×6 mm, adult female with exposed last visible ventrites. Syninclusions are represented by one fungus gnat (Insecta: Diptera: Sciaridae, Gen. sp.), one springtail (Collembola Fam. and Gen. sp.) and four fagacean stellate hairs. Specimen provided with two additional printed labels: “HOLOTYPUS” [red label with black frame] and “STEROPES eleticinoides sp. nov. det. D.Telnov and A.Bukejs, 2018”.
Type stratum. Baltic amber, Mid-Eocene to Upper Eocene.
Type locality. Yantarny settlement (formerly Palmnicken), Sambian (Samland) Peninsula, Kaliningrad Region, Russia.
Derivatio nominis. Since a) this is the second known anthicid species with ventral spines (strongly elongate lobes) on the tarsal claws and b) Steropinae are phylogenetically and morphologically similar to Eleticinae Wellman, 1910 of Meloidae (Chandler, 2010), which are known to have spinose tarsal claws (Bologna et al., 2010; Chandler, 2010), this new species is named to indicate its similarity with the Eleticinae subfamily of blister beetles.
Differential diagnosis. This is the only known Steropes species and the second taxon among the Steropinae (see also original description of Australosteropes in Telnov (2018)) with peculiar, ventrally spinose tarsal claws. Among other tenebrionoid Coleoptera spinose claws currently only known in Meloidae and Stenotrachelidae (Ivie, 2002; Bologna et al., 2010; Lawrence and Ślipiński, 2010).
 Description. Measurements: Total body length about 6 mm [exposed last visible ventrites excluded]. Head about 1.15 mm long [not possible to take exact measurements due to the position of head], across eyes 1.03 mm wide, pronotum 1.2 mm long, elytra 3.8 mm long, maximum combined width about 1.3-1.5 mm [impossible to take exact measurements due to the position of the specimen].
Description. Measurements: Total body length about 6 mm [exposed last visible ventrites excluded]. Head about 1.15 mm long [not possible to take exact measurements due to the position of head], across eyes 1.03 mm wide, pronotum 1.2 mm long, elytra 3.8 mm long, maximum combined width about 1.3-1.5 mm [impossible to take exact measurements due to the position of the specimen].
Colouration: Dorsum and venter uniformly pale brown, head darker brown with reddish mouthparts. Legs and palps yellow-brown.
Head dorsally subopaque with large and strongly prominent eyes. Eye shortly notched on anterior margin at the insertion of antenna. Head base broadly rounded posterior to eyes. Broad shallow transverse impression at place of frontoclypeal suture. Frons narrow between eyes; in female minimum interocular distance almost half of eye diameter, slightly greater than length of antennomere 1. Dorsal surface deeply punctate, intervening spaces about as large as, to smaller than punctures. Pubescence sparse, inconspicuous, suberect, directed anteriorly. Several longer and erect setae present on vertex and along inner margin of eyes. Antennae extending towards basal third of elytra; antennomeres 3–7 elongate, slightly thickened distally, antennomere 3 the longest; female antennomere 8 shortened, 9–11 greatly lengthened and thickened, antennomeres 9–10 widened distally; female terminal antennomere strongly elongate, broad, tapered distally, about 1.1× as long as combined length of two preceding antennomeres; combined length of antennomeres 1–8 and 9–10 nearly equal [exact measuring not possible due to the position of the specimen]. Maxillary palpomeres 2-3 slightly widened distally. Terminal maxillary palpomere scalene triangular, tapered distally. Neck more than 1/3 of head width, densely and coarsely punctured antennomere 7 about as long as wide, 9–10 transverse and shorter than preceding ones; terminal antennomere elongate, asymmetrically triangular, tapered distally, as long as antennomeres 9–10 combined. Terminal maxillary palpomere fusiform, medially angulate on inner margin.
Pronotum smooth dorsally, slightly constricted laterally towards narrower base, its lateral and anterior margins evenly rounded. Anterior rim lacking dorsally, well-defined laterally and ventrally. Pronotal base dorsally with a complete basal bead and antebasal sulcus originating at lateral fovea. Lateral pronotal fovea conspicuous, forming deep and narrow internally closed incision which ends shortly in a pit on both sides. Punctures dense and uniform (inclusive those of antebasal sulcus area), intervening spaces smooth, about as large as to 2× as large as punctures. Pubescence dense, inconspicuous, suberect, directed posteriorly. Scutellar shield not visible.
Elytra elongate, smooth and shiny, slightly widened around mid-length, with postbasal transverse impression hardly indicated. Humeri obsolete, rounded. Punctures large, deep and dense in basal half, becoming smaller, shallower and sparser in apical half. Punctures arranged into three groups of irregular longitudinal striae on each elytron: the inner one, sutural, initially consists of two irregular rows of punctures of which internal one discontinues in basal third but the outer adjoins the suture and continues towards elytral apex; the median one, consists of three irregular rows of punctures becoming irregular postmedium and discontinue in apical third; the outer one, consists of at least four irregular rows of punctures, almost reaching elytral apex. There are narrow unpunctured areas between these three puncture groups (Figure 5.1). Intervening spaces smaller than up to as large as punctures in basal half but becoming larger than those in apical half of elytra. Pubescence yellowish, longer and denser than on forebody (but still sparse), appressed, directed posteriad. A few scattered long erect tactile setae on disc and sides of elytra.
Metasternum and abdominal ventrites smooth, minutely punctate and with inconspicuous pubescence. Procoxal cavities open externally (presumably also internally). Mesoventrite triangular, mesepisterna almost meeting in front of it [not fully visible because of the position of specimen]. Mesosternum not fused with mesepisterna, with suture present in-between. Metacoxae narrowly separated by acute intercoxal projection. Female morphological tergite VII and sternite VII both broadly rounded on apical margin.
Legs long and slender. Femora broad, finely, sparsely inconspicuously pubescent. Tibiae covered with long setae which become denser in distal half. Tibial spurs paired, acute; spurs each with 5-6 long microspines on each side. Penultimate tarsomeres widened, indistinctly bilobate. Claws long, curved in distal half and each with a long acute ventral spine of about two-third of claw length. Female basal metatarsomere almost as long as combined length of other tarsomeres.
Sexual dimorphism. Male is unknown.
Remarks. The present distribution of Steropinae is Holarctic (Chandler, 2010), Oriental and Australian (Telnov, 2018). Steropes inhabit arid and semiarid areas of the Palaearctic, Nearctic and NE Afrotropical (Arabian Peninsula) regions (Chandler et al., 2008), with single species each in the Oriental (Telnov, 2006) and Australian (Telnov, 2018) regions. Extant Steropes, except S. hercules Telnov, 2006 from rainforests of Vietnam, seem to avoid forested areas and nowadays occur mostly in open and semi-open landscape. But the Eocene record of S. eleticinoides sp.nov. is apparently from ancient pine tree forest. In Western Palaearctic, currently northernmost records of this genus are from Romania, Moldova and Ukraine. The formerly wider distribution of Steropes correlate with worldwide warmer and uniform climate of Eocene.
Subfamily Tomoderinae Bonadona, 1961
Genus Tomoderus LaFerté-Sénectère, 1849
Remarks. The specimen considered here was assigned to the genus Tomoderus within the Tomoderinae based on the following morphological characters: (1) pronotum lacking apical rim or collar, (2) distinct lateral antebasal constriction present on pronotum, (3) metacoxae widely separated and (4) intercoxal process of 1st abdominal ventrite broadly rounded.
Tomoderus saecularis sp. nov.
Figure 5.1-2
zoobank.org/F57F11FC-6CD0-4E34-B037-CEF81D85C2BB
Type material. Holotype: sex unknown, ID "GPIH no. 4958, coll. Gröhn no. 8588” [GPIH]: beetle inclusion in a small amber piece of trapezoidal shape with dimensions 15×10×5 mm, adult specimen with all body parts present. Numerous syninclusions present: pieces of soil and plant material. Specimen provided with two printed labels: “HOLOTYPUS” [label red, black framed] and “TOMODERUS saecularis sp. nov. det. D.Telnov and A.Bukejs, 2018”.
 Type stratum. Baltic amber, Mid-Eocene to Upper Eocene.
Type stratum. Baltic amber, Mid-Eocene to Upper Eocene.
Type locality. Yantarny settlement (formerly Palmnicken), Sambian (Samland) Peninsula, Kaliningrad Region, Russia.
Derivatio nominis. This species named from Latin “saecularis” [century, 100 years old] in honour of the 100th anniversary of the Republic of Latvia (1918-2018), where Baltic amber deposits are also known.
Differential diagnosis. Known fossil congeners (T. balticus Telnov, 2012a and T. longelytratus Telnov, 2013) are both larger and have differently punctured elytra and pronotum. Among extant taxa T. saecularis sp. nov. is peculiar primarily in elytral punctures (which are aligned into three groups of irregular punctural rows on each elytron) in combination with roughly punctured pronotal disc and presence of median longitudinal impression on anterior lobe of pronotum, remarkably large eyes and short tempora.
Description. Measurements: Total body length about 2.8 mm [exposed last visible ventrites excluded], maximum combined width across middle of elytra 0.9 mm. Head about 0.5 mm long [not possible to take exact measurements due to the position of head], across eyes about 0.45 mm wide [not possible to take exact measurements due to the position of head], pronotum 0.5 mm long, maximum width about 0.55 mm [not possible to take exact measurements due to the position of pronotum], minimum width at constriction about 0.4 mm [not possible to take exact measurements due to the position of pronotum], elytra 1.3 mm long, maximum combined width about 0.8 mm [not possible to take exact measurements due to the position of elytra].
Colouration: Uniformly brown with paler (yellowish to yellow-brown) legs, maxillary palps and antennomeres 1-9; three terminal antennomeres darkened.
Head dorsally and ventrally smooth. Eyes large, prominent, almost circular, truncate opposite the insertions of antennae. Tempora much shorter than length of an eye, temporal angles broadly rounded. Head base truncate posterior to eyes. Dorsal surface with rather large and very shallow punctures, intervening spaces smooth, up to as large as punctures. Pubescence very sparse and inconspicuous; few very long erect tactile setae present at base and on tempora. Antennae extending to base of elytra; basal antennomere long, rugulose; antennomere 3 slender, 1.2× as long as antennomere 2; antennomeres 3-6 thickened distally; antennomeres 7 about as long as wide, 9-10 transverse and shorter than preceding ones; terminal antennomere elongate, asymmetrically triangular, tapered distally, as long as antennomeres 9-10 combined. Terminal maxillary palpomere fusiform, medially widened on inner margin.
Pronotum smooth dorsally, wider than head, with strong lateral postmedian constriction, truncate on anterior, evenly rounded on lateral margins. Anterior rim and antebasal sulcus not present dorsally. Posterior lobe of pronotum distinctly narrower than anterior. Anterior lobe with a track of longitudinal median impression, that reaches the postmedian lateral constriction. Punctures on disc of anterior lobe large and dense but shallow, intervening spaces smooth, smaller than punctures; those punctures situated on lateral margins are minute, with wider intervening spaces. Lateral postmedian constriction dorsally largely, coarsely punctured. Pubescence short, sparse and subdecumbent with some long erect tactile setae present on sides and on disc. Scutellar shield transverse, densely punctured, truncate apically.
Elytra elongate, smooth and shiny, slightly widened around mid-length, with postbasal transverse impression hardly indicated. Humeri obsolete, rounded. Punctures large, deep and dense in basal half, becoming smaller, shallower and sparser in apical half. Punctures arranged into three groups of irregular longitudinal striae on each elytron: the first one, sutural, initially consists of two irregular rows of punctures of which internal one discontinues in basal third but the outer adjoins the suture and continues towards elytral apex; the median one, consists of three irregular rows of punctures becoming irregular postmedium and discontinue in apical third; external one, consists of at least four irregular rows of punctures, almost reaching elytral apices. There are narrow unpunctured areas between these three puncture groups (Figure 5.1). Intervening spaces smaller than up to as large as punctures in basal half but becoming larger than those in apical half of elytra. Pubescence yellowish, longer and denser than on forebody (but still sparse), appressed, directed posteriad. A few scattered long erect tactile setae on disc and sides of elytra.
Underside of the body smooth, glabrous. Metacoxae broadly separated.
Legs slender, smooth, finely and densely appressedly pubescent on femora, denser pubescent on tibiae. Penultimate tarsomeres narrow, deeply bilobate. Claws simple, without appendages or teeth, rather long. Basal metatarsomere slightly longer than combined length of remaining tarsomeres.
Sexual dimorphism. Unknown.
Systematic Changes
Tenebrionoidea family incertae sedis
“Eurygenius” wickhami Cockerell, 1917 genus incertae sedis
Figure 6.1-2
Cockerell, 1917 (new species, initially assigned to Eurygenius (Pedilidae s. lat.), Burmese amber, 99.7–94.3 Ma); Fletcher, 1920; Zherikhin, 1980; Spahr, 1981; Keilbach, 1982; Poinar, 1992; Ross, 1998; Ross and York, 2000; Kirejtshuk, 2002-2013; Kirejtshuk and Azar, 2008; Ross et al., 2010; Wang and Zhang, 2011; Soriano et al., 2014; Peris et al., 2016 (in Pyrochroidae but without any comments on the placement); Guo et al., 2017; Kirejtshuk et al., 2019 (mentioned Eurygeniinae from Burmese amber but not species).
 Remarks. Eurygenius wickhami was originally described by Cockerell (1917) as a member of Pedilidae. From the beginning the author was unsure about the placement of his new taxon, which follows from his comment: “It may go in Eurygenius as interpreted in the broader sense, though it may hereafter be treated as the type of a distinct genus” (Cockerell, 1917: 325). As a member of the “Pseudo-Anthicites”, Eurygenius LaFerté-Sénectère, 1849 was initially placed in the Pedilidae until relatively recently, when Pedilinae were removed from the Anthicidae to Pyrochroidae by Young and Pollock (1991). Type of this genus is Eurygenius reichei LaFerté-Sénectère, 1849 by monotypy. Presently, Eurygenius is a small genus of six New World species distributed from Brazil (where the type species coming from) to the southern United States (Telnov, unpublished data).
Remarks. Eurygenius wickhami was originally described by Cockerell (1917) as a member of Pedilidae. From the beginning the author was unsure about the placement of his new taxon, which follows from his comment: “It may go in Eurygenius as interpreted in the broader sense, though it may hereafter be treated as the type of a distinct genus” (Cockerell, 1917: 325). As a member of the “Pseudo-Anthicites”, Eurygenius LaFerté-Sénectère, 1849 was initially placed in the Pedilidae until relatively recently, when Pedilinae were removed from the Anthicidae to Pyrochroidae by Young and Pollock (1991). Type of this genus is Eurygenius reichei LaFerté-Sénectère, 1849 by monotypy. Presently, Eurygenius is a small genus of six New World species distributed from Brazil (where the type species coming from) to the southern United States (Telnov, unpublished data).
After studying the holotype (BMNH) it became evident this species is neither Eurygenius nor Eurygeniinae where it was assigned initially. The following basic features of Eurygeniinae are absent in this species: head base truncate or concave except in Steriphodon Abeille de Perrin, 1895 (head base either very broadly rounded in “Eurygenius” wickhami), mandible scooped with wide apex (mandible apice unidentate with tapered apex in “Eurygenius” wickhami), pronotum anteriorly with dorsal rim flanged (except strongly protruding in Mitraelabrini Abdullah, 1969) and ventral rim much narrower (pronotum without anterior dorsal rim in “Eurygenius” wickhami), antebasal sulcus of pronotum distinct (antebasal sulcus absent dorsally or very inconspicuous in “Eurygenius” wickhami), elytra coarsely and densely punctured (elytral punctures fine and sparse in “Eurygenius” wickhami). Moreover, morphological features as visible in “Eurygenius” wickhami holotype are not sufficient to classify this taxon as the Anthicidae. Considering similarities in external morphology (and phylogenetic relationship) between Mycteridae Blanchard, 1845, Pyrochroidae Latreille, 1807 and Anthicidae, the following characters are crucial to examine prior to assigning this taxon to any of families listed afore (Ivie, 2002; Young, 2005; Chandler, 2010; Pollock, 2010; Young and Pollock, 2010): (1) antebasal sulcus of pronotum extends / not extends laterally onto hypomeron; (2) prothoracic coxal cavities open externally and closed internally / open both externally and internally / procoxal cavities closed internally, open externally; (3) compound eyes conspicuously setose between facets / not setose; (4) metathoracic coxae nearly contiguous mesally / separated; (5) intercoxal process of abdominal ventrite 1 with margins diverging at angle less than 45° / more than 45°; (6) metacoxa extending laterally to body side, completely separating metepisternum and abdominal ventrite 1 / not reaching elytron or side of body, metepisternum and abdominal ventrite 1 in contact laterad metacoxa. These features are not visible in “Eurygenius” wickhami holotype. Therefore, this taxon cannot be certainly assigned to Anthicidae, Mycteridae, or Pyrochroidae. Until additional material of “Eurygenius” wickhami Cockerell, 1917 becomes available or technical progress will allow more detailed study of the available inclusion, based on aforementioned facts, we reject the original combination and propose a new placement for “Eurygenius” wickhami as a Genus incertus and Familia incerta within the superfamily Tenebrionoidea Latreille, 1802.
Type material. Holotype Eurygenius wickhami Cockerell, 1917, sex unknown, ID “Swinhoe Collection, NHMUK PI In 19110” [BMNH]: beetle inclusion in a large amber piece of irregular form, adult specimen with metathoracic wings and tip of abdomen (tergite VII) partly exposed from under partly open elytra. The holotype specimen is in a thick slab of dark amber with numerous syninclusions (see the list and inclusion description in Ross and York, 2000), and a ventral view of this specimen is not available.
Type stratum. Burmese amber, Cretaceous.
Type locality. Hukawng Valley, Kachin State, N Burma (now Myanmar).
Re-description. We provide a supplemental description of this species (only dorsal view available, see comment above). Dorsal surface yellowish castaneous, antennomeres 4–11 darker than basal ones. Head smooth dorsally, slightly transverse. Frontoclypeal suture apparently absent. Compound eyes very large and strongly prominent, emarginate (Cockerell (1917: 324) describes the eyes as apparently not emarginate”, which is not correct) at insertions of antennae, occupying whole sides of the head. Minimum interocular distance half of the of eye diameter. Tempora absent. Head base subtruncate or very broadly rounded, with a short median notch. Mandible long and pointed, unidentate apically. Terminal maxillary palpomere elongate, not securiform (exact shape of maxillary palpomeres remains unknown due their position; Cockerell (1917: 324) describes the shape of terminal maxillary palpomere as “elongated, subtriangular”, which is of little taxonomical use). Antennal insertions exposed, widely separated. Antenna 11-segmented, filiform, very long, extending over the postbasal transverse impression of elytra. Antennomeres elongate, finely and densely pubescent. Antennomere 3 about 1.3× as long as preceding one; antennomeres 4–8 slightly widened distally; antennomeres 9–10 shorter and distally stronger widened; terminal antennomere tapered, 1.6× as long as preceding one. Neck broad, almost half of the head width.
Pronotum smooth dorsally, about 1.1× as long as head, same width or wider than it (measured at present position of the inclusion), widest at anterior half, strongly constricted laterally at posterior third. Anterior margin rounded, posterior margin straight. Anterior dorsal rim absent. Antebasal sulcus absent dorsally or very inconspicuous. Dorsal sculpture not clearly visible, appears densely and straight longitudinally strigose under certain light conditions, but definitively sparsely punctate (at least on disc) and not strigose (Cockerell (1917: 324) describes the pronotal sculpture as “consisting of irregular longitudinal grooves”, which is either a result of an optical illusion or structure of the inclusion resin over pronotum; similar but transverse “grooves” appear over basal third of pronotum). Dorsal pubescence inconspicuous, dense. Scutellar shield with broadly triangular posterior edge.
Elytra smooth dorsally, elongate, 2.4× as long as their maximum combined width and 1.5× as long as forebody (head and pronotum), slightly widened apically. Humeral angles widely rounded but distinct. Omoplates not indicated. Postbasal transverse impression shallow, present in basal third. Punctation irregular, quite dense and shallow. Pubescence uniform, moderately long and dense, suberect (possible that elytral pubescence is complex with setae of two different lengths, but this is not evident in the holotype). Metathoracic wings fully developed.
We provide a supplemental description of this species (only dorsal view available, see comment above). Dorsal surface yellowish castaneous, antennomeres 4-11 darker than basal ones. Head smooth dorsally, slightly transverse. Frontoclypeal suture apparently absent. Compound eyes very large and strongly prominent, emarginate (Cockerell (1917: 324) describes the eyes as “apparently not emarginate”, which is not correct) at insertions of antennae, occupying whole sides of the head. Minimum interocular distance half of the of eye diameter. Tempora absent. Head base subtruncate or very broadly rounded, with a short median notch. Mandible long and pointed, unidentate apically. Terminal maxillary palpomere elongate, not securiform (exact shape of maxillary palpomeres remains unknown due their position; Cockerell (1917: 324) describes the shape of terminal maxillary palpomere as “elongated, subtriangular”, which is of little taxonomical use). Antennal insertions exposed, widely separated. Antenna 11-segmented, filiform, very long, extending over the postbasal transverse impression of elytra. Antennomeres elongate, finely and densely pubescent. Antennomere 3 about 1.3× longer than preceding one; antennomeres 4-8 slightly widened distally; antennomeres 9-10 shorter and distally stronger widened; terminal antennomere tapered, 1.6× longer than preceding one. Neck broad, almost half of the head width.
Prothorax smooth dorsally, about 1.1× as long as head, same width or wider than it (measured at present position of the inclusion), widest at anterior half, strongly constricted laterally at posterior third. Anterior margin rounded, posterior margin straight. Anterior dorsal rim absent. Antebasal sulcus absent dorsally or very inconspicuous. Dorsal sculpture not clearly visible, appears densely and straight longitudinally strigose under certain light conditions, but definitively sparsely punctate (at least on disc) and not strigose (Cockerell (1917: 324) describes the pronotal sculpture as “consisting of irregular longitudinal grooves”, which is either a result of an optical illusion or structure of the inclusion resin over pronotum; similar but transverse “grooves” appear over basal third of prothorax). Dorsal pubescence inconspicuous, dense. Scutellar shield with broadly triangular posterior edge.
Elytra smooth dorsally, elongate, 2.4× longer than their maximum combined width and 1.5× as long as forebody (head and pronotum), slightly widened apically. Humeral angles widely rounded but distinct. Omoplates not indicated. Postbasal transverse impression shallow, present in basal third. Punctation irregular, quite dense and shallow. Pubescence uniform, moderately long and dense, suberect (possible that elytral pubescence is complex with setae of two different lengths, but this is not evident in the holotype). Metathoracic wings fully developed.
Legs long and slender. Tibiae straight, densely pubescent. Tibial spurs paired at least of meso- and metatibiae (possible due to the position of the inclusion, only one protibial spur visible), thin, long, not pubescent. Penultimate tarsomeres widened, bilobed. Basal metatarsomere longer than metatarsomeres 2-4 combined. Claws with blunt basal tooth.
Cucujiformia family incertae sedis
Petratypus nigri Kaddumi, 2005
Kaddumi, 2005 (new genus and species, initially assigned to Cleridae, Jordanian amber, 112.6-99.7 Ma); Kolibáč and Huang, 2016 (moved to Anthicidae).
Remarks. The holotype of this species was not available for our study. In the original description it is clearly stated “All tarsi 5-segmented” (Kaddumi, 2005: 128). There is nothing said in the description about presence or absence of anterior rim and antebasal sulcus on pronotum, but in figure 132 the holotype is depicted without any of them. Each elytron is said to be provided with “… 9-10 longitudinal rows of small punctures”. Tarsal claws as depicted on figure 130 in the original description are basally denticulate. Kolibáč and Huang (2016) proposed new placement for this species in Anthicidae considering “… the shape of the antennomeres and structure of the tarsi“. This proposal should be considered poorly justified since none of significant features of the Anthicidae and its subfamilies were assessed by the aforementioned two authors.
When the type of Petratypus nigri Kaddumi, 2005 is re-examined, the missing features have to be described. Based on the original description, tarsal formula of 5-5-5 in particular (tarsal formula is 5-5-4 in the Anthicidae and other Tenebrionoidea), we propose a new placement for this genus and species as a Familia incerta within the infraorder Cucujiformia Lameere, 1938.
New Fossil Record
Macratriinae tribe incertae sedis, genus sp.
Material examined. The examined specimen [ID “LDM G 337/35”] is labelled “MYANMAR Estado Cachin Valle Hukawng” / “MACRATRIINAE Tribe and Gen. sp. det. D.Telnov, 2018”.
One Burmese amber inclusion investigated during our study (kindly provided by F. Molino-Olmedo) contains a specimen of the Macratriinae. Unfortunately, due to the position of this specimen within amber piece and limited clearness of the amber, this specimen cannot be assigned to any of two Macratriinae tribes. Therefore, this species is not named and described here. Nevertheless, this is the oldest known Macratriinae record from Southeast Asia.
The following characters are recognizable on the specimen: eyes very large, round and strongly prominent, entire (not notched at insertions of antennae); tempora much shorter than diameter of an eye; head base rounded to subtruncate; head base shortly notched medially; antennae long with strongly lengthened three terminal antennomeres (terminal one is the longest) and shortened, distally angulate antennomeres 4-8; neck about 1/3 of head width; pronotum short and slightly transverse, anterior margin broadly rounded, laterally strongly constricted towards base, with narrow entire anterior rim and broad, deep antebasal sulcus; elytra elongate and parallel; tibial spurs paired; tibial spurs long, narrow and pointed, protarsi strongly widened; penultimate tarsomeres of all legs strongly widened and distinctly bilobate; tarsal claws simple.
The List of Fossil and Subfossil Records for Anthicidae and Ischaliidae
The list presented (Table 1) is produced as a result of bibliographical analysis and/or material examination. For each taxon the following information is given: Taxon (name, author(s) and year of description), Holotype depository (see Materials and methods for abbreviations), References, Fossil type and/or Locality information (as specified in cited source), Epoch and Age (according to Behrensmeyer and Turner, 2013-2018). In total, three Ischaliidae species of one genus and three subgenera and 17 Anthicidae species of 12 genera belonging to seven subfamilies are known from fossil deposits.
DISCUSSION
Brief Analysis of Known Fossil Records
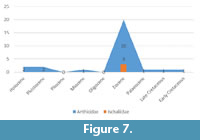 According to the summarised information (Table 1), fossil records are available for seven of eight Anthicidae subfamilies (88%). This information is detailed in Table 2. The oldest fossil representative of Anthicidae, Camelomorpha longicervix Kirejtshuk, Azar et Telnov (Macratriinae: Camelomorphini) was described from Earlier Cretaceous Lebanese amber (130.0-125.5 Ma), while the scarce fossil records of Ischaliidae come only from Upper Eocene Baltic amber (37.2-33.9 Ma).
According to the summarised information (Table 1), fossil records are available for seven of eight Anthicidae subfamilies (88%). This information is detailed in Table 2. The oldest fossil representative of Anthicidae, Camelomorpha longicervix Kirejtshuk, Azar et Telnov (Macratriinae: Camelomorphini) was described from Earlier Cretaceous Lebanese amber (130.0-125.5 Ma), while the scarce fossil records of Ischaliidae come only from Upper Eocene Baltic amber (37.2-33.9 Ma).
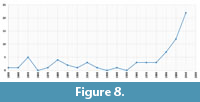
At present, known fossil records of Anthicidae and Ischaliidae from are most numerous from Eocene (Figure 7) and mostly originate from Baltic amber. The majority of these fossil records were published in the last two decades (Figure 8).
Biogeographic Analysis
The Ischaliidae, as well as subfamilies and tribes of the Anthicidae, have different distribution patterns in today’s zoogeographical regions (this account doesn’t includes two Chilean genera of uncertain placement, Apotominus Fairmaire et Germain, 1863 and Heterolobus Philippi in Philippi and Philippi, 1864 (see Chandler, 2010)). Most widespread and species rich are the Anthicinae (and Anthicini), which occur in all zoogeographical regions (Uhmann, 1976; Chandler, 2010). The Endomiini are restricted to Old World (only one species reaches Australia) as the Formicomini are (with one genus in Chile and several species in Australian and Papuan regions), while the Microhorini are known from the Palaearctic, Afrotropical and Oriental (except insular systems) regions. The monotypic Copobaeninae is a temperate South American group (Abdullah, 1969; Chandler, 2010). The subfamily Eurygeniinae is less diverse than the Anthicinae in terms of genera and species, but also reveals a cosmopolitan distribution (Chandler, 2010). Within Eurygeniinae, Eurygeniini are presently absent in Western Palaearctic, but known from here in fossil deposits (France). Representatives of Ictistygnini occur in Afrotropical, Australian and Neotropical regions, but the monotypic Mitraelabrini restricted to Chile. The Lemodinae currently distributed in Australian and Papuan regions and Chile, with fossil record from Oriental Region. The Macratriinae (and Macratriini) are cosmopolitan, while Camelomorphini only known from Western Palaearctic (Lebanese amber, fossil). The Notoxinae and Tomoderinae are cosmopolitan, with just a few genera in each group. Extant Steropinae are of Holarctic, Oriental, and Australian (Telnov, 2018) distribution.
 The present distribution of the Ischaliidae is restricted to mild climate areas of the Nearctic, E and S Palaearctic (Himalaya, China except North and West, Far East of Russia, Kurile and Japanese archipelagos, Korean Peninsula, and Taiwan) and Oriental (E India, most of continental Southeast Asia, and Greater Sunda Islands) regions (Gusakov and Telnov, 2007; Young, 2011; Alekseev and Telnov, 2016).
The present distribution of the Ischaliidae is restricted to mild climate areas of the Nearctic, E and S Palaearctic (Himalaya, China except North and West, Far East of Russia, Kurile and Japanese archipelagos, Korean Peninsula, and Taiwan) and Oriental (E India, most of continental Southeast Asia, and Greater Sunda Islands) regions (Gusakov and Telnov, 2007; Young, 2011; Alekseev and Telnov, 2016).
Fossil records illustrate that both ant-like flower beetles and false fire-coloured beetles have fossil representatives occurring outside their contemporary distribution ranges. Several studies have already demonstrated that distribution of numerous extant arthropod taxa differs significantly from their fossil distribution ranges (e.g., Eskov, 1987; 1992). The Western Palaearctic is not in the present distribution for extant Eurygeniini and Ischaliidae, but the Eocene fossil records discussed above (Table 1) illustrate that it was part of their former biogeographical range. The Lemodinae are not present in the Oriental Region today, but with Late Cretaceous record from this realm. Eskov (2002) already pointed out, that in certain cases, the extant distribution seems to be the relict of a formerly broader distribution, especially concerning arthropods and insects, in particular. The formerly wider distribution ranges of these taxa correlate well with worldwide warmer and uniform climate of Eocene (Frakes et al., 1992; Wedmann et al., 2011). Consequently, we want to emphasize that any interpretations of Coleoptera biogeography based only on extant taxa can be misleading, and therefore we strongly support similar conclusions provided earlier by Wedmann et al. (2011). The known historical biogeography of the Anthicidae and Ischaliidae is summarized in Table 3.
Discussion on Some Critical Features of the Anthicidae Subfamilies
As stated above and noted by earlier authors (e.g., Chandler, 2010), there has never been a phylogenetic analysis at the family level for Anthicidae. Abdullah (1969) presented some (18) of the critical characters and their states in the form of transformation series, dividing them into “primitive” and “derivative”. Nowadays these terms are out of methodology in phylogenetics and lack ground and explanations. We assessed selected morphological features (following Abdullah, 1967; 1969; Chandler, 2010; Telnov, 2018) from extant (text couplets below) and fossil taxa and highlighted minimum age of these features regarding data available from fossil material (Table 4). In the couplets below those features visible (available and fixed) both in fossil and extant taxa are marked with the dagger symbol (†) and enclosed into brackets, but those not yet fixed (e.g., not available or not visible due to the conditions of the fossil specimens) in fossil material but available in extant taxa are not specially marked.
Anthicinae. († head with frontoclypeal suture, which can be represented by an impression) or lacking in some subgroups; († eyes entire); († mandible apice bidentate) or unidentate; († neck narrow, smooth, 1/4 of head width or less); († antenna 11-segmented without modifications in most species); († terminal maxillary palpomere fusiform or triangular); († pronotum anteriorly with rim of equal width dorsally and ventrally); († antebasal sulcus distinct); († pronotum lacking an apical flange); mesoventrite triangular to transverse; mesepisterna separated from ventrite by distinct suture; procoxal cavities closed externally or internally; († metacoxae moderately to widely separated by strongly divergent intercoxal projection); († elytra with confused punctures in most subgroups); († tibial spurs small); († tarsal claws simple); († male genitalia usually with phallobase and tegmen distinct); ovipositor coxite 1-segmented or incompletely 2-segmented.
Eurygeniinae. († head lacking frontoclypeal suture); mandible apice usually scooped, broad; eyes emarginate in certain genera; († terminal antennomere not lengthened) or strongly lengthened (in Bactrocerus LeConte, 1866); († terminal maxillary palpomere securiform, axeform) or cultriform; († neck broad, 1/3-1/2 of head width); († pronotum anteriorly with dorsal rim strongly protruding / flanged), with ventral rim much narrower (except in Mitraelabrini Abdullah, 1969); male metasternum spinous in certain subgroups; procoxal cavities closed externally or internally; metacoxae narrowly separated by acute intercoxal projection; metathoracic wings with radial and anal cells present in some subgroups; († elytra with confused punctures); († abdominal ventrites simple) or with appendages in males of some Steriphodon Abeille de Perrin, 1895; tibial spurs long and distinct; tibial spurs microspinose or not; tibial spur formula 2-2-2; legs with ctenidia present; tarsal claws simple, appendiculate or with blunt basal tooth; ovipositor coxite incompletely 2-segmented.
Lemodinae. Head with frontoclypeal suture variably present, with transverse impression when lacking; († eyes entire); mandible apice bidentate; († antenna 11-) to 12-segmented without modifications; († neck broad, 1/3-1/2 of head width); pronotum with thin anterior rim; antebasal sulcus of pronotum only complete in Zealanthicus Werner et Chandler, 1995; procoxal cavities open externally and internally; mesoventrite triangular; mesoventrite discrimen weakly developed or absent; mesosternum and mesepisterna meet at the anterior edge of mesothorax; metacoxae moderately separated by divergent triangular intercoxal projection; († elytra with confused punctures) or arranged into rows; tibial spurs short; tibial spur formula 2-2-2; phallobase short and/or fused to tegmen.
Macratriinae. Peculiar within the Anthicidae primarily due to the mesoventrite fused to mesepisterna; († head lacking frontoclypeal suture); († mandible apice bidentate); († antenna 11-segmented); († 11th or 9-11th terminal antennomeres strongly lengthened and / or widened); († maxillary palpomeres 2-3 with angular mesal lobes); († terminal maxillary palpomere securiform, axeform) or cultriform; († neck narrow, 1/4 or less of head width); neck smooth; († pronotum with anterior collar of nearly equal width dorsally and ventrally); († antebasal sulcus distinct); († lateral fovea of pronotum extends anteroventrally at lateral margin towards procoxae towards a pit, which is situated just dorsal of procoxa); († lateral fovea and pits obscured by setation); († mesoventrite triangular, fused to mesepisterna); († procoxa comparatively short, ovoid in lateral view); († procoxal rests contiguous when available); († metacoxae narrowly separated by acute intercoxal projection); († elytra at least with tracks of puncture rows in most species); († tibial spurs long and distinct); tibial spur formula 2-2-2); († tarsal claws deeply cleft); ovipositor coxite 2-segmented.
Notoxinae. († head with frontoclypeal suture); († eyes entire in most species); († mandible apice bidentate); († antenna 11-segmented, without modifications); neck broad and short, 1/3-1/2 of head width; († pronotum with prominent anterodorsal horn and cervical notches lateral to neck); († antebasal sulcus of pronotum distinct); mesoventrite triangular; procoxal cavities closed externally or internally; metacoxae moderately to widely separated by divergent triangular intercoxal projection; († elytra with confused punctures); († tibial spurs distinct); tibial spur formula 2-2-2; tarsal claws simple; male genitalia with phallobase and separate parameres; ovipositor coxite 1-segmented or incompletely 2-segmented.
Steropinae. Considered “primitive” member of the Anthicidae by earlier authors (e.g., Abdullah, 1969) this subfamily shares many characters with Pedilinae (Pyrochroidae) and Eleticinae (Meloidae) (Chandler, 2010; Telnov, 2018). († Frontoclypeal suture) variably present, († with transverse impression when lacking); († eyes shortly notched at insertions of antennae) or entire; († maxillary palpomeres 2-3 apically rounded and slightly widened); († terminal maxillary palpomere scalene triangular); († antenna 11-segmented); († three terminal antennomeres strongly lengthened); († neck broad, 1/3-1/2 of head width); († pronotum dorsally without anterior rim; antebasal sulcus of pronotum complete); († lateral fovea present on pronotum, but ends in a pit distantly from procoxae); († lateral fovea and pits glabrous); († pronotum lacking an apical flange); mesepisterna meeting or almost meeting in front of mesosternite; heart-like sclerite separates procoxal rests; († procoxal cavities open externally) and internally; († procoxa elongate and truncate fusiform); († metacoxae nearly contiguous, narrowly separated by acute intercoxal projection); metathoracic wings with radial and anal cells present; elytra confusedly punctate; male elytra with a spot of very short and dense pubescence in basal third; († tibial spurs distinct); († tibial spur formula 2-2-2); († tibial spurs microspinose in some taxa); († tarsal claws each with long ventral spine) or shallowly toothed or medially lobed; aedeagus with distinct phallobase, penis, and separate parameres; ovipositor coxite 2-segmented.
Tomoderinae. († head with frontoclypeal suture variably present, with transverse impression when lacking); mandible apice bidentate; († antennae with a weak club of 3-4 terminal antennomeres); († neck narrow, 1/3 of head width or less); († pronotum with thin anterior rim); († antebasal sulcus only indicated laterally near hypomeron); († pronotum strongly constricted laterally postmedium), often notched in the constriction; anterior lobe denticulate or not on sides; († mesoventrite short, semicircular); († mesosternum and mesepisterna meet behind the anterior edge of mesothorax); († metacoxae widely separated); († tibial spurs very short to absent); male genitalia with phallobase usually fused to tegmen.
ACKNOWLEDGEMENTS
C. Mellish (BMNH) is sincerely thanked for providing access to the type material of “ Eurygenius ” wickhami. We are grateful to C. and H. Werner Hoffeins (Hamburg, Germany), C. Gröhn (Glinde, Germany) and F. Molino-Olmedo (Mancha Real (Jaén), Spain) for the loan of fossil material, to D.J. Mann (Oxford University Museum of Natural History, United Kingdom) for valuable help with obtaining some of references used for our study, to D.A. Pollock (Eastern New Mexico University, Portales, USA) and D.K. Young (University of Wisconsin, Madison, USA) for their valuable comments on “ Eurygenius ” wickhami. D.A. Pollock is also thanked for proofreading the manuscript.
REFERENCES
Abdullah, M. 1964. New heteromerous beetles (Coleoptera) from the Baltic amber of Eastern Prussia and gum copal of Zanzibar. Transactions of the Royal Entomological Society of London, 116:329-346.
Abdullah, M. 1965. New Anthicidae and Pyrochroidae (Coleoptera) from the Baltic amber (Oligocene). The Entomologist, 98:38-42.
Abdullah, M. 1967. Some phylogenetic conclusions on the Eurygeniinae (Coleoptera: Anthicidae), with a review of the North American species of Eurygenius including the description of a new species (E. darlingtoni) from Texas. Entomological News, 78:180-188.
Abdullah, M. 1969. The natural classification of the family Anthicidae with some ecological and ethological observations. Deutsche Entomologische Zeitschrift, 16:323-366.
Abdullah, M. 1975. The higher Classification of the insect order Coleoptera including fossil records and a classified directory of the coleopterists and Coleoptera collections of the world. Zoologische Beiträge, Neue Folge, 21:363-461.
Alekseev, V.I. 2013. The beetles (Insecta: Coleoptera) of Baltic amber: the checklist of described species and preliminary analysis of biodiversity. Zoology and Ecology, 23:5-12. https://doi.org/10.1080/21658005.2013.769717
Alekseev, V.I. 2017. Coleoptera from the middle-upper Eocene European ambers: generic composition, zoogeography and climatic implications. Zootaxa, 4290:401-443. https://doi.org/10.11646/zootaxa.4290.3.1
Alekseev, V.I. and Bukejs, A. 2017. New fossil taxa of Ischalia Pascoe (Coleoptera: Ischaliidae) from Eocene Baltic amber. Zootaxa, 4323:229-238. https://doi.org/10.11646/zootaxa.4323.2.6/10.11646/zootaxa.4323.2.6
Alekseev, V.I. and Telnov, D. 2016. First fossil record of Ischaliidae Blair, 1920 (Coleoptera) from Eocene Baltic amber. Zootaxa, 4109:595-599. https://doi.org/10.11646/zootaxa.4109.5.8/10.11646/zootaxa.4109.5.8
Arillo, A. and Ortuño, V.M. 1997. First records of the families Anthicidae and Chrysomelidae from the Oligocene of Izarra (Alava, Spain). Coleopterists' Bulletin, 51:293-297.
Azar, D., Geze, R., and Acra, F. 2010. Chapter 14. Lebanese amber, pp. 271-298. In Penney, D. (ed.), Biodiversity of Fossils in Amber from the Major World Deposits. Siri Scientific Press, Manchester.
Bachofen-Echt, A. 1949. Der Bernstein und seine Einschlüsse. Springer Verlag, Wien.
Batelka, J., Kundrata, R., and Bocak, L. 2016. Position and relationships of Ripiphoridae (Coleoptera: Tenebrionoidea) inferred from ribosomal and mitochondrial molecular markers. Annales Zoologici, 66:113-123.
Behrensmeyer, A.K. and Turner, A. 2013-2018. Taxonomic occurrences of Suidae recorded in the Paleobiology Database. Fossilworks. Updated at: http://fossilworks.org, accessed 6 February 2018.
Berendt, G.C. 1845. Die im Bernstein befindlichen organischen Reste der Vorwelt gesammelt in Verbindung mit mehreren bearbeitet. Erster Band. Nikolaische Buchhandlung, Danzig.
Bidashko, F.G., Proskurin, K.P. and Shatrovskii, A.G. 1995. Paleogeography of the lower Volga at the end of Atelian time based on entomological and botanical data. Paleontological Journal, 30:332-336.
Bologna, M.A., Turco, F. and Pinto, J.D. 2010. 11.19. Meloidae Gyllenhal, 1910, p. 681-693. In Leschen, R.A.B., Beutel, R.G., and Lawrence, J.F. (eds.), Coleoptera, Beetles. Volume 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim). Handbook of Zoology. Arthropoda: Insecta. De Gruyter, Berlin and New York.
Brasero, N., Nel, A., and Michez, D. 2009. Insects from the Early Eocene amber of Oise (France): diversity and palaeontological significance. Denisia, 26 (zugleich Kataloge der Oberösterreichischen Landesmuseen, Neue Serie, 86:41-52.
Chandler, D.S. 2010. 11.26. Anthicidae Latreille, 1819, pp. 729-741. In Leschen, R.A.B., Beutel, R.G., and Lawrence, J.F. (eds.), Coleoptera, Beetles. Volume 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim). Handbook of Zoology. Arthropoda: Insecta. De Gruyter, Berlin and New York.
Chandler, D.S., Uhmann, G., Nardi, G., and Telnov, D. 2008. Family Anthicidae Latreille, 1819, p. 421-455. In Löbl, I. and Smetana, A. (eds.), Catalogue of Palaearctic Coleoptera. Volume 5. Apollo Books, Stenstrup.
Cockerell, T.D.A. 1917. Insects in Burmese amber. Annals of the Entomological Society of America, 10:323-329.
Cockerell, T.D.A. 1926. Some Tertiary fossil insects. The Annals and Magazine of Natural History, 18:313-324.
Cockerell, T.D.A. 1936. The fauna of the Sunchal (or Margas Verdes) formation, northern Argentina. American Museum Novitates, 886:1-9.
Coope G.R., Shotton, F.W., Strachan, I., and Dance, S.P. 1961. A Late Pleistocene fauna and flora from Upton Warren, Worcestershire. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 244:379-421.
de Motschulsky, V.M. 1856. Lettres de M. de Motschulsky à M. Ménétriés. Études Entomologique s, 5:3-38.
Eskov, K.Y. 1987. A new archaeid spider (Chelicerata: Araneae) from the Jurassic of Kazakhstan, with notes on the so-called “Gondwanan” ranges of recent taxa. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 175:81-106.
Eskov, K.Y. 1992. Archaeid spiders from Eocene Baltic amber (Chelicerata: Araneida: Archaeidae) with remarks on the so-called “Gondwanan” ranges of recent taxa. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 185:311-328.
Eskov, K.Y. 2002. Geographical history of insects, p. 427-435. In Rasnitsyn, A.P. and Quicke, D.L.J. (eds.), History of Insects. Kluwer Academic Publishers, Dordrecht.
Fletcher, T.B. 1920. Indian fossil insects. Report of the Proceedings of the Third Entomological Meeting, Pusa, 1919, 3:983-990.
Frakes, L.A., Francis, J.E., and Sykes, J.I. 1992. Climate Modes of the Phanerozoic. Cambridge University Press, Cambridge.
Giebel, C.G. 1852. Deutschlands Petrefakten. Ein systematisches Verzeichniss aller in Deutschland und den angrenzenden Ländern vorkommenden Petrefacten nebst Angabe der Synonyme und Fundorte. Abel, Leipzig.
Giebel, C.G. 1856a. Fauna der Vorwelt mit steter Beruecksichtigung der lebenden Thiere. Monographisch dargestellt. Zweiter Band: Gliederthiere. Erste Abtheilung: Insecten und Spinnen. Die Insecten und Spinnen der Vorwelt mit steter Berücksichtigung der lebenden Insecten und Spinnen. F. A. Brockhaus, Leipzig.
Giebel, C.G. 1856b. Geologische Übersicht der vorweltlichen Insecten. Zeitschrift für die gesamten Naturwissenschaften, 8:174-188.
Green, C.P., Branch, N.P., Coope, G.R., Field, M.H., Keen, D.H., Wells, J.M., Schwenninger, J.L., Preece, R.C., Schreve, D.C., Canti, M.G., and Gleed-Owen, C.P. 2006. Marine Isotope Stage 9 environments of fluvial deposits at Hackney, north London, UK. Quaternary Science Reviews, 25:89-113. https://doi.org/10.1016/j.quascirev.2004.10.011/10.1016/j.quascirev.2004.10.011
Guo, M., Xing, L., Wang, B., Zhang, W., Wang, S., Shi, A., and Bai, M. 2017. A catalogue of Burmite inclusions. Zoological Systematics, 42:249-379.
Gusakov, A.A. and Telnov, D. 2007. Systematic changes and new species of Ischaliidae (Coleoptera). Folia heyrovskyana, Series A, 15:39-46.
Handlirsch, A. 1907. IV. Abschnitt. Tertiäre Insekten, p. 673-1120. In Handlirsch. A. (ed.), 1906-1908. Die Fossilen Insekten und die Phylogenie der Rezenten Formen: Ein Handbuch für Paläontologen und Zoologen. Wilhelm Engelman, Leipzig.
Helm, O. 1886. Mittheilungen über Bernstein. XIII. Über die Insecten des Bernsteins. Schriften der Naturforschenden Gesellschaft in Danzig, 6:267-277.
Helm, O. 1896. Beiträge zur Kenntniss der Insecten des Bernsteins. Schriften der Naturforschenden Gesellschaft in Danzig, Neue Folge, 9:220-231.
Hieke, F. and Pietrzeniuk, E. 1984. Die Bernstein-Käfer des Museums für Naturkunde, Berlin (Insecta, Coleoptera). Mitteilungen aus dem Zoologischen Museum Berlin, 60:297-326.
Hope, F.W. 1836. XXX. Observation on succinic insects. The Transactions of the Entomological Society of London, 1 (1834-1836), part 3:133-147.
Ivie, M.A. 2002. Keys to families of beetles in America north of Mexico, pp. 816-835. In Arnett R.H. Jr., Thomas, M.C., Skelley, P.E., and Frank, J.H. (eds.), American Beetles. Volume 2. Polyphaga: Scarabaeoidea through Curculionoidea. CRC Press, Boca Raton.
Kaddumi, H.F. 2005. Amber of Jordan: the Oldest Prehistoric Insects in Fossilized Resin. Publication of Eternal River Museum of Natural History, Amman.
Keilbach, R. 1982. Bibliographie und Liste der Arten tierischer Einschlüße in fossilen Harzen sowie ihrer Aufbewahrungsorte. Deutsche Entomologische Zeitschrift, 29:129-286, 301-391.
Kirejtshuk, A.G. 2002-2013. Catalogue of Fossil Coleoptera. Updated at: http://www.zin.ru/Animalia/Coleoptera/eng/paleosys.htm, accessed 27 December 2017.
Kirejtshuk, A.G. and Azar, D. 2008. New taxa of beetles (Insecta, Coleoptera) from Lebanese amber with evolutionary and systematic comments. Alavesia, 2:15-46.
Kirejtshuk, A.G. and Azar, D. 2013. Current knowledge of Coleoptera (Insecta) from the Lower Cretaceous Lebanese amber and taxonomical notes for some Mesozoic groups. Terrestrial Arthropod Reviews, 6:103-134. https://doi.org/10.1163/18749836-06021061
Kirejtshuk, A.G. and Nel, A. 2008. New beetles of the suborder Polyphaga from the lowermost Eocene French amber (Insecta: Coleoptera). Annales de la Société Entomologique de France (n. s.), 44:419-442. https://doi.org/10.1080/00379271.2008.10697578/10.1080/00379271.2008.10697578
Kirejtshuk, A.G. and Nel, A. 2013. Current knowledge of Coleoptera (Insecta) from the lowermost Eocene Oise amber. Insect Systematics and Evolution, 44:175-201. https://doi.org/10.1163/1876312x-04402007
Kirejtshuk, A.G., Ponomarenko, A.G., Kurochkin, A.S., Alexeev, A.V., Gratshev, V.G., Solodovnikov, A.Yu., Krell, F.-T., Soriano, C. 2019. The beetle (Coleoptera) fauna of the Insect Limestone (late Eocene), Isle of Wight, southern England. Earth and Environmental Science Transactions of the Royal Society of Edinburgh:1-88. https://doi.org/10.1017/S1755691018000865
Kiselev, S.V. and Nazarov, V.I. 2009. Late Cenozoic insects of northern Eurasia. Paleontological Journal, 43:732-850. https://doi.org/10.1134/s0031030109070016/10.1134/s0031030109070016
Klebs, R. 1889. Aufstellung und Katalog des Bernstein-Museums von Stantien and Becker, Königsberg i. Pr. nebst einer kurzen Geschichte des Bernsteins. Fiartung, Königsberg.
Klebs, R. 1910. Über Bernsteineinschlüsse in allgemeinen und die Coleopteren meiner Bernsteinsammlung. Schriften der Physikalisch-ökonomischen Gesellschaft zu Königsberg, 51:217-242.
Kolibáč, J. and Huang, D.Y. 2016. The oldest known clerid fossils from the Middle Jurassic of China, with a review of Cleridae systematics (Coleoptera). Systematic Entomology, 41:808-823. https://doi.org/10.1111/syen.12192/10.1111/syen.12192
Kubisz, D. 2000. Fossil beetles (Coleoptera) from Baltic amber in the collection of the Museum of Natural History of ISEA in Krakow. Polskie Pismo Entomologiczne, 69:225-230.
Kulicka, R. and Ślipiński, S.A. 1996. A review of the Coleoptera inclusions in the Baltic amber. Prace Muzeum Ziemi, 44:5-11.
Larsson, S.G. 1978. Baltic Amber - A Palaeobiological Study. Entomonograph 1. Scandinavian Science Press Ltd., Klampenborg, Denmark.
Lawrence, J.F., Escalona, H., and Leschen, R.A.B. 2010. 11.29. Tenebrionoidea Incertae sedis, p. 750-760. In Leschen, R.A.B., Beutel, R.G., and Lawrence. J.F. (eds.), Coleoptera, Beetles. Volume 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim). Handbook of Zoology. Arthropoda: Insecta. De Gruyter, Berlin and New York.
Lawrence, J.F. and Ślipiński, S.A. 2010. 11.17. Stenotrachelidae C.G. Thomson, 1859, p. 670-674. In Leschen, R.A.B., Beutel, R.G., and Lawrence. J.F. (eds.), Coleoptera, Beetles. Volume 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim). Handbook of Zoology. Arthropoda: Insecta. De Gruyter, Berlin and New York.
Lawrence, J.F. and Ślipiński, S.A. 2013. Australian beetles. Volume 1. Morphology, Classification and Keys. CSIRO Publishing, Collingwood.
Menge, A. 1856. Lebenszeichen vorweltlicher im Bernstein eingeschlossener Thiere. Programm Petrischule. A.W. Kafemann, Danzig.
Molino-Olmedo, F. 2017. Descripcion de Lemodicarmenia olmedoae gen. et sp. nov. del ámbar cretácico de Myanmar (Coleoptera, Anthicidae, Lemodinae). Lambillionea, 117:121-124.
Nel, A. and Brasero, N. 2010. Chanter 8. Oise amber, pp. 137-148. In Penney, D. (ed.), Biodiversity of Fossils in Amber from the Major World Deposits. Siri Scientific Press, Manchester.
Oustalet, É. 1874. Recherches sur les insectes fossiles des terrains Tertiaires de la France. 2 partie: Insectes fossiles d’Aix en Provence. Annales des Sciences Géologiques, 5:1-347.
Peris, D., Ruzzier, E., Perrichot, V., and Delclòs, X. 2016. Evolutionary and paleobiological implications of Coleoptera (Insecta) from Tethyan-influenced Cretaceous ambers. Geoscience Frontiers, 7:695-706. https://doi.org/10.1016/j.gsf.2015.12.007/10.1016/j.gsf.2015.12.007
Perkovsky, E.E., Rasnitsyn, A.P., Vlaskin, A.P., and Taraschuk, V. 2007. A comparative analysis of the Baltic and Rovno amber arthropod faunas: representative samples. African Invertebrates, 48:229-245.
Perkovsky, E.E., Zosimovich, V.Y., and Vlaskin, A.P. 2010. Chapter 7: Rovno amber, p. 116-136. In Penney, D. (ed.), Biodiversity of Fossils in Amber from the Major World Deposits. Siri Scientific Press, Manchester.
Poinar, G.O. Jr. 1992. Life in Amber. Stanford University Press, Stanford.
Pollock, D.A. 2010. 11.20. Mycteridae Blanchard, 1845, pp. 693-699. In Leschen, R.A.B., Beutel, R.G. and Lawrence, J.F. (eds), Coleoptera, Beetles. Volume 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim). Handbook of Zoology. Arthropoda: Insecta. De Gruyter, Berlin and New York.
Rasnitsyn, A.P. and Ross, A.J. 2000. A preliminary list of arthropod families present in the Burmese amber collection at The Natural History Museum, London. Bulletin of the Natural History Museum, Geology Series, 56:21-24.
Ritzkowski, S. 1990. Die Inklusen der ehemaligen Königsberger Bernsteinsammlung in Göttingen. Prace Museum Ziemi, 41:149-153.
Ross, A.J. 1998. Amber, the Natural Time Capsule. The Natural History Museum, London.
Ross, A.J. and York, P.V. 2000. A list of type and figured specimens of insects and other inclusions in Burmese amber. Bulletin of the Natural History Museum, Geology Series, 56:11-20.
Ross, A., Mellish, C., York, P., and Crighton, B. 2010. Chapter 12 Burmese amber, p. 208-235. In Penney, D. (ed.), Biodiversity of Fossils in Amber from the Major World Deposits. Siri Scientific Press, Manchester.
Schawaller, W. 1986. Fossile Käfer aus miozänen Sedimenten des Randecker Maars in Südwest-Deutschland (Insecta: Coleoptera). Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie), 126:1-9.
Scudder, S.H. 1885. Systematische Übersicht der fossilen Myriapoden, Arachnoideen und Insekten, p. 721-831. In Zittel, K.A. (ed.), Handbuch der Palaeontologie, I. Abtheilung. Palaeozoologie, Band II. München, Leipzig, Oldenbourg.
Scudder, S.H. 1886. Systematic review of our present knowledge of fossil insects including myriapods and arachnids. Bulletin of the United States Geological Survey, 31:1-128.
Scudder, S.H. 1891. Index to the known fossil insects of the world including myriapods and arachnids. Bulletin of the United States Geological Survey, 71:1-744.
Soriano, C., Pollock, D., Néraudeau, D., Nel, A., and Tafforeau, P. 2014. First fossil record of polypore fungus beetles from Lower Cretaceous amber of France. Acta Palaeontologica Polonica, 59:941-946. https://doi.org/10.4202/app.2012.0074/10.4202/app.2012.0074
Spahr, U. 1981. Systematischer Katalog der Bernstein- und Kopal-Käfer (Coleoptera). Stuttgarter Beiträge zur Naturkunde (Ser. B), 80:1-107.
Tarasov, S., Vaz-de-Mello, F.Z., Krell F.-T., and Dimitrov, D. 2016. A review and phylogeny of scarabaeine dung beetle fossils (Coleoptera: Scarabaeidae: Scarabaeinae), with the description of two Canthochilum species from Dominican amber. PeerJ:1-35, 4:e1988. https://doi.org/10.7717/peerj.1988/10.7717/peerj.1988
Telnov, D. 1999. Anthicus oustaleti Telnov, 1999 nom. nov., a new name for Anthicus melancholicus Oustalet, 1874 nec A. melancholicus LaFerté-Sénectere, 1848 (Coleoptera: Anthicidae). Mitteilungen des Internationalen Entomologischen Vereins e.V., 24:61-62.
Telnov, D. 2006. The first record of Steropinae from the Oriental region, with the description of a new Steropes from Vietnam (Coleoptera: Anthicidae). Journal of the Zoological Society Wallacea, 2:63-66.
Telnov, D. 2008. Order Coleoptera, family Anthicidae, pp. 270-292. In van Harten, A. (ed.), Arthropod Fauna of the UAE. Volume 1. Dar Al Ummah Printing, Publishing, Distribution and Advertising, Abu Dhabi.
Telnov, D. 2012a. Tomoderinae (Coleoptera: Anthicidae) of the Baltic amber. Latvijas Entomologs, 51:3-11.
Telnov, D. 2012b. Macratriinae (Coleoptera: Anthicidae) of the Baltic amber. Latvijas Entomologs, 51:28-40.
Telnov, D. 2013. A new species of Tomoderinae (Coleoptera: Anthicidae) from the Baltic amber. Latvijas Entomologs, 52:130-134.
Telnov, D. 2018. Descriptions of two new Australian genera of Anthicidae (Insecta: Coleoptera). Baltic Journal of Coleopterology, 18(2):199-218.
Telnov, D. and Ghahari, H. 2018. An annotated checklist of the Anthicidae and pediline Pyrochroidae (Insecta: Coleoptera) of Iran, with thirteen new country records. Zootaxa, 4497(4):451-491. https://doi.org/10.11646/zootaxa.4497.4.1
Uhmann, G. 1976. Die Gattungen der Anthicidae und ihre systematische Anordnung. Beitrag zur Kenntnis der Anthicidae (Coleoptera). Entomologische Blätter, 72:166-182.
Wang, B. and Zhang, H. 2011. The oldest Tenebrionoidea (Coleoptera) from the Middle Jurassic of China. Journal of Paleontology, 85:266-270. https://doi.org/10.1666/09-088.1/10.1666/09-088.1
Wedmann, S., Hörnschemeyer, T., and Schmied, H. 2011. Fossil water-penny beetles (Coleoptera: Psephenidae: Eubrianacinae) from the Eocene of Europe, with remarks on their phylogenetic position and biogeography. Palaeontology, 54:965-980. https://doi.org/10.1111/j.1475-4983.2011.01088.x/10.1111/j.1475-4983.2011.01088.x
Wickham, H.F. 1910. New fossil Coleoptera from Florissant, with notes on some already described. American Journal of Science, Series 4, 29:47-51.
Young, D.K. 2005. Taxonomic notes on South American Pogonoceromorphus Pic (Coleoptera), including transfer from Pyrochroidae (Pyrochroinae) to Anthicidae (Eurygeniinae). Proceedings of the Entomological Society of Washington, 107:357-361.
Young, D.K. 2011. A new Asian subgenus and species of Ischalia (Coleoptera: Ischaliidae) with an assessment of subgeneric concepts, revised world checklist, and keys to the subgenera and “blue elytra” species. Zootaxa, 2811:53-58.
Young, D.K. and Pollock, D.A. 1991. Description of the mature larva of Pedilus flabellatus (Coleoptera: Pyrochroidae: Pedilinae), with phylogenetic implications of the discovery. Canadian Journal of Zoology, 69:2234-2238.
Young, D.K. and Pollock, D.A. 2010. 11.24. Pyrochroidae Latreille, 1807, p. 715-721. In Leschen, R.A.B, Beutel, R.G., and Lawrence, J.F. (eds.), Coleoptera, Beetles. Volume 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim). Handbook of Zoology. Arthropoda: Insecta. De Gruyter, Berlin and New York.
Zherikhin, V.V. 1980. Class Insecta. Nasekomye [Class Insecta. Insects], p. 40-97. In Menner, V.V. (ed.), Razvitie i smena bespozvonochnykh na rubezhe Mezozoya i Kainozoya. Mshanki, chlenistonogie, iglokozhye. [Development and changes of invertebrates at the Mesozoic / Cainozoic boundary. Bryozoans, arthropods, echinoderms]. Nauka, Moscow. (In Russian)
