

 A reconsideration of the palinuroid family Synaxidae (Crustacea, Decapoda), with a new member from the Upper Jurassic of southern Poland
A reconsideration of the palinuroid family Synaxidae (Crustacea, Decapoda), with a new member from the Upper Jurassic of southern Poland
Article number: 26.2.19
https://doi.org/10.26879/1252
Copyright Palaeontological Association, June 2023
Proceedings of the 8th Symposium on Fossil Decapod Crustaceans
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 3 November 2022. Acceptance: 25 May 2023.
ABSTRACT
Representatives of the family Synaxidae, commonly referred to as furry lobsters, are rare constituents of modern-day marine communities, while their fossil record suggests that they were more common in the past, in reefal settings during the Late Jurassic across Europe, from where at least three species have been recorded to date. An overview of all known extant and extinct synaxids is presented here and a sixth fossil form is added to the list. The latter constitutes one of the earliest records to date of furry lobsters worldwide and extends the palaeogeographical range of this relatively small group of early palinuroids. It was collected from massive sponge-microbial build-ups (reefal limestones) of middle Oxfordian age (Gregoryceras transversarium ammonite Zone) near Kraków, southern Poland, and is here named Palaeosynaxes montserratae nov. gen., nov. sp.
René H.B. Fraaije, Oertijdmuseum, Bosscheweg 80, 5283 WB Boxtel, The Netherlands. directie@oertijdmuseum.nl (corresponding author)
Barry W.M. van Bakel, Oertijdmuseum, Bosscheweg 80, 5283 WB Boxtel, The Netherlands, and Department of Earth Sciences, Faculty of Geosciences, Utrecht University, Budapestlaan, 4, 3584 CD Utrecht, The Netherlands. b.w.m.vanbakel@uu.nl
John W.M. Jagt, Natuurhistorisch Museum Maastricht, De Bosquetplein 6-7, 6211 KJ Maastricht, The Netherlands. john.jagt@maastricht.nl
Michał Krobicki. AGH University of Science and Technology, Mickiewicza 30, 30-059 Kraków, Poland. krobicki@agh.edu.pl
Àlex Ossó. Llorenç de Villalonga, 17B, 1-1, 43007 Tarragona, Catalonia, Spain. aosso@tinet.cat
Ferran Palero, Institut Cavanilles de Biodiversitat i Biologia Evolutiva (ICBIBE), Carrer del Catedrátic José Beltrán Martinez, 2, 46980 Paterna, Valencia. Ferran.Palero@uv.es
Jonathan J.W. Wallaard, Oertijdmuseum, Bosscheweg 80, 5283 WB Boxtel, The Netherlands. curator@oertijdmuseum.nl
Keywords: Reptantia; Achelata; spiny lobsters; new taxa; Europe
Final citation: Fraaije, René H.B., van Bakel, Barry W.M., Jagt, John W.M., Krobicki, Michał, Ossó, Àlex, Palero, Ferran, and Wallaard, Jonathan J.W. 2023. A reconsideration of the palinuroid family Synaxidae (Crustacea, Decapoda), with a new member from the Upper Jurassic of southern Poland. Palaeontologia Electronica, 26(2):a19.
https://doi.org/10.26879/1252
palaeo-electronica.org/content/2023/3859-synaxidae-and-new-genus
Copyright: June 2023 Palaeontological Association.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/7E4423BE-D292-4407-919F-6A353AD251B3
INTRODUCTION
Furry lobsters (or coral lobsters) are small decapod crustaceans that are closely related to slipper lobsters (Scyllaridae) and spiny lobsters (Palinuridae). However, synaxids differ from spiny and slipper lobsters in lacking pronounced supra-orbital horns and in having broad-based, pronounced triangular rostra, while their body is covered in short hairs (hence the name ‘furry lobster’), and their antennae are not as enlarged as in scyllarids and palinurids. In modern-day seas, representatives of the family Synaxidae are rare constituents of benthic communities (e.g., Holthuis, 1991), but from the fossil record they were more common in the past, being known in particular from reefal settings of Late Jurassic age across Europe (Bachmayer, 1959; Förster, 1973; Fraaije et al., 2020). We herein revise the three taxa recorded to date from such levels, as well as younger forms from the mid-Cretaceous (Garassino and Pasini, 2020) and Upper Eocene (De Angeli and Garassino, 2014), and erect a new genus and species for a form from the Upper Jurassic of southern Poland.
GEOLOGICAL SETTINGS
 A road cutting into a small hill near the motorway from Kraków to Katowice, close to the village of Szklary in southern Poland, has been well known for the presence of numerous remains of middle Oxfordian decapod crustaceans (Krobicki, 1994; Krobicki and Müller, 1998a, b; Müller et al., 2000; Garassino and Krobicki, 2002; Krobicki and Zatoń, 2008; Fraaije et al., 2022).
A road cutting into a small hill near the motorway from Kraków to Katowice, close to the village of Szklary in southern Poland, has been well known for the presence of numerous remains of middle Oxfordian decapod crustaceans (Krobicki, 1994; Krobicki and Müller, 1998a, b; Müller et al., 2000; Garassino and Krobicki, 2002; Krobicki and Zatoń, 2008; Fraaije et al., 2022).  Palaeogeographically, the limestones that crop out in a small, abandoned quarry in a forest-lined valley close to Szklary belong to the Late Jurassic sponge megafacies (Matyja and Wierzbowski, 1995) of the peri-Tethyan epicontinental sea across Europe, from Portugal to Romania. Oxfordian-aged sponge bioconstructions predominated within these facies. Sedimentologically, these have been interpreted as cyanobacterial-sponge buildups (bioherms/“reefs”) that constructed massive, non-bedded limestones (Figure 1). They are surrounded by micritic, thin-bedded platy-type limestones that primarily formed in inter- and/or peri-bioherm environments (Matyszkiewicz, 1999; Krobicki and Zatoń, 2008; Matyszkiewicz et al., 2012) on a deep-neritic sea floor (Matyja and Wierzbowski, 1995). These limestones are well dated by ammonites (e.g., Matyja and Wierzbowski, 1995), also in the Kraków area in southern Poland (Głowniak, 2006; Matyja and Ziółkowski, 2014) (Figure 2).
Palaeogeographically, the limestones that crop out in a small, abandoned quarry in a forest-lined valley close to Szklary belong to the Late Jurassic sponge megafacies (Matyja and Wierzbowski, 1995) of the peri-Tethyan epicontinental sea across Europe, from Portugal to Romania. Oxfordian-aged sponge bioconstructions predominated within these facies. Sedimentologically, these have been interpreted as cyanobacterial-sponge buildups (bioherms/“reefs”) that constructed massive, non-bedded limestones (Figure 1). They are surrounded by micritic, thin-bedded platy-type limestones that primarily formed in inter- and/or peri-bioherm environments (Matyszkiewicz, 1999; Krobicki and Zatoń, 2008; Matyszkiewicz et al., 2012) on a deep-neritic sea floor (Matyja and Wierzbowski, 1995). These limestones are well dated by ammonites (e.g., Matyja and Wierzbowski, 1995), also in the Kraków area in southern Poland (Głowniak, 2006; Matyja and Ziółkowski, 2014) (Figure 2).
ACHELATAN MORPHOLOGY AND CLASSIFICATION
We here follow the classification of achelatan lobsters as based on carapace morphology (e.g., Holthuis, 1991; Martin and Davis, 2001; Lavalli and Spanier, 2010; Schweitzer et al., 2015), in which four families are recognised, namely the Cancrinidae, Palinuridae, Scyllaridae, and Synaxidae. Extant synaxids comprise only two genera: Palibythus Davie, 1990 and Palinurellus von Martens, 1878. Synaxids are differentiated from palinurids in that they lack pronounced supra-orbital horns. Moreover, they have short antennal flagella, a long and broad triangular anterior part of the carapace (i.e., rostrum to cervical groove), a prominent, broad-based rostrum and a tapered carapace that is not depressed or flattened, but tubular. They also have more robust first pereiopods than most extant achelatans. All these features may be considered ancestral in the evolutionary history of this group of decapod crustaceans. This observation is corroborated by data obtained in a detailed study of the morphology of achelatan larvae carried out by Baisre (1994), who demonstrated that Palinurellus was the most basal amongst extant palinuroids.
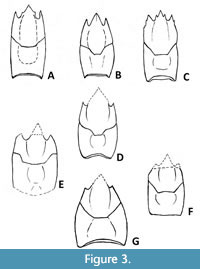 Within the family Synaxidae (Figure 3), only three extant species have been recorded to date. The first, Palibythus magnificus Davie, 1990 (Figure 3C), is known from depths between 220 and 275 metres near western Samoa in the Pacific Ocean. The other two, Palinurellus gundlachi von Martens, 1878 (Figure 3A; Caribbean Sea and Atlantic coast of South America) and P. wienecki (De Man, 1881) (Figure 3B; Indo-Pacific), both of which inhabit shallow-water reef environments at depths between 9 and 35 metres, occurring deep within holes and crevices, and apparently active only at night, which explains why they have only rarely been collected (Holthuis, 1966, 1991; Davie, 1990; Lavalli and Spanier, 2010; Idreesbabu et al., 2018).
Within the family Synaxidae (Figure 3), only three extant species have been recorded to date. The first, Palibythus magnificus Davie, 1990 (Figure 3C), is known from depths between 220 and 275 metres near western Samoa in the Pacific Ocean. The other two, Palinurellus gundlachi von Martens, 1878 (Figure 3A; Caribbean Sea and Atlantic coast of South America) and P. wienecki (De Man, 1881) (Figure 3B; Indo-Pacific), both of which inhabit shallow-water reef environments at depths between 9 and 35 metres, occurring deep within holes and crevices, and apparently active only at night, which explains why they have only rarely been collected (Holthuis, 1966, 1991; Davie, 1990; Lavalli and Spanier, 2010; Idreesbabu et al., 2018).
Previously, synaxids were interpreted either as sister group to all other achelatans or as an in-group of the Palinuridae (e.g., Davie, 1990; George, 2006; Palero et al., 2009). In recent years, an increasing number of extinct (Late Jurassic to Late Eocene; Figure 3D-E, G) synaxids have been published (De Angeli and Garassino, 2014; Fraaije et al., 2020; Garassino and Pasini, 2020), demonstrating that furry lobsters were more diverse in late Mesozoic and Paleogene marine ecosystems than they are at the present day.
Palibythus was placed by Davie (1990) in the Palinuridae on account of the presence of a well-developed stridulatory organ. Although Palinurellus lacks such an organ, Davie assigned this genus to the Palinuridae as well, interpreting it as a primitive member.
The first extinct species of Palinurellus, from the Upper Eocene (Priabonian) of north-east Italy, was recorded by De Angeli and Garassino (2014) as P. bericus (Figure 3D). The Late Jurassic genus Palaeopalinurellus (Fraaije et al., 2020) differs from Palinurellus in the scabrous nature of the ornament of its posterior carapace, the irregular tuberculation of its anterior carapace, and the more angular course of its cervical groove.
Based on a single carapace, Bachmayer (1959, p. 938, pl. 1, fig. 1) recorded a new palinurid, Palinurus strambergensis (Figure 3F), from ‘grauer Kalk, ob. Malm, Stramberger Schichten’ (i.e., Tithonian, Upper Jurassic) at Štramberk (Moravia, Czech Republic). Because of its overall morphology, Palinurus strambergensis has recently been transferred to Palaeopalinurellus by Fraaije et al. (2020). This species is also known from Kimmeridgian strata in southern Germany (Förster, 1973).
SYSTEMATIC PALAEONTOLOGY
Infraorder ACHELATA Scholtz and Richter, 1995
Superfamily PALINUROIDEA Latreille, 1802
Family SYNAXIDAE Bate, 1881
Genus Palaeosynaxes nov.
zoobank.org/F8716888-2DED-4C4A-B199-2CC638D45C6E
Type species. Palaeosynaxes montserratae nov. sp.
Etymology. From the Greek palaios for ‘old’, in reference to this being an early member of the family Synaxidae.
Diagnosis. Carapace small, oval, tubular; cervical groove V-shaped centrally; carapace densely covered with coarse, blunt, forwardly directed tubercles, most robust on two postrostral ridges on anterior part.
Palaeosynaxes montserratae nov. gen., nov. sp.
Figure 4
zoobank.org/23EEB842-EF06-4A28-8DE9-D5EDCFA00C5D
Diagnosis. As for genus.
Repository of the type specimen. MAB - Oertijdmuseum, Boxtel, The Netherlands.
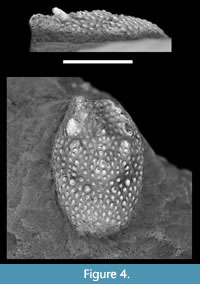
Type material. The holotype MAB13942, and sole specimen known to date, is an incomplete carapace with a maximum preserved length of 9.5 mm and a maximum preserved width of 6.0 mm, collected in June 2013 by one of us (À.O.) during a field trip to the 5th Symposium on Mesozoic and Cenozoic Decapod Crustaceans, Kraków (Poland).
Etymology. Dedicated to Montserrat Batet, partner of one of us (À.O.), as a token of thanks for her support and understanding.
Locality and stratigraphy. A road cutting on a small hill close to the village of Szklary, southern Poland, exposing upper middle Oxfordian (Gregoryceras transversarium ammonite Zone) reefal strata (Figure 1, Figure 2).
Description. Carapace small, oval, tubular, convex in transverse section; base of rostrum broad, tuberculate and concave; cervical groove V-shaped centrally, obscure laterally; carapace densely covered with coarse, blunt, forwardly directed tubercles, most robust on two postrostral ridges on anterior part; another less pronounced ridge of tubercles extending centrally on mesogastric with double circular tubercle arrangement forming a figure 8 on mesogastric area; cervical groove forming wide V in central third of carapace and extending obliquely (c. 45 degrees) toward anterolateral border; posterior part of carapace densely covered with coarse, blunt, forwardly directed tubercles, largest around cervical groove; maximum length of preserved posterior part about 35 per cent of total preserved carapace length.
Remarks. The type of ornament, general shape of the anterior carapace and length of the posterior portion, rules out assignment to the families Palinuridae, Cancrinidae, or Scyllaridae and places the present Late Jurassic taxon in the Synaxidae. The two postrostral ridges of larger tubercles on the anterior carapace of Palaeosynaxes nov. gen. could be taken as a prelude of the postfrontal horns of members of the Palinuridae. These two rows of forwardly directed tubercles and the subcircular tubercular arrangements on the central gastric area make Palaeosynaxes nov. gen. easily differentiated from all other known synaxids.
The Middle Triassic Yunnanopalinura schrami Feldmann, Schweitzer, and Zhang in Feldmann, et al., 2012 shows some similarities to synaxids according to Feldmann et al. (2012), but a more detailed comparison is impossible because the Chinese taxon is based on two specimens in which preservation of parts of the cephalothorax is too poor (Fraaije et al., 2020).
DISCUSSION
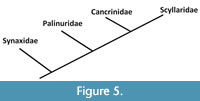 With recent additions of extinct achelatans, the evolutionary history of the group may be interpreted in greater detail. Combined palaeontological data and morphological observations on extant larvae (Baisre, 1994) demonstrate the Synaxidae to be the most basal group within the Achelata. Palero et al. (2009, p. 153) noted that, ‘Under the traditional classification scheme, the presence of a stridulating organ in Palybithus and absence in Palinurellus would imply that either this specialized structure appeared twice (once in the Synaxidae lineage and again in the Stridentes) or that it disappeared twice (once in the Synaxidae lineage and again in the Silentes)’. We opine that the stridulating organ appeared only once, namely in the Synaxidae, and that this family comprises the precursors of the family Palinuridae. Based on the data from Haug et al. (2009), and herein, we propose an evolutionary scenario of the Achelata as illustrated in Figure 5.
With recent additions of extinct achelatans, the evolutionary history of the group may be interpreted in greater detail. Combined palaeontological data and morphological observations on extant larvae (Baisre, 1994) demonstrate the Synaxidae to be the most basal group within the Achelata. Palero et al. (2009, p. 153) noted that, ‘Under the traditional classification scheme, the presence of a stridulating organ in Palybithus and absence in Palinurellus would imply that either this specialized structure appeared twice (once in the Synaxidae lineage and again in the Stridentes) or that it disappeared twice (once in the Synaxidae lineage and again in the Silentes)’. We opine that the stridulating organ appeared only once, namely in the Synaxidae, and that this family comprises the precursors of the family Palinuridae. Based on the data from Haug et al. (2009), and herein, we propose an evolutionary scenario of the Achelata as illustrated in Figure 5.
CONCLUSIONS
Palaeosynaxes montserratae nov. gen., nov. sp., from massive sponge-microbial buildups of middle Oxfordian age near Kraków (southern Poland), adds to the diversity of extinct synaxid achelatans. Type of ornament, general shape of the anterior carapace and length of the posterior portion, as well as the presence of two postrostral ridges of larger tubercles on the anterior carapace allow this taxon to be differentiated from all other extinct and modern synaxids of Late Jurassic to Holocene age. It constitutes one of the earliest records to date of furry lobsters worldwide and extends the palaeogeographical range of this relatively small group of early palinuroids. With more carapaces of extinct forms being recovered, described, and formally named, the phylogenetic history of the achelatans, so far based mainly on extant material, may be assessed more accurately, like recent developments for the anomuran superfamily Paguroidea (Fraaije et al., 2022b).
ACKNOWLEDGEMENTS
We thank the journal reviewers, Rodney Feldmann (Kent State University, USA), Alessandro Garassino (Loma Linda University, USA), Hiroaki Karasawa (Mizunami Fossil Museum, Japan), for pertinent comments that improved an earlier version of the typescript.
REFERENCES
Bachmayer, F. 1959. Neue Crustaceen aus dem Jura von Stramberg (ČSR). Sitzungsberichte der Österreichischen Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Klasse, 168 (I):937-944.
Baisre, J.A. 1994. Phyllosoma larvae and the phylogeny of the Palinuroidea (Crustacea: Decapoda): a review. Australian Journal of Marine and Freshwater Research, 45:925-944. https://doi.org/10.1071/MF9940925
Bate, C.S. 1881. On Synaxes , a new genus of Crustacea. The Annals and Magazine of Natural History, 5:220-228. https://doi.org/10.1080/00222938109459497
Davie, P.J. 1990. A new genus and species of marine crayfish, Palibythus magnificus , and new records of Palinurellus (Decapoda, Palinuridae) from the Pacific Ocean. Invertebrate Taxonomy, 4:685-695. https://doi.org/10.1071/it9900685
De Angeli, A. and Garassino, A. 2014. Palinurellus bericus n. sp. (Crustacea, Decapoda, Palinuridae) from the late Eocene (Priabonian) of San Feliciano (Orgiano, Vicenza, northeastern Italy). Natural History Sciences, Atti della Società italiana di Scienze naturali e del Museo civico di Storia naturale in Milano, 1(1):7-12. https://doi.org/10.4081/nhs.2014.58
De Man, J.G. 1881. Carcinological studies in the Leyden Museum, No. 1. Notes from the Leyden Museum, 3:121-144.
Feldmann, R.M., Schweitzer, C.E., Hu, S., Zhang, Q., Zhou, C., Xie, T., Huang, J., and Wen, W. 2012. Macrurous Decapoda from the Luoping biota (Middle Triassic) of China. Journal of Paleontology, 86(3):452-441. https://doi.org/10.1666/11-113.1
Förster, R. 1973. Untersuchungen an oberjurassischen Palinuridae. Mitteilungen aus der Bayerischen Staatssammlung für Paläontologie und historische Geologie, 13:31-46.
Fraaije, R.H.B., Van Bakel, B.W.M., Jagt, J.W.M., and Brochet, R. 2020. A new furry lobster (Crustacea, Decapoda, Synaxidae) from the Oxfordian (Upper Jurassic) of northeastern France. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 296:327-330. https://doi.org/10.1127/njgpa/2020/0909
Fraaije, R.H.B., Klompmaker, A.A., Jagt, J.W.M., Krobicki, M., and Van Bakel, B.W.M. 2022a. A new, highly diverse paguroid assemblage from the Oxfordian (Upper Jurassic) of southern Poland and its environmental distribution. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 304:1-12. https://doi.org/10.1127/njgpa/2022/1054
Fraaije, R.H.B., Van Bakel, B.W.M., Jagt, J.W.M., Charbonnier, S., Schweigert. G., Garcia, G., and Valentin, X. 2022b. The evolution of hermit crabs (Crustacea, Decapoda, Anomura, Paguroidea) on the basis of carapace morphology: a state-of-the-art-report. Geodiversitas, 44:1-16. https://doi.org/10.5252/geodiversitas2022v44a1
Garassino, A. and Krobicki, M. 2002. Galicia marianae n. gen., n. sp. (Crustacea, Decapoda, Astacidea) from the Oxfordian (Upper Jurassic) of the southern Polish Uplands. Bulletin of the Mizunami Fossil Museum, 29:51-59.
Garassino, A. and Pasini, G. 2020. A new furry lobster (Crustacea, Decapoda, Synaxidae) from the Upper Cretaceous (Cenomanian) of Lebanon. Palaeodiversity, 13:131-133. https://doi.org/10.18476/pale.v13.a10
George, R.W. 2006. Tethys origin and subsequent radiation of the spiny lobsters (Palinuridae). Crustaceana, 79:397-422. https://doi.org/10.1163/156854006777554848
Głowniak, E. 2006. The Platysphinctes immigration event: biostratigraphic and palaeobiogeographic implications for the Middle Oxfordian (Late Jurassic) seas of Central Europe (NW Germany and Poland). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 241:155-2 01. https://doi.org/10.1127/njgpa/241/2006/155
Haug, J.T., Haug, C., Waloszek, D., Maas, A., Wulf, M., and Schweigert, G. 2009. Development in Mesozoic scyllarids and implications for the evolution of Achelata (Reptantia, Decapoda, Crustacea). Palaeodiversity, 2:97-110.
Holthuis, L.B. 1966. On spiny lobsters of the genera Palinurellus , Linuparus and Puerulus (Crustacea, Decapoda, Palinuridae). Symposium on Crustacea, Marine Biological Association of India, 1:260-278.
Holthuis, L.B. 1991. Marine lobsters of the world: An annotated and illustrated catalogue of species of interest to fishes known to date. FAO Fisheries Synopsis, 125:1-292.
Idreesbabu, K.K., Shanis, C.P.R., and Sureshkumar, S. 2018. First record of the rare furry lobster Palinurellus wieneckii (De Man, 1881) (Decapoda: Palinuridae) from the Arabian Sea. Journal of Threatened Taxa, 10:12986-12989. https://doi.org/10.11609/jott.4166.10.15.12986-12989
Krobicki, M. 1994. Decapod Crustacea in the Oxfordian limestones in the vicinity of Cracow. 3rd International Meeting of IGCP N° 43, Excursion Guidebook, 52.
Krobicki, M. and Müller, P. 1998a. Jurassic primitive crabs (Prosopidae) - their palaeoecology and biogeography. 4th International Crustacean Congress, Proceedings and Abstracts, Amsterdam, 76-77.
Krobicki, M. and Müller, P. 1998b. Palaeoecology and biogeography of Jurassic primitive crabs (Brachyura, Prosopidae). 5th International Symposium on the Jurassic System, Abstracts and Program, Vancouver, 50-51.
Krobicki, M. and Zatoń, M. 2008. Middle and Late Jurassic roots of brachyuran crabs: palaeoenvironmental distribution during their early evolution. Palaeogeography, Palaeoclimatology, Palaeoecology, 263:30-43. https://doi.org/10.1016/j.palaeo.2008.01.025
Latreille, P.A. 1802. Histoire naturelle, générale et particulière des Crustacés et des Insectes. Ouvrage faisant suite à l’histoire naturelle générale et particulière, composée par Leclerc de Buffon, et rédigée par C.S. Sonnini, membre de plusieures sociétés savantes. Familles naturelles des genres, 3:1-467. F. Dufart, Paris.
Lavalli, K.J. and Spanier, E. 2010. Infraorder Palinura Latreille, 1802. In Schram, F.R. and Von Vaupel Klein, J.C. (eds.), Treatise on Zoology - Anatomy, Taxonomy, Biology. The Crustacea. Volume 9, Part A. Eucarida: Euphausiacea, Amphionidacea, and Decapoda (partim):425-531. E.J. Brill, Leiden/Boston. https://doi.org/10.1163/ej.9789004164413.i-562.80
Martin, J.W. and Davis, G.E. 2001. An updated classification of the recent Crustacea. Natural History Museum of Los Angeles County, Science Series, 39:1-124. https://doi.org/10.1086/345208
Matyja, B.A. and Wierzbowski, A. 1995. Biogeographic differentiation of the Oxfordian and early Kimmeridgian ammonite fauna of Europe, and its stratigraphic consequences. Acta Geologica Polonica, 45(1-2):1-8.
Matyja, B.A. and Ziółkowski, P. 2014. Upper Jurassic stratigraphy and facies differentiation - a case study of the Cianowice 2 and Trojanowice 2 boreholes (Olkusz Upland and Krzeszowice graben borderland). Biuletyn Państwowego Instytutu Geologicznego, 459:93-108 (In Polish, with English abstract).
Matyszkiewicz, J. 1999. Sea-bottom relief versus differential compaction in ancient platform carbonates: a critical reassessment of an example from Upper Jurassic of the Cracow-Wieluń Upland. Annales Societatis Geologorum Poloniae, 69:63-79.
Matyszkiewicz, J., Kochman, A., and Duś, A. 2012. Influence of local sedimentary conditions on development of microbialites in the Oxfordian carbonate buildups from the southern part of the Kraków-Częstochowa Upland (south Poland). Sedimentary Geology, 263-264:109-132. https://doi.org/10.1016/j.sedgeo.2011.08.005
Müller, P., Krobicki, M., and Wehner, G. 2000. Jurassic and Cretaceous primitive crabs of the family Prosopidae (Decapoda: Brachyura) - their taxonomy, ecology and biogeography. Annales Societatis Geologorum Poloniae, 70:49-79.
Palero, F., Crandall, K.A., Abello, P., Macpherson, E., and Pascual, M. 2009. Phylogenetic relationships between spiny, slipper and coral lobsters (Crustacea, Decapoda, Achelata). Molecular Phylogenetics and Evolution, 50:152-162. https://doi.org/10.1016/j.ympev.2008.10.003
Scholtz, G. and Richter, S. 1995. Phylogenetic systematics of the reptantian Decapoda (Crustacea, Malacostraca). Zoological Journal of the Linnean Society, 113:289-328. https://doi.org/10.1006/zjls.1995.0011
Schweitzer, C.E., Feldmann, R.M., Karasawa, H., and Garassino, A. 2015. Part R, Revised, Volume 1, Chapter 8H: Systematic descriptions: Infraorder Achelata. Treatise Online, 67:1-17. https://doi.org/10.17161/to.v0i0.4987
von Martens, E. 1878. Ueber einige Crustaceen und Mollusken, welche das Zoologische Museum in letzter Zeit erhalten hat. Sitzungsberichte der Gesellschaft naturforschender Freunde zu Berlin, 1878:31-135.
