

 On the oldest Mongolian moschids (Mammalia, Ruminantia) and the early moschid evolution
On the oldest Mongolian moschids (Mammalia, Ruminantia) and the early moschid evolution
Article number: 22.2.53
https://doi.org/10.26879/959
Copyright Paleontological Society, August 2019
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 10 January 2019. Acceptance: 10 July 2019.
{flike id=2670}
ABSTRACT
In contrast to Europe, the fossil record of Miocene Moschidae in Asia remains extremely scarce (so far exclusively from China). The description of at least two moschids from the same locality of Ulan Tolgoi (Valley of Lakes, Mongolia) provides new insights into the early evolution of this ruminant family. The two different species (Moschidae indet. and Micromeryx cf. primaevus) are larger than the oldest known European Moschidae. The Moschidae indet. specimens belong to a Moschidae combining characteristics of Micromeryx and Hispanomeryx. However, the scarcity of the material (three teeth) does not allow the erection of a new taxon. A detailed study of all potential moschid specimens from Asia and further field work would be necessary to solve this issue and to better understand the moschid fossil diversity in Asia.
Bastien Mennecart. Naturhistorisches Museum Wien, Burgring 7, 1010 Vienna, Austria and Naturhistorisches Museum Basel, Augustinergasse 2, 4001 Basel, Switzerland mennecartbastien@gmail.com
Manuela Aiglstorfer. Staatliches Museum für Naturkunde Stuttgart, Rosenstein 1, 70191 Stuttgart, Germany. manuela.aiglstorfer@smns-bw.de
Ursula B. Göhlich. Naturhistorisches Museum Wien, Burgring 7, 1010 Vienna, Austria. ursula.goehlich@nhm-wien.ac.at
Gudrun Daxner-Höck. Naturhistorisches Museum Wien, Burgring 7, 1010 Vienna, Austria. gudrun.hoeck@sbg.at and gudrun.hoeck@nhm-wien.ac.at
Keywords: systematics; musk deer; Moschidae; Micromeryx; early middle Miocene; Asia
Mennecart, Bastien, Aiglstorfer, Manuela, Göhlich, Ursula B., and Daxner-Höck, Gudrun. 2019. On the oldest Mongolian moschids (Mammalia, Ruminantia) and the early moschid evolution. Palaeontologia Electronica 22.2.53 1-17. https://doi.org/10.26879/959
palaeo-electronica.org/content/2019/2670-oldest-mongolian-moschidae
Copyright: August 2019 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
The Moschidae are a family of “derived ruminants” (Pecora). Today, they are only represented by the genus Moschus (or musk deer), but were far more diverse during their evolutionary history. Recent molecular (Hassanin et al., 2012), morphological (Sánchez et al., 2010), and combined (Sánchez et al., 2015) phylogenetic analyses point to a close relationship with bovids. However, there is no record of cranial outgrowths in the family Moschidae. Instead, the moschid males possess saber-like upper canines that are used both in display and for male-to-male combat. Today, the genus Moschus naturally occurs in Asia (from Afghanistan to Siberia) in mountainous and hilly areas (Groves, 2011; Groves and Grubb, 2011). All current musk deer populations are listed in Appendices I and II of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES, 2017) and the IUCN Red List of Threatened Species. There are six endangered and one vulnerable species in the IUCN Red list (IUCN, 2018). In 1975, over 44,000 musk deer (Moschus moschiferus) were living in Mongolia (Damiran, 2005). Around three million musk deer lived in China in the 1950s. Today most likely fewer than 220 to 320 thousand remain (Groves, 2011). One of the main causes for the strong decline is hunting to gain musk (a fragrance produced in the male gland and highly valued in perfume industry and traditional Chinese medicine). The present overexploitation may finally lead to the disappearance of populations in Siberia and China.
During their evolutionary history, moschids were far more diverse and widespread over Eurasia (e.g., Sánchez and Morales, 2006, 2008; Sánchez et al. 2009, 2010, 2011; Wang et al., 2015; Aiglstorfer et al., 2017, 2018). However, only little is known on the origin and early evolution of this family, especially in Asia. Several late Oligocene and early Miocene hornless ruminants have been included into this family, mainly based on plesiomorphic characters including the absence of cranial outgrowths and the presence of saber-like upper canines (e.g., Amphitragulus, Pomelomeryx, Bedenomeryx, Dremotherium, and Blastomerycinae in Prothero, 2007, and Liu and Groves, 2016). However, recent studies demonstrate that some of these taxa can be assigned to other clades (e.g., Dremotherium included in the Cervoidea, Sánchez et al., 2015; Bedenomeryx included in the Giraffomorpha, Mennecart et al., 2019). For the other above-mentioned taxa, we consider that an open nomenclature at the family level is the best to use at the present time (Mennecart, 2012; Mennecart et al., 2012). As a result, only Micromeryx and Hispanomeryx remain as undisputable extinct moschid genera.
According to Sánchez et al. (2010), the Moschidae is defined as the clade “comprising Micromeryx, Hispanomeryx, Moschus, and ‘ Moschus’ grandaevus, their more recent common ancestor and all of its descendants” (Sánchez et al., 2010, p. 1039) and are diagnosed morphologically “by a foramen ovale separated from the infratemporal fossa by a low and small crest [, a] highly derived p4 with a mesolingual conid that joins with the [transverse cristid] through the posterolingual cristid and projects a very developed anterolingual cristid that closes, totally or almost totally, the anterior valley [, an] m3 with a bicuspidate third lobe displaying a well-developed postentoconulidcristid [, the] presence of a posterior metatarsal tuberosity [, and a] long and wide capitular facet in the radius” (Sánchez et al., 2010, p. 1025).
Central Asia is often proposed as the origin of the diversification of pecoran families (Trofimov, 1957; Blondel, 1997; Mennecart, 2015; Mennecart et al., 2017). However, the current oldest record of the Moschidae so far is from southeastern Europe (a small moschid assigned to Micromeryx sp. from Sibnica 4, Serbia; unit MN4 of the European biochronological system based on mammals; older than 16 Ma; Alaburic and Radovic, 2016). Liu and Groves (2016) named Micromeryx sp. from Sihong in China as a record of similar age. However, in their review of the locality, Qiu and Qiu (2013) state that the record is unconfirmed, as the original specimens are lost or the identification cannot be verified. The determination of the ruminant remains defined as Micromeryx from the early Miocene locality Damiao DM16 (Wang et al., 2015) by Zhang et al. (2011) has been questioned due to the fragmentary state of the specimens (Wang et al., 2015). No less than six species of Micromeryx and three species of Hispanomeryx are known in Western and Central Europe (Thenius, 1950; Morales et al., 1981; Azanza, 1986; Vislobokova, 2007; Sánchez and Morales, 2008; Sánchez et al., 2009; Sánchez et al., 2010; Liu and Groves, 2016; Aiglstorfer et al., 2017). In contrast, only a few remains of Micromeryx and Hispanomeryx are currently known from Asia, comprising one described Hispanomeryx species: Hispanomeryx abdrewsi from the Chinese Wolf Camp, Tunggur Formation (Sánchez et al., 2011). Most of the other specimens are undetermined at the species level (Wang et al., 2015; Aiglstorfer et al., 2018).
From 1995 to 2012, the Mongolian Academy of Sciences and the Natural History Museum of Vienna carried out an extensive program of field work in Oligocene and Miocene deposits of the Valley of Lakes region in Mongolia (Daxner-Höck and Göhlich, 2017). This project’s outreach comprises several scientific publications, the identification of about 19,000 fossils, the description of 175 fossil mammal species (including 32 newly erected species), and the first precise integrated stratigraphy of the Mongolian Oligocene and Miocene combining biostratigraphy and radiometric dating. In 2002, Vislobokova and Daxner-Höck published the ruminants discovered during the first missions (1995-1997). Very few ruminant species and specimens were described from the Ulan Tolgoi Miocene deposits then (Eotragus sp. and Lagomeryx sp.). Here we re-evaluate the fossils assigned to Lagomeryx by Vislobokova and Daxner-Höck (2002) and describe two additional teeth of a small-sized ruminant. We consider these fossils to be the first Miocene moschid remains from Mongolia. The very scarce fossils (only four specimens) belong to at least two different species, including one that cannot be assigned to any currently known moschid.
GEOLOGIC SETTING AND AGE
 The locality Ulan Tolgoi is situated in the northeast of the Taatsiin Tsagaan Nuur Basin, which is part of the Valley of Lakes in Central Mongolia (Figure 1). From bottom to top, the basin is filled with Mesozoic and Cenozoic continental sediments. The Cenozoic sequences comprise the Tsagan Ovo, Hsanda Gol, and Loh formations and range from the Eocene to the Miocene. After the first field investigations (Mongolian-Austrian research project 1995 - 1997), an integrated stratigraphy of this region was published based on lithology and biostratigraphy of the fossil bearing sediments and radiometric ages of intercalated basalt flows (basalts I - III) (Höck et al., 1999).
The locality Ulan Tolgoi is situated in the northeast of the Taatsiin Tsagaan Nuur Basin, which is part of the Valley of Lakes in Central Mongolia (Figure 1). From bottom to top, the basin is filled with Mesozoic and Cenozoic continental sediments. The Cenozoic sequences comprise the Tsagan Ovo, Hsanda Gol, and Loh formations and range from the Eocene to the Miocene. After the first field investigations (Mongolian-Austrian research project 1995 - 1997), an integrated stratigraphy of this region was published based on lithology and biostratigraphy of the fossil bearing sediments and radiometric ages of intercalated basalt flows (basalts I - III) (Höck et al., 1999).
Later (2011 - 2012), detailed investigations of more than 20 geological sections ranging from Eocene to Oligocene and earliest Miocene strata of the Taatsiin Tsagaan Nuur Basin followed, yielding around 19,000 fossils from 70 fossil beds. This huge fossil database, the 40Ar/39 Ar ages of basalt I (~ 31.5 Ma) and basalt II (~ 27 Ma), and magnetostratigraphic data enable a composite age chronology for deposits ranging across the Eocene/Oligocene boundary, the Oligocene (Rupelian and Chattian stages / Hsandagolian and Tabenbulukian CMA = Chinese Mammal Ages) and earliest Miocene (Aquitanian stage / Xiejian CMA). The updated data allow age estimations of the hitherto informal Mongolian biozones A - D. The biozones A - B correlate to the Rupelian, biozones C, C1, C1-D correlate to the Chattian, and biozone D to the Aquitanian (Daxner-Höck et al., 2017). Moreover, the informal biozones (A - D) were formalized according to the International Stratigraphic Guide (Harzhauser et al., 2017).
Younger deposits, of the early Miocene (Burdigalian stage / Shanwangian CMA) and/or middle Miocene age (Langhian and Serravallian stages / Tunggurian CMA) were not resampled and not studied in detail during the 2011-2012 field investigations. The associated fossils (indicating the informal biozones D1/1 and D1/2) provide inadequate numbers and quality of data. They belong to early collections of 1995 - 1997, when fossils were collected from surface and from small test samples (Höck et al., 1999; Daxner-Höck and Badamgarav, 2007; Daxner-Höck et al., 2013).
Biozone D1/1 was defined on the basis of a few small mammal teeth from the localities Uolon Ovoony Khurem (samples: ODO-A/2- 6 and ODO-B/1) and Luugar Khudag (sample: LOG-B/1). The composite faunal list is: Alloptox gobiensis, Alloptox cf. minor*, Bellatona sp*, Parvericius buk, Heterosminthus mongoliensis, Democricetodon tongi, Democricetodon aff. lindsayi°, Megacricetodon aff. sinensis ° (identified / updated by *, °: Daxner-Höck et al., 2013 or Erbajeva and Daxner-Höck, 2014 *; Maridet et al., 2014a °). Located in the northwest of the basin, these strata consist of brick-red to brown silty claystone of the Loh Formation. The outcrops are located below basalt III (in this region the ages of basalt III range from ~13 to15 Ma). The estimated age is likely early Miocene (Burdigalian / Shanwangian) (Daxner-Höck et al., 2013; Erbajeva and Daxner-Höck, 2014), although Maridet et al. (2014a) suggested a middle Miocene age according to the evolutionary stage of Cricetidae. In fact, due to the poor fossil evidence final stratigraphic resolutions are still missing.
Biozone D1/2 was defined on the basis of mammals from the localities Ulan Tolgoi (samples: UTO-a, b, c = UTO-A/3, 5, 6; Table 1, Figure 1) and Loh (sample: LOH-A/2), about 80 km east of the localities of biozone D1/1 (Daxner-Höck and Badamgarav, 2007; Daxner-Höck et al., 2013).
Ulan Tolgoi (UTO)
Geographic setting and lithostratigraphy. The locality is situated in the northeast of the basin, close to its northern margin. Here, a ~ 50 m thick sediment sequence of the Loh Formation is exposed along a southwest to northeast striking ridge (Figure 1). From bottom to top, the beds are: (1) reddish-brown sandy siltstone (sample UTO-a = UTO-A/3; GPS position: N 45°20’49” E 101°50’16”), followed by (2) whitish-grey sandstone and siltstone with gravel lenses (scattered bones included). The sandy whitish part is locally covered by (3) a reddish-brown silty claystone (sample UTO-b = UTO-A/5; GPS position: N 45°20’41” E 101°50’28”), followed by (4) a rose-grey siltstone layer (sample UTO-c = UTO-A/6; GPS position: N 45°20’34” E 101°50’13”). Towards the top of the sequence (5) brown silt, and (6) quaternary gravels follow.
Mammal composition. The Ulan Tolgoi fossil collection is composed of small and large mammals. Small mammals were selected from three screen washed samples (UTO-a, b, c), and scattered large mammal remains were collected from surface in the vicinity of the sample places. Though the fauna comprises 21 genera (Table 1), species identification is almost impossible, because the taxa are represented by only one or a few specimens.
Biostratigraphy and age estimation. The age of the Ulan Tolgoi fauna can only be inferred from biostratigraphic data, because no Miocene basalt is exposed in this region, and no magnetostratigraphic data are available so far. The UTO-fauna is characteristic of biozone D1/2, and the initial age estimation was late early Miocene or early middle Miocene (Höck et al., 1999; Daxner-Höck and Badamgarav, 2007). A late early Miocene age was favoured by Vislobokova and Daxner-Höck (2002) and Daxner-Höck et al. (2013). Later, Maridet et al. (2014a, 2014b, 2017) and Erbajeva and Nadezhda (2013) dated the Ulan Tolgoi fauna as middle Miocene.
Recent comparisons of the Ulan Tolgoi fauna to mammals from Early and middle Miocene faunas from Inner Mongolia in China (Qiu et al., 2013, figure 5.2, table 5.1; Wang et al., 2003, table I) show that most genera listed from Ulan Tolgoi have their lowermost occurrences in the early Miocene and beginning of the middle Miocene (late Shanwangian - early Tunggurian): i.e., Parvericius, Keramidomys, Eomyops (= Leptodontomys), Sayimys, Microdyromys, Megacricetodon, Cricetodon, Gobicricetodon, Gomphotherium / Platybelodon, Micromeryx. Only a smaller part may be traced back to the late Oligocene (Tabenbulukian) or earliest Miocene (Xiejian) (Qiu, et al., 2013, table 5.1).
The Tunggur and Tarium Nur faunas of central Inner Mongolia provide a dense fossil record, first of all the Moergen faunas (= former “Tunggur faunas”). This allows us to observe evolutionary changes of individual lineages and changes in the entire faunal composition during the Tunggurian CMA (Qiu, 1996; Qiu, et al., 2013; Wang et al., 2003). Moreover, magnetostratigraphic data along the composite Tunggur section (sections of Tunggur and Tarium Nur) were studied and correlated with the geomagnetic polarity time scale (Wang et al., 2003) and later several times reinterpreted (Qiu et al., 2013). So far, the final correlation of the fossil bearing layers in the Moergen section are assumed to have an age of 13.7 - 11.8 Ma. In case of the Tarium Nur section, the age may be extended downward to 15 Ma (Qiu et al., 2013, p.172, figure 5.3). All of these data indicate a middle Miocene age (Tunggurian) for the Tarium Nur and Moergen faunas.
The composition of the Ulan Tolgoi fauna is incomplete, and it is apparent that most taxa could not be identified on species level because of the extremely low number of fossils due to sampling biases. The absence of Protalactaga, of the high crowned Plesiodipus, and of the advanced Gobicricetodon robustus, on the other hand, suggest an early middle Miocene age of Ulan Tolgoi (~ 16 - 13 Ma; upper Langhian stage / late Shanwangian to early Tunggurian CMA) rather than a younger age.
MATERIAL AND METHODS
Material
The moschid material presented in this work comes from Ulan Tolgoi (UTO, Valley of Lakes, Mongolia). Most of the specimens have been discovered during the first field expeditions (1995-1997), and they are housed at the Naturhistorisches Museum Wien (NHMW) in Austria in agreement with the Mongolian competent authorities under the repository numbers NHMW 2001z0119/0001, NHMW 2001z0120/0001, and NHMW 2001z0120/0002. One specimen has been recently found and is housed at the Institute of Paleontology and Geology of the Mongolian Academy of Sciences (MPC; MPC/0192/0001). Only NHMW 2001z0119/0001 and MPC/0192/0001 possess a specific location in the stratigraphy (UTO-A/5 and UTO-A/6, respectively). Nevertheless, all UTO localities are of similar age (Daxner-Höck et al., 2013). Pictures of the teeth are multi-focus photographs (z-stacking of 10 pictures).
Nomenclature and Measurements
The dental nomenclature (Figure 2) follows Bärmann and Rössner (2011). The measurements were done using a calliper with a precision of 0.2 mm. Systematic position of the Lagomerycidae is still unsolved (Mennecart et al., 2017). We will refer to them in the text as Lagomerycidae as a distinct family from Cervidae.
Institutional Abbreviations
AMNH, American Museum of Natural History, New York, United States of America; MNHN, Muséum National d’Histoire Naturelle, Paris, France; MPC, Institute of Paleontology and Geology of the Mongolian Academy of Sciences, Ulaanbaatar, Mongolia; NHMW, Naturhistorisches Museum Wien, Vienna, Austria; NMB, Naturhistorisches Museum Basel, Basel, Switzerland; SMNS, Staatliches Museum für Naturkunde Stuttgart, Stuttgart, Germany.
Other Abbreviations
UTO, Ulan Tolgoi; p, lower premolar; m, lower molar.
SYSTEMATIC PALAEONTOLOGY
Class MAMMALIA Linnaeus, 1758
Order CETARTIODACTYLA Montgelard, Catzeflis and Douzery, 1997
Sub-order RUMINANTIA Scopoli, 1777
Family MOSCHIDAE Gray, 1821
Moschidae indet. Figure 2.1-2.9
Material
 Right p2 with preserved roots (MPC/0192/0001; size 5.8x2.9) from UTO-A/6, right p4 with preserved roots (NHMW 2001z0119/0001; size 8.6x4.9; formerly identified as Lagomeryx sp. by Vislobokova and Daxner-Höck in 2002) from UTO-A/5, and right m1? (NHMW 2001z0120/0002; size 7.8x5.4) from UTO (Ulan Tolgoi, Valley of Lakes, Mongolia), Loh Formation.
Right p2 with preserved roots (MPC/0192/0001; size 5.8x2.9) from UTO-A/6, right p4 with preserved roots (NHMW 2001z0119/0001; size 8.6x4.9; formerly identified as Lagomeryx sp. by Vislobokova and Daxner-Höck in 2002) from UTO-A/5, and right m1? (NHMW 2001z0120/0002; size 7.8x5.4) from UTO (Ulan Tolgoi, Valley of Lakes, Mongolia), Loh Formation.
Description
In the right p2, the enamel is slightly pleated on the lingual side. The anterior conid is located anteromedian, forming a pointed anterior part of the tooth. The straight anterolabial cristid starts from the apex of the mesolabial conid and joins the anterior conid on its posterolabial side. The anterior conid is low and does not form any labial relief. The large mesolabial conid is the highest cuspid. It is located central and median. It possesses a broad posterior part. Since the posteriolingual conid is labiolingually oriented, it forms a narrow and deep posterior valley. The posterolingual conid is long, reaching to the lingual side of the tooth, forming a small protrusion. On its labial face, we can observe a vertical depression. The posterolabial cristid, reaching from the mesolabial conid apex to the labial part of the posterolingual conid, is curved. The posterolabial conid is quite distinct. It forms the posterolabial corner of the tooth. As there is no posterior stylid, the back valley is widely open. The posterolingual conid is higher than the anterior conid. There is no cingulid.
The p4 is a little worn, high crowned, and the enamel is slightly wrinkled. There is a short anterolabial cingulid and a vestigial anterolingual cingulid. The mesolabial conid is located median. It is smaller than the mesolingual conid. The latter is slightly shifted to anterior and is laterally compressed. It is the highest conid. The very deep anterior valley is closed by the connection of the anterolingual cristid and an enlarged anterior conid. The anterior conid shows an additional minute anterior bifurcation. The anterior stylid forms the anterolingual edge of the tooth. The oblique transverse cristid fuses with the posterior end of the posterolingual cristid building the posterior wall of the anterior valley. Transverse cristid and posterolingual conid are oriented parallel, as are posterior and back valley. The valleys are both deep and oriented obliquely. The posterior valley is open lingually. The posterior cristid turns posteriorly at the base and almost reaches the posterolingual edge of the tooth closing the back valley basally. The posterolabial cristid is very short, and the posterolabial conid is well-marked. There is a distinct labial depression anterior to it. The posterior stylid is very weak. The posterior interdental contact surface implies that the p4 was considerably intruded by the m1.
The medium-sized molar has a fully developed selenodonty. The enamel is slightly wrinkled. The lingual side of the tooth is flattened. The conids are not fully aligned, comprising an intermediate situation between the condition of bovids and cervids/palaeomerycids, as observed in some moschid taxa. All cristids are more or less straight. Preprotocristid and prehypocristid are parallel and oriented obliquely to the main axis of the tooth. The same applies to internal postprotocristid and posthypocristid. The prehypocristid is lower and terminates in the posterior wall of the internal postprotocristid. It does not meet the connection between internal postprotocristid, preentocristid, and postmetacristid. A well-developed, long and slender, external postprotocristid starting from the apex of the protoconid forms a deep “ Palaeomeryx fold”. The metastylid is weak and the entostylid is absent. Postentocristid and posthypocristid are fused and close the posterior wall. The ectostylid is very well-developed. The anterior cingulid is weakly developed. Due to the strong “ Palaeomeryx fold” and the still marked anterior cingulid, we consider the tooth to be a m1.
Taxonomic Discussion
As the p2 (MPC/0192/0001) shows no distinct taxonomic characters it is attributed to the Moschidae here mainly based on size and general morphology.
The p4 (NHMW 2001z0119/0001) was first determined as the lagomerycid Lagomeryx sp. (Vislobokova and Daxner-Höck, 2002). However, the tooth bears a character combination that clearly distinguishes it from Cervidae / Lagomerycidae. First of all, the relatively high crown of the tooth is neither present in early Cervidae, from the early and middle Miocene, nor in Lagomerycidae, (e.g., Azanza and Ginsburg, 1997; Rössner, 2010). Moreover, the split of anterior stylid and anterior conid, the closure of the anterior valley with participation of an anterolingual cristid, an elongated mesolingual conid lacking a strong lingual rib and fusing with an oblique cristid obliqua at the posterior tip of the posterolingual cristid, as well as an open posterior valley in the p4 is characteristic for the Moschidae among all Oligocene to middle Miocene Pecora (e.g., Sánchez and Morales, 2008; Sánchez et al., 2010, 2011; Aiglstorfer and Costeur, 2013; Aiglstorfer et al., 2017; and see Mennecart, 2012, 2015 for late Oligocene to early Miocene ruminants; Azanza and Ginsburg, 1997; Ginsburg and Chevrier, 2003; Rössner, 2010 for Lagomerycidae; van der Made, 2012; Köhler, 1987; Suraprasit et al., 2013 for middle Miocene Bovidae from Eurasia). In the North American “Blastomerycidae” there is a slight tendency to close the anterior valley in the p4 as well (personal observation on the material housed at the AMNH). However, there is no “blastomerycid” taxon that combines a fully closed anterior valley, a mesolingual conid with distinct cristids, and an obliquely transverse cristid in the p4 (Frick, 1937; Prothero, 2007). Other taxa with a closed or nearly closed p4 differ from the p4 described here by morphology and size. Tuscomeryx is an enigmatic insular ruminant possessing an anteriorly closed p4 (Abbazzi, 2001). Its moschid attribution (Abbazzi, 2001; Angelone and Rook, 2011) is questioned today (Sánchez et al., 2010; Fejfar et al., 2015). Its p4s are shorter and anteroposteriorly more compressed than the specimen described here and distinctly differ in morphology (e.g., mesolingual conid with strong rib; for further details on the morphology see Abbazzi, 2001). Although the anterolingual cristid of the p4 described here is not as strongly developed as is usually the case in the middle Miocene Moschidae, it is still in the range of variation of what we observed in the rich moschid populations from Steinheim am Albuch (Germany) or Sansan (France; both middle Miocene; specimens housed at MNHN and SMNS, Aiglstorfer et al., 2017). The strong relief in the described p4 formed by a pronounced posterolabial conid and a distinct incision is similar to the condition in Hispanomeryx. The relief is often more enhanced in this genus than in Micromeryx, although there is variability (pers. obs. and pictures in Azanza, 1986; Sánchez and Morales, 2008; Sánchez et al., 2010, 2011). The p4 from Ulan Tolgoi differs from Hispanomeryx duriensis by having a transverse cristid, which is absent in the latter, and from the other two species H. daamsi and H. aragonensis clearly by a larger size. Its size is closest to the Asian Hispanomeryx andrewsi (Sánchez et al., 2011; see Figure 3). The Mongolian form furthermore shares with H. andrewsi the quite pronounced split in anterior conid and stylid. Hispanomeryx andrewsi is currently only known from the Chinese Wolf Camp, Tunggur Formation (Sánchez et al., 2011; Wang et al., 2015), which is dated from 11.8 to 13 Mya.
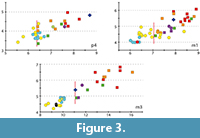 The lower molar (m1?, NHMW 2001z0120/0002) from Ulan Tolgoi also shows clear characteristics of the Moschidae. Its lingual wall is too flat to belong to any Miocene Cervidae or Lagomerycidae or even to any more primitive Pecora (Amphitragulus and Dremotherium). The mesostylid in the Ulan Tolgoi specimen is not as developed as in Cervidae or Amphitragulus and Dremotherium (see above mentioned articles). In contrast to the condition observed in the North American “Blastomerycidae”, which are already more hypsodont than most contemporary Eurasian ruminants, the posterior fossa is posteriorly closed in the Ulan Tolgoi specimen by a fusion of the posthypocristid and a well-pronounced postentocristid. This recalls the condition observed in Bovoidea (Sánchez et al., 2010). From bovids the molar can be clearly distinguished by the presence of a strong external postprotocristid. On the other hand, this cristid is shared with many ruminants as in Tuscomeryx. However, in contrast to Tuscomeryx (Abbazzi, 2001) the lingual wall is more flattened, the conids are less bulky with weaker ribs, and the posterior valley is closed in the m1? from Mongolia, as it is the case in other Moschidae. The presence of a strong external postprotocristid and the marked lingual elements of the Ulan Tolgoi molar also prevents an attribution to Hispanomeryx. The specimen, furthermore, differs from the usual condition in Hispanomeryx, as the inner walls of the conids are less steep and the fossae are wider. In Micromeryx, an external postprotocristid is a common feature (except for Micromeryx azanzae), but in this genus the teeth are less flattened lingually, less aligned and the lingual crown elements have a more pyramidal shape than in the Ulan Tolgoi molar (Sánchez and Morales, 2008; Sánchez et al., 2009; Aiglstorfer et al., 2017). Furthermore, Micromeryx is usually lower crowned than the Ulan Tolgoi molar (Sánchez et al., 2011; Aiglstorfer et al., 2017) and smaller (Figure 3, Figure 4). The ribs in the Mongolian specimen, despite the clearly present rib at the metaconid, are not as bulgy, and the stylids not as prominent as they are usually in Micromeryx flourensianus from Sansan and Steinheim am Albuch (Aiglstorfer et al., 2017). The external postprotocristid in the Mongolian specimen is stronger than in Micromeryx mirus (Vislobokova, 2007) and Micromeryx soriae (Sánchez et al., 2009). The lingual wall of the entoconid is less rounded in the specimen from Mongolia than in these Micromeryx species, as well as in Micromeryx styriacus (Thenius, 1950). The cingulid in the Mongolian specimen is quite weak in comparison to Micromeryx, and even in the more hypsodont M. soriae it seems to be stronger (Sánchez et al., 2009). However, there seems to be even intraspecific variability in this character. In Micromeryx ? eiselei, it also reaches quite far to the lingual side, which is uncommon in M. flourensianus (Aiglstorfer et al., 2017). In any case, the cingulid is usually shorter/less distinct in Hispanomeryx than in Micromeryx. Summing up, the general morphology of the Mongolian molar differs from Hispanomeryx by the presence of a strong external postprotocristid, the absence of this character being diagnostic for the genus Hispanomeryx (Sánchez et al., 2010). The degree of the alignment of the lingual conids represents a condition in between Hispanomeryx species and Micromeryx. The unique combination of characters suggests a new taxon.
The lower molar (m1?, NHMW 2001z0120/0002) from Ulan Tolgoi also shows clear characteristics of the Moschidae. Its lingual wall is too flat to belong to any Miocene Cervidae or Lagomerycidae or even to any more primitive Pecora (Amphitragulus and Dremotherium). The mesostylid in the Ulan Tolgoi specimen is not as developed as in Cervidae or Amphitragulus and Dremotherium (see above mentioned articles). In contrast to the condition observed in the North American “Blastomerycidae”, which are already more hypsodont than most contemporary Eurasian ruminants, the posterior fossa is posteriorly closed in the Ulan Tolgoi specimen by a fusion of the posthypocristid and a well-pronounced postentocristid. This recalls the condition observed in Bovoidea (Sánchez et al., 2010). From bovids the molar can be clearly distinguished by the presence of a strong external postprotocristid. On the other hand, this cristid is shared with many ruminants as in Tuscomeryx. However, in contrast to Tuscomeryx (Abbazzi, 2001) the lingual wall is more flattened, the conids are less bulky with weaker ribs, and the posterior valley is closed in the m1? from Mongolia, as it is the case in other Moschidae. The presence of a strong external postprotocristid and the marked lingual elements of the Ulan Tolgoi molar also prevents an attribution to Hispanomeryx. The specimen, furthermore, differs from the usual condition in Hispanomeryx, as the inner walls of the conids are less steep and the fossae are wider. In Micromeryx, an external postprotocristid is a common feature (except for Micromeryx azanzae), but in this genus the teeth are less flattened lingually, less aligned and the lingual crown elements have a more pyramidal shape than in the Ulan Tolgoi molar (Sánchez and Morales, 2008; Sánchez et al., 2009; Aiglstorfer et al., 2017). Furthermore, Micromeryx is usually lower crowned than the Ulan Tolgoi molar (Sánchez et al., 2011; Aiglstorfer et al., 2017) and smaller (Figure 3, Figure 4). The ribs in the Mongolian specimen, despite the clearly present rib at the metaconid, are not as bulgy, and the stylids not as prominent as they are usually in Micromeryx flourensianus from Sansan and Steinheim am Albuch (Aiglstorfer et al., 2017). The external postprotocristid in the Mongolian specimen is stronger than in Micromeryx mirus (Vislobokova, 2007) and Micromeryx soriae (Sánchez et al., 2009). The lingual wall of the entoconid is less rounded in the specimen from Mongolia than in these Micromeryx species, as well as in Micromeryx styriacus (Thenius, 1950). The cingulid in the Mongolian specimen is quite weak in comparison to Micromeryx, and even in the more hypsodont M. soriae it seems to be stronger (Sánchez et al., 2009). However, there seems to be even intraspecific variability in this character. In Micromeryx ? eiselei, it also reaches quite far to the lingual side, which is uncommon in M. flourensianus (Aiglstorfer et al., 2017). In any case, the cingulid is usually shorter/less distinct in Hispanomeryx than in Micromeryx. Summing up, the general morphology of the Mongolian molar differs from Hispanomeryx by the presence of a strong external postprotocristid, the absence of this character being diagnostic for the genus Hispanomeryx (Sánchez et al., 2010). The degree of the alignment of the lingual conids represents a condition in between Hispanomeryx species and Micromeryx. The unique combination of characters suggests a new taxon.
 The material presented here comprises only a few teeth, and the record of Miocene Moschidae in Asia is generally sparse. Thus, for the moment, it is impossible to evaluate inter- and intraspecific variability among Miocene moschids in this area. Therefore, we cannot determine if the three teeth can indeed be assigned to one taxon, or if they belong to different species. This is especially difficult for the p2, as it is not a very diagnostic tooth. Nevertheless, based on size and shape, we associate the p2 (MPC/0192/0001), the p4 (NHMW 2001z0119/0001) and the m1? (NHMW 2001z0120/0002) to a large form of Moschidae. If indeed p4 and m1? from Ulan Tolgoi belong to the same taxon, the length in the two tooth positions was quite similar, a feature that is similar to the condition observed in the genus Micromeryx (Figure 4). Usually, Hispanomeryx possesses an m1 that is clearly longer than the p4 (Figure 4). Since the p4 is bigger than the m1, some doubt remains on the attribution of the two specimens to the same taxon. However, intraspecific size variation of more than 1 mm between p4 and m1 is still recorded for moschids (e.g., in Hispanomeryx andrewsi and Hispanomeryx daamsi, Sánchez et al., 2010, 2011). Moreover, while two sympatric moschid species are rather common (e.g., Sánchez et al., 2009, 2010; Aiglstorfer et al., 2018), it is very rare to find localities with three coexisting species (only Batallones-1 and Los Valles de Fuertidueña in Spain currently known, Sánchez and Morales, 2006). Therefore, due to the sparsity of the material we refrain from splitting these specimens into more than one species and leave them all in open nomenclature as Moschidae indet.
The material presented here comprises only a few teeth, and the record of Miocene Moschidae in Asia is generally sparse. Thus, for the moment, it is impossible to evaluate inter- and intraspecific variability among Miocene moschids in this area. Therefore, we cannot determine if the three teeth can indeed be assigned to one taxon, or if they belong to different species. This is especially difficult for the p2, as it is not a very diagnostic tooth. Nevertheless, based on size and shape, we associate the p2 (MPC/0192/0001), the p4 (NHMW 2001z0119/0001) and the m1? (NHMW 2001z0120/0002) to a large form of Moschidae. If indeed p4 and m1? from Ulan Tolgoi belong to the same taxon, the length in the two tooth positions was quite similar, a feature that is similar to the condition observed in the genus Micromeryx (Figure 4). Usually, Hispanomeryx possesses an m1 that is clearly longer than the p4 (Figure 4). Since the p4 is bigger than the m1, some doubt remains on the attribution of the two specimens to the same taxon. However, intraspecific size variation of more than 1 mm between p4 and m1 is still recorded for moschids (e.g., in Hispanomeryx andrewsi and Hispanomeryx daamsi, Sánchez et al., 2010, 2011). Moreover, while two sympatric moschid species are rather common (e.g., Sánchez et al., 2009, 2010; Aiglstorfer et al., 2018), it is very rare to find localities with three coexisting species (only Batallones-1 and Los Valles de Fuertidueña in Spain currently known, Sánchez and Morales, 2006). Therefore, due to the sparsity of the material we refrain from splitting these specimens into more than one species and leave them all in open nomenclature as Moschidae indet.
Other Asian fossils falsely attributed to Lagomerycidae may also belong to this lineage of Moschidae indet. Confusion between the Lagomerycidae and Moschidae might actually extend to the works of Roger (1885, 1896), who, on a false comprehension of Micromeryx, described some Lagomeryx remains as Micromeryx. This might be one of the causes that led to a different understanding of moschids and lagomerycids in the literature in the past. The specimens figured in Vislobokova (1983, figure 10 and 11) as Lagomeryx primaevus and ? Lagomeryx triacuminatus from Mongolia, in fact clearly show moschid features, while specimens attributed to Micromeryx sp. in the same article (Vislobokova, 1983, figure 19) are more disputable as they represent only deciduous teeth. It is interesting to note that Vislobokova (1990) also attributed several moschid-like teeth from the Tagay Bay of Ol’Khon Island (Lake Baykal, Russia) to Lagomeryx parvulus. The morphology of the p4 from Tagay Bay (e.g., anterior valley fully closed, oblique transverse cristid fused with the posterolingual cristid) and the size are very similar to the p4 from Ulan Tolgoi (NMW 2001z0119/0001). However, we have not seen the original fossil material from Tagay Bay of Ol’Khon Island, and we cannot rule out a posteriorly extended posterolingual cristid (which would be untypical for the Miocene Moschidae) based on the figures. However, there are also m3 from the Tagay Bay of Ol’Khon Island (Vislobokova, 1990, figure k and q), which bear a closed third basin with a well-developed entoconulid contributing to the lingual wall, as it is typical for the Moschidae. New biostratigraphical dating of the Tagay Bay of Ol’Khon Island fauna led to an age correlating the biozone D1/1 (Langhian), and thus a little older than the Ulan Tolgoi locality (Erbajeva and Nadezhda, 2013; Klementiev and Sizov, 2015). There are further records in Eurasia of alleged “lagomerycids”, but bearing quite distinct moschid morphology (Vislobokova, 1983; Lungu, 1984; see also discussion in Aiglstorfer and Costeur, 2013). Hopefully, with new findings, it will be possible to evaluate the specimens described here and other moschid-like ruminants found in the Miocene fossil record from Asia.
Genus MICROMERYX Lartet, 1851
Type species. Micromeryx flourensianus Lartet, 1851.
Micromeryx cf. primaevus Figure 2.10-12
Material
Left m3 (NHMW 2001z0120/0001; size 11.1x5.4; formerly identified as Lagomeryx sp. by Vislobokova and Daxner-Höck (2002) from UTO (Ulan Tolgoi, Valley of Lakes, Mongolia), Loh Formation.
Description
The small-sized m3 is selenodont. The enamel is slightly wrinkled. The lingual wall in the m3 is more bulgy than in the m1? (NHMW 2001z0120/0002) described above, and the lingual cuspids are less aligned. The metastylid is very strong. The prehypocristid nearly meets the connection between internal postprotocristid, preentocristid, and postmetacristid. A well-developed, long, and bulky external postprotocristid, starting from the apex of the protoconid, forms a deep “ Palaeomeryx fold” with the internal one. There is a quite distinct entostylid sitting lingual on the developed entoconulid. The preentoconulidcristid connects the posthypocristid and the entostylid. The back fossa of the m3 is oblique. The selenodont and quite large hypoconulid forms the posterolabial corner of the m3. The ectostylid and the posterior ectostylid are very well developed. The anterior cingulid is weakly developed.
Taxonomic Discussion
The lower m3 from Mongolia shows another interesting combination of characters. It represents a smaller taxon than the specimens described above (Figure 3) and differs in morphology (Figure 2). It has a less flattened lingual wall and more pronounced lingual elements. There is a clearly developed rib at the metaconid, and a weaker one at the entoconid. Furthermore, the tooth bears a very strong metastylid. This morphology is similar to the general condition in Cervidae and Lagomerycidae. However, neither Miocene Cervidae nor Lagomerycidae possess a back fossa closed by a high entoconulid and a postentoconulidcristid. In Lagomerycidae this feature is only known in the tiny Lagomeryx pumilio, whose phylogenetic relationships are not yet fully resolved (it is the only lagomerycid where no cranial appendages are known until now; Rössner, 2010). In any case, L. pumilio can be distinguished from the Ulan Tolgoi m3 by morphology and size. The tooth crown height is lower, the lingual wall is less aligned and less closed, and it is smaller (Rössner, 2010).
The tooth also differs from other non-moschid taxa. The metastylid of the m3 from Ulan Tolgoi is not as developed as in Amphitragulus and Dremotherium (Mennecart, 2012). The type specimen of the North American “blastomerycid” Blastomeryx olcotti (AMNH 13224) does possess an additional stylid closing the back basin of the m3 (Matthew, 1908), but the basin is not as developed as in our specimen, and the crown of the tooth is clearly lower. The Ulan Tolgoi m3 shares with Tuscomeryx, as with many ruminants, the clearly present external postprotocristid, but differs by a stronger metastylid and a fully closed back fossa with a high entoconulid and a postentoconulidcristid (Abbazzi, 2001). Furthermore, there is no strongly developed postentocristid / entostylid in Tuscomeryx. The ectostylid and the posterior ectostylid are very strong in the Mongolian specimen and similar to the condition in Amphimoschus, which also shares with the Mongolian specimen a biconid third lobe. Amphimoschus, however, clearly differs by the lack of an external postprotocristid from the Mongolian m3. This feature also distinguishes the Ulan Tolgoi specimen from both Bovidae and the moschid Hispanomeryx. Additionally, the m3 from Ulan Tolgoi differs from the currently known Hispanomeryx in its general morphology, e.g., by its less slender crown shape, the less flattened lingual conids, and the more structured lingual wall. In morphology the Ulan Tolgoi specimen is most similar to the moschid Micromeryx, but it is in the upper size range of what is known of this genus from the middle Miocene of Europe (Figure 3). Contrary to Hispanomeryx (except in H. aragonensis and H. andrewsi, where the metastylids are more pronounced than in other Hispanomeryx species; Sánchez et al., 2010, 2011), the molars of Micromeryx flourensianus do possess a fairly strong and salient metastylid, as well as a “ Palaeomeryx fold”. However, the metastylid in the Ulan Tolgoi m3 is still stronger than in M. flourensianus from Sansan and Steinheim am Albuch (Aiglstorfer et al., 2017). This clearly distinguishes the Ulan Tolgoi specimen also from all other Micromeryx species. It furthermore differs from M. azanzae by the presence of the external postprotocristid. Micromeryx ? eiselei is of similar size as the Ulan Tolgoi m3 and possesses a pronounced metastylid (Aiglstorfer et al., 2017). However, the metastylid in this species is still smaller than in the Mongolian specimen, and the relief on the lingual wall is flatter. Furthermore, the anterior cingulid extends to the lingual wall in M.? eiselei, which is not the case in the Ulan Tolgoi specimen.
Theilhard de Chardin (1926) described Moschus primaevus (also written Moschus primigenius in the captions of the figure 8, pl. 4 and figure 1, pl. 5) from the “Terres rouges pontiennes du Chiton-gol” (latest Miocene/Pliocene, China). Qiu et al. (2013) reattributed this locality to the Huitenghe locality (China; Ertemte and Harr Obo faunas in their appendix). Four different sedimentological layers were identified by Theilhard de Chardin (1926). Moschus primaevus has been recovered in the layer B in association with very well-preserved mandibles of Ochotona lagreli (Theilhard de Chardin, 1926). This lagomorph is characteristic for late Miocene deposits, associated to the Mongolian biozone E (Erbajeva, 2007; Erbajeva et al., 2011; Cermák, 2016). Due to the recently found abundant fauna this locality has been attributed to the Bahean stage, late Miocene, ca. 9 Mya (Qiu et al., 2013). As we do not see any characteristic features of Moschus (high anterior cingulid forming a “goat fold”, hypsodont teeth, lack of external postprotocristid, fold like structure at the metaconid) in M. primaevus, but find characteristics for the genus Micromeryx (p4 morphology, presence of external postprotocristid (except of M. azanzae), mesodont teeth, less aligned lingual conids in molars) we agree with Sánchez et al. (2010) and reassess the species as belonging to the genus Micromeryx.
NHMW 2001z0120/0001 is relatively worn in comparison to the type specimen of Micromeryx primaevus. Nevertheless, they both share bulky lingual cuspids with strong stylids, a prominent external postprotocristid, and are relatively lower crown than Hispanomeryx. Furthermore, they are of similar size (Figure 3). However, it cannot be completely resolved if the m3 of Ulan Tolgoi and M. primaevus are conspecific. The material from Ulan Tolgoi is limited, and we can only refer to the drawings of Theilhard de Chardin (1926) for the dental morphology in M. primaevus. In these drawings, there is a structure indicated at the anterior base of the lingual molar wall that is not present in the Ulan Tolgoi specimen. It could either represent the anterior cingulid, proceeding on the lingual wall as in M.? eiselei, or just an intention to highlight the concavity on the lingual wall below the mesostylid. Furthermore, there is a large age difference between the Ulan Tolgoi and the Huitenghe localities. As a consequence of these uncertainties, we determine the specimen NHMW 20001z0120/0001 in open nomenclature as M. cf. primaevus.
BIOSTRATIGRAPHY AND PALEOBIOGEOGRAPHY OF THE OLDEST KNOWN MOSCHIDAE
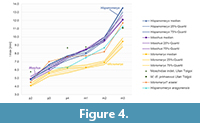
Thanks to recent works on the Miocene Moschidae (e.g., Sánchez and Morales, 2006, 2008; Vislobokova 2007; Sánchez et al. 2009, 2010, 2011; Aiglstorfer and Costeur, 2013; Wang et al. 2015; Aiglstorfer et al., 2017, 2018), we have a better understanding of the early evolutionary history of the Moschidae. Based on combined DNA and morphological character analyses, Mennecart et al. (2017) and Sánchez et al. (2015) predicted an origin of the Moschidae at about 20 Mya. However, no fossil of this age can firmly be attributed to this family (see introduction). Despite the fact that the fossil record is abundant in Central and Western Europe (especially in Spain), no Moschidae is known there before about 15 Mya (European unit MN5; Sánchez and Morales, 2006; Aiglstorfer et al., 2018).
 The oldest recognized moschid is from at least 16 Mya (European unit MN4) and comes from Serbia (see also discussion in introduction; Figure 5; Alaburic and Radovic, 2016). It has been attributed to Micromeryx based on general shape and size. The Balkans were probably separated from Western Europe by the Paratethys at that time and may have been connected to Anatolia (Berger et al., 2005). Unfortunately, data on Asian ruminants from this period are still too limited to allow speculation on the presence of moschids there.
The oldest recognized moschid is from at least 16 Mya (European unit MN4) and comes from Serbia (see also discussion in introduction; Figure 5; Alaburic and Radovic, 2016). It has been attributed to Micromeryx based on general shape and size. The Balkans were probably separated from Western Europe by the Paratethys at that time and may have been connected to Anatolia (Berger et al., 2005). Unfortunately, data on Asian ruminants from this period are still too limited to allow speculation on the presence of moschids there.
A more abundant fossil record of the Moschidae is observed in Europe between 15 and 14 Mya (during European unit MN5, see Figure 5). The retreat of the Paratethys may have permitted Micromeryx to spread all over Eurasia (Aiglstorfer et al., 2018). The dispersal of Hispanomeryx, on the other hand, is different. It is known in Asia between 15 and 14 Mya (Hispanomeryx sp. from Pa ş alar, Lengshuigou, and Lierbao; Wang et al., 2015; Aiglstorfer et al., 2018) and appeared in Europe between 14 and 13 Mya, where it became quite abundant in Spain (Figure 5; Sánchez et al., 2010).
The Moschidae indet. specimens described in this work are a first step to better track the early Asian radiation of the family with probably new genera to be described. With new Asian material we hope to better understand the early evolution and origin of the Moschidae.
CONCLUSIONS
We describe the first unambiguous Miocene Moschidae from Mongolia. They come from the early middle Miocene locality Ulan Tolgoi and belong to at least two different taxa: Micromeryx cf. primaevus and Moschidae indet. The fossils from Ulan Tolgoi are one of the oldest known moschids and the oldest moschids from Mongolia so far. They help to better understand the early evolution of the family Moschidae. However, further data is needed to better understand the broad evolutionary history of these enigmatic ruminants with sabre-like canines.
ACKNOWLEDGMENTS
All our gratitude goes to the the Austrian Science Fund (FWF P-10505-GEO and P-23061-N19) and the Swiss National Science Foundation (projects P300P2_161065 and P3P3P2_161066) who granted this research. N. Ichinnorov, K. Tsogtbaatar, and S. Ulziitseren are also thanked for their permission and help to study the fossils from the collection of the Pal.-Geol. Inst. Mongolian Acad. Sci. Ulaanbaatar. We thank C. Argot (MNHN) and R. Ziegler (SMNS) for access to comparison material. MA visited the collections at the MNHN with the support of the SYNTHESYS Project http://www.synthesys.info (FR-TAF-1892, FR-TAF-6335), which is financed by European Community Research Infrastructure Action under the FP7 “Capacities” Program. Thanks to Nyanka and Zurück for their help during BM's stay in Mongolia. Furthermore, we thank the reviewers I. Sánchez and S. Wang as well as the editors D. Hembree and J. Louys for helping to improve our manuscript.
REFERENCES
Abbazzi, L. 2001. Cervidea and Moschidae (Mammalia, Artiodactyla) from the Baccinello V-3 Assemblage (Late Miocene, Late Turolian, Grosseto, Italy). Rivista Italiana di Paleontologia e Stratigrafia (Research in Paleontology and Stratigraphy), 107(1):107-123. https://doi.org/10.13130/2039-4942/5426
Aiglstorfer, M. and Costeur, L. 2013. The Moschidae of Dorn-Dürkheim 1 (Germany). Palaeobiodiversity and Palaeoenvironments, 93:207-215. https://doi.org/10.1007/s12549-013-0117-9
Aiglstorfer, M., Costeur, L., Mennecart, B., and Heizmann, E.P.J. 2017. Micromeryx ? eiselei—A new moschid species from Steinheim am Albuch, Germany, and the first comprehensive description of moschid cranial material from the Miocene of Central Europe. PLOS ONE, 12(10):e0185679. https://doi.org/10.1371/journal.pone.0185679
Aiglstorfer, M., Mayda, S., and Heizmann, E.P.J. 2018. First record of late middle Miocene Moschidae from Turkey: Micromeryx and Hispanomeryx from Catakbağyaka (Muğla, SW Turkey). Comptes Rendus Palevol, 17:178-188. https://doi.org/10.1016/j.crpv.2017.10.001
Alaburic, S. and Radovic, P. 2016. An early record of the genus Micromeryx (Mammalia, Ruminantia), p. 141-148. In Marković Z. and Milovojević M. (eds.), Life on the Shore - Geological and Paleontological Research in the Neogene of Sibnica and Vicinity (Levač Basin, Central Serbia) Part 1. Natural History Museum in Belgrade.
Angelone, C. and Rook, L. 2011. Alilepus meini nov. sp. (Leporidae, Lagomorpha) from the Early Messinian of Tuscany (central-western Italy). Geobios, 44:151-156. https://doi.org/10.1016/j.geobios.2010.11.003
Azanza, B. 1986. Estudio geológico y paleontológico del Mioceno del sector oeste de la Comarca de Borja. Cuadernos de Estudios Borjanos, XVII-XVIII:63-126.
Azanza, B. and Ginsburg, L. 1997. A revision of the large lagomerycid artiodactyls of Europe. Palaeontology, 40:461-485.
Bärmann, E.V. and Rössner, G.E. 2011. Dental nomenclature in Ruminantia: Towards a standard terminological framework. Mammalian Biology, 76(6):762-768. https://doi.org/10.1016/j.mambio.2011.07.002
Berger, J.-P., Reichenbacher, B., Becker, D., Grimm, M., Grimm, K., Picot, L., Storni, A., Pirkenseer, C., Derer, C., and Schaefer, A. 2005. Paleogeography of the Upper Rhine Graben (URG) and the Swiss Molasse Basin (SMB) from Eocene to Pliocene. International Journal of Earth Sciences, 94:697-710. https://doi.org/10.1007/s00531-005-0475-2
Blondel, C. 1997. Les ruminants de Pech Desse et de Pech du Fraysse (Quercy, MP 28); évolution des ruminants de l’Oligocène d’Europe. Geobios, 30:573-591.
Cermák, S. 2016. The late Miocene species Ochotona kalfense (Mammalia, Lagomorpha) of Moldova: The oldest European record of the genus in the context of the earliest Ochotoninae. Comptes Rendus Palevol, 14:927-940. https://doi.org/10.1016/j.crpv.2016.04.010
CITES 2017. https://cites.org/fra/node/23148 (consulted 10/2017)
Damiran, D. 2005. Palatability of Mongolian rangeland plants. Union, OR, USA: Eastern Oregon Agricultural Research Center, Oregon State University Circular of Information, 3:1-91.
Daxner-Höck, G. 2001. New zapodids (Rodentia) from Oligocene-Miocene deposits in Mongolia. Part 1. Senckenbergiana Lethara, 81(2):359-389. https://doi.org/10.1007/BF03042790
Daxner-Höck, G. and Badamgarav, D. 2007. Geological and stratigraphical setting. Annalen des Naturhistorischen Museums in Wien, 108A:1-24.
Daxner-Höck, G., Badamgarav, D., Barsbold, R., Bayarmaa, B., Erbajeva, M.A., Göhlich, U.B., Harzhauser, M., Höck, V., Höck, E., Ichinnorov, N., Khand, Y., López-Guerrero, P., Maridet, O., Neubauer, T.A., Oliver, A., Piller, W.E., Tsogtbaatar, K., and Ziegler, R. 2017. Oligocene stratigraphy across the Eocene and Miocene boundaries in the Valley of Lakes (Mongolia). Palaeobiodiversity and Palaeoenvironments, 97(1):111-218. https://doi.org/10.1007/s12549-016-0257-9
Daxner-Höck, D., Badamgarav, D., Erbajeva, M., and Göhlich, U.B. 2013. Miocene mammal biostratigraphy of Central Mongolia (Valley of Lakes): new results, p. 477-494. In Wang, X.-M., Flynn, L.J., and Fortelius, M. (eds.), Fossil Mammals of Asia. Columbia University Press, New York.
Daxner-Höck, G. and Göhlich, U. B. 2017. Introduction to the special issue “The Valley of Lakes in Mongolia, a key area of Cenozoic mammal evolution and stratigraphy.” Palaeobiodiversisity and Palaeoenvironments, 97:1-9. https://doi.org/10.1007/s12549-016-0270-z
Erbajeva, M.A. 2007. 5. Lagomorpha (Mammalia): preliminary results. Annalen des Naturhistorischen Museums in Wien, 108(A):165-171.
Erbajeva, M.A. and Daxner-Höck, G. 2014. The most prominent Lagomorpha from the Oligocene and early Miocene of Mongolia. Annalen des Naturhistorischen Museums in Wien, 116:215-245.
Erbajeva, M.A., Mead, J.I., Alexeeva, N.V., Angelone, C., and Swift, S.L. 2011. Taxonomic diversity of Late Cenozoic Asian and North American ochotonid (an overview). Palaeontologia Electronica, 14.3.42A:1-9.
https://palaeo-electronica.org/2011_3/25_erbajeva/index.html
Erbajeva, M.A. and Nadezhda, A. 2013. Late Cenozoic mammal faunas of the Baikalian Region: composition, biochronology, dispersal, correlation with Central Asia, p. 495-507. In Wang, X.-M., Flynn, L.J. and Fortelius, M. (eds.), Fossil Mammals of Asia. Columbia University Press, New York.
Fejfar, O., Sabil, M., and Ďurišová, A. 2015. Pliocene vertebrates from Ivanovce and Hajnáčka (Slovakia). IX. A small-sized cervoid pecoran from Ivanovce. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, 275(3):347-356. https://doi.org/10.1127/njgpa/2015/0468
Frick, C. 1937. Horned ruminants of North America. American Museum of Natural History Bulletin, 59:1-669.
Ginsburg, L. and Chevrier, F. 2003. Les Lagomerycidae (Artiodactyla, Mammalia) de France. Annales de Paléontologie, 89(4):253-268.
Göhlich, U.B. 2007. Gomphotheriidae (Proboscidea, Mammalia). Annalen des Naturhistorischen Museums Wien, 108A:273-289.
Gray, J.E. 1821. On the natural arrangement of vertebrose animals. London Medical Repository, 15:297-310.
Groves, C.P. 2011. Family Moschidae (musk-deer), p. 336-348. In Wilson, D.E. and Mittermeier, R.A. (eds.), Handbook of the Mammals of the World, Vol. 2. Lynx Edicions, Barcelona.
Groves, C. and Grubb, P. 2011. Ungulate Taxonomy. Johns Hopkins University Press, Baltimore.
Harzhauser, M., Daxner-Höck, G., Erbajeva, M.A., López-Guerrero, P., Maridet, O., Oliver, A., Piller, W.E., Göhlich, U.B., and Ziegler, R. 2017. Oligocene and early Miocene mammal biostratigraphy of the Valley of Lakes in Mongolia. Palaeobiodiversity and Palaeoenvironments, 97/1:219-231. https://doi.org/10.1007/s12549-016-0264-x
Hassanin, A., Delsuc, F., Ropiquet, A., Hammer, C., Jansen van Vuuren, B., Matthee, C., Ruiz-Garcia, M., Catzeflis, F., Areskoug, V., Nguyen, T.T., and Couloux, A. 2012. Pattern and timing of diversification of Cetartiodactyla (Mammalia, Laurasiatheria), as revealed by a comprehensive analysis of mitochondrial genomes. Comptes Rendus Biologie, 335(1):32-50. https://doi.org/10.1016/j.crvi.2011.11.002
Heissig, K. 2007. Rhinocerotidae (Perissodactyla, Mammalia). Annalen des Naturhistorischen Museums Wien, 108A:233-269.
Höck, V., Daxner-Höck, G., Schmid, H. P., Badamgarav, D., Frank, W., Furtmüller, G., Montag, O., Barsbold, R., Khand, Y., and Sodov, J. 1999. Oligocene-Miocene sediments, fossils and basalts from the Valley of Lakes (Central Mongolia) - An integrated study. Mitteilungen der Geologischen Gesellschaft Wien, 90:83-125.
IUCN 2018. http://www.iucnredlist.org (consulted 08/2018)
Klementiev, A.M. and Sizof, A.V. 2015. New record of anchithere (Anchitherium aurelianense) in the Miocene of Eastern Siberia, Russia. Russian Journal of Theriology, 14(2):133-143. https://doi.org/10.15298/rusjtheriol.14.2.02
Köhler, M. 1987. Boviden des türkischen Miozäns (Känozoikum und Braunkohlen der Türkei. 28). Paleontologia i Evolució, 21:133-246.
Lartet, E. 1851. Notice sur la Colline de Sansan, Suivie d’une Récapitulation des Diverses Espèces d’Animaux Vertébrés Fossiles Trouvés soit à Sansan, soit dans d’autres Gisements du Terrain Tertiaire Miocène dans le Bassin Sous-Pyrénéen. J.-A. Portes, Auch.
Liu, Z. and Groves, C. 2016. Dispersing or contracting: a perspective on the evolutionary history and population conservation of musk deer. American Journal of Life Sciences, 4:20-30. https://doi.org/10.11648/j.ajls.20160402.12
Lungu, A.N. 1984. The Middle Sarmatian Hipparion Fauna from Moldavia (Hoofed Mammals). Stiintsa, Kishinev. [in Russian]
van der Made, J. 2012. Eotragus clavatus (Artiodactyla, Bovidae, Boselaphini) de Sansan. Mémoire du Muséum National d’Histoire Naturelle, 203:145-199.
Maridet, O., Daxner-Höck, G., Badamgarav, D., and Göhlich, U.B. 2014a. Cricetidae (Rodentia, Mammalia) from the Valley of Lakes (Central Mongolia): focus on the Miocene record. Annalen des Naturhistorischen Museums in Wien, 116:247-269.
Maridet, O., Daxner-Höck, G., Badamgarav, D., and Göhlich, U.B. 2014b. New discoveries of Sciurids (Rodentia, Mammalia) from the Valley of Lakes (Central Mongolia). Annalen des Naturhistorischen Museums in Wien, 116:271-291.
Maridet, O., Daxner-Höck, G., López-Guerrero, P., and Göhlich, U.B. 2017. The record of Aplodontidae (Rodentia, Mammalia) in the Oligocene and Miocene of the Valley of Lakes (Central Mongolia) with some comments on the morphologic variability. Palaeobiodiversisity and Palaeoenvironments; 97:25-49. https://doi.org/10.1007/s12549-016-0255-y
Matthew, W.D. 1908. Osteology of Blastomeryx and phylogeny of the American Cervidae. Bulletin of the American Museum of Natural History, 24:535-562.
Mennecart, B. 2012. The Ruminantia (Mammalia, Cetartiodactyla) from the Oligocene to the early Miocene of Western Europe: systematics, palaeoecology and palaeobiogeography. Geofocus, 32:1-263.
Mennecart, B. 2015. The European ruminants during the “ Microbunodon Event” (MP28, latest Oligocene): impact of climate changes and faunal event on the ruminant evolution. PLOS ONE, 10:e0116830. https://doi.org/10.1371/journal.pone.0116830
Mennecart, B., DeMiguel, D., Bibi, F., Rössner, G.E., Métais, G., Neenan, J.M., Wang, S., Schultz, G., Müller, B., and Costeur, L. 2017. Bony labyrinth morphology clarifies the origin and evolution of deer. Scientific Reports 7:13176. https://doi.org/10.1038/s41598-017-12848-9
Mennecart, B., Scherler, L., Hiard, F., Becker, D., and Berger, J.P. 2012. Ungulates from Rickenbach (type locality for MP29, late Oligocene, Switzerland): palaeoecological and palaeoenvironmental implications. Swiss Journal of Palaeontology, 131:161-181.
Mennecart, B., Zoboli, D., Costeur, L., and Pillola, G.L. 2019. On the systematic position of the oldest insular ruminant Sardomeryx oschiriensis (Mammalia, Ruminantia) and the early evolution of the Giraffomorpha. Journal of Systematic Palaeontology, 17(8):691-704. https://doi.org/10.1080/14772019.2018.1472145
Montgelard, C., Catzeflis, F.M., and Douzery, E. 1997. Phylogenetic relationships of artiodactyls and cetaceans as deduced from the comparison of cytochrome b and 12S rRNA mitochondrial sequences. Molecular Biology and Evolution, 14(5):550-559.
Morales, J., Moyà-Solà, S., and Soria, D. 1981. Presencia de la familia Moschidae (Artiodactyla, Mammalia) en el Vallesiense de Espana: Hispanomeryx duriensis novo gen. nova sp. Estudios Geologicos, 37:467-475.
Prothero, D.R. 2007. Family Moschidae, p. 221-226. In Prothero, D.R. and Foss, S.C. (eds.), The Evolution of Artiodactyls. The Johns Hopkins University Press, Baltimore.
Qiu, Z.-D. 1996. Middle Miocene micromammalian Fauna from Tunggur, Nei Mongol. Science Press, Beijing. (In Chinese with English summary)
Qiu, Z.-D. and Qiu, Z.-X. 2013. Early Miocene Xiejiahe and Sihong fossil localities and their faunas, Eastern China, p. 142-154. In Wang, X.-M., Flynn, L.J. and Fortelius, M. (eds.), Fossil Mammals of Asia. Columbia University Press, New York.
Qiu, Z.-D., Wang, X., and Li, Q. 2013. Neogene faunal succession and biochronology of Central Nei Mongol (Inner Mongolia), p. 155-186. In Wang, X.-M., Flynn, L.J., and Fortelius, M. (eds.), Fossil Mammals of Asia. Columbia University Press, New York.
Roger, O. 1885. Kleine paläontologische Mitteilungen. Bericht des Naturwissenschaftlichen Vereins für Schwaben und Neuburg in Augsburg, 28:93-118.
Roger, O. 1896. Vorläufige Mitteilungen über Säugethierreste aus dem Dinotheriensand von Stätzling bei Augsburg. Bericht des Naturwissenschaftlichen Vereins von Schwaben und Neuburg, 32:547-552
Rössner, G. 2010. Systematics and palaeoecology of Ruminantia (Artiodactyla, Mammalia) from the Miocene of Sandelzhausen (southern Germany, Northern Alpine Foreland Basin). Paläontologische Zeitschrift, 84:123-162. https://doi.org/10.1007/s12542-010-0052-2
Sánchez, I.M., Cantalapiedra, J.L., Ríos, M., Quiralte, V., and Morales, J. 2015. Systematics and evolution of the Miocene three-horned palaeomerycid ruminants (Mammalia, Cetartiodactyla). PLOS ONE, 10(12):e0143034. https://doi.org/10.1371/journal.pone.0143034
Sánchez, I.M., DeMiguel, D., Quiralte, V., and Morales, J. 2011. The first known Asian Hispanomeryx (Mammalia, Ruminantia, Moschidae). Journal of Vertebrate Paleontology, 31(6):1397-1403. https://doi.org/10.1080/02724634.2011.618155
Sánchez, I.M., Domingo, M.S., and Morales, J. 2009. New data on the Moschidae (Mammalia, Ruminantia) from the upper Miocene of Spain (MN 10-MN 11). Journal of Vertebrate Paleontology, 29:567-575. https://doi.org/10.1671/039.029.0223
Sánchez, I.M., Domingo, M.S., and Morales, J. 2010. The genus Hispanomeryx (Mammalia, Ruminantia, Moschidae) and its bearing on musk deer phylogeny and systematics. Palaeontology, 53:1023-1047. https://doi.org/10.1111/j.1475-4983.2010.00992.x
Sánchez, I.M. and Morales, J. 2006. Distribucion biocronologica de los Moschidae (Mammalia; Ruminantia) en Espana. Estudios Geologicos, 62:533-546. https://doi.org/10.3989/egeol.0662153
Sánchez, I.M. and Morales, J. 2008. Micromeryx azanzae sp. nov. (Ruminantia: Moschidae) from the middle upper Miocene of Spain, and the first description of the cranium of Micromeryx. Journal of Vertebrate Paleontology, 28:873-885. https://doi.org/10.1671/0272-4634(2008)28[873:MASNRM]2.0.CO;2
Scopoli, G.A. 1777. Introductio ad Historiam Naturalem Sistens Genera Lapidum, Plantarum, et Animalium: Hactenus Detecta, Caracteribus Essentialibus Donata, in Tribus Divisa, Subinde ad Leges Naturae. Apud Wolfgangum Gerle, Pragae.
Suraprasit, K., Chaimanee, Y., Chavasseau, O., and Jaeger, J.-J. 2013. Middle Miocene bovids from Mae Moh Basin, Northern Thailand: The first record of the genus Eotragus from Southeast Asia. Acta Palaeontologica Polonica, 60:67-78. https://doi.org/10.4202/app.2012.0061
Theilhard de Chardin, P. 1926. Description des mammifères tertiaires de Chine et de Mongolie. Annales de Paléontologie, XV:1-56.
Thenius, E. 1950. Die tertiären Lagomeryciden und Cerviden der Steiermark. Sitzungsbericht der Österreichischen Akademie der Wissenschaften, Mathematisch-naturwissenschaftlichen Klasse I, 159:219-254.
Trofimov, B.A. 1957. Nouvelles données sur les Ruminantia les plus anciens d’Asie. Cursillos y Conferiences del Instituto Lucas Mallada, 4:137-141.
Vislobokova, I.A. 1983. The fossil deer of Mongolia [mainly in Russian]. The Joint Soviet-Mongolian Palaeontological Expedition Transaction, 23:1-74.
Vislobokova, I.A. 1990. Artiodactyls from the lower Miocene of Tagay Bay of Ol’Khon island (Lake Baykal). Paleontological Journal, 24(2):139-143.
Vislobokova, I.A. 2007. New data on late Miocene mammals of Kohfidisch, Austria. Paleontological Journal, 41(4):451-460. https://doi.org/10.1134/S0031030107040119
Vislobokova, I.A. and Daxner-Höck, G. 2002. Oligocene-early Miocene ruminants from the Valley of Lakes (Central Mongolia). Annalen des Naturhistorischen Museums in Wien, 103(A):213-235.
von Linnaeus, C. 1758. Systema Naturæ per Regna Tria Naturæ, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Editio decima reformat.
Wang, S., Shi, Q., Hui, Z., Li, Y., Zhang, J., and Peng, T. 2015. Diversity of Moschidae (Ruminantia, Artiodactyla, Mammalia) in the middle Miocene of China. Paleontological Research, 19:143-155. https://doi.org/10.2517/2014PR032
Wang X., Qiu Z., and Opdyke N. 2003. Litho-, bio-, and magnetostratigraphy and paleoenvironment of Tunggur Formation (middle Miocene) in central Inner Mongolia, China. American Museum Novitates, 3411:1-31.
Zazhigin, V.S. and Lopatin, A.V. 2000. The history of the Dipodoidea (Rodentia, Mammalia) in the Miocene of Asia: 1. Heterosminthus (Lophocricetinae). Palaeontological Journal, 34:319-332.
Zhang, Z.-Q., Wang, L.-H., Kaakinen, A., Liu, L.-P., and Fortelius, M. 2011. Miocene mammalian faunal succession from Damiao, central Nei Mongol and the environmental changes. Quaternary Sciences, 31:608-613. (In Chinese with English abstract)
Ziegler, R., Dahlmann,T., and Storch, G. 2007. Marsupialia, Erinaceomorpha and Soricomorpha. Annalen des Naturhistorischen Museums Wien, 108A:53-164.
