

 Continuous character variation within the Glossopleura-Anoria-Sonoraspis plexus: Dolichometopid trilobites from the Cadiz Formation (Cambrian: Miaolingian, Wuliuan), California
Continuous character variation within the Glossopleura-Anoria-Sonoraspis plexus: Dolichometopid trilobites from the Cadiz Formation (Cambrian: Miaolingian, Wuliuan), California
Article number: 25.2.a18
https://doi.org/10.26879/1173
Copyright Paleontological Society, June 2022
Author biography
Plain-language and multi-lingual abstracts
PDF version
Appendix
Submission: 27 June 2021. Acceptance: 17 May 2022
ABSTRACT
Restudy of type, referred, and new material of the dolichometopid trilobites belonging to Anoria, Glossopleura, and Sonoraspis from the middle Cambrian Cadiz Formation of California, USA, along with material from other formations of the Glossopleura Assemblage Biozone in western North America, demonstrates that only two dolichometopid species are present in the Cadiz Formation: Glossopleura mohavensis and G. lodensis. Sonoraspis californica is confirmed as a junior subjective synonym of G. mohavensis, and Sonoraspis in general is a junior synonym of Glossopleura, as proposed previously. Anoria is valid but should be restricted to the species A. tontoensis. Anoria baton should be within Glossopleura, and the generic position of Anoria bessus is unresolved. Most of the qualitative characters used in the past to separate Anoria, Glossopleura, and Sonoraspis (e.g., palpebral lobe, glabella) prove to be continuously variable with no statistical separation to allow discrete character assignment. Similarly, postcranidial characters, such as thoracic segment numbers, axial spine presence or absence, and enlarged or normal T5 pleural spines, are so variable between genera and within samples as to be unreliable for generic separation, although some characters may assist species diagnoses as non-autapomorphic, “character combination” descriptive elements.
John R. Foster. Utah Field House of Natural History State Park Museum, 496 East Main St., Vernal, Utah 84078 USA. johnfoster@utah.gov
Keywords: Trilobites; Corynexochida; Dolichometopidae; Glossopleura; Cambrian; systematics
Final citation: Foster, John R. 2022. Continuous character variation within the Glossopleura-Anoria-Sonoraspis plexus: Dolichometopid trilobites from the Cadiz Formation (Cambrian: Miaolingian, Wuliuan), California. Palaeontologia Electronica, 25(2):a18. https://doi.org/10.26879/1173
palaeo-electronica.org/content/2022/3617-cadiz-dolichometopids
Copyright: June 2022 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
The Cadiz Formation is a lower-middle (Stage 4-Wuliuan) Cambrian unit exposed in the Marble, Providence, Bristol, and Ship mountains of the Mojave Desert in southeastern California, USA (Hazzard and Crickmay, 1933; Hazzard and Mason, 1936; Fuller, 1980; Foster, 1994, 2011a; Kenney, 2011; Stone et al., 2017). Among the middle Cambrian fossils previously reported from the formation are several species of the dolichometopid trilobite genera Glossopleura, Anoria, and Sonoraspis (Resser, 1928, 1935; Stoyanow and Susuki, 1955). These genera have been regarded as separate by some (Walcott, 1924; Poulsen, 1927; Resser, 1935; Stoyanow, 1952; Stoyanow and Susuki, 1955) but Lochman (in Cooper et al. 1952) and Palmer (1954) regarded Sonoraspis as a junior subjective synonym of Glossopleura.
Anoria was named by Walcott (1924) from the Bright Angel Formation of Grand Canyon. Poulsen (1927) named Glossopleura based on Dolichometopus boccar from the Stephen Formation, plus the new species G. walcotti from the Cape Wood Formation, Greenland. Glossopleura was differentiated from Anoria based on longer eyes and a lack of axial nodes (i.e., “median tubercles” or spine bases) on the thoracic segments or pygidium, plus the lack of an enlarged T5 pleural spine (Poulsen, 1927). Sonoraspis was named by Stoyanow (1952) based on material from the Arrojo Formation of Mexico, with a purported mix of Anoria - and Glossopleura -like characters. Including the dolichometopid Bathyuriscus, which is outside the dolichometopid plexus being studied here, at least 10 genus-species combinations have been assigned to what are probably a total of only three species of dolichometopids from the Cadiz Formation.
This study seeks to: 1) determine the validity of Sonoraspis and Anoria, relative to the more widely reported Glossopleura, by quantitatively testing characters used historically to separate the genera; and 2) determine which genera and species of dolichometopids are then actually present in the Cadiz Formation, based on historic type and referred specimens, plus new material collected in recent years. These results will have implications for other species of these genera from other formations as well.
MATERIALS AND METHODS
Materials studied included type material from numerous institutions, plus published specimens, and newly collected field specimens from the Cadiz Formation (UCR locality 7359). Specimens were identified using type specimens of Glossopleura boccar (Walcott); G. mckeei Resser, 1935; G. mohavensis Resser, 1935; Anoria tontoensis Walcott, 1924; and A. lodensis (Resser, 1935) from the USNM, and Sonoraspis californica Stoyanow and Susuki, 1955, from the LACM, plus a number of other specimens (total N=60; Table 1) and additional references such as Oldroyd (1973), Schwimmer (1973), and Campbell (1974). Based on data in Poulsen (1964), all specimens studied were holaspid stage.
As noted above, qualitative characters used historically to separate the genera Glossopleura, Anoria, and Sonoraspis (and sometimes species of Glossopleura) were tested to help determine the relationships of specimens from the Cadiz Formation. These analyses involved other species from other formations, and in some cases the results had implications for these taxa and units as well.
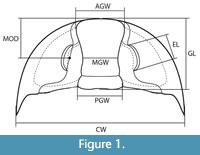 To test the distinction between long and short eyes, and between more anteriorly positioned and posteriorly positioned eyes, in Glossopleura, Anoria, and Sonoraspis, 46 complete or nearly complete specimens and several isolated cranidia were examined (Table 1). Three aspects were measured for these specimens: 1) eye length (EL), more specifically as a proxy for palpebral lobe length; 2) glabella length (GL); and 3) mid-ocular distance (MOD), anterior-posterior distance between level of mid-point of eye length and front edge of glabella, measured parallel to long axis of glabella (Figure 1; Appendix Table 1). Relative eye length (EL/GL) and relative eye position (MOD/GL) were plotted. Total-sample specimen size range was limited and meraspids were not identified; according to criteria of Poulsen (1964), and as noted above, the specimens examined were holaspids, so ontogenetic change in relative eye size or position was not a significant factor. Additionally, there is no indication of ontogenetic change in at least two of the aspects studied.
To test the distinction between long and short eyes, and between more anteriorly positioned and posteriorly positioned eyes, in Glossopleura, Anoria, and Sonoraspis, 46 complete or nearly complete specimens and several isolated cranidia were examined (Table 1). Three aspects were measured for these specimens: 1) eye length (EL), more specifically as a proxy for palpebral lobe length; 2) glabella length (GL); and 3) mid-ocular distance (MOD), anterior-posterior distance between level of mid-point of eye length and front edge of glabella, measured parallel to long axis of glabella (Figure 1; Appendix Table 1). Relative eye length (EL/GL) and relative eye position (MOD/GL) were plotted. Total-sample specimen size range was limited and meraspids were not identified; according to criteria of Poulsen (1964), and as noted above, the specimens examined were holaspids, so ontogenetic change in relative eye size or position was not a significant factor. Additionally, there is no indication of ontogenetic change in at least two of the aspects studied.
Relative expansion of the anterior glabella was assessed by measuring and comparing the width of the anterior glabella (AGW) versus the mid-glabella width (MGW) and versus the posterior glabella width (PGW) (Figure 1). The cephalon width (CW) of specimens with articulated librigenae was also measured. The relative width of the glabella was assessed by comparing the ratio of PGW/CW.
Analysis of the results included calculation of means, standard deviations, 95% confidence ranges, and others. Kolmogorov-Smirnov tests with the data sets were by calculators on Social Science Statistics (socscistatistics.com) and binomial tests were through StatTrek (stattrek.com). Principal components analysis (PCA) of cranidium measurements was with PAST 4.03. Phylogenetic analysis of species within the three dolichometopid genera was in TNT 1.5 (Goloboff and Catalano, 2016) and Mesquite.
Abbreviations—BPM, Back to the Past Museum, Cancún, Mexico; DMNH, Denver Museum of Nature and Science, Denver, Colorado; FHPR, Utah Field House of Natural History State Park Museum, Vernal, Utah; GRCA, Grand Canyon National Park Museum, South Rim, Arizona; KUMIP, University of Kansas Collection of Invertebrate Paleontology, Lawrence, Kansas; LACMIP, Natural History Museum of Los Angeles County, Invertebrate Paleontology, Los Angeles, California; MWC, Museums of Western Colorado, Fruita, Colorado; SGDS, Saint George Dinosaur Discovery Site, Saint George, Utah; UCR, University of California Riverside, Riverside, California; USNM, National Museum of Natural History, Smithsonian Institution, Washington, D.C.; UU, University of Utah Department of Geology & Geophysics, Salt Lake City, Utah.
RESULTS
Characters and Continuous Variation
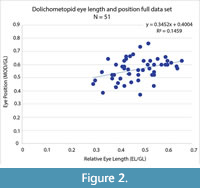 Eye (palpebral lobe) length and position. Historically, Anoria and Sonoraspis have been distinguished from Glossopleura in part based on a qualitative trend of smaller and more anteriorly positioned eyes, and a wider and less anteriorly expanded glabella versus the long and posteriorly positioned eyes, and narrow more anteriorly expanded glabella, of Glossopleura (e.g., Stoyanow and Susuki, 1955; Harrington et al., 1959). Results of relative eye length and position comparison between Anoria, Glossopleura, and Sonoraspis indicate continuous variation in all these parameters. For the entire data set, there is only a weak trend of anterior positioning with smaller eye size and larger eye size with most posterior positioning (Figure 2; slope = 0.3452) and extremely low correlation (R2 = 0.1459). Eye position in particular varies greatly within the total sample (Figure 3) and, more importantly, within the samples assigned to each of the genera. Support for the eye position trend in these dolichometopids in general is weak (Figure 4), and its utility in distinguishing the genera at all is questionable. Figure 4 also indicates no significant change in eye length or position, at least with increasing size within the holaspid sample studied.
Eye (palpebral lobe) length and position. Historically, Anoria and Sonoraspis have been distinguished from Glossopleura in part based on a qualitative trend of smaller and more anteriorly positioned eyes, and a wider and less anteriorly expanded glabella versus the long and posteriorly positioned eyes, and narrow more anteriorly expanded glabella, of Glossopleura (e.g., Stoyanow and Susuki, 1955; Harrington et al., 1959). Results of relative eye length and position comparison between Anoria, Glossopleura, and Sonoraspis indicate continuous variation in all these parameters. For the entire data set, there is only a weak trend of anterior positioning with smaller eye size and larger eye size with most posterior positioning (Figure 2; slope = 0.3452) and extremely low correlation (R2 = 0.1459). Eye position in particular varies greatly within the total sample (Figure 3) and, more importantly, within the samples assigned to each of the genera. Support for the eye position trend in these dolichometopids in general is weak (Figure 4), and its utility in distinguishing the genera at all is questionable. Figure 4 also indicates no significant change in eye length or position, at least with increasing size within the holaspid sample studied.
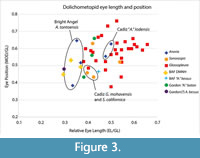 Similarly, eye length relative to glabella length is continuously variable, with Anoria and Glossopleura populations intermixed in the mid-ranges (Figure 3 and Figure 4). In fact, short eyes tend to be associated more with the subset of Bright Angel Formation specimens assigned to Anoria (6 of 7 with shortest eyes), although specimens assigned to G. mckeei consistently do have longer eyes (Figure 3).
Similarly, eye length relative to glabella length is continuously variable, with Anoria and Glossopleura populations intermixed in the mid-ranges (Figure 3 and Figure 4). In fact, short eyes tend to be associated more with the subset of Bright Angel Formation specimens assigned to Anoria (6 of 7 with shortest eyes), although specimens assigned to G. mckeei consistently do have longer eyes (Figure 3). 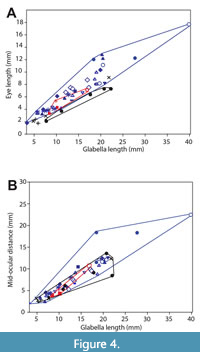 The distinction breaks down in other formations, however, as Sonoraspis and Anoria lodensis (from the Cadiz Formation) both plot among traditional Glossopleura, with the latter Cadiz species actually longer-eyed and none from the Cadiz as small-eyed as Anoria tontoensis (Figure 3). If there is a distinction between Anoria and Glossopleura in relative eye length it would be at 0.380 of glabella length (Anoria EL/GL <0.380 and Glossopleura EL/GL >0.380; Figure 3). This would be an entirely arbitrary distinction, however, based on historical assignment of type and referred Anoria material, mostly from the Bright Angel Formation.
The distinction breaks down in other formations, however, as Sonoraspis and Anoria lodensis (from the Cadiz Formation) both plot among traditional Glossopleura, with the latter Cadiz species actually longer-eyed and none from the Cadiz as small-eyed as Anoria tontoensis (Figure 3). If there is a distinction between Anoria and Glossopleura in relative eye length it would be at 0.380 of glabella length (Anoria EL/GL <0.380 and Glossopleura EL/GL >0.380; Figure 3). This would be an entirely arbitrary distinction, however, based on historical assignment of type and referred Anoria material, mostly from the Bright Angel Formation.
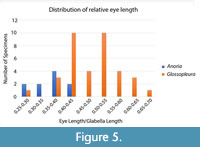 Comparisons of relative eye length for Glossopleura and Anoria indicate little support for a clear distinction between the genera based on this character, however. Both the total sample and the subsamples for each genus are normally distributed (Figure 5), including according to Kolmogorov-Smirnov tests for normality (D=0.11886 for Anoria, N =10; D=0.09329 for Glossopleura, N=34; D=0.09421 for combined data, N=44). Means for the relative eye lengths for Anoria and Glossopleura are distinct, but the 95% confidence ranges overlap. The mean EL/GL for Anoria is 0.3575, and that for Glossopleura is 0.4931. Based on the normal distributions, the 95% confidence ranges are 0.2662-0.4488 for Anoria and 0.3210-0.6652 for Glossopleura, an overlap of 32.0%. Thus, to retain the distinction between Anoria and Glossopleura, based on an arbitrary break in relative eye length or on continuous variation within glabella anterior expansion or relative width, would be, as noted by Schwimmer (1973) for the separation of Anoria and Glossopleura, more of a functional or practical consideration than a natural and statistically-based separation.
Comparisons of relative eye length for Glossopleura and Anoria indicate little support for a clear distinction between the genera based on this character, however. Both the total sample and the subsamples for each genus are normally distributed (Figure 5), including according to Kolmogorov-Smirnov tests for normality (D=0.11886 for Anoria, N =10; D=0.09329 for Glossopleura, N=34; D=0.09421 for combined data, N=44). Means for the relative eye lengths for Anoria and Glossopleura are distinct, but the 95% confidence ranges overlap. The mean EL/GL for Anoria is 0.3575, and that for Glossopleura is 0.4931. Based on the normal distributions, the 95% confidence ranges are 0.2662-0.4488 for Anoria and 0.3210-0.6652 for Glossopleura, an overlap of 32.0%. Thus, to retain the distinction between Anoria and Glossopleura, based on an arbitrary break in relative eye length or on continuous variation within glabella anterior expansion or relative width, would be, as noted by Schwimmer (1973) for the separation of Anoria and Glossopleura, more of a functional or practical consideration than a natural and statistically-based separation.
Specimens featuring distinctly small pygidia and moderately long pleural spines on all thoracic segments (mostly assigned to A. baton and A. bessus from the Gordon Shale of Montana, Deiss, 1939, plus two specimens from the Bright Angel) plot at a range of eye lengths from among A. tontoensis on the short end up to traditional long-eyed Glossopleura, too (Figure 3).
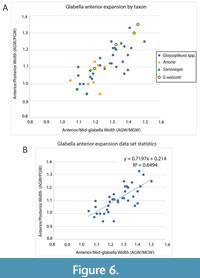 Glabella shape. Comparisons of the anterior expansion of the glabella relative to the mid-width and posterior width, another qualitative character used for distinction of species and genera, show this trend to have essentially continuous variation also, with Glossopleura and other genera intermixed. Although species of Anoria and Sonoraspis do have relatively low ratios, indicating less anterior expansion than in Glossopleura (Figure 6A), the trend is weak and the R2 is moderate at just 0.6494 (Figure 6B). Similarly, the ratio of the posterior glabella width to the full cephalon width in complete specimens does show a mean glabella width broader relative to the cephalon in Anoria (0.305) compared to Glossopleura (0.281), but with 70% overlap of the two-standard deviation 95% confidence intervals. These anterior glabella expansion and relative glabella width characters are, along with relative eye positioning, additional continuously varying features that defy discrete character state assignment for individual genera and sometimes species.
Glabella shape. Comparisons of the anterior expansion of the glabella relative to the mid-width and posterior width, another qualitative character used for distinction of species and genera, show this trend to have essentially continuous variation also, with Glossopleura and other genera intermixed. Although species of Anoria and Sonoraspis do have relatively low ratios, indicating less anterior expansion than in Glossopleura (Figure 6A), the trend is weak and the R2 is moderate at just 0.6494 (Figure 6B). Similarly, the ratio of the posterior glabella width to the full cephalon width in complete specimens does show a mean glabella width broader relative to the cephalon in Anoria (0.305) compared to Glossopleura (0.281), but with 70% overlap of the two-standard deviation 95% confidence intervals. These anterior glabella expansion and relative glabella width characters are, along with relative eye positioning, additional continuously varying features that defy discrete character state assignment for individual genera and sometimes species.
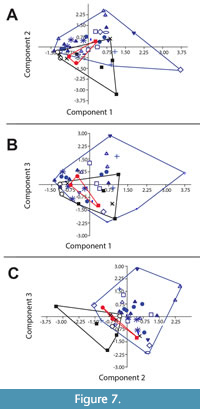 PCA on quantifiable characters. PCA incorporating eye length, eye position, glabella length, and anterior, mid, and posterior glabella width indicates significant overlap between Glossopleura and Sonoraspis for all components (Figure 7). Anoria significantly overlaps the other two as well (Figure 7B) but to a lesser degree (e.g., Figure 7A), especially comparing components 2 and 3 (Figure 7C), in which overall size (effectively accounted for in PC1) is not a factor. Breaking the PCA down by species (see Figure 7 caption), it is clear that A. tontoensis is most separated from Glossopleura, Sonoraspis, and other species assigned to Anoria. PCA thus supports the distinction of Anoria tontoensis of the Bright Angel Formation from other dolichometopids and the broad overlap of Glossopleura, Sonoraspis, and other species of Anoria.
PCA on quantifiable characters. PCA incorporating eye length, eye position, glabella length, and anterior, mid, and posterior glabella width indicates significant overlap between Glossopleura and Sonoraspis for all components (Figure 7). Anoria significantly overlaps the other two as well (Figure 7B) but to a lesser degree (e.g., Figure 7A), especially comparing components 2 and 3 (Figure 7C), in which overall size (effectively accounted for in PC1) is not a factor. Breaking the PCA down by species (see Figure 7 caption), it is clear that A. tontoensis is most separated from Glossopleura, Sonoraspis, and other species assigned to Anoria. PCA thus supports the distinction of Anoria tontoensis of the Bright Angel Formation from other dolichometopids and the broad overlap of Glossopleura, Sonoraspis, and other species of Anoria.
Axial nodes. Thoracic axial nodes (probable spine bases) are highly variable in being present in species assigned to all three genera but only in just less than half of specimens (Table 1); in the others it is taphonomically equivocal as to whether the absence of axial nodes is due to lack of preservation or to true absence. In only a few specimens (e.g., a G. bion from the Spence Shale) was it clear that the dorsal surface of the exoskeleton lacked axial nodes. At least one sample from the Bright Angel Formation of the Grand Canyon has two clearly different forms represented by thoracic segments with smooth axial rings and others with defined nodes. These nodes occur also on pygidia, as they do in some Glossopleura (Sundberg, 2005), but there are several untubercled pygidia in the sample as well.
Elongate pleural spines. The length of pleural spines on the thoracic segments has been used to distinguish some genera and species, particularly the elongate spine on the fifth thoracic (almost rivalling the T3 on olenellids) for Anoria (Anoria tontoensis and A. lodensis) and moderately long spines on most of the thoracic segments in A. bessus and A. baton. However, the elongate T5 spines are only present in three specimens of the total sample studied here (Table 1).
Thoracic segments number. Most specimens of the included genera have seven thoracic segments, but there appear to be rare exceptions in which individuals have eight (Table 1). However, it appears that specimens from the Cadiz Formation, which historically have been assigned to all three genera, consistently have eight thoracic segments (again with one or two possible exceptions), and Spence Shale specimens of Glossopleura mostly have eight thoracic segments (Table 1). Having eight thoracic segments appears to be characteristic more of populations in some formations, and thus geographic regions, than it is consistent for genera, as the character seems to cut across taxa assigned based on eye length and position and other characters.
Phylogenetic Analysis
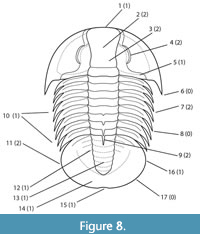 Phylogenetic analysis of 12 dolichometopid taxa for which there are relatively complete specimens, and utilizing 17 characters of the cephalon (1-6), thorax (7-10), and pygidium (11-17) (Appendix; Appendix Table 2; Figure 8), returned 40 most parsimonious trees (MPTs) of 41 steps, a strict consensus of which was a polytomy
Phylogenetic analysis of 12 dolichometopid taxa for which there are relatively complete specimens, and utilizing 17 characters of the cephalon (1-6), thorax (7-10), and pygidium (11-17) (Appendix; Appendix Table 2; Figure 8), returned 40 most parsimonious trees (MPTs) of 41 steps, a strict consensus of which was a polytomy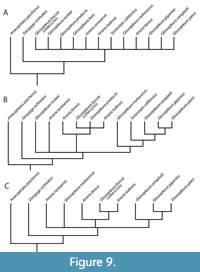 (Figure 9A). However, in many of the trees there were consistent sister taxa pairs: G. boccar + G. producta; G. gigantea + G. yatesi; and G. mohavensis + Sonoraspis californica, the latter pairing reflecting findings presented above. Anoria tontoensis was most frequently basal or at least separate from other taxa, but A. lodensis and A. bessus moved around among species of Glossopleura. Removal of the less well-represented taxon G. bion resulted in an analysis returning four MPTs with 38 steps, the strict consensus of which gave slightly more resolution (Figure 9B). All four MPTs again paired G. gigantea + G. yatesi and also G. boccar + G. producta, both pairings being reflected in the strict consensus tree as well (Figure 9B). The strict consensus also suggested a clade of Spence Shale Glossopleura species and the Cadiz Formation’s G. mohavensis and S. californica, if the latter species is separate. Another possible clade includes Anoria bessus, G. boccar, G. producta, and A. lodensis. Relationships of these clades to the Bright Angel taxa Anoria tontoensis and G. mckeei are unresolved. If we assume a few of the synonymies suggested above, and in some cases elsewhere, too (G. mckeei = G. boccar; S. californica = G. mohavensis; and G. producta = G. boccar) and rerun the analysis, we end up with two MPTs of 35 steps and another clade of Spence Glossopleura and a clade of Anoria bessus +(G. boccar + G. lodensis), both in an unresolved polytomy with G. mohavensis and A. tontoensis (Figure 9C).
(Figure 9A). However, in many of the trees there were consistent sister taxa pairs: G. boccar + G. producta; G. gigantea + G. yatesi; and G. mohavensis + Sonoraspis californica, the latter pairing reflecting findings presented above. Anoria tontoensis was most frequently basal or at least separate from other taxa, but A. lodensis and A. bessus moved around among species of Glossopleura. Removal of the less well-represented taxon G. bion resulted in an analysis returning four MPTs with 38 steps, the strict consensus of which gave slightly more resolution (Figure 9B). All four MPTs again paired G. gigantea + G. yatesi and also G. boccar + G. producta, both pairings being reflected in the strict consensus tree as well (Figure 9B). The strict consensus also suggested a clade of Spence Shale Glossopleura species and the Cadiz Formation’s G. mohavensis and S. californica, if the latter species is separate. Another possible clade includes Anoria bessus, G. boccar, G. producta, and A. lodensis. Relationships of these clades to the Bright Angel taxa Anoria tontoensis and G. mckeei are unresolved. If we assume a few of the synonymies suggested above, and in some cases elsewhere, too (G. mckeei = G. boccar; S. californica = G. mohavensis; and G. producta = G. boccar) and rerun the analysis, we end up with two MPTs of 35 steps and another clade of Spence Glossopleura and a clade of Anoria bessus +(G. boccar + G. lodensis), both in an unresolved polytomy with G. mohavensis and A. tontoensis (Figure 9C).
DISCUSSION
Phylogenetic Analysis
Phylogenetic results suggest: 1) that Sonoraspis californica may be a junior synonym of Glossopleura mohavensis; 2) confirmation that “Anoria” lodensis is a species of Glossopleura (G. lodensis); and 3) that Anoria tontoensis may be distinct. Additionally, and more broadly, the results may indicate: 4) possible synonymy of G. producta with G. boccar; 5) Anoria baton and A. bessus may belong within Glossopleura; and 6) Glossopleura species from the Spence Shale may form a distinct clade, although this latter conclusion may be in part an artifact of positive preservation bias (better specimens) allowing clearer scoring of specimens.
Characters Relative to Populations and Paleobiogeography
The number of thoracic segments in the studied dolichometopids is variable between seven and eight within species but more frequently between formations, with the Cadiz, Spence, and Arrojo formations consistently yielding numerous specimens with eight thoracic segments and sometimes from more than one species. Similarly, the Cadiz, Bright Angel, and Arrojo formations produce species with moderate to short eyes (“S. californica”, Anoria tontoensis, and “S. gomezi”, respectively). Formations such as the Chisholm, Spence, Stephen, and Ophir appear to yield mainly long- to moderate-eyed forms. The range of eye size within A.? bessus from the Gordon Shale and Bright Angel Formation is anomalous. That the smaller-eyed forms on the Glossopleura-Anoria spectrum (Figure 3 and Figure 4) are mostly from more southern formations (and in the case of the Anoria tontoensis, from more proximal settings on the shelf) may be coincidental, but this may be a pattern worth further investigation.
Cadiz Formation Dolichometopidae
Specimens collected from UCR 7359 for this project include numerous dolichometopid cranidia and pygidia plus two partial, articulated specimens of G. mohavensis. Type material of dolichometopids such as Glossopleura boccar and G. stephenensis, plus those of geographically closer-sourced taxa such as G. mckeei and Anoria tontoensis from the Bright Angel Formation and G. bion and others from the Spence Shale helped put the Cadiz Formation species in context (Table 1). The articulated specimens from the Cadiz Formation illustrated by Resser (1928), one assigned to G. mohavensis and the other to G. lodensis (Resser, 1935), appear to show medium- and long-eyed forms, one with and one without enlarged fifth thoracic pleural spines; the nature of the axial rings of these specimens cannot be determined.
Glossopleura lodensis. “Long-eyed” forms from the Cadiz Formation (as shown above) have twice previously been assigned to Anoria lodensis (Mason, 1935; Hazzard and Mason, 1936). Palmer and Halley (1979) note relatively short eyes in the description of Glossopleura lodensis, based on Resser’s (1928, 1935) material, but as confirmed in Figure 3 here, the eyes of G. lodensis are actually long and very typical of Glossopleura.
Glossopleura mohavensis. Recent collecting since 2011, at locality UCR 7359 (Fuller, 1980) in the Marble Mountains near Section B of Foster (1994), recovered additional Glossopleura mohavensis specimens. Among medium-eyed dolichometopid forms without deep S1 or occipital furrows, as preserved, Glossopleura mohavensis admittedly cannot be distinguished from the type Glossopleura boccar except by the shorter eyes and consistent occurrence of eight thoracic segments in specimens from the Cadiz Formation. Unfortunately, Resser (1935) designated G. mohavensis with no diagnosis. Considering the medium-length eyes and eight thoracic segments consistently found in G. mohavensis, it is retained as separate from G. boccar.
In the Cadiz Formation, and in addition to Glossopleura and Anoria, Stoyanow and Susuki (1955) described “Sonoraspis californica” based on two specimens from another site in the Cadiz Formation, near Marble Mountains Section A of Foster (1994). Susuki and Lumsden (1962) described another specimen they attributed to “S. californica” from the Pole Canyon Formation of Nye County, Nevada. The genus Sonoraspis was designated by Stoyanow (1952, in Cooper et al., 1952) from the Arrojo Formation of Sonora, Mexico, with two species designated, “S. torresi” and “S. gomezi”.
“Sonoraspis” is generally similar to Anoria tontoensis except in the lack of elongate fifth thoracic pleural spines and in having larger eyes, and the former has been regarded as a junior subjective synonym of Glossopleura (Lochman in Cooper et al., 1952; Palmer, 1954; Jell and Adrain, 2002). “Sonoraspis californica” from the Cadiz Formation was erected based on eight thoracic segments and “mixed characteristics of” Glossopleura and Anoria (Stoyanow and Susuki, 1955, p. 467). They specified “tubercules” (axial nodes) on the anterior thoracic segments and the “course of the facial sutures” (related to eye length and position) as similar to Anoria and the lack of an elongate fifth thoracic pleural spine as similar to Glossopleura. The lengths of the eyes in the “S. californica” type and paratype specimens are, however, somewhat intermediate between the long eyes of most Glossopleura specimens and the short eyes of the type specimen of Anoria tontoensis (USNM 62685); the eyes of the type “S. californica” material are most similar to G. mohavensis (Figure 3).
Glossopleura walcotti was first identified in the Cadiz Formation by Fuller (1980), but the specimen he illustrated was rather a moderate-eye-length form that would graph close to Sonoraspis and G. mohavensis in Figure 3 (EL/GL of 0.384). Considering this and the variability in occipital and S1 furrow depth among a range of specimens in the formation, Fuller’s (1980) G. walcotti is here regarded as G. mohavensis.
Broader Implications for Dolichometopidae
Anoria. Anoria tontoensis from the Bright Angel Formation of the Grand Canyon was defined by Walcott (1924) based on the lack of a frontal border, a feature shared with Dolichometopus boccar (type species of the not-yet-named Glossopleura Poulsen 1927). Walcott (1925) later described other important aspects of Anoria: 1) elongate fifth thoracic pleural spines, a feature not found in all Bright Angel material referred to Anoria (in fact, it is only present in the type specimen USNM 62685), and present in only two other dolichometopid specimens (of “A.” lodensis from the Cadiz Formation; Table 1); 2) “axial nodes” (likely poorly preserved spine bases) on the thoracic segments, a feature variable within samples of Glossopleura and Anoria (Table 1); and 3) seven thoracic segments, also shared by most Glossopleura (Table 1). The enlarged fifth thoracic, number of thoracic segments, and presence or absence of axial nodes or spines appear to be too variable as characters to be of much value distinguishing Glossopleura and Anoria, although, as noted above, eye length may differentiate the two.
Distinguishing Anoria and Glossopleura except by nearly complete specimens or very well preserved cranidia, therefore, is difficult and unreliable. This results also from the fact that axial nodes extend on to the pygidium in some specimens of Anoria (but also in some of Glossopleura), but do not in all specimens, even from the same quarry. This makes pygidia of Glossopleura and Anoria very similar and difficult to distinguish.
At this time, the only species previously assigned to Anoria that can be confidently separated from Glossopleura, and thus kept in the former genus, is A. tontoensis from the Bright Angel Formation (Walcott, 1916a; Foster, 2011b).
Glossopleura. Poulsen (1927) designated Dolichometopus boccar as the type species of Glossopleura and then named the new species G. walcotti and described its features, some of which are marginally distinctive, but he did not directly address its differentiation from G. boccar. Overall, the two are quite similar in the cranidia and pygidia, although G. walcotti is believed to have more pronounced S1 glabellar and occipital furrows (Poulsen, 1927; Sundberg, 2005). Morphological differences between (and within) populations of Glossopleura are subtle but variable (Sundberg, 2005) and may account for much of what has been attributed to species delineations.
The distinction of longer and more posteriorly positioned eyes in Glossopleura than Anoria (Poulsen, 1927; Harrington et al., 1959; Schwimmer, 1973) seems to be one of the few distinguishing characters potentially useful to separate the two genera, although eye length is only a marginal differentiation and eye position is of little value at all.
Palmer (1954) suggested the synonymy of “Sonoraspis” with Glossopleura due to variability in the number of thoracic segments in samples of many dolichometopid genera, a fact demonstrated in the sample studied here (Table 1) and supplemented by the other analyses above.
Characters. In the years since the original establishments of Anoria, Glossopleura, and “Sonoraspis”, additional characteristics have been used to separate them also, but few of these appear to be clearly distinguishing. As outlined above, the anterior expansion of the glabella and the relative glabella width vary continuously and do not exhibit discrete character states among the genera studied here. Poulsen’s (1927) type material of G. walcotti demonstrates a range of anterior expansion of the glabella relative to the mid-glabella width, but this is within the range of Glossopleura species from other formations as well (Figure 6A; e.g., Cadiz and Spence). Palmer and Halley’s (1979) referred G. walcotti from the Carrara Formation demonstrate significant anterior expansion of the glabella, but the AGW:MGW ratios of these two specimens are not the highest recorded in this study and are comparable to specimens of other Glossopleura species in formations such as the Spence, Bright Angel, and Gordon (Table 1).
An additional apparent difference between Glossopleura boccar and G. walcotti is that the latter has a slight and very shallow mid-line indentation or flat section in the pygidial outline in dorsal view. This indentation or flat section in the otherwise rounded pygidial rim is only identifiable in two specimens from the current sample. Otherwise, the pygidia all strongly resemble G. boccar in pygidial outline, in the width of the pygidial border, and in the length and structure of the pygidial axis. In fact, the pygidial indentation is variably present even among the type material of G. walcotti (Poulsen, 1927). The slight indentation occurs in some specimens of other species of Glossopleura, too (Campbell, 1974; Robison et al., 2015). In addition, the slight flattening of the posterior border of the pygidium seen in G. walcotti also occurs to some degree in the pygidium of the lectotype of Anoria tontoensis (USNM 62685), suggesting again that even generic assignment of the isolated pygidia may be difficult.
These characters are all highly variable, and glabella width, anterior glabella expansion, and eye position in particular appear to be continuously variable features among the Glossopleura-Anoria-“Sonoraspis” plexus. Relative eye length is continuously variable also but an arbitrary distinction between Anoria and Glossopleura at least can be made for use in the above preliminary phylogenetic analysis and other analyses.
With eye length used as an almost sole distinction between Anoria and Glossopleura (Anoria EL/GL <0.380 and Glossopleura EL/GL >0.380), and in the absence of other reliable characters (as outlined above), some consequences are: 1) “Sonoraspis” from the Cadiz Formation is confirmed as a junior synonym of Glossopleura (eye length among medium-eyed Glossopleura); 2) the historic “Anoria” lodensis from the Cadiz is confirmed as within Glossopleura and is a longer-eyed form than “Sonoraspis”; 3) Glossopleura mohavensis and the synonymous “S. californica” form a distinct species of medium-eyed Glossopleura in the Cadiz Formation (G. mohavensis); and 4) dolichometopids with small pygidia and medium-length pleural spines on all thoracic segments, historically assigned to “Anoria” baton and A. bessus (and mostly from the Gordon Shale but with at least two specimens in the Bright Angel), appear to range from small-eyed (Anoria range) to medium-large eyed (Glossopleura range).
SYSTEMATIC PALEONTOLOGY
Order Corynexochida Kobayashi, 1935
Family Dolichometopidae Walcott, 1916
Genus Glossopleura Poulsen, 1927
Type species. Dolichometopus boccar Walcott, 1916a, from the Stephen Formation, British Columbia.
Glossopleura lodensis (Clark, 1921)
Figure 10A
Bathyuriscus howelli var. lodensis Clark, 1921, p. 6
 Dolichometopus ? lodensis Resser, 1928, p. 10, pl. 3, fig. 9.
Dolichometopus ? lodensis Resser, 1928, p. 10, pl. 3, fig. 9.
Dolichometopus lodensis Hazzard and Crickmay, 1933, p. 73.
Anoria lodensis Mason, 1935, p. 109-110, pl. 15, figs. 11, 12.
Glossopleura lodensis Resser, 1935, p. 34
Anoria lodensis Hazzard and Mason, 1936, p. 233.
Glossopleura lodensis Stoyanow, 1956, p. 680, figs. 1, 2.
Glossopleura lodensis Palmer and Halley, 1979, p. 78-79, pl. 16, figs. 1-5, 9, 10.
Glossopleura lodensis Fuller, 1980, p. 68.
Type specimens. Holotype USNM 78400a, complete specimen.
Occurrence. Cadiz Formation, Marble Mountains, California; Carrara Formation, Eagle Mountain, California, and Striped Hills, Nevada; all Glossopleura Assemblage Biozone, Interval Biozone undesignated (Morgan, 2021) (Wuliuan, Miaolingian).
Material examined. USNM 78400a, USNM 208250, USNM 208251, LACMIP 2471-11.
Remarks. Species features long eyes, a long pleural spine on the fifth thoracic segment, and eight thoracic segments in two complete specimens (USNM 78400a, LACMIP 2471-11).
Glossopleura mohavensis Resser, 1935
Figure 10B-F, Figure 11G and 11H
Dolichometopus productus Resser, 1928, p. 10, pl. 3, fig. 9.
Dolichometopus productus Hazzard and Crickmay, 1933, p. 73.
 Dolichometopus productus Mason, 1935, p. 113-114.
Dolichometopus productus Mason, 1935, p. 113-114.
Glossopleura mohavensis Resser, 1935, p. 34
Glossopleura mohavensis Hazzard and Mason, 1936, p. 233.
Sonoraspis californica Stoyanow and Susuki, 1955, p. 468-469, pl. 1, figs. 1, 2.
Sonoraspis mohavensis Stoyanow, 1956, p. 680-681, fig. 1.
Sonoraspis nelsoni Stoyanow, 1958, p. 348-349, pl. 1, fig. 3.
Sonoraspis californica Susuki and Lumsden, 1962, p. 234-237, fig. 2.
Glossopleura californica Fuller, 1980, p. 68.
Type specimens. Holotype USNM 78400b, nearly complete specimen.
Occurrence. Cadiz Formation, Marble Mountains, California; Monola Formation, Inyo Mountains, California; Pole Canyon Formation, Currant Gap, Nevada. Glossopleura Assemblage Biozone, Interval Biozone (Morgan, 2021) undesignated, (Wuliuan, Miaolingian).
Material examined. USNM 78400b, LACMIP 2469, LACMIP 10785, LACMIP 10786, LACMIP 10782, MWC 7779, MWC 7769.
Remarks. Species features moderately long eyes, short pleural spines on all thoracic segments, and eight thoracic segments in seven complete specimens (USNM 78400b, LACMIP 2469, LACMIP 10782, LACMIP 10785, LACMIP 10786, MWC 7779, MWC 7769).
Glossopleura boccar (Walcott, 1916a)
Figure 11A-B and 11D-E
Dolichometopus boccar Walcott, 1916a, p. 363-365, pl. 52, figs. 1a-1f.
Dolichometopus productus (Hall and Whitfield, 1877) (part) Walcott, 1916a, p. 369-372, pl. 53, figs. 4, 4a.
Glossopleura boccar (Walcott) Poulsen, 1927, p. 268.
Glossopleura mckeei Resser, 1935, p. 33.
Glossopleura bosworthensis Resser, 1935, p. 31.
Glossopleura nitida Resser, 1935, p. 31.
Glossopleura ? stephenensis Resser, 1935, p. 31.
Glossopleura boccar (Walcott) Rasetti, 1951, p. 164, pl. 24, figs. 1-6.
Glossopleura mckeei (Resser) Rasetti, 1951, p. 165, pl. 24, figs. 9-12, 18.
Glossopleura boccar (Walcott) Sundberg, 2005, p. 65-66, fig. 6.10-6.14.
Glossopleura boccar (Walcott) Foster, 2011b, p. 105-106, fig. 4.1, 4.2, 4.3.
Glossopleura boccar (Walcott) Morgan, 2021, p. 94-97, figs. 3.5.1, 3.5.2, 3.5.4-3.5.11
Type specimens. USNM 62703, USNM 62705, and USNM 62707, cotypes.
Occurrence. Stephen Formation, Mount Bosworth and Mount Stephen, British Columbia; Mount Cap Formation, Northwest Territories; Chisholm Formation, Half Moon Mine, Pioche, Nevada; Bright Angel Formation, Grand Canyon, Arizona; all Glossopleura Assemblage Biozone, Glossopleura boccar Interval Biozone (Morgan, 2021), (Wuliuan, Miaolingian).
Material examined. USNM 62703, USNM 62702, USNM 62705, USNM 62707, USNM 62713, USNM 62714, USNM 108592a, USNM 108592b, USNM 108592c, SGDS 1973-1976.
Remarks. This is the type species of the genus Glossopleura, and although it has not been identified in the Cadiz Formation, it is close in form to G. mohavensis except for having longer eyes and only seven thoracic segments.
Morgan (2021) proposed restricting Glossopleura boccar to USNM 62703 as a lectotype and USNM 62705 as a syntype/paratype, with USNM 62707 as a hypotype of G. walcotti.
Glossopleura walcotti Poulsen, 1927
Glossopleura walcotti Poulsen, 1927, p. 268, pl. 16, figs. 20-30.
Glossopleura expansa Poulsen, 1927, p. 269, pl. 16, figs. 31, 32.
Glossopleura sulcata Poulsen, 1927, p. 272, pl. 16, fig. 39.
Glossopleura longifrons Poulsen, 1927, p. 272, pl. 17, fig. 10 [not figs. 8, 9].
Glossopleura walcotti Poulsen, 1964, p. 25-29, pl. 1, figs. 2-4.
Glossopleura walcotti Palmer and Halley, 1979, p. 79, pl. 16, figs. 6-8, 11-19.
Glossopleura walcotti Babcock, 1994, p. 94, fig. 12.
Glossopleura aff. leona Lochman Bordonaro and Banchig, 1995, pl. 2, figs. 14, 15.
Glossopleura walcotti Benedetto et al., 2009, fig. 2g.
Glossopleura walcotti Foglia and Vaccari, 2010, p. 434, figs. 4.10-4.23.
Glossopleura walcotti ? Foster, 2011b, p. 106, figs. 4.4, 4.5.
Glossopleura walcotti Sundberg, 2018, p. 19-20, fig. 13.
Glossopleura walcotti Morgan, 2021, p. 96, fig. 3.5.3.
Type specimens. Poulsen (1927) unnumbered, cranidia; USNM 62707, hypotype (Morgan, 2021), thoracics and pygidium.
Occurrence. Cape Wood Formation and Henson Gletscher Formation, Greenland; Stephen Formation, British Columbia; Carrara Formation, California and Nevada; Emigrant Formation, Split Mountain, Nevada; La Laja Formation, San Juan, Argentina; possibly Bright Angel Formation, Grand Canyon, Arizona; all Glossopleura Assemblage Biozone, Glossopleura walcotti Interval Biozone (Morgan, 2021), (Wuliuan, Miaolingian).
Material examined. USNM 62707 (see Morgan, 2021); USNM 208256, USNM 208258; unnumbered specimens in Poulsen, 1927, pl. 16.
Remarks. Glossopleura walcotti is identified mainly by deep S1 glabellar and occipital furrows, although these can of course vary due to taphonomic factors quite independently from actual morphology. The presence of G. walcotti in the Cadiz Formation was proposed by Fuller (1980), but this could not be confirmed by the present study and most specimens are likely G. mohavensis.
Glossopleura producta (Hall and Whitfield, 1877)
Ogyia producta Hall and Whitfield, 1877, p. 244, pl. 2, figs. 31-34.
Bathyuriscus productus Walcott, 1886, p. 217, pl. 30, fig. 1f.
Dolichometopus productus (part) Walcott, 1916a, p. 369-372, pl. 53, figs. 2, 2a-e, 3, 3a-b.
Dolichometopus productus Resser, 1928, p. 10, pl. 3, fig. 9.
Glossopleura producta Resser, 1935, p. 31-32.
Glossopleura producta Palmer, 1954, pl. 13, figs. 7, 9-13.
Type specimens. USNM 15456, USNM 15459, cotypes.
Occurrence. Ophir Formation, Oquirrh Mountains, Utah; Glossopleura Assemblage Biozone, Interval Biozone (Morgan, 2021) undesignated, (Wuliuan, Miaolingian).
Material examined. USNM 15459, USNM 123356, FHPR 16738, FHPR 16739, FHPR 11285, DMNH EPI.42919.
Remarks. Much previous material from the Cadiz Formation, once identified as Dolichometopus productus, would at later stages have been considered Glossopleura producta; however, Palmer (1954) and Sundberg (2005) restricted G. producta to the type material from the Ophir Formation of Utah only. Most Cadiz specimens previously assigned to Dolichometopus (G.) producta are more likely G. mohavensis.
Glossopleura baton (Walcott, 1916a)
Dolichometopus baton Walcott, 1916a, p. 362, pl. 51, figs. 2, 2a-b.
Anoria baton Resser, 1935, p. 10.
Anoria baton (part) Schwimmer, 1973, pl. 6, figs. 4, 5.
Type specimens. USNM 62696-62698, partial to nearly complete specimens.
Occurrence. Gordon Shale, Montana; Glossopleura Assemblage Biozone, Interval Biozone (Morgan, 2021) undesignated, (Wuliuan, Miaolingian).
Material examined. USNM 62696-62698; USNM uncataloged in Schwimmer (1973).
Remarks. Most characters of these specimens are typical of various species of Glossopleura; most importantly, however, the eye length of the studied specimens falls within the range of Glossopleura, and the specimens cannot clearly be placed within Anoria even by this possibly more reliable character.
Glossopleura sp.
Sonoraspis torresi Stoyanow, 1952 (in Cooper et al., 1952), p. 53-54, pl. 14, figs. 1-4.
Sonoraspis gomezi Stoyanow, 1952 (in Cooper et al., 1952), p. 54-55, pl. 14, figs. 5, 6.
Glossopleura sp. Lochman, 1952 (in Cooper et al., 1952), p. 137, pl. 31, figs. 8-14.
Glossopleura leona (part) Cuen-Romero et al., 2018, p. 230.
Occurrence. Arrojo Formation, Sonora; Glossopleura Assemblage Biozone, Interval Biozone (Morgan, 2021) undesignated, (Wuliuan, Miaolingian).
Material examined. USNM 116333, USNM 116349, USNM 116348, USNM 116350 (all originally S. torresi by Stoyanow, 1952); USNM 116335, USNM 116334, both originally S. gomezi by Stoyanow, 1952), all nearly complete specimens.
Remarks. These specimens compare well with those of “S. californica” (and thus Glossopleura mohavensis), especially in the number of thoracic segments, length of pleural spines, and medium length eyes in those preserving the structures. The preservation, features, and axial nodes of USNM 116334 especially are similar to “S. californica”. These specimens are then rather similar to G. mohavensis from the Cadiz Formation, although they are not formally assigned to that species here. Comparison with Glossopleura leona is more difficult given the preservation of nearly complete specimens in shale for the present specimens versus cranidia and pygidia only in limestone for G. leona (Lochman in Cooper et al., 1952).
Genus Anoria Walcott, 1924
Type species. Dolichometopus tontoensis Walcott, 1916a, from the Bright Angel Formation, Grand Canyon, Arizona.
Anoria tontoensis Walcott, 1924
Figure 11F
Dolichometopus tontoensis Walcott, 1916a, p. 373, pl. 51, figs. 1, 1a-1h.
Anoria tontoensis (Walcott, 1916a) Walcott, 1924, p. 54, p. 54, pl. 9, fig. 2.
“Anoria” tontoensis Foster, 2011b, p. 107-110. figs. 5, 6.
Type specimens. USNM 62685, lectotype, part and counterpart of a complete specimen; USNM 62686, paralectotype, nearly complete specimen.
Occurrence. Bright Angel Formation, Grand Canyon, Arizona; Glossopleura Assemblage Biozone, possibly Glossopleura walcotti Interval Biozone (Morgan, 2021) if identification of G. walcotti in same beds (Foster, 2011b) is correct, (Wuliuan, Miaolingian).
Material examined. USNM 62685, USNM 62686, GRCA 2797, GRCA 11520, GRCA 123303, GRCA 123304, DMNH EPI.42917.
Remarks. This is the type and only clearly valid species of Anoria studied in this report; previously reported specimens of other Anoria species from the Cadiz Formation are confirmed as belonging within Glossopleura.
Anoria? bessus (Walcott, 1916a)
Dolichometopus ? bessus Walcott, 1916a, p. 362-363, pl. 51, figs. 3, 3a-c.
Anoria bessus (Walcott) Resser, 1935, p. 10, p. 54, pl. 9, fig. 2.
Anoria baton (part) Schwimmer, 1973, pl. 6, fig. 3.
Type specimens. USNM 62699, nearly complete specimen; USNM 62700, pygidium; USNM 62701, cephalon with hypostome.
Occurrence. Gordon Shale, Montana; Bright Angel Formation, Grand Canyon (Grand Wash Cliffs), Arizona; Glossopleura Assemblage Biozone, Interval Biozone (Morgan, 2021) undesignated, (Wuliuan, Miaolingian).
Material examined. USNM 62699, USNM 62700, USNM 62701, DMNH EPI.42915, DMNH EPI.42916, USNM uncataloged (in Schwimmer, 1973).
Remarks. Species characterized by short to long eyes, reduced oval pygidium smaller than cephalon, and moderately elongate pleural spines on all thoracic segments. This latter character contrasts with the short pleural spines of most species of Glossopleura and the elongate fifth thoracic pleural spines of G. lodensis and the type material of Anoria tontoensis. These features are pronounced in USNM 62699 and the uncataloged specimen illustrated by Schwimmer (1973). Despite qualitatively moderately short eyes in the Gordon Shale specimens, most specimens have moderate to long eyes except USNM 62699 (EL/GL of 0.298 versus 0.428 in Schwimmer’s, 1973, USNM uncataloged and 0.446 in DMNH EPI.42915). Thus, all but one specimen featuring moderately elongate pleural spines and a small, oval pygidium from the Gordon and Bright Angel formations, typical of Anoria bessus, actually have elongate eyes that fit within the range of Glossopleura. Because the single anomalous, short-eyed specimen (USNM 62699) is the most complete of the cotypes, all the studied material here is designated Anoria ? bessus. The question of whether the species is better placed in Glossopleura (despite the short-eyed USNM 62699 and considering the results of this study regarding eye length) is left for a separate investigation.
CONCLUSIONS
Based on the material studied here (Table 1), in the Cadiz Formation: 1) the type of Glossopleura mohavensis (USNM 78400b) is retained as a valid species, distinguished from other species by the consistent occurrence of eight thoracic segments (in Cadiz, Pole Canyon, and Monola formations), moderate eye length (EL/GL just >0.380), and a lack of elongate T5 pleural spines; other than the number of thoracic segments, however, this species is similar to G. boccar; 2) the type and paratype of “Sonoraspis californica” (LACM 10785 and 10786) are confirmed as being included within G. mohavensis: there are no previously featured differences between the two taxa that are consistent, and their eye lengths and positions all match well and are among the Glossopleura range, and they share eight thoracic segments each; this, by qualitative and PC analysis, lends quantitative support to the idea that at the genus level “Sonoraspis” is a junior synonym of Glossopleura (Lochman in Cooper et al., 1952; Palmer, 1954); and 3) “Anoria” lodensis of Mason (1935; USNM 78400a) and Stoyanow (1956) is confirmed as a species of Glossopleura (G. lodensis) as listed by Resser (1935), characterized by elongate T5 pleural spines and frequently eight thoracic segments; USNM 78400a and LACM 2471-11 demonstrate that G. lodensis has the long eyes typical of many members of that genus. Thus, there appear to be just two species of Glossopleura in the Cadiz Formation: G. mohavensis and G. lodensis. As demonstrated here and in Mason (1935) and based on cranidial and complete material, both species occur at UCR 7359.
Additionally, study of non-Cadiz material in this report suggests: 4) Anoria may be a distinct genus but should be restricted to A. tontoensis (currently known only from the Bright Angel Formation), as the most consistently small-eyed (EL/GL <0.380) and PCA-separated dolichometopid; 5) in Montana, the species “A.” baton should be Glossopleura baton based on the type material; and 6) A.? bessus specimens from the Gordon Shale and Bright Angel Formation, united by reduced, oval pygidia and moderately long pleural spines on all thoracic segments, demonstrate a wide range of relative eye lengths, from short (graphing among Anoria specimens in USNM 62699) up to approximately average for Glossopleura (in DMNH EPI.42915 from the Bright Angel Formation). This latter, broad range of relative eye lengths, in a species (A.? bessus) otherwise distinguished from all others by more clear differences in the pygidium and thorax, is a conundrum best left to future projects involving more specimens, but it may undermine the utility of even quantitative measurements of relative eye length for genus assignment in dolichometopids. Measurement of many of the previously qualitative characters used historically to separate Anoria, Glossopleura, and “Sonoraspis” vary too much along continuous spectra for the characters to be reliable for generic separation.
ACKNOWLEDGMENTS
Thanks to the curators and collections managers at the DMNH, GRCA, KUMIP, LACM, MWC, SGDS, UCR, and USNM for loan of material and facilitating collections visits. Special thanks to D. Schwimmer, A.R.C. Milner, F. Sundberg, J. Hagadorn, and R. Irmis for additional information, material, and assistance with TNT, Mesquite, and the PCA, and to S. Gon III for the images for Figure 1 and Figure 8. Some of the new specimens from the Cadiz, Ophir, and Chisholm, now at the MWC, DMNH, FHPR, and SGDS, were found by J. Hagadorn, A.R.C. Milner, R. Hunt-Foster, and Z. Miracle. Special thanks to C. Morgan and two anonymous reviewers for their comments which improved the manuscript. Field work was facilitated by: the Bureau of Land Management under permit CA-08-00-002P (Cadiz), L. Ault and J. Lehane (Ophir), and National Park Service permit GRCA-2019-SCI-0029 and a National Science Foundation Earth Sciences grant to J. Hagadorn and K. Karlstrom (Bright Angel).
REFERENCES
Babcock, L.E. 1994. Systematics and phylogenetics of polymeroid trilobites from the Henson Gletscher Kap Stanton formations (Middle Cambrian), North Greenland. Bulletin Grønlands Geologiske Undersøgelse, 169:79-127. https://doi.org/10.34194/bullggu.v169.6727
Benedetto, J.L., Vaccari, N.E., Waisfeld, B.G., Sánchez, T.M., and Foglia, R.D. 2009. Cambrian and Ordovician biogeography of the South American margin of Gondwana and accreted terranes. In Basset, M.G. (ed.), Early Paleozoic Peri-Gondwana Terranes: New Insights from Tectonics and Biogeography. Geological Society of London, Special Publications, 325:201-232. https://doi.org/10.1144/SP325.11
Bordonaro, O.L. and Banchig, A.L. 1995. Trilobites Laurenticos en el Cámbrico de la Precordillera Argentina. VI Congreso Argentino de Paleontología y Bioestratigrafía, Actas, p. 59-65.
Campbell, D.P. 1974. Biostratigraphy of the Albertella and Glossopleura zones (Lower Middle Cambrian) of northern Utah and southern Idaho. Unpublished MSc thesis, University of Utah, Salt Lake City, 295 p.
Clark, C.W. 1921. Lower and Middle Cambrian formations of the Mohave Desert. University of California Publications, Bulletin of the Department of Geological Sciences, 13:1-7.
Cooper, G.A. 1952. Cambrian stratigraphy and paleontology near Caborca, northwestern Sonora, Mexico. Smithsonian Miscellaneous Collections, 119(1):1-181.
Cuen-Romero, F.J., Valdez-Holguín, J.E., Buitró-Sánchez, B.E., Monreal, R., Enríquez-Ocaña, L.F., Aguirre-Hinojosa, E., Ochoa-Granillo, J.A., and Palafox-Reyes, J.J. 2018. Trilobite-based biostratigraphy (Arthropoda-Trilobita) and related faunas of the Cambrian from Sonora, Mexico. Journal of South American Earth Sciences, 83:227-236. https://doi.org/10.1016/j.jsames.2018.03.002
Deiss, C. 1939. Cambrian stratigraphy and trilobites of northwestern Montana. Geological Society of America Special Papers, 18:1-135. https://doi.org/10.1130/SPE18-p1
Foglia, R.D. and Vaccari, N.E. 2010. Delamaran trilobites from the La Laja Formation, San Juan, Argentina. Ameghiniana, 47:431-445. https://doi.org/10.5710/AMGH.v47i4.2
Foster, J.R. 1994. A note on depositional environments of the Lower-Middle Cambrian Cadiz Formation, Marble Mountains, California. The Mountain Geologist, 31:29-36.
Foster, J.R. 2011a. A short review of the geology and paleontology of the Cambrian sedimentary rocks of the southern Marble Mountains, Mojave Desert, California. New Mexico Museum of Natural History and Science Bulletin, 53:38-51.
Foster, J.R. 2011b. Trilobites and other fauna from two quarries in the Bright Angel Shale (middle Cambrian, Series 3; Delamaran), Grand Canyon National Park, Arizona. Museum of Northern Arizona Bulletin, 67:99-120.
Fuller, J.E. 1980. A Middle Cambrian fossil locality in the Cadiz Formation, Marble Mountains, California. Southern California Paleontological Society Special Publication, 2:30-33.
Goloboff, P.A. and Catalano, S.A. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics, 32:221-238. https://doi.org/10.1111/cla.12160
Harrington, H.J., Henningsmoen, G., Howell, B.F., Jaanusson, V., Lochman-Balk, C., Moore, R.C., Poulsen, C., Rasetti, F., Richter, E., Richter, R., Schmidt, H., Sdzuy, K., Struve, W., Størmer, L., Stubblefield, C.J., Tripp, R., Weller, J.M., and Whittington, H.B. 1959. Arthropoda - General features, Protarthropoda, Euarthropoda - General features, Trilobitomorpha. Treatise on Invertebrate Paleontology, Part O, Arthropoda 1, Geological Society of America and University of Kansas Press, 560 p.
Hazzard, J.C. and Crickmay, C.H. 1933. Notes on the Cambrian rocks of the eastern Mojave Desert, California, with a paleontological report. University of California Publications in Geological Sciences, 23:57-77.
Hazzard, J.C. and Mason, J.F. 1936. Middle Cambrian formations of the Providence and Marble Mountains, California. Geological Society of America Bulletin, 47:229-240. https://doi.org/10.1130/GSAB-47-229
Jell, P.A. and Adrain, J.M. 2002. Available generic names for trilobites. Memoirs of the Queensland Museum, 48:331-553.
Kenny, M. 2011. Geologic structural evaluation of the Fenner Gap region located between the southern Marble Mountains and Ship Mountains, San Bernardino County, California. Santa Margarita Water District Cadiz Valley Water Conservation, Recovery, and Storage Project, Draft Environmental Impact Report, Appendix B, 81 p.
Lochman, C. 1952. Trilobites. In, Cooper, G.A., Cambrian stratigraphy and paleontology near Caborca, northwestern Sonora, Mexico. Smithsonian Miscellaneous Collections, 119(1):1-181.
Mason, J.F. 1935. Fauna of the Cambrian Cadiz Formation, Marble Mountains, California. Bulletin of the Southern California Academy of Sciences, 34:97-114.
McKee, E.D. and Resser, C.E. 1945. Cambrian history of the Grand Canyon region. Carnegie Institution of Washington Publication 563, 232 p.
Morgan, C.A. 2021. Quantitative trilobite biostratigraphy of the middle Cambrian (Wuliuan Stage; Miaolingian Series) and microfacies analysis of the middle Cambrian Stephen Formation, southern Canadian Rocky Mountains. Unpublished PhD dissertation, University of Calgary, Alberta, 445 p. https://doi.org/10.11575/PRISM/39018
Oldroyd, J.D. 1973. Biostratigraphy of the Cambrian Glossopleura zone, west-central Utah. Unpublished MSc thesis, University of Utah, Salt Lake City, 140 p.
Palmer, A.R. 1954. An appraisal of the Great Basin Middle Cambrian trilobites described before 1900. United States Geological Survey Professional Paper 264-D, p. 55-86. https://doi.org/10.3133/pp264d
Palmer, A.R. and Halley, R.B. 1979. Physical stratigraphy and trilobite biostratigraphy of the Carrara Formation (Lower and Middle Cambrian) in the southern Great Basin. United States Geological Survey Professional Paper 1047, 131 p. https://doi.org/10.3133/pp1047
Poulsen, C. 1927. The Cambrian, Ozarkian, and Canadian faunas of northwest Greenland. Meddelelser om Grønland, 70:233-343.
Poulsen, V. 1964. Contribution to the Lower and Middle Cambrian paleontology and stratigraphy of northwest Greenland. Meddelelser om Grønland, 164(6):1-105.
Rasetti, F. 1951. Middle Cambrian stratigraphy and faunas of the Canadian Rocky Mountains. Smithsonian Miscellaneous Collections, 116(5):1-277.
Resser, C.E. 1928. Cambrian fossils from the Mohave Desert. Smithsonian Miscellaneous Collections, 81(2):1-14.
Resser, C.E. 1935. Nomenclature of some Cambrian trilobites. Smithsonian Miscellaneous Collections, 93(5):1-46.
Resser, C.E. 1939. The Spence Shale and its fauna. Smithsonian Miscellaneous Collections, 97(12):1-29.
Robison, R.A. and Babcock, L.E. 2011. Systematics, paleobiology, and taphonomy of some exceptionally preserved trilobites from Cambrian Lagerstätten of Utah. Paleontological Contributions, University of Kansas, 5:1-47. https://doi.org/10.17161/PC.1808.8543
Robison, R.A., Babcock, L.E., and Gunther, V.G. 2015. Exceptional Cambrian fossils from Utah: a window into the age of trilobites. Utah Geological Survey Miscellaneous Publication 15-1, 97 p. https://doi.org/10.34191/mp-15-1
Schwimmer, D.R. 1973. The Middle Cambrian biostratigraphy of Montana and Wyoming. Unpublished PhD dissertation, State University of New York, Stony Brook, 452 p.
Susuki, T. and Lumsden, W.W. 1962. The occurrence of Sonoraspis californica from east-central Nevada. Bulletin of the Southern California Academy of Sciences, 61:233-239.
Stone, P., Miller, D.M., Stevens, C.H., Rosario, J., Vazquez, J.A., Wan, E., Priest, S.S., and Valin, Z.C. 2017. Geologic map of the Providence Mountains in parts of the Fountain Peak and adjacent 7.5’ quadrangles, San Bernardino County, California. United States Geological Survey, Scientific Investigations Map 3376. https://doi.org/10.3133/sim3376
Stoyanow, A. 1952. The original collection of Cambrian trilobites from Sonora. In, Cooper, G.A., Cambrian stratigraphy and paleontology near Caborca, northwestern Sonora, Mexico. Smithsonian Miscellaneous Collections, 119(1):1-181.
Stoyanow, A. 1956. Types of Bathyuriscus howelli var. lodensis Clark. Bulletin of the Geological Society of America, 67:679-682. https://doi.org/10.1130/0016-7606(1956)67[679:TOBHVL]2.0.CO;2
Stoyanow, A. 1958. Sonoraspis and Albertella in the Inyo Mountains, California. Bulletin of the Geological Society of America, 69:347-352. https://doi.org/10.1130/0016-7606(1958)69[347:SAAITI]2.0.CO;2
Stoyanow, A. and Susuki, T. 1955. Discovery of Sonoraspis in southern California. Bulletin of the Geological Society of America, 66:467-470. https://doi.org/10.1130/0016-7606(1955)66[467:dosisc]2.0.co;2
Sundberg, F.A. 2005. The Topazan stage, a new Laurentian stage (Lincolnian series - “Middle” Cambrian). Journal of Paleontology, 79:63-71. https://doi.org/10.1666/0022-3360(2005)079<0063:TTSANL>2.0.CO;2
Sundberg, F.A. 2018. Trilobite biostratigraphy of the Cambrian 5 and Drumian stages, Series 3 (Laurentian Delamaran, Topazan, and Marjuman stages, Lincolnian Series) of the lower Emigrant Formation at Clayton Ridge, Esmeralda County, Nevada. Journal of Paleontology, 92 (Memoir 76):1-44. https://doi.org/10.1017/jpa.2017.130
Walcott, C.D. 1916a. Cambrian geology and paleontology, III, n. 5, Cambrian trilobites. Smithsonian Miscellaneous Collections, 64(5):303-456.
Walcott, C.D. 1916b. Cambrian geology and paleontology, III, n. 3, Cambrian trilobites. Smithsonian Miscellaneous Collections, 64(3):157-258.
Walcott, C.D. 1924. Cambrian geology and paleontology, V, n. 2, Cambrian and lower Ozarkian trilobites. Smithsonian Miscellaneous Collections, 75(2):53-60.
Walcott, C.D. 1925. Cambrian geology and paleontology, V, n. 3, Cambrian and Ozarkian trilobites. Smithsonian Miscellaneous Collections, 75(3):61-146.
