

 Resurrecting the taxon Diatryma: A review of the giant flightless Eocene Gastornithiformes (Aves), with a report of the first skull of Diatryma geiselensis
Resurrecting the taxon Diatryma: A review of the giant flightless Eocene Gastornithiformes (Aves), with a report of the first skull of Diatryma geiselensis
Article number: 27.3.a57
https://doi.org/10.26879/1438
Copyright Palaeontological Association, December 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 16 August 2024. Acceptance: 7 December 2024.
ABSTRACT
The middle Eocene coal seams of the Geisel valley in Germany have yielded several remains of giant flightless birds of the taxon Gastornithiformes. These fossils were described as Diatryma geiselensis Fischer, 1978 and represent the latest occurrence of gastornithiforms. Initiated by the discovery of a previously unrecognized, largely complete albeit strongly flattened skull, we revisit the gastornithiform material from the Geisel valley. Most recent authors assigned D. geiselensis and all other gastornithiforms to the taxon Gastornis Hébert, 1855, which has taxonomic precedence over Diatryma Cope, 1876. In this paper we show that D. geiselensis distinctly differs from the late Paleocene and early Eocene Gastornis parisiensis Hébert, 1855 in the morphologies of the scapulocoracoid and tarsometatarsus. We therefore re-establish the taxon Diatryma and comment on its oldest European record, based on a new tarsometatarsus from the British London Clay. The new D. geiselensis skull from the Geisel valley represents only the second skull of a gastornithiform known so far. The upper beak of this specimen differs from a previously reported partial beak of D. geiselensis in a less vaulted culmen. Together with pronounced size differences, this may indicate that D. geiselensis was sexually dimorphic. We also report the first in situ preserved gastrolith in a specimen of D. geiselensis and comment on some aspects of the paleobiology of gastornithiforms.
Gerald Mayr. Ornithological Section, Senckenberg Research Institute and Natural History Museum Frankfurt, Senckenberganlage 25, 60325 Frankfurt am Main, Germany. Corresponding author. Gerald.Mayr@senckenberg.de
Cécile Mourer-Chauviré. Université Claude Bernard Lyon 1, Laboratoire de Géologie de Lyon: Terre, Planètes, Environnement, UMR CNRS 5276, F-69622 Villeurbanne, France. cecile.mourer@gmail.com
Estelle Bourdon. Laboratoire Informatique et Systématique, Institut de Systématique, Évolution, Biodiversité, UMR CNRS 7205, F-75005 Paris, France. esguelay@gmail.com
Michael Stache. Natural Sciences Collections, Geiseltalmuseum of Martin-Luther-Universität Halle-Wittenberg, Halle/Saale, Germany. michael.stache@zns.uni-halle.de
Keywords: evolution; fossil birds; Gastornis; paleobiology; systematics; taxonomy
Final citation: Mayr, Gerald, Mourer-Chauviré, Cécile, Bourdon, Estelle, and Stache, Michael. 2024. Resurrecting the taxon Diatryma: A review of the giant flightless Eocene Gastornithiformes (Aves), with a report of the first skull of Diatryma geiselensis. Palaeontologia Electronica, 28(3):a57.
https://doi.org/10.26879/1438
palaeo-electronica.org/content/2024/5393-review-of-gastornithiformes
Copyright: December 2024 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Some of the most unusual birds that populated the Northern Hemisphere in the early Cenozoic belong to the Gastornithiformes. Except for a distal tibiotarsus from the early Eocene of central China, which was described as Zhongyuanus xichuanensis by Hou (1980), all fossils of these large flightless birds stem from the late Paleocene and Eocene of Europe and North America. Until recently (see below), these latter specimens were assigned to the taxa Gastornis Hébert, 1855 and Diatryma Cope, 1876, within two distinct higher-level clades, Gastornithidae Fürbringer, 1888 and Diatrymidae Shufeldt, 1913 (e.g., Lambrecht, 1933; Brodkorb, 1967; Andors, 1992).
Gastornithiforms became known to science in 1855, when Hébert described a femur and a tibiotarsus (the lectotype) of a giant flightless bird from the early Eocene (earliest Ypresian; MP 7) of Meudon near Paris as Gastornis parisiensis Hébert, 1855, with the genus name honoring the finder of the fossil, Gaston Planté. Further remains of G. parisiensis from the early Eocene of Passy, also near Paris, were reported by Milne-Edwards (1867) and include fragments of the tarsometatarsus.
Only a few years later, Cope (1876) described a proximal tarsometatarsus and two tarsometatarsal trochleae of a similar bird from early Eocene strata of North America as Diatryma gigantea Cope, 1876. Since then, a fairly comprehensive fossil record of gastornithiforms was assembled from North American fossil sites, which was reviewed by Andors (1988). All North American species stem from early Eocene (Wasatchian) sites and were assigned to the taxon Diatryma. Andors (1988, 1992) considered two species to be valid, Diatryma gigantea and D. regens (Marsh, 1894). Of D. gigantea, a nearly complete skeleton, initially described as Diatryma steini Matthew and Granger, 1917, is known and well figured (Matthew and Granger, 1917), upon which much of our knowledge of the anatomy of gastornithiforms is based; in addition, multiple isolated bones of this species were found. By contrast, D. regens is represented by a single pedal phalanx, which constitutes the holotype of this species. A pedal phalanx of a gastornithiform was also found in the early Eocene of Ellesmere Island in Canada (West and Dawson, 1978; Stidham and Eberle, 2016).
After Hébert’s (1855) description of the Gastornis parisiensis material from the Paris area, gastornithiform remains were also found in other late Paleocene and Eocene localities in Europe. However, for a long time the European fossil record of these birds mainly consisted of fragmentary bones and was afflicted with erroneous assumptions and reconstructions of earlier authors. In particular, much confusion was introduced by Lemoine (1878, 1881), who erected a second species of Gastornis, G. edwardsii Lemoine, 1878, on the basis of various fragmentary bones from the late Paleocene (Thanetian, MP 6) of Cernay-lès-Reims in France. Martin (1992) showed that many of these specimens were non-avian and that Lemoine’s reconstruction of Gastornis was patently incorrect, which prevented earlier authors from recognizing the close similarities of the fossils assigned to Diatryma and Gastornis. Additional gastornithiform bones from the Reims area (Cernay-lès-Reims and Mont de Berru) were collected in later years, and the material now known from these localities constitutes one of the richest late Paleocene assemblages of Gastornis (Martin, 1992; Buffetaut, 1997; Angst and Buffetaut, 2013).
A distal end of a femur from the early Eocene of Belgium was assigned to Gastornis edwardsii by Dollo (1883), and proximal femora and distal tibiotarsi from the early Eocene (earliest Ypresian; MP 7) Reading Formation of Croydon near London were described as G. klaasseni Newton, 1885 (Newton, 1885, 1886; Harrison and Walker, 1977). Gastornis edwardsii and G. klaasseni were synonymized with G. parisiensis by Martin (1992), who described a much smaller species from the late Paleocene of Mont de Berru as G. russelli Martin, 1992. This species may be conspecific with G. minor, which was erected by Lemoine (1878) on the basis of a fragmentary distal tibiotarsus from Cernay-lès-Reims and was considered a nomen dubium by Martin (1992).
Mourer-Chauviré and Bourdon (2016) reported a third, as yet unnamed late Paleocene gastornithiform species from Louvois in the Reims area, which is larger than Gastornis russelli and differs from G. parisiensis in some morphological details. A small fragment of a putative gastornithiform vertebra from the late Paleocene of Rivecourt in northern France was described by Buffetaut and de Ploëg (2020).
The material from the Reims area includes the oldest formally named gastornithiforms, but somewhat older remains of comparatively small species stem from the late Paleocene (Selandian) of Walbeck in Germany (Weigelt, 1939; Mayr, 2007) and Maret in Belgium (Mayr and Smith, 2019). These unnamed species are merely represented by a coracoid (from Walbeck) and a fragmentary femur (from Maret), but Weigelt (1939: 525) also reported “ungual phalanges and vertebrae of the ostrich-sized Gastornis” [our translation] from Walbeck, which could not be located and may be lost.
Recently, a new gastornithiform species from the early Eocene (Ypresian; MP 8‒9) of the La Borie site near Saint-Papoul in southern France was described as Gastornis laurenti by Mourer-Chauviré and Bourdon (2020). Unlike other early Eocene gastornithiforms from Europe, it is well represented by skeletal elements from different body parts including various skull remains. Bones of this species were initially assigned to G. parisiensis (Buffetaut, 2008; Bourdon et al., 2016), and Mourer-Chauviré et al. (2024) reported further material, which support its distinctness.
Martin (1992) considered Gastornis and Diatryma to be distinct taxa, but only identified rather subtle characters of the tibiotarsus as distinguishing features. Buffetaut (1997) emphasized the similarities between Gastornis and Diatryma, and the latter was synonymized with the former by Mlíkovský (2002: 95). Subsequent authors followed this proposal and assigned all gastornithiforms (including Zhongyuanus xichuanensis) to the taxon Gastornis (Buffetaut, 2008, 2013; Mayr, 2009, 2022; Hellmund, 2013; Bourdon et al., 2016; Mourer-Chauviré and Bourdon, 2016, 2020; Angst and Buffetaut, 2017; Worthy et al., 2017; Agnolin, 2021; Mourer-Chauviré et al., 2024). As detailed below, we no longer consider this classification to be justified and resurrect the taxon Diatryma.
European records of Diatryma stem from late early and middle Eocene sites. A partial distal tarsometatarsus from the early Eocene (late Ypresian; MP 10) of Monthelon (France), which lacks the trochlea metatarsi IV, was described as ? Diatryma sarasini by Schaub (1929); the question marks indicate that the assignment to the taxon Diatryma was considered uncertain in the original publication, but gastornithiform affinities of the specimen are uncontested. The latter is not true for “Diatryma” cotei Gaillard, 1937, which is based on a distal tarsometatarsus and some pedal phalanges from the middle Eocene of France (Gaillard, 1937) and was - presumably erroneously (Andors, 1988; Mayr, 2009, 2022) - referred to the Gastornithiformes; the species was likened to the Phorusrhacidae by Angst et al. (2013), but this classification was likewise challenged (Mayr, 2022).
A natural mold of a femur from the latest early or earliest middle Eocene (Lutetian; MP 11) of Messel in Germany was reported by Berg (1965). This singular find is unusual in its preservation as a natural mold in a concretion, which distinguishes it from other fossils from the locality. Berg (1965) assigned the Messel gastornithiform to “Diatryma steini” (= D. gigantea), but Peters (1988) considered that it possibly belongs to the coeval D. geiselensis.
Apart from middle and late Eocene footprints of disputed identity (Buffetaut, 2004; Patterson and Lockley, 2004; see also Mustoe et al., 2012), Diatryma geiselensis is the youngest record of the Gastornithiformes. The species was described by Fischer (1978) from the middle Eocene (Lutetian; MP 11‒13) coal seams of the Geisel valley (Geiseltal) in Germany. Various localities of this area yielded well-preserved gastornithiform material, which was excavated in the 1950s and 1960s and studied by Fischer (1962, 1978). Mlíkovský (2002) synonymized D. geiselensis with D. sarasini, listing it as Gastornis sarasini (Schaub, 1929), but most recent authors, e.g., Hellmund (2013), did not follow this taxonomic proposal and detailed comparisons between the material assigned to D. geiselensis and D. sarasini have not yet been performed.
The gastornithiform remains were omitted from previous revisions of the middle Eocene avifauna from the Geisel valley (Mayr, 2002, 2020), but a survey of the material was published by Hellmund (2013), who amended Fisher’s earlier studies by giving a comprehensive overview of the Diatryma geiselensis fossils and by assigning several of the bones to particular individuals. Here, we report an as yet unrecognized largely complete skull of D. geiselensis from the Geisel valley, which is only the second skull of a gastornithiform known so far. We revise the taxonomic status and phylogenetic affinities of D. geiselensis and comment on previously unnoticed aspects of the gastornithiform material of the Geisel valley, which are of paleobiological interest.
MATERIAL AND METHODS
The fossils are deposited in the collections of the American Museum of Natural History, New York, USA (AMNH); the Natural Sciences Collections, Geiseltalmuseum of Martin-Luther-Universität Halle-Wittenberg, Halle/Saale, Germany (GMH); the Natural Sciences Collections, Geowissenschaftliche Sammlung of Martin-Luther-Universität Halle-Wittenberg, Halle/Saale, Germany (MLU_GeoS); the Muséum National d’Histoire Naturelle, Paris, France (MNHN); the Muséum d’Histoire Naturelle de Toulouse, France (MHNT), and the National Museum of Natural History, Smithsonian Institution, Washington D.C., USA (USNM).
The anatomical terminology follows Baumel and Witmer (1993). Three-dimensional (3D) surface scans are made available for three of the Geisel valley gastornithiforms: GMH XVIII-1178-1958 (https://doi.org/10.25673/117207), GMH XLI-200[15]-1968 (https://doi.org/10.25673/117208), and GMH XLI-200[14]-1968: (https://doi.org/10.25673/117209).
Roman numerals in the inventory numbers of the vertebrate fossils from the Geisel valley in GMH indicate particular stratigraphic sections. For the Gastornis fossils, six localities are of relevance, which span a stratigraphic range from MP 11 to MP 13 (early to late Lutetian), that is, have an age of about 47.5 to 43.5 million years ago [Ma] (Vandenberghe et al., 2012) and therefore cover an interval of about four million years. The “Unterkohle” (“basal coal seam”; MP 11) occurred at sites XIII and XIV of the Neumark West mine, the “untere Mittelkohle” (“lower middle coal seam”; MP 12) was quarried at sites XVIII of Neumark Süd and IL of Geiselröhlitz, and the “obere Mittelkohle” (“upper middle coal seam”; MP 13) occurred at sites XXXV of Neumark Süd and XLI of Geiselröhlitz (Krumbiegel et al., 1983; Hellmund, 1996).
SYSTEMATIC PALEONTOLOGY
Class AVES Linnaeus, 1758
Order GASTORNITHIFORMES Stejneger, 1885
Family GASTORNITHIDAE Fürbringer, 1888
Genus DIATRYMA Cope, 1876
Type species. Diatryma gigantea Cope, 1876, by monotypy.
Emended differential diagnosis. Differs from Gastornis Hébert, 1855 (as exemplified by the specimens from the Reims area, which were originally assigned to G. edwardsii) in that the scapulocoracoid is straight, with essentially no mediolateral or dorsoventral curvature; the body of the coracoid is dorsoventrally flattened (curved in Gastornis) and pierced by a large foramen (which may or may not be homologous to the foramen nervi supracoracoidei; see description); the tarsometatarsus is stouter, with an elongation index (ratio length of bone to mediolateral width of proximal end) of less than 2.8 (2.9‒3.8 in the species of Gastornis), the proximal and distal ends are much wider mediolaterally than the shaft, the midsection of the shaft shows a mediolateral constriction, and the trochlea metatarsi IV is more laterally splayed and the incisura intertrochlearis lateralis is wider.
Remarks. The above diagnosis is based on material referred to Diatryma gigantea, even though all of the above-listed diagnostic features also pertain to D. geiselensis. For Gastornis parisiensis, the type species of the taxon Gastornis, comparisons are made with material originally assigned to G. edwardsii.
Diatryma geiselensis Fischer, 1978
Differential diagnosis. The gastornithiform species from the Geisel valley differs from:
Gastornis parisiensis Hébert, 1855 in: mandibular rami dorsoventrally deeper and processus coronoideus better developed; rudiments of furcula absent; trochlea metatarsi II reaching farther distally (other differences concerning the morphology of the scapulocoracoid and the tarsometatarsus are mentioned in the differential diagnosis of the taxon Diatryma).
Gastornis russelli Martin, 1992 in: being much larger; trochlea metatarsi II reaching farther distally (other differences concerning the morphology of the tarsometatarsus are mentioned in the differential diagnosis of the taxon Diatryma).
Gastornis laurenti Mourer-Chauviré and Bourdon, 2020 in: upper beak proportionally longer; mandibular rami dorsoventrally deeper and processus coronoideus better developed; trochlea metatarsi II reaching farther distally (other differences concerning the morphology of the tarsometatarsus are mentioned in the differential diagnosis of the taxon Diatryma).
Diatryma sarasini Schaub, 1929 in: trochlea metatarsi II proportionally larger and reaching farther distally; trochlea metatarsi III not asymmetrical (in D. sarasini the lateral rim of this trochlea reaches farther distally than the medial one).
Diatryma gigantea Cope, 1876 in: mandible dorsoventrally deeper; furcula absent or more strongly co-ossified with scapulocoracoid (see description); body of coracoid pierced by a large foramen; tarsometatarsus stouter; trochlea metatarsi II proportionally larger.
Remarks. Meaningful comparisons with Diatryma regens, of which only a single pedal phalanx is known (Marsh, 1894), are not possible. Zhongyuanus xichuanensis is based on a distal tibiotarsus (Hou, 1980). This element exhibits a similar morphology in all gastornithiforms and shows some damage owing to compression and breakage in all D. geiselensis specimens, so that only limited comparisons with Z. xichuanensis are possible.
Referred specimens. Fischer (1962, 1978) made only few attempts to assign to particular individuals the various isolated bones he described. Hellmund (2013) presented a clearer overview of the association of these remains and estimated that they represent nine individuals (the new skull described in the present study was not included). Because some specimens lack exact stratigraphic data, there remains a possibility that they stem from individuals with such data, and we determine a minimum number of eight Diatryma geiselensis individuals in the collection of GMH (including the new skull).
GMH Dia 2 (holotype scapulocoracoid); there are no exact stratigraphic data, but Fischer (1962, 1978) noted that the specimen is likely to be from site XIII, so that it possibly belongs to the same individual as GMH XIII-40-1954.
GMH Dia 3 (right humerus lacking distal end); there are no exact stratigraphic data, but Fischer (1962) noted that the specimen may be from site XIII, so that it possibly belongs to the same individual as GMH XIII-40-1954.
GMH Dia 8 (left femur); Hellmund (2013) considered the exact provenance of this specimen to be uncertain, but Fischer (1962) indicated that it stems from the “Unterkohle” of Neumark West and may belong to the same individual as GMH XIII-40-1954.
GMH Dia 12 (left tarsometatarsus); no exact stratigraphic data exist, but the specimen may stem from one of the individuals found in the “Unterkohle” of Neumark West.
GMH XIII-40-1954 (right femur, right tibiotarsus); from Neumark West (“Unterkohle”, early Lutetian, MP 11), found in 1954.
GMH XIII-108-1954 (rib); from Neumark West (“Unterkohle”, early Lutetian, MP 11), found in 1954; whether the specimen belongs to the same individual as GMH XIII-40-1954 cannot be determined.
GMH XIV-258-1954 (proximal end of left tibiotarsus); from Neumark West (“Unterkohle”, early Lutetian, MP 11), found in 1954.
GMH XIV-2476-1955 (proximal and distal ends of right femur); from Neumark West (“Unterkohle”, early Lutetian, MP 11), found in 1955.
GMH XIV-4730-1956 (partial mandible), GMH XIV-658-1956 (partial pelvis), GMH XIV-2042-1956 (left femur), GMH XIV-4628-1956 (right femur), GMH XIV-2091-1956 (left tibiotarsus), GMH XIV-4005-1956 (pedal phalanx), from Neumark West (“Unterkohle”, early Lutetian, MP 11), found in 1956.
GMH XVIII-1178-1958 (skull); from Neumark Süd (“untere Mittelkohle”, middle Lutetian, MP 12), found in 1958.
GMH IL-38-1969 (pedal phalanges); from Geiselröhlitz (“untere Mittelkohle”, middle Lutetian, MP 12), found in 1969.
GMH XXXV-481-1963 (right femur, left tibiotarsus) and GMH XXXV-202-1963 (right tibiotarsus); from Neumark Süd (“obere Mittelkohle”, late Lutetian, MP 13), found in 1963; contrary to Hellmund (2013), we consider it likely that both tibiotarsi are from the same individual.
GMH XLI-200-1968 (partial upper beak, mandible, atlas and three further vertebrae, rib, partial scapulocoracoid, partial pelvis, right femur and partial left femur, right tibiotarsus and distal portion of left tibiotarsus, right tarsometatarsus, six pedal phalanges); from Geiselröhlitz (“obere Mittelkohle”, late Lutetian, MP 13), found in 1968.
Remarks. Fischer (1967) reported a putative partial gastornithiform sternum from the Geisel valley (GMH XIV-3979c-1956), but this specimen was misidentified and actually is a right ilium of the perissodactyl mammal Lophiodon. Incidentally, the specimen was published in a study, in which metatarsals of Lophiodon were introduced as a new avian taxon (“Saurornis matthewsi”); this latter identification was corrected by Fischer (1987), whereas the sternum was still assigned to the Gastornithiformes by Hellmund (2013). A putative gastornithiform humerus (GMH XIV-3979a-1956), which was listed by Hellmund (2013), is the right radius of Lophiodon. A putative scapulocoracoid (GMH XIV-3979b-1956), also listed by Hellmund (2013), is likewise misidentified even though its true identity is elusive.
 Notes on the comparative osteology of Diatryma geiselensis. Fischer (1978) described a partial upper beak (rostrum) of D. geiselensis, which belongs to the most complete individual GMH XLI-200-1968 (Figure 1C-F). Together with two partial mandibles, this specimen until now constituted the only skull remains of D. geiselensis. Here we report a nearly complete but strongly flattened skull from the collection of GMH, which was stored in two parts (neurocranium and upper beak) that have now been reassembled (Figure 1A-B). The specimen (GMH XVIII-1178-1958) was mistaken for crocodilian remains ‒ as per the index card of the fossil ‒ and is from a smaller individual than the previously known D. geiselensis beak and mandibles. The fossil is only the second complete skull of a gastornithiform known so far, with the other being that of the D. gigantea skeleton described by Matthew and Granger (1917). Unfortunately, GMH XVIII-1178-1958 is strongly flattened and the bone surfaces of the specimen are shattered throughout, so that subtle osteological details cannot be recognized. The maximum length of GMH XVIII-1178-1958, from the caudal end of the left processus paroccipitalis to the tip of the beak, is 347 mm.
Notes on the comparative osteology of Diatryma geiselensis. Fischer (1978) described a partial upper beak (rostrum) of D. geiselensis, which belongs to the most complete individual GMH XLI-200-1968 (Figure 1C-F). Together with two partial mandibles, this specimen until now constituted the only skull remains of D. geiselensis. Here we report a nearly complete but strongly flattened skull from the collection of GMH, which was stored in two parts (neurocranium and upper beak) that have now been reassembled (Figure 1A-B). The specimen (GMH XVIII-1178-1958) was mistaken for crocodilian remains ‒ as per the index card of the fossil ‒ and is from a smaller individual than the previously known D. geiselensis beak and mandibles. The fossil is only the second complete skull of a gastornithiform known so far, with the other being that of the D. gigantea skeleton described by Matthew and Granger (1917). Unfortunately, GMH XVIII-1178-1958 is strongly flattened and the bone surfaces of the specimen are shattered throughout, so that subtle osteological details cannot be recognized. The maximum length of GMH XVIII-1178-1958, from the caudal end of the left processus paroccipitalis to the tip of the beak, is 347 mm.
 The culmen (dorsal ridge) of the upper beak of GMH XVIII-1178-1958 is less dorsally vaulted than in Diatryma gigantea and the tomia (cutting edges) are more strongly sigmoidally curved (Figure 2A, C). The beak of this specimen is also less strongly dorsally vaulted than in the partial beak GMH XLI-200-1968 (Figure 1G), which was described by Fischer (1978). The rostrum of the latter specimen (GMH XLI-200-1968) is mediolaterally wider than in D. gigantea (Figure 1F, H); owing to the flattening of GMH XVIII-1178-1958, the width of its beak cannot be reliably assessed. The bone surface of the D. geiselensis beak exhibits numerous neurovascular foramina and vascular grooves. Mourer-Chauviré and Bourdon (2020) already commented on the shape of the nostril of D. geiselensis, which is preserved on the right side of specimen GMH XLI-200-1968, but was not figured by Fischer (1978). The nostrils of GMH XVIII-1178-1958 conform to those of GMH XLI-200-1968. They are of similar size to the nostrils of D. gigantea and open rostrally into shallow furrows at mid-height on the rostrum level with the quadratojugal bar (Figure 1D); these furrows were also noted for G. laurenti and D. gigantea by Bourdon et al. (2016). The damaged surface of the upper beak of GMH XLI-200-1968 shows a “spongy” interior.
The culmen (dorsal ridge) of the upper beak of GMH XVIII-1178-1958 is less dorsally vaulted than in Diatryma gigantea and the tomia (cutting edges) are more strongly sigmoidally curved (Figure 2A, C). The beak of this specimen is also less strongly dorsally vaulted than in the partial beak GMH XLI-200-1968 (Figure 1G), which was described by Fischer (1978). The rostrum of the latter specimen (GMH XLI-200-1968) is mediolaterally wider than in D. gigantea (Figure 1F, H); owing to the flattening of GMH XVIII-1178-1958, the width of its beak cannot be reliably assessed. The bone surface of the D. geiselensis beak exhibits numerous neurovascular foramina and vascular grooves. Mourer-Chauviré and Bourdon (2020) already commented on the shape of the nostril of D. geiselensis, which is preserved on the right side of specimen GMH XLI-200-1968, but was not figured by Fischer (1978). The nostrils of GMH XVIII-1178-1958 conform to those of GMH XLI-200-1968. They are of similar size to the nostrils of D. gigantea and open rostrally into shallow furrows at mid-height on the rostrum level with the quadratojugal bar (Figure 1D); these furrows were also noted for G. laurenti and D. gigantea by Bourdon et al. (2016). The damaged surface of the upper beak of GMH XLI-200-1968 shows a “spongy” interior.
As in Gastornis laurenti and Diatryma gigantea (see Bourdon et al., 2016), there are a distinct nasofrontal hinge and lateral projections at the base of the upper beak. GMH XVIII-1178-1958 lacks the bones of the palate and the jugal bars are likewise not preserved. The orbits are not clearly visible in the specimen, but must have been situated rostral to the large processus postorbitalis (Figure 2A). As in D. gigantea, the neurocranium is strikingly short relative to the entire length of the skull. Andors (1988: 135) noted a peculiar rostral restriction of the orbit of D. gigantea, which he considered to possibly be the result of a hyperdevelopment of the fossae temporales. The fossae temporales of GMH XVIII-1178-1958 are likewise very deep. In D. gigantea, a massive bony bridge connects the processus zygomaticus with the processus postorbitalis and forms a secondary temporal opening. This orbitozygomatic junction (sensu Elzanowski and Mayr, 2018) is not visible in GMH XVIII-1178-1958 and appears to be broken. The condylus occipitalis is very large and caudally protruding, as it also is in D. gigantea.
 Two gastornithiform mandibles, which are from specimens GMH XIV-4730-1956 and GMH XLI-200-1968, are known from the Geisel valley (Figure 3A-E). In both, the rami are dorsoventrally deeper than in Gastornis laurenti, G. parisiensis (Figure 3F-G), and Diatryma gigantea (Figure 2C). In both specimens from the Geisel valley, the symphysis mandibulae is wider than in G. laurenti, G. parisiensis, and D. gigantea; whether this is an artefact of preservation is difficult to ascertain. The processus coronoideus is more prominent than in G. parisiensis and G. laurenti (see Mourer-Chauviré and Bourdon, 2020: figure 1) and narrower than in D. gigantea. As noted by Fischer (1962, 1978), the mandibular surface is distinctly pitted by neurovascular foramina and covered with vascular grooves. In both specimens, the rostral tip of the mandible is more strongly raised dorsally than in G. parisiensis.
Two gastornithiform mandibles, which are from specimens GMH XIV-4730-1956 and GMH XLI-200-1968, are known from the Geisel valley (Figure 3A-E). In both, the rami are dorsoventrally deeper than in Gastornis laurenti, G. parisiensis (Figure 3F-G), and Diatryma gigantea (Figure 2C). In both specimens from the Geisel valley, the symphysis mandibulae is wider than in G. laurenti, G. parisiensis, and D. gigantea; whether this is an artefact of preservation is difficult to ascertain. The processus coronoideus is more prominent than in G. parisiensis and G. laurenti (see Mourer-Chauviré and Bourdon, 2020: figure 1) and narrower than in D. gigantea. As noted by Fischer (1962, 1978), the mandibular surface is distinctly pitted by neurovascular foramina and covered with vascular grooves. In both specimens, the rostral tip of the mandible is more strongly raised dorsally than in G. parisiensis.
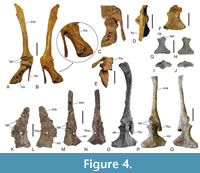 The holotype of Diatryma geiselensis is a scapulocoracoid (Figure 4M-N), which lacks exact collection data but is likely to stem from the “Unterkohle” of Neumark West (Fischer, 1978) and therefore possibly belongs to specimen GMH XIII-40-1954. We identified a second, previously unrecognized partial scapulocoracoid in the collection of GMH, which is from the individual GMH XLI-200-1968 and was stored amongst unsorted material (Figure 4K-L). The specimen is distinctly larger than the scapulocoracoid of the holotype but otherwise shows an identical morphology. Fischer (1962, 1978) noted the presence of a foramen nervi supracoracoidei in D. geiselensis and its absence in D. gigantea; furthermore, unlike in D. geiselensis, there is an elongated process extending from the cranial end of the scapula on the medial side of the scapulocoracoid of D. gigantea (Figure 4O‒Q). This process was considered to be the acromion of the scapula by Matthew and Granger (1917). At the time of Fischer’s studies, no correctly identified scapulocoracoid of Gastornis was known. Martin (1992) showed that the shoulder girdle bones identified by Lemoine (1881) do not belong to Gastornis and are not even of avian origin. Martin (1992) identified a previously undescribed scapulocoracoid of G. parisiensis from Mont de Berru, and photographs of this compound bone, which has a distinctive “boot-shaped” outline, are here for the first time shown (Figure 4A-F). Unlike in D. geiselensis and D. gigantea, the scapula and coracoid of G. parisiensis are angled relative to each other, whereas both bones are situated in the same axis and plane in the Diatryma species. The coracoid of G. parisiensis exhibits a small foramen nervi supracoracoidei, and there is a narrow, rod-shaped process that is likely to represent the shaft of the furcula (Figure 4A-B). Based on comparisons with the scapulocoracoid of G. parisiensis, the stouter process in the same area of the D. gigantea coracoid (which was longer at the time of the discovery of the specimen but has since lost its tip; compare Figure 4O and 4P) probably also represents the omal portion of a co-ossified furcula. Because there is a deep notch in the coracoid of D. gigantea (Figure 4O), it is unclear whether the large foramen piercing the scapulocoracoid of D. geiselensis is indeed a foramen nervi supracoracoidei or whether it represents an opening that formed by fusion of the remnants of the furcular shaft with the coracoid (which would explain the absence of a furcula in D. geiselensis). The facies articularis humeralis of D. geiselensis is less deeply incised into the body of the scapulocoracoid than in D. gigantea. As in G. parisiensis, a slit-like gap separates the ventral portions of the facies articulares humerales of the coracoid and scapula in the holotype coracoid of D. geiselensis. In both, D. geiselensis and D. gigantea, there is a marked longitudinal ridge along the midline of the ventral surface of the corpus of the coracoid.
The holotype of Diatryma geiselensis is a scapulocoracoid (Figure 4M-N), which lacks exact collection data but is likely to stem from the “Unterkohle” of Neumark West (Fischer, 1978) and therefore possibly belongs to specimen GMH XIII-40-1954. We identified a second, previously unrecognized partial scapulocoracoid in the collection of GMH, which is from the individual GMH XLI-200-1968 and was stored amongst unsorted material (Figure 4K-L). The specimen is distinctly larger than the scapulocoracoid of the holotype but otherwise shows an identical morphology. Fischer (1962, 1978) noted the presence of a foramen nervi supracoracoidei in D. geiselensis and its absence in D. gigantea; furthermore, unlike in D. geiselensis, there is an elongated process extending from the cranial end of the scapula on the medial side of the scapulocoracoid of D. gigantea (Figure 4O‒Q). This process was considered to be the acromion of the scapula by Matthew and Granger (1917). At the time of Fischer’s studies, no correctly identified scapulocoracoid of Gastornis was known. Martin (1992) showed that the shoulder girdle bones identified by Lemoine (1881) do not belong to Gastornis and are not even of avian origin. Martin (1992) identified a previously undescribed scapulocoracoid of G. parisiensis from Mont de Berru, and photographs of this compound bone, which has a distinctive “boot-shaped” outline, are here for the first time shown (Figure 4A-F). Unlike in D. geiselensis and D. gigantea, the scapula and coracoid of G. parisiensis are angled relative to each other, whereas both bones are situated in the same axis and plane in the Diatryma species. The coracoid of G. parisiensis exhibits a small foramen nervi supracoracoidei, and there is a narrow, rod-shaped process that is likely to represent the shaft of the furcula (Figure 4A-B). Based on comparisons with the scapulocoracoid of G. parisiensis, the stouter process in the same area of the D. gigantea coracoid (which was longer at the time of the discovery of the specimen but has since lost its tip; compare Figure 4O and 4P) probably also represents the omal portion of a co-ossified furcula. Because there is a deep notch in the coracoid of D. gigantea (Figure 4O), it is unclear whether the large foramen piercing the scapulocoracoid of D. geiselensis is indeed a foramen nervi supracoracoidei or whether it represents an opening that formed by fusion of the remnants of the furcular shaft with the coracoid (which would explain the absence of a furcula in D. geiselensis). The facies articularis humeralis of D. geiselensis is less deeply incised into the body of the scapulocoracoid than in D. gigantea. As in G. parisiensis, a slit-like gap separates the ventral portions of the facies articulares humerales of the coracoid and scapula in the holotype coracoid of D. geiselensis. In both, D. geiselensis and D. gigantea, there is a marked longitudinal ridge along the midline of the ventral surface of the corpus of the coracoid.
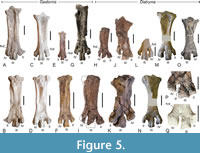 Even though fossils from the Geisel valley are often deformed by diagenetic flattening, this does not severely affect the two Diatryma geiselensis tarsometatarsi, which are largely three-dimensionally preserved, with only moderate dorsoplantar compression. These two tarsometatarsi are stouter than those of the Gastornis species, of which a complete tarsometatarsus is known (G. parisiensis, G. russelli, and G. laurenti; Figure 5). The elongation indices (ratio length of bone to mediolateral width of proximal end) of the two tarsometatarsi from the Geisel valley are ~2.21 (GMH XLI-200-1968) and ~2.78 (GMH Dia 12). For G. parisiensis, this index is 2.96 (MNHN L3092) and 3.32 (MNHN L3093), whereas it is 2.79 in D. gigantea (based on a complete tarsometatarsus figured by Andors, 1988: pl. 17) and 2.52 in a Diatryma -like tarsometatarsus from the London Clay of the Isle of Sheppey (see discussion). The ratio of the proximal (mediolateral) width of the bone to the width of the midsection of the shaft is 1.58‒1.77 in G. parisiensis (MNHN L3092 and MNHN L3093) and 1.97‒2.46 in D. gigantea (the larger value is based on the tarsometatarsus figured by Andors, 1988: pl. 17, the smaller value is from USNM 15118 [based on a photo provided by T. Worthy]). For D. geiselensis, this ratio is difficult to estimate, because part of the proximal end of the tarsometatarsus is broken in GMH Dia 12, whereas the shaft is damaged in GMH XLI-200-1968; for the latter specimen the ratio is ~2.24. The tarsometatarsus of D. geiselensis differs from the distal tarsometatarsus of the D. sarasini holotype in that the trochlea metatarsi II reaches farther distally and the trochlea metatarsi III has a symmetric shape (in G. sarasini, the lateral rim projects farther distally than the medial one). The two largely complete tarsometatarsi of D. geiselensis differ in size and some morphological features (Figure 5I, K). Whereas the foramen vasculare distale is well delimited in GMH Dia 12, it appears to be distally open in GMH XLI-200-1968, and the trochlea metatarsi II is proportionally larger in the smaller specimen GMH Dia 12 than in the larger GMH XLI-200-1968. In both specimens the trochlea metatarsi III is proportionally larger than in the Gastornis tarsometatarsi, and the trochlea metatarsi IV is mediolaterally wider. In GMH XLI-200-1968, the trochlea metatarsi IV reaches less far distally than the trochlea metatarsi II (the condition in GMH Dia 12 cannot be established, because the distal portion of the trochlea metatarsi IV is broken).
Even though fossils from the Geisel valley are often deformed by diagenetic flattening, this does not severely affect the two Diatryma geiselensis tarsometatarsi, which are largely three-dimensionally preserved, with only moderate dorsoplantar compression. These two tarsometatarsi are stouter than those of the Gastornis species, of which a complete tarsometatarsus is known (G. parisiensis, G. russelli, and G. laurenti; Figure 5). The elongation indices (ratio length of bone to mediolateral width of proximal end) of the two tarsometatarsi from the Geisel valley are ~2.21 (GMH XLI-200-1968) and ~2.78 (GMH Dia 12). For G. parisiensis, this index is 2.96 (MNHN L3092) and 3.32 (MNHN L3093), whereas it is 2.79 in D. gigantea (based on a complete tarsometatarsus figured by Andors, 1988: pl. 17) and 2.52 in a Diatryma -like tarsometatarsus from the London Clay of the Isle of Sheppey (see discussion). The ratio of the proximal (mediolateral) width of the bone to the width of the midsection of the shaft is 1.58‒1.77 in G. parisiensis (MNHN L3092 and MNHN L3093) and 1.97‒2.46 in D. gigantea (the larger value is based on the tarsometatarsus figured by Andors, 1988: pl. 17, the smaller value is from USNM 15118 [based on a photo provided by T. Worthy]). For D. geiselensis, this ratio is difficult to estimate, because part of the proximal end of the tarsometatarsus is broken in GMH Dia 12, whereas the shaft is damaged in GMH XLI-200-1968; for the latter specimen the ratio is ~2.24. The tarsometatarsus of D. geiselensis differs from the distal tarsometatarsus of the D. sarasini holotype in that the trochlea metatarsi II reaches farther distally and the trochlea metatarsi III has a symmetric shape (in G. sarasini, the lateral rim projects farther distally than the medial one). The two largely complete tarsometatarsi of D. geiselensis differ in size and some morphological features (Figure 5I, K). Whereas the foramen vasculare distale is well delimited in GMH Dia 12, it appears to be distally open in GMH XLI-200-1968, and the trochlea metatarsi II is proportionally larger in the smaller specimen GMH Dia 12 than in the larger GMH XLI-200-1968. In both specimens the trochlea metatarsi III is proportionally larger than in the Gastornis tarsometatarsi, and the trochlea metatarsi IV is mediolaterally wider. In GMH XLI-200-1968, the trochlea metatarsi IV reaches less far distally than the trochlea metatarsi II (the condition in GMH Dia 12 cannot be established, because the distal portion of the trochlea metatarsi IV is broken).
DISCUSSION
Taxonomy, Interspecific Relationships, and Biogeographic History
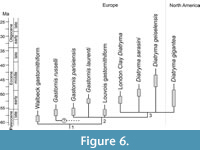 As detailed above, Diatryma geiselensis and the North American D. gigantea differ distinctly from Gastornis parisiensis in features of the coracoid and tarsometatarsus. A clade including D. geiselensis and D. gigantea to the exclusion of G. parisiensis is supported by the straight and dorsoventrally plane scapulocoracoid and the morphology of the tarsometatarsus, which is stouter than in Gastornis, with wider proximal and distal ends and an elongation index of less than 2.8, a mediolaterally waisted midsection of the shaft, a large trochlea metatarsi III, and a wide incisura intertrochlearis lateralis. The latter two features also suggest an assignment of D. sarasini to the Diatryma clade (Figure 6). Outgroup comparisons with other neornithine birds show the above tarsometatarsus morphologies to be derived, and the differences are similar to those distinguishing different genera of moas (e.g., Cracraft, 1976).
As detailed above, Diatryma geiselensis and the North American D. gigantea differ distinctly from Gastornis parisiensis in features of the coracoid and tarsometatarsus. A clade including D. geiselensis and D. gigantea to the exclusion of G. parisiensis is supported by the straight and dorsoventrally plane scapulocoracoid and the morphology of the tarsometatarsus, which is stouter than in Gastornis, with wider proximal and distal ends and an elongation index of less than 2.8, a mediolaterally waisted midsection of the shaft, a large trochlea metatarsi III, and a wide incisura intertrochlearis lateralis. The latter two features also suggest an assignment of D. sarasini to the Diatryma clade (Figure 6). Outgroup comparisons with other neornithine birds show the above tarsometatarsus morphologies to be derived, and the differences are similar to those distinguishing different genera of moas (e.g., Cracraft, 1976).
Our phylogenetic hypothesis is not based on a formal analysis, which is impeded by the fact that some gastornithiforms are represented by non-overlapping skeletal material and fragmentary remains (of the Walbeck species, for example, only a coracoid was described). However, the main conclusion of our study is that the Geisel valley gastornithiform is more closely related to the North American forms than to Gastornis parisiensis. The above-listed character evidence for this placement shows no homoplasy within gastornithiforms, and there exists no morphological evidence in support of a clade including G. parisiensis and Diatryma geiselensis to the exclusion of D. gigantea. By itself, this phylogenetic hypothesis does not necessarily entail taxonomic consequences, but if all gastornithiform species were assigned to the taxon Gastornis, information on the interrelationship of gastornithiforms would not be properly reflected. This is why we advocate re-establishment of the taxon Diatryma for a well-defined subclade of gastornithiforms.
It is less straightforward to assess the interrelationships of other Paleocene and early Eocene Gastornithiformes from Europe, which are currently assigned to the taxon Gastornis. The incomplete femur from Maret in Belgium described by Mayr and Smith (2019) is from a very small species and may belong to G. russelli. The small size of these fossils may be a plesiomorphic characteristic, which supports a position of the Maret gastornithiform and G. russelli outside a clade formed by G. parisiensis, G. laurenti, the Louvois gastornithiform, and the Diatryma species. This hypothesis conforms to the fact that the Maret gastornithiform and G. russelli are among the oldest known gastornithiforms and - if confirmed in future studies - would require an assignment of G. russelli to a new genus-level taxon. Martin (1992) tentatively assigned a beak fragment from the late Paleocene of the Reims area to G. russelli, which differs from other gastornithiform beaks in having very large nostrils (Figure 2E). However, according to previous authors (Martin, 1992; Louchart et al., 2021), this latter specimen is from a juvenile individual, so that the size of the nostrils may have been subjected to ontogenetic changes in gastornithiforms.
The gastornithiform from Walbeck is even older, but it is only known from a coracoid. This bone is not co-ossified with the scapula, which suggests that the Walbeck species is outside a clade formed by Gastornis and Diatryma (Figure 6). However, the bone surface of the Walbeck coracoid has a somewhat porous texture, so that the specimen may stem from an immature individual, in which the coracoid and scapula were not yet fused.
The phylogeny outlined above and shown in Figure 6 supports an Old World origin of the Gastornithiformes. As suggested by earlier authors (Andors, 1992; Buffetaut, 1997; Mayr, 2009; Buffetaut, 2013; Buffetaut and Angst, 2014), these birds are likely to have dispersed into North America via high latitude land corridors. This hypothesis conforms to the discovery of gastornithiform remains on Ellesmere Island in Canada (West and Dawson, 1978; Stidham and Eberle, 2016), but it conflicts with the fact that current records of Diatryma from Europe are significantly younger than those from North America.
The so far oldest Old World gastornithiform that was assigned to the taxon Diatryma is D. sarasini. The holotype and only known specimen of this species is from the late Ypresian (MP 10) of France and therefore dates back some 51‒48 Ma (Vandenberghe et al., 2012). The specimen is still five to eight million years younger than the earliest North American records of the Gastornithiformes, which stem from the early Wasatchian, some 56 Ma.
However, there exists a previously unpublished tarsometatarsus from the early Eocene London Clay of the Isle of Sheppey in a private collection, of which a photograph (Figure 5O) is here shown by permission of its owner, Scott Moore-Fay. This bone is much stouter than the tarsometatarsus of Gastornis parisiensis and shows the distinctive proportions of the tarsometatarsus of the Diatryma species. The London Clay Formation of the Isle of Sheppey spans a time interval from about 53.5 to ca. 51.5 Ma (Ypresian, MP 8‒9; “upper Division C to F”; Collinson et al., 2016). Depending on its exact stratigraphic provenance, the London Clay gastornithiform is therefore at least 3 million younger than the gastornithiform from the Reading Formation (early Ypresian, MP 7; 56 Ma, Vandenberghe et al., 2012) of Croydon near London, which was described as G. klaasseni and is now assigned to G. parisiensis. If the London Clay gastornithiform were conspecific with that from the Reading Formation, G. klaasseni would have to be removed from synonymy with G. parisiensis and transferred to the taxon Diatryma.
Because Diatryma occurs in North America already in the earliest Wasatchian, some 56 Ma (Andors, 1992), the Diatryma lineage is likely to extend into the late Paleocene. A potential candidate taxon is the unnamed late Paleocene Louvois gastornithiform, for which Mourer-Chauviré and Bourdon (2016) noted some resemblance in tarsometatarsus morphology to D. sarasini. Currently, however, the fragmentary material known of the Louvois gastornithiform does not allow an unambiguous phylogenetic placement.
The distal tibiotarsus from the early Eocene of China described as Zhongyuanus xichuanensis by Hou (1980) also cannot be reliably assigned to either Gastornis or Diatryma. Buffetaut (2013) synonymized Zhongyuanus Hou, 1980 with Gastornis Hébert, 1855, which he recognized as the only genus-level taxon within gastornithiforms. Now that Diatryma has been re-established, it is more appropriate to maintain the Chinese species in the taxon Zhongyuanus until its affinities can be clarified.
Agnolin (2007, 2021) suggested close affinities between gastornithiforms and Brontornis burmeisteri from the late Oligocene of Bolivia (Buffetaut, 2017) and the late early Miocene of Patagonia (Argentina). This latter species is usually assigned to the Phorusrhacidae, but it differs from typical phorusrhacids in a much stouter tarsometatarsus, which is of very similar proportions to that of Diatryma (compare 5M, O with Agnolin, 2021: figure 7). The sparse material known of Brontornis does not allow an unambiguous determination of its affinities, but the species differs from gastornithiforms in that the quadrate has a condylus caudalis (Worthy et al., 2017). The rostral portion of the mandible of Brontornis is dorsoventrally deeper than in Gastornis and Diatryma, and unlike in gastornithiforms the distal end of the Brontornis tibiotarsus lacks a pons supratendineus. The latest North American record of Diatryma is from the early Eocene, so that a dispersal to South America must have occurred at that time or before. However, no early Cenozoic land bridges are known that would have facilitated the dispersal of large flightless birds from North to South America. Palaeogeography therefore also challenges close affinities between Brontornis and gastornithiforms.
Paleobiology
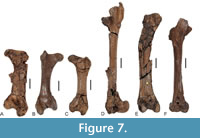 The Gastornis remains from Louvois in France show disparate sizes, which Mourer-Chauviré and Bourdon (2016) considered to be indicative of a sexual size dimorphism of this as yet unnamed species. Angst and Buffetaut (2017) stated that a dimorphism in size has not been demonstrated for other gastornithiforms, but the hindlimb bones of Diatryma geiselensis likewise exhibit pronounced differences in size, and differently-sized individuals were found in the same stratigraphic horizon (e.g., GMH XLI-200-1968 and GMH XXXV-481-1963; Figure 7). Rather than assuming the coexistence of two large flightless birds in the same habitat, we consider it more likely that D. geiselensis was sexually dimorphic in size. The culmen of the upper beak is more vaulted in the larger specimen GMH XLI-200-1968 than in the smaller GMH XVIII-1178-1958 (Figure 1G), so that D. geiselensis also appears to have shown sexual dimorphism in beak shape.
The Gastornis remains from Louvois in France show disparate sizes, which Mourer-Chauviré and Bourdon (2016) considered to be indicative of a sexual size dimorphism of this as yet unnamed species. Angst and Buffetaut (2017) stated that a dimorphism in size has not been demonstrated for other gastornithiforms, but the hindlimb bones of Diatryma geiselensis likewise exhibit pronounced differences in size, and differently-sized individuals were found in the same stratigraphic horizon (e.g., GMH XLI-200-1968 and GMH XXXV-481-1963; Figure 7). Rather than assuming the coexistence of two large flightless birds in the same habitat, we consider it more likely that D. geiselensis was sexually dimorphic in size. The culmen of the upper beak is more vaulted in the larger specimen GMH XLI-200-1968 than in the smaller GMH XVIII-1178-1958 (Figure 1G), so that D. geiselensis also appears to have shown sexual dimorphism in beak shape.
 Gastornithiforms are now generally regarded as having been herbivorous birds (Andors, 1992; Mayr, 2009; Angst et al., 2014). In this regard, it is notable that a conspicuous rounded quartz pebble, which was not mentioned by Fischer (1978), is situated on the ventral surface of the synsacrum of GMH XLI-200-1968 (Figure 8). Isolated pebbles are uncommon in the coal seams of the Geisel valley, and based on its shape, size, and location, we interpret the specimen as a gastrolith. Possible Gastornis gastroliths were reported before from southern France, where they were found in association with putative gastornithiform eggshell fragments (Angst and Buffetaut, 2017). Gastroliths often occur in herbivorous birds, where they serve to aid digestion of coarse plant matter, but they are also found in omnivorous or carnivorous birds and fulfill different functions across birds that ingest them (Wings, 2007). The partial pelvis of GMH XLI-200-1968 preserves the first in situ gastrolith, which conforms to an herbivorous diet of gastornithiforms, but by itself does not constitute unequivocal evidence for a particular dietary preference.
Gastornithiforms are now generally regarded as having been herbivorous birds (Andors, 1992; Mayr, 2009; Angst et al., 2014). In this regard, it is notable that a conspicuous rounded quartz pebble, which was not mentioned by Fischer (1978), is situated on the ventral surface of the synsacrum of GMH XLI-200-1968 (Figure 8). Isolated pebbles are uncommon in the coal seams of the Geisel valley, and based on its shape, size, and location, we interpret the specimen as a gastrolith. Possible Gastornis gastroliths were reported before from southern France, where they were found in association with putative gastornithiform eggshell fragments (Angst and Buffetaut, 2017). Gastroliths often occur in herbivorous birds, where they serve to aid digestion of coarse plant matter, but they are also found in omnivorous or carnivorous birds and fulfill different functions across birds that ingest them (Wings, 2007). The partial pelvis of GMH XLI-200-1968 preserves the first in situ gastrolith, which conforms to an herbivorous diet of gastornithiforms, but by itself does not constitute unequivocal evidence for a particular dietary preference.
Gastornithiforms existed over at least 16 million years, from the late Paleocene, some 60 Ma, to the middle Eocene, about 44 Ma. During that period, they were exposed to selective forces, which altered their body plan. In particular, gastornithiforms show notable differences in the shapes of their beak, which are indicative of disparate feeding ecologies. The upper beaks of both Diatryma geiselensis and D. gigantea are proportionally much longer than the beak of Gastornis laurenti (Figure 2D; Bourdon et al., 2016). The upper beak of D. geiselensis is mediolaterally wider than that of D. gigantea, and the mandibular rami are dorsoventrally deeper; in GMH XVIII-1178-1958 the tomia are more strongly curved than in D. gigantea. Disparate feeding ecologies of gastornithiforms are also suggested by differences in the shapes of the quadrate, which has a dorsoventrally wider processus orbitalis in D. gigantea than in G. laurenti (compare Matthew and Granger, 1917: pl. XXII with Bourdon et al., 2016: figure 2).
Gastornithiforms are considered to have been graviportal and probably had rather low walking speeds (Fischer, 1978; Angst et al., 2016). The stouter tarsometatarsus proportions probably indicates that the species Diatryma were heavier than those of Gastornis or that they had different walking or feeding postures.
The coracoid of gastornithiforms is unusually wide and lacks a processus acrocoracoideus. This morphology strongly departs from the typical neoavian condition and is likely to be the result of paedomorphosis, that is, the retention of juvenile character, which is a characteristic trait of the development of the pectoral girdle and wing bones of flightless birds (e.g., Livezey, 1993; Cubo and Arthur, 2000). Progressive paedomorphosis - rather than functional adaptations - may also explain the differences in the morphology of the scapulocoracoid of Gastornis parisiensis and the species of Diatryma.
ACKNOWLEDGMENTS
M. Rabi is thanked for drawing our attention to the new Diatryma geiselensis skull. Access to specimens was enabled by R. Rozzi (MLU) and C. Mehling (AMNH). L. Grimm and J. Bolte are acknowledged for taking some of the photographs of the D. geiselensis material. Finally, we are grateful to S. Moore-Fay for providing photos of the gastornithiform from the Isle of Sheppey. Comments from T. Worthy and two anonymous reviewers improved the manuscript.
REFERENCES
Agnolin, F. 2007. Brontornis burmeisteri Moreno and Mercerat, un Anseriformes (Aves) gigante del Mioceno Medio de Patagonia, Argentina. Revista del museo argentino de ciencias naturales nueva serie, 9:15‒25.
https://doi.org/10.22179/REVMACN.9.361
Agnolin, F.L. 2021. Reappraisal on the phylogenetic relationships of the enigmatic flightless bird (Brontornis burmeisteri) Moreno and Mercerat, 1891. Diversity, 13(2):90.
https://doi.org/10.3390/d13020090
Andors, A.V. 1988. Giant groundbirds of North America (Aves, Diatrymidae). Unpublished Ph.D. dissertation, Columbia University, New York.
Andors, A. 1992. Reappraisal of the Eocene groundbird Diatryma (Aves: Anserimorphae), p. 109‒125. In Campbell, K.E. (ed.), Papers in avian paleontology honoring Pierce Brodkorb. Natural History Museum of Los Angeles County, Science Series, 36.
Angst, D. and Buffetaut, E. 2013. The first mandible of Gastornis Hébert, 1855 (Aves, Gastornithidae) from the Thanetian (Paleocene) of Mont-de-Berru (France). Revue de Paléobiologie, 32:423‒432.
Angst, D., Buffetaut, E., Lécuyer, C., and Amiot, R. 2013. “Terror birds” (Phorusrhacidae) from the Eocene of Europe imply trans-Tethys dispersal. PLoS ONE 8(11):e80357.
https://doi.org/10.1371/journal.pone.0080357
Angst, D., Lécuyer, C., Amiot, R., Buffetaut, E., Fourel, F., Martineau, F., Legendre, S., Abourachid, A., and Herrel, A. 2014. Isotopic and anatomical evidence of an herbivorous diet in the Early Tertiary giant bird Gastornis. Implications for the structure of Paleocene terrestrial ecosystems. Naturwissenschaften 101:313‒322.
https://doi.org/10.1007/s00114-014-1158-2
Angst, D., Buffetaut, E., Lecuyer, C., and Amiot, R. 2016. A new method for estimating locomotion type in large ground birds. Palaeontology, 59:217‒223.
https://doi.org/10.1111/pala.12224
Angst, D. and Buffetaut, E. 2017. Paleobiology of giant flightless birds. ISTE Press, London.
Baumel, J.J. and Witmer, L.M. 1993. Osteologia. In Baumel, J.J., King. A.S., Breazile. J.E., Evans, H.E., Vanden Berge, J.C. (eds.), Handbook of Avian Anatomy: Nomina Anatomica Avium, 2nd edition. Publications of the Nuttall Ornithological Club, 23:45-132.
Berg, D.E. 1965. Nachweis des Riesenlaufvogels Diatryma im Eozän von Messel bei Darmstadt/Hessen. Notizblatt des hessischen Landesamtes für Bodenforschung, 93:68‒72.
Bourdon, E., Mourer-Chauviré, C., and Laurent, Y. 2016. Early Eocene birds from La Borie, southern France. Acta Palaeontologica Polonica, 61:175‒190.
https://doi.org/10.1016/j.geobios.2022.10.004
Brodkorb, P. 1967. Catalogue of fossil birds. Part 3 (Ralliformes, Ichthyornithiformes, Charadriiformes). Bulletin of the Florida State Museum, Biological Sciences, 11:99-220.
https://doi.org/10.58782/flmnh.koax3014
Buffetaut, E. 1997. New remains of the giant bird Gastornis from the Upper Paleocene of the eastern Paris Basin and the relationships between Gastornis and Diatryma. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 1997:179‒190.
https://doi.org/10.1127/njgpm/1997/1997/179
Buffetaut, E. 2004. Footprints of giant birds from the Upper Eocene of the Paris basin: an ichnological enigma. Ichnos, 11:357‒362.
https://doi.org/10.1080/10420940490442287
Buffetaut, E. 2008. First evidence of the giant bird Gastornis from southern Europe: a tibiotarsus from the Lower Eocene of Saint-Papoul (Aude, southern France). Oryctos, 7:75‒82.
Buffetaut, E. 2013. The giant bird Gastornis in Asia: a revision of Zhongyuanus xichuanensis Hou, 1980, from the early Eocene of China. Paleontological Journal, 47:1302‒1307.
https://doi.org/10.1134/S0031030113110051
Buffetaut, E. 2017. A brontornithid from the Deseadan (Oligocene) of Bolivia. Contribuciones del Museo Argentino de Ciencias Naturales, 7:39-47.
Buffetaut, E. and Angst, D. 2014. Stratigraphic distribution of large flightless birds in the Palaeogene of Europe and its palaeobiological and palaeogeographical implications. Earth-Science Reviews, 138:394‒408.
https://doi.org/10.1016/j.earscirev.2014.07.001
Buffetaut, E. and de Ploëg, G. 2020. Giant birds from the uppermost Paleocene of Rivecourt (Oise, northern France). Boletim do Centro Português de Geo-História e Pré-História, 2:29‒33.
Collinson, M.E., Adams, N.F., Manchester, S.R., Stull, G.W., Herrera, F., Smith, S.Y., Andrew, M.J., Kenrick, P., and Sykes, D. 2016. X-ray micro-computed tomography (micro-CT) of pyrite-permineralized fruits and seeds from the London Clay Formation (Ypresian) conserved in silicone oil: a critical evaluation. Botany, 94:697-711.
https://doi.org/10.1139/cjb-2016-0078
Cope, E.D. 1876. On a gigantic bird from the Eocene of New Mexico. Proceedings of the Acadmy of Natural Sciences Philadelphia, 28:10‒11.
Cracraft, J. 1976. The hindlimb elements of the moas (Aves, Dinornithidae): A multivariate assessment of size and shape. Journal of Morphology, 150:495‒526.
https://doi.org/10.1002/jmor.1976.150.2.495
Cubo, J. and Arthur, W. 2000. Patterns of correlated character evolution in flightless birds: a phylogenetic approach. Evolutionary Ecology, 14:693‒702.
https://doi.org/10.1023/A:1011695406277
Dollo, L. 1883. Note sur la présence du Gastornis Edwardsii, Lemoine, dans l’assise inférieure de l’étage Landénien à Mesvin, près Mons. Bulletin du Musée royal d’Histoire naturelle de Belgique, 2:297-305.
Elzanowski, A. and Mayr, G. 2018. Multiple origins of secondary temporal fenestrae and orbitozygomatic junctions in birds. Journal of Zoological Systematics and Evolutionary Research, 56:248‒269.
https://doi.org/10.1111/jzs.12196
Fischer, K. 1962. Der Riesenlaufvogel Diatryma aus der eozänen Braunkohle des Geiseltales. Hallesches Jahrbuch für mitteldeutsche Erdgeschichte, 4:26‒33.
Fischer, K. 1967. Ein neuer großer Laufvogel aus dem Eozän des Geiseltales bei Halle (Saale). Berichte der deutschen Gesellschaft für geologische Wissenschaften, Reihe A, Geologie und Paläontologie, 12:601-605.
Fischer, K. 1978. Neue Reste des Riesenlaufvogels Diatryma aus dem Eozän des Geiseltals bei Halle (DDR). Mitteilungen aus dem Zoologischen Museums in Berlin, 54, Supplement: Annalen für Ornithologie, 2:133‒144.
Fischer, K. 1987. Eulenreste (Eoglaucidium pallas nov. gen., nov. sp., Strigiformes, Aves) aus der mitteleozänen Braunkohle des Geiseltals bei Halle (DDR). Mitteilungen aus dem Zoologischen Museums in Berlin, 63, Supplement: Annalen für Ornithologie, 11:137‒142.
Fürbringer, M. 1888. Untersuchungen zur Morphologie und Systematik der Vögel, zugleich ein Beitrag zur Anatomie der Stütz- und Bewegungsorgane. Van Holkema, Amsterdam.
Gaillard, C. 1937. Un oiseau géant dans les dépots éocènes du Mont d’Or lyonnais. Annales de la Société linnéenne de Lyon (Nouvelle Série), 80:111-126.
Harrison, C.J.O. and Walker, C.A. 1977. Birds of the British Lower Eocene. Tertiary Research Special Paper, 3:1‒52.
https://doi.org/10.1163/9789004611443
Hébert, E. 1855. Note sur le tibia du Gastornis pariensis [sic]. Comptes Rendus de l’Académie des Sciences, 40:579-582.
Hellmund, M. 1996. Letzte Grabungsaktivitäten im südwestlichen Geiseltal bei Halle (Sachsen-Anhalt, Deutschland) in den Jahren 1992 und 1993. Hercynia - Ökologie und Umwelt in Mitteleuropa, 30:163‒176.
Hellmund, M. 2013. Reappraisal of the bone inventory of Gastornis geiselensis (Fischer, 1978) from the Eocene “Geiseltal Fossillagerstätte”(Saxony-Anhalt, Germany). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 269:203‒220.
https://doi.org/10.1127/0077-7749/2013/0345
Hou, L.-H. 1980. [New form of the Gastornithidae from the Lower Eocene of the Xichuan, Honan]. Vertebrata PalAsiatica, 18:111‒115 [in Chinese]
Krumbiegel, G., Rüffle, L., and Haubold, H. 1983. Das eozäne Geiseltal. Die Neue Brehm-Bücherei, 237. A. Ziemsen, Wittenberg.
Lambrecht, K. 1933. Handbuch der Palaeornithologie. Gebrüder Borntraeger, Berlin
Lemoine, V. 1878. Recherches sur les oiseaux fossiles des terrains tertiaires inférieurs des environs de Reims. F. Keller, Reims.
https://doi.org/10.5962/bhl.title.105275
Lemoine, V. 1881. Recherches sur les oiseaux fossiles des terrains tertiaires inférieurs des environs de Reims, deuxième partie. Matot-Braine, Reims.
Linnaeus, C. 1758. Systema naturae per regna tria naturae. 10th edition, 2 vols. L. Salmii, Holmiae.
Livezey, B.C. 1993. An ecomorphological review of the dodo (Raphus cucullatus) and solitaire (Pezophaps solitaria), flightless Columbiformes of the Mascarene Islands. Journal of Zoology, 230:247‒292.
https://doi.org/10.1111/j.1469-7998.1993.tb02686.x
Louchart, A., Bhullar, B.A., Riamon, S., and Field, D.J. 2021. The true identity of putative tooth alveoli in a Cenozoic crown bird, the gastornithid Omorhamphus. Frontiers in Earth Science, 9:661699.
https://doi.org/10.3389/feart.2021.661699
Marsh, O.C. 1894. A gigantic bird from the Eocene of New Jersey. American Journal of Science, Series 3, 48:344.
https://doi.org/10.2475/ajs.s3-48.286.344
Martin, L.D. 1992. The status of the Late Paleocene birds Gastornis and Remiornis, p. 97‒108. In Campbell, K.E. (ed.), Papers in avian paleontology honoring Pierce Brodkorb. Natural History Museum of Los Angeles County, Science Series, 36.
Matthew, W.D. and Granger, W. 1917. The skeleton of Diatryma, a gigantic bird from the Lower Eocene of Wyoming. Bulletin of the American Museum of Natural History, 37:307‒326
Mayr, G. 2002. Avian Remains from the Middle Eocene of the Geiseltal (Sachsen-Anhalt, Germany), p. 77‒96. In Zhou, Z. and Zhang, F. (eds.), Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution, Beijing, 1-4 June 2000. Science Press, Beijing.
Mayr, G. 2007. The birds from the Paleocene fissure filling of Walbeck (Germany). Journal of Vertebrate Paleontology, 27:394‒408.
https://doi.org/10.1671/0272-4634(2007)27[394:TBFTPF]2.0.CO;2
Mayr, G. 2009. Paleogene Fossil Birds. Springer, Heidelberg.
https://doi.org/10.1007/978-3-540-89628-9
Mayr, G. 2020. An updated review of the middle Eocene avifauna from the Geiseltal (Germany), with comments on the unusual taphonomy of some bird remains. Geobios, 62:45‒59.
https://doi.org/10.1016/j.geobios.2020.06.011
Mayr, G. 2022. Paleogene Fossil Birds, 2nd edition. Springer, Cham, Switzerland.
https://doi.org/10.1007/978-3-030-87645-6
Mayr, G. and Smith, T. 2019. New Paleocene bird fossils from the North Sea Basin in Belgium and France. New Paleocene bird fossils from the North Sea Basin in Belgium and France. Geologica Belgica, 22:35‒46.
https://doi.org/10.20341/gb.2019.003
Milne-Edwards, A. 1867. Etude sur les rapports zoologiques du Gastornis parisiensis. Annales des sciences naturelles, Zoologie et Paléontologie, 7:217‒227.
Mlíkovský, J. 2002. Cenozoic Birds of the World. Part 1: Europe. Ninox Press, Praha
Mourer-Chauviré, C. and Bourdon, E. 2016. The Gastornis (Aves, Gastornithidae) from the Late Paleocene of Louvois (Marne, France). Swiss Journal of Palaeontology, 135:327‒341.
https://doi.org/10.1007/s13358-015-0097-7
Mourer-Chauviré, C. and Bourdon, E. 2020. Description of a new species of Gastornis (Aves, Gastornithiformes) from the early Eocene of La Borie, southwestern France. Geobios, 63:39‒46.
https://doi.org/10.1016/j.geobios.2020.10.002
Mourer-Chauviré, C., Bourdon, E., Duffaud, S., Le Roux, G., and Laurent, Y. 2024. New avian remains from the early Eocene of La Borie, southern France. Geobios, 83:61‒84.
https://doi.org/10.1016/j.geobios.2022.10.004
Mustoe, G.E., Tucker, D.S., and Kemplin, K.L. 2012. Giant Eocene bird footprints from northwest Washington, USA. Palaeontology, 55:1293‒1305.
https://doi.org/10.1111/j.1475-4983.2012.01195.x
Newton, E.T. 1885. Gastornis klaasseni, Newton, a gigantic bird from the Lower Eocene of Croydon. Geological Magazine, (3) 2:362‒364.
Newton, E.T. 1886. VI. On the remains of a gigantic species of bird (Gastornis klaasseni, n. sp.) from the Lower Eocene beds near Croydon. Transactions of the Zoological Society of London, 12:143-160.
https://doi.org/10.1111/j.1096-3642.1886.tb00009.x
Patterson, J. and Lockley, M.G. 2004. A probable Diatryma track from the Eocene of Washington: an intriguing case of controversy and skepticism. Ichnos, 11:341‒347.
https://doi.org/10.1080/10420940490442278
Peters, D.S. 1988. Die Messel-Vögel - eine Landvogelfauna, p. 135-151. In Schaal, S. and Ziegler, W. (eds.), Messel - ein Schaufenster in die Geschichte der Erde und des Lebens. Kramer, Frankfurt.
Schaub, S. 1929. Über eocäne Ratitenreste in der osteologischen Sammlung des Basler Museums. Verhandlungen der Naturforschenden Gesellschaft zu Basel, 40:588-598.
Shufeldt, R.W. 1913. Further studies of fossil birds with descriptions of new and extinct species. Bulletin of the American Museum of Natural History, 32:285-306.
Stejneger, L. 1885. Gastornithes, p. 54-55. In Kingsley, J.S. (ed.), The Standard Natural History, vol. 4, Birds. S.E. Cassino, Boston, MA.
Stidham, T.A. and Eberle, J.J. 2016. The palaeobiology of high latitude birds from the early Eocene greenhouse of Ellesmere Island, Arctic Canada. Scientific Reports, 6:20912.
https://doi.org/10.1038/srep20912
Vandenberghe, N., Hilgen, F.J., and Speijer, R.P., with contributions by Ogg, J.G., Gradstein, F.M., Hammer, O., Hollis, C.J., and Hooker J.J. 2012. The Paleogene period, p. 855-921. In Gradstein, F.M., Ogg, J.G., Schmitz, M., and Ogg, G. (eds.), The Geologic Time Scale. Elsevier, Amsterdam.
https://doi.org/10.1016/B978-0-444-59425-9.00028-7
Weigelt, J. 1939. Die Aufdeckung der bisher ältesten tertiären Säugetierfauna Deutschlands. Nova Acta Leopoldina, 7:515‒528.
West, R.M. and Dawson, M.R. 1978. Vertebrate paleontology and the Cenozoic history of the North Atlantic region. Polarforschung, 48:103‒119.
Wings, O. 2007. A review of gastrolith function with implications for fossil vertebrates and a revised classification. Acta Palaeontologica Polonica, 52:1‒16.
Worthy, T.H., Degrange, F.J., Handley, W.D., and Lee, M.S. 2017. The evolution of giant flightless birds and novel phylogenetic relationships for extinct fowl (Aves, Galloanseres). Royal Society Open Science, 4(10):170975.
https://doi.org/10.1098/rsos.170975
