

 Ammonoids and their biozonation across the Santonian-Campanian boundary in north-eastern Coahuila, Mexico
Ammonoids and their biozonation across the Santonian-Campanian boundary in north-eastern Coahuila, Mexico
Article number: 24.3.a34
https://doi.org/10.26879/1046
Copyright Palaeontological Association, October 2021
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 19 November 2019. Acceptance: 7 September 2021.
ABSTRACT
Fossil-rich shelf carbonates of Santonian-Campanian age are superbly exposed at Tepeyac near Jiménez, northern Coahuila, north-eastern Mexico. In this paper we describe 24 species of ammonoids assigned to 12 genera from this section. The 330 specimens were mostly recorded in situ and allow for a detailed biozonation of the section. We propose the first formal biozonation across the Santonian-Campanian boundary in the Gulf Coast region. The section yields abundant Parapuzosia assigned to P. (P.) seppenradensis and may thus correspond to one of the historical sites from which Böse and Cavins documented this giant ammonite ca. 90 years ago. The faunal assemblage provides evidence for repeated and increasing faunal exchange with Europe and southern latitudes, but also includes intervals of faunal cut-off and endemic evolution. The abundance of fossils, the excellent outcrop conditions, and biozonal completeness favor the use of the Tepeyac section as a potential reference section for the Santonian-Campanian boundary in the Americas.
Christina Ifrim. Institut für Geowissenschaften, Ruprecht-Karls-Universität, Im Neuenheimer Feld 234, 69120 Heidelberg, Germany
current address: SNSB Jura-Museum, Burgstr. 19, 85072 Eichstätt, Germany, ifrim@snsb.de
Wolfgang Stinnesbeck. Institut für Geowissenschaften, Ruprecht-Karls-Universität, Im Neuenheimer Feld 234, 69120 Heidelberg, Germany. wolfgang.stinnesbeck@geow.uni-heidelberg.de
Keywords: Upper Cretaceous; Santonian-Campanian boundary; ammonoids; biozonation; Gulf of Mexico
Final citation: Ifrim, Christina and Stinnesbeck, Wolfgang. 2021. Ammonoids and their biozonation across the Santonian-Campanian boundary in north-eastern Coahuila, Mexico. Palaeontologia Electronica, 24(3):a34. https://doi.org/10.26879/1046
palaeo-electronica.org/content/2021/3462-campanian-ammonoid-biozonation
Copyright: October 2021 Palaeontological Association.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
To date, the only description of a Mexican locality with Parapuzosia is by Böse (1927). According to these authors, the horizon providing evidence for this giant Upper Cretaceous (Santonian-lower Campanian) ammonite “is very well developed in the Rio de San Rodrigo near Piedras Negras, and still better in the Arroyo (= creek) del Tecolote and Arroyo Blanco near Jimenez, Coahuila, where it contains an enormous number of fossils, especially ammonites. Some of the Pachydiscus there measure over 1.5 meters in diameter and ammonites of a diameter of between one-half meter and 1 meter are quite common.” Böse and Cavins (1927) further stated that the “lower Santonian consists of a white to gray chalky limestone with beds of hard gray limestone and light gray marls. A suitable place to measure the thickness has not been found, but it is probably not over one hundred meters and possibly even less. 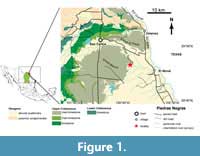 The horizon is very fossiliferous; the most common fossils are: Mortoniceras texanum Roemer and related members of the group, Pachydiscus flaccidicosta Roemer, Baculites sp.,
The horizon is very fossiliferous; the most common fossils are: Mortoniceras texanum Roemer and related members of the group, Pachydiscus flaccidicosta Roemer, Baculites sp.,  large undescribed Inoceramus related to I. regularis d'Orbigny; locally it contains great numbers of echinoderms, small Exogyra, Gryphaea sp., etc. The frequency of Mortoniceras texanum shows that the horizon represents the lower Santonian. We have never found a single Placenticeras in these beds although several species of this genus have been cited from Texas”.
large undescribed Inoceramus related to I. regularis d'Orbigny; locally it contains great numbers of echinoderms, small Exogyra, Gryphaea sp., etc. The frequency of Mortoniceras texanum shows that the horizon represents the lower Santonian. We have never found a single Placenticeras in these beds although several species of this genus have been cited from Texas”.
In this paper we describe ammonoid specimens from one of the historical sites originally documented by Böse (1927), Renz (1936), and Böse and Cavins (1927) as Arroyo Blanco. This site was relocated by us near the village of Tepeyac, about 40 km north of the Piedras Negras and 15 km SSW of Jiménez, Coahuila, in the bed of the intermittent Arroyo Blanco river, a tributary to the Rio Grande, and close to the mouth of the Arroyo Tecolote (Figure 1). We logged a section of 28.9 m of rhythmically bedded bright grey to light yellow-coloured layers of chalk and limestone, and intercalated units of marl and marly clay (Figure 2). Both litho- and biofacies indicate that these sediments were deposited on the shelf of the ancient Gulf of Mexico.The ammonite assemblage documented here consists of about 220 specimens mostly found in situ, i.e., in their bed of origin. Additionally, ca. 100 determinable moulds were documented in the field. This study thus comprises ca. 320 stratified specimens, providing a solid base for biozonation across the Santonian-Campanian boundary (Figure 2).
SYSTEMATIC PALEONOLOGY
Higher Systematics, Taxonomy, and Nomenclature
The systematics of ammonites majorly follows the Treatise of Invertebrate Paleontology (Wright et al., 1996). Exceptions refer to Texanitinae, for which the generic definition of Klinger and Kennedy (1980) was used, to Puzosiinae, which are here used in the sense of Kennedy (2019), and to Amanpondella Klinger and Kennedy, 1997. Synonymy lists are abbreviated to the most important citations and references used for determination. More complete synonymies are indicated where available.
Ornament is described in the sense of Ifrim et al. (2018).
Abbreviations
Dimensions. All dimensions are given in mm; dimensions in brackets are % of diameter. D: diameter; WH: whorl height; U: umbilical diameter. L: undeformed length in heteromorphs taken dorsally. Asterisks mark estimated values or those from deformed or weathered individuals.
Collections. Specimens described here are inventorized in the CPC: Colección de Paleontología de Coahuila, housed at the Museo del Desierto, Saltillo, Coahuila, Mexico and in the IGM: Instituto de Geología de México of the Universidad Autónoma de Mexicoc (UNAM), housed in the Museo María del Carmen Perrilliat M., Colección Nacional de Paleontología, UNAM, Mexico City.. Other collections mentioned are: BEG: collections of the Bureau of Economic Geology, University of Texas, Austin, Texas; GPIB: Geologisch-Paläontologisches Institut der Universität Bonn, Germany; NMBE: Naturhistorisches Museum Bern, Switzerland; SAM: South African Museum, Cape Town, South Africa; TMM: Texas Memorial Museum, Austin, Texas, USA; USNM: United States Museum of Natural History, Washington, D.C., USA; UT: Department of Geology, University of Texas, Austin, Texas; WSA: W. S. Adkins collection of the University of Texas at Austin,Texas.
Phylum MOLLUSCA
Class CEPHALOPODA Cuvier, 1797
Order AMMONOIDEA Zittel, 1884
Suborder AMMONITINA Hyatt, 1889
Superfamily ACANTHOCERATOIDEA de Grossouvre, 1894
Family COLLIGNONICERATIDAE Wright and Wright, 1951
Subfamily TEXANITINAE Collignon, 1948
This subfamily is discussed in detail by Klinger and Kennedy (1980). We follow these authors and consider Texanites and Plesiotexanites as separate genera instead of subgenera.
Genus PLESIOTEXANITES Matsumoto, 1970
The genus is discussed in detail by Klinger and Kennedy (1980).
Plesiotexanites shiloensis Young, 1963
Figure 3A-G, Table 1
1952 Texanites internodosus Young and Marks, p. 477 ff [ non plate 1, figure 4 = Texanites internodosus Renz]
 1963 Texanites shiloensis n. sp.; Young, p. 89, plate 46, figures 1-4; plate 54, figures 4-7; plate 70, figures 5, 6, 8; text-figure 24d
1963 Texanites shiloensis n. sp.; Young, p. 89, plate 46, figures 1-4; plate 54, figures 4-7; plate 70, figures 5, 6, 8; text-figure 24d
1970 Texanites sp. cf. Texanites (Plesiotexanites) shiloensis Young; Matsumoto, p. 296, plate 47[51], figure 1; text-figure 26 [100]
1980 Plesiotexanites shiloensis (Young); Klinger and Kennedy, p. 65
1991 Texanites (Plesiotexanites) shiloensis Young, 1963; Kennedy and Cobban, p. 179, figure 8.30
1997 Texanites (Plesiotexanites) shiloensis Young, 1963; Kennedy and Cobban, p. 18, figure 13
Type. The holotype is TMM 1986 by original designation by Young (1963, p. 89, plate 46, figures 1-4, plate 70, figure 5; text-figure 24d). It is from the Dessau Chalk on Brushy Creek, 23.2 km south of Hutto, Williamson County, Texas.
Material. CPC-2363, -2371, -2376, -2378, -2384, -2385, -2543, IGM 12795, and uncollected specimens in beds TPY 23 to TPY 46.
Description. Evolute, with U ranging from 0.31 to 0.44. Whorl section moderately compressed, with WB/WH of between 0.55 and 0.75, increasing with D. The umbilical wall is undercut and bends in a narrow arch into flattened flanks. Rib index is 27 to 32 at D > 200 mm. Ribs strengthen across the umbilical wall into sharp, closely spaced umbilical bullae that project into the umbilicus. Blunt, straight, slightly prorsiradiate ribs arise from these bullae. A row of weak, rounded tubercles is located at midflank, while a submarginal row of weaker tubercles is close to a marginal row of somewhat stronger clavate tubercles, each of which is linked by a low, broad rib to a strong, external clavus. A broad, smooth zone separates the external clavi from a strong siphonal keel.
Dimensions. Table 1.
Remarks. The species was included into Plesiotexanites by Matsumoto (1970), a view accepted by Kennedy et al. (1997a) and here, based on tuberculation, i.e., the presence of umbilical, lateral, submarginal, marginal, and external tubercles. The compressed, dense, and finely ribbed juvenile stage of the species (e.g., Figure 3B) distinguish P. shiloensis from all other North American Plesiotexanites. Texanites s.s. is distinguished by tuberculation and details in ribbing.
Occurrence. Upper Santonian to lower Campanian of the Jiménez area of Coahuila, Mexico, the central Texas and Big Bend areas of Texas as well as the state of Mississippi in the US, and Hokkaido, Japan.
Genus TEXANITES Spath, 1932
Texanites lonsdalei Young, 1963
Figure 4A-G, Table 2
1963 Texanites lonsdalei n. sp.; Young, 1963, p. 90, plate 34, figure 1; plate 51, figures 3-7; plate 58, figures 5, 6; text-figures 22a, d
 1967 Texanites lonsdalei Young, 1963; Wolleben, p. 1152, p1. 147, figures 4, 5; text-figures 8c,d
1967 Texanites lonsdalei Young, 1963; Wolleben, p. 1152, p1. 147, figures 4, 5; text-figures 8c,d
1997 Texanites (Texanites) lonsdalei Young, 1963; Kennedy and Cobban, p. 17, figures 9-12
Type. The holotype is TMM-UT 30474 by original designation by Young (1963: 90, plate 34, figure 1; plate 51, figures 3-7; plate 58, figures 5, 6; text-figures 22a, d). It is from the Dessau Chalk of Kitchens Ranch, Travis County, Texas.
Material. CPC-2375, -2377, -2584, IGM 12796, and 12797, and uncollected specimens from beds TPY 21 to TPY 53.
Description. Evolute shell with U/D of about 0.4. 28 to 33 sharp ribs rise on the umbilical wall and strengthen into pointed bullae that project into the umbilicus. These bullae give rise to sharp, straight, moderately prorsiradiate ribs, which are occasionally intercalated by secondary ribs. A row of strong, lateral tubercles is located dorsal of midflank and separated from the umbilical row of bullae. A third slightly weaker row of tubercles is present ventrolaterally; they are rounded to feebly clavate. A fourth row of larger tubercles, respectively coarse clavi, is seen on the venter, next to a siphonal keel from which they are separated by a smooth zone. The secondary ribs also show these outer rows of tubercles, resulting in 36 ventral clavi per whorl. The keel and lateral clavi disappear at largest diameters, and ribs in these large-sized specimens cross the venter in a moderately narrow arch.
Dimensions. Table 2.
Remarks. Klinger and Kennedy (1980) casted doubt on the validity of Texanites lonsdalei, but subsequently, Kennedy et al. (1997a) discussed the species in detail and presented characters for distinction from similar taxa. According to these latter authors, T. lonsdalei is distinguished by sharper, straighter ribs than coeval species of Plesiotexanites or Menabites (Delawarella).
Occurrence. Uppermost Santonian and lowermost Campanian Menabites (Delawarella) tequesquitense Zone of Texas. At Tepeyac, the species occurs in the uppermost Plesiotexanites shiloensis zone of the Santonian and in the top of the lower Campanian M. (D.) delawarensis Zone.
Genus BEVAHITES Collignon, 1939
The genus is discussed in detail by Klinger and Kennedy (1980).
Bevahites bevahensis Collignon, 1948
Figure 5A-B, G-J, Table 3
1928 Mortoniceras sp.; Adkins, plate 34, figure 1
 1948 Behavites [sic] bevahensis nov. sp.; Collignon, p. 84, plate 11 (5), figures 3, 3a, 3b
1948 Behavites [sic] bevahensis nov. sp.; Collignon, p. 84, plate 11 (5), figures 3, 3a, 3b
p. 1952 Texanites densinodosus; Young and Marks, p. 480, 482-483 [not plate 1, figure 1]
1963 Bevahites bevahensis Collignon, 1948; Young, p. 94, plate 53, figures 1-7; text-figures 15c, 21d, 27b
1967 Bevahites bevahensis Collignon, 1948; Wolleben, p. 1153, plate 148, figure 4; text-figures 8a, b
Type. The holotype is the original of Collignon (plate 11, figures 3, 3a, b) by monotypy. It is from the upper Santonian of Bevaho, Madagascar, and stored in the Collections de l'École Nationale Supérieure des Mines, Paris, currently housed in the Villeurbanne campus, Université Lyon.
Material. Two specimens, CPC-2374 and -2547, from bed TPY26.
Description. Moderately involute, with U/D between 0.30 and 0.39. Whorl section suboval; WB/WH = 0.56, with WB largest at mid-flank. Ribs moderately sinuous. One or two ribs arise from prominent, strong umbilical tubercles, although ribbing is moderately inequidistant. Ribs are moderately prorsiradiate and flex back in a wide arch to become straight and rursiradiate ventrolaterally. Bullae are located on the ribs at mid-flank, while broad, faint ventrolateral tubercles are connected to weak clavi next to the venter.
Dimensions. Table 3.
Remarks. Ribbing and whorl section readily distinguish Bevahites bevahensis from other Texanitinae. The adult stage of this species is discussed in detail by Wolleben (1967).
Occurrence. Uppermost Santonian of westernmost to central Texas, Chihuahua, Tepeyac in Coahuila, Madagascar.
Bevahites costatus Collignon, 1948 coahuilaensis Young, 1963
Figure 5D-F, Table 4
1963 Bevahites costatus Collignon, 1948, subsp. coahuilaensis, n. subsp.; Young, p. 96, plate 47, figures 1-4, plate 71, figure 5; text-figures 34b
1967 Bevahites costatus coahuilaensis Young, 1963; Wolleben, p. 1156, plate 150, figure 8
Type. The holotype is BEG-20288 through original designation by Young (1963, p. 96, plate 47, figures 1-4, plate 71, figure 5, text-figure 34b), from Arroyo Tecolote near Jiménez, Coahuila.
Material. Three specimens, CPC-2372, -2546, and -2548 from beds TPY27 to TPY44.
Description. Moderately involute, with subrectangular whorl section and a maximum WB at mid-flank. Blunt ribs arise from umbilical tubercles and reach to mid-flank where they bifurcate at a rounded tubercle. From here, secondaries reach to the venter and wear two rows of shallow, ventrolateral tubercles, as well as a third ventrolateral row of clavi. The venter is keeled, with a smooth area between the ventrolateral clavi and the keel.
Dimensions. Table 4.
Remarks. Young’s (1963) type specimens are from the Arroyo Tecolote, located at about 5 km north of Tepeyac. Wolleben (1967) noted that this subspecies of Bevahites costatus is rare in the area, but that it occurs in several stratigraphic levels. It differs from Bevahites behavensis in a stout whorl section and ribs bifurcating at D of about 130 mm.
Occurrence. Bevahites costatus coahuilaensis occurs at about the Santonian-Campanian boundary in the Ojinaga Formation of eastern Chihuahua and in the Jiménez area of Coahuila, Mexico, as well as in western Texas. At Tepeyac, it was identified in beds 27, 36, and 44-46, i.e., in the interval across the Santonian-Campanian boundary. The species was also reported from the lower Campanian of Madagascar (Collignon, 1969).
Genus MENABITES Collignon, 1948
Subgenus DELAWARELLA Collignon, 1948
Many species described below were earlier included into Submortoniceras based on quinque-tuberculation seen in lower Campanian Texanitinae from Texas, at least during part of their ontogeny, while other species were based on juveniles. However, Submortoniceras is based on the South African Mortoniceras woodsi Spath, 1921. This taxon defined by Spath (1926) was interpreted to have evolved from Texanites soutoni soutoni in South Africa at about the Santonian-Campanian boundary, there producing a large number of endemic species (Klinger and Kennedy, 1980). Menabites (Delawarella) tequesquitense (Morton, 1830), on the other hand, was interpreted to have evolved during the late Santonian from Plesioexanites shiloensis (Young, 1963), or alternatively, from the South African soutoni lineage (Klinger and Kennedy, 1980); it cannot be assigned to the younger South African genus Submortoniceras. In the North American Menabites (Delawarelly) group two lineages are distinguished: M. (D.) tequesquitense gave rise to the tequesquitense - delawarensis - danei lineage. The proximity between early Campanian Texanitinae from Texas and South Africa suggests a close relationship. The other lineage departs from M. (D.) vanuxemi giving rise to M. (D.) mariscalense and M. (D.) uddeni (Young, 1963) and some more compressed species of this genus endemic to Texas. They are here all included into Menabites (Delawarella).
Menabites (Delawarella) tequesquitense (Young, 1963)
Figure 6A-B, Figure 7A-C, Figure 8, Figure 9, Figure 10A-C, Table 5
 1963 Submortoniceras tequesquitense, n. sp.; Young, p. 97, plate 28, figure 1; plate 42, figures 1, 2; plate 44, figures 4, 5; plate 51, figures 1, 2; plate 52, figures 1-4; plate 57, figure 4; plate 70, figure 1; text-figures 12b, 28b
1963 Submortoniceras tequesquitense, n. sp.; Young, p. 97, plate 28, figure 1; plate 42, figures 1, 2; plate 44, figures 4, 5; plate 51, figures 1, 2; plate 52, figures 1-4; plate 57, figure 4; plate 70, figure 1; text-figures 12b, 28b
1991 Submortoniceras tequesquitense Young, 1963; Cobban and Kennedy, p. B3, plate 1, figures 1, 2
1997 Submortoniceras tequesquitense Young, 1963; Kennedy et al., p.23, figures 14, 15
 Type. The holotype is TMM 34742 by original designation of Young (1963, p. 97, plate 42, figures 1-2; plate 52, figures 1-2, 4, text-figure 28b). It is from the upper part of the Dessau Chalk, "downstream from the concrete spillway of the new (1932) Del Rio-Eagle Pass highway crossing on Tequesquite Creek, Kinney County, Texas" (Young, 1963, p. 97).
Type. The holotype is TMM 34742 by original designation of Young (1963, p. 97, plate 42, figures 1-2; plate 52, figures 1-2, 4, text-figure 28b). It is from the upper part of the Dessau Chalk, "downstream from the concrete spillway of the new (1932) Del Rio-Eagle Pass highway crossing on Tequesquite Creek, Kinney County, Texas" (Young, 1963, p. 97).
 Material. Eight specimens, CPC-2381,-2383, -2387, -2422, -2550,-2551, -2564,-2567, in addition to uncollected specimens, from beds TPY 25 to TPY 50.
Material. Eight specimens, CPC-2381,-2383, -2387, -2422, -2550,-2551, -2564,-2567, in addition to uncollected specimens, from beds TPY 25 to TPY 50.
Description. Evolute shell with a moderately high expansion rate and a relatively wide U (U/D of 0.29-0.41). The umbilical wall seems to be undercut. 24 sharp umbilical bullae are present per whorl and project into the umbilicus. Sharp single or paired ribs arise from these bullae. They are broad and low, straight to slightly prorsiradiate, and efface markedly on the flanks. A row of sharp lateral tubercles is situated near mid-flank and separated by a row of weaker, ventrolateral tubercles. Each of these tubercles is connected by a low and broad rib to stronger elongate clavi; the latter are separated from the strong siphonal keel by a smooth sulcate-shaped zone. The external clavi thus flank the bisulcate-carinate venter. In adult specimens the clavi and keel are replaced by weaker, broad rounded tubercles, and the ribs cross a moderately wide rounded venter.
Dimensions. Table 5.
 Remarks. The well-developed ribs on the inner whorls readily distinguish this species from all other Gulf Coast representatives of the genus described by Young (1963) as Submortoniceras, including S. sancarlosense, S. mariscalense, S. uddeni, and S. vandaliaense.
Remarks. The well-developed ribs on the inner whorls readily distinguish this species from all other Gulf Coast representatives of the genus described by Young (1963) as Submortoniceras, including S. sancarlosense, S. mariscalense, S. uddeni, and S. vandaliaense.
Klinger and Kennedy (1980) suggested Menabites (Delawarella) tequesquitense to be closely allied if not conspecific with Submortoniceras woodsi, because dimensions of the two species overlap. Apart from a slightly narrower whorl section, denser and more forwardly curved costation, there seems to be little difference between M. (D.) tequesquitense and S. woodsi.  However, this difference is here considered to be sufficient for specific differentiation. Young (1963, p. 98) suggested that M. (D.) tequesquitense could be derived from Plesiotexanites shiloensis. However, affinities of Menabites (Delawarella) tequesquitense with the South African T. soutoni seem closer than with P. shiloensis. In this view M. (D.) tequesquitense would also be derived from T. soutoni and then give rise to the North American succession of species. It nevertheless appears that both lineages are closely connected and that a repeated interchange of these species occurred between the South African - Madagascar Southern Ocean and the Ancient Gulf of Mexico.
However, this difference is here considered to be sufficient for specific differentiation. Young (1963, p. 98) suggested that M. (D.) tequesquitense could be derived from Plesiotexanites shiloensis. However, affinities of Menabites (Delawarella) tequesquitense with the South African T. soutoni seem closer than with P. shiloensis. In this view M. (D.) tequesquitense would also be derived from T. soutoni and then give rise to the North American succession of species. It nevertheless appears that both lineages are closely connected and that a repeated interchange of these species occurred between the South African - Madagascar Southern Ocean and the Ancient Gulf of Mexico.
Occurrence. Menabites (Delawarella) tequesquitense is the index fossil of the identical uppermost Santonian-lowermost Campanian ammonite zone, introduced for the Gulf Coast region by Young (1963). The zone has been recorded in Mississippi, Alabama, and Texas (Young, 1963; Kennedy et al., 1997a). A single specimen documented by Kennedy et al. (1997a) from New Mexico, has been collected from the Scaphites (Scaphites) hippocrepis I zone.
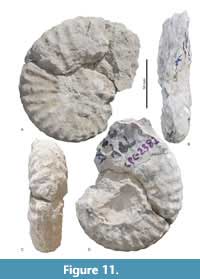
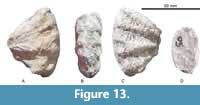
Menabites (Delawarella) delawarensis (Morton, 1830)
Figure 11, Figure 12, Figure 13, Table 6
1830 Ammonites delawarensis Morton, p. 244, plate 2, figure 4.
 1948 Menabites (Delawarella) delawarensis Morton; Collignon, p. 29 (86) (with synonymy)
1948 Menabites (Delawarella) delawarensis Morton; Collignon, p. 29 (86) (with synonymy)
1986 Menabites (Delawarella) delawarensis (Morton, 1830); Kennedy, p. 81, plate 3, figures 3, 4 (with synonymy)
1992a Menabites (Delawarella) delawarensis (Morton, 1830); Cobban and Kennedy, p. 73, plate 6, figure 9
1992b Menabites (Delawarella) delawarensis (Morton, 1830); Cobban and Kennedy, p. 445, figures 4.6-4.9, 5.1, 5.2, 5.6, 5.7, 5.11-5.13
1993a Menabites (Delawarella) delawarensis (Morton, 1830); Kennedy and Cobban, p. 836, figures 5.10, 5.11, 9.5-9.13, 10.3, 11.1, 11.2, 11.4- 11.6, 11.8, 11.9, 11.11
 1996 Menabites (Delawarella) delawarensis (Morton); Wiese, plate 1, figure 1a-b
1996 Menabites (Delawarella) delawarensis (Morton); Wiese, plate 1, figure 1a-b
1997 Menabites (Delawarella) delawarensis (Morton, 1830); Kennedy et al., p. 23, figures 16, 17, 18d-e
Types. The types are lost (see Reeside, 1962 p. 132, and Kennedy, 1986 p. 81).
Material. Fourteen specimens, CPC-2382,-2386, -2388, -2389, -2550, -2551, -2552, -2564, -2568,-2569, -2573,-2584, -2585, and -2586 from beds TPY33 to above TPY 60, in addition to uncollected specimens.
Description. Evolute coiling, with U/D between 0.32 and 0.51 and a depressed whorl section. Greatest WB occurs below mid-flank intercostally and at the lateral tubercles costally. The umbilicus is deep, with a flat, vertical wall and narrowly rounded umbilical shoulder. The inner flanks are flattened and subparallel, the outer flanks broadly rounded and convex, and the venter is broadly arched. Sharp umbilical bullae on the umbilical shoulder give rise to 27 to 36 broad, slightly rursiradiate single or paired ribs, their number increasing with diameter. These bullae give rise to strong, straight prorsiradiate ribs. Each submarginal tubercle splits into a marginal clavus and conical submarginal tubercle, while a lateral tubercle is situated near mid-flank. A broad, smooth sulcate zone separates the external clavi from the siphonal keel, which is only preserved in smaller specimens.
 Dimensions. Table 6.
Dimensions. Table 6.
Discussion. Menabites (Delawarella) delawarensis is similar to M. (D.) tequesquitense from which it differs by coarser ribbing. M. (D.) danei Young, 1963 is characterized by even coarser ribbing, stronger umbilical, submarginal, and external tubercles only. M. (D.) vanuxemi (Morton, 1834), also described below, is more compressed and involute, and tubercles are subordinate to ribs.
Our specimens present a slightly lower WB/WH ratio than specimens published by other authors. This may partially be due to diagenetic compression but majorly reflects wide intraspecific variation, as known for Schloenbachia varians (Wilmsen and Mosavinia, 2011) and Pseudoschloenbachia mexicana (see Ifrim et al., 2013).
Occurrence. Middle lower Campanian Menabites (Delawarella) delawarensis zone in Delaware and New Jersey, Texas, and Oklahoma (Young, 1963; Cobban and Kennedy, 1992b; Kennedy and Cobban, 1993a; Kennedy et al., 1997a; Kennedy et al., 1997b). Isolated specimens are also reported from the lower Campanian of Aquitaine, France, from Spain, the lower Campanian of Zululand (South Africa) and, possibly, Madagascar (Besaire, 1930; Klinger and Kennedy, 1980; Kennedy, 1986), but not in western Coahuila (Ifrim et al., 2013).
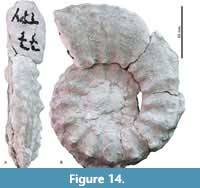
Menabites (Delawarella) danei (Young, 1963)
Figure 14A-B
1929 Mortoniceras delawarensis (Morton), Dane, 1929, pI. 10, figures 1, 2
 1963 Delawarella danei, n. sp.; Young, 1963, p. 114, plate 57, figure 6, plate 62, figures 1, 2; p1. 64, figures 1, 5; p1. 65, figures 1, 2; plate 66, figures 3, 4; text-figures 24e, 33b
1963 Delawarella danei, n. sp.; Young, 1963, p. 114, plate 57, figure 6, plate 62, figures 1, 2; p1. 64, figures 1, 5; p1. 65, figures 1, 2; plate 66, figures 3, 4; text-figures 24e, 33b
1992a Menabites (Delawarella) danei (Young, 1963); Cobban and Kennedy, p. 74, plate 2, figures 5-8; plate 3, figures 1-3; plate 4, figures 1, 12
1992b Menabites (Delawarella) danei (Young, 1963); Cobban and Kennedy, p. 448, figures 3.2-3.4, 5.5, 5.10
1997 Menabites (Delawarella) danei (Young, 1963); Kenned y et al., p. 29, figures 19, 20
Type. The holotype is UT-30646 by original designation by Young (1963, p. 114, plate 57, figure 6; plate 62, figures 1, 2; plate 65, figure 1; plate 66, figure 4; text-figure 33b). It is from “a formation equivalent in age to the Gober chalk,” in McCurtain County, Oklahoma, 1 mile west of the Oklahoma-Arkansas state boundary on Route 3 from Foreman, Arkansas, to Tom, Oklahoma.
Material. Three specimens, CPC-2409 and -2421 from beds TPY 75 and TPY 77, CPC-2590 from bed TPY83, in addition to an uncollected specimen from bed 50.
Description. Evolute coiling, with an U/D ratio of 0.43. The WB/WH ratio is 0.67, with greatest breadth at about mid-flank. Seventeen strong umbilical bullae per whorl give rise to coarse, blunt, prorsiradiate ribs, which efface at mid-flank and again strengthen on the outer flank, where they develop coarse, submarginal tubercles. Broad, coarse ribs link to a row of clavi, with about the same number of additional clavi intercalated. A deep ventral groove separates the row of clavi from the siphonal keel. Faint tubercles at mid-rib are present on the last half whorl, and a second row of ventrolateral tubercles is seen on the last three ribs.
Dimensions. CPC-2421, D: 117, WB: 29 (0.25), WH: 43 (0.37), WB/WH: 0.67, U: 50 (0.43). CPC-2590, WB: 16, WH: 24, WB/WH: 0.67.
Remarks. Specimens of this species show a prolonged Australiella stage and a transition to a typically penta-tuberculate Delawarella ornamentation at large diameters. Menabites (Delawarella) danei is distinguished from all other American Delawarella by the unique coarseness of tuberculation and ribbing. The species is closely related to M. (D.) sabinalensis (Young, 1963), which has no tubercles and intercalated secondaries, instead of the bifurcating ribs and strong tuberculation of M. (D.) danei.
Traces of septate spines as described by Kennedy et al. (1997a) cannot be identified in the Tepeyac specimens, although the tubercles seem to be extended.
Occurrence. Upper lower Campanian of Oklahoma, Texas, Colorado (zone of Baculites obtusus), and now north-eastern Mexico.
Menabites (Delawarella) sp.
Figure 15A-C
Material. Two specimens, CPC-2588 from bed TPY77 and -2590 from bed TPY80.
 Dimensions. CPC-2588, D: 140, WB: 40 (0.29), WH: 62 (0.44), WB/WH: 0.65, U: 43 (0.31), ribs per whorl: 34.
Dimensions. CPC-2588, D: 140, WB: 40 (0.29), WH: 62 (0.44), WB/WH: 0.65, U: 43 (0.31), ribs per whorl: 34.
Remarks. This morphotype has no tubercles and strong, simple ribs, contrasting with all other species in this genus. The two specimens may or may not belong to the same species. CPC-2590 has has a polygonal whorl section comparable to M. (D.) danei but has more convergent flanks, and ribbing is finer. Menabites (Delawarella) sabinalensis (Young, 1963, p. 112, PI. 54, figure 2, PI. 63, figures 1, 3, 4, text-figures 20c, 2 le, 26c) is a boldly and distantly ribbed species, with weakly developed tubercles and quadri-tuberculate ornamentation in adults. However, tubercles are absent in our specimens.
Occurrence. Only known from the upper Tepeyac section close to the early-middle Campanian boundary.
Menabites (Delawarella) vanuxemi (Morton, 1830)
Figure 16A-F, Table 7
1830 Ammonites vanuxemi Morton, p. 244, plate 3, figures 2-4.
 1962 Submortoniceras vanuxemi (Morton); Reeside, p. 133, plate 72, figures 4, 5 (with synonymy)
1962 Submortoniceras vanuxemi (Morton); Reeside, p. 133, plate 72, figures 4, 5 (with synonymy)
1963 Submortoniceras vanuxemi (Morton); Young, p. 98, plate 54, figure 3; plate 56, figure 2; plate 57, figure 7; plate 58, figure 3; plate 67, figure 3; plate 69, figure 1, 2, 6; text-figures 12c, 12e, 26d- 26e (with synonymy)
1967 Submortoniceras vanuxemi (Morton); Wolleben, p. 1156, plate 17, figure 1, plate 148, figure 3
1992b Menabites (Delawarella) vanuxemi (Morton); Cobban and Kennedy, p. 76, plate 2, figures 2-4 (with additional synonymy)
1993a Menabites (Delawarella) vanuxemi (Morton,1830); Kennedy and Cobban, p. 838, figures 11.3, 11.7, 11.10, 11.11, 11.12, 12.5, 12.6, 12.17-12.19, 13.1
1997a Menabites (Delawarella) vanuxemi (Morton, 1830); Kennedy et al., p. 27, figure 18a
2013 Menabites (Delawarella) vanuxemi (Morton, 1830); Ifrim et al., p. 889, figures 3.8, 3.9, 4.5, 4.6
Types. The holotype, by monotypy, is No. 19492 in the collection of the Academy of Natural Sciences in Philadelphia (Morton, 1830, plate 3, figures 2-4; refigured by Reeside, 1962, plate 72, figures 4, 5). It is from the Chesapeake and Delaware Canal in Delaware, USA.
Material. Eight specimens, CPC-2379, -2390, -2391, -2575, -2578, -2579, -2587, and -2591, and numerous uncollected specimens from bed TPY33 to TPY77.
Description. Moderately involute shell with compressed whorls; the WB/WH ratio is between 0.43 and 0.67 and descreases upsection; U/D is between 0.16 and 0.27. The umbilical wall bends narrowly into parallel, then convergent flanks; greatest whorl width is below mid-flank. The ventrolateral shoulders are narrowly rounded. Eight to nine umbilical bullae per half whorl give rise to pairs of straight, moderately prorsiradiate ribs. Secondaries are intercalated. Weak but pointed tubercles are present on the ventrolateral shoulder. Ribs terminate at small external clavi; there is a smooth zone between these and the siphonal ridge.
Dimensions. Table 7.
Remarks. Among the finely ornamented species of this subgenus, this is the least compressed form. Menabites (Delawarella) mariscalense is almost smooth at similar diameters, M. (D.) uddeni and the other compressed species differ in their type of ribbing at similar diameters.
Occurrence. Records are from the upper lower Campanian of the Atlantic Coast, northern and western Gulf Coastal Plain, and the Western Interior of North America (Reeside, 1962; Young, 1963; Wolleben, 1967, 1977; Cobban and Kennedy, 1992a, 1992b; Larson et al., 1997; Ifrim et al., 2013). The species was described to be restricted to the upper lower Campanian, but at Tepeyac it was identified near the base of the Campanian.
Menabites (Delawarella) mariscalense (Young, 1963)
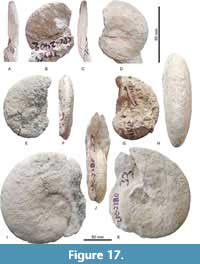
Figure 17A-K, Table 8
1963 Submortoniceras mariscalense, n. sp.; Young, p. 104, plate 59, figure 3; plate 60, figures 1, 4-6; text-figure 14b, f
 Type. The holotype is BEG-20478 by original designation by Young (1963, p. 104, plate 59, figure 3; plate 60, figures 1, 4-6; text-figure 14b, f). It is from “the concretion horizon on the west side of Mariscal Mountain, Big Bend National Park, Brewster County,” Texas.
Type. The holotype is BEG-20478 by original designation by Young (1963, p. 104, plate 59, figure 3; plate 60, figures 1, 4-6; text-figure 14b, f). It is from “the concretion horizon on the west side of Mariscal Mountain, Big Bend National Park, Brewster County,” Texas.
Material. Five specimens, CPC-2380, -2401, -2553, -2565, and -2566.
Description. Moderately compressed whorl with involute coiling, U/D is 0.16 to 0.19, WB/WH is 0.43 to 0.58, descreasing throughout ontogeny. The ornament is vanuxemi -like, with fine ribs rising from small umbilical tubercles, but ribs efface on the flanks so that the outer whorl is almost smooth. At D > 50 mm, flanks and venter are smooth, and the venter is moderately wide and rounded. Only umbilical tubercles persist into this largest stage.
Dimensions. Table 8.
Remarks. Submortoniceras mariscalense Young, 1963 (p. 104, plate 59, figure 3, p1. 60, figures 1, 4-6, text-figures 14b, f) is based on a single specimen to which five more are added here. It is is more evolute and smoother than all other species of the discoidal Menabites (Delawarella) group.
The species was hitherto included into Submortoniceras, but is here interpeted to have evolved from Menabites (Delawarella) vanuxemi. It is therefore here assigned to this genus. However, both are also close to compressed species from Madagascar, potentially indicating another relationship.
Occurrence. This species is endemic to the lower Campanian of western Texas and across the border into northeastern Mexico.
Menabites (Delawarella) vandaliense (Young, 1963)
Figure 18A-B
1963 Submortoniceras vandaliense, n. sp.; Young, p. 102, p1. 55, figures 6, 7; text-figures 26a
?1993a Submortoniceras uddeni Young, 1963; Kennedy and Cobban, p. 842, figures 10.1, 12.3, 12.4, 12.7, 12.8
 Type. The holotype is UT-30638, by original designation by Young (1963, p. 102, plate 55, figures 6, 7; text-figure 26a). It is from the Blossom Sand in Red River County, Texas.
Type. The holotype is UT-30638, by original designation by Young (1963, p. 102, plate 55, figures 6, 7; text-figure 26a). It is from the Blossom Sand in Red River County, Texas.
Material. One specimen, CPC-2576 from bed TPY77.
Description. Involute. Whorl section compressed, with greatest width at mid-flank. Flanks converge moderately towards the subrounded venter. Nineteen pointed umbilical tubercles give rise to branching prorsiradiate ribs which efface on the flanks. They strengthen on the outer flank and form numerous fine oblique bullae on the ventrolateral shoulder. They terminate before reaching the siphonal keel so that the latter is accompanied by narrow smooth grooves.
Dimensions. CPC-2576, D: 90, WB: 12 (0.17), WH: 29 (0.42), WB/WH: 0.41, U: 11 (0.16), number of ribs: 50.
Remarks. This rare species was based on a single specimen with fine ribbing presented by Young (1963), to which another is added here. It differs from Menabites (Delawarella) vanuxemi in the weakening of the outer tubercles at larger diameters and lateral effacing of the ribs. In this aspect it is similar to M. (D.) uddeni, but ventrolateral ornament is finer. Young (1963) already discussed that Submortoniceras vandaliense and S. sancarlosense may be conspecific, but this question cannot be answered here.
The specimen described as Submortoniceras uddeni Young, 1963, by Kennedy and Cobban (1993a, p. 842, figures 10.1, 12.3, 12.4, 12.7, 12.8) has finer ornament than the nominate species and is better placed in Menabites (Delawarella) vandaliense. It is from the Merchantville Formation exposed on the flanks of the Chesapeake and Delaware Canal 1.65 km (1 mi) east of Summit Bridge on the north side of the canal, New Castle County, Delaware, and thus lacks precise stratification.
Occurrence. Upper Santonian of Texas (but see discussion on the age of the Blossom Sand below), Merchantville Formation of New Castle County, Delaware, USA, and lower Campanian of Tepeyac, Mexico. Where precisely dated the species is from the lower Campanian.
Menabites (Delawarella) uddeni (Young, 1963)
Figure 18, Table 9
1963 Submortoniceras uddeni, n. sp.; Young, p. 105, p1. 59, figures 1, 2, 4-9; pI. 60, figures 2, 3, 7, 9, 10; text-figures 14d, e, 28c
non 1993a Submortoniceras uddeni Young, 1963; Kennedy and Cobban, p. 842, figures 10.1, 12.3, 12.4, 12.7, 12.8 [= possibly M. (D.) vandaliense).
Type. The holotype is USNM-130739, by original designation of Young (1963, p. 105, plate 59, figures 5, 7-9; text-figure 14e) from near the top of the Terlingua Formation near the east end of Maverick Mountain, Brewster County, Texas.
Material. Eight specimens, CPC-2396, -2399, -2400, -2402, -2405, -2415, -2416, and -2574.
Description. Involute shell, U/D is between 0.2 and 0.1 and decreases with growth. The whorl section is compressed, with WB/WH ranging between 0.58 and 0.29. Rounded umbilical tubercles give rise to moderately prorsiradiate ribs, which efface on the flanks. Fifty-eight ribs are present on the last whorl at D=80 mm.
Dimensions. Table 9.
Remarks. This species is closely related to Menabites (Delawarella) mariscalense, but differs from the latter by a stronger and more prorsiradiate ventrolateral ornament and the absence of a rounded venter near the aperture. M. (D.) vandaliense is also closely related but differs by more delicate, more abundant, and more rounded and less prominent ventrolateral tubercles.
According to Young (1963), the species is restricted to the upper Menabites (Delawarella) tequesquitense zone, but at Tepeyac it occurs at higher levels of the lower Campanian.
Occurrence. Lower lower Campanian in Texas, upper lower Campanian at Tepeyac.
Subgenus BERERELLA Collignon, 1948
Type species. Menabites (Bererella) bererensis Collignon, 1948. Widely umbilicate and flat conchs with few, widely spaced ribs; discussed by Klinger and Kennedy (1980).
Menabites (Bererella) walnutensis (Young, 1963)
Figure 19A-G, Figure 20A-F, Figure 21 A-F, Figure 22, Figure 23 A-C, Table 10
 1963 Menabites s.l. walnutensis n. sp.; Young, p.109, plate 58, figures 1, 4; text-figures 20e-f, 26k
1963 Menabites s.l. walnutensis n. sp.; Young, p.109, plate 58, figures 1, 4; text-figures 20e-f, 26k
 1963 Submortoniceras chicoense (Trask, 1856), Young, p. 106, plate 57, figures 13; text-figures 11e-f, 12d
1963 Submortoniceras chicoense (Trask, 1856), Young, p. 106, plate 57, figures 13; text-figures 11e-f, 12d
1993 ? Menabites (Bererella) sp., Kennedy and Cobban, p. 841, figures 14.36, 14.38, 14.39
1997b Menabites (Bererella) walnutensis (Young, 1963); Kennedy et al., p. 4, figure 3a-k, figures 4a-f, 5a-k, 6a-c
Type. The holotype is UT-18, by original designation by Young (1963, p. 109, plate 58, figures 1, 4; text-figures 20ef, 26k). It is from Bureau of Economic Geology locality 226-T-4 in Travis County, Texas.
Material. Twenty-eight specimens, CPC-2392, -2394, -2395, -2398, -2403, -2406 to -2414, -2417 to -2420, -2549, -2551, -2570, -2572, -2573, -2577, -2580, -2581, -2583, and -2589 from beds TPY46 upwards, but mostly in beds TPY73 to TPY77.
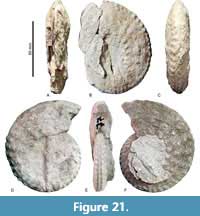 Description. Medium-sized, moderately involute shells (U/D between 0.21 and 0.46, increasing with D). The fifth, submarginal row of tubercles is always weak.
Description. Medium-sized, moderately involute shells (U/D between 0.21 and 0.46, increasing with D). The fifth, submarginal row of tubercles is always weak.
Dimensions. Table 10.
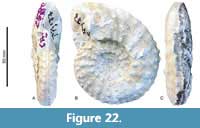 Remarks. Menabites (Bererella) walnutensis (Young, 1963) is based on a stout specimen (WB/WH ca. 0.9) with coarse ribbing and strong tuberculation. Nevertheless, Kennedy et al. (1997b) demonstrated a wide intraspecific variation and morphological dimorphism between stout and gracile forms of this species. Our specimens thus majorly correspond to the stout specimens and show a similarly wide range of morphologies (Figure 23). This suggests a wide intraspecific variation, as reported for Pseudoschloenbachia (P.) mexicana (see Ifrim et al., 2013). With
Remarks. Menabites (Bererella) walnutensis (Young, 1963) is based on a stout specimen (WB/WH ca. 0.9) with coarse ribbing and strong tuberculation. Nevertheless, Kennedy et al. (1997b) demonstrated a wide intraspecific variation and morphological dimorphism between stout and gracile forms of this species. Our specimens thus majorly correspond to the stout specimens and show a similarly wide range of morphologies (Figure 23). This suggests a wide intraspecific variation, as reported for Pseudoschloenbachia (P.) mexicana (see Ifrim et al., 2013). With 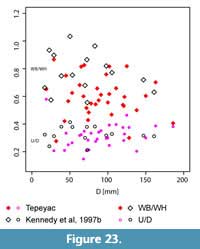 this in mind we assign the Tepeyac specimens to M. (B.) walnutensis.
this in mind we assign the Tepeyac specimens to M. (B.) walnutensis.
Occurrence. Merchantville Formation. New Jersey; central and western Texas (Young, 1963; Kennedy et al., 1997b), and now northeastern Coahuila.
Superfamily DESMOCERATOIDEA von Zittel, 1895
Family MUNIERICERATIDAE Wright, 1952
Genus and subgenus PSEUDOSCHLOENBACHIA Spath, 1921
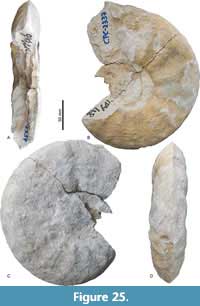
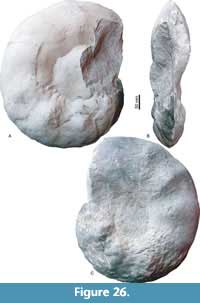
Pseudoschloenbachia (Pseudoschloenbachia) mexicana Renz, 1936
Figure 24, Figure 25, Figure 26, Table 11
1936 Schloenbachia bertrandi de Grossouvre var. mexicana Renz, p. 6, plate 1, figure 2; plate 3, figure 1
 1963 Pseudoschloenbachia mexicana Renz; Young, p. 121, plate 29, figures 3-4; plate 30, figures 1-7; plate 31, figures 1, 3-9; plate 32, figures 1-6; plate 33, figures 1-3, 5-7, plate 44, figure 1; text-figures 13e, 28d, 29b, d
1963 Pseudoschloenbachia mexicana Renz; Young, p. 121, plate 29, figures 3-4; plate 30, figures 1-7; plate 31, figures 1, 3-9; plate 32, figures 1-6; plate 33, figures 1-3, 5-7, plate 44, figure 1; text-figures 13e, 28d, 29b, d
1963 Pseudoschloenbachia sp. juv. cf. P. mexicana Renz; Young, plate 30, figure 9; plate 31, figure 2; plate 33, figure 4
 1967 Pseudoschloenbachia mexicana Renz; Wolleben, p. 1157, plate 149, figures 4-5
1967 Pseudoschloenbachia mexicana Renz; Wolleben, p. 1157, plate 149, figures 4-5
1991 Pseudoschloenbachia (Pseudoschloenbachia) mexicana Renz; Kennedy and Cobban, p. 172, figures 6.1-2, 7.5-39, 10.17-19, 10.22-23, 12.4
2013 Pseudoschloenbachia (Pseudoschloenbachia) mexicana (Renz, 1936); Ifrim et al., p. 885, figures 4.9-4.17, 5.1-5.11, 6.1, 7
Types. Lectotype is NMBE 5024066, designated as holotype by Young (1963, p. 121). It is the original of Renz (1936, pl. 3, figure 1). The paralectotype, which corresponds to the original of Renz (1936, pl. 1, figure 2), was lost (U. Menkveld, NMBE, personal comm. 2018). The type specimens are from the Arroyo Tecolote and Arroyo del Fresno, i.e., a few to a maximum of 10 km north of our working area in northeastern Coahuila, Mexico.
 Material. Ten internal moulds, CPC-2334 to -2339 from beds TPY13 to TPY53; CPC-2447, -2450, and -2451 collected earlier from the working area.
Material. Ten internal moulds, CPC-2334 to -2339 from beds TPY13 to TPY53; CPC-2447, -2450, and -2451 collected earlier from the working area.
Description. Oxyxcone, moderately involute. U is mostly covered with sediment, U/D is about 0.25, except for the largest specimen for which it is >670 mm. WB/WH ratio ranges from 0.4 to 0.9. The umbilical wall is short and steep. It bends towards the flanks in a narrow arch in juveniles and in a wide arch in adults. Greatest whorl width is at mid-flank in juveniles and in the umbilical half in adults. Flanks bend towards the acute venter ventrolaterally in a moderately wide arch in juveniles, in a wide arch in late juveniles, and they are flat and convergent in adults. Juvenile ornament consists of an umbilical row of distant tubercles that give rise to seven straight primary ribs per half whorl. In between, secondary ribs are intercalated rising at mid-flank. Tubercles along the ventrolateral shoulder accentuate this narrow edge in jveniles. Ventral to this row of tubercles, all ribs show equal strength and are prorsiradiate and straight, fading towards the venter. The narrow ventrolateral shoulder and the tubercles disappear at larger D, and ribs bifurcate; in adults the ribs are simple, broad,and shallow, with a broad tubercle in the center. The suture line shows an asymmetrically trifid L which exceeds E in depth. Auxiliary lobes are also asymmetrically trifid.
Dimensions. Table 11.
Remarks. An exceedingly wide morphological variability of Pseudoschloenbachia (P.) mexicana (Renz, 1936) was documented by Ifrim et al. (2013) based on abundant pyritized moulds from the Union y Progreso site in western Coahuila. This full range of morphological variability is not identified in the Tepeyac material; while CPC-2339 is a rather stout specimen, most other individuals conform discoidal end members. The differences in the morphological habitus may relate to the larger sizes of Tepeyac specimens in comparison to the juveniles documented by Ifrim et al. (2013).
Occurrence. Pseudoschloenbachia (P.) mexicana Renz is restricted to the upper Santonian and lower Campanian of Texas, Chihuahua (Renz, 1936; Young, 1963; Wolleben, 1967; Kennedy and Cobban, 1991; Becker et al., 1998), western (Ifrim et al., 2013) and north-eastern Coahuila.
Family DESMOCERATIDAE von Zittel, 1895
Subfamily PUZOSIINAE Spath, 1922
Synonymy of this subfamily after Wright and Kennedy (1984-1996). Their interpretation includes the Hauericeratinae Matsumoto, 1938.
Genus and subgenus PARAPUZOSIA Nowak, 1913
Parapuzosia (Parapuzosia) leptophylla Sharpe, 1857
This species was recently re-described by Kennedy (2019). The reader is referred to this paper for systematic description. Occurrences in Northeastern Mexico have been described by Ifrim et al. (in press)..
Occurrence. Lower Santonian records of Parapuzosia (Parapuzosia) leptophylla exist from southern France (Musée de Paléontologie de Provence, 2010), Austria (Summesberger, 1978), and from England (Kennedy, 2019). In the upper Santonian, the species is distributed in England and Mexico (Ifrim et al., in press).
Parapuzosia (Parapuzosia) seppenradensis Landois, 1895
This species was recently re-described by Ifrim et al. (in press). The reader is referred to this publication. All Parapuzosia (P.) specimens >> 1 m are exclusively Campanian in age.
Occurrence. Uppermost Santonian of the north-western Gulf of Mexico, lower Campanian of Germany, northern and western Gulf of Mexico and England. The age of the specimen from Montana is unknown. Parapuzosia (Parapuzosia) seppenradensis is abundant in the Tepeyac section and ranges from the Uppermost Santonian throughout the lower Campanian.
Family PACHYDISCIDAE Spath, 1922
Genus and subgenus PACHYDISCUS Spath, 1922
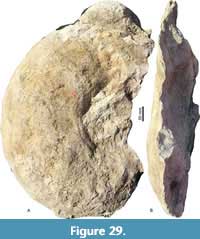
Pachydiscus (Pachydiscus) duelmensis (Schlüter, 1872)
Figure 27A-B, Figure 28A-B, Figure 29A-B, Figure 30B-E, Figure 31A, Figure 32A-B, Table 12
1872 Ammonites dülmensis Schlüter. p. 52. plate 16. figure 1-2
1894 Pachydiscus dülmensis Schlüter sp.; de Grossouvre. p. 199. plate 20
1920 Pachydiscus dülmenensis Schlüter; Köplitz. p. 69. plate 8. figure 25
1936 Pachydiscus (Parapachydiscus) jimenezi Renz. p. 3. plate 2. figure 4, 4a
1963 Eupachydiscus jimenezi Young. p. 59, plate 14, figures 1, 5; plate 16, figure 4; text-figure 10k
1963 Parapuzosia paulsoni Young. p. 53, plate 11. figures 3-5; plate 12. figures 1-4; plate 15. figure 10; plate 17. figure 9; plate 19. figures 3-4; text-figures 8a-b. 9g, j, r
1986 Pachydiscus (Pachydiscus) duelmensis (Schlüter. 1872); Kennedy. p. 35. text-figure 12
1989 Pachydiscus (Pachydiscus) duelmensis (Schlüter. 1872); Jagt. p. 7. plate 1-5. plate 6. figure 1
1992a Pachydiscus (Pachydiscus) paulsoni (Young. 1963); Cobban and Kennedy. p. 69, plate 1, figures 4-7; plate 4. figures 7-9; plate 5, figures 7-8; text-figures 3a-c, 4b
1992b Pachydiscus cf. P. paulsoni (Young. 1963); Kennedy et al., p. 441
1993a Pachydiscus sp.; Kennedy and Cobban. p. 831., figures 3-4
1995 Pachydiscus (Pachydiscus) duelmenensis (Schlüter. 1872); Wippich, p. 52., plate 2. figures 1-2; plate 4. figures 1-2
1995 Pachydiscus (Pachydiscus) duelmensis (Schlüter. 1872); Kennedy and Kaplan, p. 27, plate 9. figures 1-2; plate 10, figures 1-2; plate 11; plate 12, figures 1, 2 (with synonymy)
1997 Pachydiscus cf. P. paulsoni (Young, 1963); Kennedy et al., p. 9
2005 Pachydiscus (Pachydiscus) duelmenensis (Schlüter, 1872); Kaplan et al., p. 62, plate 6, figures 6-7; plate 10
2006 Pachydiscus (Pachydiscus) duelmensis (Schlüter, 1872); Kaplan et al., p. 34, plate 2, figures 1-3
 |
 |
 |
Type. The lectotype is GPIB 47, the original of Schlüter (1872, pl. 16, figures 1-2) by subsequent designation by (Jagt, 1989, p. 7). It is from the lower Campanian Dülmen Formation, Westphalia, Germany, and was re-figured by Kennedy (1986, figure 12) and Jagt (1989, pl. 5).
Material. Five collected specimens, CPC-2193, -2195, -2197, -2545,and -2561 from bed TPY40 and upwards.
 Description. Involute shell (U/D between 0.24 and 0.36) with compressed oval whorl section (WB/WH is around 1 but with variation). The shell is ornamented by narrow, sharp, shallow, moderately sinuous primary ribs separated by one or two long, narrow secondary ribs. The primaries show elongated bullae from the umbilical shoulder to the inner part of flank. All ribs cross the venter with forward arching. Distance of ribs is constant, with increasing D, so that the number of ribs increases linear with D.
Description. Involute shell (U/D between 0.24 and 0.36) with compressed oval whorl section (WB/WH is around 1 but with variation). The shell is ornamented by narrow, sharp, shallow, moderately sinuous primary ribs separated by one or two long, narrow secondary ribs. The primaries show elongated bullae from the umbilical shoulder to the inner part of flank. All ribs cross the venter with forward arching. Distance of ribs is constant, with increasing D, so that the number of ribs increases linear with D.
Dimensions. Table 12.
Remarks. The specimens included here are characterized by numerous sharp, shallow primary and secondary ribs. These are also present on shells Pachydiscus (Pachydiscus) jimenezi (Renz, 1936), P. (P.) paulsoni Young, 1963 and Pachydiscus (Pachydiscus) duelmensis (Schlüter, 1872) from the lower Campaniain of Mexico, Texas, and Germany.
Specimens from the Tepeyac-Jimenez area were first described by Renz (1936). The holotype of Pachydiscus (P.) jimenezi is the specimen of Renz (1936, p. 3, pl. 2, figure 4, 4a) by monotypy. Renz (op. cit.) states that it is specimen No. 1 in the Collection Böse-Staub, housed in the Geological Institute of the University of Bern. This collection has been transferred to the Naturhistorisches Museum Bern, but the holotype is untraceable there and is here regarded as lost.
 The morphology described here is also seen in Parapuzosia paulsoni Young, 1963 (p. 53ff). The holotype of Parapuzosia paulsoni is TMM 30625, from the “Gober chalk” in McCurtain County, Oklahoma. Specimens from Texas were described by Cobban and Kennedy (1992b), in addition to unfigured specimens from Texas Cobban and Kennedy (1992a) and Alabama (Kennedy et al. (1997a). Cobban and Kennedy (1992a) shortly mention similarities of P. paulsoni Young, 1963 with Pachydiscus (Pachydiscus) duelmensis (Schlüter, 1872, p. 52, P1. 16, figures 1-2; see Kennedy, 1986a, text-figure 12). They suggested that the American species differed by weak bullae, a more compressed whorl section, and fewer, wider spaced ribs. However, these differences appear to be related to different ontogenetic stages. When plotting the morphometric data of all three species (Figure 31A–B), it becomes evident that no notable differences exist between the three closely related species regarding whorl dimensions and ribbing.
The morphology described here is also seen in Parapuzosia paulsoni Young, 1963 (p. 53ff). The holotype of Parapuzosia paulsoni is TMM 30625, from the “Gober chalk” in McCurtain County, Oklahoma. Specimens from Texas were described by Cobban and Kennedy (1992b), in addition to unfigured specimens from Texas Cobban and Kennedy (1992a) and Alabama (Kennedy et al. (1997a). Cobban and Kennedy (1992a) shortly mention similarities of P. paulsoni Young, 1963 with Pachydiscus (Pachydiscus) duelmensis (Schlüter, 1872, p. 52, P1. 16, figures 1-2; see Kennedy, 1986a, text-figure 12). They suggested that the American species differed by weak bullae, a more compressed whorl section, and fewer, wider spaced ribs. However, these differences appear to be related to different ontogenetic stages. When plotting the morphometric data of all three species (Figure 31A–B), it becomes evident that no notable differences exist between the three closely related species regarding whorl dimensions and ribbing.  Rather, each of the species is morphologically variable (Cobban and Kennedy, 1992a; Kaplan et al., 2005; Kaplan et al., 2006; Kennedy, 1986; Kennedy et al., 1997a; Kennedy and Kaplan, 1995). In fact, the holotypes of Pachydiscus (P.) duelmensis (Schlüter, 1872), P. (P.) jimenezi (Renz, 1936), and P. (P.) paulsoni (Young, 1963) plot together in all morphometric aspects (Figure 31A–B). As no other unequivocal characteristics exist between them, the three are considered to be conspecific, with Pachydiscus (P.) duelmensis as the homonym.
Rather, each of the species is morphologically variable (Cobban and Kennedy, 1992a; Kaplan et al., 2005; Kaplan et al., 2006; Kennedy, 1986; Kennedy et al., 1997a; Kennedy and Kaplan, 1995). In fact, the holotypes of Pachydiscus (P.) duelmensis (Schlüter, 1872), P. (P.) jimenezi (Renz, 1936), and P. (P.) paulsoni (Young, 1963) plot together in all morphometric aspects (Figure 31A–B). As no other unequivocal characteristics exist between them, the three are considered to be conspecific, with Pachydiscus (P.) duelmensis as the homonym.
The low rib index in CPC-2193 is probably due to abrasion of the outer flanks and venter, also allowing for destruction of the secondaries. Specimen CPC-2545 shows a smooth adult body chamber, which exludes an assignation to the genus Eupachydiscus, because the latter presents prominent ribs on the terminal whorl.
Morphometric relations of Menuites gordoni Young, 1963 resemble those of Pachydiscus (P.) duelmensis. However, ribbing differs significantly from the conspicious ornament of P. (P.) duelmensis, and separation of the two taxa is here maintained.
Occurrence. Basal to middle lower Campanian of Germany, Belgium, Netherlands, southwestern France, Oklahoma, Texas, Alabama, and Mexico. In the Tepeyac section, the taxon occurs in bed TPY40 and above this layer, i.e., in the Menabites (Delawarella) tequesquitense, M. (D.) delawarensis and Baculites taylorensis zones, and thus throughout the lower Campanian.
Genus MENUITES Spath, 1922
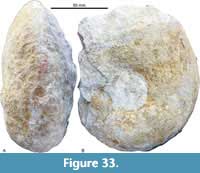
Menuites stephensoni Young, 1963
Figure 31A–B; Figure 32C-F; Figure 33A-B, Table 13
1932 Menuites n. sp., Adkins, p. 467
 1963 Menuites stephensoni, n. sp.; Young, p. 57, p1. 15, figures 1-2, text-figures 7o, 9n
1963 Menuites stephensoni, n. sp.; Young, p. 57, p1. 15, figures 1-2, text-figures 7o, 9n
1963 Menuites juv. sp., Young, p. 58, plate 15, figures 6, 7, 9, 11, 12; plate 20, figures 11, 11; text-figure 9q
1999 Menuites sp.; Kennedy and Cobban, p. 70
p.2005 Menuites portlocki portlocki (Sharpe, 1855); Kaplan et al, p. 76, plate 33, figure 3 only
p.2005 Pachydiscus pseudostobaei (Moberg [sic], 1885); Kaplan, et al., p. 66, plate 31, figures 1-2 only
Type. The holotpe is WSA-69 by original designation by Young (1963, p. 57, plate 15, figures 1, 2; text-figures 7o, 9n). It is from a horizon with calcareous concretions in the upper Taylor Marl in Travis County, Texas.
Material. Four specimens, CPC-2194,-2349, -2554, and -2562, from beds TPY17, 50, 52, and 62.
Description. Involute (U/D with moderate growth rate). Juveniles with rounded whorl sections and 28 to 32 distant, sharp, straight to slightly prorosiradiate ribs. At D >120 mm lateral and lateroventral rounded bullae develop and whorl section is gradually subrectangular. In large adults ribs become irregular and finally disappear. Suture line with a trifid L that is equal in depth to E.
Dimensions. Table 13.
Remarks. The taxon is characterized by high morphologic variability. Juveniles assigned to Menuites juv. sp. by (Ifrim et al., 2013) may differ from Menuites stephensoni with respect to their U/D ratio and rib index (Figure 31A–B) and are kept separately until intermediate growth stages are known.
Specimen RE A 4214 from questionable lower Campanian sediments of Germany was described as Menuites portlocki portlocki (Sharpe, 1855) by Kaplan et al. (2005), but rather agrees with M. stephensoni than with the upper Campanian species (compare plate 32, op. cit.).
Specimen RE A 4193 from the lower Campanian of Germany described as Pachydiscus pseudostobaei (Möberg, 1885) by Kaplan et al. (2005) also rather shows characteristics of a juvenile Menuites stephensoni as understood here.
The species is closely related to, and most probably the ancestor, of the upper Campanian Menuites portlocki (Sharpe, 1855) known from North America. Menuites portlocki differs from M. stephensoni by a stronger ornamentation (compare e.g., Błaszkiewicz, 1980), although morphometry is similar between the two species.
Occurrence. Upper Santonian to lower Campanian of the Tepeyac section. Young (1963, p. 57) states that the holotype from Texas is upper Campanian in age, but without providing further details or age calibration. In central to western Europe the taxon is known from the lower Campanian. The record from Tepeyac is thus the oldest.
Menuites sp.
Figure 34 A-D, Table 14
Material. Four specimens, CPC-2196, -2544, -2554, and -2562.
 Description. The shells are moderately involute with a moderate expansion rate. Ornament in the inner whorls of CPC-2544 is simlar to Pachydiscus (P.) duelmensis at similar D, but ribs are more numerous and more irregular, and the shell is more involute. Each specimen presents a deep constriction.
Description. The shells are moderately involute with a moderate expansion rate. Ornament in the inner whorls of CPC-2544 is simlar to Pachydiscus (P.) duelmensis at similar D, but ribs are more numerous and more irregular, and the shell is more involute. Each specimen presents a deep constriction.
Dimensions. Table 14.
Remarks. This morphotype differs from Menuites stephensoni by the absence of ribbing except at small diameters and by the presence of deep constrictions. The material may represent more than one species but specimens are not comparable as they represent different ontogenetic stages.
Suborder Ancyloceratina Wiedmann, 1966
Superfamily Turrilitoidea Gill, 1871
Family DIPLOMOCERATIDAE Spath, 1926
Subfamily DIPLOMOCERATIDAE Spath, 1926
Genus GLYPTOXOCERAS Spath, 1925
Shafts are more or less curved, no straight shafts, simple ribs, circular to oval cross section
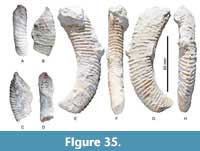
Glyptoxoceras texanum
Kennedy, Landman, and Cobban, 2001
Figure 35A-H
1992 Glyptoxoceras sp.; Cobban and Kennedy, p. 80, plate 4, figures 10-11; plate 6, figures 5-6; plate 7, figures 7-8
 1997a Glyptoxoceras sp.; Kennedy et al., p. 35, figure 22c-e
1997a Glyptoxoceras sp.; Kennedy et al., p. 35, figure 22c-e
2001 Glyptoxoceras texanum Kennedy, Landman and Cobban, p. 7, figure 4
Type. The holotype is USNM 490970, by original designation by Kennedy, Landman and Cobban (2001, figure 4c-e). It is from the top of the Blossom Sand at USGS Mesozoic locality 18120, between Detroit and Bagwell, Red River County, Texas
Material. Two fragments, CPC-2349 from bed TPY48-50 and CPC-2610 from bed TPY73.
Description. Open coil with different intensities of coiling, moderately helicoidal. Whorl section is ovoid, WB/WH is between 0.60 and 0.74, but the ratio may be distorted due to crushing of the shell. Ribs are closely spaced, with a rib index of between 8 and 10, increasing with WH. Ribs are weak and straight across the dorsum, strong and slightly convex across the dorsolateral margin, strong, straight, and rursiradiate on the flanks, and transverse on the venter where they reach their maximum height. CPC-2349 shows a deep constriction which is flanked by a flared adapertural rib.
Dimensions. CPC-2346, WB: 14, WH: 19, WB/WH 0.74, L: 15, ribs per WH: 8. CPC-2610, smaller end,WB: 12, WH: 20, WB/WH: 0.60, ribs per WH: 10, L: 65 to larger end, there WB: 15, WH: 24, WB/WH 0.63.
Remarks. Our specimens are closely resemble the lower Campanian Glyptoxoceras sp. described by Kennedy et al. (1997b) and Cobban and Kennedy (1992b), which are here included in the present species.
Glyptoxoceras retrorsum (Schlüter, 1972) from the lower Campanian of Germany is similar to our specimens with respect to ribbing and dimensions near the aperture (e.g., Kaplan and Kennedy 2005, p. 310, plate 46, figures 1-3; plate 49, figures 1-2), but lacks flared ribs. Glyptoxoceras novimexicanus (Reeside, 1927) is morphologically indistinguishable from G. retrorsum and is here regarded as a synonym.
The present material is ribbed more finely than the upper Santonian Glyptoxoceras sp. of (Kennedy and Cobban, 1991). Glyptoxoceras aquisgranense (Schlüter, 1972) from the lower Campanian of Germany has a lower rib index and a more rounded whorl section (Kennedy and Kaplan, 1995). This species was also recorded from lower Campanian of the Atlantic Coast (Kennedy et al., 1997b). A more circular section and significantly coarser ribs are present in Glyptoxoceras ellisoni Young, 1963 and Glyptoxoceras rubei (Reeside, 1927).
Two groups may thus be distinguished within lower Campanian members of the genus Glyptoxoceras, i.e., delicately ribbed taxa including Glyptoxoceras retrorsum and G. texanum, and coarsely ribbed ones such as G. aquisgranense and G. ellisoni. The presence, or absence, of constrictions may distinguish species in these two groups of Glyptoxoceras, but this cannot be decided to date.
Occurrence. Uppermost Santonian of northeastern Texas, Menabites (Delawarella) delawarensis zone of New Jersey, lower Campanian of northeastern Coahuila, and middle Campanian of Texas.
Family Nostoceratidae
Genus AMAPONDELLA Klinger and Kennedy, 1997
Type species. Heteroceras amapondense van Hoepen, 1921 by original designation of Klinger and Kennedy (1997).
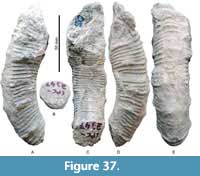
Amapondella amapondense (van Hoepen, 1921)
Figure 36A-E, Figure 37A-E, Table 15
1921 Heteroceras amapondense van Hoepen, p. 17, plate 4, figures 1, 2
 1969 Anaklinoceras (?) stephensoni Collignon, p. 50, plate 532 (figure 2096)
1969 Anaklinoceras (?) stephensoni Collignon, p. 50, plate 532 (figure 2096)
1969 Hyphantoceras reussianum d’Orb.; Collignon, p. 29, plate 523 (figure 2064), p. 38, plate 527 (figures 2079-2080); p. 48, plate 531 (figure 2095)
1978 Hyphantoceras (Madagascarites ?) amapondense (van Hoepen); Summesberger, p. 123, plate 3, figure 19; text-figure 13
1983 Hyphantoceras amapondense (van Hoepen); Lewy, p. 24, figures 1-6 (with synonymy)
1985 Hypthantoceras (Madagascarites) amapondense (van Hoepen, 1921); Klinger, p. 6, figure 41-k
1991 Hyphantoceras (?) amapondense (van Hoepen, 1921); Kennedy, p. 181, figure 9.9, 9.10
1997 Eubostrychoceras (Amapondella) amapondense van Hoepen; Klinger and Kennedy, p. 244, figures 12- 14a, 15a-d, 17a-c
2003 Eubostrychoceras (Amapondella) amapondense (van Hoepen); Klinger and Kennedy p. 235, figures 5-6, 7a, 8a-d, 9a-c (with synonymy)
 2007 Amapondella amapondense (van Hoepen, 1921); Klinger and Kennedy, p. 101, figures 5a-c, 10i-j, 11, 12b-i, 13a-e, m
2007 Amapondella amapondense (van Hoepen, 1921); Klinger and Kennedy, p. 101, figures 5a-c, 10i-j, 11, 12b-i, 13a-e, m
Type. Holotype is the specimen figured by van Hoepen (1921, plate 4, figures 1-2) by original designation. It is from the upper Santonian or lower Campanian of the Mzamba Formation at the type section (locality 1), Eastern Cape Province, South Africa. SAM-PCZ18722 is a cast of the holotype.
Material. Five specimens, CPC-2345 to 2347 from bed TPY50, CPC-2351 to -2352 from bed TPY25.
Description. Open, slightly helicoidal, more or less even coiling, U/D is ca. 0.47. Circular to more subquadrate whorl sections at larger D, while ovoid whorl sections may result from deformation. Ribs are weak, feebly prorsiradiate, sweep back and strengthen on both upper and lower whorl faces; they are markedly oblique and reach their maximum strength on the outer whorl face. Every fourth to sixth rib is strengthened, forming a pronounced flare; these are conspicuous on the lateral, ventrolateral and ventral whorls.
Dimensions. Table 15.
Remarks. This species was extensively discussed by Lewy (1983), and additional comments were provided by Klinger and Kennedy (2003) and Klinger et al. (2007). The latter authors discussed dimorphism in this species.
Occurrence. Middle Santonian to lower upper Santonian of France, middle Santonian to lower Campanian of South Africa, upper Santonian of Israel, Austria, and Mississippi, uppermost Santonian to lower Campanian of Coahuila, lower Campanian of Madagascar.
Family Baculitidae Gill, 1871
Genus Baculites Lamarck, 1799
Baculites taylorensis Adkins, 1929
Figure 38A-B, G-H, Table 16
1929 Baculites taylorensis Adkins, p. 204, plate 5, figures 9-11
 1993a Baculites taylorensis Adkins; Kenney and Cobban, p. 93, figures 10.1-9, 10.11-12, 10.16, 10.18-19, 11.1-2 (Kennedy and Cobban, 1993b)
1993a Baculites taylorensis Adkins; Kenney and Cobban, p. 93, figures 10.1-9, 10.11-12, 10.16, 10.18-19, 11.1-2 (Kennedy and Cobban, 1993b)
1993b Baculites taylorensis Adkins; Kennedy and Cobban, p. 143, plate 6, figures 1-9, plate 7, figures 1-6, 10-13; text-figure 8b, d (Kennedy and Cobban, 1993c)
1994 Baculites taylorensis Adkins; Cobban and Kennedy, p. D6, plate 3, figures 5-19; text-figure 7 (Cobban and Kennedy, 1994)
2001 Baculites taylorensis Adkins; Kennedy and Cobban, p. 25, plate 6, figures 8-9 (Kennedy and Cobban, 2001) (with synonymy)
2013 Baculites taylorensis Adkins, 1929; Ifrim et al., p. 234, figure 5b-f, figure 6f-y, figure 7a-l, figure 8a-k, figure 9a-l
Type. The holotype is TMM 21014 by monotypy. This specimen is from the basal Pecan Gap Chalk, Travis County, Texas.
Material. Two specimens, CPC-2198 and -2199, both internal moulds, from beds TPY64 and 77, respectively.
Description. Shell substraight with a low expansion rate. Whorl section is stout and ovate, with a widely-rounded dorsum, dorsolaterally parallel and ventrolaterally convergent flanks which terminate in a moderately narrow rounded venter. Rib index is 1 to 0.5. These nodes arise at the dorsolateral shoulder and increase in size on the dorsal third to half of the flank; they are rounded and weak in CPC-2183 and stronger in CPC-2199. Ribs and nodes fade markedly on both the dorsum and the ventral portion of the flank.
Dimensions. Table 16.
Remarks. The two specimens and their ornamentation match the abundant specimens described from Arkansas, Texas and Coahuila.
Occurrence. Lower-upper Campanian of the northern and north-western Gulf coast. The FAD of the species marks the base for an uppermost lower Campanian biozone in this region.
Baculites cf. haresi Reeside, 1927
Figure 38C-F, I-L
1927 Baculites ovatus var. haresi, Reeside, p. 10. plate 6, figures 5-10, plate 7, figures 9-10 (with synonymy)
1992a Baculites haresi Reeside; Cobban and Kennedy, p. 449. figures 6.5-6.8, 6.11-6.13, 6.16-6.24, 7.1 (with additional synonymy)
1993a Baculites haresi Reeside, 1927; Kennedy and Cobban, p. 844, figures 14.18-14.35, 14.37, 15.2, 16.1-16.6 (with additional synonymy)
1997 Baculites haresi, Reeside; Larson, p. 20
2013 Baculites haresi Reeside, 1927; Ifrim et al., p. 890, figures 8.1-8.13, 9.1-9.6, 10.1-10.7, table 5
Type. The lectotype is USNM 73296, the original of Reeside (1927, pl. 6, figs 5-6), by subsequent designation of Cobban and Kennedy (1992a, p. 449). It comes from the Elk Basin Sandstone Member of the Telegraph Creek Formation, Wyoming, USA.
Material. Four specimens, CPC-2604 to -2607, from beds TPY52, 57, and 77.
Description. Compressed whorl section with rounded dorsum and venter. The shell is smooth or ornamented by faint bullae and falcate ribs in dorsolateral position.
Remarks. Our specimens are deformed. Their characteristics correspond to the lectotype of Reeside (1927, pl. 6, figures 5-6) with respect to whorl section, as far as reconstructible, and the faint ornament. The suture is little incised. Superficially, B. haresi Reeside, 1927 resembles B. scotti Cobban, 1958 and B. eliasi Cobban, 1958, but the latter two species differ in the shape and higher complexity of their sutural elements.
Occurrence. Baculites haresi is widely distributed in lower Campanian strata of the Western Interior Seaway, from Wyoming to Texas (Larson et al., 1997), and also present in Coahuila (Ifrim et al., 2013, and herein). Isolated records also exist from the lower Campanian of New Jersey (fide Klinger and Kennedy, 2001) and Greenland (Birkelund, 1965).
Superfamily SCAPHITOIDEA Gill, 1871
Family SCAPHITIDAE Gill, 1871
Subfamily SCAPHITINAE Spath, 1926
Genus and subgenus SCAPHITES Parkinson, 1811
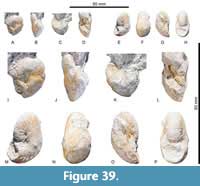
Scaphites (Scaphites) hippocrepis (DeKay, 1828) III Cobban, 1969
Figure 39A-P
1828 Ammonites hippocrepis Dekay, p. 273, plate 5, figure 5.
 1969 Scaphites hippocrepis (DeKay) III; Cobban, plate 3, figures 1-25; plate 4, figures 35-49; plate 5, figures 36-40; text figures 2, 4, 10, 11 (with synonymy)
1969 Scaphites hippocrepis (DeKay) III; Cobban, plate 3, figures 1-25; plate 4, figures 35-49; plate 5, figures 36-40; text figures 2, 4, 10, 11 (with synonymy)
1986 Scaphites (Scaphites) hippocrepis (Dekay, 1828); Kennedy, p. 118, plate 13, figure 5; plate 19, figures 1-34; plate 20, figures 1-6, 9, 1 O; plate 21, figures 1-11; plate 23, figures 2, 3 (with synonymy)
1992 Scaphites (Scaphites) hippocrepis (Dekay) III; Cobban and Kennedy, p. 451, figures 4.1-4.3, 4.13-4.20, 4.22-4.23, 4.28, 4.92, 6.1-6.4, 6.10, 7.2
1993 Scaphites (Scaphites) hippocrepis (Dekay, 1828); Kennedy and Cobban, p. 845, figures 15.1, 17.1-17.32
1995 Scaphites (Scaphites) hippocrepis (Dekay, 1828) form III; Jagt, Burnett and Kennedy, p. 57, plate 3, figure 3
1996 Scaphites (Scaphites) hippocrepis (Dekay, 1828), form III of Cobban, 1969; Kennedy and Christensen, p. 115, figure 32
2000a Scaphites hippocrepis (Dekay) III; Küchler, plate 8, figures 3-6
2004 Scaphites (Scaphites) hippocrepis (Dekay, 1828), III sensu Cobban, 1969; Jagt, Walaszczyk, Yazykova, and Zaton, p. 577, plate 1, figure 4
2005 Scaphites hippocrepis (Dekay, 1828), Kaplan et al., p. 138, plate 57, figures 1-30; plate 58, figures 1-15; plate 62, figure 5 (with synonymy)
?2013 Scaphites sp. ex gr. S. hippocrepis (DeKay, 1828); Ifrim et al., p. 895, figures 9.23-9.28, 12.1-12.15, table 7
Type. Neotype, designated by (Kennedy, 1986, p. 118), is no. 19483 in the collections of the Philadelphia Academy of Sciences. It is the holotype of Scaphites cuvieri Morton, 1834, p. 109, p1. 7, figure 1, refigured by Whitfield, 1892, p1. 44, figure 8, Richards, 1953, text-figure 224, and Reeside, 1962, p1. 71, figures 1-5. The specimen is from the deep cut of the Chesapeake and Delaware Canal, Delaware.
Material. Two specimens, CPC-2599 and -2600, both from bed TPY732.
Description. Inflated to globular whorl section. WB/WH is <1.76 in juveniles but decreases to 1.03 in shells with D >16 mm. Maximum width is reached at mid-flank. The expansion rate is high in early growth stages but decreases at larger diameters, as indicated by similar WBs of the early and later whorl. The final whorl section grades towards a subtriangular shape. U is small, with U/D decreasing from 0.2 in early juveniles to 0.1 at large D. The umbilicus is deep. Umbilical walls are rounded and grade into rounded flanks, which are short due to a widely rounded venter. Eight tubercles are present per half whorl and give rise to bullae that each grade into pairs of moderately prorsiradiate convex ribs. These are occasionally intercalated by secondaries, resulting in ca. 17 primaries and secondaries per half whorl. The suture is simple, with shallow, subrectangular, asymmetrically bifid elements. Only E/L and L are asymmetrically trifid.
Dimensions. CPC-2599, D: 17.0; CPC-2600, D: 21.5, WB: 12.0 (0.6), WH: 8.0 (0.4), WB/WH: 1.50, U: 6.0 (0.28), ribs per whorl: 38.
Remarks. Ornament, particularly the type of ribbing, relates our specimens to morphotype III of S. hippocrepis. This morphotype is the most widespread which provides good biostratigraphic control in both North America and Europe. Size differences exist between CPC-2599 and -2600 and suggest that the two represent a micro- and a macroconch.
Occurrence. In North America S. hippocrepis III is known from the middle lower Campanian of the Western Interior, Gulf Coast and Atlantic Coast Plan (Cobban, 1969; Cobban and Kennedy, 1992a, 1992b), western Coahuila (Ifrim et al., 2013) (Pervinquière, 1907), and the North American Pacific coast (Anderson, 1958). Records are also from France (Kennedy, 1986), Poland, Germany, and Netherlands (Jagt et al., 2004; Kaplan et al., 2005), Sweden (Kennedy and Christensen, 1997), and Tunisia. Lower middle Campanian records of S. hippocrepis III also exist from Spain (Küchler, 2000) and Germany (Kaplan et al., 2005).
AMMONID ZONATION AND THE SANTONIAN-CAMPANIAN BOUNDARY
 Young (1963, 1985) and Kennedy et al. (1997a) used the first occurrence (FO) of Menabites (Delawarella) tequesquitense to define the base of the Campanian in Texas (Figure 40), but their biozonation within the lower Campanian provides slight differences (Figure 40). Gale et al. (2008) noted, that the FO of Uintacrinus anglicus was to be placed slightly above the FO of M. (D.) tequesquitense, within this ammonite zone. This crinoid is another good index for the base of the Campanian, similar to the LO of Marsupites testudinarius (Gale et al., 1995; Kennedy et al., 1997a; Gale et al., 2008), which is also placed within the M. (D.) tequesquitense zone (Kennedy et al., 1997a). Another good marker for the base of the Campanian is provided by the stable carbon isotope curve which shifts from a generally increasing towards a decreasing trend in the Santonian-Campanian Boundary Event (SCBE, Thibault, 2016, and Figure 2). This excursion is the best marker for the base of the Campanian in bed 30 of the Tepeyac section.
Young (1963, 1985) and Kennedy et al. (1997a) used the first occurrence (FO) of Menabites (Delawarella) tequesquitense to define the base of the Campanian in Texas (Figure 40), but their biozonation within the lower Campanian provides slight differences (Figure 40). Gale et al. (2008) noted, that the FO of Uintacrinus anglicus was to be placed slightly above the FO of M. (D.) tequesquitense, within this ammonite zone. This crinoid is another good index for the base of the Campanian, similar to the LO of Marsupites testudinarius (Gale et al., 1995; Kennedy et al., 1997a; Gale et al., 2008), which is also placed within the M. (D.) tequesquitense zone (Kennedy et al., 1997a). Another good marker for the base of the Campanian is provided by the stable carbon isotope curve which shifts from a generally increasing towards a decreasing trend in the Santonian-Campanian Boundary Event (SCBE, Thibault, 2016, and Figure 2). This excursion is the best marker for the base of the Campanian in bed 30 of the Tepeyac section.
The FO of the giant ammonite Parapuzosia (Parapuzosia) seppenradensis provides to be another event close to the Santonian-Campanian boundary, which was identified, at Tepeyac, in bed 30 (Ifrim et al., in press) and at the base of the Campanian. At Waxahachie, Texas, however, a small specimen of P. (P.) seppenradensis was found below the Santonian-Campanian boundary, i.e., 0.7 m below the LO of Marsupites testudinarius and the FO of Uintacrinus anglicus (Gale et al., 2008; but see discussion in Ifrim et al., in press). At Tepeyac, a gap exists between the LO of Parapuzosia (P.) leptophylla (loose specimen near bed 12) and the FO of P. (P.) seppenradensis in bed 30. The FO of this giant ammonite thus also seems to be a suitable secondary marker. However, this exceptional ammonite is abundant in the upper half of the Tepeyac section but rare elsewhere (Ifrim et al., in press); it thus does not meet general abundance as requirement for a reliable index ammonite.
The original definition of the base of the Campanian of de Grossouvre (1894) was the lowest level with Placenticeras bidorsatum. The FO of this ammonite provides a good appoximation for the Santonian-Campanian boundary level in central Europe, but the taxon has not been found elsewhere (Kennedy, 1986; Kennedy and Kaplan, 1995). In North America, Placenticeras syrtale [including the endemic subspecies of Chihuahua and adjacent Trans-Pecos-Texas described by Hyatt (1903) and Wolleben (1967)] occurs below and above the boundary in Chihuahua, Texas, Alabama, Mississippi, New Mexico, Colorado, Wyoming, Montana, and Utah. The taxon seems to be a good proxy to determine the boundary in clastic facies of the Western Interior, but not a single specimen of Placenticeras was found at Tepeyac, likely due to its absence in carbonate environments of the Gulf of Mexico, so other index species are needed there.
The ammonoid biozonations for the upper Santonian and lower Campanian stages in Texas and northeastern Mexico (Tepeyac) are summarized in Figure 40. Young (1963: 18, table 7) first presented a biozonation for the Santonian and Campanian of the Gulf Coast region (Figure 40). Based on data from central Texas to Tepeyac, the Menabites (Delawarella) tequesquitense zone represents the lowermost Campanian and underlies the Menabites (Delawarella) delawarensis zone. Subsequent work has shown that Menabites (Delawarella) delawarensis has a long range in the Gulf Coast region (Cobban and Kennedy, 1992a, 1992b). Therefore, the delawarensis zone was extended to encompass the entire interval between Young's (1963) zones of tequesquitense and marroti, with the latter renamed Baculites taylorensis zone by Kennedy et al. (1997a). The Texanites shiloensis zone was changed to the B. bevahensis zone by Young (1985), due to the wide geographical occurrence of this species from Texas to Madagascar. Although not explicitely stated, the authors appear to have used these zones in the sense of ranges, thus allowing them to modify their zonation as shown in Figure 40, but actually, these zones have never been defined formally.
Collignon (1948, p. 85) states that Bevahites bevahensis occurs at the boundary between the uppermost Santonian and the lower Campanian. This matches the occurrence of B. bevahensis in the lower tequesquitense zone at Tepeyac. Texanites shiloensis occurs lower in the section, notably below the FO of tequesquitense in Chihuahua (Wolleben, 1967), although it was only found in a single bed. Young (1959) suggested to use B. bevahensis as index species for the upper Santonian in Texas. Elsewhere, its occurrence is, however, not calibrated with other index species. A single specimen of Bevahites bevahensis was discovered in bed TPY31, i.e., from the lowermost Menabites (Delawarella) tequesquitense zone. We maintain the Plesiotexanites shiloensis zone of Young (1963) and Cobban and Kennedy (1992a, 1992b), which is documented by various specimens.
Given the numerous different zonations, the absence of a formal biozonal definition, and the fact that Tepeyac is the only section in the Americas where ammonoid zonation across the Santonian-Campanian boundary is calibrated by a stable carbon isotope curve (Ifrim et al., in press), we propose the following ammonoid zonation for this interval of the Upper Cretaceous in the northern and northwestern Gulf Coast (Figure 2):
Plesiotexanites shiloensis Interval Zone
The base of this zone is defined by the first occurrence (FO) of the index species. At Tepeyac, this species first occurs in bed 12, but layers below this level are little exposed and the fossil record is scarce. We suggest that the FO of Plesiotexanites shiloensis may even reach to below the base of the Tepeyac section. Species co-occurring with Plesiotexanites shiloensis are Parapuzosia (P.) leptophylla, Pseudoschloenbachia (P.) mexicana, and Texanites lonsdalei, whereas Menuites stephensoni has its FO in this zone.
Menabites (Delawarella) tequesquitense Interval Zone
The base of this zone is defined by the first occurrence (FO) of the index species in bed 25 at 10.7 m of the Tepeyac section. The zone is 5.3 m thick. Co-occurring species are Amapondella amapondense, Plesiotexantes shiloenis, Pseudoschloenbachia (Pseudoschloenbachia) mexicana, Menuites stephensoni, and Texanites lonsdalei. Bevahites bevahensis, Menabites (Delawarella) mariscalense, Menabites (Delawarella) vanuxemi, and Parapuzosia (Parapuzosia) seppenradensis have their FOs within this zone. The index is an endemic species. Even though, Menabites (Delawarella) tequesquitense is chosen as the index here because it occurs in several beds, whereas B. bevahensis, the other potential zonal marker, is scarce and only found in bed 33, at 14.5 m of the section.
Menabites (Delawarella) delawarensis Interval Zone
The base of this zone is defined by the first occurrence (FO) of the index species in bed 35 at 16.0 m of the section. The zone is 6.6 m thick. Amapondella amapondense, Bevahites costatus coahuilaensis, Menabites (Delawarella) tequesquitense, Menuites stephensoni, Plesiotexantes shiloenis, Texanites lonsdalei, and the index species have their last occurrences (LO) in this zone, while Parapuzosia (Parapuzosia) seppenradensis, Pseudoschloenbachia (Pseudoschloenbachia) mexicana, Menabites (Delawarella) mariscalense, and Menabites (Delawarella) vanuxemi range throughout the zone, and Glyptoxoceras texanum, Baculites haresi, Menabites (Bererella) walnutensis, Menabites (Delawarella) danei, Pachydiscus (P.) duelmensis, and Menuites sp. have their first occurrences (FO). This zone may correspond to a full range of the index fossil, but we suggest definition as an interval zone.
Baculites taylorensis Interval Zone
The base of this zone is defined by the first occurrence (FO) of the index species in bed 64, at 22.6 m of the section. The zone is 5.7 m thick at Tepeyac and cut off upsection by a disconformity. Co-occurring taxa are Glyptoxoceras texanum, Baculites haresi, M. (B.) walnutensis, Menabites (Bererella) walnutensis, Menabites (Delawarella) danei, Menabites (Delawarella) mariscalense, Menabites (Delawarella) uddeni, Menabites (Delawarella) vandaliense, Menabites (Delawarella) vanuxemi, Menuites sp., P achydiscus (P.) duelmensis, Parapuzosia (Parapuzosia) seppenradensis, Pseudoschloenbachia (Pseudoschloenbachia) mexicana and Scaphites (S.) hippocrepis III.
Other Occurrences
Submortoniceras uddeni was previously known only from a single specimen collected by L.W. Stephenson, in 1932, along the Chesapeake and Delaware Canal. Its occurrence was suggested to correlate with the lowest Campanian Submortoniceras tequesquitense zone, documented for Texas fide Kennedy et al. (1997b), based on a statement by Young (1963, p. 106), who nevertheless did not refer to this correlation explicitly. The view of Kennedy et al. (1997b) is also challenged here because the species is recorded, at Tepeyac, from beds yielding Scaphites (S.) hippocrepis III and above, and thus not from the lowest Campanian, nor co-occurring with Menabites (Delawarella) tequesquitense.
Sohl and Mello (in Owens et al., 1970, table 5) documented Submortoniceras uddeni and Menabites (Delawarella) delawarensis from spoil heaps along the Atlantic seaboard of New Jersey and Maryland. In figure 23 of the authors, the two species are interpreted as co-occurring in strata stratigraphically older than those containing S. hippocrepis III. At Tepeyac, however, they clearly co-occur with Scaphites (S.) hippocrepis III.
A single specimen of Menabites (Delawarella) vandaliense was recorded from the Blossom Sand, Texas. The unit was assigend to the upper Santonian by Kennedy et al. (2001), although Glyptoxoceras texanum, Placenticeras syrtale and Baculites cf. haresi oppose this view. These species do not reliably indicate a Santonian age, and correlation of the Blossom Sand to the upper Santonian therefore appears questionable. At Tepeyac, G. texanum and Baculites haresi first appear in the early Campanian, along with Menabites (Delawarella) vandaliense, while Placenticeras syrtale is absent. The Blossom Sand deposit may therefore be early Campanian in age. The occurrence of Young’s (1963) high-whorled Submortoniceras in the Blossom Sand deposit would then be coeval to the upper part of the lower Campanian at Tepeyac, where these morphotypes also occur in large numbers.
PALAEOBIOGEOGRAPHIC RELATIONS
Upper Santonian and lower Campanian Placenticeras syrtale and Placenticeras placenta are absent at Tepeyac. These widely distributed species developed lower Campanian endemic subspecies in the Ojinaga region of Chihuahua, about 300 km west of Tepeyac, an area previously identified as an endemic center for bivalves (Kauffman, 1973). Restriction of Placenticeras to siliciclastic facies, as prevailing in Chihuahua and western and central Texas (Hyatt, 1903; Young, 1963; Wolleben, 1967; Kennedy et al., 2001; Kennedy et al., 2004) and the northern Gulf Coast (Kennedy et al., 1997a), has already been suggested. High-whorled morphotypes, such as Placenticeras, are frequently interpreted to have adapted to wave movements and shallow water (Jacobs and Chamberlain, 1996; Ifrim and Stinnesbeck, 2010). The absence of Placenticeras in the Tepeyac limestones may therefore reflect palaeoenvironmental factors.
The “high-whorled” oxycone and smooth Menabites (Delawarella) sancarlosense (Young, 1963) from the upper Santonian of western Texas and Ojinaga, Chihuahua, and its potential descendents M. (D.) candelariae (Young, 1963), M. (D.) mariscalense (Young, 1963) and M. (D.) uddeni (Young, 1963) from the lower Campanian of the Big Bend area, Texas, but also the coarsely ornamented M. (D.) densinodosus (Young, 1963) from the Lower Campanian of the northern Gulf area of Texas, seem to be endemic species of the subgenus Delawarella with occurrences in a restricted area reaching from southern Texas to eastern Chihuahua. This interpretation of an early Campanian endemic event is supported by the absence of coarsely ornamented Menabites (Australiella) (Ifrim et al., 2013) and of M. (D.) sabinalensis, an index species for a upper part of the lower Campanian which may also be endemic to Texas. Texanites candelariae, originally documented from the Big Bend area of southern Texas, and Submortonicers punctatum from the Atlantic seaboard of New Jersey and Maryland, are further endemics to the region of their first description. There was little geographical interchange between texaniids of the region during the late Santonian and early Campanian.
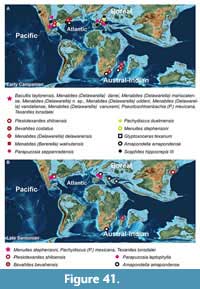 On the other hand, there is also evidence for faunal connections between Tepeyac and other palaeobiogeographic provinces, as summarized in Figure 41. The genus Menuites evolved in high southern latitudes (see dicussion in Ifrim et al., 2013). An early Campanian Menuites from western Coahuila was interpreted by Ifrim et al. (2013) to be the earliest representative in North America, but some specimens from Tepeyac are even older. This includes late Santonian occurrences of Menuites stephensoni, a species previously known only from the early Campanian of southern North America and Central Europe, but also Parapuzosia (Parapuzosia) seppenradensis, which is here documented from the basal Campanian on and thus earlier than in Europe (Ifrim et al., in press). Faunal exchange must have existed with Europe, although these connections have been little evident to date. Parapuzosia (P.) leptophylla is known from the lower Santonian of South France, the Santonian of Austria, and the upper Santonian of England. Its appearance in the northwestern Gulf of Mexico is now dated to the late Santonian. Although frequent at Tepeyac, the species has never been recorded in the Chihuahua area, at only a few hundred kilometres to the west along the same broad shelf. Parapuzosia (P.) leptophylla evolved into the even larger species Parapuzosia (P.) seppenradensis at the end of the Santonian, a species now known to be present on both sides of the North Atlantic (Ifrim et al., in press). This latter gigantic species may have been mobile enough to cross the North Atlantic into the European Basin. Nevertheless, there are also other species recorded from both sides of the North Atlantic, such as Amapondella amanpondense, Menuites stephensoni, and Pachydiscus (P.) duelmensis. These taxa are known from the Lower Campanian of Germany and Tepeyac. Glyptoxoceras texanum occurs in Texas and adjacent areas in the late Santonian but was also found at the Atlanic coast of New Jersey in the early Campanian. It is a close relative of Neoglyptoxoceras retrorsum (Schlüter, 1972) from the lower Campanian of Germany. However, N. retrorsum and its synonym N. novimexicanus (Reeside, 1927) occur in Germany (Kaplan et al., 2005), Austria (Kennedy and Summesberger, 1984; Summesberger et al., 2017), Poland (Błaszkiewicz, 1980; Machalski et al., 2004), the Crimea (Arkadiev et al., 2000), Belgium (Kennedy, 1986; Kennedy, 1993), France (Kennedy, 1986), but also the US Western Interior (Reeside, 1927). Glyptoxoceras texanum appears at Tepeyac shortly after Pachydiscus (P.) duelmensis, i.e., during the lower Campanium, which further supports the interpretation of a temporal connection to Europe. After this phase, FOs at Tepeyac are from endemic and North American species, which indicates that the connection to Europe was again lost.
On the other hand, there is also evidence for faunal connections between Tepeyac and other palaeobiogeographic provinces, as summarized in Figure 41. The genus Menuites evolved in high southern latitudes (see dicussion in Ifrim et al., 2013). An early Campanian Menuites from western Coahuila was interpreted by Ifrim et al. (2013) to be the earliest representative in North America, but some specimens from Tepeyac are even older. This includes late Santonian occurrences of Menuites stephensoni, a species previously known only from the early Campanian of southern North America and Central Europe, but also Parapuzosia (Parapuzosia) seppenradensis, which is here documented from the basal Campanian on and thus earlier than in Europe (Ifrim et al., in press). Faunal exchange must have existed with Europe, although these connections have been little evident to date. Parapuzosia (P.) leptophylla is known from the lower Santonian of South France, the Santonian of Austria, and the upper Santonian of England. Its appearance in the northwestern Gulf of Mexico is now dated to the late Santonian. Although frequent at Tepeyac, the species has never been recorded in the Chihuahua area, at only a few hundred kilometres to the west along the same broad shelf. Parapuzosia (P.) leptophylla evolved into the even larger species Parapuzosia (P.) seppenradensis at the end of the Santonian, a species now known to be present on both sides of the North Atlantic (Ifrim et al., in press). This latter gigantic species may have been mobile enough to cross the North Atlantic into the European Basin. Nevertheless, there are also other species recorded from both sides of the North Atlantic, such as Amapondella amanpondense, Menuites stephensoni, and Pachydiscus (P.) duelmensis. These taxa are known from the Lower Campanian of Germany and Tepeyac. Glyptoxoceras texanum occurs in Texas and adjacent areas in the late Santonian but was also found at the Atlanic coast of New Jersey in the early Campanian. It is a close relative of Neoglyptoxoceras retrorsum (Schlüter, 1972) from the lower Campanian of Germany. However, N. retrorsum and its synonym N. novimexicanus (Reeside, 1927) occur in Germany (Kaplan et al., 2005), Austria (Kennedy and Summesberger, 1984; Summesberger et al., 2017), Poland (Błaszkiewicz, 1980; Machalski et al., 2004), the Crimea (Arkadiev et al., 2000), Belgium (Kennedy, 1986; Kennedy, 1993), France (Kennedy, 1986), but also the US Western Interior (Reeside, 1927). Glyptoxoceras texanum appears at Tepeyac shortly after Pachydiscus (P.) duelmensis, i.e., during the lower Campanium, which further supports the interpretation of a temporal connection to Europe. After this phase, FOs at Tepeyac are from endemic and North American species, which indicates that the connection to Europe was again lost.
Other faunal connections existed with the Southern hemisphere, and in particular with South Africa and Madagascar. This is indicated by the genus Pseudoschloenbachia, known from endemic upper Santonian and lower Campanian species of Texas, Coahuila, and Chihuahua, but also from a variety of lower Campanian species from Madagascar (Collignon, 1969).
The two Submortoniceras - Delawarella lineages of South Africa - Madagascar on one side, and the Gulf of Mexico on the other, are closely connected. Interchange between the two is seen in the late Santonian, based on the close relation between the Texanites soutoni - Submortoniceras woodsi lineage and the emergence of Menabites (Delawarella) tequesquitense. Amapondella amanpondense, Bevahites costatus, and Menuites stephensoni first appear in these levels in both Tepeyac and Madagascar, and possibly Bevahites bevahensis in Texas. During the early Campanian, Menabites (Delawarella) delawarensis dispersed to Europe and is documented for this interval from France and Spain. Menabites (Bererella) walnutensis (Young, 1963), present in the Gulf of Mexico region including Tepeyac, and along the North American Atlantic coast, also has equivalents in the lower Campanian of Madagascar (Collignon, 1969). The subgenus may there even be more diverse than in Texas, New Jersey (Kennedy et al., 1997b), and Coahuila.
Amanpondiella amanpondense is another widespread taxon known from the Santonian of Israel (Lewy, 1983), France (Kennedy et al., 1995), South Africa (Klinger and Kennedy, 2003) and Madagascar (Klinger et al., 2007), and the Campanian of Germany, and now Coahuila.
Scaphites (S.) hippocrepis III is even wider distributed. The taxon evolved in the Western Interior but dispersed into the northern Gulf (Kennedy et al., 1997a) and to the Atlantic Coast of New Jersey (Kennedy et al., 1997b). It crossed to Europe and is documented from Spain (Küchler, 2000), France (Kennedy, 1986), Poland, Germany, the Netherlands (Jagt et al., 2004; Kaplan et al., 2005), and Sweden (Kennedy and Christensen, 1997). Beyond Europe, the species occurs in late early Campanian sediments of Tunisia (Pervinquière, 1907) and the North American Pacific coast (Anderson, 1958).
The Tepeyac faunal assemblage thus provides evidence for repeated and increasing faunal exchange with Europe and southern latitudes, but also includes intervals of faunal cut-off and endemic evolution. It is difficult to date to explain these unusual faunal connections of the Gulf of Mexico ammonite assemblage with Europe, South Africa, and even Madagascar, but we suggest that these may have been triggered by geotectonic changes during the Santonian-Campanian stages. During this period of the Late Cretaceous, the Cretaceous Supergreenhouse ended and was followed by a long-term gradual cooling. This cooling trend has been interpreted to be a consequence of changes in the global ocean circulation system due to the opening of the Equatorial Atlantic Gateway in the late Santonian to early Campanian (Wagner and Pletsch, 1999; Pletsch et al., 2001). The sediments exposed at Tepeyac formed during this phase, and the palaeobiogeographic analysis reflects this palaeoceanographic event. Faunal exchange with other provinces, especially with the Southern Ocean, i.e., Madagascar and Antarctica, increased considerably during this phase. Faunal exchange across the Atlantic was also intensified. It is likely that the final deep ocean opening of the Atlantic Gateway between Brazil and North Africa allowed for the connection to the South and immigration of southern hemisphere taxa during the late Santonian. Intensified oceanic circulation may then have improved the connection with Europe during the early Campanian. Intercontinental faunal exchange in the late Santonian - early Campanian was thus significantly more intense than hitherto known (compare Ifrim et al., 2015a).
CONCLUSIONS
Twenty-four species of ammonites assigned to 12 genera are described from Tepeyac near Jiménez, northern Coahuila, north-eastern Mexico. The faunal assemblage is late Santonian-early Campanian (Upper Cretaceous) in age and allows for detailed biozonation for Gulf coast ammonoids across the stage boundary. Outstanding outcrop conditions, the abundance and diversity of ammonites in the new section, and their biozonal completeness distinguish Tepeyac as a potential reference section for the Santonian-Campanian boundary in the Americas.
Three phases of faunal interchange are identified in the Tepeyac section: (1) In the late Santonian, with the FO of M. (D.) tequesquitense along with other species or genera from the Southern Ocean. (2) In the early Campanian. FO of M. (D.) delawarensis. Regional to endemic lineages evolved from immigrating ammonoids, thus indicating a subsequent phase of cut-off. (3) The dispersal of Scaphites (S.) hippocrepis III ended the endemic phase during the late early Campanian and led to a phase of increasing dispersal of species and better connectivity between provinces.
Faunal connections between Tepeyac and the Southern hemisphere (e.g., South Africa, Madagascar) may have resulted from the opening of the Atlantic Gateway during the late Santonian, while early Campanian exchange with Europe is interpreted as related to intensified oceanic circulation, another consequence of rapid opening of the Atlantic.
Placenticeras, a genus frequently used as a proxy to determine the Santonian-Campanian boundary, is absent at Tepeyac, but this is explained as facies-controlled. This genus appears to have preferred clastic depositional environments and avoided carbonate-dominated environments, as present in Tepeyac.
ACKNOWLEDGEMENTS
Financial support of this research was provided by the DFG (IF61/11-1, STI128/40-1) and is gratefully acknowledged. We are grateful to J. González (Mexico City) and E. Acévez (Playa del Carmen) for assistance in outcrop documentation, to J.M. Bedmar and M.A. Argaez Martínez (both Universidad Nacional Autónoma de México, Mexico City) for technical support and company in the field, to J. Madrazo (Saltillo, Coahuila), C. Wollny and N. Schorndorf (both Heidelberg University) for assistance during fieldwork, and J.M. Padilla Gutiérrez for access to the CPC collection in the Museo del Desierto (Saltillo, Coah.). H. Fuentes (Piedras Negras) provided logistic support in the Piedras Negras area, J.J. López (Tepeyac) gave access to his property, A.H. González González and J.M. Padilla Gutiérrez (Museo del Desierto, Saltillo, México) are acknowledged for logistic support in the field and access to CPC collections, while U. Menkveld (Naturhistorisches Museum Bern, Switzerland) provided information on the Collection Böse-Straub, i.e., the originals of Renz. The palaeogeographic map in Figure 41 is courtesy of the Paleomap project and was kindly provided by C. Scotese.We also thank the anonymous reviewers and handling editor R. Bicknell.
REFERENCES
Adkins, W.S. 1928. Handbook of Cretaceous fossils. University of Texas Bulletin, 2838:1-303.
Adkins, W.S. 1929. Some Upper Taylor ammonites from Texas. University of Texas Bulletin, 2901:203-211.
Adkins, W.S. 1932. Mesozoic systems in Texas. The Geology of Texas: vol 1, Stratigraphy. University of Texas Bulletin, 3232:239-518.
Anderson, F.M. 1958. Upper Cretaceous of the Pacific Coast. Geological Society of America, Memoir 71:1-378. https://doi.org/10.1130/MEM71
Arkadiev, V.V., Atabekian, A.A., Baraboshkin, E.Y., and Bogdanova, T.N. 2000. Stratigraphy and ammonites of Cretaceous deposits of South-West Crimea. Palaeontographica, A255:85-128.
Becker, M.A., Slattery, W., and Chamberlain Jr., J.A. 1998. Mixing of Santonian and Campanian chondrichthyan and ammonite macrofossils along a transgressive lag deposit, Greene County, western Alabama. Southeastern Geology, 37:205-216.
Besaire, H. 1930. Recherches géologiques à Madagascar. Contributions à l'étude des ressources minérales. Imprimerie H. Basuyau & Cie, Toulouse.
Birkelund, T. 1965. Ammonites from the Upper Cretaceous of West Greenland. Meddeleser om Grønland, 179:1-192.
Błaszkiewicz, A. 1980. Campanian and Maastrichtian ammonites of the Middle Vistula Valley, Poland: a stratigraphic-paleontological study. Prace Instytutu Geologicznego, 92:1-63.
Böse, E. 1927. Cretaceous ammonites from Texas and northern Mexico. University of Texas Bulletin, 2748:143-357.
Böse, E. and O.A. Cavins. 1927. The Cretaceous and Tertiary of southern Texas and northern Mexico. University of Texas Bulletin, 2748:7-142.
Cobban, W.A. 1958. Two new species of Baculites fom the Western Interior region. Journal of Paleontology, 32:660-665.
Cobban, W.A. 1969. The Late Cretaceous ammonites Scaphites leei Reeside and Scaphites hippocrepis (DeKay) in the Western Interior of the United States. United States Geological Survey Professional Paper, 619:1-17.
Cobban, W.A. and Kennedy, W.J. 1992a. Campanian ammonites from the Upper Cretaceous Gober Chalk of Lamar County, Texas. Journal of Paleontology,66:440-454.
Cobban, W.A. and Kennedy, W.J. 1992b. Campanian Trachyscaphites spiniger ammonite fauna in north-east Texas. Palaeontology, 35:63-93.
Cobban, W.A. and Kennedy, W.J. 1994. Middle Campanian (Upper Cretaceous) ammonites from the Pecan Gap Chalk of central and northeastern Texas. United States Geological Survey Bulletin, 2073:D1-D9.
Collignon, M. 1938. Ammonites Campaniennes et Maastrichtiennes de l'ouest et du sud de Madagascar. Annales Géologiques du Service des Mines de Madagascar, 9:55-118.
Collignon, M. 1948. Ammonites néocrétacées du Ménabe (Madagascar). I. Les Texanitidae. Annales Geologiques du Service des Mines, Madagascar, 13:47-107.
Collignon, M. 1969. Atlas des fossiles caractéristiques de Madagascar (Ammonites). Fascicule 15 (Campanien inférieur). Service Géologique, Tananarive.
de Grossouvre, A. 1894. Recherches sur la Craie Supérieure, 2, Paléontologie. Les ammonites de la Craie Supérieure. Imprimerie Nationale, Paris.
DeKay, J.E. 1828. Report on several fossil multilocular shells from the state of Delaware: wih observations on a second specimen of the new fossil genus Eurypterus. Annales of the Lyceum of Natural History, 2:273-279.
de Lamarck, J.B.P.A.M. 1799. Prodrome d’une nouvelle classification des coquilles. Mémoirs du Musée de l'Histoire Naturelle. 1:63-90.
Gale, A., Montgomery, P., Kennedy, W., Hancock, J., Burnett, J., and McArthur, J. 1995. Definition and global correlation of the Santonian-Campanian boundary. Terra Nova, 7:611-622.
Gale, A.S., Hancock, J.M., Kennedy, W.J., Petrizzo, M.R., Lees, J.A., Walaszczyk, I., and Wray, D.S. 2008. An integrated study (geochemistry, stable oxygen and carbon isotopes, nannofossils, planktonic foraminifera, inoceramid bivalves, ammonites and crinoids) of the Waxahachie Dam Spillway section, north Texas: a possible boundary stratotype for the base of the Campanian stage. Cretaceous Research, 29:131-167.
https://doi.org/10.1016/j.cretres.2007.04.006
Gill, T. 1871. Arrangement of the families of mollusks. Smithsonian Institution, Washington D.C., U.S.A.
Hancock, J.M. 1993. Sea-level changes around the Cenomanian-Turonian boundary. Cretaceous Research, 14:553-562. https://doi.org/10.1006/cres.1993.1039
Hyatt, A. 1889. Genesis of the Arietidae, Smithsonian Contributions to Knowledge, 673:502-592.
Hyatt, A. 1903. Pseudoceratites of the Cretaceous. Government Printing Office, Washington.
Ifrim, C. and Stinnesbeck, W. 2010. Migration pathways of the late Campanian and Maastrichtian shallow facies ammonite Sphenodiscus in North America. Palaeogeography, Palaeoclimatology, Palaeoecology, 292:96-102. https://doi.org/10.1016/j.palaeo.2010.03.030
Ifrim, C., Stinnesbeck, W.,and Ventura, J.F. 2013. An endemic cephalopod assemblage from the lower Campanian (Late Cretaceous) Parras Shale, western Coahuila, Mexico. Journal of Paleontology, 87:881-901. https://doi.org/10.1666/12-123
Ifrim, C., Lehmann, J., and Ward, P.D. 2015a. Paleobiogeography of Late Cretaceous Ammonoids, p. 259-274. In Klug, C., Korn, D., De Baets, K., Kruta, I., and Mapes, R.H. (eds.), Ammonoid Paleobiology - From Macroevolution to Paleogeography.Volume 44. Springer, Heidelberg. https://doi.org/10.1007/978-94-017-9633-0_10
Ifrim, C., Stinnesbeck, W., Ventura, J.F., and Espinosa, B. 2015b. Upper Campanian (Upper Cretaceous) cephalopods from the Parras Shale at Saucedas, Coahuila, Mexico. Journal of South American Earth Sciences, 64:229-257. https://doi.org/10.1016/j.jsames.2015.07.013
Ifrim, C., Bengtson, P., and Schweigert, G. 2018. Growth and function of spines in Jurassic and Cretaceous ammonites. Cretaceous Research, 88:62-78. https://doi.org/10.1016/j.cretres.2017.05.003
Ifrim, C., Gale, A.S., Stinnesbeck, W., Schorndorf, N., and González González, A.H. In press. Ontogeny, evolution and palaeobiogeographic distribution of Parapuzosia (P.) seppenradensis, the world’s largest ammonite. PlosOne.
Jacobs, D.K. and Chamberlain, J.A. 1996. Buoyancy and hydrodynamics in ammonoids, p. 169-224. In Landman, N., Tanabe, K., and Davis, R.A. (eds.), Ammonoid Paleobiology. Volume 13. Plenum Press, New York. https://doi.org/10.1007/978-1-4757-9153-2_7
Jagt, J.W. 1989. Ammonites from the early Campanian Vaals Formation at the CPL Quarry (Haccourt, Liege, Belgium) and their stratigraphic implications. Mededelingen Rijks Geologische Dienst, 43:1-33.
Jagt, J.W.M. 2002. Late Cretaceous ammonite faunas of the Maastrichtian type area, p. 509-522. In Summesberger, H., Histon, K., and Daurer, A. (eds.), Cephalopods - Present and Past. Geologische Bundesanstalt, Wien.
Jagt, J.W.M., Burnett, J.A., and Kennedy, W.J. 1995. Upper Campanian ammonites and nannofossils from South Limburg. Mededelingen Rijks Geologische Dienst, 53:49-63.
Jagt, J.M.W., Walaszczyk, I., Yazykova, E.A., and Zaton, M. 2004. Linking southern Poland and northern Germany: Campanian cephalopods, inoceramid bivalves and echinoids. Acta Geologica Polonica, 54:573-586.
Kaplan, U., Kennedy W.J., and Hiß, M. 2005. Stratigraphie und Ammonitenfaunen des Campan im nordwestlichen und zentralen Münsterland. Geologie und Paläontologie in Westfalen, 64:1-171.
Kaplan, U., Kennedy, W.J., and Scheer, U. 2006. Ammoniten der Bottrop-Formation, Campanium, westliches Münsterland. Geologie und Paläontologie in Westfalen, 67:1-71.
Kauffman, E.G. 1973. Cretaceous Bivalvia, p. 353-383. In Hallam, A. (ed.), Atlas of Paleobiogeography. Elsevier, Amsterdam.
Kennedy, J. and Cobban, W.A. 1991. Upper Cretaceous (upper Santonian) Boehmoceras fauna from the Gulf Coast region of the United States. Geological Magazine, 128:167-189.
Kennedy, W. and Cobban, W. 1993a. Lower Campanian (Upper Cretaceous) ammonites from the Merchantville Formation of New Jersey, Maryland, and Delaware. Journal of Paleontology, 67:828-849.
Kennedy, W.J. 1986. Campanian - Maastrichtian ammonites from northern Aquitaine, France. Special Papers in Palaeontology, 36:1-145.
Kennedy, W.J. 1993. Campanian and Maastrichtian ammonites from the Mons Basin and adjacent areas (Belgium). Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, 63:99-131.
Kennedy, W.J. 2019. The Ammonoidea of the Upper Chalk. Part 1. Monograph of the Palaeontographical Society, London, 173:1-207. https://doi.org/10.1080/02693445.2019.1642002
Kennedy, W.J. and Summesberger, H. 1984. Upper Campanian Ammonites from the Gschliefgraben (Ultrahelvetic, Upper Austria). Beiträge zur Paläontologie von Österreich, 11:149-206.
Kennedy, W.J. and Cobban, W.A. 1993b. Upper Campanian ammonites from the Ozan-Annona Formation boundary in Southwestern Arkansas. Bulletin of the Geological Society of Denmark, 40:115-148.
Kennedy, W.J. and Cobban, W.A. 1993c. Campanian ammonites from the Annona Chalk near Yancy, Arkansas. Journal of Paleontology, 67:83-97.
Kennedy, W.J. and Kaplan, U. 1995. Parapuzosia (Parapuzosia) seppenradensis (Landois) und die Ammonitenfauna der Dülmener Schichten, unteres Unter-Campan, Westfalen. Geologie und Paläontologie in Westfalen, 33:1-127.
Kennedy, W.J. and Christensen, W.K. 1997. Santonian to Maastrichtian ammonites from Scania, southern Sweden. Fossils and Strata, 44:75-128.
Kennedy, W.J. and Cobban, W.A. 1999. Campanian (Late Cretaceous) ammonites from the Bergstrom Formation in central Texas. Acta Geologica Polonica, 49:67-80.
Kennedy, W.J. and Cobban, W.A. 2001. Campanian (Late Cretaceous) ammonites from the upper part of the Anacacho Limestone in South-Central Texas. Acta Geologica Polonica, 51:15-30.
Kennedy, W.J., Bilotte, M., and Melchior, P. 1995. Ammonite faunas, biostratigraphy and sequence stratigraphy of the Coniacian-Santonian of the Corbières (NE Pyrénées). Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, 19:377-499.
Kennedy, W.J., Cobban, W.A., and Landman, N.H. 1997a. Campanian ammonites from the Tombigbee Sand Member of the Eutaw Formation, the Mooreville Formation, and the basal part of the Demopolis Formation in Mississippi and Alabama. American Museum Novitates, 3201:1-44.
Kennedy, W.J., Landman, N.H., and Cobban, W.A. 2001. Santonian ammonites from the Blossom Sand in Northeast Texas. American Museum Novitates, 3332:1-9.
Kennedy, W.J., Cobban, W.A., Landman, N.H., and Johnson, R. 1997b. New ammonoid records from the Merchantville Formation (Upper Cretaceous) of Maryland and New Jersey. American Museum Novitates, 3193:1-17.
Kennedy, W.J., Hancock, J.M., Cobban, W.A., and Landman, N.H. 2004. A revision of the ammonite types described in F. Roemer's ‘Die Kreidebildungen von Texas und ihre organischen Einschlüsse’ (1852). Acta Geologica Polonica, 54:433-445.
Klinger, H. 1985. Upper Cretaceous Cephalopoda from offshore deposits of the Natal South Coast, South Africa. Palaeoontologia Africana, 26:1-12.
Klinger, H.C. and Kennedy, W.J. 1980. Cretaceous faunas from Zululand and Natal, South Africa. The ammonite subfamily Texanitinae Collignon, 1948. Annals of the South African Museum, 80:1-357.
Klinger, H.C. and Kennedy, W.J. 1997. On the affinities of Madagascaites andimakensis Collignon, 1966, and allied Upper Cretaceous heteromorph ammonites. Annals of the South African Museum, 105:227-247.
Klinger, H.C. and Kennedy, W.J. 2001. Stratigraphic and geographic distribution, phylogenetic trends and general comments on the ammonite family Baculitidae Gill, 1871 (with and annotated list of species referred to the family). Annals of the South African Museum, 107:1-290.
Klinger, H.C. and Kennedy, W.J. 2003. Cretaceous faunas from Zululand and Natal, South Africa. The ammonite families Nostoceratidae Hyatt, 1894 and Diplomoceratidae Spath, 1926. Annals of the South African Museum, 110:219-334.
Klinger, H.C., Kennedy, W.J., and Grulke, W.E. 2007. New and little-known Nostoceratidae and Diplomoceratidae (Cephalopoda: Ammonoidea) from Madagascar. African Natural History, 3:89-115.
Köplitz, W. 1920. Über die Fauna des oberen Untersenon im Seppenrade-Dülmener Höhenzuge. Dissertation, Westfälische Wilhelms-Universität, Münster..
Küchler, T. 2000. Scaphites hippocrepis (Dekay) IV, a new chronological subspecies from the Lower - Upper Campanian (Upper Cretaceous) boundary interval of northern Spain. Acta Geologica Polonica, 50:161-167.
Landois, H. 1895. Die Riesenammoniten von Seppenrade: Pachydiscus ZITTEL Seppenradensis H. LANDOIS. Jahresbericht des Westfälischen Provinzialvereins für Wissenschaft und Kunst, 23:99-108.
Larson, N.L., Jorgensen, S.D., Farrar, R.A., and Larson, P.L. 1997. Ammonites and the Other Cephalopods of the Pierre Seaway: An Identification Guide. Geoscience Press Inc., Tuscon, Arizona, U.S.A.
Lewy, Z. 1983. A well-preserved Upper Santonian heteromorph ammonite from Israel. Geological Survey of Israel, Current Research, 1982:24-27.
Machalski, M., Kennedy, W.J., and Kin, A. 2004. Early Late Campanian ammonite fauna from Busko Zdrój (Nida Trough, southern Poland). Acta Geologica Polonica, 54:447-471.
Matsumoto, T. 1938. A biostratigraphic study on the Cretaceous deposits of the Naibuti Valley, South Karahuto. Proceedings of the Imperial Academy Japan, 14:190-194.
Matsumoto, T. 1970. A monograph of the Collignoniceratidae from Hokkaido, Part IV. Memoirs of the Faculty of Science, Kyushu University, Series D, Geology, 20:225-304.
Michailov, N.P. 1951. Upper Cretaceous ammonites from the southern part of the USSR and their importance for zonal stratigraphy (Campanian, Maastrichtian). Trudy Instituta Geologicheskikh Akademia Nauk SSSR, 129, Geology Series 50.
Morton, S.G. 1830. Synopsis of the organic remains of the ferruginous sand formation of the United States, with geological remarks. American Journal of Science, 18:243-250.
Morton, S.G. 1834. Synopsis of the organic remains of the Cretaceous Group of the United States. Key & Biddle, Philadelphia.
Musée de Paléontologie de Provence. 2010. Parapuzosia leptophylla (Sharpe), Galerie de fossiles. Musée de Paléontologie de Provence. http://mdp.cerege.fr/galerie.php?page=ammogeante#pas, accessed August 2021.
Nowak, J. 1913. Untersuchungen über die Cephalopoden der oberen Kreide in Polen III. Teil. Bulletin international de l’Académie des Sciences de Cracovie, Classe des Sciences mathématiques et naturelles, Série B, Sciences naturelles, 1913:335-415.
Owens, J.P., Minard, J., Sohl, N., and Mello, J. 1970. Stratigraphy of the outcropping post-Magothy Upper Cretaceous formations in southern New Jersey and northern Delmarva Peninsula, Delaware, and Maryland. United States Geological Survey Professional Paper, 674:1-60.
Parkinson, J. 1811. The Fossil Starfish, Echini Shells, Insects, Amphibia, Mammalia & c. The Organic Remains of a Former World, vol. 3. Sherwood, Neely and Jones, London.
Pervinquière, L. 1907. Études de paléontologie tunisienne 1: Céphalopodes des terrains sécondaires. De Rudeval, Paris.
Pletsch, T., Erbacher, J., Holbourn, A.E.L., Kuhnt, W., Moullade, M., Oboh-Ikuenobede, F.E., Söding, E., and Wagner, T. 2001. Cretaceous separation of Africa and South America: the view from the West African margin (ODP Leg 159). Journal of South American Earth Sciences, 14:147-174.
Reeside, J.B. 1927. The cephalopods of the Eagle Sandstone and related formations in the Western Interior of the United States. United States Geological Survey Professional Paper, 151:1-87.
Reeside, J.B. 1962. The Cretaceous ammonites of New Jersey. New Jersey Geological Survey Bulletin, 61:113-137.
Renz, H.H. 1936. Neue Cephalopoden aus der oberen Kreide von Rio Grande del Norte (Mexico und Texas). Abhandlungen der Schweizerischen Paläontoloogischen Gesellschaft, 57:1-16.
Richards, H.G. 1953. Record of the rocks - the geological story of Eastern North America. Ronald Press Company, New York..
Schlüter, C. 1872. Cephalopoden der oberen deutschen Kreide I. Palaeontographica, 21:1-120.
Scotese, C.R. 2021. Paleomap project. http://scotese.com
Servicio Geológico de México. 2008. Carta Geológica Minera 1: 250 000, H14-7 and H14-10, México D.F., México.
Sharpe, D. 1853-1857. Description of the fossil remains of Mollusca found in the Chalk of England. I, Cephalopoda. Monograph of the Palaeontographical Society, London:1-68.
Shumard, B.F. 1861. Descriptions of new Cretaceous fossils from Texas. Boston Society of Natural History Proceedings, 8:188-205.
Spath, L.F. 1921. On Cretaceous Cephalopoda from Zululand. Annals of the South African Museum, 12:217-321.
Spath, L.F. 1922. On the Senonian ammonite fauna of Pondoland. Transactions of the Royal Society of South Africa, 10:113-148.
Spath, L.F. 1925. On Senonian Ammonoidea from Jamaica. Geological Magazine, 62:28-32.
Spath, L.F. 1926. On new ammonites from the English Chalk. Geological Magazine, 3:77-83.
Spath, L.F. 1932. A Monograph of the Ammonoidea of the Gault, part 9. Palaeontographical Society, 84:379-410.
Summesberger, H. 1978. Eine obersantone Ammonitenfauna aus dem Becken von Gosau (Oberösterreich). Annalen des Naturhistorischen Museums in Wien:109-176.
Summesberger, H., Kennedy, W.J., and Skoumal, P. 2017. Early and middle Santonian Cephalopods from the Gosau Group (Upper Cretaceous, Austria) 2. Heteromorph Ammonoidea. Abhandlungen der Geologischen Bundesanstalt, 71:101-149.
van Hoepen, E.C.N.v. 1921. Cretaceous Cephalopoda from Pondoland. Annals of the Transvaal Museum, 8:1-48.
von Zittel, K.A. 1884. Handbuch der Palæontologie 1. Abtheilung. Palæozoologie 2. Band. Mollusca und Arthropoda. R. Oldenbourg, München, 2nd edition.
von Zittel, K.A. 1895. Grundzüge der Paläontologie. R. Oldenbourg, München, Germany.
Wagner, T. and Pletsch, T. 1999. Tectono-sedimentary controls on Cretaceous black shale deposition along the opening Equatorial Atlantic Gateway (ODP Leg 159). Geological Society, London, Special Publications, 153:241-265.
Whitfield, R.P. 1892. Gastropods and Cephalopoda of the Raritan Clays and Greensand Marls of New Jersey. United States Geological Society Monograph, 18:1-402.
Wiese, F., Brüning, J., and Otto, A. 1996. First record of Libycoceras ismaelis (Zittel, 1885) (Cretaceous Ammonoidea) in Europe (Campanian of the Santander area, Cantabria, northern Spain). Acta Geologica Polonica, 46:105-116.
Wilmsen, M. and Mosavinia, A. 2011. Phenotypic plasticity and taxonomy of Schloenbachia varians (J. Sowerby, 1817) (Cretaceous Ammonoidea). Paläontologische Zeitschrift, 85:169-184. https://doi.org/10.1007/s12542-010-0086-5
Wolleben, J.A. 1967. Senonian (Cretaceous) mollusca from Trans-Pecos Texas and northeastern Chihuahua, Mexico. Journal of Paleontology, 41:1150-1165.
Wolleben, J.A. 1977. Paleontology of the Difunta Group (Upper Cretaceous-Tertiary) in northeastern Mexico. Journal of Paleontology, 51:373-398.
Wright, C. and Kennedy, W. 1978. The ammonite Stoliczkaia from the Cenomanian of England and northern France. Palaeontology, 21:393-409.
Wright, C.W. 1952. A classification of the Cretaceous ammonites. Journal of Paleontology, 26:213-222.
Wright, C.W. and Wright, E.V. 1951. A survey of the fossil Cephalopoda of the Chalk of Great Britain. primarily a nomenclatorial Revision of Daniel Sharpe's monograph (1853-57). Monograph of the Palaeontographical Society, London, 453/457:1-148.
Wright, C.W. and Kennedy, J. 1984-1996. The ammonoidea of the Lower Chalk. Monograph of the Palaeontographical Society, London:pp. 1-126, pl. 121-140 (1984); 1127-1218, pl. 1941-1955 (1987); 1219-1294, pl. 1956-1986 (1991); 1295-1319, pl. 1987-1994 (1995); 1320-1403, pl. 1995-1124 (1996); 1404-1459, pl. 1125-1145.
Wright, C.W., Calloman [sic], J.H., and Howarth, M.K. 1996. Cretaceous Ammonoidea, Treatise on Invertebrate Paleontology. Volume Part L, Mollusca 4, revised. University of Kansas, Lawrence and Geological Society of America, Boulder.
Young, K. 1959. Techniques of mollusc zonation in Texas Cretaceous. American Journal of Science, 257:752-769.
Young, K. 1963. Upper Cretaceous ammonites from the Gulf Coast of the United States. University of Texas Bulletin, 6304:1-373.
Young, K. 1985. The Austin division of central Texas, p. 3-52. In Young, K. and Woodruff, C.M. (eds.), Austin Chalk in its type area; Stratigraphy and Structure. Volume 7, Austin Geological Society.
Young, K. and Marks, E. 1952. Zonation of Upper Cretaceous Austin chalk and Burditt marl, Williamson County, Texas. American Association of Petroleum Geologists, Bulletin, 36:477-488.
