

 A diverse Campanian biota from the Bozeș Formation (Petrești, Romania): Insights into the paleontology and paleoecology of a transitional sequence
A diverse Campanian biota from the Bozeș Formation (Petrești, Romania): Insights into the paleontology and paleoecology of a transitional sequence
Article number: 28.2.a32
https://doi.org/10.26879/1536
Copyright Society of Vertebrate Paleontology, July 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Supplementary material
Submission: 24 January 2025. Acceptance: 9 July 2025.
ABSTRACT
Campanian estuarine deposits are rare across Europe, and the assemblages described herein represent a paleontologically significant exception. Systematic collection efforts conducted over the past eight years have resulted in the identification of 102 taxa spanning a broad spectrum of organisms, including calcareous nannoplankton, seeds, fruits, bivalves, gastropods, ammonites, corals, echinoids, asteroids, selachians, actinopterygians, anurans, squamates, crocodyliforms, and putative theropod dinosaurs, the latter inferred from eggshell fragments. Notably, a new percomorph fish taxon is described — Paraplesiopoma transylvanica n. gen. et n. sp. (Trif and Schwarzhans). In addition, amber — a rare occurrence in European Cretaceous strata — was recovered from the site. Sedimentological and paleontological evidence indicates a marked paleoenvironmental transition from fully marine to brackish, estuarine ecosystem. The presence of continental taxa within the assemblage offers valuable insights into the composition of the adjacent terrestrial ecosystems during the Campanian. This assemblage constitutes a unique mixture of autochthonous marine and aquatic elements with parautochthonous and allochtonous continental/terrestrial components, situated within the Alpine Tethyan realm, during the Late Cretaceous. Furthermore, it documents a diverse array of terrestrial organisms predating the well-known Maastrichtian vertebrate faunas of the so-called “Hațeg Island”, thereby providing an important window into the pre-Maastrichtian terrestrial biota of this region.
Nicolae Trif. Brukenthal National Museum, Natural History Museum, Cetății St. 1-550160, Sibiu, Romania; and Research Center for Integrated Geological Studies, Mihail Kogălniceanu St. 1-4000084, Cluj-Napoca, Romania (corresponding author). nicolae.trif@gmail.com
Roxana Pirnea. Independent researcher. roxana.pirnea@gmail.com
Andrej Čerňanský. Department of Ecology, Laboratory of Evolutionary Biology, Faculty of Natural Sciences, Comenius University in Bratislava, Mlynská dolina, Ilkovičova St. 6-84215, Bratislava, Slovakia; and Institute of Evolutionary Biology, Faculty of Biology, University of Warsaw, Żwirki i Wigury 101, 02-089 Warsaw, Poland. cernansky.paleontology@gmail.com
Werner Schwarzhans. Zoological Museum, Natural History Museum of Denmark, Universitetsparken 15-2100 Copenhagen, Denmark. wwschwarz@aol.com
Ramona Bălc. Babeș-Bolyai University, Faculty of Environmental Science and Engineering, Fântânele St. 30-400294, Cluj-Napoca, Romania; and Interdisciplinary Research Institute on Bio-Nano Sciences, Babeș-Bolyai University, Treboniu Laurian St. 42-400271, Cluj-Napoca, Romania; and Center for Risk Studies, Space Modeling and Dynamics of Terrestrial and Coastal Systems, University of Bucharest, Nicolae Bălcescu St. 1-010041, Bucharest, Romania. ramona.balc@ubbcluj.ro
Sreepat Jain. Department of Geology, School of Applied Natural Science, Adama Science and Technology University, Adama St. 1888, Oromia, Ethiopia. sreepatjain@gmail.com
Andrzej Kaim. Institute of Paleobiology, Polish Academy of Sciences, ul. Twarda 51/55, 00-818, Warsaw, Poland. kaim@twarda.pan.pl
Sofia Bakayeva. Institute of Paleobiology, Polish Academy of Sciences, ul. Twarda 51/55, 00-818, Warsaw, Poland; and State Museum of Natural History, National Academy of Sciences of Ukraine, Teatralna St. 18, 79008, Lviv, Ukraine. sofiyabakayeva@gmail.com
Krzysztof Hryniewicz. Institute of Paleobiology, Polish Academy of Sciences, ul. Twarda 51/55, 00-818, Warsaw, Poland. krzyszth@twarda.pan.pl
Keywords: vertebrates; invertebrates; plants; amber; paleoecology; Upper Cretaceous
https://zoobank.org/531B0560-BF59-48C6-ADC5-23EFF837346B
Final citation: Trif, Nicolae, Pirnea, Roxana, Čerňanský, Andrej, Schwarzhans, Werner, Bălc, Ramona, Jain, Sreepat, Kaim, Andrzej, Bakayeva, Sofia, and Hryniewicz, Krzysztof. 2025. A diverse Campanian biota from the Bozeș Formation (Petrești, Romania): Insights into the paleontology and paleoecology of a transitional sequence. Palaeontologia Electronica, 28(2):a32.
https://doi.org/10.26879/1536
palaeo-electronica.org/content/2025/5577-bozes-formation-campanian-biota
Copyright: July 2025 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
The Upper Cretaceous continental formations of Romania have garnered increasing international scientific interest over recent decades, largely due to the discovery of the so-called “dwarf dinosaurs” in the Hațeg Basin. These iconic fossils were first described in the late nineteenth and early twentieth centuries by Nopcsa (1897, 1902, 1914a, 1914b, 1915, 1923, 1925, 1929). Following these pioneering studies, research on Late Cretaceous vertebrates in Romania experienced a prolonged period of neglect, with only sporadic references in reports by geologists of the Geological Institute of Romania (e.g., Mamulea, 1953a, 1953b; Dincă et al., 1972). A revival of systematic paleontological investigations occurred during the 1980s, leading to the discovery of a remarkably diverse vertebrate assemblage. These excavations expanded the known faunal spectrum beyond dinosaurs to include mammals, pterosaurs, lissamphibians, and squamates (Grigorescu, 1983; Grigorescu and Hahn, 1987; Grigorescu et al., 1985, 1990, 1999; Jianu et al., 1997). Since then, paleontological prospecting has expanded significantly beyond the classical localities of the Hațeg Basin, with new vertebrate-bearing sites being discovered or rediscovered, thereby enlarging the known spatial distribution of fossiliferous Upper Cretaceous deposits in Romania (Csiki-Sava et al., 2016). Noteworthy additional localities include outcrops in the Sebeș region, near the village of Rusca Montană, and at Someș-Odorhei in the Jibou area, which have yielded significant fossil assemblages and have been the focus of renewed attention (Codrea et al., 2010b, 2012; Vremir et al., 2014, 2015).
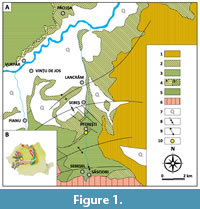 The area surrounding Sebeș (Alba County, Romania) hold considerable paleontological significance, as it represents the only currently known continuous Upper Cretaceous transition from marine to continental sedimentation in Romania. This transition is preserved at the lithostratigraphic contact between the Bozeș and Sebeș formations (Vremir, 2010; Vremir et al., 2014; Csiki-Sava et al., 2016; Țabără et al., 2022; Bălc et al., 2024) (Figure 1). The site, located near the locality of Petrești, was first reported by Codrea et al. (2010a), who documented a bone fragment tentatively attributed to an indeterminate dinosaur. Subsequent investigations quickly followed, leading to the discovery of additional vertebrate remains attributed to dinosaurs, turtles, mammals, and pterosaurs. These fossils have been recovered predominantly from the Sebeș Formation, though some occurrences have also been documented within the underlying Bozeș Formation (Vremir, 2010; Vremir et al., 2014, 2015; Brusatte et al., 2013).
The area surrounding Sebeș (Alba County, Romania) hold considerable paleontological significance, as it represents the only currently known continuous Upper Cretaceous transition from marine to continental sedimentation in Romania. This transition is preserved at the lithostratigraphic contact between the Bozeș and Sebeș formations (Vremir, 2010; Vremir et al., 2014; Csiki-Sava et al., 2016; Țabără et al., 2022; Bălc et al., 2024) (Figure 1). The site, located near the locality of Petrești, was first reported by Codrea et al. (2010a), who documented a bone fragment tentatively attributed to an indeterminate dinosaur. Subsequent investigations quickly followed, leading to the discovery of additional vertebrate remains attributed to dinosaurs, turtles, mammals, and pterosaurs. These fossils have been recovered predominantly from the Sebeș Formation, though some occurrences have also been documented within the underlying Bozeș Formation (Vremir, 2010; Vremir et al., 2014, 2015; Brusatte et al., 2013).
In subsequent years, research efforts focused predominantly on the Sebeș Formation, particularly at an outcrop located several kilometers north of Petrești. These investigations have yielded a diverse assemblage of vertebrate fossils, including representatives of dinosaurs, pterosaurs, birds, crocodyliforms, turtles, anuran amphibians, multituberculate mammals, and actinopterygian fishes (Csiki et al., 2010; Codrea and Jipa, 2011; Dyke et al., 2012; Vremir et al., 2014; Fernández et al., 2019; Solomon et al., 2020, 2022a, 2022b; Vasile et al., 2021, 2022; Trif and Codrea, 2022). These taxa are interpreted as components of the fauna that inhabited the so-called “Hațeg Island”.
While research on the Sebeș Formation has increasingly focused on macrofossils, investigations of the underlying Bozeș Formation and the intervening transitional estuarine interval have primarily concentrated on microfossil assemblages. These studies have emphasized calcareous nannoplankton, palynomorphs, and have incorporated organic geochemical analyses to reconstruct paleoenvironmental conditions (e.g., Bălc and Chira, 2002; Bălc et al., 2007, 2012, 2024; Țabără et al., 2022). Nevertheless, historical studies have also documented certain macrofossil groups from the Bozeș Formation, notably mollusks and isolated plant remains (Staub, 1889; Pálfy, 1902).
Given the rarity of such transitional estuarine environments in Romania, we conducted an extensive, multidisciplinary investigation over an eight-year period to better understand the sedimentological and paleontological characteristics of this unique Upper Cretaceous sequence. Fieldwork was carried out along the right bank of the Sebeș River, near the village of Petrești, a northern suburb of the city of Sebeș (Alba County). This site yielded a diverse assemblage of invertebrate fossils, as well as both terrestrial and aquatic microvertebrate remains, eggshell fragments, plant seeds, leaf impressions, and amber — all of which form the basis of this study.
GEOLOGICAL SETTING
The studied section is situated in the Sebeș area, in the southwestern part of the Transylvanian Basin, between the Metaliferi Mountains (a subunit of the Apuseni Mountains) and the Șureanu Mountains (part of the Southern Carpathians) (Krézsek and Bally, 2006) (Figure 1).
The Transylvanian Basin is a Late Cretaceous-Paleogene sedimentary basin that developed atop a complex structural substrate comprising a Paleozoic crystalline basement, ophiolitic assemblages, island-arc volcanic units, and Triassic to mid-Cretaceous sedimentary sequences (Săndulescu and Visarion, 1978; Paraschiv, 1979; Bortolotti et al., 2004; Krézsek and Bally, 2006). The basin’s sedimentary infill is subdivided into four major tectonostratigraphic megasequences: Upper Cretaceous, Paleogene, Lower Miocene, and Middle to Upper Miocene (Krézsek and Bally, 2006). Of particular relevance to this study are the Upper Cretaceous deposits, which comprise the marine Bozeș Formation and the overlaying continental Sebeș Formation. These units record a rare, continuous marine-to-continental transition, making the region a key area for reconstructing Late Cretaceous paleoenvironmental dynamics in the Carpathian realm.
During the Late Cretaceous, numerous sedimentary basins developed across the region, accumulating a wide range of deposits from deep- and shallow-marine to continental settings. The latter are frequently characterized by red beds (Willingshofer et al., 2001; Krézsek and Bally, 2006). In the Sebeș area, the marine succession is represented by the Bozeș Formation, a typical flysch unit with an estimated thickness of approximately 3000 m. This formation comprises a varied lithological assemblage, including sandstones, calcareous sandstones, silty marls, microconglomerates, and conglomerates, particularly in its upper intervals (Dimian and Popa-Dimian, 1964; Bălc et al., 2007). Initially, the Bozeș Formation was assigned a Santonian to early Maastrichtian age based on macrofossil evidence (Dimian and Popa-Dimian, 1964; Tomescu et al., 1969). However, more recent biostratigraphic analysis utilizing calcareous nannoplankton, foraminiferal, and palynological assemblages have revised this age to the mid- to late Campanian (Bălc et al., 2007, 2024; Țabără et al., 2022). While certain ammonite taxa previously suggested a possible Maastrichtian age (Macovei and Atanasiu, 1934; Tomescu et al., 1969), this interpretation remains unconfirmed by current biostratigraphic frameworks (Bălc et al., 2007). At the top of the Bozeș Formation, within the Petrești section, a relatively thin but distinct brackish-estuarine interval is preserved, marking a transitional phase between marine and continental deposition (Vremir et al., 2014, Bălc et al., 2024).
The Sebeș Formation conformably overlies the Bozeș Formation and has been interpreted as Maastrichtian in age (Vremir et al., 2014, 2015). An alternative stratigraphic framework and sedimentological interpretation was proposed by Codrea and Dica (2005), who suggested the presence of a distinct lithostratigraphic unit. However, this interpretation has since been refuted. The lithological units proposed in that model are now regarded as facies variations within a single depositional system, reflecting lateral changes due to variable sediment sources (for a detailed discussion see Vremir, 2010; Vremir et al., 2015). In the Petrești section, the Sebeș Formation is characterized by transitional wetland to lacustrine deposits, as well as fluvial mudstones and channel-fill sediments (Vremir et al., 2014).
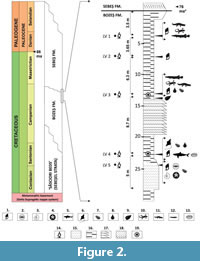 The study site corresponds to the uppermost stratigraphic interval of the Bozeș Formation, encompassing its final meter prior to the transition into the overlying Sebeș Formation. The succession consists of alternating sandy marls, marls, and sandstones, with a total measured thickness of 27 m. At its base, the sequence begins with approximately 5 m of fully marine deposits, characterized by compacted sands grading into sandstones and yielding a diverse assemblage of marine invertebrates, including corals, gastropods, bivalves, ammonites, rudists, asteroids, and echinoids. These marine beds are overlain by approximately 22 m marls, sandy marls, and sandstones interpreted as representing a transitional estuarine depositional environment. Within this composite section, five discrete fossiliferous horizons have been identified (designated LV1 through LV5), with individual thicknesses ranging from 0.2 to 0.4 m (LV1 to LV4), and up to 5 m in the case of LV5 (Figure 2).
The study site corresponds to the uppermost stratigraphic interval of the Bozeș Formation, encompassing its final meter prior to the transition into the overlying Sebeș Formation. The succession consists of alternating sandy marls, marls, and sandstones, with a total measured thickness of 27 m. At its base, the sequence begins with approximately 5 m of fully marine deposits, characterized by compacted sands grading into sandstones and yielding a diverse assemblage of marine invertebrates, including corals, gastropods, bivalves, ammonites, rudists, asteroids, and echinoids. These marine beds are overlain by approximately 22 m marls, sandy marls, and sandstones interpreted as representing a transitional estuarine depositional environment. Within this composite section, five discrete fossiliferous horizons have been identified (designated LV1 through LV5), with individual thicknesses ranging from 0.2 to 0.4 m (LV1 to LV4), and up to 5 m in the case of LV5 (Figure 2).
These deposits are overlain by a thick succession of well-consolidated conglomerates and sandstones, which are interpreted as representing the lowermost stratigraphic interval of the Sebeș Formation (see Figure 3).
MATERIALS AND METHODS
 The site was systematically investigated between 2016 and 2024. Fieldwork and sampling for both macro- and micropaleontological analyses were conducted across five stratigraphic levels.
The site was systematically investigated between 2016 and 2024. Fieldwork and sampling for both macro- and micropaleontological analyses were conducted across five stratigraphic levels.
To assess the fossil content, a total of 272 kg of marl and sandy marl was collected from these levels, where fossil concentrations were visually noted to be higher. The samples underwent a disaggregation procedure involving controlled drying to a residual humidity of 5-10%, followed by rapid rehydration using water heated to 100°C. The resulting disaggregated sediment was then wet-sieved using a 0.5 mm mesh. The residue retained on the sieve was subsequently dried and examined under a stereomicroscope. This protocol enabled the recovery of a diverse assemblage of microfossils, including microvertebrate remains, plant seeds, eggshell fragments, small mollusks, and amber.
An additional collection technique was employed specifically for the recovery of fragile mollusk specimens. Large, intact fragments of wet sediment were carefully extracted in the field and transported to the laboratory under controlled conditions to maintain their original moisture content. In the laboratory, mollusk specimens were meticulously extracted from the matrix using fine needles and scalpels. To facilitate the cleaning and to stabilize the fragile shell structures, a 5% polyvinyl acetate (PVA) solution was applied with a fine brush to remove residual sediments and consolidate any existing fractures. The treated specimens were then allowed to dry gradually at ambient room temperature.
The otoliths classification follows the phylogenetic framework of Betancur-R. et al. (2017), while the morphological terminology is based on Koken (1884), with modifications by Chaine and Duvergier (1934) and Schwarzhans (1978). All otoliths are illustrated from the right side; left-side specimens were mirrored digitally and are clearly indicated as such in figure captions.
Macroscopic specimens, including mollusks, corals, and echinoids, were photographed using a Nikon D5300 digital camera fitted with a Sigma 105 mm macro lens. Microvertebrate remains and plant seeds were imaged with an AmScope 10X Plan Achromatic objective mounted on a 200 mm host lens. To enhance image sharpness and depth field, focus staking was performed using CombineZP software (version1.0). Eggshell fragments were examined and photographed using a TescanVega II SBH Scanning Electronic Microscope.
Calcareous nannoplankton analysis was conducted on five samples collected from the uppermost part of the marine sequence to refine the biostratigraphic age of the studied section. Smear slides were prepared following the standard technique outlined by Bown and Young (1998), and analyzed under a light microscope (Axiolab A) at 1000x magnification. Four traverses were completed per slide (~700 fields of view), and all identifiable taxa were counted to determine relative abundance. Selected specimens were imaged using an AxioCam ERCc5s digital microscope camera for illustration.
Fossil specimens exceeding 1 mm in maximum dimension were measured using a Profi Scale Precise PS 7215 electronic caliper, which has a reported measurement accuracy of ±0.01 mm. Smaller specimens were measured digitally using ImageJ software (version 1.54g), calibrated with a graphical scale included in each image alongside the specimen.
All collected specimens have been deposited in the Paleontological Collection of the Natural History Museum in Sibiu, part of the Brukenthal National Museum, and are referenced hereafter with the abbreviation NHMS.
Cluster analysis was conducted using Past software (version 0.45), employing the paired group (UPGMA) algorithm and Jaccard similarity index to assess taxonomic associations among the fossil assemblages.
Abbreviations used in the text: LV1 — level 1; LV2 — level 2; LV3 — level 3; LV4 — level 4; LV5 — level 5; OL — otolith length; OH — otolith height; OT — otolith thickness; OsL — ostium length; OsH — ostium height; CaL — cauda length; CaH — cauda height; U/D — umbilical width to shell diameter ratio; NHMS — Natural History Museum, Sibiu.
RESULTS AND DISCUSSION
1. Flora
Microflora – Calcareous Nannoplankton
The calcareous nannofossil assemblage is characterized by poor to moderate preservation and comprises 36 species, exhibiting very low relative abundances (ranging from 0.02 and 0.21 specimens per field of view). Both species richness and total abundance decrease progressively from sample LV5 to sample LV1, with the number of recorded species declining from 22 to 7. Detailed counts and species occurrences for each stratigraphic level are provided in Table 1.
The most dominant taxon within the assemblage is Watznaueria barnesiae (Black in Black and Barnes, 1959; Perch-Nielsen, 1968), representing between 29.6 and 55.5% of the total assemblage. Other common species include Retecapsa crenulata (Bramlette and Martini, 1964; Grün in Grün and Allemann, 1975), Eiffellithus eximius (Stover 1966; Perch-Nielsen, 1968), Prediscosphaera cretacea (Arkhangelsky, 1912; Gartner, 1968), and Cribrosphaerella ehrenbergii (Arkhangelsky, 1912; Deflandre in Piveteau, 1952).
Key taxa identified within the recovered calcareous nannofossil assemblage, relevant for biostratigraphic age determination, include Broinsonia parca constricta (Hattner et al., 1980), Reinhardtites anthophorus (Deflandre, 1959; Perch-Nielsen, 1968), and Eiffellithus eximius.
 All of these species are illustrated in Figure S1 of Supplementary materials, Section A. It is noteworthy, however, that several biostratigraphically diagnostic taxa typically indicative of the Campanian are absent from the assemblage, thereby limiting the resolution of age assignment. Nevertheless, based on the presence of the aforementioned taxa, the age of the deposits can be constrained. The first occurrence of Broinsonia parca constricta marks the base of the UC 14b Biozone (Burnett, 1998), corresponding to approximately 81.4 Ma (Young et al., 2022), thus indicating a maximum (oldest possible) age no earlier than the early Campanian. Furthermore, the last occurrence of Reinhardtites anthophorus and Eiffellithus eximius fall within the UC15e subzone of Burnett (1998), dated to the later part of the late Campanian, thereby providing a minimum (youngest possible) age constraint. Consequently, the stratigraphic interval under study is tentatively assigned to a timespan between the early and late Campanian.
All of these species are illustrated in Figure S1 of Supplementary materials, Section A. It is noteworthy, however, that several biostratigraphically diagnostic taxa typically indicative of the Campanian are absent from the assemblage, thereby limiting the resolution of age assignment. Nevertheless, based on the presence of the aforementioned taxa, the age of the deposits can be constrained. The first occurrence of Broinsonia parca constricta marks the base of the UC 14b Biozone (Burnett, 1998), corresponding to approximately 81.4 Ma (Young et al., 2022), thus indicating a maximum (oldest possible) age no earlier than the early Campanian. Furthermore, the last occurrence of Reinhardtites anthophorus and Eiffellithus eximius fall within the UC15e subzone of Burnett (1998), dated to the later part of the late Campanian, thereby providing a minimum (youngest possible) age constraint. Consequently, the stratigraphic interval under study is tentatively assigned to a timespan between the early and late Campanian.
Our results are consistent with the findings of Bălc et al. (2024), who also reported the presence of calcareous nannoplankton assemblages in the uppermost part of the Bozes Formation. This concordance between studies enhances the robustness of the proposed age model. Our study reinforces the conclusions of Bălc et al. (2024) and independently confirms the assignment of this stratigraphic interval to the Late Campanian.
Mesoflora
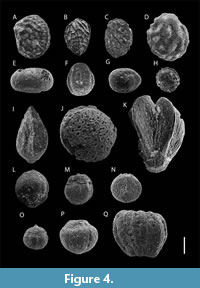 A mesofloral assemblage was recovered from fossiliferous levels LV3 and LV4 of the Bozeș Formation. The specimens are generally well-preserved; however, most exhibit varying degrees of abrasion and cracking. Seventeen distinct types of angiosperm seeds and fruits (Figure 4A-Q) were identified and recorded as separate taxa. Due to compression and mechanical weathering, precise measurements of seed dimensions were often difficult to take.
A mesofloral assemblage was recovered from fossiliferous levels LV3 and LV4 of the Bozeș Formation. The specimens are generally well-preserved; however, most exhibit varying degrees of abrasion and cracking. Seventeen distinct types of angiosperm seeds and fruits (Figure 4A-Q) were identified and recorded as separate taxa. Due to compression and mechanical weathering, precise measurements of seed dimensions were often difficult to take.
Nine types of small anatropous seeds were recognized within the mesofloral material. Four of these (Taxa 1-4, Figure 4A-D) exhibit ornamented surfaces composed of polygonal facets forming distinctive seed coats. These seeds are sub-rounded to elliptical in shape ranging from 2.2 to 2.7 mm in length and 1.6 to 2.4 mm in breadth. A narrow circumferential furrow is consistently present in these specimens. One oblate seed with a shallow depression at each pole was identified as Taxon 5 (Figure 4E). It measures 0.7 mm in length and 1.2 mm in breadth, and its surface is covered with very fine linear striations connecting the two poles. Taxon 6 (Figure 4F) is represented by an elliptical seed with a surface pattern of fine concentric circles, lacking any discernible anatomical features. It measures 1.9 mm in length and 1.6 mm in breadth. Two additional rounded seeds with little to no surface ornamentation were documented as Taxa 7 and 8 (Figure 4G-H). Despite clear signs of weathering, both depressions and projections are evident on each grain. Taxon 7 measures 1.4 mm in length and 1.9 mm in breadth, while Taxon 8 is 1.4 mm in both dimensions. The last analyzed seed, designated as Taxon 9 (Figure 4I), has a distinct lenticular shape, lacks surface ornamentation, and measures 2.2 mm in length and 2 mm in breadth.
Two fruit specimens, designated as Taxon 10 and Taxon 11, exhibit morphological features characteristic of the Normapolles group (Figure 4J-K), including a distinct triangular shape and tricolpate apertures in polar view. Taxon 10 measures 6.3 mm in length and 3.6 mm in breadth, while Taxon 11 measures 1.9 mm in length and 2.2 mm in breadth. Similar to previously described specimens, these could not be assigned to specific lower-level taxa. The term “Normapolles” generally refers to fossil pollen grains from the Late Cretaceous to early Paleogene, characterized by a distinctive triangular amb and typically triporate structure (Pflug, 1953; Góczán et al., 1967; Tschudy, 1975; Polette and Batten, 2017). These forms have often been associated with members of the order Fagales.
In addition to the aforementioned seeds and fruits, the Petrești mesoflora includes six additional taxa of capsular fruits whit currently uncertain systematic affinities. These fruits exhibit varying numbers of capsules and are preserved at an early, indehiscent stage of maturity. No petals were observed in association with the fruits, and no style were preserved, likely due to abrasion. Taxon 12 (Figure 4L) is a pentacarpellate fruit with a rounded side composed of three locules and a flattened side with two locules. It measures 1.5 mm in both length and breadth. Although the surface is heavily abraded, the locules and their ornamentation remain discernible. Taxon 13 (Figure 4M) consists of four preserved locules, but its overall architecture suggests it is also pentacarpellate. This is the largest fruit specimen recorded, measuring approximately 4.4 mm in length and 3.2 mm in breadth. It lacks ornamentation, but the calyx is partially preserved. Taxon 14 (Figure 4N) is arguably the best-preserved specimen among the capsular fruits. It comprises six locules with an ornamented surface on one side and two flattened locules on the opposite side. The specimen measures 2.9 mm in length and 3.2 mm in breadth. Taxon 15 (Figure 4O) represents a fragment of a spherical fruit with well-preserved surface ornamentation. Although the specimen cracked during handling, it measures approximately 3.1 mm in diameter. Taxon 16 (Figure 4P) is a spherical fruit composed of four locules, measuring 1.4 mm in length and 1.5 mm in breadth. It shows no surface ornamentation, although the calyx is preserved. Taxon 17 (Figure 4Q) is strongly compressed and displays extensive fissuring. It measures 1.9 mm in length and 1.8 mm in breadth, a sub-spherical shape, and appears to consist of four locules. The calyx is partially preserved, and no ornamentation is visible.
The overall composition of the mesofloral assemblage recovered from the Bozeș Formation bears a general resemblance to other Late Cretaceous mesofloras from Europe. Similar to the Maastrichtian Budurone mesoflora of the Hațeg Basin, Romania (Lindfors et al., 2010), the specimens described herein may represent new, previously undescribed taxa, potentially reflecting the biogeographic isolation of the Petrești flora. Given the limited number of specimens available for each taxon, no formal systematic assignments are proposed at this time. Consequently, this study should be regarded as a preliminary report on the mesofloral content of the Bozeș Formation. The challenges in assigning these specimens to established genera or families highlight the need for further systematic investigation and comparative analysis.
Macroflora
 The macrofloral assemblage identified at Petrești is represented by a single taxon, collected from fossiliferous level LV3. The material belongs to the class Polypodiopsida and is tentatively assigned to cf. Sphenopteris ungeri Kvaček and Herman (in Herman and Kvaček, 2010) (Figure 5A-B).
The macrofloral assemblage identified at Petrești is represented by a single taxon, collected from fossiliferous level LV3. The material belongs to the class Polypodiopsida and is tentatively assigned to cf. Sphenopteris ungeri Kvaček and Herman (in Herman and Kvaček, 2010) (Figure 5A-B).
The specimens consist of sterile, entire-margined, lanceolate pinnules with attenuate apices and decurrent bases. In one specimen, the pinnules appear to be basally fused (Figure 5B). The venation is pinnate, with a slender midvein extending towards the apex and numerous, thin secondary veins that branch dichotomously near the margin.
Although the material is fragmentary and poorly preserved, the morphological features closely resemble the apical pinnules of Sphenopteris ungeri described from the Late Cretaceous flora of Grünbach, Austria (Herman and Kvaček, 2007, 2010). This species was originally recorded in the Cretaceous flora of Austria under the name Pecopteris striata Presl in Sternberg (Unger, 1867). Cuticle fragments are present in the Petrești specimens; however, because of the poor state of preservation, cuticle extraction was not possible. Such preservation limitations are common for the plant macrofossils from the Petrești site.
Sphenopteris ungeri is frequently encountered in Late Cretaceous floras of Europe and is represented as a component of riparian vegetation, commonly occurring alongside Ettingshausenia (Herman and Kvaček, 2010).
Amber
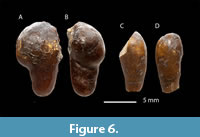 Sediment sieving yielded several amber fragments, all of which were recovered from fossiliferous levels LV3 and LV4. Most fragments are small, ranging from 1 to 3 mm in size, although a few larger specimens measure between 8 and 11 mm (see Figure 6). The amber exhibits an external coloration ranging from brown to yellow-brown, with a distinctly lighter interior that varies from white to light yellow or light brown. The outer surfaces are generally smooth, though some specimens display fine scratches and surface cracking. Morphologically, the amber fragments are predominantly cylindrical, rounded, or teardrop-shaped. The preservation of these delicate morphologies — especially in such fragments — suggests minimal transport or deposition within a calm aquatic environment near the site of resin exudation.
Sediment sieving yielded several amber fragments, all of which were recovered from fossiliferous levels LV3 and LV4. Most fragments are small, ranging from 1 to 3 mm in size, although a few larger specimens measure between 8 and 11 mm (see Figure 6). The amber exhibits an external coloration ranging from brown to yellow-brown, with a distinctly lighter interior that varies from white to light yellow or light brown. The outer surfaces are generally smooth, though some specimens display fine scratches and surface cracking. Morphologically, the amber fragments are predominantly cylindrical, rounded, or teardrop-shaped. The preservation of these delicate morphologies — especially in such fragments — suggests minimal transport or deposition within a calm aquatic environment near the site of resin exudation.
Cretaceous amber occurrences in Romania are poorly documented and limited to sporadic, often outdated reports. The earliest known reference originates from the unpublished manuscript of Paul Maria Joseph Partsch, who described in 1826 the amber from Săsciori, Transylvania (approximately 6 km south of Petrești) in his Tagebücher seiner Reisen in Siebenbürgen (“Diaries of his travels in Transylvania”). This account was later cited by Hauer and Stache (1863), bringing it into the formal scientific literature. Subsequent studies, including those by Zechmeister and Vrabely (1927), identified and described the amber from Alba County as a purported new mineral species, named Telegdit. Later mentions are brief and general, such as those by Protesco (1937) and Rabichon (1938), in broader reviews of Romanian amber deposits. The precise age of the Săsciori amber remains unresolved. Both Santonian and Campanian sedimentary units have been identified in the region (Marincaș, 1966, 1970; Tomescu, 1968), but the stratigraphic origin of the amber-bearing layers remains uncertain. Notably, the amber from Săsciori housed in the Brukenthal National Museum in Sibiu differs markedly from the Petrești material in terms of color, clarity, and fracture characteristics (NT, personal observation).
Codrea et al. (2012) briefly reported the presence of “yellowish amber-like resin” in the Rusca Montană Basin, potentially representing an occurrence of Maastrichtian amber.
Upper Cretaceous amber deposits are relatively uncommon in Europe. Nevertheless, several occurrences have been documented, including sites in Austria (Turonian-Santonian; Ősi et al., 2021), France (uppermost Albian-lowermost Cenomanian; Néraudeau et al., 2008; Adl et al., 2011), and Hungary (Santonian; Szabó, 1871; Ősi et al., 2012). The recent identification of amber from Petrești, dated to the Upper Campanian, expands this geographical and stratigraphic distribution and provides a valuable addition to the existing reports.
It is noteworthy that the amber from Petrești was recovered as rather a by-product of microvertebrate and micropaleontological sampling, and not through targeted collection efforts. Future investigations employing specialized recovery techniques, such as flotation in dense liquids, may facilitate the retrieval of a greater quantity of amber specimens, thereby enabling comprehensive analyses.
2. Invertebrates
Mollusks
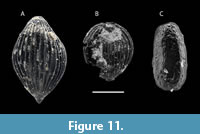
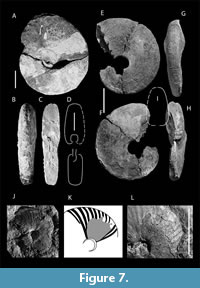 Cephalopods. Three specimens of Pachydiscus (Pachydiscus) cf. haldemsis (Schlüter, 1867) (M) were recovered from LV5 (Figure 7 ). These specimens closely resemble Pachydiscus (Pachydiscus) cf. haldemsis in their compressed shell morphology and whorl section. However, P. (P.) cf. haldemsis is somewhat more evolute, exhibiting a U/D ratio = ~0.30 at 126 mm, compared to a U/D ratio of 0.25 at 237 mm reported in previous studies (Kennedy and Kaplan, 1997; Kaplan et al., 2005; Kennedy et al., 2007). The macroconch of Pachydiscus (Pachydiscus) oldhami (Sharpe, 1853) is also morphologically comparable, but differs by displaying a weaker and more crowded ribbing pattern. In P. (P.) oldhami, ornamentation diminishes in ontogeny, becoming largely absent on the late phragmocone stage, particularly on the inner flank (Kennedy and Summesberger, 1984; Kennedy, 1986a; Kennedy and Kaplan, 1997). A general comparison can also be drawn with the upper Campanian Pachydiscus (Pachydiscus) colligatus (Binkhorst, 1861) (see Kennedy, 1986a, 1986b, 1986c, 1986d; Martínez, 1997); however, the Petrești specimens are relatively more compressed and exhibit a narrow, rounded venter, in contrast to the broader rounded venter characteristic of P. (P.) colligatus (Kennedy, 1986a, 1986b, 1986c, 1986d). Given the incomplete, fragmentary, and somewhat crushed condition of the material, the specimens are provisionally assigned to Pachydiscus (Pachydiscus) cf. haldemsis (Schlüter, 1867) (M). This species has previously been reported from a wide geographic range, including Northern Ireland, Westphalia (Germany), the Gschliefgraben (Upper Austria), Poland, Sweden, Donbass (Ukraine), Kopet Dag Basin (Iran), Turkmenia, and Kastamonu (Turkey).
Cephalopods. Three specimens of Pachydiscus (Pachydiscus) cf. haldemsis (Schlüter, 1867) (M) were recovered from LV5 (Figure 7 ). These specimens closely resemble Pachydiscus (Pachydiscus) cf. haldemsis in their compressed shell morphology and whorl section. However, P. (P.) cf. haldemsis is somewhat more evolute, exhibiting a U/D ratio = ~0.30 at 126 mm, compared to a U/D ratio of 0.25 at 237 mm reported in previous studies (Kennedy and Kaplan, 1997; Kaplan et al., 2005; Kennedy et al., 2007). The macroconch of Pachydiscus (Pachydiscus) oldhami (Sharpe, 1853) is also morphologically comparable, but differs by displaying a weaker and more crowded ribbing pattern. In P. (P.) oldhami, ornamentation diminishes in ontogeny, becoming largely absent on the late phragmocone stage, particularly on the inner flank (Kennedy and Summesberger, 1984; Kennedy, 1986a; Kennedy and Kaplan, 1997). A general comparison can also be drawn with the upper Campanian Pachydiscus (Pachydiscus) colligatus (Binkhorst, 1861) (see Kennedy, 1986a, 1986b, 1986c, 1986d; Martínez, 1997); however, the Petrești specimens are relatively more compressed and exhibit a narrow, rounded venter, in contrast to the broader rounded venter characteristic of P. (P.) colligatus (Kennedy, 1986a, 1986b, 1986c, 1986d). Given the incomplete, fragmentary, and somewhat crushed condition of the material, the specimens are provisionally assigned to Pachydiscus (Pachydiscus) cf. haldemsis (Schlüter, 1867) (M). This species has previously been reported from a wide geographic range, including Northern Ireland, Westphalia (Germany), the Gschliefgraben (Upper Austria), Poland, Sweden, Donbass (Ukraine), Kopet Dag Basin (Iran), Turkmenia, and Kastamonu (Turkey).
For detailed descriptions, morphometric data, taxonomy, additional remarks, and supplementary imagery, refer to Supplementary materials, Section B.
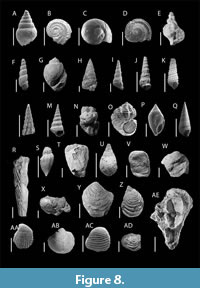 Gastropods. A substantial assemblage of gastropods and bivalves was recovered from all five sampled stratigraphic levels. Gastropods dominate the mollusk fauna and encompass representatives from several higher taxonomic groups, including Seguenziida, Cycloneritida, Littorinimorpha, Sorbeoconcha, Neogastropoda, and Heterobranchia. Within Seguenziida, specimens belong to the family Chilodontaidae, represented by Onkospira sp. Figure 8A. The order Cycloneritida includes taxa from the families Neritidae — namely Neritoptyx spinosa (Pálfy, 1902) Figure 8B and Neritoplica sp. Figure 8C — and Deianiridae, represented by Deianira bicarinata (Zekeli, 1852) Figure 8D. Littorinimorpha is represented by Capulidae, possibly Trichotropis sp. Figure 8E, and Hydrobiidae, represented by Parateinostoma sp. Figure 8F. The most taxonomically diverse group is Sorbeoconcha, comprising multiple families: Ampullinidae (Pseudamaura alkenyeriensis [Pálfy, 1902]); Figure 8G, Cassiopidae (Cassiope obvoluta Schlotheim, 1820); Figure 8H), Procerithiidae (? Rhabdocolpus sp. and Ageria sp.; Figure 8I-J), Potamididae Pirenella alvincziense (Pálfy, 1902); Figure 8L), Pleuroceridae (Pleurocera sp.; Figure 8M), Turritellidae (?Nairiella sp.; Figure 8N), Hemisinidae (Pyrgulifera boeckhi Pálfy, 1902; Figure 8O), and Melanopsidae (Melanopsis crastina [Vidal, 1874] and Campylostylus galloprovincialis [Matheron, 1843]); Figure 8P-Q. Neogastropoda is represented by the family Volutidae, with specimens assigned to Volutilithes sp. Figure 8S. Additionally, two heterobranchs taxa were identified: one assigned to Acteonellidae (Trochactaeon sp.; Figure 8T), and another classified as Acteonoidea indet. (Figure 8U) due to poor preservation.
Gastropods. A substantial assemblage of gastropods and bivalves was recovered from all five sampled stratigraphic levels. Gastropods dominate the mollusk fauna and encompass representatives from several higher taxonomic groups, including Seguenziida, Cycloneritida, Littorinimorpha, Sorbeoconcha, Neogastropoda, and Heterobranchia. Within Seguenziida, specimens belong to the family Chilodontaidae, represented by Onkospira sp. Figure 8A. The order Cycloneritida includes taxa from the families Neritidae — namely Neritoptyx spinosa (Pálfy, 1902) Figure 8B and Neritoplica sp. Figure 8C — and Deianiridae, represented by Deianira bicarinata (Zekeli, 1852) Figure 8D. Littorinimorpha is represented by Capulidae, possibly Trichotropis sp. Figure 8E, and Hydrobiidae, represented by Parateinostoma sp. Figure 8F. The most taxonomically diverse group is Sorbeoconcha, comprising multiple families: Ampullinidae (Pseudamaura alkenyeriensis [Pálfy, 1902]); Figure 8G, Cassiopidae (Cassiope obvoluta Schlotheim, 1820); Figure 8H), Procerithiidae (? Rhabdocolpus sp. and Ageria sp.; Figure 8I-J), Potamididae Pirenella alvincziense (Pálfy, 1902); Figure 8L), Pleuroceridae (Pleurocera sp.; Figure 8M), Turritellidae (?Nairiella sp.; Figure 8N), Hemisinidae (Pyrgulifera boeckhi Pálfy, 1902; Figure 8O), and Melanopsidae (Melanopsis crastina [Vidal, 1874] and Campylostylus galloprovincialis [Matheron, 1843]); Figure 8P-Q. Neogastropoda is represented by the family Volutidae, with specimens assigned to Volutilithes sp. Figure 8S. Additionally, two heterobranchs taxa were identified: one assigned to Acteonellidae (Trochactaeon sp.; Figure 8T), and another classified as Acteonoidea indet. (Figure 8U) due to poor preservation.
All gastropods recorded from LVs 1-3 are known from brackish or freshwater environments in other regions (Zekeli, 1852; Pálfy, 1902; Bandel and Riedel, 1994), whereas the gastropods from LVs 4-5 are fully marine. Genera such as Volutilithes, Onkospira, Trochactaeon, and Nairiella documented in these upper levels are widely distributed in Cretaceous marine sediments (e.g., Akopian, 1976; Sohl and Kollmann, 1985; Bakayeva, 2011). This contrast strongly indicates a pronounced T shift in water salinity occurring between LV4 and LV3.
The gastropod assemblage from LVs 1-3 shows strong compositional affinity with the Santonian gastropod fauna of the Ajka Formation in Hungary, which is interpreted as deposited in lacustrine-lagoonal to lagoonal-brackish water environments (Czabalay, 1983). Bandel and Riedel (1994) further characterized the Ajka assemblage as indicative of freshwater to brackish conditions. This paleoenvironmental interpretation is supported by analogous fauna from the Lower Cretaceous Weald Clay Formation (Radley, 1999; Radley and Allen, 2012).
The marine gastropod fauna from LVs 4-5 warrants more detailed investigation, particularly focusing on smaller-sized faunal elements. Nevertheless, the occurrence of Trochactaeon sp. suggests affinities with warm, shallow-water Tethyan faunas (Sohl and Kollmann, 1985).
Bivalves. The bivalve assemblage is almost entirely confined to the lower part of the investigated section, specifically LVs 4-5. The greatest diversity is observed in the lowermost level (LV5), which yields at least seven species. This assemblage includes inoceramid bivalves (?Platyceramus sp.; Figure 8Y), the pinnid Pinna sp. (Figure 8R), an indetermined anomiid (Figure 8X), the gryphaeid oyster Pycnodonte sp. (Figure 8W), a cardiid, possibly Granocardium sp. (Figure 8AB); the hiatellid Panopea sp. (Figure 8D), and a radiolitoid rudist, Sauvagesia sp. (Figure 8AE). In the overlying LV4, species richness is notably reduced, with only three taxa recorded: a gryphaeid oyster (tentatively Exogyra sp.; Figure 8V), a cardiid (cf. Venericardia sp.; Figure 8AA), and a corbulid (possibly Corbula sp.; Figure 8AC).
Echinoids and Asteroids
 Echinoderms are represented by only three specimens within the investigated section. Two echinoid specimens, both poorly preserved and preserved solely as internal molds with minor remnants of the original tests, were recovered from LV5. Specimen NHMS 57412-LV5 displays a broad, flattened test with long, relatively narrow ambulacra (Figure 9A). In contrast, specimen NHMS 57411-LV5 is more globular in shape, with comparatively broader ambulacrum than in the other specimen (Figure 9B). These morphological distinctions suggest that the two individuals may represent two different species. Based on their overall test morphology, both specimens may be attributed to the family Hemiasteridae (J. Gallemí, personal comm., 14 January 2024). Asteroidea are represented in the Petrești deposits by two associated ossicles recovered from LV5 (specimen NHMS 57462). This specimen is poorly preserved, likely due to mechanical abrasion (Figure 9C-E).
Echinoderms are represented by only three specimens within the investigated section. Two echinoid specimens, both poorly preserved and preserved solely as internal molds with minor remnants of the original tests, were recovered from LV5. Specimen NHMS 57412-LV5 displays a broad, flattened test with long, relatively narrow ambulacra (Figure 9A). In contrast, specimen NHMS 57411-LV5 is more globular in shape, with comparatively broader ambulacrum than in the other specimen (Figure 9B). These morphological distinctions suggest that the two individuals may represent two different species. Based on their overall test morphology, both specimens may be attributed to the family Hemiasteridae (J. Gallemí, personal comm., 14 January 2024). Asteroidea are represented in the Petrești deposits by two associated ossicles recovered from LV5 (specimen NHMS 57462). This specimen is poorly preserved, likely due to mechanical abrasion (Figure 9C-E).
Corals
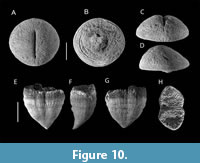 Although relatively rare, a few specimens belonging to the families Cunnolitidae and Phyllosmiliidae were identified in LV5. Six individuals of the genus Cunnolites Alloiteau, 1952, ranging in diameter from 25 to 55 mm, were successfully recovered (Figure 10A-D). These specimens are well preserved, exhibiting no evidence of basal attachment structures or overgrowth margins. While several subgenera and species of Cunnolites have been described (e.g., El-Asa’ad, 1990; Gameil et al., 2018), other authors have emphasized the taxonomic challenges posed by the high degree of morphological variability within the genus (Baron-Szabo, 1998, 1999), which complicates species-level identification.
Although relatively rare, a few specimens belonging to the families Cunnolitidae and Phyllosmiliidae were identified in LV5. Six individuals of the genus Cunnolites Alloiteau, 1952, ranging in diameter from 25 to 55 mm, were successfully recovered (Figure 10A-D). These specimens are well preserved, exhibiting no evidence of basal attachment structures or overgrowth margins. While several subgenera and species of Cunnolites have been described (e.g., El-Asa’ad, 1990; Gameil et al., 2018), other authors have emphasized the taxonomic challenges posed by the high degree of morphological variability within the genus (Baron-Szabo, 1998, 1999), which complicates species-level identification.
In addition to Cunnolites, two conical coral specimens were recovered from the same level, measuring 40 and 69 mm in height, respectively, and exhibiting an elliptical apical outline (Figure 10E-H). These specimens have been assigned to the genus Aulosmilia Alloiteau, 1952, based on morphological features (H. Löser, personal comm., 20 February 2024).
Insects
In addition to plant and reptile remains, the terrestrial biotic elements of the Petrești fossil assemblage also include certain fossil morphotypes interpreted as insect ichnofossils.
 Ovoid-shaped microfossils with prominent longitudinal ridges were recovered in association with mesofloral material. These structures range in length from 1.8 to 2.8 mm, and in width from 1.6 and 1.9 mm. The ridges covering their surface are triangular in cross-section, and conspicuous projections are present at both the apical and basal ends of the specimens (Figure 11A-B). Initially, microfossils of this morphology were attributed to the angiosperm seed species Spirellea kvacekii (Knobloch) Knobloch and Mai, 1983 (Knobloch and Mai, 1983, 1986). However, Heřmanová et al. (2013) argued that the terminal projections and absence of a micropyle scar are diagnostic of insect eggs. Consequently, this morphotype was reassigned to Knoblochia cretacea Heřmanová et al., 2013, an ichnotaxon related to insects. While the insect origin of K. cretacea is now widely accepted, its precise systematic placement remains uncertain. Similar remains have previously been reported from the Maastrichtian continental deposits of the Hațeg Basin (Bodor et al., 2014).
Ovoid-shaped microfossils with prominent longitudinal ridges were recovered in association with mesofloral material. These structures range in length from 1.8 to 2.8 mm, and in width from 1.6 and 1.9 mm. The ridges covering their surface are triangular in cross-section, and conspicuous projections are present at both the apical and basal ends of the specimens (Figure 11A-B). Initially, microfossils of this morphology were attributed to the angiosperm seed species Spirellea kvacekii (Knobloch) Knobloch and Mai, 1983 (Knobloch and Mai, 1983, 1986). However, Heřmanová et al. (2013) argued that the terminal projections and absence of a micropyle scar are diagnostic of insect eggs. Consequently, this morphotype was reassigned to Knoblochia cretacea Heřmanová et al., 2013, an ichnotaxon related to insects. While the insect origin of K. cretacea is now widely accepted, its precise systematic placement remains uncertain. Similar remains have previously been reported from the Maastrichtian continental deposits of the Hațeg Basin (Bodor et al., 2014).
Another morphotype identified in the Petreşti assemblage consists of elongated granules with a hexagonal cross-section, approximately 1.8 mm in length and 1.0 mm in width (Figure 11C). These microfossils have been identified as termite coprolites based on their strong morphological similarity to the fecal pellets of extant termites. They have been widely documented across global sites ranging from the Lower Cretaceous to the Pleistocene, typically attributed to Isoptera and described under the ichnospecies Microcarpolithes hexagonalis Vangerow 1954 (Colin et al., 2011; Heřmanová et al., 2021) or more generally as termite coprolites (Ősi et al., 2021). Fragments of M. hexagonalis are particularly abundant in the Petreşti deposits, especially within levels LV4 and LV3. This ichnotaxon has also been reported in large numbers from the Maastrichtian deposits of the Hațeg and Rusca Montană basins in Romania, as well as from the Santonian deposits of Iharkút, Hungary (Vasile et al., 2013).
3. Vertebrates
Fish
 Although relatively scarce, fish remains are present throughout all analyzed stratigraphic levels at the site. A total of 38 identifiable specimens — including teeth, otoliths, and scales — were recovered. Part of the material was assigned to the Class Chondrichthyes (cartilaginous fishes), representing both the Division Selachii, Order Lamniformes (aff. Lamniformes indet.; Figure 12A-C) and the Division Batomorphii, Family Rhinobatidae (Paratrygonorrhina amblysoda Kriwet et al., 2007; Figure 12D-H). In addition to chondrichthyans, representatives of the Class Actinopterygii (ray-finned fishes) were also identified. These include the orders Lepisosteiformes (Atractosteus sp.; Figure 12I-N, and Lepisosteus sp.; Figure S2 in the Supplementary materials), Aulopiformes ( Enchodus petrosus Cope, 1874; Figure 12O-Q), Elopiformes ( Paralbula casei Estes, 1969; Figure 12R-T), and Actinopterygii indet. (Figure 12U-W). Additionally, a new genus and species of basal percomorph fish is described from this locality by Trif and Schwarzhans as Paraplesiopoma transylvanica n. gen. et n. sp., representing Percomorpha incertae sedis (see description below).
Although relatively scarce, fish remains are present throughout all analyzed stratigraphic levels at the site. A total of 38 identifiable specimens — including teeth, otoliths, and scales — were recovered. Part of the material was assigned to the Class Chondrichthyes (cartilaginous fishes), representing both the Division Selachii, Order Lamniformes (aff. Lamniformes indet.; Figure 12A-C) and the Division Batomorphii, Family Rhinobatidae (Paratrygonorrhina amblysoda Kriwet et al., 2007; Figure 12D-H). In addition to chondrichthyans, representatives of the Class Actinopterygii (ray-finned fishes) were also identified. These include the orders Lepisosteiformes (Atractosteus sp.; Figure 12I-N, and Lepisosteus sp.; Figure S2 in the Supplementary materials), Aulopiformes ( Enchodus petrosus Cope, 1874; Figure 12O-Q), Elopiformes ( Paralbula casei Estes, 1969; Figure 12R-T), and Actinopterygii indet. (Figure 12U-W). Additionally, a new genus and species of basal percomorph fish is described from this locality by Trif and Schwarzhans as Paraplesiopoma transylvanica n. gen. et n. sp., representing Percomorpha incertae sedis (see description below).
SYSTEMATIC PALEONTOLOGY
Infraclass TELEOSTEI Müller, 1846
Subsection ACANTHOMORPHA Rosen, 1973
Division ACANTHOPTERYGII Artedi, 1738
Subdivision PERCOMORPHA Hay, 1903
Basal PERCOMORPHA incertae sedis
Genus PARAPLESIOPOMA n. gen. Trif and Schwarzhans
Figure 13
zoobank.org/54D1553C-7595-438C-9DBD-3645CC04CB7C
 Type species. Paraplesiopoma transylvanica n. sp. Trif and Schwarzhans
Type species. Paraplesiopoma transylvanica n. sp. Trif and Schwarzhans
Remarks. The otolith terminology follows Koken (1884).
Etymology. The genus name, Paraplesiopoma, refers to its morphological resemblance to the Late Cretaceous otolith-based genus Plesiopoma Schwarzhans, 2010.
Diagnosis. Fossil otolith-based genus tentatively associated with basal Percomorpha. Otoliths small (maximum not exceeding 3.5 mm in length), elongate oval in shape with rounded posterior tip and projecting, rounded rostrum. Otolith length to height ratio (OL/OH) 1.40-1.75, height to thickness ratio (OH/OT) 2.2-2.7. Antirostrum and excisura minute. Inner face distinctly convex. Sulcus long, clearly divided into ostium and cauda. Ratio cauda length (CaL) to ostium length (OsL) 1.40-1.55; ratio ostium height (OsH) to cauda height (CaH) 1.5-2.2. Ostium open anterior-dorsally, ventrally expanded, and with clearly defined ostial colliculum; cauda slightly flexed along entire course; no ventral furrow.
Comparison. Paraplesipoma exhibits an otolith morphology reminiscent of several percomorph, particularly eupercarian, taxa distinguished by a broad, short ostium and a slender, slightly curved cauda. This configuration, referred to as “heterosulcoid” by Schwarzhans (1978) is widespread among numerous unrelated teleost clades. However, Paraplesiopoma lacks additional diagnostic features that would allow for a more precise taxonomic placement beyond basal Percomorpha. Its generalized oval shape and absence of a ventral furrow are considered plesiomorphic, paralleling conditions observed in more basal teleost groups such as Elopiformes, Albuliformes, and Aulopiformes (see Schwarzhans, 2018 for comparative data). For instance, Elopiformes otoliths tend to be slenderer with a less pronounced ventral expansion of the ostium; Albuliform otoliths typically display a diagonally positioned sulcus (e.g., in Pterothrissidae) or a sharply curved caudal tip (e.g., in Albulidae). In Aulopiformes such as Paraulopidae, the dorsal margin of the ostium is characteristically expanded both upward and backward. It cannot be excluded, however, that in a phylogenetic sense Paraplesiopoma is of sub-percomorph affinities.
As previously noted, Paraplesiopoma closely resembles otoliths of the fossil basal percomorph otolith-based genus Plesiopoma Schwarzhans, 2010 known from several species from Coniacian to early Paleocene (Schwarzhans, 2010, 2012). Plesiopoma was initially placed within the family Acropomatidae, but has more recently been reinterpreted as Percomorpha incertae sedis (Schwarzhans et al., 2024). The two genera are differentiated by the presence of a ventral furrow in Plesiopoma — a derived percomorph character, and by its ostium and cauda being of nearly equal length.
Species. The type species is Paraplesiopoma transylvanica sp. nov. from the Campanian of Romania. Additional species include P. bagassiana (Nolf, 2003), originally described as “genus Acropomatidarum bagassianus”, and P. santoniana (Nolf, 2003), originally described as “genus Haemulidarum santonianus”, both from the Santonian of Spain.
Paraplesiopoma transylvanica n. sp.
Trif and Schwarzhans
Figure 13
zoobank.org/D9BC31EB-7B86-423E-A65E-ABC4A3BA9A62
Type locality. Petrești, Alba County, Romania.
Stratum typicum. Level 4 (see Figure 2 and Figure 3A).
Age. Campanian.
Etymology. Named after the Transylvanian Basin.
Holotype. NHMS 57413/LV4 (Figure 13A-D).
Paratypes. Specimens NHMS 57414 (Figure 13E), 57415 (Figure 13F-G), 57416 (Figure 13H), and 54718 (Figure 13I-J), all originating from the same stratigraphic level and locality as the holotype.
Diagnosis. OL:OH = 1.55-1.65; OH:OT = 2.4-2.7. Maximal observed length ca 3.3 mm. Dorsal and ventral rims regularly curved without angles. Ostium distinctly shorter and wider than cauda, slightly upward directed, mainly ventrally expanded; CaL:OsL = 1.4-1.5; OsH:CaH = 1.5-1.8.
Description. Otoliths are small, elongate-oval, and moderately thick, with maximum observed length reaching 3.3 mm (holotype: 2.75 mm long). The dorsal rim is regularly curved, slightly and irregularly undulating. The ventral rim is regularly curved, smooth or with fine crenulation, particularly in small specimens (Figure 13I-J); in the larger specimens the crenulation is less intense and primarily along preventral rim (Figure 13H). The anterior tip displays a massive, moderately pointed to rounded rostrum that is positioned slightly above the otolith axis; the rostrum length is about 15-20% of OL. The antirostrum and excisura are minute. The posterior rim is rounded, positioned slightly inferior to the level of the rostrum.
The inner face is distinctly convex with a supramedially positioned, long, somewhat deepened sulcus. The ostium is anteriorly opening to the predorsal rim, its relatively flat dorsal margin is about half the length of the deeply expanded ventral margin; the ostium is slightly upward inclined at 7-10°, which is best visible along the straight central part of the ventral margin of the ostium. The cauda is longer than the ostium and distinctly narrower, slightly flexed along the entire course and reaches close to the posterior rim of the otolith. CaL:OsL = 1.4-1.5; OsH:CaH = 1.5-1.8. The dorsal depression is long, narrow, ventrally bound by a distinct crista superior towards the sulcus, and its dorsal margin is gradual, indistinct. There is no ventral furrow discernible. The outer face is flat to convex, relatively smooth except for the presence of occasional short radial furrows along its ventral rim.
Discussion. Paraplesiopoma transylvanica is the only otolith-based species identified in the transitional brackish deposits of Petrești. Its occurrence suggests adaptation to such marginal environment. In contrast, the stratigraphically slightly older Santonian congeners, P. santoniana and P. bagassiana a re known from fully marine deposits (Nolf, 2003). Paraplesiopoma santoniana differs from P. transylvanica in the more pointed posterior tip of the otolith and the slightly downward inclined cauda (versus straight and slightly flexed). Paraplesiopoma bagassiana is more compact than P. transylvanica (OL:OH = 1.4-1.6 vs 1.55-1.65) and thicker (OH:OT = 2.2 vs 2.4-2.7). Paraplesiopoma transylvanica differs from both its congeners in the dorsal margin of the ostium being less expanded, which is reflected in a lower OsH:CaH ratio (1.5-1.8 measured from drawings in Nolf, 2003 versus 1.9-2.3 in the Petrești specimens), and in the ostium being slightly upward inclined (versus horizontal in the new species).
A comprehensive reconstruction of the ichthyofauna at Petreşti remains challenging due to the low species richness. Nonetheless, comparative evidence offers insights into paleoecological conditions. Paratrygonorrhina amblysoda (level LV4), previously recorded only in the Spanish and French Pyrenees, is interpreted as a nektobenthic ray associated with marginal littoral environments (Gheerbrant et al., 1997; Kriwet et al., 2007). Similarly, Paralbula casei (LV4 and LV5) and Enchodus cf. petrosus (LV4) are known from shallow marine and coastal plain environments, often linked to fluvially influenced estuarine or deltaic systems (Becker et al., 2010; Schubert et al., 2017). The indeterminate aff. Lamniformes indet. material from LV4 is too fragmentary for ecological inference.
The presence of the gars (Atractosteus sp. and Lepisosteus sp.) in levels LV1, LV2, and LV3 may indicate a stronger continental influence. At least one of these genera is already documented in Maastrichtian continental deposits from Romania (e.g., Csiki et al., 2008; Solomon et al., 2022b).
For additional taxonomic details, measurements, discussions, and supporting figures, refer to Supplementary materials, Section C.
Herpetofauna
 The herpetofaunal assemblage recovered from various stratigraphic levels is relatively limited in both diversity and abundance. The identified material includes two urostyles, several incomplete jaw elements, a fragment of a vertebral neural arch, and a number of isolated teeth. These remains were assigned to representatives of amphibians (Anura), and reptiles (Squamata and Crocodylomorpha). The amphibian remains were attributed to the anuran clade Alytidae (?Alytidae indet., specimen NHMS 57446, levelLV1), represented by a urostyle characterized by distinctly bipartite cotyles and well-developed transverse processes (Figure 14A-F). Additional anuran remains — comprising a urostyle, a partial dentary, and a vertebral fragment — are referred to Anura indet. (Figure 14G-O).
The herpetofaunal assemblage recovered from various stratigraphic levels is relatively limited in both diversity and abundance. The identified material includes two urostyles, several incomplete jaw elements, a fragment of a vertebral neural arch, and a number of isolated teeth. These remains were assigned to representatives of amphibians (Anura), and reptiles (Squamata and Crocodylomorpha). The amphibian remains were attributed to the anuran clade Alytidae (?Alytidae indet., specimen NHMS 57446, levelLV1), represented by a urostyle characterized by distinctly bipartite cotyles and well-developed transverse processes (Figure 14A-F). Additional anuran remains — comprising a urostyle, a partial dentary, and a vertebral fragment — are referred to Anura indet. (Figure 14G-O).
The reptilian material includes both squamates and crocodylomorphs. A partially preserved lower jaw was attributed to Scleroglossa indet., potentially representing a scincoid taxon (Figure 14P-R). The crocodylomorph remains are represented by a possible ziphosuchian, tentatively identified as cf. Doratodon sp. (Figure 14S-W), and an isolated tooth is attributed to cf. Allodaposuchidae indet. (Figure 14X-Z). Additionally, the presence of non-avian theropod dinosaurs is inferred based on the occurrence of eggshell fragments (Figure 15A-F), although no osteological remains were identified.
 The amphibian material provides evidence for the presence of at least two distinct anuran taxa: ?Alytidae and Anura indet. Alytidae is an anuran clade with many primitive features that includes a group of species associated with aquatic habitats (Discoglossus and Latonia) and another stoutly built group with semi-fossorial habits (Alytes). Their natural habitats are temperate forests, Mediterranean-type shrubby vegetation, rivers, freshwater marshes, freshwater springs, and rocky areas (Sanchiz, 1998; Dubois, 2005; Blackburn and Wake, 2011; Beukema et al., 2013).
The amphibian material provides evidence for the presence of at least two distinct anuran taxa: ?Alytidae and Anura indet. Alytidae is an anuran clade with many primitive features that includes a group of species associated with aquatic habitats (Discoglossus and Latonia) and another stoutly built group with semi-fossorial habits (Alytes). Their natural habitats are temperate forests, Mediterranean-type shrubby vegetation, rivers, freshwater marshes, freshwater springs, and rocky areas (Sanchiz, 1998; Dubois, 2005; Blackburn and Wake, 2011; Beukema et al., 2013).
The reptilian assemblage includes both lizard and crocodylomorph remains. The lizard material is represented by a specimen assigned to Scleroglossa indet., potentially within the clade Scincoidea. The crocodylomorphs appear to comprise two taxa. Based on the typical tooth morphology (labiolingually flat, ziphodonty), a Doratodon -like crocodylomorph can be identified (note, however, that caution is needed — identification of ziphosuchians and atoposaurids below family level, based on a single isolated tooth, might be doubtful, because several distinct tooth morphotypes can be present in a single individual. Moreover, besides the zipchosuchian Doratodon, at least two taxa of atoposaurids are present in the Transylvanian landmass (e.g., Venczel and Codrea, 2019). The second tooth is rather generalized in morphology. The crenulation of the second tooth together with the general morphology resemble a tooth from the posterior region of the jaw of a semi-aquatic eusuchian, being comparable with members of Allodaposuchidae. Thus, it is tentatively assigned to cf. Allodaposuchidae indet. In this group, the enamel surface usually bears some apicobasal coarse ridges (e.g., Narváez et al., 2020; note, however, that it can be smooth in some cases, this is a variable character). Allodaposuchidae is a clade of basal eusuchians known from the Late Cretaceous (Campanian and Maastrichtian) of Europe, being an endemic group of the European archipelago (Narváez et al., 2015, 2016, 2020; Blanco and Brochu, 2017) and even the type material of Allodaposuchus precedens is previously reported from Romania (the lower Maastrichtian of Vălioara) (Nopcsa, 1928; Narváez et al., 2020).
Doratodon is an enigmatic extinct crocodyliform that inhabited the western Tethyan archipelago during the Late Cretaceous, corresponding to present-day Romania, Hungary, southern France, and Spain. Members of this genus were relatively small-bodied, characterized by ziphodont dentition and inferred to have had a terrestrial lifestyle with adaptations consistent with active predation (Martin et al., 2006, 2010; Martin, 2007; Vasile, 2008; Vasile and Csiki, 2011; Rabi and Sebők, 2015).
Modern crocodiles are typically confined to tropical and subtropical regions, largely due to their limited reproductive success in colder climates (Alderton, 1991). As such, fossil representative of Crocodyliformes serve as valuable proxies for reconstructing paleoclimatic and paleoecological conditions (Berg, 1965; Markwick, 1998). Analyses of extant crocodilian distribution suggest that temperature is a critical factor governing their global range, with the minimum average annual temperature for current species being approximately 14.2°C (Markwick, 1998). It is important to emphasize, however, that this threshold applies to crown-group crocodilians. The ecological tolerances and niche requirements of the more diverse Mesozoic crocodylomorphs were likely broader, and may have differed substantially from those of modern forms (see the discussion in Mannion et al. (2015) regarding long-term biodiversity decline). Carvalho et al. (2010) documented spatial and temporal patterns in the distribution of terrestrial Crocodyliformes throughout the Cretaceous, noting an expansion of medium- to large-bodied taxa during the Late Cretaceous. These taxa were often active terrestrial predators, and their distribution appears to have been significantly influenced by paleotemperature. Notably, large non-marine radiations of notosuchians and eusuchians are well documented during this interval (e.g., Carvalho et al., 2010; Bronzati et al., 2015). However, a decline in crocodyliforms biodiversity is observed from the Campanian to the Maastrichtian in both Europe and North America, which may reflect a long-term global cooling trend during the Late Cretaceous (Mannion et al., 2015). Nevertheless, de Celis et al. (2018) proposed that additional biotic and abiotic factors — beyond paleotemperature alone — may have significantly influenced crocodyliform macroevolution during the Mesozoic, suggesting a more complex evolutionary narrative than previously recognized.
For comprehensive descriptions, morphometric data, taxonomic revisions, and supplementary visual material, refer to Supplementary materials, Section D.
Eggshells
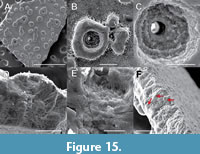
A limited number of eggshell fragments exhibiting comparable morphological characteristics were recovered from the LV1 and LV2 levels (specimens NHMS 57456-57458). The external surface of these fragments is sculptured, displaying moderately spaced tubercles with irregular — occasionally subcircular — outlines (Figure 15A), imparting a nodular texture to the shell surface. Most tubercles exhibit a relatively smooth surface; however, some display minute pitting, and a subset — typically the larger tubercles — are perforated by funnel-shaped channels (Figure 15B-E). The diameter of these perforations ranges from 2.2 to 4.1 μm. The thickness of the eggshells, measured in the intertubercular regions, ranges from 173 to 247 μm. A distinctive feature observable in radial sections of all three specimens is the presence of tabular structures oriented parallel to the eggshell surface (Figure 15F). The functional or structural significance of these features remains uncertain at this stage (Figure 15F). Notably, no vertical zoning, layering, or columnar structures are evident within the examined samples.
Remarks. Eggshells exhibiting similar morphological characteristics have previously been reported as “geckonoid eggshell fragment” or “geckoid eggshell fragments” from the Maastrichtian deposits of the Hațeg and Rusca Montană basins (Vasile and Csiki, 2011; Csiki-Sava et al., 2016). Other thin-shelled eggs from the Hațeg Basin, sometimes referred to as the “thin egg type” (Codrea et al., 2002), have also been interpreted as having geckkotan affinities and were assigned to the geckoid morphotype sensu Garcia (2000), based on the morphological classification proposed therein. Additionally, Maastrichtian eggshells from the Transylvanian Basin (Oarda de Jos locality) were tentatively identified as ?Pseudogeckoolithus (Codrea et al., 2010a). However, due to the lack of detailed descriptions and illustrative documentation, it remains uncertain whether these latter two reports pertain to the same morphotype.
Recent investigations by Choi et al. (2020) on similar eggshells from Upper Cretaceous deposits of Europe have revealed distinct crystallographic features, particularly in the orientation of calcite grain axes. These microstructural criteria have proven to be useful in differentiating fossil and extant eggshells that share a superficially similar outer nodular ornamentation. Based on these crystallographic analyses, Choi et al. (2020) concluded that the so-called “geckoid” eggshells from the European Upper Cretaceous are more accurately attributed to maniraptoran theropods, and are assignable to the oogenus Pseudogeckoolithus. At present, two, and possibly three, oospecies within this genus are recognized from the Upper Cretaceous deposits of Romania (Choi et al., 2020). Given that crystallographic analysis was not conducted in the present study, and considering the limited number of specimens and insufficient data to assess variability in surface ornamentation, we tentatively assign our eggshell material to Pseudogeckoolithus sp.
For a concise summary of the taxonomic composition and stratigraphic distribution of fossils across the five levels of the Petrești section refer to Table 1.
PALEOENVIRONMENTAL PERSPECTIVES
Based on the fossil assemblages recovered from this transitional sequence, three distinct latest Cretaceous paleoenvironments can be inferred, each reconstructed to varying degrees of detail: marine, brackish-estuarine, and continental. A gradual increase in continental influence is evident from the lowermost level (LV5) to the uppermost level (LV1) (see Figure 3).
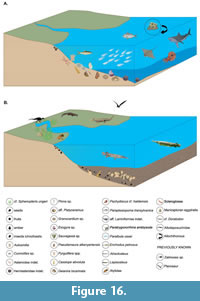 Two of these paleoenvironments — the marine and brackish-estuarine settings — are illustrated in Figure 16A-B.
Two of these paleoenvironments — the marine and brackish-estuarine settings — are illustrated in Figure 16A-B.
The marine paleoenvironment (Figure 16A) is interpreted as a shallow coastal setting characterized by a mixed assemblage of marine taxa, indicative of relatively stable salinity conditions. The presence of stenohaline organisms such as ammonites (e.g., Pachydiscus), echinoids, and asteroids supports this interpretation. However, the accompanying ichthyofauna — including Paratrygonornina, Paralbula, and Echodus — suggests fluvial influence, consistent with a nearshore environment subject to terrestrial input. The presence of plant debris and amber further corroborates this interpretation. This marine assemblage is best represented at stratigraphic level LV5. The subsequent level, LV4, likely marks the onset of environmental transition.
Sedimentation rates at LV5 appear to have been relatively high. Evidence supporting this interpretation includes the occurrence and preservation of the solitary coral Cunnolites, a cosmopolitan genus typically associated with tropical to warm-temperate environments. Cunnolites is commonly found in non-reefal settings, favoring sandy to sandy-silty substrates — conditions inferred for the Petreşti section. Notably, the absence of basal pedestals, macroboring traces, and overgrowth features on our Cunnolites specimens implies rapid sedimentation, as such features typically form under conditions of slower deposition that permit vertical coral growth and substrate colonization (Sanders and Baron-Szabo, 2005, 2008). In addition, the relatively small size of our specimens (maximum diameter of 55 mm) compared to the known maximum size for the taxon (~400 mm) suggests abbreviated lifespans, likely attributable to the stress of high-sedimentation rates (Sanders and Baron-Szabo, 2005).
The progressive reduction in bivalve diversity observed upward through the Petreşti section is interpreted as a response to increasingly marginal marine conditions. Fully marine assemblages appear to be restricted to the lowermost beds, particularly those exposed at level LV5. In contrast, levels LV4, LV3, and LV1 yield assemblages dominated by more tolerant taxa, such as corbulid bivalves. The presence of corbulids in these strata may indicate low sedimentation rates and episodic influence from low-salinity waters, as members of this group are known to tolerate brackish conditions better than most other marine bivalves (Lewy and Samtleben, 1979). The bivalve assemblage is notably depauperate in epifaunal taxa, with the absence of typically common forms such as arcoids, pectinids, and mytilids. Additionally, several infaunal groups frequently encountered in coeval marine deposits — such as crassatellids and veneroids — are also absent or rare. The recorded diversity among infaunal bivalves may reflect a combination of low sedimentation rates and suboptimal conditions within the substrate, possibly including fluctuating salinity, low oxygen levels, or unstable sedimentary regimes.
The occurrence of Sauvagesia sp. within this assemblage is considered allochthonous. The valves are broken, compressed, and show signs of erosion, suggesting post-mortem transport from a different depositional setting. It is likely that these specimens were introduced into the Petreşti environment by wave or current action (L. Săsăran, pers. comm., 06 December 2024).
The occurrence of corbulid bivalves in LV4 at Petrești may indicate periodic influence from low-salinity waters — conditions that corbulids are known to tolerate more effectively than most other marine bivalves (Lewy and Samtleben, 1979). The recorded low diversity among infaunal bivalves may also reflect reduced sedimentation rates and potentially unfavorable substrate conditions.
The brackish, estuarine paleoenvironment reconstructed for portions of the Petrești section (Figure 16A) is characterized by the complete absence of ammonites and corals, indicating a departure from fully marine conditions. The gastropod assemblage exhibits a marked shift in taxonomic composition, with Pyrgulifera and Campylostylus often forming dense accumulations of shells. The bivalve component is notably absent in these facies. The localized concentration of mollusk remains suggests episodic high-energy depositional event, likely associated with increased freshwater influx and river discharge, which may have transported and concentrated shells during periods of enhanced hydrodynamic activity.
An increasing terrestrial influence up the section is also reflected in the ichthyofaunal assemblage. In the upper stratigraphic levels (LV3-LV1, Lepisosteiformes — particularly Atractosteus and Lepisosteus — replace the fish taxa that dominate the lower, marine levels (e.g., Echodus, Paralbula, and the chondrichthyan taxa). This faunal turnover coincides with a marked increase in the abundance of plant debris and amber, particularly in levels LV3 through LV1, supporting the interpretation of a brackish to estuarine depositional environment for these strata.
The nature of the continental paleoenvironment at Petreşti remains relatively poorly constrained, as it is currently inferred from sparse and predominantly allochthonous remains. The occurrence of the fern Sphenopteris ungeri suggests the presence of riparian vegetation, a hypothesis further supported by the associated reptilian assemblage, which includes crocodyliforms. Additional evidence of terrestrial conditions is provided by fragmentary remains of frogs and lizards, although the finds are limited and do not yet allow for a detailed ecological reconstruction. The floral assemblage, primarily composed of seeds and fruits, is characterized by relatively low taxonomic diversity when compared to other Cretaceous floras (e.g., Knobloch and Mai, 1986). However, it is broadly comparable in richness to the Maastrichtian mesoflora of the Hațeg Basin (Lindfors et al., 2010). Expanded sampling and systematic analysis of plant material would be necessary to obtain a more comprehensive picture of mesofloral diversity and composition. Previous discoveries from the region (e.g., Vremir 2010; Vremir et al., 2015) provide additional support for the presence of a diverse terrestrial ecosystem during the latest Cretaceous, despite the taphonomic and sampling limitations currently affecting the Petreşti record.
To support the interpretative framework for the ecological transition from marine to brackish environments, a basic statistical analysis was conducted, incorporating cluster analysis and the construction of a species distribution chart (for the data used refer to Supplementary materials, Section E and F).
Cluster analysis based on Jaccard similarity index and employing the Unweighted Pair Group Method with Arithmetic Mean (UPGMA) was performed to assess the relationships among stratigraphic sampling levels, using presence/absence data of recorded taxa. Given the potential of calcareous nannoplankton to obscure genuine taxonomic and ecological patterns — particularly within dynamic marine-estuarine transitional zones — analyses were carried out on two separate datasets to distinguish their potential influence.
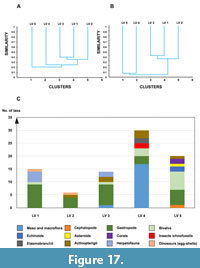 The initial analysis, encompassing the complete assemblage of taxa identified at the Petrești site, yielded a main cluster with a basal similarity of 0.21. Within this cluster, a distinct subcluster was formed by level LV4 (similarity: 0.26), while a second subcluster included levels LV1, LV2, and LV3, which exhibited pairwise similarities ranging from 0.36 to 0.39 (Figure 17A).
The initial analysis, encompassing the complete assemblage of taxa identified at the Petrești site, yielded a main cluster with a basal similarity of 0.21. Within this cluster, a distinct subcluster was formed by level LV4 (similarity: 0.26), while a second subcluster included levels LV1, LV2, and LV3, which exhibited pairwise similarities ranging from 0.36 to 0.39 (Figure 17A).
Although a superficial interpretation of these clustering results might suggest a relatively high degree of faunal similarity — and, by implication, a homogeneous ecological composition across the sampled levels — this interpretation is likely misleading. The apparent homogeneity is primarily attributed to the inclusion of calcareous nannoplankton in the dataset. In marine-estuarine transitional environments, the distribution pattern of nannoplankton may not accurately reflect local ecological gradients. These organisms, being susceptible to advective transport due to water mass mixing, can be episodically introduced into brackish or even freshwater-influenced settings. As a result, their presence may mask ecologically meaningful signals associated with more environmentally sensitive taxa.
The second dataset, which excludes calcareous nannoplankton taxa, produced a clustering pattern that diverges significantly from that obtained in the initial analysis. Although the overall topology of the resulting dendrogram may appear similar to that of the complete dataset, the distinctions revealed are considered to offer deeper insights into the underlying ecological and biological dynamics.
In this refined analysis, levels LV1, LV2, and LV3 form a coherent cluster, characterized by relatively high Jaccard similarity indices, ranging from 0.33 to 0.40 (Figure 17B). This pattern indicates considerable taxonomic overlap and implies relatively stable environmental conditions across these stratigraphic intervals. By contrast, levels LV4 and LV5 diverge much earlier in the dendrogram, at substantially lower similarity values (Jaccard index >0.1), reflecting a faunal composition markedly different from that of the LV1-LV3 cluster. Although LV4 and LV5 form a weakly defined subgroup, the low degree of similarity suggests limited taxonomic overlap, with their apparent association largely driven by the shared presence of a single species.
The hierarchical structure of the dendrogram suggests a two-step faunal turnover: first, a compositional shift between LV5 to LV4, followed by a more pronounced transition between LV4 and the LV1-LV3 cluster. The tight clustering of levels LV1-LV3 is interpreted as indicative of a phase of relative palaeoecological stability, in the uppermost part of the Bozeș Formation.
Integration of the cluster analysis results with the taxonomic distribution patterns, as depicted in the species distribution chart (Figure 17C), supports the interpretation that the grouping of levels LV1-LV3 is primarily driven by the consistent occurrence of gastropod and bivalve taxa. In contrast, the assemblages of LV4 and LV5 exhibit greater taxonomic heterogeneity, incorporating mesofloral remains, cephalopods, asteroids, corals, echinoids, and fish. This broader faunal diversity likely accounts for the distinct separation of LV4 and LV5 in the cluster analysis, and is interpreted as reflective of more variable and potentially transitional environmental conditions at these stratigraphic levels.
CONCLUSIONS
This study provides critical insights into the paleoenvironmental and paleobiological evolution of the upper Campanian Bozeş Formation at Petreşti, within the southern Transylvanian Basin. The main conclusions are as follows:
1. A major paleoenvironmental shift from marine to brackish conditions has been precisely documented and correlated with the stratigraphic horizon LV4. While the transition had been recognized in prior research, our study provides a more detailed documentation and confirms its stratigraphic position through a well-characterized fossil assemblage.
2. The study presents important new paleontological data that refine and expand the known faunal and floral composition of the upper estuarine deposits of the Campanian Bozeș Formation at Petrești, located in the southern Transylvanian Basin.
3. The environmental transition is interpreted as gradual, progressing from a marine to an estuarine setting with increasing continental influence. There is no evidence supporting an abrupt or catastrophic shift.
4. A revised taxonomic reassessment of the ammonite assemblage has led to the reclassification of specimens previously assigned to Pachydiscus neubergicus and P. colligatus which are now referred to as Pachydiscus cf. haldemsis, based on newly collected and better-preserved material.
5. The diversity of gastropods and bivalves, formerly limited to a few genera such as Cerithium, Pyrgulifera, and Barbotella, has been substantially revised, with at least 30 taxa now recognized. This expanded diversity underscores the need for continued systematic investigation in the region.
6. This study documents, for the first time within the Bozeş Formation, the occurrence of echinoids, asteroids, plant seeds, fruits, amber, and the first recorded impressions of plants associated with the brackish transition. Additional significant discoveries include insect eggs and coprolites preserved alongside amber.
7. The identification of vertebrate remains — including fishes, anurans, scleroglossan lizards, crocodyliforms, and maniraptoran theropods — further augments the vertebrate diversity previously known from the formation. Notably, the Anura and Scleroglossa specimens represent the oldest known Cretaceous occurrences of these clades in Romania.
8. A new percomorph bony fish taxon, Paraplesiopoma transylvanica n. gen. n. sp., is described herein.
9. From a paleoenvironmental standpoint, a marked compositional shift is evident between LVs 4-5 and LVs 1-3. percomorph The fossil assemblage in LV4 and LV5, dominated by marine invertebrates and vertebrates (including ammonites, gastropods, bivalves, starfish, echinoids, and fish), is indicative of normal salinity conditions, whereas the assemblage in LVs 1-3 reflects a transition toward brackish and continental influence. This is evidenced by a reduction in bivalve diversity (from over seven to three species) and the appearance of terrestrial indicators such as seeds, fruits, amber, and fragments of carbonized wood, although these occur in relatively low abundance.
10. Several avenues for future research are proposed, including more detailed investigations of meso- and macrofloral assemblages, comprehensive analyses of amber, and refined taxonomic work on gastropod and bivalve assemblages.
11. Collectively, this study significantly advances our understanding of the late Campanian biodiversity in marginal marine environments, highlighting the ecological dynamics at the marine-terrestrial interface during the Late Cretaceous in the Transylvanian Basin.
ACKNOWLEDGMENTS
We would like to express our sincere gratitude to D. Vasilyan (JURASSICA Museum, Switzerland) and J. Martin (Université Lyon, France) for their helpful advice. S.J. acknowledges the significant contributions of M. Machalski (Warszawa, Poland) and C. Ifrim (Eichstätt, Germany) for their expert insights on ammonite identification and literature guidance. We extend our thanks to W.J. Kennedy (Oxford University Museum of Natural History, UK) for confirming the ammonite identifications. We are also indebted to H. Löser (Sonora, Mexico) for identifying the genus Aulosmilia, to J. Gallemí (Barcelona, Spain) for identifying Echinoidea specimens as cf. Hemiasteriae, and to L. Săsăran for identifying the Sauvagesia radiolitoid. We would like to acknowledge the valuable contributions of the Natural History Museum, Sibiu volunteers, A. Sandu, D. Cocîrlea, and C. Nemeș, as well as the museum curators M. Stănciugelu, M. Sasu, and S. Țicu for their diligent efforts in collecting, transporting, and washing the sediments from Petrești. We also thank Ș. Olaru from the Faculty of Biology, “Alexandru Ioan Cuza” University of Iași, for the SEM images of the eggshells. The two anonymous reviewers are thanked for their suggestions.
Funding Sources
For Andrej Čerňanský this research was supported by the Scientific Grant Agency of the Ministry of Education of Slovak Republic and Slovak Academy of Sciences, Grant Nr. 1/0160/24.
For Andrzej Kaim, Sofia Bakayeva, Krzysztof Hryniewicz this research was supported by the National Science Centre, Poland, grant number 2018/31/B/ST10/03415.
Sofia Bakayeva was also supported by the International Education’s Scholar Rescue Fund Fellowship.
Nicolae Trif, Roxana Pirnea, Werner Schwarzhans, Ramona Bălc, and Sreepat Jain did not receive any specific grants from funding agencies in the public, commercial, or not-for-profit sectors.
Author Contributions
NT: Conceptualization, Methodology, Investigation, Resources, Supervision, Visualization, Writing — Original Draft, Writing — Review and Editing; RP: Conceptualization, Investigation, Visualization, Writing — Original Draft, Writing — Review and Editing; AČ: Investigation, Visualization, Writing — Original Draft, Writing — Review and Editing, Funding acquisition; WS: Investigation, Methodology, Visualization, Writing — Original Draft, Writing — Review and Editing; RB: Investigation, Methodology, Visualization, Writing — Original Draft, Writing — Review and Editing; SJ: Investigation, Visualization, Writing — Original Draft, Writing — Review and Editing; AK: Investigation, Visualization, Writing — Original Draft, Writing — Review and Editing, Funding acquisition; SB: Investigation, Writing — Original Draft, Writing — Review and Editing, Funding acquisition; KH: Investigation, Visualization, Writing — Original Draft, Writing — Review and Editing.
REFERENCES
Adl, S., Girard, V., Breton, G., Lak, M., Maharning, A., Mills, A., Perrichot, V., Trionnaire, M., Vullo, R. and Neraudeau, D. 2011. Reconstructing the soil food web of a 100 million-year-old forest: The case of the mid-Cretaceous fossils in the amber of Charentes (SW France). Soil Biology and Biochemistry, 43:726–735.
https://doi.org/10.1016/j.soilbio.2010.12.003
Akopian, V.T. 1976. Pozdniemielowyje gastropody Armianskoj SSR [Late Cretaceous gastropods of Armenia SSR]. AN Armianskoj SSR, Yerevan. (In Russian)
Alderton, D. 1991. Crocodiles & Alligators of the World. Blandford Publishing, London.
Alloiteau, J. 1952. Madreporaires post-Paleozoiques, p. 639–782. In Piveteau, J. (ed.), Traite de Paleontologie. Masson, Paris.
Arkhangelsky, A.D. 1912. Upper Cretaceous deposits of east European Russia. Materialien zur Geologie Russlands, 25:1–631.
Artedi, P. 1738. Ichthyologia sive opera omnia de piscibus, scilicet: Bibliotheca ichthyologica. Philosophia ichthyologica. Genera piscium. Synonymia specierum. Descriptiones specierum. Omnia in hoc genere perfectiora, quam antea ulla. Lugduni Batavorum, apud Conradum Wishoff.
https://doi.org/10.5962/bhl.title.111064
Bakayeva, S. 2011. Pizniokreydovi cherevonohi moliusky Volyno-Podillia [Late Cretaceous gastropods from Volyno-Podillia]. Naukova dumka, Kyiv. (In Ukrainian)
Bălc, R. and Chira, C. 2002. The preliminary study of the Upper Cretaceous calcareous nannofossils from Alba Iulia area, Transylvania. Studia Universitatis Babeş-Bolyai, Geologia, 1:97–111.
Bălc, R., Suciu-Krausz, E., and Borbei, F. 2007. Sedimentary deposition of Bozeş Formation (Apuseni Mts., Romania) – detrital zircon dating and micropaleontological ages. Studia Universitatis Babeş-Bolyai, Geologia, 52:37–43.
Bălc, R., Silye, L., and Zaharia, L. 2012. Calcareous nannofossils and sedimentary facies in the Upper Cretaceous Bozeş Formation (Southern Apuseni Mountains, Romania). Studia Universitatis Babeș-Bolyai, Geologia, 57:23–32.
https://doi.org/10.5038/1937-8602.57.1.3
Bălc, R., Bindiu-Haitonic, R., Kövecsi, S.A., Vremir, M., Ducea, M., Csiki-Sava, Z., Țabără, D., and Vasile, Ș. 2024. Integrated biostratigraphy of Upper Cretaceous deposits from an exceptional continental vertebrate-bearing marine section (Transylvanian Basin, Romania) provides new constraints on the advent of “dwarf dinosaur” faunas in Eastern Europe. Marine Micropaleontology, 187:102328.
https://doi.org/10.1016/j.marmicro.2023.102328
Bandel, K. and Riedel, F. 1994. The Late Cretaceous gastropod fauna from Ajka (Bakony Mountains, Hungary): a revision. Annalen des Naturhistorischen Museums in Wien A, 96:1–65.
Baron-Szabo, R.C. 1998. A new coral fauna of the Campanian from north Spain (Torallola village, Prov. Llèida). Geologisch-Paläontologische Mitteilungen Innsbruck, 23:127–191.
Baron-Szabo, R.C. 1999. Taxonomy of Upper Cretaceous scleractinian corals of the Gosau Group (Weissenbachalm, Steiermark, Austria). Abhandlungen der Geologischen Bundesanstalt, 56:441–464.
Becker, M.A., Mallery, C.S., and Chamberlain, J.A. 2010. Osteichthyans from an Arkadelphia Formation–Midway Group lag deposit (Late Maastrichtian–Paleocene), Hot Spring County, Arkansas, U.S.A. Journal of Vertebrate Paleontology, 30:1019–1036.
https://doi.org/10.1080/02724634.2010.483603
Berg, D. 1965. Krokodile als Klimazeugen. Geologische Rundschau, 54:328–333.
Betancur-R, R., Wiley, E.O., Arratia, G., Acero, A., Bailly, N., Miya, M., Lecointre, G., and Orti, G. 2017. Phylogenetic classification of bony fishes. BMC Evolutionary Biology 17:162.
https://doi.org/10.1186/s12862-017-0958-3
Beukema, W., De Pous, P., Donaire-Barroso, D., Bogaerts, S., Garcia-Porta, J., Escoriza, D., Arribas, O.J., Mouden, H.E., and Carranza, S. 2013. Review of the systematics, distribution, biogeography and natural history of Moroccan amphibians. Zootaxa, 3661:1–60.
https://doi.org/10.11646/ZOOTAXA.3661.1.1
Binkhorst, J.T. 1861. Monographie des gastropodes et des Céphalopodes de la Craie Supérieure du Limbourg. Muller Frères, Maastricht.
Black, M. and Barnes, B. 1959. The structure of Coccoliths from the English Chalk. Geological Magazine, 96(5):321–328.
Blackburn, D.C. and Wake, D.B. 2011. Class Amphibia Gray, 1825. Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, 3148(1):39–55.
https://doi.org/10.11646/zootaxa.3148.1.8
Blanco, A. and Brochu, C.A. 2017. Intra- and interspecific variability in allodaposuchid crocodylomorphs and the status of western European taxa. Historical Biology, 29:495–508.
https://doi.org/10.1080/08912963.2016.1201081
Bodor, E.R., Kovács, J., Vasile, Ș., Csiki-Sava, Z., and Váchová, Z. 2014. Fossil insect eggs from the Maastrichtian of the Haţeg Basin (Romania) – employment of morphometrics in taxonomical assessment, p. 148. In Popa, L.O., Adam, C., Chișmera, G., Iorgu, E., Murariu, D., and Popa, O.P. (eds.), International Zoological Congress of “Grigore Antipa” Museum – Book of Abstracts. Bucharest, Romania.
Bortolotti, V., Marroni, M., Nicolae, I., Pandolfi, L., Principi, G., and Saccani, E. 2004. An update of the Jurassic ophiolites and associated calc-alkaline rocks in the South Apuseni Mountains (Western Romania). Ofioliti, 29:5–18.
Bown, P.R. and Young, J.R. 1998. Techniques, p. 16–28. In Bown, P.R. (ed.), Calcareous Nannofossil Biostratigraphy. Chapman and Hall, London.
Bramlette, M.N. and Martini, E. 1964. The great change in calcareous nannoplankton fossils between the Maestrichtian and Danian. Micropaleontology, 10(2):291–322.
Bronzati, M., Montefeltro, F.C., and Langer, M.C. 2015. Diversification events and the effects of mass extinctions on Crocodyliformes evolutionary history. Royal Society Open Science, 2:140385.
https://doi.org/10.1098/rsos.140385
Brusatte, S.L., Vremir, M., Watanabe, A., Csiki-Sava, Z., Naish, D., Dyke, G., Erickson, G.M., and Norell, M.A. 2013. An Infant Ornithopod Dinosaur Tibia from the Late Cretaceous of Sebeş, Romania. Terra Sebus, 5:627–644.
Burnett, J.A. 1998. Upper Cretaceous, p. 132–199. In Bown, P.R. (ed.), Calcareous Nannofossil Biostratigraphy. Chapman and Hall, London.
Carvalho, I.S., Gasparini, Z.B., Salgado, L., Vasconcellos, F.M., and Marinho, T.S. 2010. Climate’s role in the distribution of the Cretaceous terrestrial Crocodyliformes throughout Gondwana. Palaeogeography, Palaeoclimatology, Palaeoecology, 297:252–262.
https://doi.org/10.1016/j.palaeo.2010.08.003
Chaine, J. and Duvergier, J. 1934. Recherches sur les otolithes des poissons. Etude descriptive et comparative de la sagitta des Teléostéens. Actes de la Societé Linnéenne de Bordeaux, 86:5–256.
Choi, S., Moreno-Azanza, M., Csiki-Sava, Z., Prondvai, E., and Lee, Y. 2020. Comparative crystallography suggests maniraptoran theropod affinities for latest Cretaceous European “geckoid” eggshell. Papers in Palaeontology, 6(2):265–292.
https://doi.org/10.1002/spp2.1294
Codrea, V., Smith, T., Dica, P., Folie, A., Garcia., G., Godefroit, P., and Itterbeeck, J.V. 2002. Dinosaur egg nests, mammals and other vertebrates from a new Maastrichtian site of the Hațeg Basin (Romania). Comptes Rendus Palevol, 1:173–180.
https://doi.org/10.1016/S1631-0683(02)00021-0
Codrea, V.A. and Dica, E.P. 2005. Upper Cretaceous–lowermost Miocene lithostratigraphic units exposed in Alba Iulia–Sebeș–Vințu de Jos area (SW Transylvanian basin). Studia Universitatis Babeș-Bolyai, Geologia, 50:19–26.
https://doi.org/10.5038/1937-8602.50.1.3
Codrea, V., Barbu, O., and Jipa-Murzea, C. 2010a. Upper Cretaceous (Maastrichtian) landvertebrate diversity in Alba District (Romania). Bulletin of the Geological Society of Greece, 43:594–601.
https://doi.org/10.12681/bgsg.11221
Codrea, V., Vremir, M., Jipa, C., Godefroit, P., Csiki, Z., Smith, T., and Fărcaș, C. 2010b. More than just Nopcsa’s Transylvanian dinosaurs: A look outside the Hațeg Basin. Palaeogeography, Palaeoclimatology, Palaeoecology, 293:391–405.
https://doi.org/10.1016/j.palaeo.2009.10.027
Codrea, V. and Jipa, C. 2011. New data on the Maastrichtian fishes (Lepisosteidae and Characiformes) from Transylvania. 9th Annual Meeting of the EAVP, Heraklion Abstract Volume, p. 19.
Codrea, V., Godefroit, P., and Smith, T. 2012. First Discovery of Maastrichtian (Latest Cretaceous) Terrestrial Vertebrates in Rusca Montană Basin (Romania), p. 571–581. In Godefroit, P. (ed.), Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems. Indiana University Press, Bloomington.
Colin, J.-P., Néraudeau, D., Nel, A., and Perrichot, V. 2011. Termite coprolites (Insecta: Isoptera) from the Cretaceous of western France: a palaeoecological insight. Revue de Micropaléontologie, 54:129–139.
https://doi.org/10.1016/j.revmic.2011.06.001
Cope, E.D. 1874. Review of the vertebrata of the Cretaceous period found west of the Mississippi River. U.S. Geological Survey of the Territories Bulletin, 1:3–48.
Csiki, Z., Ionescu, A., and Grigorescu, D., 2008. The Budurone microvertebrate fossil site from the Maastrichtian of the Haţeg Basin – flora, fauna, taphonomy and paleoenvironment. Acta Palaeontologica Romaniae, 6:49–66.
Csiki, Z., Vremir, M., Brusatte, S.L., and Norell, M.A. 2010. An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania. Proceedings of the National Academy of Sciences, 107:15357–15361.
https://doi.org/10.1073/pnas.1006970107
Csiki -Sava, Z., Vremir, M., Vasile, S., Brusatte, S.L., Dyke, G., Naish, D., Norell, M.A., and Totoianu, R. 2016. The East Side Story – The Transylvanian latest Cretaceous continental vertebrate record and its implications for understanding Cretaceous–Paleogene boundary events. Cretaceous Research, 57:662–698.
https://doi.org/10.1016/j.cretres.2015.09.003
Czabalay, L. 1983. Faunen des Senons im Bakony-Gebirge und ihre Beziehungen zu den Senon-Faunen der Ostalpen und anderer Gebiete. Zitteliana, 10:183–190.
de Celis, A., Narváez, I., and Ortega, F. 2018. Assessing the abiotic drivers of palaeobiodiversity in Eusuchia (Crocodyliformes), p 127–130. In Vaz, N. and Sá, A.A. (eds.), Yacimientos paleontológicos excepcionales en la península Ibérica. Cuadernos del Museo Geominero. Instituto Geológico y Minero de España, Madrid.
Deflandre, G. 1959. Sur les nannofossiles calcaires et leur systématique. Revue de Micropaléontologie, 2:127–152.
Dimian, E. and Popa-Dimian, M. 1964. Stratigraphic and sedimentologic data on Cretaceous formations between Mureș Valley and Ampoi Valley. Dări de Seamă ale Institului de Geologie, 50:103–107.
Dincă, A., Tocorjescu, M., and Stilla, A. 1972. Despre vârsta depozitelor continentale cu dinozaurieni din Bazinele Haţeg şi Rusca Montană. Dări de Seamă ale Institutului Geologic Român, 58:83–94.
Dubois, A. 2005. Amphibia Mundi. 1.1. An ergotaxonomy of recent amphibians. Alytes, 23:1–24.
Dyke, G., Vremir, M., Kaiser, G., and Naish, D. 2012. A drowned Mesozoic bird breeding colony from the Late Cretaceous of Transylvania. Naturwissenschaften, 99:435–442.
https://doi.org/10.1007/s00114-012-0917-1
El -Asa’ad, G.M.A. 1990. Maastrichtian species of the coral genus Cunnolites from Saudi Arabia. Journal of African Earth Sciences (and the Middle East), 10:633–642.
https://doi.org/10.1016/0899-5362(90)90030-I
Estes, R. 1969. Studies on fossil phyllodont fishes: interrelationships and evolution in the Phyllodontidae. Copeia, 1969:317–331.
Fernández, M.S., Wang, X., Vremir, M., Laurent, C., Naish, D., Kaiser, G., and Dyke, G. 2019. A mixed vertebrate eggshell assemblage from the Transylvanian Late Cretaceous. Scientific Reports, 9:1944. https://doi.org/10.1038/s41598-018-36305-3
Gameil, M., El-Sorogy, A.S., and Al-Kahtany, K. 2018. Solitary corals of the Campanian Hajajah Limestone Member, Aruma Formation, Central Saudi Arabia. Historical Biology, 31:1–17.
https://doi.org/10.1080/08912963.2018.1461217
Garcia, G. 2000. Diversité des coquilles “Minces” d’oeufs fossils du Crétacé supérieur du Sud de la France. Geobios, 33:113–126.
https://doi.org/10.1016/S0016-6995(00)80155-2
Gartner, S. 1968. Coccoliths and related calcareous nannofossils from Upper Cretaceous deposits of Texas and Arkansas. University of Kansas Paleontological Contributions, Articles, 48(1):1–56.
Gheerbrant, E., Abrial, C., and Cappetta, H. 1997. Nouveaux sites à microvertébrés continentaux du Crétacé terminal des Petites Pyrénées (Haute-Garonne et Ariège, France). Geobios, 30:257–269.
https://doi.org/10.1016/S0016-6995(97)80031-9
Góczán, F., Groot, J.J., Krutzsch, W., and Pacltová, B. 1967. Die Gattungen des “Stemma Normapolles Pflug 1953b” (Angiospermae) – Neubeschreibungen und Revision europäischer Formen (Oberkreide bis Eozän). Palaeontographica Abteilung B, 2:429–540.
Grigorescu, D. 1983. A stratigraphic, taphonomic and paleoecologic approach to a “forgotten land”: the dinosaur-bearing deposits of the Haţeg Basin (Transylvania, Romania). Acta Palaeontologica Polonica, 28:103–121.
Grigorescu, D. and Hahn, G. 1987. The first multituberculate teeth from the Upper Cretaceous of Europe (Romania). Geologica et Palaeontologica, 21:237–243.
Grigorescu, D., Hartenberger, J.-L., Rădulescu, C., Samson, P., and Sudre, J. 1985. Découverte de mammifères et de dinosaures dans le Crétacé supérieur de Pui (Roumanie). Comptes rendus de l'Académie des Sciences de Paris, 301:1365–1368.
Grigorescu, D., Şeclăman, M., Norman, D.B., and Weishampel, D.B. 1990. Dinosaur eggs from Romania. Nature, 346:417.
https://doi.org/10.1038/346417a0
Grigorescu, D., Venczel, M., Csiki, Z., and Limberea, R. 1999. New latest Cretaceous microvertebrate fossil assemblages from the Haţeg Basin (Romania). Geologie en Mijnbouw/Netherlands Journal of Geosciences, 98:310–314.
https://doi.org/10.1023/A:1003890913328
Grün, W. and Allemann, F. 1975. The Lower Cretaceous of Caravaca (Spain): Berriasian Calcareous Nannoplankton of the Miravetes Section (Subbetic Zone, Prov. of Murcia). Eclogae Geologicae Helvetiae, 68:147–211.
Hattner, J.G., Wind, F.H., and Wise, S.W. 1980. The Santonian-Campanian boundary: comparison of nearshore-offshore calcareous nannofossil assemblages. Cahiers de Micropaléontologie. 3:9–26.
Hauer, F. and Stache, G. 1863. Geologie Siebenburgens. Wilhelm Braumuller, Wien.
Hay, O.P. 1903. On a collection of upper Cretaceous fishes from Mount Lebanon, Syria, with descriptions of four new genera and nineteen new species. Bulletin of the American Museum of Natural History, 19:395–452
Herman, A.B. and Kvaček, J. 2007. Early Campanian Grünbach flora of Austria: systematic composition and palaeoclimatic interpretations. Acta Palaeobotanica, 47:37–55.
Herman, A.B. and Kvaček, J. 2010. Late Cretaceous Grünbach flora of Austria. Naturhistorisches Museum Wien, Wien.
Heřmanová, Z., Bodor, E., and Kvaček, J. 2013. Knoblochia cretacea, Late Cretaceous insect eggs from Central Europe. Cretaceous Research, 45:7–15.
https://doi.org/10.1016/j.cretres.2013.07.001
Heřmanová, Z., Kvaček, J., and Friis, E.M. 2021. Plant mesofossils from the Late Cretaceous Klikov Formation, the Czech Republic. Fossil Imprint, 77:256–270.
https://doi.org/10.37520/fi.2021.018
Integration and Application Network, 2024. Media Library
https://ian.umces.edu/media-library/
Jianu, C.M., Weishampel, D.B., and Ştiucă, E. 1997. Old and new pterosaur material from the Haţeg Basin (Late Cretaceous) of western Romania and comments about pterosaur diversity in the Late Cretaceous of Europe. Second European Workshop on Vertebrate Palaeontology (Espéraza – Quillan), Abstracts book, p. 1.
Kaplan, U., Kennedy, W.J., and Hiß, M. 2005. Stratigraphie und Ammonitenfaunen des Campan im nordwestlichen und zentralen Münsterland. Geologie und Paleaontologie in Westfalen, 64:1–171.
Kennedy, W.J. 1986a. The ammonite fauna of the Calcaire à Baculites (upper Maastrichtian) of the Cotentin Peninsula (Manche, France). Palaeontology, 29:25–83.
Kennedy, W.J. 1986b. The Campanian–Maastrichtian ammonite sequence in the environs of Maastricht (Limburg, The Netherlands), Limburg and Liège provinces (Belgium). Newsletters on Stratigraphy, 16:149–168.
Kennedy, W.J. 1986c. Campanian and Maastrichtian ammonites from northern Aquitaine, France. Palaeontological Association, London.
Kennedy, W.J. 1986d. The ammonite fauna of the type Maastrichtian with a revision of Ammonites colligatus Binkhorst, 1861. Bulletin de l’ Institut Royal des Sciences Naturelles de Belgique: Sciences de la Terre, 556:151–267.
Kennedy, W.J. and Summesberger, H. 1984. Upper Campanian ammonites from the Gschliefgraben (Ultrahelvetic, Upper Austria). Beiträge zur Paläontologie Österreich, 11:149–206.
Kennedy, W.J. and Kaplan, U. 1997. Ammoniten aus dem Campan des Stemweder Berges, Dammer Oberkreidemulde, NW–Deutschland. Geologie und Paläontologie in Westfalen, 50:31–245.
Kennedy, W.J., Tunoğlu, C., Walaszczyk, I., and Ertekí, I.K. 2007. Ammonite and inoceramid bivalve faunas from the Davutlar Formation of the Devrekanie–Kastamonu area, northern Turkey, and their biostratigraphical significance. Cretaceous Research 28:861–894.
https://doi.org/10.1016/J.CRETRES.2006.12.007
Knobloch, E. and Mai, D.H. 1983. Carbonized seeds and fruits from the Cretaceous of Bohemia and Moravia and their stratigraphical significance. Knihovniča zemního plynu a nafty, 4:305–322.
Knobloch, E. and Mai, D.H. 1986. Monographie der Früchte und Samen in der Kreide von Mitteleuropa. Nakladatelství Československé Akademie Věd Praha, Praha.
Koken, E. 1884. Über Fisch-Otolithen, insbesondere über diejenigen der norddeutschen Oligocän-Ablagerungen. Zeitschrift der Deutschen Geologischen Gesellschaft, 36:500–565.
Krézsek, C. and Bally, A.W. 2006. The Transylvanian Basin (Romania) and its relation to the Carpathian fold and thrust belt: insights in gravitational salt tectonics. Marine and Petroleum Geology, 23:405–442.
https://doi.org/10.1016/j.marpetgeo.2006.03.003
Kriwet, J., Soler-Gijón, R., and López-Martínez, N. 2007. Neoselachians from the upper Campanian and lower Maastrichtian (Upper Cretaceous) of the southern Pyrenees, northern Spain. Palaeontology, 50:1051–1071.
https://doi.org/10.1111/j.1475-4983.2007.00695.x
Lewy, Z. and Samtleben, C. 1979. Functional morphology and palaeontological significance of conchiolin layers in corbulid pelecypods. Lethaia, 12:341–351.
https://doi.org/10.1111/j.1502-3931.1979.tb01019.x
Lindfors, S.M., Csiki, Z., Grigorescu, D., and Friis, E.M. 2010. Preliminary account of plant mesofossils from the Maastrichtian Budurone microvertebrate site of the Haţeg Basin, Romania. Palaeogeography, Palaeoclimatology, Palaeoecology, 293:353–359.
https://doi.org/10.1016/j.palaeo.2009.10.018
Macovei, G. and Atanasiu, I. 1934. L’évolution géologique de la Roumanie. Crétacé. Anuarul Institutului Geologic, 16:63–280.
Mamulea, M.A. 1953a. Cercetări geologice în partea de Vest a Bazinului Haţeg (Regiunea Sarmisegetuza – Răchitova). Dări de Seamă ale Comitetului Geologic Român, 37:142–148.
Mamulea, M.A. 1953b. Studii geologice în regiunea Sânpetru – Pui (Bazinul Haţegului). Anuarul Comitetului Geologic Român, 25:211–174.
Mannion, P.D., Roger, B.J., Benson, M.T., Carrano, J.P., Tennant, J.J., and Richard, J.B. 2015. Climate constrains the evolutionary history and biodiversity of crocodylians. Nature Communications, 6:8438.
https://doi.org/ 10.1038/ncomms9438
Marincaș, V. 1966. Revizuirea acteonelelor din regiunea Sebeș. Studia Universitatis Babeș-Bolyai. Series Geologia-Geographia, 11:21–31.
Marincaș, V. 1970. Orizontarea depozitelor senoniene din regiunea Sebeș-Pianuri pe baze de micro și macrofauna. Sargetia, 7:289–295.
Markwick, P.J. 1998. Fossil crocodylians as indicators of Late Cretaceous and Cenozoic climates: implications for using palaeontological data in reconstructing palaeoclimate. Palaeogeography, Palaeoclimatology, Palaeoecology, 137:205–271.
https://doi.org/10.1016/S0031-0182(97)00108-9
Martin, J.E. 2007. New material of the Late Cretaceous globidontan Acynodon iberoccitanus (Crocodylia) from southern France. Journal of Vertebrate Paleontology, 27(2):326–372.
https://doi.org/10.1671/0272-4634(2007)27[362:NMOTLC]2.0.CO;2
Martin, J.E., Csiki, Z., Grigorescu, D., and Buffetaut, E. 2006. Late Cretaceous crocodilian diversity in Haţeg Basin, Romania. Hantkeniana, 5:31–37.
Martin, J.E., Rabi, M., and Csiki, Z. 2010. Survival of Theriosuchus (Mesoeucrocodylia: Atoposauridae) in a Late Cretaceous archipelago: a new species from the Maastrichtian of Romania. Naturwissenschaften, 97(9):845–854.
https://doi.org/ 10.1007/s00114-010-0702-y
Martínez, R. 1997. Campanian and Maastrichtian ammonites from southeast Spain. Cretaceous Research, 18:373–384.
https://doi.org/10.1006/cres.1997.0066
Matheron, P. 1843. Catalogue méthodique et descriptif des Corpsorganisés fossiles du département des Bouches-du-Rhône et lieuxcirconvoisins; précédé d’un Mémoire sur les terrains supérieurs augrès bigarré du S.E. de la France. Répertoire des travaux de la Société de Statistique de Marseille, 6:81–341.
Müller, J. 1846. Über den Bau und die Grenzen der Ganoiden, und über das natürliche System der Fische. Physikalisch-mathematische Abhandlungen der königlichen Akademie der Wissenschaften zu Berlin, 1846:117–216.
Narváez, I., Brochu, C.A., Escaso, F., Pérez-García, A., and Ortega, F. 2015. New crocodyliforms from southwestern Europe and definition of a diverse clade of European uppermost Cretaceous basal eusuchians. PLOS One, 10:e0140679.
https://doi.org/10.1371/journal.pone.0140679
Narváez, I., Brochu, C.A., Escaso, F., Pérez-García, A., and Ortega, F. 2016. New Spanish Late Cretaceous eusuchian reveals the synchronic and sympatric presence of two allodaposuchids. Cretaceous Research, 65:112–125.
https://doi.org/10.1016/j.cretres.2016.04.018
Narváez, I., Brochu, C.A., De Celis, A., Codrea, V., Escaso, F., Pérez-García, A., and Ortega, F. 2020. New diagnosis for Allodaposuchus precedens, the type species of the European Upper Cretaceous clade Allodaposuchidae. Zoological Journal of the Linnean Society, 189(2):618634.
https://doi.org/10.1093/zoolinnean/zlz029
Néraudeau, D., Perrichot, V., Colin, J.-P., Girard, V., Gomez, B., Guillocheau, F., Masure, E., Peyrot, D., Tostain, F., Videt, B., and Vullo, R. 2008. A new amber deposit from the Cretaceous (Upper Albian–Lower Cenomanian) of SW France. Cretaceous Research, 29:925–929.
https://doi.org/10.1016/j.cretres.2008.05.009
Nolf, D. 2003. Fish otoliths from the Santonian of the Pyrenean faunal province, and an overview of all otolith-documented North Atlantic Late Cretaceous teleosts. Bulletin de l’Institute royal des Sciences Naturelles de Belgique, Sciences de la Terre, 73:155–173.
Nopcsa, F. 1897. Vorläufiger Bericht über das Auftreten von oberer Kreide im Hátzeger Thale in Siebenbürgen. Verhandlungen des geologischen Reichsanstalt Wien, 1897:273–274.
Nopcsa, F. 1902. Dinosaurierreste aus Siebenbürgen. II. Schädelreste von Mochlodon. Denkschriften der Kaiserlichen Akademie der Wissenschaften, 72:149–175.
Nopcsa, F. 1914a. Die Lebensbedingungen der obercretacischen Dinosaurier Siebenbürgens. Centralbaltt für Mineralogie, Geologie, und Paläontologie, 1914:564–574.
Nopcsa, F. 1914b. Über das Vorkommen der Dinosaurier in Siebenbürgen. Verhandlungen der zoologische-botanischen Gesellschaft Wien, 54:12–14.
Nopcsa, F. 1915. Die Dinosaurier der Siebenbürgischen Landesteile Ungarns. Mitteilungen aus dem Jahrbuche der königlisch Ungarischen Geologischen Reichsanstalt, 23:1–24.
Nopcsa, F. 1923. On the geological importance of the primitive reptilian fauna of the uppermost Cretaceous of Hungary: with a description of a new tortoise (Kallokibotium). Quarterly Journal of the Geological Society of London, 79:100–116.
Nopcsa, F. 1925. Dinosaurierreste aus Siebenbürgen IV. Wirbelsaule von Rhabdodon und Orthomerus. Palaeontologia Hungarica, 1:273–304.
Nopcsa, F. 1928. Palaeontological notes on Reptilia. 7. Classification of the Crocodilia. Geologica Hungarica, Series Palaeontologica, 1:75–84.
Nopcsa, F. 1929. Dinosaurierreste aus Siebenbürgen V. Geologica Hungarica, Series Palaeontologica, 4:1–76.
Ősi, A., Rabi, M., Makádi, L., Szentesi, Z., Botfalvai, G., and Gulyás, P. 2012. The Late Cretaceous continental vertebrate fauna from Iharkút (Western Hungary): a review, p. 532–569. In Godefroit, P. (ed.), Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems. Indiana University Press, Bloomington.
Ősi, A., Szabó, M., Tóth, E., Bodor, E., Lobitzer, H., Kvaček, J., Svobodová, M., Szente, I., Wagreich, M., Trabelsi, K., Sames, B., Magyar, J., Makádi, L., Berning, B., and Botfalvai, G. 2021. A brackish to non-marine aquatic and terrestrial fossil assemblage with vertebrates from the lower Coniacian (Upper Cretaceous) Gosau Group of the Tiefengraben locality near St. Wolfgang im Salzkammergut, Austria. Cretaceous Research, 127:104938.
https://doi.org/10.1016/j.cretres.2021.104938
Pálfy, M. 1902. Die oberen Kreideschichten in der Umgebung von Alvincz. Mitteilungen aus dem Jahrbuche der Königlich Ungarischen Geologischen Anstalt, 13:241–348.
Paraschiv, D. 1979. Romanian Oil and Gas Fields. Institutul de Geologie şi Geofizică Studii Tehnice şi Economice, 13:1–397.
Perch-Nielsen, K. 1968. Der Feinbau und die Klassifikation der Coccolithen aus dem Maastrichtien von Danemark. Biologiske Skrifter, Kongelige Danske Videnskabernes Selskab, 16:1–96.
Pflug, H.D. 1953. Zur Entstehung und Entwicklung des angiospermiden Pollens in der Erdgeschichte. Palaeontographica Abteilung B, 95:60–171.
Piveteau, J. 1952. Traité de Paléontologie, p. 107–115. In Grassé, P.P. (ed.), Traité de Zoologie. Anatomie, systématique, biologie, 1, part 1, Phylogenie. Protozoaires: généralités. Flagellés. Masson and Cie, Paris.
Polette, F. and Batten, D.J. 2017. Fundamental reassessment of the taxonomy of five Normapolles pollen genera. Review of Palaeobotany and Palynology, 243:47–91.
https://doi.org/10.1016/j.revpalbo.2017.04.001
Protesco, O. 1937. Étude géologique et paléobiologique de l'ambre roumain. Les inclusions organiques de l'ambre de Buzău. Buletinul Societății Române de Geologie, 3:67–110.
Rabi, M. and Sebők, N. 2015. A revised Eurogondwana model: Late Cretaceous notosuchian crocodyliforms and other vertebrate taxa suggest the retention of episodic faunal links between Europe and Gondwana during most of the Cretaceous. Gondwana Research, 28(3):1197–1211.
Rabichon, A. 1938. Chihlimbarul. Cunoștințe folositoare. Știință pentru toți. Seria A, 57:1–32.
Radley, J.D. 1999. Weald Clay (Lower Cretaceous) palaeoenvironments in southeast England: molluscan evidence. Cretaceous Research, 20:365–368.
https://doi.org/10.1006/cres.1999.0156
Radley, J.D. and Allen, P. 2012. The Wealden (non-marine Lower Cretaceous) of the Weald Sub-basin, southern England. Proceedings of the Geologists’ Association, 123:245–318.
https://doi.org/10.1016/j.pgeola.2012.01.003
Rosen, D. E. 1973. Interrelationships of higher euteleostean fishes, p. 397–513. In Greenwood, P.H., Miles R.S., and Patterson C. (eds.), Interrelationships of Fishes. Academic Press, London.
Sanchiz, B. 1998. Salientia. Handbuch der Paläoherpetologie (Encyclopedia of Paleoherpetology). Teil 4, Verlag Dr. Friedrich Pfeil, München.
Sanders, D. and Baron-Szabo, R.C. 2005. Scleractinian assemblages under sediment input: their characteristics and relation to the nutrient input concept. Palaeogeography, Palaeoclimatology, Palaeoecology, 216:139–181.
https://doi.org/10.1016/j.palaeo.2004.10.008
Sanders, D. and Baron-Szabo, R. 2008. Palaeoecology of solitary corals in soft-substrate habitats: the example of Cunnolites (upper Santonian, Eastern Alps). Lethaia, 41:1–14.
https://doi.org/10.1111/j.1502-3931.2007.00039.x
Săndulescu, M. and Visarion, M. 1978. Considérations sur la structure tectonique du soubassement de la Dépression de Transylvanie. Dări de Seamă Institutul de Geologie şi Geofizică, 64:153–173.
Schlotheim, E.F. 1820. Die Petrefactenkunde auf ihrem jetzigen Standpunkte durch die Beschreibung seiner Sammlung versteinerter und fossiler Überreste des Thier- und Pflanzenreichs der Vorwelt. Becker, Gotha.
Schlüter, C.A. 1867. Beitrag zur Kenntnis der jüngsten Ammoneen Norddeutschlands. 1. Heft. Ammoniten der Senonbildungen, Bonn, 36 p.
Schubert, J.A., Wick, S.L., and Lehman, T.M. 2017. An Upper Cretaceous (middle Campanian) marine chondrichthyan and osteichthyan fauna from the Rattlesnake Mountain sandstone member of the Aguja Formation in West Texas. Cretaceous Research, 69:6–33.
https://doi.org/10.1016/j.cretres.2016.08.008
Schwarzhans, W. 1978. Otolith-morphology and its usage for higher systematical units with special reference to the Myctophiformes. Mededelingen van de Werkgroep voor Tertiaire en Kwartaire Geologie, 15:167–185.
Schwarzhans, W. 2010. Otolithen aus den Gerhartsreiter Schichten (Oberkreide: Maastricht) des Gerhartsreiter Grabens (Oberbayern). Palaeo Ichthyologica, 10:1–100.
https://doi.org/10.13140/2.1.4019.2641
Schwarzhans, W. 2012. Fish otoliths from the Paleocene of Bavaria (Kressenberg) and Austria (Kroisbach and Oiching-Graben). Palaeo Ichthyologica, 12:1–88.
Schwarzhans, W. 2018. A review of Jurassic and Early Cretaceous otoliths and the development of early morphological diversity in otoliths. Neues Jahrbuch Geologie und Paläontologie, Abhandlungen, 287:75–121.
https://doi.org/10.1127/njgpa/2018/0707
Schwarzhans, W., Carnevale, G., and Stringer, G.L. 2024. The diversity of teleost fishes during the terminal Cretaceous and the consequences of the K/Pg boundary extinction event. Netherlands Journal of Geosciences, 103:e5.
https://doi.org/10.1017/njg.2024.1
Sharpe, D. 1853. Description of the Fossil Remains of Mollusca Found in the Chalk of England. Part I. Cephalopoda. Monographs of the Palaeontographical Society, 7:1–26.
Sohl, N.F. and Kollmann, H.A. 1985. Cretaceous Actaeonellid Gastropods from the Western Hemisphere. Geological Survey Professional Paper, 1304:1–104.
Solomon, A., Codrea, V., Venczel, M., and Grellet-Tinner, G. 2020. A new species of large-sized pterosaur from the Maastrichtian of Transylvania (Romania). Cretaceous Research, 110:104316. https://doi.org/10.1016/j.cretres.2019.104316
Solomon, A., Codrea, V., Venczel, M., and Smith, T. 2022a. New data on Barbatodon oardaensis, the smallest Late Cretaceous multituberculate mammal from Europe. Comptes Rendus Palevol, 21:253–271.
https://doi.org/10.5852/cr-palevol2022v21a13
Solomon, A., Codrea, V., Venczel, M., Bordeianu, M., Trif, N., and Fărcaș, C. 2022b. Good or bad luck? – an “ox-bow” deposit from Oarda de Jos (Alba County, Romania): preliminary results. Brukenthal. Acta Musei, 17:447–464.
Staub, M. 1889. Sabal major Unger a Maros völgyéből. Földtani Közlöny, 19:258–264.
Stover, L.E. 1966. Cretaceous coccoliths and associated nannofossils from France and the Netherlands. Micropaleontology, 12(2):133–167.
Szabó, J. 1871. Az ajkai kőszéntelep a Bakonyban. Földtani Közlöny, 1:124–130.
Țabără, D., Vasile, Ș., Csiki-Sava, Z., Bălc, R., Vremir, M., and Chelariu, M. 2022. Palynological and organic geochemical analyses of the Upper Cretaceous Bozeș Formation at Petrești (southwestern Transylvanian Basin) – biostratigraphic and palaeoenvironmental implications. Cretaceous Research, 134:105148.
https://doi.org/10.1016/j.cretres.2022.105148
Tomescu, C. 1968. Orizontarea Santonianului de la Săsciori (Sebeș) pe baza echinoidelor. Dări de Seamă ale Ședințelor, 55:95–105.
Tomescu, C., Panin, Ş., Georgescu, F., Mantea, G., and Antonescu, E. 1969. Contributions to the Neocretaceous stratigraphy of deposits from Southern Apuseni Mountains. Studii și Cercetări de Geologie, Geofizică, Geografie, (Geologie), 14:239–252.
Trif, N. and Codrea, V. 2022. New data on Maastrichtian fishes of the ‘Hațeg Island’. Annales Geologiques de la Peninsule Balkanique, 83:1–12.
https://doi.org/10.2298/GABP220424001T
Tschudy, R.H. 1975. Normapolles pollen from the Mississippi Embayment. United States Geological Survey Professional Papers, 865:1–42.
Unger, F. 1867. Kreidepflanzen aus Österreich. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften. Mathematisch-Naturwissenschaftliche Classe, 55:642–654.
Vangerow, E.F. 1954. Megasporen und andere pflanzliche Mikrofossilien aus der Aachener Kreide. Palaeontographica, 96:24–36.
Vasile, Ș., 2008. A new microvertebrate site from the Upper Cretaceous (Maastrichtian) deposits of the Haţeg Basin. Sargetia. Acta Musei Devensis, 21:5–15.
Vasile, Ș. and Csiki, Z. 2011. New Maastrictian microvertebrates from the Rusca Montană Basin (Romania). Oltenia. Studii şi comunicări. Ştiinţele Naturii, 27:221–230.
Vasile, Ș., Bodor, E.R., Csiki-Sava, Z., and Szentesi, Z. 2013. Isopteran trace fossils from the Upper Cretaceous of Central-Eastern Europe, p. 88–89. In Țabără, D. (ed.), Ninth Romanian Symposium on Paleontology. Abstract Book. Editura Universități “Alexandru Ioan Cuza” Iași.
Vasile, Ș., Csiki-Sava, Z., Vremir, M., Norell, M.A., Totoianu, R., Brusatte, S.L., Bălc, R., and Țabără, D. 2021. New data on the Late Cretaceous microvertebrate assemblage from Petrești-Arini (SW Transylvanian Basin, Romania), p. 51–52. In Ionesi, V., Miclăuș, C., and Țabără, D. (eds.), Thirteenth Romanian Symposium on Paleontology. Tehnopress, Iași.
Vasile, Ș., Csiki-Sava, Z., Vremir, M., Țabără, D., Bălc, R., Bindiu-Haitonic, R., and Kövecsi, S.-A. 2022. The Late Campanian microvertebrate assemblage from Petrești (Transylvanian Basin, Romania), a new window into the evolution of European Cretaceous insular faunas, p. 214–215. In Belvedere, M., Mecozzi, B., Amore, O., and Sardella, R. (eds.), The XIX Annual Conference of the European Association of Vertebrate Palaeontologists, Benevento/Pietraroia.
Venczel, M. and Codrea, V. 2019. A new Theriosuchus -like crocodyliform from the Maastrichtian of Romania. Cretaceous Research, 100:24–38.
https://doi.org/10.1016/j.cretres.2019.03.018
Vidal, L.M. 1874. Datos para el conocimiento del terreno Garumniense de Cataluña. Boletin de la Comisión del Mapa Geológico de Espana, 1:209–247.
Vremir, M. 2010. New faunal elements from the Late Cretaceous (Maastrichtian) continental deposits of Sebeș area (Transylvania). Terra Sebus. Acta Musei Sabesiensis, 2:635–684.
Vremir, M., Bălc, R., Csiki-Sava, Z., Brusatte, S.L., Dyke, G., Naish, D., and Norell, M.A. 2014. Petrești-Arini – an important but ephemeral Upper Cretaceous continental vertebrate site in the southwestern Transylvanian Basin, Romania. Cretaceous Research, 49:13–38.
https://doi.org/10.1016/j.cretres.2014.02.002
Vremir, M., Dyke, G., and Totoianu, R., 2015. Repertoire of the Late Cretaceous vertebrate localities from Sebeș area, Alba County (Romania). Terra Sebus. Acta Musei Sabesiensis 7:695–724.
Willingshofer, E., Andriessen, P., Cloething, S., and Neubauer, F. 2001. Detrital fission track thermochronology of Upper Cretaceous syn-orogenic sediments in the South Carpathians (Romania): inferences on the tectonic evolution of a collisional hinterland. Basin Research, 13:379–395.
https://doi.org/10.1046/j.0950-091x.2001.00156.x
Young, J.R., Bown, P.R., and Lees, J.A., 2022. Nannotax3 Website: http://www.mikrotax.org/. International Nannoplankton Association.
Zechmeister, L. and Vrabely, V. 1927. Ueber Telegdit, ein fossiles Harz aus Siebenbürgen. Centralblatt für Mineralogie, Geologie und Paläontologie Abteilung A, 8:287–290.
Zekeli, F.L. 1852. Die Gasteropoden der Gosaugebilde in den Nordöstlichen Alpen. Abhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt, 1:1–124.
