

 The first late middle Pleistocene Stegodon (Proboscidea, Stegodontidae) evidence found in Peninsular Malaysia
The first late middle Pleistocene Stegodon (Proboscidea, Stegodontidae) evidence found in Peninsular Malaysia
Article number: 28.3.43
https://doi.org/10.26879/1432
Copyright Palaeontological Association, September 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 23 July 2024. Acceptance: 27 August 2025.
ABSTRACT
A maxillary left milk molar (dp3) of a Stegodon was discovered in a cave in northern Peninsular Malaysia. This finding represents the southernmost record of Stegodontidae in continental Asia. The half-exposed tooth was embedded in breccia attached to the cave wall. A micro-computed tomography (μCT) scanning was employed to virtually isolate the specimen from the matrix and to reconstruct a 3D model of the fossil for accurate measurements. No sign of wear or mechanical abrasion was observed in the specimen. Age determination on the matrix based on post-infrared infrared luminescence (pIR-IRSL) dating and U-series revealed a late middle Pleistocene age of between 233 ± 31 to 199 ± 28 ka. Detailed comparisons with specimens of Stegodon orientalis and Stegodon trigonocephalus revealed a number of differences in size and morphologies (development of posterior cingulum, presence of enamel tubercles at posterior cingulum, tapering of each plate lingual-buccally, thickness of the plates, and depth of the valley between plates) which are different to either of these species. The discovery of Stegodon in Peninsular Malaysia provides a better understanding of the distribution of this genus in the Sundaic subregion during the middle Pleistocene.
Amir Hakim Amiruddin. Department of Geology, Universiti Malaya, 50603, Kuala Lumpur, Malaysia. amirhakim081197@gmail.com
Lim Tze Tshen. Department of Geology, Universiti Malaya, 50603, Kuala Lumpur, Malaysia. limtzetshen@yahoo.com
Alexandra van der Geer. Naturalis, Darwinweg 2, 2333 CR Leiden, Netherlands. alexandra.vandergeer@naturalis.nl
Kira Westaway. School of Natural Sciences, Faculty of Science and Engineering, Macquarie University, NSW 2109, Australia. kira.westaway@mq.edu.au
Chiang Hongwei. Research Center for Environment Changes, Academia Sinica Taipei 115201, Taiwan, ROC. hwchiang@ntu.edu.tw
Ru Smith. Department of Geology, Universiti Malaya, 50603, Kuala Lumpur, Malaysia. ru.d.a.smith@gmail.com
Hijaz Kamal Hasnan. Department of Geology, Universiti Malaya, 50603, Kuala Lumpur, Malaysia. hijazzains@um.edu.my
Norliza Ibrahim. Department of Oral & Maxillofacial Clinical Sciences, Faculty of Dentistry, Universiti Malaya, 50603 Kuala Lumpur, Malaysia. norlizaibrahim@um.edu.my
Mohd Azmi Abdul Razak. Department of Restorative Dentistry, Faculty of Dentistry, Universiti Malaya, 50603 Kuala Lumpur, Malaysia. azmirazak@um.edu.my
Nguyen Thi Mai Huong. Institute of Archaeology, Hanoi, Vietnam. maihuong72@gmail.com
Nguyen Anh Tuan. Institute of Archaeology, Hanoi, Vietnam. nguyenanhtuanvkch@gmail.com
Ros Fatihah Muhammad. Department of Geology, Universiti Malaya, 50603, Kuala Lumpur, Malaysia (corresponding author). rosfmuhammad@um.edu.my
Keywords: juvenile; karst fossil; 3D computed tomography; Quaternary; luminescence dating
Final citation: Amiruddin, Amir Hakim, Lim, Tze Tshen, van der Geer, Alexandra, Westaway, Kira E., Hongwei, Chiang, Smith, Ru, Hasnan, Hijaz K., Ibrahim, Norliza, Razak, Mohd Azmi Abdul, Nguyen, Thi Mai Huong, Nguyen, Anh Tuan, and Muhammad, Ros Fatihah. 2025. The first late middle Pleistocene Stegodon (Proboscidea, Stegodontidae) evidence found in Peninsular Malaysia. Palaeontologia Electronica, 28(3):a43.
https://doi.org/10.26879/1432
palaeo-electronica.org/content/2025/5637-stegodon-in-peninsular-malaysia
Copyright: September 2025 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
The genus Stegodon is a key taxon in the studies of the Pleistocene mammalian faunas in mainland Southeast Asia and south China that were characterized by the occurrence of the early to late Pleistocene faunal assemblage “Ailuropoda-Stegodon faunal complex” (Kahlke, 1961; Rink et al., 2008; Suraprasit et al., 2016).
Fossil records suggested that this genus was widely distributed in parts of central and east Africa and south and Southeast Asia. Thus far, fossils recorded from mainland and insular Asia were reported from China, Taiwan, Japan, India, Pakistan, Nepal, Myanmar, Thailand, Laos, Vietnam, Indonesia and Philippines, with ages ranging from the late Miocene to the late Pleistocene or Holocene (O’ Reagan et al., 2005; Ao et al., 2016). Miocene and Pliocene representatives of the genus in Asia such as S. zhaotongensis, S. licenti, S. clifti, S. bombifrons, S. zdanskyi, S. officinalis, S. aurorae, S. miensis, and S. insignis were found in east and south Asia, with the exception of S. sompoensis, which was recorded only in Sulawesi island (Indonesia); later forms, including S. chiai, S. huananensis, S. wushanensis, S. preorientalis, S. sinensis, and S. elephantoides were found from earlier Pleistocene deposits in China, Taiwan, Myanmar, and Laos, with no known records dated later than that. Large- and medium-sized Stegodon, such as S. orientalis and S. trigonocephalus, as well as dwarf species (S. hypsilophus, S. sumbaensis, S. timorensis, S. sondaari, S. florensis, and S. cf. luzonensis) thrived in various places in continental and insular Southeast Asia (including, Sundaic subregion in the west and Wallacea in the east) and Luzon island in the Philippines during the middle Pleistocene times. Even though S. orientalis and S. trigonocephalus were known to have existed in earlier Pleistocene, these two species persisted through middle Pleistocene until at least to the late Pleistocene (Table 1). The genus was generally recorded as globally extinct during terminal Pleistocene (van den Bergh et al., 2008; Turvey et al., 2013), with a few controversial Holocene records from mainland China (Tong and Liu, 2004).
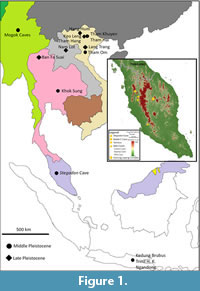 Locations of middle to late Pleistocene sites in mainland Southeast Asia and Java are shown in Figure 1. Within the Malay Peninsula, the geographically closest known occurrence of Stegodon is from Satun province in peninsular Thailand (Duangkrayom et al., 2017). However, as of the time of this study, those specimens have not yet been dated. This is an in-depth analysis of a Stegodon fossil found from a cave in northern Peninsular Malaysia. The finding was briefly reported by Muhammad et al. (2020) and represents the southernmost distribution of the genus in mainland Southeast Asia.
Locations of middle to late Pleistocene sites in mainland Southeast Asia and Java are shown in Figure 1. Within the Malay Peninsula, the geographically closest known occurrence of Stegodon is from Satun province in peninsular Thailand (Duangkrayom et al., 2017). However, as of the time of this study, those specimens have not yet been dated. This is an in-depth analysis of a Stegodon fossil found from a cave in northern Peninsular Malaysia. The finding was briefly reported by Muhammad et al. (2020) and represents the southernmost distribution of the genus in mainland Southeast Asia.
The present work aims to provide a detailed description of the new Stegodon fossil from Peninsular Malaysia based on μCT scans. We also report the results of the geochronological dating based on pIR-IRSL dating of feldspar grains from the surrounding breccia matrix and U-series of a flowstone that overlies the Stegodon fossil, in this new Pleistocene fossil site in Malaysia.
GEOLOGICAL SETTING
The study area is located in Gopeng (Kinta Valley), a district in the state of Perak, about 180 km north of Kuala Lumpur in the west part of Peninsular Malaysia (04°28'16.1503", 101°08'57.4313") at 50 m above mean sea level (Figure 1). The small hill and cave network where the Stegodon fossil was found were previously unmarked on regional topographical maps consulted (Muhammad et al., 2020). The cave was subsequently named Stegodon Cave, and this name will be used in this article.
The karst system in the valley consist of the ± 1550 m thick Kinta Limestone that was earlier assigned a Lower Devonian to Middle Permian age (Suntharalinggam, 1968) and later a Middle Ordovician to Middle Permian age (Henri and Amnan, 1994). It forms typical tropical karst towers that protrude above the vast valley (Muhammad, 2018) while most of it in the form of dissected subsurface platforms. The valley is sandwiched in between two I-type granitic highlands (Ghani et al., 2013), the Main Range in the east and Kledang Range on the west. Heavy drainage from the highlands brought down the alluvium, the products of weathered granite into the floodplain. Walker (1965) named the main stratigraphic units of the Cenozoic cover in Kinta Valley as Boulder Beds, Old Alluvium and Young Alluvium. The sediments were reassessed and Old Alluvium renamed as Simpang Formation and Young Alluvium as Beruas Formation by Suntharalinggam (1983).
The tin-rich alluvium that blankets the valley is associated with the Simpang Formation while the younger Beruas Formation fills up V-shaped valleys. Even though the earlier Simpang Formation was assigned to the Pleistocene, the underlying Cenozoic sediments that cannot be differentiated from the Quaternary sediments (Suntharalinggam, 1983) imply that this formation could extend into the Neogene. This has also been indicated earlier by Batchelor (1979a, 1979b).
 Stegodon Cave
Stegodon Cave
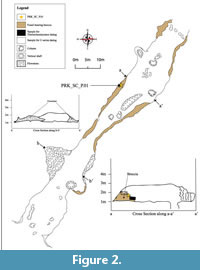
This cave forms a single northeast-southwest orientated 94 m passage that cuts through the small unnamed karst hill, with its floor located about 6 m above the general ground. The average height of the ceiling is about 2 m throughout the cave, and width of not more than 10 m. Two distinctive vertical shafts connect the passage to the lower passage, which lies on the general ground (Figure 2).
Remnants of highly lithified breccia up to 80 cm thick line both walls, and certain parts over the speleothems. The breccia is oligomictic, and primarily comprises fine sands with some muds, along with streaky crystalline carbonate intercalations and a few subangular rock pebbles. The tooth of the Stegodon was located about 16 m from the northeast mouth, on the northwestern wall, approximately 70 cm above the floor (Figure 3). Scattered fossil-bearing unconsolidated sediments embed the walls and ceiling of the lower passage.
MATERIALS AND METHODS
Abbreviations
 PRK Perak, Malaysia
PRK Perak, Malaysia
SC Stegodon Cave
KL Keo Leng, Vietnam
LKCNHM Lee Kong Chian Natural History Museum, National University of Singapore, Singapore.
LF Lamellar frequency
Description and measurements of the Stegodon tooth
Results from visual observation of the external morphological characters were documented. A complete cave passageway was mapped and drawn using simple traversing with laser rangefinder and a compass.
All measurements were taken using hand-held vernier callipers (model: H+W: 150 x 0.05mm) for the width of the plates, length and height of the molars. Morphological terms for cheek teeth and methods of measuring and tabulating biometrical parameters are largely based on those used by Maglio (1973: 8-13), Roth and Shoshani (1988: 572-574) and van den Bergh (1999: 28-30). Dimensions used in the measurement include:
Length (L) - Maximum length of a cheek tooth, measured along the longitudinal axis of the tooth parallel to crown base.
Width (W) - Maximum width of a cheek tooth, measured across the widest plate (including the covering cementum layer, if any) with the jaws of the calliper parallel to the anterior and posterior plates.
Height (H) - Maximum height of the highest plate, measured from crown base (not including root mass) to the top of the plate.
Height/Width Index (H/W Index) - Also known as Hypsodonty Index (HI) or Relative Crown Height (RCH), in which the maximum height (H) of a tooth is expressed as a percentage of its maximum width (W) and calculated by multiplying the height-width ratio by 100.
Plate number or Plate formula - The total number of plates or lamellae present or preserved as viewed from the side of a tooth which includes the small half-plate at the front end of a tooth and also plates partially covered within cementum.
Lamellar Frequency (LF) - (Total number of plates x 100)/ Maximum length.
This paper follows the common practice among modern palaeontologists to label the six cheek teeth of Stegodon and Elephas specimens as dp2-dp3-dp4-M1-M2-M3, instead of using Roman numerals I to VI, to indicate the order of the tooth growth from the back to the front of each jaw quadrant.
The Stegodon specimen (Catalogue No.: PRK_SC_P.01) is temporarily housed in the Geology Department of Universiti Malaya. Direct comparisons were made with one specimen of S. orientalis from Keo Leng, Vietnam (Institute of Archaeology, Hanoi) and one specimen of S. trigonocephalus (CD 11649) from Java, Indonesia (Naturalis Biodiversity Center, Leiden). The S. orientalis specimen from Vietnam does not seem to have any catalogue number, so it is here referred to as ‘KL DP3’. Both of these comparative fossil specimens have been identified as maxillary milk teeth (dp3). For lamellar frequency comparison we also used a selected group of fossil and modern dp3s: Stegodon trigonocephalus, CD 11647a and CD 11647b (from Hooijer, 1955), and modern Asian elephant Elephas maximus, ZRC 4.1669 from the Lee Kong Chian Natural History Museum, Singapore.
Terminology for anatomical parts
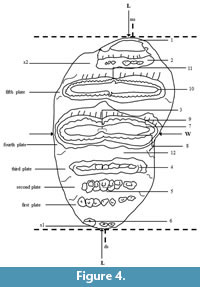 The terminology and measurement of Stegodon tooth (see Figure 4) were modified from van den Bergh (1999).
The terminology and measurement of Stegodon tooth (see Figure 4) were modified from van den Bergh (1999).
The plate formula used:
x N x
x before the number N = the anterior cingulum.
N = total number of plates.
x after the number N = the posterior cingulum.
MicroCT scanning
Given that PRK_SC_P.01 is a deciduous tooth and potentially fragile, special care has been taken during its preparation. The specimen is partially embedded in hard carbonate cave breccia, which bubbles under an acid test and reveals only about half of the specimen. Based on previous fossil extraction experience, where a heavy-duty battery-operated drill was used, it was determined that the breccia is too indurated for routine airbrushing techniques. The standard practice of soaking the sediment in acid to loosen it was also avoided due to concerns about damaging the delicate tooth. The use of micro-CT (μCT) technology significantly advances palaeontology by reducing the time and effort required for cleaning, especially with fragile specimens like this one.
The scanning was conducted at the scanning facility of the Universiti Malaya, using a Zeiss Xradia Versa 520 machine with a nanofocus transmission tube and moveable detector. The voxel size of the reconstructed volume is 63 µm in three dimensions. 3D rendering, meshes and movies were made using 3DSlicer software (Kikinis et al., 2014). Variations in the material density are visible through changes of greyscale from dark (low density, low attenuation) to light (high density, high attenuation). Segmentation of the volume was achieved with a combination of threshold and manual paint methods. Using the virtually isolated specimen, a set of measurements could be confidently obtained from the images.
pIR-IRSL dating of feldspars. A block sample of the surrounding breccia matrix was sampled in-situ using subdued red lights in dark cave conditions and wrapped in black plastic (Figure 3). Within subdued red-light conditions, the light-exposed outer layer was removed using a chisel and hammer and was retained as the dosimetry sample. These layers were broken up using a pestle and mortar and oven dried, then the entire fraction was milled and used for environmental dose estimation. The unexposed inner core was also gently broken up using a small hammer and was processed using the standard sample purification procedures for feldspar separation (Aitken, 1998) including a 10% wash in hydrofluoric acid for 10 minutes to remove the external alpha-dosed rinds (Rizal et al., 2020). All luminescence analysis was conducted at the ‘Traps’ luminescence dating facility at Macquarie University in Sydney, Australia.
To obtain an estimate of the environmental dose rate for each of the samples, we first measured beta dose rates using a Geiger-Muller multi-counter beta counting of dried and powdered sediment samples (Botter-Jensen and Mejdahl, 1988) in the laboratory. Allowance was made for the effect of sample moisture content, different grain sizes (Brennan, 2003) and HF etching on attenuation of the beta dose, and the total beta dose-rate contribution was calculated by comparing the beta count rate to a standard beta source (SHAP with a dose rate of 5.99 Gy/kyr) and magnesium oxide as a non-beta emitting background material. Secondly, thick source alpha counting using a Daybreak 583 intelligent alpha counter was used to obtain estimates of Uranium and Thorium (Wang and Xia, 1991) to estimate the gamma dose rate, and thirdly the difference between beta and alpha counting was used to estimate potassium values. These estimates were then converted to gamma dose rates using the conversion factors of Guerin et al. (2011). Allowance was made for the effect of sample moisture content (Aitken, 1988) on the external beta and gamma dose rates using a long-term water content of between 2 ± 0.2 %, which is similar to the measured (field) water content of between 2.2% and allows for an initial period of saturation when first deposited in the karst environment.
The total dose rate was then calculated using an effective internal beta dose rate of 0.84 Gy kyr-1 (Huntley and Baril, 1997; Huntley and Hancock, 2001) for the 180-212 µm feldspar samples (due to the radioactive decay of 40K and 87Rb), which were made assuming K (12.5 ± 0.5%) and 87Rb (400 ± 100 μg g-1) concentrations and included in the total dose rate. Cosmic-ray dose rates were estimated from published relationships (Prescott and Hutton, 1994), making allowance for the thickness of limestone above the cave (~10 m with an assumed density of 1.2 g/cm3), sediment overburden at the sample locality (1.0 m with an assumed density of 2.0 g/cm3), the altitude (~20 m above sea level) and geographic latitude and longitude (4°S and 101°E) of the sampling site.
We used the 90-180 μm size fraction and stainless steel single aliquot discs for all the procedural tests using the following preheat and IR stimulation combinations: 1) 250 and 225°C (pIR-IRSL50,225)28,29; 2) 280 and 250°C (pIR-IRSL50,250)30; 3) 300 and 270°C (pIR-IRSL50,270)30; 4) 320 and 290°C (pIR-IRSL50,290)31,32; and as the expected De is >450 Gy, we also tested 5) 320 and 200/290°C (pIR-IRSL200.290)33. All tests followed these procedures 1) Preheat plateau and dose recovery tests - different preheat/IR laser stimulation combination was applied to each disc then a surrogate dose of 20 Gy was recovered, 2) Bleaching tests - a dose of 20 Gy was applied and bleached in a solar simulator for 4 hrs 3) Fading tests - testing anomalous fading in single-aliquots of feldspars.
The tests revealed that the flattest plateau was provided by the pIR-IRSL50,270, pIR-IRSL50,290 and pIR-IRSL200,290 signals, while the pIR-IRSL50,250 signal provided the best recovery of the surrogate dose (with a dose recovery ratio of 0.995) and lowest residual values after bleaching (<10 Gy) (although calculated, these residual doses were not subtracted from the De). Thus, the pIR-IRSL50,250 was chosen as the most suitable procedure for this sample with a fading correction of 1.7 % applied.
Single aliquots of potassium feldspar were processed first to gauge suitability for a subsequent single grain approach. Thus, single aliquot discs loaded with 90-125 µm feldspar grains were loaded onto a carousel and processed in a Riso TL-DA-20 containing an automated Detection and Stimulation Head (DASH) set up with a Dual laser single grain attachment with a Blue/UV sensitive Electron Tube PMT (PDM9107Q-AP-TTL-03) with maximum detection efficiency between 200 and 400 nm. The filters in the automated detection changer were set on the blue filter pack (Schott BG-39 and Corning 7-59 filters to transmit wavelengths of 320-480 nm (Hutt et al., 1988). The grains were stimulated using IR (830 nm) diodes for 250 s with the first 15 s as the signal and last the 100 s as background.
In addition, 180-212 µm feldspar grains were also processed but using aluminium single grain discs. They were loaded in the same reader with the same filter combination, but this time stimulated using a 830 nm 140 mW TTL modulated IR laser for 2.5 s, first at 50 °C and secondly at 270 °C after a 300°C preheat according to the procedures of the pIR-IRSL protocol selected. The signal was integrated over the first 0.21-0.46 s with the last 1.88-2.5 s used as a background, with a standard exponential fit and monte carlo simulation for error determination.
U-series dating. We collected in situ flowstone stratigraphically above and below the Stegodon (Figure 3), which offers an opportunity to apply U-Th dating to bracket the Stegodon’s age. Back in the lab, we carefully identified and selected the best calcite crystals, then cleaned them using ultrasonication with pure water. Procedures for U and Th chemical separation and purification are similar to those described in Edwards et al. (1987) and Shen et al. (2002). In short, calcite grains were dissolved in 7N nitric acid, mixed with an appropriate amount of 229Th-233U-236 U triple spike and dissolved iron. Then, ammonium hydroxide was added dropwise to increase the solution’s pH until the appearance of iron oxyhydroxide precipitation. After removing the major Ca2+ ions with three times of centrifuging, the AG1-X8 resin (Bio-Rad) is used to separate and purify U/Th. All the chemical steps were performed on laminar-flow benches in a calss-1000 cleanroom. Uranium and thorium isotopic compositions were measured on a multi-collector inductively coupled plasma mass spectrometer (MC-ICP-MS), Thermo Scientific NEPTUNE, housed at the Department of Geosciences, National Taiwan University (Shen et al., 2012). Decay constants of 9.1705 × 10−6 yr-1 for 230Th, 2.8221 × 10−6 yr−1 for 234U (Cheng et al., 2013) and 1.55125 × 10−6 yr−1 for 238 U (Jaffey et al., 1971) were adopted. Age (relative to 1950 CE) was corrected using an initial 230Th/232 Th atomic ratio of 4 ± 2 × 10−6. The uncertainties from all the sources were propagated through the U-Th age calculation and presented at the 2σ level or two standard deviations of the mean (2σm) unless otherwise noted. The detailed off-line data deduction and 230Th date calculation were described in Shen et al. (2002). We tried two subsamples individually from above and below the Stegodon, in total four.
SYSTEMATIC PALAEONTOLOGY
Order PROBOSCIDEA Illiger, 1811
Suborder ELEPHANTIFORMES Tassy, 1988
Superfamily ELEPHANTOIDEA Osborn, 1921
Family STEGODONTIDAE Osborn, 1918
Genus STEGODON Falconer and Cautley, 1847
Stegodon sp.
Figure 5, Figure 6
PRK_SC_P.01. A cheek tooth embedded in a conglomerate matrix.
Morphological descriptions
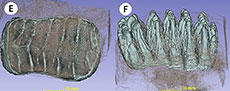
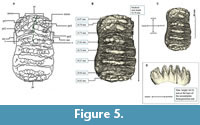 Peninsular Malaysia Stegodon (PRK_SC_P.01). The plate formula was determined to be x6x. Both the anterior and posterior cingula are preserved which indicate that this is a complete dp3 (see Figure 5). The tooth is brachydont and all the plates are unworn. From the exposed lateral view, the crown is longitudinally convex, the plates diverge apically, and constriction on the mesial end indicates that this is an upper tooth. The lateral convergence of plates suggested that the exposed side is lingual. All these characters show that this specimen is a left maxillary tooth.
Peninsular Malaysia Stegodon (PRK_SC_P.01). The plate formula was determined to be x6x. Both the anterior and posterior cingula are preserved which indicate that this is a complete dp3 (see Figure 5). The tooth is brachydont and all the plates are unworn. From the exposed lateral view, the crown is longitudinally convex, the plates diverge apically, and constriction on the mesial end indicates that this is an upper tooth. The lateral convergence of plates suggested that the exposed side is lingual. All these characters show that this specimen is a left maxillary tooth.
In occlusal view, the posterior outline is broader compared to the anterior outline of the specimen. Each plate consists of cusps that are called ‘conelets’ and separated by longitudinal grooves. The median sulcus is well-developed in at least the anterior plates of the molar and does not reach the crown base. This median sulcus is prominent along the first to the third plates of the mesial end which divided the postrite and pretrite halves of the specimen. The mesial constriction is well-developed between the second and third plates of the mesial end. The lingual part is concave and the buccal end slightly convex in shape (see Figure 5).
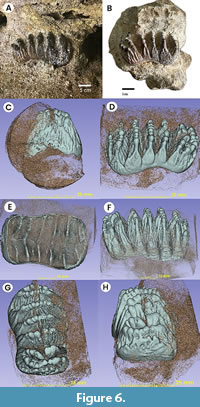 In lingual view, the plates are bulbous towards the direction of the root, taper and thin towards the apex and take the shape of “roof”. The valleys that separate the plates are Y-shaped, and the spacing of the plates increases anteriorly. Three plates at one end of the tooth are arranged more vertically which indicates that this is the posterior end of the tooth; the rest of the plates lean and converging apically which indicate that this is the anterior end of the tooth. The crown is higher towards the midline compared to the posterior and anterior ends. Both cingula are closely attached to the respective first plate at the anterior and the posterior ends of the tooth. There are no observable roots from this specimen (see Figure 6E), a fact which indicates that the roots were either not well developed or not preserved. The posterior cingulum consists of an estimated seven conelets. The first plate from the posterior end contains an estimated eight conelets. The penultimate and third plate (both from the posterior end) consist of an estimated eight and seven conelets, respectively. The fourth and fifth plate (both from the posterior end) both consist of five conelets. The sixth plate from the posterior end has an estimated nine conelets and contains an anterior cingulum. This anterior cingulum consists of an estimated six conelets and is fused with the first plate of the mesial end towards the middle.
In lingual view, the plates are bulbous towards the direction of the root, taper and thin towards the apex and take the shape of “roof”. The valleys that separate the plates are Y-shaped, and the spacing of the plates increases anteriorly. Three plates at one end of the tooth are arranged more vertically which indicates that this is the posterior end of the tooth; the rest of the plates lean and converging apically which indicate that this is the anterior end of the tooth. The crown is higher towards the midline compared to the posterior and anterior ends. Both cingula are closely attached to the respective first plate at the anterior and the posterior ends of the tooth. There are no observable roots from this specimen (see Figure 6E), a fact which indicates that the roots were either not well developed or not preserved. The posterior cingulum consists of an estimated seven conelets. The first plate from the posterior end contains an estimated eight conelets. The penultimate and third plate (both from the posterior end) consist of an estimated eight and seven conelets, respectively. The fourth and fifth plate (both from the posterior end) both consist of five conelets. The sixth plate from the posterior end has an estimated nine conelets and contains an anterior cingulum. This anterior cingulum consists of an estimated six conelets and is fused with the first plate of the mesial end towards the middle.
Comparison materials
S. orientalis (KL DP3). The plate formula was determined to be x5x, and the crown is brachydont. The posterior end is complete with smooth texture while the anterior end is incomplete, judging from the hard texture and abrasion seen at this end. This tooth is slightly used, the first three plates from the posterior end and posterior cingulum are not used. However, the fifth plate from the posterior end is heavily used with a single large enamel island and exposed dentine. The median sulcus that separates the postrite and pretrite halves is most observable at the third and fourth plates from the posterior end. From the sagittal section, the crown is longitudinally convex, and the plates diverge apically, and there is a constriction on the mesial end which indicates that this is an upper cheek tooth. All these characters show that this specimen is a tooth from the right maxilla.
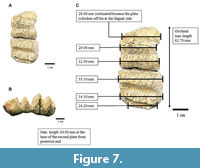 From the lateral view, the plates are bulbous towards direction of the root, taper and thin towards the apex and “roof” shaped (see Figure 7). The valleys that separate the plates are Y-shaped, and the spacing of the plates increases anteriorly. Two plates at one end of the tooth are arranged more vertically which indicates that this is the posterior end of the tooth; the rest of the plates leaning and converging apically which indicate that this is the anterior end of the tooth. The crown is higher towards the posterior end compared to the anterior. The buccal side of the anterior cingulum is damaged and only the lingual side of the anterior cingulum can be observed from this specimen. There are no observable roots from this specimen which indicates that the roots were either not well developed or not preserved.
From the lateral view, the plates are bulbous towards direction of the root, taper and thin towards the apex and “roof” shaped (see Figure 7). The valleys that separate the plates are Y-shaped, and the spacing of the plates increases anteriorly. Two plates at one end of the tooth are arranged more vertically which indicates that this is the posterior end of the tooth; the rest of the plates leaning and converging apically which indicate that this is the anterior end of the tooth. The crown is higher towards the posterior end compared to the anterior. The buccal side of the anterior cingulum is damaged and only the lingual side of the anterior cingulum can be observed from this specimen. There are no observable roots from this specimen which indicates that the roots were either not well developed or not preserved.
The posterior cingulum has nine unworn conelets. The first, second and third plates (all from the posterior end) have an estimated 14, 11 and 12 conelets, respectively, all not in use. Mesoconelets between the sixth and seventh conelets from the lingual side can be observed from the posterior end. The fourth plate from the posterior end shows the division of the postrite and pretrite sides, where the lingual side is slightly used, and the conelets are fused together to form one single large enamel island. The fifth plate from the posterior end has enamel fused to form one single large enamel island. The anterior cingulum is not fully preserved.
The tooth is more convex at the posterior end, tapered at the anterior end at the third plate from the posterior end (mesial constriction) and convex again at the anterior end.
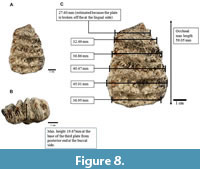 S. trigonocephalus (CD 11649). The plate formula of this specimen was determined to be (+)5(+)x. The posterior end of the specimen is complete, but it is damaged at the mesial-lingual side (see Figure 8). The specimen is a slightly worn left upper dp3. The first two plates from the posterior end have a high number of conelets (10 or more) compared to the subsequent plates (which have less than 10 conelets). These two plates are not in use. Formation of small enamel islands (digitation) can be seen at the third and fourth plates from the posterior end and larger enamel islands can be seen at the lingual side of the fifth plate from the posterior end. Enamel wrinkles are most obvious at the fourth and fifth plates from the posterior end. A prominent median sulcus which divides the postrite and pretrite halves can be observed towards the posterior end. An obvious subdivision in two parts can be seen at the second plate from the posterior end, of which the lingual side is wider than the buccal side.
S. trigonocephalus (CD 11649). The plate formula of this specimen was determined to be (+)5(+)x. The posterior end of the specimen is complete, but it is damaged at the mesial-lingual side (see Figure 8). The specimen is a slightly worn left upper dp3. The first two plates from the posterior end have a high number of conelets (10 or more) compared to the subsequent plates (which have less than 10 conelets). These two plates are not in use. Formation of small enamel islands (digitation) can be seen at the third and fourth plates from the posterior end and larger enamel islands can be seen at the lingual side of the fifth plate from the posterior end. Enamel wrinkles are most obvious at the fourth and fifth plates from the posterior end. A prominent median sulcus which divides the postrite and pretrite halves can be observed towards the posterior end. An obvious subdivision in two parts can be seen at the second plate from the posterior end, of which the lingual side is wider than the buccal side.
From the sagittal section, the crown is longitudinally convex, the plates diverge apically, and there is constriction on the mesial end which indicates that this is an upper cheek tooth. All these characters show that this specimen is a tooth from the left maxilla.
From the lateral view, the plates are bulbous towards direction of the root, taper and thinner towards the apex, and “roof” shaped. The valleys that separate the plates are Y-shaped, and the spacing of the plates increases anteriorly. Three plates at one end of the tooth are arranged more vertically which indicates that this is the posterior end of the tooth; the rest of the plates lean and converge apically which indicates that this is the anterior end of the tooth. The crown is higher at the second and third plates from the posterior end. It is difficult to determine if the cingulum is present as the anterior end is broken. However, a cingulum is observed fully attached to the first plate from the posterior end. There are no observable roots from this specimen. Plate dimensions of all three specimens are summarized in Table 2.
Geochronology
The pIR-IRSL dating of the breccia matrix constrained the tooth to 233 ± 31 ka according to 12 single aliquots of feldspar and 199 ± 28 ka based on 116 single grains of feldspars. These results are coeval with error margins and reflect the difference in approaches between the aliquots and single grains methods. Due to the large number of grains on single aliquot discs an averaging effect occurs on the luminescence signal. Thus, the single aliquot result due to its opportunity to observe partially bleached grains is usually slightly younger and more reliable for age determination.
 Only one age was obtained (see Table 3) from the U-series dated sample, since the other ones contain too much detritus to give any reasonable results. The concentrations of 238U and 232Th in the sample are ~171 ppb and ~114 ppb respectively. The yield age is 35184 ± 9485 yr BP.
Only one age was obtained (see Table 3) from the U-series dated sample, since the other ones contain too much detritus to give any reasonable results. The concentrations of 238U and 232Th in the sample are ~171 ppb and ~114 ppb respectively. The yield age is 35184 ± 9485 yr BP.
The combined in-situ sediment dating of the breccia matrix and overlying flowstone reveal that the fossil was deposited in this location before 35 ± 10 ka (U-series dating of flowstone, see Table 3) and between 171-264 ka (233 ± 31-199 ± 28 ka) (pIR-IRSL dating of feldspars, see Table 4) with examples of the pIR-IRSL dating of feldspars data presented in Figure 9. The two dating results agree with each other and are consistent with their stratigraphic location. Due to the error margin associated with the pIR-IRSL age estimate there is a good chance that the Stegodon individual died within this age range and was later deposited within the cave breccia.
DISCUSSION
PRK_SC_P.01 remains the only and most well-preserved Stegodon specimen from Peninsular Malaysia. The discovery of the Stegodon fossil here is a testimony to the wide distribution range of this genus during middle Pleistocene time in the Sundaic subregion. Even though large parts of the tooth are still embedded in matrix, the use of micro-CT is helpful in revealing the full morphology of the specimen for comparison to be made with related middle Pleistocene non-dwarf species of Stegodon found in the Sundaic subregion (Table 1). The age range for this individual based on the pIR-IRSL and U-series chronology lies within the expected age range for Stegodon found on mainland Southeast Asia.
Differences and similarities between PRK_SC_P.01 (Stegodon sp.) and the dp3s of S. trigonocephalus and S. orientalis
The posterior cingulum of KL DP3 is only very low and weakly developed, while in PRK_SC_P.01 and the S. trigonocephalus specimen examined, the posterior cingulum almost resembles all the other true plates. However, this might be due to individual variation in the development of these terminal plates or talons. KL DP3 may resemble some specimens from mainland China in having five ridges and a small talon (posterior cingulum), but PRK_SC_P.01 and CD 11649 seem to resemble the condition reported for some dp4 specimens from Yenchingkou and Paxian Dadong, in which the cingulum is so well developed that it can be considered as a fully developed plate (e.g., A.M.N.H. no. 18705, described in Osborn, 1942; Colbert and Hooijer, 1953; Schepartz et al., 2005). However, the anterior or posterior cingulum in these Chinese dp4 specimens do not seem to have any associated row of small enamel tubercles as is in PRK_SC_P.01.
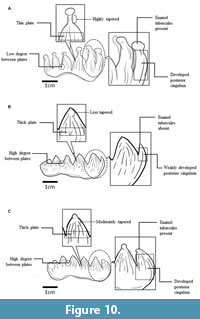 The enamel tubercles on the apical of the plates in PRK_SC_P.01 are larger as compared to those of S. trigonocephalus but these tubercles are smaller and less developed in the S. orientalis specimen examined. Towards the lower end of the enamel tubercles, the plates of PRK_SC_P.01 are highly tapered, less so in the S. trigonocephalus and S. orientalis specimens. PRK_SC_P.01 conelets are smaller and less bulbous compared to those of S. orientalis. From lingual view, PRK_SC_P.01 has gracile plates and higher crowns as compared to the S. trigonocephalus and S. orientalis specimens. The valley between the plates of PRK_SC_P.01 is shallower compared to both S. trigonocephalus and S. orientalis. These features are summarized in Table 5 and Figure 10.
The enamel tubercles on the apical of the plates in PRK_SC_P.01 are larger as compared to those of S. trigonocephalus but these tubercles are smaller and less developed in the S. orientalis specimen examined. Towards the lower end of the enamel tubercles, the plates of PRK_SC_P.01 are highly tapered, less so in the S. trigonocephalus and S. orientalis specimens. PRK_SC_P.01 conelets are smaller and less bulbous compared to those of S. orientalis. From lingual view, PRK_SC_P.01 has gracile plates and higher crowns as compared to the S. trigonocephalus and S. orientalis specimens. The valley between the plates of PRK_SC_P.01 is shallower compared to both S. trigonocephalus and S. orientalis. These features are summarized in Table 5 and Figure 10.
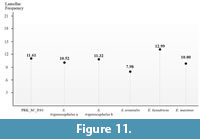 PRK_SC_P.01 shared some of the common morphological features with S. trigonocephalus and S. orientalis – greater number of plates than those of primitive Stegodon species such as S. zdanskyi), widely spaced ridges, vertical grooves along the lingual and buccal surfaces, and relatively low crowned. However, metrical analysis shows that PRK_SC_P.01 has different parameters and measurements from these two species. PRK_SC_P.01 has high lamellar frequency values (LF), 11.61 (Figure 11) which is quite different from the dp3 of S. orientalis (KL DP3) that has a relatively low value of 7.98. However, as compared to both S. trigonocephalus specimens from Java (CD 11647a and CD 11647b), PRK_SC_P.01 shows almost similar in terms of LF values as both Java specimens which have high LF, 10.52 and 11.32, respectively. As compared to the genus Elephas, PRK_SC_P.01 shows a higher value than the living Asian elephant (E. maximus) but lower than the extinct E. hysudricus.
PRK_SC_P.01 shared some of the common morphological features with S. trigonocephalus and S. orientalis – greater number of plates than those of primitive Stegodon species such as S. zdanskyi), widely spaced ridges, vertical grooves along the lingual and buccal surfaces, and relatively low crowned. However, metrical analysis shows that PRK_SC_P.01 has different parameters and measurements from these two species. PRK_SC_P.01 has high lamellar frequency values (LF), 11.61 (Figure 11) which is quite different from the dp3 of S. orientalis (KL DP3) that has a relatively low value of 7.98. However, as compared to both S. trigonocephalus specimens from Java (CD 11647a and CD 11647b), PRK_SC_P.01 shows almost similar in terms of LF values as both Java specimens which have high LF, 10.52 and 11.32, respectively. As compared to the genus Elephas, PRK_SC_P.01 shows a higher value than the living Asian elephant (E. maximus) but lower than the extinct E. hysudricus.
From comparative morphological studies with other Stegodon specimens (S. trigonocephalus and S. orientalis) PRK_SC_P.01 differs in morphology from S. orientalis and less so from S. trigonocephalus. Due to this limitation and the observed morphological differences, it is currently not possible to assign a definite species to PRK_SC_P.01. Further comparisons with other Stegodon species both continental and island species are needed to address the exact taxonomic status of this specimen.
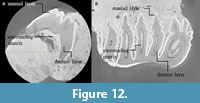 The lack of cementum from the μCT images of PRK_SC_P.01 might indicate that this layer is yet to be developed as it is still a milk tooth. The absence of cementum, a common occurrence in the early stages of tooth development in mammals, occurs because this layer is formed much later than the enamel and dentin layers (Foster, 2017). The cementum has a close anatomical relationship to the root dentine, which is not yet fully developed in PRK_SC_P.01 (see Figure 12).
The lack of cementum from the μCT images of PRK_SC_P.01 might indicate that this layer is yet to be developed as it is still a milk tooth. The absence of cementum, a common occurrence in the early stages of tooth development in mammals, occurs because this layer is formed much later than the enamel and dentin layers (Foster, 2017). The cementum has a close anatomical relationship to the root dentine, which is not yet fully developed in PRK_SC_P.01 (see Figure 12).
Most of the continental Stegodon fossils that were discovered in middle to late Pleistocene of Southeast Asia were assigned either definitely or tentatively to the species S. orientalis, except for some examples discovered in a middle Pleistocene site in Tam Nang, Laos (Arambourg and Fromaget, 1938), which were identified as belonging to S. sinensis. According to Chen (1999), remains of Stegodon from southern China were mostly assigned to S. orientalis. Our results of morphological comparison indicated that PRK_SC_P.01 exhibits morphological characters that are different from S. orientalis. This implies that either the species is highly variable in the characters considered here (Table 5) or, alternatively, there might be a greater level of taxonomoic diversity of Stegodon in middle to late Pleistocene continental Southeast Asia that has yet to be fully recognized.
Biogeographic implications
Stegodon remains have been found on mainland Southeast Asia, ranging in age from earlier Pleistocene, such as in the Irrawaddy beds, Myanmar (Colbert, 1943), to late Pleistocene deposits in Vietnam, Myanmar, and Laos (Olsen and Ciochon, 1990; Zin-Maung-Maung-Thein et al., 2011; Bacon et. al, 2018). The oldest documented evidence of S. orientalis comes from Dayakou, located in southwest China, while S. trigonocephalus was first recorded in Ci Saat, Java (Sondaar, 1984; van den Bergh et al., 2001a). These two species of Stegodon were, therefore, determined to have evolved during the earlier phase of Pleistocene period. As for the Stegodon from island Southeast Asia, their remains were found from early Pleistocene in Ci Saat up to the late Pleistocene deposits in Liang Bua (Flores) (van den Bergh et al., 2001a, 2008).The age range for PRK_SC_P.01 based on the pIR-IRSL and U-series chronology lies within the expected age range for Stegodon found on mainland Southeast Asia during the middle Pleistocene. The middle Pleistocene of the Tham Khuyen and Tham Hai in Vietnam, where S. orientalis specimens were discovered, indicate between roughly 300,000 to 200,000 years old based on the dating of biostratigraphic ground (Long and Du, 1981). Remains of S. trigonocephalus had also been reported from middle Pleistocene sites in Java, including Kedung Brubus (0.8 - 0.7 Ma, based on the faunal assemblages [van den Bergh et al., 2001b]), Ngandong (142 - 92 ka, according to thermoluminescence, pIR-IRSL, and 40Ar/39Ar methods [Rizal et al., 2020]), Trinil HK (0.54 ± 0.10 Ma - 0.43 ± 0.05 Ma, based on 40Ar/39 Ar, palaeomagnetic and luminescence dating method [Joordens et al., 2015]), and Trinil HK Bone Bearing Channel 2 (450 ± 110 ka - 430 ± 50 ka [Hilgen et al., 2023]).
The discovery of Stegodon sp. reported here represents the geologically youngest fossil evidence of Stegodon found in the Malay Peninsula, specifically in the southern region below the Kra Isthmus. This isthmus serves as a demarcation between the Indochinese and Sundaic subdivisions of the contemporary Southeast Asian zoogeographical regions (Corbet and Hill, 1992). PRK_SC_P.01 represents the southernmost Stegodon in continental Asia, situated amidst two prominent groups of Stegodon. Although the Stegodon specimen from Peninsular Malaysia exhibits distinct morphologies as compared to the other two main taxa, it is challenging to ascertain whether this specimen represents a transitional species between the continental and island forms of Stegodon. This difficulty arises due to the limited information and the singular nature of the Stegodon specimen from Peninsular Malaysia. PRK_SC_P.01 does not show any distinctive morphologies that can help to identify whether it is a continental or insular species from the Sundaland during the middle Pleistocene. Regardless, by comparing the size of PRK_SC_P.01 to both S. orientalis and S. trigonocephalus specimens, PRK_SC_P.01 does not exhibit any dwarfism in size. PRK_SC_P.01 affirms that there were populations of this genus in the region, however, it remains unknown whether they were the direct descendants of either S. orientalis (continental species) or S. trigonocephalus (insular species), or from a transitional species as both of these species were found in early Pleistocene times.
Stegodon and palaeoenvironment
Dental microwear analysis of S. orientalis from southern China shows that the species was primarily browsers in a more densely forested landscape (Zhang et al., 2017). This is different from the S. trigonocephalus of Java which shows that the species was mostly grazers in an environment dominated by a mix of open savannah woodlands and evergreen forests (Puspannigrum et. al, 2020). Securely dated middle Pleistocene fossil sites from Peninsular Malaysia are few in number, but all have yielded mammalian remains from taxa that are known to have included forest environment as part of their natural habitats in modern times (Ibrahim et al., 2013). Indicative taxon frequently used for palaeoenvironmental reconstruction such as Pongo had been reported from all of these sites in Peninsular Malaysia (Ibrahim et al., 2013) and also from the Stegodon Cave (Muhammad et al., 2020). The presence of Pongo among fossil mammal assemblages was previously taken as a definite indication of an environment that was predominantly forest (Tougard, 1998; Storm et al., 2005). However, there is growing evidence from recent studies which showed that modern representatives of the genus can not only live in continuous closed-canopy rainforests but are also capable of surviving in altered landscapes dominated by secondary forests and farmlands (Wich et al., 2008; Oram, 2018; Sabah Wildlife Department, 2020; Ancrenaz et al., 2021), with some of the fossil Pongo exhibited isotopic signals indicating broader dietary spectrum and being discovered in drier environments from the end of the Last Interglacial until the late Holocene (Westaway et al., 2007; Louys et al., 2021, 2022). As a result, the presence of Pongo within the same stratum of PRK_SC_P.01 cannot now in itself be regarded as an unequivocal evidence of the predominant occurrence of closed-canopy tropical rainforests (to the exclusion of other similar environments) in the area during middle Pleistocene times. However, based on our current knowledge of the ecological tolerance of modern Pongo and isotopic evidence of fossil Pongo and Stegodon from elsewhere in Southeast Asia and adjacent areas, it is reasonable to suggest that the palaeoenvironment of the Stegodon Cave site during middle Pleistocene may have been either under continuous forest cover or a mixed landscape with patches of forests interspersed among more opened vegetation. Nevertheless, more comprehensive studies are needed to properly reconstruct the detailed palaeoenvironment in this region, since mammal assemblages that were discovered in Stegodon Cave were the results of upbringing from the stream that flows along the periphery of the cliff. Further analysis of the palaeoenvironment could enhance in identifying the correlation of PRK_SC_P.01 to the S. orientalis or S. trigonocephalus, or perhaps a species that transitions between the continental or islandic species.
CONCLUSIONS
The specimen PRK_SC_P.01 represents the only known record of Stegodon from Peninsular Malaysia. This late middle Pleistocene specimen shows a unique combination of characters as compared to two species of Stegodon found in the region, namely S. orientalis and S. trigonocephalus, which lived during the early to late Pleistocene. Characters unique to PRK_SC_P.01 include a strongly developed posterior cingulum as compared to S. orientalis from Vietnam, but slightly less developed than in S. trigonocephalus from Java. It also has enamel tubercles present at the posterior cingulum, and highly tapered plates at the lingual side of the tooth. As compared to both S. orientalis and S. trigonocephalus, the specimen from Peninsular Malaysia has thinner plates with shallower valleys between them. The discovery of Stegodon in Peninsular Malaysia indicates there were representatives of this genus in the region at least before 199 ± 28 and 233 ± 31 ka. It remains unclear whether this middle Pleistocene Stegodon represented the direct descendants of the continental species from the north or the insular species from the south.
ACKNOWLEDGEMENTS
This research was supported by GPF021B-2018: Studies On Middle-Late Pleistocene Vertebrate Teeth Fossils In Karst In Peninsular Malaysia and IIRG004A-2020FNW: Top Down: Evolutionary Dynamics Study Of Quaternary Faunal Remains From Karstic Landscapes Of Peninsular Malaysia, both provided by Universiti Malaya awarded to RFM. The luminescence dating aspects of the project were funded by an ARC Discovery grant DP230100440 to KEW.
Our team would like to thank members of Kinta Valley Watch (KVW), especially B.T. Ching , C.Y. Tee, Y.L. Nang, J.C. Chan, and J. F. Chan who first discovered the fossil and reported to the authority. Appreciation goes to the 5th Earl of Cranbrook (G. Gathorne-Hardy), J. de Vos, G. van den Bergh, G. Drawhorn, C. Chang, P. Piper, A. Wattanapituksakul, S.Z.H. Wen, J. Ma, and J. Duangkrayom for their help with earlier identification of the specimen from Peninsular Malaysia. We sincerely acknowledge Dr. C.-C. Shen of the Department of Geosciences, National Taiwan University, for the approval of U-Th dating facilities. AHA expresses his gratitude to the Vietnam Institute of Archaeology (Hanoi) for the opportunity to examine their Stegodon fossils, to L.K. Chian Natural History Museum (LKCNHM, National University of Singapore) for the opportunity to examine their Elephas maximus specimen, to M. Tablizo, M. Puspaningrum, and H.Q. Le for their valuable insights concerning Stegodon in the Philippines, Indonesia and Vietnam, and to Z. Zahari for his assistance in arranging and formatting the figures in this manuscript. RFM is grateful to N.I. Taib for the help with the fossil 3D model printing, F.A. Zulkifli for helping with the GIS, F.M. Razif and Z. Kem who kindly helped in the field and to M.N.A. Rahman, N.A. Md Nur from the Department of Mineral and Geoscience Malaysia for research permission and S.R. Hussein and A.M. Yazrol from the Perak State Parks Corporation for facilitating the fossil extraction. LTT would like to thank Vereniging Nederland-Maleisië (VNM) for a research grant to the Netherlands, Naturalis Biodiversity Center for permission to study their Stegodon collection, and to H. van de Bunte, N. den Ouden, and A. van Westbroek for their hospitality, technical support and assistance.
REFERENCES
Aiba, H., Baba, K., and Matsukawa, M. 2010. A new species of Stegodon (Mammalia, Proboscidea) from the Kazusa Group (lower Pleistocene), Hachioji City, Tokyo, Japan and its evolutionary morphodynamics. Palaeontology, 53:47-490.
https://doi.org/10.1111/j.1475-4983.2010.00953.x
Aitken, M.J. 1998. An Introduction to Optical Dating: the Dating of Quaternary Sediments by the Use of Photon-Stimulated Luminescence. Oxford University Press, Oxford, United Kingdom.
https://doi.org/10.1093/oso/9780198540922.001.0001
An, Z., Gao, W., Zhu, Y., Kan, X., Wang, J., Sun, J., and Wei, M. 1990. Magnetostratigraphic dates of Lantian Homo erectus. Acta Anthropologica Sinica, 9:1-7.
Ancrenaz, M., Oram, F., Nardiyono, N., Silmi, M., Jopony, M.E.M., Voigt, M., Seaman, D.J.I., Sherman, J., Lackman, I., Traeholt, C., Wich, S.A., Santika, T., Struebig, M.J., and Meijaard, E. 2021. Importance of small forest fragments in agricultural landscapes for maintaining orangutan metapopulations. Frontiers in Forests and Global Change, 4:560944.
https://doi.org/10.3389/ffgc.2021.560944
Ao, H., Zhang, P., Dekkers, M.J., Roberts, A.P., Zhinseng A., Yongxiang L., Fengyan L., Lin S., and Li, X. 2016. New magnetochronology of Late Miocene mammal fauna, NE Tibetan Plateau, China: mammal migration and paleoenvironments. Earth and Planetary Science Letters, 434:220-230.
https://doi.org/10.1016/j.epsl.2015.11.019
Arambourg, C. and Fromaget, J. 1938. Le gisement quaternaire de Tam Nang (Chaîne Annamitique septentrionale). Sa stratigraphie et ses faunes. Comptes Rendus de l'Academie des Sciences, 207:793-795.
Aslam, S., Iqbal, A., Khan, A., Akbar khan, M., and Akhtar, M. 2015. New fossil remains of Stegodon bombifrons (Proboscideans) from the Pinjor Stage Soan Formation of Pakistan. Biologia (Lahore, Pakistan), 61:101-106.
Bacon, A.M., Demeter, F., Tougard, C., de Vos, J., Sayavongkhamdy, T., Antoine, P.O., Bouasisengpaseuth, B., and Sichanthongtip, P. 2008. Redécouverte d’une faune Pléistocène dans les remplissages karstiques de Tam Hang au Laos: premiers résultats. Comptes Rendus Palevol, 7:277-288.
https://doi.org/10.1016/j.crpv.2008.03.009
Bacon, A.M., Duringer, P., Antoine, P.O., Demeter, F., Shackelford, L., Sayavongkhamdy, T., Sichanthongtip, P., Khamdalavong, P., Nokhamaomphu, S., Sysuphanh, V., Patole-Edoumba, E., Chabaux, F., and Pelt, E. 2011. The Middle Pleistocene mammalian fauna from Tam Hang karstic deposit, northern Laos: new data and evolutionary hypothesis. Quaternary International, 245:315-332.
https://doi.org/10.1016/j.quaint.2010.11.024
Bacon A.M., Demeter, F., Duringer, P., Patole-Edoumba, E., Sayavongkhamdy, T., Coupey, A.S., Shackelford, L., Westaway, K.E., Ponche, J.L., Antoine, P.O., and Sichanthongtip, P. 2012. Les sites de Tam Hang, Nam Lot et Tam Pà Ling au nord du Laos: des gisements à vertébrés du Pléistocène aux origines des hommes modernes. CNRS Editions:149.
Bacon, A.M., Westaway, K.E., Antoine, P.O., Duringer, P., Blin, A., Demeter, F., Ponche, J.L., Zhao, J.X., Barnes, L.M., Sayavonkhamdy, T., Thuy, N.T.K., Long, V.T., Patole-Edoumba, E., and Shackelford, L. 2015. Late Pleistocene mammalian assemblages of Southeast Asia: new dating, mortality profiles and evolution of the predator-prey relationships in an environmental context. Palaeogeography, Palaeoclimatology, Palaeoecology, 422:101-127.
https://doi.org/10.1016/j. palaeo.2015.01.011
Bacon, A.M., Bourgon, N., Dufour, E., Zanolli, C., Duringer, P., Ponche, J.L., Antoine, P.O., Shackelford, L., Huong, N.T.M., and Sayavonkhamdy, T. 2018. Nam lot (MIS 5) and Duoi U’Oi (MIS 4) Southeast Asian sites revisited: zooarchaeological and isotopic evidences. Palaeogeography, Palaeoclimatology, Palaeoecology, 512:132-144.
https://doi.org/10.1016/j.palaeo.2018.03.034
Batchelor, B.C. 1979a. Discontinuously rising Late Cainozoic eustatic sea-levels with special reference to Sundaland, Southeast Asia. Geologie en Mijnbouw, 58:1-20.
https://doi.org/10.1016/0198-0254(79)90729-5
Batchelor, B.C. 1979b. Geological characteristics of certain coastal and offshore placers as essential guides for tin exploration in Sundaland, Southeast Asia. Bulletin of the Geological Society of Malaysia, 11:283-313.
https://doi.org/10.7186/bgsm11197913
Batchelor, D.A. 2015. Clarification of stratigraphic correlation and dating of Late Cainozoic alluvial units in Peninsular Malaysia. Bulletin of the Geological Society of Malaysia, 61:75-84.
https://doi.org/10.7186/bgsm61201508
Brennan, B.J. 2003. Beta doses to spherical grains. Radiation Measurements, 37:299-303.
https://doi.org/10.1016/s1350-4487(03)00011-8
Bøtter-Jensen, L. and Mejdahl, V. 1988. Assessment of beta dose-rate using a GM multicounter system. International Journal of Radiation Applications and Instrumentation. Part D. Nuclear Tracks and Radiation Measurements, 14:187-191.
https://doi.org/10.1016/1359-0189(88)90062-3
Chen, G. 1999. Sinomastodon Tobien, et al., 1986 (Proboscidea, Mammalia) from the Pliocene and Early-Middle Pleistocene of China. Proceedings of the Seventh Annual Meeting of the Chinese Society of Vertebrate Paleontology, Beijing, China, p. 179-187. (in Chinese with English summary)
Chen, G. 2021. Palaeovertebrata Sinica. Volume III Basal Synapsids and Mammals. Fascicle 10 (Serial No. 23) Hyracoidea and Proboscidea. Science Press, Beijing.
Cheng, H., Edwards, R.L., Shen, C.C., Polyak, V.J., Asmerom, Y., Woodhead, J., Hellstrom, J., Wang, Y.J., Kong, X., Spotl, C., Wang, X.F., and Alexander Jr., E. 2013. Improvements in 230Th dating, 230Th and 234U half-life values, and U-Th isotopic measurements by multi-collector inductively coupled plasma mass spectrometry. Earth and Planetary Science Letters, 371-372:82-91.
https://doi.org/10.1016/j.epsl.2013.04.006
Ciochon, R.L. 2009. The mystery ape of Pleistocene Asia. Nature, 459:910-911.
https://doi.org/10.1038/459910a
Clift, W. 1828. On the fossil remains of two new species of Mastodon, and of other vertebrated animals, found on the left bank of the Irawadi. Transactions of the Geological Society of London, 2:369-376.
https://doi.org/10.1144/transgslb.2.3.369
Colbert, E.H. 1943. Pleistocene vertebrates collected in Burma by the American Southeast Asia Expedition. Transactions of the American Philosophical Society, 32:395-429.
https://doi.org/10.70249/9798893983203-006
Colbert, E.H. and Hooijer, D.A. 1953. Pleistocene mammals from the limestone fissures of Szechwan, China. Bulletin of the American Museum of Natural History, 10:7-134.
Corbet, G.B. and Hill, J.E. 1992. The Mammals of the Indomalayan Region. Oxford University Press, USA.
Cornivus, G. 1988. The Mio-Plio-Pleistocene litho- and biostratigraphy of the Surai Khola Siwaliks in West Nepal: first results. Comptes Rendus de l'Académie des Sciences Paris, 306:1471-1477.
Cuong, N.L. 1985. Fossile menschenfunde aus nordvietnam, p. 96-102. In Herrmann J. and Ullrich, H. (eds.), Menschwerdung-Biotischer und Gesellschaftlicher Entwicklungsprozess. Akademieverlag, Berlin, Germany.
Davison, G.W.H. 1994. Some remarks on vertebrate remains from the excavation at Gua Gunung Runtuh, Perak, p. 141-148. In Majid, Z. (ed.), The Excavation of Gua Gunung Runtuh and the Discovery of the Perak Man in Malaysia. Department of Museums and Antiquity Malaysia, Kuala Lumpur.
Delfino, M. and de Vos, J. 2010. A revision of the Dubois crocodylians, Gavialis bengawanicus and Crocodylus ossifragus, from the Pleistocene Homo erectus beds of Java. Journal of Vertebrate Paleontology, 30:427-441.
https://doi.org/10.1080/02724631003617910
de Vos, J. 1995. The migration of Homo erectus and Homo sapiens in South East Asia and the Indonesian archipelago, p. 239-260. In Bower, J.R.F and Sartono, S. (eds.), Human Evolution on the Ecological Context, Volume I, Evolution and Ecology of Homo erectus. Pithecanthropus Centennial Foundation, Leiden University, Netherlands.
de Vos, J. 2007. Vertebrate records: Mid-Pleistocene of southern Asia, p. 3232-3249. In Elias, S.A. (ed.), Encyclopedia of Quaternary Science. Elsevier, Oxford.
https://doi.org/10.1016/b0-44-452747-8/00256-8
de Vos, J. and Long, V.T. 1993. Systematic discussion of the Lang Trang fauna. Unpublished report.
de Vos, J. and Long, V.T. 2001. First settlements: relations between continental and insular Southeast Asia, p. 225-249. In Sémah, F., Falguères, C., Grimaud-Hervé, D., and Sémah, A.M. (eds.) Origine des Peuplements et Chronologie des Cultures Paléolithiques dans le Sud-Est Asiatique. Semenanjung/Artcom, Paris.
de Vos, J., van den Hoek Ostende, L.W., and van den Bergh, G.D. 2007. Patterns in insular evolution of mammals: a key to island palaeogeography, p. 315-345. In Renema, W. (ed.), Biogeography, Time and Place: Distributions, Barriers and Islands. Springer, Dordrecht, Netherlands.
https://doi.org/10.1007/978-1-4020-6374-9_10
Dietrich, W.O. 1926. Zur Altersbestimmung der Pithecanthropus Schichten. Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin for 1924, p. 134-139.
Duangkrayom, J., Nishioka, Y., Saokan, C., Jintasakul, P., Thungprue, N., and Worawansongkham, R. 2017. Proboscidean fossils (Mammalia) from the Quaternary deposits on Stegodon Cave, Thungwa, Satun Province, southern Thailand. Waseda Institute for Advanced Study Discussion Paper, No.2018-001:1-19.
Dubois, E. 1908. Das geologischer Alter der Kendeng- oder Trinil-Fauna. Tijdschrift van het Koninklijk Nederlandsch Aardrijskunding Genootschap, 2:1235-1270.
Edwards, R.L., Chen, J.H., and Wasserburg, G.J. 1987. 238U-234U-230Th-232Th systematics and the precise measurement of time over the past 500,000 years. Earth and Planetary Science Letters, 81:175-192.
https://doi.org/10.1016/0012-821X(87)90154-3
Falconer, H. 1857. On the species of Mastodon and elephant occurring in the fossil state in Great Britain, Part I, Mastodon. Quarterly Journal of the Geological Society of London, 13: 307-360.
https://doi.org/10.1144/gsl.jgs.1857.013.01-02.43
Falconer, H. and Cautley, P.T. 1846. Fauna Antiqua Sivalensis, Part I, Proboscidea. Smith, Elder and Co., London.
https://doi.org/10.5962/bhl.title.61447
Foster, B.L. 2017. On the discovery of cementum. Journal of Periodontal Research, 52: 666-685.
https://doi.org/10.1111/jre.12444
Ghani, A.A., Searle, M., Robb, M.L., and Sun-Lin Chung, S. 2013. Transitional I S type characteristic in the Main Range Granite, Peninsular Malaysia. Journal of Asian Earth Sciences, 76:225-240.
https://doi.org/10.1016/j.jseaes.2013.05.013
Guérin, G., Mercier, N., and Adamiec, G. 2011. Dose rate conversion factors: update. Ancient TL, 29:5-11.
https://doi.org/10.26034/la.atl.2011.443
Hassan, K., Nakamura, T., Price, D.M., Woodroffe, C.D., and Fujii, S. 1993. Radiocarbon and thermoluminescence dating of the Old Alluvium from a coastal site in Perak, Malaysia. Sedimentary Geology, 83:199-210.
https://doi.org/10.1016/0037-0738(93)90013-u
Henri, F. and Amnan, I. 1994. Biostratigraphy of Kinta Valley. Bulletin of the Geological Society of Malaysia, 58:159-172.
https://doi.org/10.7186/bgsm38199515
Hilgen, S.L., Pop, E., Adhityatama, S., Veldkamp, T.A., Berghuis, H.W.K., Sutisna, I., Yur-naldi, D., Dupont-Nivet, G., Reimann, T., Nowaczyk, N., Kuiper, K.F., Krijgsman, W., Vonhof, H.B., Ekowati, D.R., Alink, G., Hafsari, N.L.G.D.M., Drespriputra, O., Verpoorte, A., Bos, R., Simanjuntak, T., Prasetyo, B., and Joordens, J.C.A. 2023. Revised age and stratigraphy of the classic Homo erectus -bearing succession at Trinil (Java, Indonesia). Quaternary Science Reviews, 301:107908.
https://doi.org/10.1016/j.quascirev.2022.107908
Hooijer, D.A. 1954. Pleistocene Vertebrates from Celebes XI. Molars and a tusked mandible of Archidiskodon celebensis Hooijer. Zoologische Mededelingen, 33:103-120.
Hooijer, D.A. 1955. Fossil Proboscidea from the Malay Archipelago and India. Zoologische Verhandelingen, 28:1-146.
Hooijer, D.A. 1962. Report upon a collection of Pleistocene mammals from tin-bearing deposits in a limestone cave near Ipoh, Kinta Valley, Perak. Federation Museums Journal, 7:1-5.
Huntley, D.J. and Baril, M.R. 1997. The K content of the K-feldspars being measured in optical dating or in thermoluminescence dating. Ancient TL, 15:11-13.
https://doi.org/10.26034/la.atl.1997.271
Huntley, D.J. and Hancock, R. 2001. The Rb contents of the K-feldspar grains being measured in optical dating. Ancient TL, 19:43-46.
https://doi.org/10.26034/la.atl.2001.333
Hutt, G., Jaek, I., and Tchonka, J. 1988. Optical dating: K-feldspars optical response stimulation spectra. Quaternary Science Reviews, 7:381-385.
https://doi.org/10.1016/0277-3791(88)90033-9
Ibrahim, K.I., Lim, T.T., Westaway, K.E., of Cranbrook, E., Humphrey, L., Muhammad, R.F., and Peng, L.C. 2013. First discovery of Pleistocene orangutan (Pongo sp.) fossils in Peninsular Malaysia: biogeographic and paleoenvironmental implications. Journal of Human Evolution, 65:770-797.
https://doi.org/10.1016/j.jhevol.2013.09.005
Ishlahuda, H.S., Lim, T.T., Muhammad, R.F., Nur, S.I.A.T., and Mohammad, A.A.A. 2019. First systematic study of Late Pleistocene rat fossils from Batu Caves: new record of extinct species and biogeography implications. Sains Malaysiana, 48:2613-2622.
https://doi.org/10.17576/jsm-2019-4812-02
Ingicco, T., van den Bergh, G.D., Jago-on, C., Bahain, J.-J., Chacón, M.G., Amano, N., Forestier, H., King, C., Manalo, K., Nomade, S., Pereira, A., Reyes, M. C., Semah, A. M., Shao, Q., Voinchet, P., Falgueres, C., Albers, P.C.H., Lising, M., Lyras, G., Yurnaldi, D., Rochette, P., Bautista, A., and de Vos, J. 2018. Earliest known hominin activity in the Philippines by 709 thousand years ago. Nature, 557:233-237.
https://doi.org/10.1038/s41586-018-0072-8
Jaffey, A.H., Flynn, K.F., Glendenin, L.E., Bentley, W.C., and Essling, A.M. 1971. Precision measurement of half-lives and specific activities of 235U and 238U. Physical Review C, 4:1889-1906.
https://doi.org/10.1103/PhysRevC.4.1889
Janensch, W. 1911. Die Proboscidier-Schaedel der Trinil-Expeditions-Sammlung, p. 151-195. In Selenka, L. and Blanckenhorn, M. (eds.), Die Pithecanthropus -Schichten auf Java. Leipzig, Engelmann.
https://doi.org/10.5962/bhl.title.60936
Jensen, B.J.L., Dufrane, A., Mark, D., Zaim, Y., Rizal, Y., Aswan, A., Hascary, A., Ciochon, R., Gunnell, G., Larick, R., and Zonneveld, J.P. 2017. Newly described tephra provide age constraints to Stegodon fossils in west (Indonesian) Timor. American Geophysical Union, Fall Meeting.
Joordens, J.C.A., d’Errico, F., Wesselingh, F.P., Munro, S., de Vos, J., Wallinga, J., Ankjærgaard, C., Reimann, T., Wijbrans, J.R., Kuiper, K.F., Mücher, H.J., Coqueugniot, H., Prié, V., Joosten, I., van Os, B., Schulp, A.S., Panuel, M., van der Haas, V., Lustenhouwer, W., Reijmer, J.G.J., and Roebroeks, W. 2015. Homo erectus at Trinil on Java used shells for tool production and engraving. Nature, 518:228-231.
https://doi.org/10.1038/nature13962
Kahlke, H.D. 1961. On the complex of the Stegodon-Ailuropoda fauna of southern China and the chronological position of Gigantopithecus blacki v. Koenigswald. Vertebrata PalAsiatica, 6:83-108.
Kha, L.T. 1976. First remarks on the Quaternary fossil fauna of northern Vietnam. Vietnamese Studies, 46:107-126.
Kikinis, R., Pieper, S.D., and Vosburgh, K.G. 2014. 3D slicer: a platform for subject-specific image analysis, visualization, and clinical support, p. 277-289. In Jolesz, F. (ed.), Intraoperative Imaging and Image-Guided Therapy. Springer, New York.
https://doi.org/10.1007/978-1-4614-7657-3_19
Konishi, S. and Yoshikawa, S. 1999. Immigration times of the two proboscidean species, Stegodon orientalis and Palaeoloxodon naumanni, into the Japanese islands and the formation of land bridge. Earth Science (Chikyu Kagaku), 53:125-134 (in Japanese).
Kundal, S., Bhadur, G., and Kumar, S. 2017. A Late Pliocene baby Stegodon cf. Stegodon insignis (Proboscidea) from Upper Siwalik of Samba District, Jammu and Kashmir, India. Earth Science India, 10:82-93.
Lambard, J., Pereira, A., Voinchet, P., Guillou, H., Reyes, M.C., Nomade, S., Gallet, X., Belarmino, M., Bahain, J., de Vos, J., Falguères, C., Cosalan, A., and Ingicco, T. 2024. Geochronological advances in human and proboscideans first arrival date in the Philippines archipelago (Cagayan valley, Luzon Island). Quaternary Geochronology, 84:101597.
https://doi.org/10.1016/j.quageo.2024.101597
Lim, T.T. 2013. Quaternary Elephas from Peninsular Malaysia: historical overview and new material. Raffles Bulletin of Zoology, Supplement No. 29:139-153.
Lin, C.C. 1963. Geology and ecology of Taiwan prehistory. Asian Perspectives, 7:203-213.
Long, V.T. and Du, H.V. 1981. Zoological species belonging to the Pleistocene and the geochronology of sediments containing them in caves and grottos in northern Viet Nam. Khao Co Hoc, 1:16-19 (in Vietnamese).
Long, V.T., de Vos, J., and Ciochon, R.S. 1996. The fossil mammal fauna of the Lang Trang caves, Vietnam, compared with Southeast Asian fossil and recent mammal faunas: the geographical implications. Bulletin of the Indo-Pacific Prehistory Association, 14:101-109.
Louys, J., Curnoe, D., and Tong, H.W. 2007. Characteristics of Pleistocene megafauna extinctions in Southeast Asia. Palaeogeography, Palaeoclimatology, Palaeoecology, 243:152-173.
https://doi.org/10.1016/j.palaeo.2006.07.011
Louys, J., Price, G.J., and O’Connor, S. 2016. Direct dating of Pleistocene Stegodon from Timor island, East Nusa Tenggara. PeerJ, 4:1788.
https://doi.org/10.7717/peerj.1788
Louys, J., Zaim, Y., Rizal, Y., Aswan, Puspaningrum, M., Trihascaryo, A., Price G.J., Petherick, A., Scholtz, E., and DeSantis, L.R.G. 2021. Sumatran orangutan diets in the Late Pleistocene as inferred from dental microwear texture analysis. Quaternary International, 603:74-81.
https://doi.org/10.1016/j.quaint.2020.08.040
Louys, J., Duval, M., Price, G.J., Westaway, K., Zaim, Y., Rizal, Y., Aswan, Puspaningrum, M., Trihascaryo, A., Breitenbach, S.F.M., Kwiecien, O., Cai, Y., Higgins, P., Albers, P.C.H., de Vos, J., and Roberts, P. 2022. Speleological and environmental history of Lida Ajer cave, western Sumatra. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 377:20200494.
https://doi.org/10.1098/rstb.2020.0494
Ma, A. and Tang, H. 1992. On discovery and significance of a Holocene Ailuropoda-Stegodon fauna from Jinhua, Zhejiang. Vertebrata PalAsiatica, 30:295-312.
Maglio, V.J. 1973. Origin and evolution of the Elephantidae. Transactions of the American Philosophical Society New Series, 63:1-149.
https://doi.org/10.2307/1006229
Makiyama, J. 1938. Japonic Proboscidea. Memoirs of the College of Science, Kyoto Imperial University, Series B, 14:1-59.
Marwick, B. 2009. Biogeography of Middle Pleistocene hominins in mainland Southeast Asia: a review of current evidence. Quaternary International, 202:51-58.
https://doi.org/10.1016/j.quaint.2008.01.012
Muhammad, R.F. 2010. The geomorphology and origin of Gua Tempurung, Perak, west Malaysia. Bulletin of the Geological Society of Malaysia, 56:127-132.
https://doi.org/10.7186/bgsm56201018
Muhammad, R.F. 2018. The extreme karstification of Kinta Valley Karst, west Malaysia. Proceedings of the 15th Multidisciplinary Conference on Sinkholes and the Engineering and Environmental Impacts of Karst and the 3rd Appalachian Karst Symposium, p. 297-305.
https://doi.org/10.5038/9780991000982.1063
Muhammad, R.F. and Yeap, E.B. 2000. Proposed conservation of Badak Cave C, Lenggong as extraordinary vertebrate fossil site. Proceeding Geological Society Malaysia Annual Geological Conference, Pulau Pinang, Malaysia, p. 189-196.
Muhammad, R.F., Yoshida, D., Tani, A, and Smart, P.L. 2002. Implications of Electron Spin Resonance and Uranium-series dating techniques on speleothems in the Kinta and Lenggong Valleys, west Malaysia. Advances in ESR Applications, 18:19-26.
Muhammad, R.F., Lim, T.T., Sahak, I.H., Azhar, N.N K., Razif, F.M., Hassan, M.H.A., Basir, M.L., Mamat, M.R., Zin, M.Z.A.N., and Zoo, S. 2019. High potential new Quaternary fossil cave sites in Merapoh (Pahang), with new geographic records for orangutan and Asian black bear. Warta Geologi, 45:400-404.
https://doi.org/10.7186/wg454201903
Muhammad, R.F., Lim T.T, Ibrahim, N., Razak, M.A.A., Razif, F.M, Kem, Z., Ching, B.T., Tee C.Y., Nang, Y.L., Chan, J.C., Chan, J.F., Rahman, M.N.A., and Hussein, S.R. 2020. First discovery of Stegodon (Proboscidea) in Malaysia. Warta Geologi, 46:196-198.
https://doi.org/10.7186/wg463202004
Nanda, A.C. 1980. A new species of Stegodon from the Upper Siwalik subgroup of Ambala, India. Geological Survey of India, Special Publication, 11:365-370.
Naumann, E. 1890. Stegodon mindanensis, eine neue Art von Uebergangs-Mastodonten. Zeitschrift der Deutschen Geologischen Gesellschaft, 42: 166-169.
Olsen, J.W. and Ciochon, R.L. 1990. A review of evidence for postulated Middle Pleistocene occupations in Viet Nam. Journal of Human Evolution, 19:761-788.
https://doi.org/10.1016/0047-2484(90)90020-c
Oram, F. 2018. Abundance, feeding and behavioural ecology of orangutans (Pongo pygmaeus morio) in the fragmented forests of the Kinabatangan floodplain. Unpublished PhD Thesis, University Malaysia Sabah (ITBC), Kota Kinabalu, Sabah, Malaysia.
O’Regan, H.J., Bishop, L., Lamb, A., Elton, S., and Turner, A. 2005. Large mammal turnover in Africa and the Levant between 1.0 and 0.5 Ma. Geological Society, London, Special Publications, 247:231-249.
https://doi.org/10.1144/gsl.sp.2005.247.01.13
Osborn, H.F. 1929. New Eurasiatic and American proboscideans. American Museum Novitates, 393:1-23.
Osborn, H.F. 1942. Proboscidea: A Monograph of the Discovery, Evolution, Migration and Extinction of the Mastodonts and Elephants of the World. Volume II: Stegodontoidea, Elephantoidea. The American Museum Press, New York.
Otsuka, H. 1984. Stratigraphic position of the Chochen Vertebrate Fauna of the T’ouk’oushan Group in the environs of the Chochen district, southwest Taiwan, with special references to its geologic age. Journal of the Taiwan Museum, 37:37-56.
Puspaningrum, M.R., van den Bergh, G.D., Chivas, A.R., Setiabudi, E., and Kurniawan, I. 2020. Isotopic reconstruction of Proboscidean habitats and diets on Java since the Early Pleistocene: implications for adaptation and extinction. Quaternary Science Reviews, 228:106007.
https://doi.org/10.1016/j.quascirev.2019.106007
Prescott, J.R. and Hutton, J.T. 1994. Cosmic ray contributions to dose rates for luminescence and ESR dating: large depths and long-term time variations. Radiation Measurements, 23:497-500.
https://doi.org/10.1016/1350-4487(94)90086-8
Rink, W.J., Wei, W., Bekken, D., and Jones, H.L. 2008. Geochronology of Ailuropoda-Stegodon fauna and Gigantopithecus in Guangxi province, southern China. Quaternary Research, 69:377-387.
https://doi.org/10.1016/j.yqres.2008.02.008
Rizal, Y. Westaway, K.E., Zaim, Y., van den Bergh G.D., Bettis E.A., Morwood, M.J., Huffman, O.F., Grün, R., Joannes-Boyau, R., Bailry, R.M., Sidarto, Westaway, M.C., Kurniawan, I., Moore, M.W., Storey, M., Aziz, F., Suminto, Zhao, J., Aswan, Sipola, M.E., Larick, R., Zonneveld, J., Scott, R., Putt, S., and Ciochon R.L. 2020. Last appearance of Homo erectus at Ngandong, Java, 117,000-108,000 years ago. Nature, 577:381-385.
https://doi.org/10.1038/s41586-019-1863-2
Roth, V.L. and Shoshani, J. 1988. Dental identification and age determination in Elephas maximus. Journal of Zoology (London), 214:567-588.
https://doi.org/10.1111/j.1469-7998.1988.tb03760.x
Sabah Wildlife Department. 2020. Orangutan Action Plan for Sabah 2020-2029. Kota Kinabalu, Sabah, Malaysia.
Saegusa, H. 1996. Stegodontidae: evolutionary relationships, p. 178-190. In Shoshani, J. and Tassy, P. (eds.), The Proboscidea: Evolution and Paleoecology of Elephants and Their Relatives. Oxford University Press, Oxford, United Kingdom.
https://doi.org/10.1093/oso/9780198546528.003.0018
Saegusa, H. 2001. Comparisons of Stegodon and Elephantid abundances in the Late Pleistocene of southern China. Proceedings of the First International Congress, Rome, p. 345-349.
Saegusa, H., Thasod, Y., and Ratanasthien, B. 2005. Notes on Asian stegodontids. Quaternary International, 126-128:31-48.
https://doi.org/10.1016/j.quaint.2004.04.013
Schepartz, L.A., Stoutamire S., and Bekken, D.A. 2005. Stegodon orientalis from Panxian Dadong, a Middle Pleistocene archaeological site in Guizhou, South China: taphonomy, population structure and evidence for human interactions. Quaternary International, 126-128:271-282.
https://doi.org/10.1016/j.quaint.2004.04.026
Sharma, C.K. and Singh, Y.L. 1966. Two vertebrate fossils from Nepal. Unpublished report, Nepal Bureau of Mines, Lainchaur, Kathmandu, Nepal.
Shen, C.C., Edwards, R.L., Cheng, H., Dorale, J.A., Thomas, R.B., Moran, S.B., Weinstein, S.E., and Edmonds, H.N. 2002. Uranium and thorium isotopic and concentration measurements by magnetic sector inductively coupled plasma mass spectrometry. Chemical Geology, 185:165-178.
https://doi.org/10.1016/s0009-2541(01)00404-1
Shen, C.C., Wu, C.C., Cheng, H., Edwards, R.L., Hsieh, Y.T., Gallet, S., Chang, C.C., Li, T.Y., Lam, D.D., Kano, A., Hori, M., and Spotl, C. 2012. High-precision and high-resolution carbonate 230Th dating by MC-ICP-MS with SEM protocols. Geochimica et Cosmochimica Acta, 99:71-86.
https://doi.org/10.1016/j.gca.2012.09.018
Shikama, T. 1936. On the genus Parastegodon. Transactions and Proceedings of the Palaeontological Society of Japan, 4:62-69.
https://doi.org/10.5575/geosoc.43.557
Shikama, T., Otsuka, H., and Tomida, Y. 1975. Fossil Proboscidea from Taiwan. Science Reports of Yokohama National University, 2:13-62.
Sondaar, P.Y. 1984. Faunal evolution and the mammalian biostratigraphy of Java. Courrier Forschung Senckenberg, 69:219-235.
Stauffer, P.H. 1973. Cenozoic, p. 143-176. In Gobbett, D.J. and Hutchison, C.S. (eds.), Geology of the Malay Peninsula (West Malaysia and Singapore). Wiley-Interscience, New York.
Storm, P., Aziz, F., de Vos, J., Kosasih, D., Baskoro, S., Ngaliman, and van den Hoek Ostende, L.W. 2005. Late Pleistocene Homo sapiens in a tropical rainforest fauna in east Java. Journal of Human Evolution, 49:536-545.
https://doi.org/10.1016/j.jhevol.2005.06.003
Suraprasit, K., Jaeger, J.J., Chaimanee, Y., Chavasseau, O., Yamee, C., Tian, P., and Panha, S., 2016. The Middle Pleistocene vertebrate fauna from Khok Sung (Nakhon Ratchasima, Thailand): biochronological and paleobiogeographical implications. ZooKeys, 613:1-157.
https://doi.org/10.3897/zookeys.613.8309
Takahashi, K. and Chang, C.H. 2001. Proboscidean fossils from the Japanese archipelago and Taiwan islands and their relationship with the Chinese mainland. The World of Elephants - International Congress, Rome, p. 148-151.
Takai, M., Saegusa, H., Htike, T., and Maung-Thein, Z. 2006. Neogene mammalian fauna in Myanmar. Asian Paleoprimatology, 4:143-172.
Tjia, H.D. 2000. A large vertebrate fossil at Naga Mas Cave, Bukit Lanno, Kinta Valley, p. 209-218. In series, Warisan Geologi Malaysia 3. Resource Development for Conservation and Nature Tourism. Lestari, Universiti Kebangsaan Malaysia, Malaysia.
Tong, H. and Liu, J. 2004. The Pleistocene-Holocene extinctions of mammals in China, p. 111-119. In Dong, W. (ed.), Proceedings of the Ninth Annual Symposium of the Chinese Society of Vertebrate Paleontology. China Ocean Press, Beijing.
Tong, H. and Patou-Mathis, M. 2003. Mammoth and other proboscideans in China during the Late Pleistocene. Deinsea, 9:421-428.
Tougard, C. 1998. Les faunes de grands mammifères du Pléistocène moyen terminal de Thaïlande dans leur cadre phylogénétique, paléoécologique et biochronologique. Unpublished PhD Dissertation, Université de Montpellier II, Montpellier, France.
Tougard, C. 2001. Biogeography and migration routes of large mammal faunas in South-East Asia during the Late Middle Pleistocene: focus on the fossil and extant faunas from Thailand. Palaeogeography, Palaeoclimatology, Palaeoecology, 168:337-358.
https://doi.org/10.1016/S0031-0182(00)00243-1
Turvey, S., Tong, H., Stuart, A., and Lister, A. 2013. Holocene survival of Late Pleistocene megafauna in China: a critical review of the evidence. Quaternary Science Reviews, 76:156-166.
https://doi.org/10.1016/j.quascirev.2013.06.030
van den Bergh, G.D. 1999. The Late Neogene elephantoid-bearing faunas of Indonesia and their palaeozoogeographic implications: a study of the terrestrial faunal succession of Sulawesi, Flores, and Java, including evidence for early hominid dispersal east of Wallace’s Line. Scripta Geologica, 117:1-419.
van den Bergh, G., de Vos, J., Aziz, F., and Morwood, M.J. 2001a. Elephantoidea in the Indonesian region: new Stegodon findings from Flores. Proceedings of the First International Congress, Rome, p. 623-627
van den Bergh, G., de Vos, J., and Sondaar, P. 2001b. The Late Quaternary palaeogeography of mammal evolution in the Indonesian Archipelago. Palaeogeography, Palaeoclimatology, Palaeoecology, 171:385-408.
https://doi.org/10.1016/S0031-0182(01)00255-3
van den Bergh, G.D., Rokhus D.A., Morwood, M.J., Sutikna, T., Jatmiko, and Saptomo, E.W. 2008. The youngest Stegodon remains in Southeast Asia from the Late Pleistocene archaeological site Liang Bua, Flores, Indonesia. Quaternary International, 182:16- 48.
https://doi.org/10.1016/j.quaint.2007.02.001
van der Geer, A.A.E., van den Bergh, G.D., Lyras, G.A., Prasetyo, U.W., Due, R.A., Setiyabudi, E., and Drinia, H. 2016. The effect of area and isolation on insular dwarf proboscideans. Journal of Biogeography, 43:1656-1666.
https://doi.org/10.1111/jbi.12743
von Koenigswald, G.H.R. 1933. Beitrag zur Kenntnis der fossilen Wirbeltiere Javas. Dienst van den Mijnbouw Nederlandsch-Indië: Wetenschappelikke MededeelingenI. Teil, 23:1-127.
von Koenigswald, G.H.R. 1935. Die fossilen Saugertier Fauna Javas. Proceeding Koninklijke Nederlandsche Akademie van Wetenschappen, 38:188-198.
Walker, D. 1956. Studies on the Quaternary of the Malay Peninsula 1: alluvial deposits of Perak and the relative levels of land and sea. Federation Museums Journal, 1 and 2:19-34.
Wang, W. and Xia, J. 1991. The measurement of annual dose from Th series and U-series decay chains by thick source alpha counting. Nuclear Techniques, 14:101-108.
Wang, W., Potts, R., Baoyin, Y., Huang, W., Cheng, H., Edwards, R.L., and Ditchfield, P. 2007. Sequence of mammalian fossils, including hominoid teeth, from the Bubing Basin caves, South China. Journal of Human Evolution, 52:370-379.
https://doi.org/10.1016/j.jhevol.2006.10.003
Wang, W., Liao, W., Li, D., and Tian, F. 2014. Early Pleistocene large-mammal fauna associated with Gigantopithecus at Mohui Cave, Bubing Basin, South China. Quaternary International, 354:122-130.
https://doi.org/10.1016/j.quaint.2014.06.036
West, R.M. and Munthe, J. 1981. Neogene vertebrate palaeontology and stratigraphy of Nepal. Nepal Geological Society, 1:1-14.
https://doi.org/10.3126/jngs.v1i1.32664
Westaway, K., Morwood, M., Roberts, R., Rokus, A.D., Zhao, J., Storm, P., Aziz, F., van den Bergh, G., Hadi, P., Jatmiko, and de Vos, J. 2007. Age and biostratigraphic significance of the Punung Rainforest Fauna, East Java, Indonesia, and implications for Pongo and Homo. Journal of Human Evolution, 53:709-717.
https://doi.org/10.1016/j.jhevol.2007.06.002
Wich, S.A., Meijaard, E., Marshall, A.J., Husson, S., Ancrenaz, M., Lacy, R.C., van Schaik, C.P., Sugardjito, J., Simorangkir, T., Traylor-Holzer, K., Doughty, M., Supriatna, J., Dennis, R., Gumal, M., Knott, C.D., and Singleton, I. 2008. Distribution and conservation status of the orang-utan (spp.) on Borneo and Sumatra: how many remain? Oryx, 42:329-339.
https://doi.org/10.1017/s003060530800197x
Zeitoun, V., Seveau, A., Forestier, H., Thomas, H., Lenoble, A., Laudet, F., Antoine, P.O., Debruyne, R., Ginsburg, L., Mein, P., Winayalai, C., Chumdee, N., Doyasa, T., Kijngam, A., and Nakbunlung, S. 2005. Découverte d’un assemblage faunique à Stegodon-Ailuropoda dans une grotte du Nord de la Thaïlande (Ban Fa Suai, Chiang Dao). Comptes Rendus Palevol, 4:255-264.
https://doi.org/10.1016/j.crpv.2004.11.013
Zeitoun, V., Lenoble, A., Laudet, F., Thompson, J., Rink, W.J., Mallye, J.B., and Chinnawut, W. 2010. The Cave of the Monk (Ban Fa Suai, Chiang Dao wildlife sanctuary, northern Thailand). Quaternary International, 220:160-173.
https://doi.org/10.1016/j.quaint.2009.11.022
Zhang, H., Wang, Y., Janis, C.M., Goodall, R.H., Purnell, and M.A. 2017. An examination of feeding ecology in Pleistocene proboscideans from southern China (Sinomastodon, Stegodon, Elephas), by means of dental microwear texture analysis. Quaternary International, 445:60-70.
https://doi.org/10.1016/j.quaint.2016.07.011
Zin-Maung-Maung-Thein, Takai, M., Uno, H., Wynn, J.G., Egi, N., Tsubamoto, T.,Thaung-Htike, Aung-Naing-Soe, Maung-Maung, Nishimura, T., and Yoneda, M. 2011. Stable isotope analysis of the tooth enamel of Chaingzauk mammalian fauna (late Neogene, Myanmar) and its implication to paleoenvironment and paleogeography. Palaeogeography, Palaeoclimatology, Palaeoecology, 300:11-22.
https://doi.org/10.1016/j.palaeo.2010.11.016
Zolkurnian, H. 1998. Urutan kebudayaan Holosen di Lembah Lenggong. Unpublished Masters Thesis, University of Science Malaysia, Pulau Pinang, Malaysia.
Zong, G. 1995. On new material of Stegodon with recollections of the classification of Stegodon in China. Vertebrata PalAsiatica, 33:216-230.
Zuraina, M. 1994. The Excavation of Gua Gunung Runtuh and the Discovery of the Perak Man in Malaysia. Department of Museums and Antiquity Malaysia, Kuala Lumpur.
