

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
 Anna Jerve. Biology Department, Imperial College London, Silwood Park Campus, Buckhurst Road, Ascot, SL5 7PY, United Kingdom. a.jerve@ic.ac.uk
Anna Jerve. Biology Department, Imperial College London, Silwood Park Campus, Buckhurst Road, Ascot, SL5 7PY, United Kingdom. a.jerve@ic.ac.uk
Anna Jerve received her PhD in 2016 from Uppsala University in Uppsala, Sweden, where she studied fin spine histology from extant and fossil taxa. She is now a research associate at Imperial College London, focusing on fossil preparation.

 Oskar Bremer. Department of Organismal Biology, Uppsala University, Norbyvägen 18A, 752 36, Uppsala, Sweden. Corresponding author. bremer.oskar@gmail.com
Oskar Bremer. Department of Organismal Biology, Uppsala University, Norbyvägen 18A, 752 36, Uppsala, Sweden. Corresponding author. bremer.oskar@gmail.com
Oskar Bremer received his PhD in 2017 from the Department of Organismal Biology, Uppsala University, Sweden. His research focuses on environmental preferences among Silurian vertebrates, and subsequently how this affects their fossil record.
 Sophie Sanchez. Science for Life Laboratory and Uppsala University, Department of Organismal Biology, Norbyvägen 18A, 752 36 Uppsala, Sweden and European Synchrotron Radiation Facility, 71 Avenue des Martyrs, 38000 Grenoble, France. sophie.sanchez@ebc.uu.se
Sophie Sanchez. Science for Life Laboratory and Uppsala University, Department of Organismal Biology, Norbyvägen 18A, 752 36 Uppsala, Sweden and European Synchrotron Radiation Facility, 71 Avenue des Martyrs, 38000 Grenoble, France. sophie.sanchez@ebc.uu.se
Sophie Sanchez is an associate senior lecturer at Uppsala University. She completed her PhD in 2008 at the Muséum national d'Histoire naturelle in Paris, where she investigated the palaeobiology of temnospondyls and seymouriamorphs by studying their bone histology. In 2009 she joined in Per Ahlberg’s lab in Uppsala and develops tools for analyzing three-dimensional early-vertebrate bone histology in collaborations with P. Tafforeau at the European Synchrotron Radiation Facility (France).
 Per E. Ahlberg. Department of Organismal Biology, Uppsala University, Norbyvägen 18A, 752 36, Uppsala, Sweden. Per.Ahlberg@ebc.uu.se
Per E. Ahlberg. Department of Organismal Biology, Uppsala University, Norbyvägen 18A, 752 36, Uppsala, Sweden. Per.Ahlberg@ebc.uu.se
Per Ahlberg is Professor of Evolutionary Organismal Biology at Uppsala University. He gained his PhD in 1989 from the University of Cambridge, and worked at University of Oxford and the Natural History Museum, London, before moving to Uppsala University in 2003. He is a member of the Royal Swedish Academy of Sciences. Per Ahlberg's research focuses on early vertebrate evolution, including the early diversification of gnathostomes and the origin of tetrapods.
FIGURE 1. Map modified from Mehlqvist et al. (2014) showing the location of the Ramsåsa area (1), and the Ramsåsa localities of Grönwall (1897) with an arrow indicating site E (2) adapted from Vergoossen (2004).
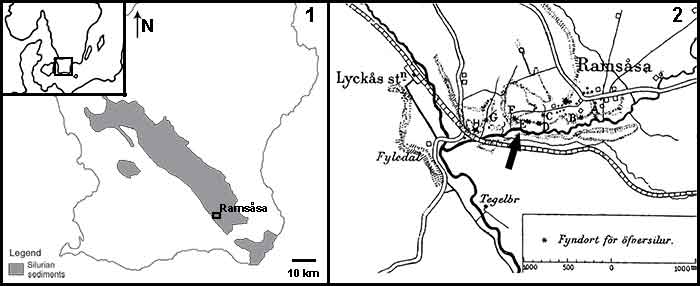
FIGURE 2. Photographs and illustrations of the seven fin spine morphotypes (A-G) from the Ramsåsa locality in Skåne, southern Sweden. (1), morphotype A (NRM-PZ P. 16173); (2), morphotype B (NRM-PZ P. 16174); (3), morphotype C (NRM-PZ P. 16175); (4), morphotype D (NRM-PZ P. 16176); (5), morphotype E (NRM-PZ P. 16177); (6), morphotype F (NRM-PZ P. 16178); and (7), morphotype G (NRM-PZ P. 16179). Each spine is shown in dorsal (top), ventral (middle), and left-lateral (bottom) views. A dotted line indicates the spine midlines. Abbreviations: ax, apex; b, base; cc, central cavity; den, denticle; i, insertion; k, keel/anterior rib; n, node; r, rib; s, sulcus. Color coding indicates the keel: red; insertion: yellow; posteriorly facing nodes/denticles: green; longitudinal sulcus along the posterior surface: orange; wide posterior surface: blue.
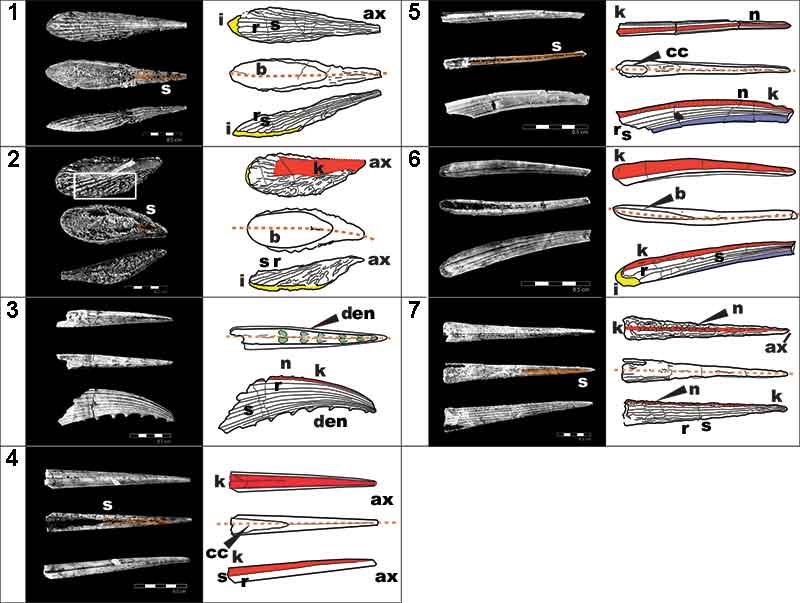
FIGURE 3. High-resolution (voxel size: 0.678 µm) synchrotron data of fin spine morphotype A. (1), virtual thin section (slice 1535) showing the general morphology and internal organization of the vascular canals and osteocyte lacunae. (2,3), segmented data illustrating the organization of vascular canals and cell spaces, and, (4,5), the reconstructed data deconstructed to show each component. (6), virtual thin sections of magnified portion of ribbing, showing the ribbing canals from slices 820, 831, and 840. Red box in (1) shows the position of the magnified portion of the fossil. Abbreviations: bvc1,2, bone vascular canals (layers 1 or 2); dvc, dentine vascular canals; o, osteocyte spaces; r, rib; rib.c, ribbing canals; s, sulcus. Scale bars equal 250 µm.
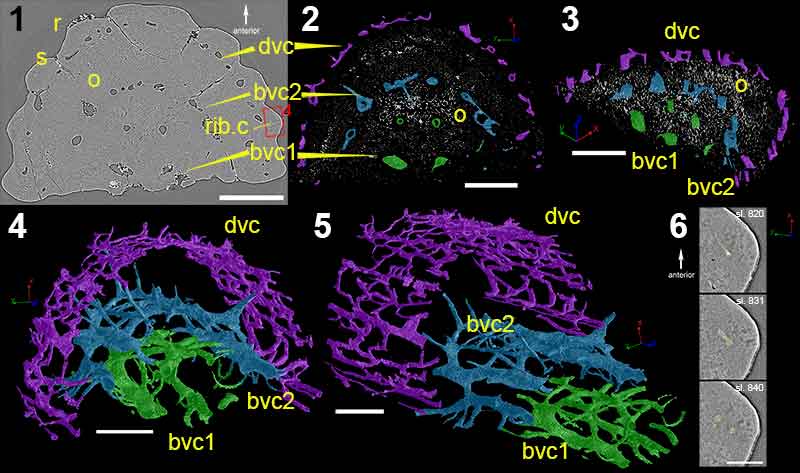
FIGURE 4. High-resolution (voxel size: 0.678 µm) synchrotron data of fin spine morphotype C, including virtual thin sections of (1), slice 1377; (2), slice 992; and (3), slice 12. (4-6) Reconstructions of the scanned fin spine showing the ornamentation. (7), Fin spine vascularization deconstructed into different parts and, (8), show all parts of the vascularization together, (9), in cross-sectional view with all canals from the scan, and (10), in cross-sectional view of isolated number of canals. (7-10) use the same colors. Scale bars equal 300 µm. Abbreviations: bas.c, basal canal; den, denticle; cc, central cavity; med.c, median canal; nod.c, node canals; r, ribbing; rib.c, ribbing canals; s, sulcus; trab.c, trabecular dentine canals.
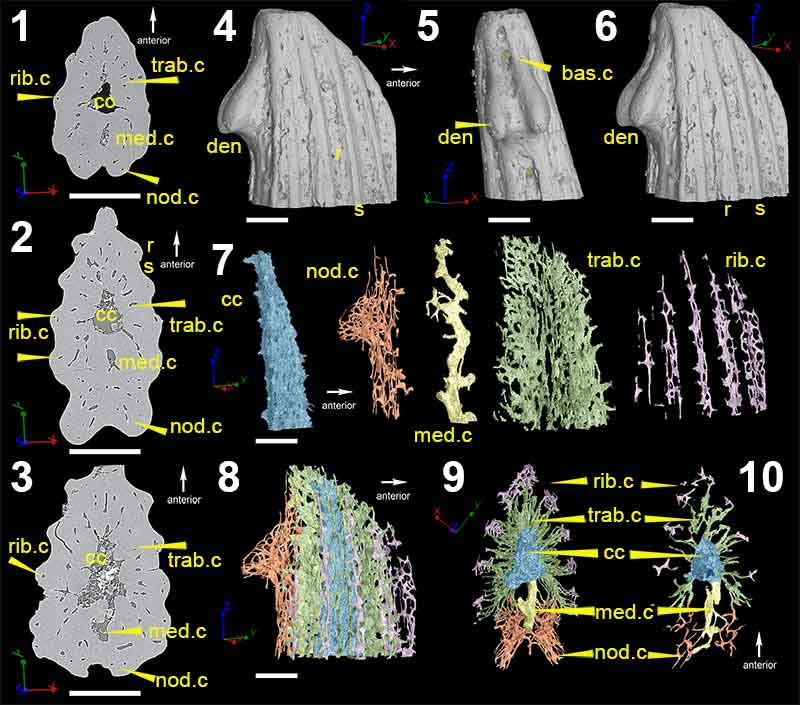
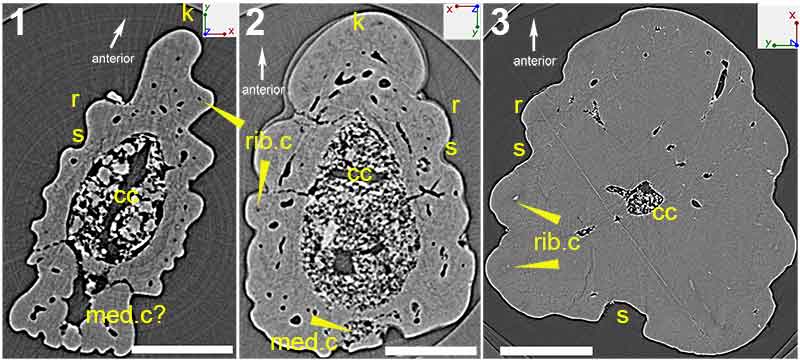
FIGURE 5. Low-resolution (voxel size: 5 µm) synchrotron data of fin spine morphotype D, including virtual thin sections of slice (1), 2596; (2), slice 2155; (3), slice 1524; (4), slice 886 and; (5), slice 222. (6), Segmented data that approximate to the scan slices 1-5, illustrating the vascularization and composition of the fin spine. (7), High-resolution (voxel size: 0.678 µm) synchrotron data 3D reconstruction illustrating the vascular organization of the fin spine. Scale bars equal 250 µm. Abbreviations: b, basal layer; cb; compact bone; cc, central canal; med.c, median canal; trab.d, trabecular dentine; vc, vascular canal.
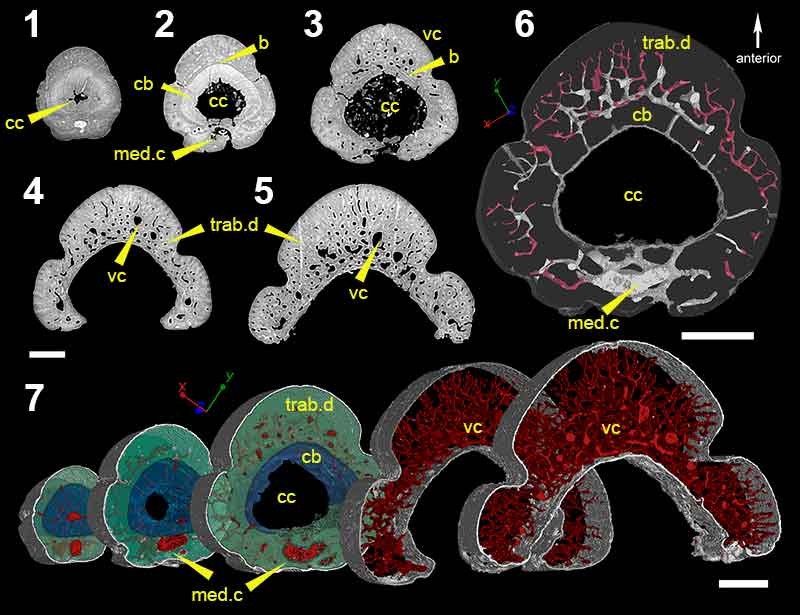
FIGURE 6. Virtual thin sections of (1), morphotype E; (2), morphotype F; and, (3), morphotype G. Scale bars equal 450 µm (1 and 2) and 200 µm (3). Abbreviations: cc, central cavity; k, keel; med.c, median canal; r, ribbing; rib.c, ribbing canals; s, sulcus.
Morphology and histology of acanthodian fin spines from the late Silurian Ramsåsa E locality, Skåne, Sweden
Plain Language Abstract
In this paper, we describe seven isolated fin spines from Silurian (ca. 420 million years old) sediments in southern Sweden. The spines come from a group of now extinct, Paleozoic fishes called acanthodians that have recently been considered as early relatives of today’s cartilaginous fishes (sharks, rays, and ratfishes) by some authors. Acanthodians generally had spines in front of all paired and midline fins (except the tail fin), as well as several more along their body, while cartilaginous fishes today only have them in front of one or both midline fins along the back. We tentatively assigned the material taxonomically to two different acanthodian groups based on earlier studies that described fish scales from the same locality. This was further facilitated by comparing the external anatomy of the spines to previously described, and more completely preserved, acanthodian fishes. This also gave an indication of their positions on the body, and most of the spines are interpreted as midline spines. Three-dimensional reconstructions based on data from high energy, synchrotron-microtomography enabled us to make detailed investigations of the histology and draw conclusions about how the spines grew. Comparisons of the tissue organization and presumed growth of the Ramsåsa spines to dermal spines in both extinct and now living fishes generally agree with the current view of acanthodians as being more closely related to sharks and their kin.
Resumen en Español
Morfología e histología de espinas de la aleta de acantodios de la localidad de Ramsåsa E, Skåne, del Silúrico tardío de Suecia
Las comparaciones de los acantodios con los gnatóstomos actuales a menudo se ven obstaculizadas por la escasez de estructuras mineralizadas en su endoesqueleto, lo que limita la potencial conservación de rasgos filogenéticamente informativos. Las espinas dorsales, estructuras dérmicas mineralizadas que se asientan delante de las aletas, se encuentran en los gnatóstomos tanto troncales como corona, y representan una potencial fuente adicional de datos comparativos para estudiar las relaciones de los acantodios con los otros grupos de gnatóstomos tempranos. Una asociación de espinas de aleta de acantodios aisladas procedente de la localidad de Ramsåsa E (sur de Suecia) ha sido reconstruida en 3D mediante el uso de microtomografía de rayos-X con radiación de sincrotrón (propagación en contraste de fase) o PPC-SRμCT. El objetivo es proporcionar afinidades morfológicas y taxonómicas para las espinas combinando la morfología y la histología con el marco taxonómico previamente establecido para el yacimiento basado principalmente en escamas aisladas. Las digitalizaciones de alta resolución también permiten investigar la composición y crecimiento de las espinas de aleta de acantodios en comparación con lo publicado en estudios similares de gnatóstomos extintos y actuales. En total, se describen siete morfotipos de espina dorsal que tienen afinidad tanto con Ischnacanthiformes Berg, 1940 como con Climatiidae Berg, 1940. La mayoría se interpreta como espinas medias de la aleta, pero también se identifican tres posibles espinas pares. Las espinas muestran diferencias en sus composiciones, pero en general coinciden con las presentadas para los Ischnacanthiformes y Climatiidae en estudios previos. Los modos de crecimiento inferidos parecen ser más similares a los de las espinas de aleta de condrictios fósiles y actuales que a los descritos para placodermos y osteíctios troncales, lo cual es congruente con la consideración emergente de que los acantodios son condrictios troncales.
Palabras clave: espinas de la aleta; histología; acantodios; reconstrucciones 3D; Silúrico; Cuenca báltica
Traducción: Enrique Peñalver (Sociedad Española de Paleontología) or Diana Elizabeth Fernández
Résumé en Français
Morphologie et histologie des épines des nageoires des acanthodiens du Silurien récent de la localité Ramsåsa E, Skåne, Suède
Les comparaisons entre les acanthodiens et les gnathostomes actuels sont souvent compliquées par la pauvreté des structures minéralisées de leur endosquelette, ce qui limite le potentiel de préservation des traits informatifs pour la phylogénie. Les épines des nageoires, des structures dermiques minéralisées situées antérieurement aux nageoires, sont présentes à la fois dans le groupe-couronne des gnathostomes et chez les gnathostomes plus basaux (en dehors du groupe-couronne), et représentent une potentielle source additionnelle de données comparatives pour étudier les relations des acanthodiens avec les autres groupes de gnathostomes anciens. Un assemblage d’épines isolées de nageoires d’acanthodiens du Silurien récent de la localité Ramsåsa site E (sud de la Suède) a été reconstruit en 3D grâce à la microtomodensitométrie par rayonnement Synchrotron utilisant le contraste de phase par propagation (PPC-SRµCT). Le but est de retrouver les affinités taxinomiques et morphologiques des épines en combinant la morphologie et l’histologie avec le cadre taxinomique précédemment établi pour le site, principalement à partir des écailles isolées. Les scans à haute résolution permettent aussi d’étudier la composition et la croissance des épines des nageoires des acanthodiens en les comparant à des études similaires sur les gnathostomes actuels et éteints. Au total, sept morphotypes d’épines des nageoires sont décrits. Ils ont des affinités à la fois avec les Climatiidae Berg 1940 et les Ischnacanthiformes Berg 1940. La majorité des épines sont interprétées comme des épines des nageoires médianes, mais trois épines possiblement paires sont aussi identifiées. Les épines montrent des différences dans leurs compositions, mais les données s’accordent généralement bien avec celles présentées pour les climatiidés et les ischnacanthiformes dans les études précédentes. Leurs modes de croissance déduits apparaissent plus similaires à ceux des épines des nageoires des chondrichthyens actuels et fossiles qu’à ceux décrits chez des placodermes et des ostéichthyens basaux (en dehors du groupe-couronne), ce qui est congruent avec la vue émergente des acanthodiens comme des chondrichthyens basaux (en dehors du groupe-couronne).
Mots-clés : épines des nageoires ; acanthodiens ; reconstructions 3D ; Silurien ; bassin de la Baltique
Translator: Antoine Souron
Deutsche Zusammenfassung
Morphologie und Histologie der Flossenstacheln acanthoder Fische aus der spätsilurischen Ramsåsa E Lokalität von Schonen, Schweden
Vergleiche von acanthoden Fischen mit heutigen Gnathostomen sind schwer, da mineralisierte Strukturen im Endoskelett sehr selten sind, was die potentielle Erhaltung phylogenetisch aussagefähiger Merkmale limitiert. Flossenstacheln, mineralisierte dermale Strukturen, die sich anterior zu den Flossen befinden, kommen sowohl bei Stammgruppen- also auch bei Kronengruppen-Gnathostomen vor. Sie können als zusätzliche Vergleichsdaten zur Untersuchung von Verwandtschaftsbeziehungen bei acanthoden Fischen mit anderen Gruppen von frühen Gnathostomen genutzt werden. Mittels Ausbreitungs-Phasenkontrast-Synchotron Microtomographie (PPC-SRµCT) wurde eine Assemblage isolierter Flossenstacheln acanthoder Fische aus dem späten Silur der Ramsåsa E Lokalität (Südschweden) rekonstruiert. Ziel ist es morphologische und taxonomische Ähnlichkeiten für die Stacheln herauszustellen, indem Morphologie und Histologie mit dem taxonomischen Rahmen kombiniert werden, der zuvor für die Lokalität anhand isolierter Schuppen aufgestellt wurde. Die hochauflösenden Aufnahmen ermöglichen auch Untersuchungen über die Zusammensetzung und das Wachstum der Flossenstacheln im Vergleich mit ähnlichen Untersuchungen über ausgestorbene und heutige Gnathostomen. Insgesamt werden sieben Flossenstacheln beschrieben, die Ähnlichkeiten mit sowohl den Climatiidae Berg 1940, als auch den Ischnacanthiformes Berg 1940, aufweisen. Die Mehrzahl wird als mediane Flossenstacheln interpretiert, es wurden jedoch auch drei möglicherweise paarige Stacheln identifiziert. Sie weisen Unterschiede in ihrer Zusammensetzung auf, sind aber im Allgemeinen mit denen der Climatiiden und Ischnacanthiformen aus früheren Untersuchungen konform. Ihre angenommene Wachstumsform scheint den Flossenstacheln der fossilen und heutigen Chondnichthyes ähnlicher zu sein, als der beschriebenen Form bei Placodermen und Stamm- Osteichthyern, was mit der aufkommenden Sicht der Acanthodia als Stamm- Chondrichthyes einhergeht.
Schlüsselwörter: Flossenstacheln; Histologie; Acanthodia; 3D-Rekonstruktion; Silur; Baltisches Becken
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024