

 Pinniped (Mammalia: Carnivora) fossils from Black Rock, a new late Neogene vertebrate locality in Victoria, Australia
Pinniped (Mammalia: Carnivora) fossils from Black Rock, a new late Neogene vertebrate locality in Victoria, Australia
Article number: 25.3.a32
https://doi.org/10.26879/1235
Copyright Paleontological Society, November 2022
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 7 August 2022. Acceptance: 12 November 2022.
ABSTRACT
The fossil record of true seals (Family Phocidae) is notoriously poorly preserved, most notably in the Southern Hemisphere. This fossil record bias has made it difficult to assess whether populations of phocid species in the Southern Hemisphere remained resident in particular regions throughout deep time, with only the eastern South Pacific preserving a near-continuous fossil record. Here we report two new seal fossil remains from the late Miocene-early Pliocene of southeastern Australia, discovered from a new site, Black Rock. This is the fourth phocid fossil site in Australia, and the seventh in Australasia. The two fossils are referrable to the clades Pinnipedia and Monachinae. Combined with other recent fossil phocid finds in Australasia, this suggests that true seals were continuously present in the region at least throughout the Pliocene. This lends further support for the importance of Australasia in the global evolution of true seals.
James P. Rule. School of Biological Sciences, Monash University, Victoria 3800, Australia and Museums Victoria Research Institute, Museums Victoria, Melbourne, Victoria 3001, Australia. jrule.palaeo@gmail.com
Erich M.G. Fitzgerald. School of Biological Sciences, Monash University, Victoria 3800, Australia and Museums Victoria Research Institute, Museums Victoria, Melbourne, Victoria 3001, Australia and National Museum of Natural History, Smithsonian Institution, Washington DC 20013-7012, U.S.A. and Department of Life Sciences, Natural History Museum, London SW7 5BD, UK. efitzgerald@museum.vic.gov.au
Keywords: pinniped; true seal; Phocidae; Monachinae; Black Rock; biogeography
Final citation: Rule, James P. and Fitzgerald, Erich M.G. 2022. Pinniped (Mammalia: Carnivora) fossils from Black Rock, a new late Neogene vertebrate locality in Victoria, Australia. Palaeontologia Electronica, 25(3):a32. https://doi.org/10.26879/1235
palaeo-electronica.org/content/2022/3723-black-rock-pinniped-fossils
Copyright: November 2022 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
Since the early Miocene, true seals (family Phocidae) have dispersed around the world, while the remaining groups of pinnipeds spent most of their evolutionary history in the North Pacific (Berta et al., 2018). True seal fossils have been found on the margins of the North Pacific, North Atlantic, Paratethys, South Pacific, and South Atlantic oceans. As a result, investigations into the biogeographic history of phocids through the expansion of this global fossil record is one of the key areas of investigation in contemporary pinniped palaeontology.
Many fossil phocids have been found in the Northern Hemisphere (Valenzuela-Toro and Pyenson, 2019), and studies of new fossils often focus on expanding the geographic range of extinct species across oceans (Koretsky and Ray, 2008; Rahmat et al., 2018). However, cosmopolitan records are usually based off fragmentary fossils (Valenzuela-Toro and Pyenson, 2019) and often are not taxonomically stable (Dewaele et al., 2018; Churchill and Uhen, 2019; Rule et al., 2020a). With such a focus on expanding the geographic range of extinct species, there has been relatively little attention on expanding the chronological range of extinct true seals; in other words, whether populations of a fossil taxon in a particular area were resident in the region throughout deep time. The best-known example of long-term occupancy is that of true seal species from Miocene sites in Peru and Chile (Muizon, 1981; Walsh and Naish, 2002; Valenzuela-Toro et al., 2016; Bosio et al. 2021), which suggest an established presence of populations of the monospecific genera Piscophoca, Acrophoca, and Australophoca in the eastern South Pacific. These sites are located in the Southern Hemisphere, a region that has historically been neglected in pinniped palaeontology (Valenzuela-Toro and Pyenson, 2019).
Recent research has expanded upon the fossil record of true seals in another region in the Southern Hemisphere; Australasia (Fitzgerald, 2005; Boessenecker and Churchill, 2016; Rule et al., 2020b; Rule et al., 2020c; Rule et al., 2020d; Rule et al., 2021a). It is becoming clear that Australasia was a critical region in the evolution of the monachines (the southern true seals), with many modern groups, such as monk seals and elephant seals, originating there (Rule et al., 2020c). However, the distribution of true seals in the region throughout deep time has received little attention, and it is currently unclear if true seal populations in Australasia were resident breeding populations or represent migrants and one-off dispersals to the region. Here we describe two phocid fossils from a new site, Black Rock, in south-eastern Australia, and discuss the possibility of long-term resident true seal populations in Australasia during the past.
MATERIALS AND METHODS
The two fossils (NMV P254178 and NMV P254995) are housed in the collections of Museums Victoria (NMV), Melbourne, Victoria, Australia. Measurements were taken to the nearest 0.01 mm using digital callipers and are reported in Table 1. To assist with illustrating preserved morphology, NMV P254995 was scanned using an Artec Space Spider surface scanner and meshed using Artec Studio 15 (Artec Group, Luxembourg). Comparative specimens (Table 2) were studied from the mammalogy collection of Museums Victoria, as well as the Natural History Museum London (NHMUK, United Kingdom), the Smithsonian Institution (USNM, Washington D.C., United States of America), and the palaeontology collection of Muséum national d'Histoire naturelle (MNHN, Paris, France).
LOCALITY AND AGE
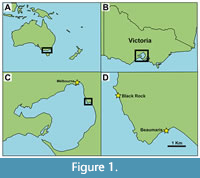 The pinniped fossils were collected ex situ on the seabed, below low-tide mark, off Black Rock Beach, Black Rock, Port Phillip Basin, Victoria, Australia; near 37°58’26” S, 145°00’48” E (Figure 1; precise locality coordinates available from Museums Victoria). Black Rock is situated on the eastern shoreline of Port Phillip Bay, about 18 km southeast of the central business district of Melbourne, and approximately 3 km northwest of the Beaumaris Bay fossil locality (Figure 1). Along the Black Rock coastline, the shallow marine upper Miocene-lower Pliocene Black Rock Sandstone (= Sandringham Sandstone: VandenBerg, 2016) is exposed as a shore platform at: the northern end of Black Rock Beach, southeast of Black Rock Point (37°58’16.99”S, 145°00’39.87”E); and further south at Quiet Corner (37°58’54.85”S, 145°01’04.52”E) (Gill, 1957; Ter and Buckeridge, 2012; VandenBerg, 2016). At the latter localities, the lithology in intertidal outcrop consists of sparsely fossiliferous, non-calcareous, ferruginous sandstone containing invertebrate burrows infilled by goethite (Gill, 1957; Abele et al., 1988; Ter and Buckeridge, 2012; VandenBerg, 2016). This lithology represents the top ~8.5 m of the Black Rock Sandstone (Abele et al., 1988). Stratigraphically underlying that top half of the Black Rock Sandstone, the lower ~6.5 m of the formation comprises fossiliferous calcareous sandstone, with a <0.3 m thick phosphatic nodule bed marking the base of the unit (Abele et al., 1988). This lower half of the Black Rock Sandstone is best exposed on- and offshore along Beaumaris Bay (see stratigraphic column in figure 1 of Rule et al., 2020b). The middle-upper Miocene Fyansford Formation (grey calcareous clay) unconformably underlies the Black Rock Sandstone in the Port Phillip Basin (Mallett and Holdgate, 1985).
The pinniped fossils were collected ex situ on the seabed, below low-tide mark, off Black Rock Beach, Black Rock, Port Phillip Basin, Victoria, Australia; near 37°58’26” S, 145°00’48” E (Figure 1; precise locality coordinates available from Museums Victoria). Black Rock is situated on the eastern shoreline of Port Phillip Bay, about 18 km southeast of the central business district of Melbourne, and approximately 3 km northwest of the Beaumaris Bay fossil locality (Figure 1). Along the Black Rock coastline, the shallow marine upper Miocene-lower Pliocene Black Rock Sandstone (= Sandringham Sandstone: VandenBerg, 2016) is exposed as a shore platform at: the northern end of Black Rock Beach, southeast of Black Rock Point (37°58’16.99”S, 145°00’39.87”E); and further south at Quiet Corner (37°58’54.85”S, 145°01’04.52”E) (Gill, 1957; Ter and Buckeridge, 2012; VandenBerg, 2016). At the latter localities, the lithology in intertidal outcrop consists of sparsely fossiliferous, non-calcareous, ferruginous sandstone containing invertebrate burrows infilled by goethite (Gill, 1957; Abele et al., 1988; Ter and Buckeridge, 2012; VandenBerg, 2016). This lithology represents the top ~8.5 m of the Black Rock Sandstone (Abele et al., 1988). Stratigraphically underlying that top half of the Black Rock Sandstone, the lower ~6.5 m of the formation comprises fossiliferous calcareous sandstone, with a <0.3 m thick phosphatic nodule bed marking the base of the unit (Abele et al., 1988). This lower half of the Black Rock Sandstone is best exposed on- and offshore along Beaumaris Bay (see stratigraphic column in figure 1 of Rule et al., 2020b). The middle-upper Miocene Fyansford Formation (grey calcareous clay) unconformably underlies the Black Rock Sandstone in the Port Phillip Basin (Mallett and Holdgate, 1985).
Sedimentary matrix adhering to the fossils described here comprises Fe-stained orange-light brown fine calcareous sandstone, containing rounded quartz grains and microfossils (including mollusc shell fragments). This associated matrix is identical to lithology within the lower 6.5 m of the Black Rock Sandstone but can be differentiated from the sediments of the basal nodule bed, which are distinctly yellow, phosphatic, and rich in quartz pebbles (Dickinson and Wallace, 2009; Rule et al., 2020b). It is, therefore, deduced that: (1) the pinniped fossils originated from a horizon within the lower 6.5 m of the Black Rock Sandstone, but stratigraphically above the basal phosphatic nodule bed; and (2) that the stratigraphic section at Black Rock Beach includes at least part of the lower 6.5 m of the Black Rock Sandstone, exposed as offshore submarine outcrop.
The Black Rock Sandstone was deposited between the late Miocene (Messinian) and early Pliocene (Zanclean) (Fitzgerald and Kool, 2015; Rule et al., 2020b). Measured 87Sr/86 Sr isotope ratios of francolite in phosphates from the basal nodule bed gave estimated ages of 6.24-5.38 Ma, suggesting a maximum age for the Black Rock Sandstone of latest Miocene (Dickinson and Wallace, 2009). 87Sr/86 Sr ratios from molluscan/brachiopod carbonate at horizons approximately 1.0 m and 2.6 m above the basal nodule bed, produced ages of 5.98-5.59 Ma (late Miocene) and 4.91 Ma (early Pliocene), respectively (Dickinson and Wallace, 2009). These data indicate that the Miocene-Pliocene boundary lies within the lowest 2.6 m of the Black Rock Sandstone. However, due to the scarcity, if not absence, of age-indicative fossils in the upper ~12 m of the Black Rock Sandstone, its minimum age is unresolved. Within the Port Phillip Basin, basalts of the Newer Volcanics Province overlie the Neogene marine succession (including the Black Rock Sandstone) (Abele et al., 1988; Wallace et al., 2005). The minimum radiometric age for the commencement of this volcanism is 4.62 Ma (Hare et al., 2005). VandenBerg (2016) inferred that the onset of this volcanism coincided with the end of Neogene marine deposition in the Port Phillip Basin (including the Black Rock Sandstone) but cautioned that the stratigraphically lowest basalt flows have not been dated. Nevertheless, we concur with VandenBerg (2016) in using these data to conservatively estimate deposition of the Black Rock Sandstone between 6.24 and 4.62 Ma; with the Black Rock section of the Black Rock Sandstone (and pinniped fossils described herein) deposited between ~5.98 and 4.62 Ma, late Miocene to early Pliocene.
Until now, the vertebrate palaeontology of the Black Rock Sandstone has been almost exclusively represented by the rich Beaumaris Local Fauna of Beaumaris Bay, comprised of: chondrichthyan and actinopterygian fish (Kemp, 1991; Fitzgerald, 2004); marine and freshwater turtles (Fitzgerald and Kool, 2015; Rule et al., 2021b); penguins (Park and Fitzgerald, 2012a), pelagornithid birds (Fitzgerald et al., 2012), an albatross (Wilkinson, 1969), a dromornithid (Park and Fitzgerald, 2012b); marsupials (Piper et al., 2006); a sirenian (Fitzgerald et al., 2013); phocid seals (Rule et al., 2020b, 2021a); and cetaceans (Fordyce, 1982; Fitzgerald, 2004, 2012; Marx et al., 2018). The only prior documentation of a vertebrate fossil from Black Rock is a zygomaturine diprotodontid left maxilla (erroneously reported as a dentary from Beaumaris) figured by Vickers-Rich and Rich (1993:177) and Warne et al. (2003:638). In addition to the pinniped material described here, recently collected vertebrate fossils from the Black Rock locality include isolated specimens of Diprotodontidae, Cetacea, Spheniscidae, Pelagornithidae, and chondrichthyans, suggesting general equivalence between the Black Rock assemblage and the Beaumaris Local Fauna.
SYSTEMATIC PALAEONTOLOGY
Order CARNIVORA Bowdich, 1821
Suborder PINNIPEDIA Illiger, 1811
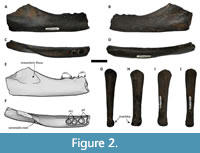 Referred specimen. NMV P254178, an isolated phalanx collected by B.S.J. Francischelli in 2019 (Figure 2).
Referred specimen. NMV P254178, an isolated phalanx collected by B.S.J. Francischelli in 2019 (Figure 2).
Diagnosis. An intermediate or proximal phalanx from the manus (from either digits II, III, or IV) of a pinniped, due to the straight medial and lateral surfaces, a distal end narrower than a proximal end, and a slightly concave ventral surface.
Description. The proximal end is rounded, and is missing the articular surface (Figure 2G-J). The distal surface is trochleated (Figure 2G-H), and larger medio-laterally than dorso-ventrally. The shaft is sub-circular in cross section. Due to the lack of diagnostic features beyond Pinnipedia, we are unable to make meaningful comparisons to other species.
Family PHOCIDAE Gray, 1821
Subfamily MONACHINAE Gray, 1869
Referred specimen. NMV P254995, a fragmentary right mandible collected by B.S.J. Francischelli in 2019 (Figure 2).
Diagnosis. A fragmentary right mandible based on the position of the masseteric fossa. Referred to Pinnipedia based on a transversely flattened mandibular body, and a coronoid crest that does not ascend vertically from the toothrow. Referred to Phocidae based on a double rooted m1 with clearly separated anterior and posterior alveoli. Referred to Monachinae based on a shallow coronoid crest that ascends below a 45° angle.
Description. NMV P254995 is missing the proximal portion of the mandible, including the articular surface and a portion of the coronoid crest, and the distal portion from the p4 onwards. The body of the mandible is thick (12 mm) compared to the low height (23 mm), making the medial-lateral thickness roughly half of the dorsal-ventral length. The medial surface of the body is flat compared to the lateral surface, which is slightly dorso-ventrally curved (Figure 2D). While the preserved body of the mandible is antero-posteriorly straight, the coronoid crest curves medially. The masseteric fossa is shallow (Figure 2E), and barely visible on the lateral surface of the coronoid. The coronoid crest ascends shallowly from the toothrow (<45°), with a short space separating the crest from the posterior margin of the toothrow. Both alveoli of the m1 and the preserved posterior alveolus of the p4 are round, with the p4 alveolus slightly larger than the m1 (Figure 2F). A short diastema separates the m1 and p4. The posterior portion of the anterior alveolus of the p4 is preserved (Figure 2F).
Comparison. The shallow coronoid crest of NMV P254995 is similar to that condition seen in Neomonachus, Mirounga, Acrophoca, and Lobodontini, and is therefore mostly consistent with Monachinae (Phocinae have a steep coronoid crest). The only exceptions are Monachus, Homiphoca, Piscophoca, and Hadrokirus. The toothrow is separated from the coronoid crest by a short space (less than twice the length of the m1), similar to the monachines Lobodon, Hydrurga, Hadrokirus, and tribe Monachini; and similar to phocines such as Erignathus, Halichoerus, Phoca, and Pusa.
The body of the mandible NMV P254995 below the toothrow is thick, similar to Monachini and Hadrokirus. All other phocids, including other monachines, have either thin mandibles below the toothrow (width less than half the height), or mandibles that grade from thin posteriorly to thick anteriorly. While most phocids have a mandible that is fairly straight antero-posteriorly, NMV P254995 has a slightly curved mandible similar to Ommatophoca, Neomonachus, and Erignathus. Overall, the preserved morphology of NMV P254995 has the most similarities with the genus Neomonachus.
Two other fragmentary fossil pinniped mandibles are known from Australia: NMV P251642 (Beaumaris: Rule et al., 2020b) and NMV P218465 (Portland: Fitzgerald, 2005). The shallow coronoid crest and round profile of the posterior m1 alveolus of NMV P254995 is shared with NMV P251642 and NMV P218465. Further, several additional characters are shared between NMV P254995 and NMV P218465: a posterior p4 alveolus larger than both m1 alveoli (also in common with other phocids), and an m1 alveoli at a slightly imbricated angle compared to the p4 alveoli. There are no discernible differences between NMV P254995, NMV P251642, and NMV P218465. The similarities between NMV P254995 and NMV P218465 are enough to confidently revise the identification of NMV P218465 to Phocidae cf. Monachinae, confirming the tentative diagnosis of Phocidae of Fitzgerald (2005).
DISCUSSION
True seals are currently thought to have originated in either the North Atlantic or Paratethys, where they are known from multiple localities covering most of the Neogene (Berta et al., 2018). However, in the North Pacific and the Global South, true seals are usually only known from very few sites. The best record in the Southern Hemisphere is the Neogene of the eastern South Pacific, with multiple fossil sites from Peru and Chile, preserving multiple species from several stratigraphic units (Muizon, 1981; Walsh and Naish, 2002; Amson and Muizon, 2014; Valenzuela-Toro et al., 2016; Benites-Palomino et al., 2022). This is potentially sufficient evidence that phocids were resident to the eastern South Pacific during the late Miocene to early Pliocene. Outside of this region, the record of phocids is sparse, (Valenzuela-Toro and Pyenson, 2019). For example, Argentina has two species from two formations (Muizon and Bond, 1982; Cozzuol, 2001) dated to the middle-late Miocene, and South Africa has many fossils of one species represented from a few early Pliocene sites (Hendey and Repenning, 1972; Muizon and Hendey, 1980; Govender, 2015). As a result, not many biogeographic regions (other than the eastern South Pacific) have phocid fossils present across a wide span of time and space.
Despite the apparent constraints on the global fossil record of true seals (Valenzuela-Toro and Pyenson, 2019), Australasia is emerging as a significant region in true seal evolutionary history due to an increase of reporting of fossil finds (Boessenecker and Churchill, 2016; Rule et al., 2020b; Rule et al., 2020c; Rule et al., 2020d; Rule et al., 2021a). These new finds span from the late Miocene to the early Pleistocene, from both south-eastern Australia and the north island of New Zealand. Together with the new fossils reported here from Black Rock, these records suggest that phocids may have had resident breeding populations in Australasia throughout the Pliocene.
U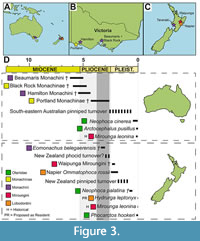 ntil recently, it was unknown which ancient true seals were present in Australasia during the Neogene, with the most specific identification being Monachinae (Fordyce and Flannery, 1983). Since then, recent studies have revealed that monk seals (tribe Monachini) existed in the Neogene of Australasia (Figure 3). Specifically, monk seal temporals are known from Beaumaris and Hamilton in Australia (Rule et al., 2021a), and skulls are known from Taranaki in New Zealand (Rule et al., 2020c). While none of the Black Rock specimens are referrable to Monachini, it is notable that NMV P254995 is most similar to the genus Neomonachus. The presence of monk seals in Australasia, from at least three sites spanning a time period of over 3 million years, is unexpected and unprecedented, as monk seals were traditionally considered a northern clade. This implies that Austral latitudes presented ideal environmental conditions for monk seal fauna to inhabit during at least the Pliocene.
ntil recently, it was unknown which ancient true seals were present in Australasia during the Neogene, with the most specific identification being Monachinae (Fordyce and Flannery, 1983). Since then, recent studies have revealed that monk seals (tribe Monachini) existed in the Neogene of Australasia (Figure 3). Specifically, monk seal temporals are known from Beaumaris and Hamilton in Australia (Rule et al., 2021a), and skulls are known from Taranaki in New Zealand (Rule et al., 2020c). While none of the Black Rock specimens are referrable to Monachini, it is notable that NMV P254995 is most similar to the genus Neomonachus. The presence of monk seals in Australasia, from at least three sites spanning a time period of over 3 million years, is unexpected and unprecedented, as monk seals were traditionally considered a northern clade. This implies that Austral latitudes presented ideal environmental conditions for monk seal fauna to inhabit during at least the Pliocene.
These records seem to indicate that monk seals were widespread around Australasia during the Pliocene, only to go extinct during the marine megafauna extinction (3.8-2.4 Ma; Boessenecker, 2013; Pimiento et al., 2017). This extinction coincided with the late Pliocene pinniped turnover events (Avery and Klein, 2011; Valenzuela-Toro et al., 2013; Rule et al., 2020d; Benites-Palomino et al., 2022), which saw the disappearance of most archaic phocids in the Southern Hemisphere. After this extinction, fossils representing lobodontins (Antarctic true seals) and miroungins (elephant seals) are known from the early Pleistocene of New Zealand (King, 1973; Boessenecker and Churchill, 2016). It is remarkable that the three tribes of Monachinae (monk, elephant, and Antarctic true seals) were present so close in time to each other in Australasia (Figure 3). Eomonachus (the New Zealand monk seal) is separated from the Napier Ross seal and Waipunga elephant seal by only 500,000-600,000 years (Figure 3; King, 1973; Boessenecker and Churchill, 2016). While this represents a very narrow separation in geological time, the two occurrences are separated from each other by the marine megafauna extinction (Pimiento et al., 2017). In addition, monk, elephant, and Antarctic seals have distinct ecologies and life histories, which influence their geographic distribution today. As such, there is no reason to postulate that the three tribes once co-occurred in the same fauna. Furthermore, elephant and Antarctic seals co-occur in the Southern Ocean today, so the early Pleistocene records of Lobodontini and Miroungini in New Zealand may simply represent the earliest onset of the Southern Ocean fauna (Figure 3). There is also the possibility that this may indicate another cryptic pinniped turnover event in New Zealand during the early Pleistocene (Figure 3), although additional fossils of these phocids should be discovered to ensure they were resident in the region (either for breeding or hauling-out to rest).
By the Late Pleistocene (Figure 3), true seals were no longer the dominant pinnipeds in Australasia. By then, fur seals and sea lions had dispersed south from the North Pacific Ocean (Yonezawa et al., 2009; Churchill et al., 2014) and become resident in Australia (McCoy, 1877) and New Zealand (Churchill and Boessenecker, 2016). The only true seals present in the region were Southern elephant seals (Mirounga leonina) in Bass Strait (Bryden et al., 1999), which remained until they became locally extinct from overhunting. This was reflective of the post-pinniped turnover fauna present elsewhere in the Southern Hemisphere, including South Africa (Avery and Klein, 2011), South America (Valenzuela-Toro et al., 2015), and sub-Antarctic islands (Berta et al., 2018). This modern pinniped fauna relied on cooler, more productive waters near the southern continents. Without a doubt, a transition from a more archaic pinniped fauna to the modern pinniped fauna occurred sometime between the Late Pliocene and Late Pleistocene.
True seals are now known from four fossil sites in Australia and three fossil sites in New Zealand. These sites encompass the entirety of the Pliocene and the beginning of the Pleistocene, indicating that exploration of other sites representing this time period in Australasia will yield more phocid fossils. We recommend further exploration of late Miocene and early-middle Pleistocene aged marine fossil sites in Australasia, as further finds of fossil phocids would expand the chronological range of the group in this region. By extension, Pleistocene marine fossil sites in Australasia could further expand the fossil record of otariids in the region. Therefore, any pre-Holocene pinniped fossils will be critical for refining the timing of the pinniped turnover in Australasia.
CONCLUSIONS
The fossil record of phocids from Australasia has greatly expanded in recent years. As a result, it seems likely that true seals were resident in the region throughout the Pliocene period. This confirms Australasia as an important region for true seal evolution in the past, with the potential to inform about the diversification and ultimate extinction of ancient true seals in the Southern Hemisphere.
ACKNOWLEDGEMENTS
We acknowledge the Bunurong People of the Kulin Nation as the Traditional Owners and Custodians of the land and waterways on which these fossils were found. J. Rule is funded by an Australian Research Council Discovery Project (DP180101797). B.S.J. Francischelli discovered the two fossils described in the manuscript. Overseas comparative material was studied thanks to the support of a Monash University Graduate Research Travel Grant awarded to J. Rule during his PhD studies. Some of the discussion and figures in this manuscript are modified from J. Rule's PhD thesis, for which he was funded by the Australian Government Research Training Program and a Robert Blackwood Partnership Award. The following museum staff are thanked for providing collections access: T. Ziegler, K. Roberts, K. Date, R-L. Erickson, and K. Rowe (NMV); T. Park, R. Miguez, and P. Jenkins (NHML); C. de Muizon and G. Billet (MNHN); J. Mead, J. Ososky, M. McGowen, D. Lunde, D. Bohaska, and N. Pyenson (USNM). This research on the Lost World of Bayside is supported by Bayside City Council, Beaumaris Motor Yacht Squadron, Sandringham Community Bank Branch of Bendigo Bank, Sandringham Foreshore Association, Bayside Earth Sciences Society and generous community donations. Thanks to the editors, Leonard Dewaele, Ana Valenzuela Toro, and one anonymous reviewer for comments that improved this manuscript.
REFERENCES
Abele, C., Gloe, C.S., Hocking, J.B., Holdgate, G., Kenley, P.R., Lawrence, C.R., Ripper, D., Threlfall, W.F., and Bolger, P.F. 1988. Tertiary, p. 251-350. In Douglas, J.G., and Ferguson, J.A. (eds.), Geology of Victoria. Victorian Division, Geological Society of Australia, Fitzroy.
Amson, E. and Muizon, C. de. 2014. A new durophagous phocid (Mammalia: Carnivora) from the late Neogene of Peru and considerations on monachine seals phylogeny. Journal of Systematic Palaeontology, 12:523-548. https://doi.org/10.1080/14772019.2013.799610
Avery, G. and Klein, R.G. 2011. Review of fossil phocid and otariid seals from the southern and western coasts of South Africa. Transactions of the Royal Society of South Africa, 66:14-24. https://doi.org/10.1080/0035919x.2011.564490
Benites-Palomino, A., Valenzuela-Toro, A.M., Figueroa-Bravo, C., Varas-Malca, R.M., Nielsen, S.N., Gutstein, C.S., and Carrillo-Biceño, J.D. 2022. A new marine mammal assemblage from central Chile reveals the Pliocene survival of true seals in South America. Historical Biology, 1-13. https://doi.org/10.1080/08912963.2021.2007528
Berta, A., Churchill, M., and Boessenecker, R.W. 2018. The Origin and Evolutionary Biology of Pinnipeds: Seals, Sea Lions, and Walruses. Annual Review of Earth and Planetary Sciences, 46; 203-228. https://doi.org/10.1146/annurev-earth-082517-010009
Boessenecker, R. and Churchill., M. 2016. The origin of elephant seals: implications of a fragmentary late Pliocene seal (Phocidae: Miroungini) from New Zealand. New Zealand Journal of Geology and Geophysics, 59:544-550. https://doi.org/10.1080/00288306.2016.1199437
Boessenecker, R.W. 2013. A new marine vertebrate assemblage from the Late Neogene Purisima Formation in Central California, part II: Pinnipeds and Cetaceans. Geodiversitas, 35:815-940. https://doi.org/10.5252/g2013n4a5
Bosio, G., Collareta, A., Celma, C.D., Lambert, O., Marx, F.G., Muizion, C. de., Gioncada, A., Gariboldi, K., Malinverno, E., Malca, R.V., Urbina, M., and Bianucci, G. 2021. Taphonomy of marine vertebrates of the Pisco Formation (Miocene, Peru): Insights into the origin of an outstanding Fossil-Lagerstätte. PLoS ONE, 16: e0254395. https://doi.org/10.1371/journal.pone.0254395
Bowdich, T. 1821. An analysis of the natural classifications of Mammalia for the use of students and travellers. J. Smith, Paris.
Bryden, M., O'Connor, S., and Jones, R. 1999. Archaeological evidence for the extinction of a breeding population of elephant seals in Tasmania in prehistoric times. International Journal of Osteoarchaeology, 9:430-437. https://doi.org/10.1002/(sici)1099-1212(199911/12)9:6<430::aid-oa512>3.0.co;2-p
Churchill, M., Boessenecker, R.W., and Clementz, M.T. 2014. Colonization of the Southern Hemisphere by fur seals and sea lions (Carnivora: Otariidae) revealed by combined evidence phylogenetic and Bayesian biogeographical analysis. Zoological Journal of the Linnean Society, 172:200-225. https://doi.org/10.1111/zoj12163
Churchill, M. and Boessenecker, R.W. 2016. Taxonomy and biogeography of the Pleistocene New Zealand sea lion Neophoca palatina (Carnivora: Otariidae). Journal of Paleontology, 90:375-388. https://doi.org/10.1017/jpa.2016.15
Churchill, M. and Uhen, M.D. 2019. Taxonomic implications of morphometric analysis of earless seal limb bones. Acta Palaeontologica Polonica, 64: 213-230. https://doi.org/10.4202/app.00607.2019
Cozzuol, M.A. 2001. A “northern” seal from the Miocene of Argentina: implications for phocid phylogeny and biogeography. Journal of Vertebrate Paleontology, 21:415-421. https://doi.org/10.1671/0272-4634(2001)021[0415:ansftm]2.0.co;2
Dewaele, L., Peredo, C.M., Meyvisch, P., and Louwye, S. 2018. Diversity of late Neogene Monachinae (Carnivora, Phocidae) from the North Atlantic, with the description of two new species. Royal Society open science, 5:172437. https://doi.org/10.1098/rsos.172437
Dickinson, J. A. and Wallace, M.W. 2009. Phosphate-rich deposits associated with the Mio-Pliocene unconformity in south-east Australia. Sedimentology, 56:547-565. https://doi.org/10.1111/j.1365-3091.2008.00985.x
Fitzgerald, E.M.G. 2004. A review of the Tertiary fossil Cetacea (Mammalia) localities in Australia. Memoirs of Museum Victoria, 61:183-208. https://doi.org/10.24199/j.mmv.2004.61.12
Fitzgerald, E.M.G. 2005. Pliocene marine mammals from the Whalers Bluff Formation of Portland, Victoria, Australia. Memoirs of Museum Victoria, 62:67-89. https://doi.org/10.24199/j.mmv.2005.62.2
Fitzgerald, E.M.G. 2012. Possible neobalaenid from the Miocene of Australia implies a long evolutionary history for the pygmy right whale Caperea marginata (Cetacea, Mysticeti). Journal of Vertebrate Paleontology, 32:976-980. https://doi.org/10.1080/02724634.2012.669803
Fitzgerald, E.M.G., Park, T., and Worthy, T.H. 2012. First giant bony-toothed bird (Pelagornithidae) from Australia. Journal of Vertebrate Paleontology, 32:971-974. https://doi.org/10.1080/02724634.2012.664596
Fitzgerald, E.M.G., Velez-Juarbe, J., and Wells, R.T. 2013. Miocene sea cow (Sirenia) from Papua New Guinea sheds light on sirenian evolution in the Indo-Pacific. Journal of Vertebrate Paleontology, 33:956-963. https://doi.org/10.1080/02724634.2013.753081
Fitzgerald, E.M.G. and Kool, L. 2015. The first fossil sea turtles (Testudines: Cheloniidae) from the Cenozoic of Australia. Alcheringa, 39:142-148. https://doi.org/10.1080/03115518.2015.964047
Fordyce, R.E. 1982. A review of Australian fossil Cetacea. Memoirs of the National Museum of Victoria, 43:43-58. https://doi.org/10.24199/j.mmv.1982.43.04
Fordyce, R. E. and Flannery, T. 1983. Fossil phocid seals from the late Tertiary of Victoria. Proceedings of the Royal Society of Victoria, 95:99-100.
Gill, E.D. 1957. The stratigraphical occurrence and palaeoecology of some Australian Tertiary marsupials. Memoirs of the National Museum of Victoria, 21:135-203. https://doi.org/10.24199/j.mmv.1957.21.10
Govender, R. 2015. Preliminary phylogenetics and biogeographic history of the Pliocene seal, Homiphoca capensis from Langebaanweg, South Africa. Transactions of the Royal Society of South Africa, 70:25-39. https://doi.org/10.1080/0035919x.2014.984258
Gray, J.E. 1821. On the natural arrangement of vertebrose animals. London Medical Repository, 15:296-310.
Gray, J.E. 1869. XLI.--Notes on seals (Phocidæ) and the changes in the form of their lower jaw during growth. Journal of Natural History 4:342-346. https://doi.org/10.1080/00222936908696068
Hare, A.G., Cas, R.A.F., Musgrave, R., and Phillips, D. 2005. Magnetic and chemical stratigraphy for the Werribee Plains basaltic lava flow-field, Newer Volcanics Province, southeast Australia: implications for eruption frequency. Australian Journal of Earth Sciences, 52:41-57. https://doi.org/10.1080/08120090500100069
Hendey, Q.B. and Repenning, C.A. 1972. A pliocene phocid from South Africa. Annals of the South African Museum, 59:79-98.
Illiger, J.K.W. 1811. Prodomus systematis mammalium et avium additis terminis zoographicii utriusque classis eorumque versione germanica. C. Salfeld, Berlin. https://doi.org/10.5962/bhl.title.106965
Kemp, N.R. 1991. Chondrichthyans in the Cretaceous and Tertiary of Australia, p. 497-568. In Vickers-Rich, P., Monaghan, J.M., Baird, R.F., and Rich, T.H. (eds.), Vertebrate Palaeontology of Australasia. Pioneer Design Studio in cooperation with the Monash University Publications Committee, Melbourne.
King, J.E. 1973. Pleistocene Ross seal (Ommatophoca rossi) from New Zealand (Note). New Zealand Journal of Marine and Freshwater Research, 7:391-397. https://doi.org/10.1080/00288330.1973.9515483
Koretsky, I.A. and Ray, C.E. 2008. Phocidae of the Pliocene of eastern USA. Virginia Museum of Natural History Special Publication, 14:81-140.
Mallett, C.W. and Holdgate, G.R. 1985. Subsurface Neogene stratigraphy of Nepean Peninsula, Victoria. Special Publication, South Australian Department of Mines and Energy, 5:233-245.
Marx, F.G., Park, T., Fitzgerald, E.M.G., and Evans, A.R. 2018. A Miocene pygmy right whale fossil from Australia. PeerJ 6:e5025. https://doi.org/10.7717/peerj.5025
McCoy, F. 1877. Prodromus of the Palaeontology of Victoria, Decade 5. Geological Survey of Victoria, 5:283-284. https://doi.org/10.1017/s0016756800150368
Muizon, C. de. 1981. Les vertébrés fossiles de la formation Pisco (Pérou). Première partie: deux nouveaux Monachinae (Phocidae, Mammalia) du Pliocène de Sud-Sacaco. Travaux de l’Institut français d’Études andines, 22:1-161.
Muizon, C. de. and Hendey, Q.B. 1980. Late tertiary seals of the South Atlantic Ocean. Annals of the South African Museum, 82:91-128.
Muizon, C. de. and Bond, M. 1982. Le Phocidae (Mammalia) Miocene de la formation Parana (Entre Rios, Argentine). Bulletin du Muséum National d’Histoire Naturelle, section C, 4eme série, 4:165-207.
Park, T. and Fitzgerald, E.M.G. 2012a. A review of Australian fossil penguins (Aves: Sphenisciformes). Memoirs of Museum Victoria, 69:309-325. https://doi.org/10.24199/j.mmv.2012.69.06
Park, T. and Fitzgerald, E.M.G. 2012b. A late Miocene-early Pliocene Mihirung bird (Aves: Dromornithidae) from Victoria, southeast Australia. Alcheringa, 36:419-422. https://doi.org/10.1080/03115518.2012.663572
Pimiento, C., Griffin, J.N., Clements, C.F., Silvestro, D., Varela, S., Uhen, M.D., and Jaramillo, C. 2017. The Pliocene marine megafauna extinction and its impact on functional diversity. Nature ecology & evolution 1:1100-1106. https://doi.org/10.1038/s41559-017-0223-6
Piper, K.J., Fitzgerald, E.M.G., and Rich, T.H. 2006. Mesozoic to early Quaternary mammal faunas of Victoria, south-east Australia. Palaeontology, 49:1237-1262. https://doi.org/10.1111/j.1475-4983.2006.00595.x
Rahmat, S., Muñiz, F., Toscano, A., Esperante, R., and Koretsky, I. 2018. First European record of Homiphoca (Phocidae: Monachinae: Lobodontini) and its bearing on the paleobiogeography of the genus. Historical Biology, 32:561-569. https://doi.org/10.1080/08912963.2018.1507030
Rule, J.P., Adams, J.W., Rovinsky, D.S., Hocking, D.P., Evans, A.R., and Fitzgerald, E.M.G. 2020a. A new large-bodied Pliocene seal with unusual cutting teeth. Royal Society Open Science 7:201591. https://doi.org/10.1098/rsos.201591
Rule, J.P., Adams, J.W., and Fitzgerald, E.M.G. 2020b. Colonization of the ancient southern oceans by small-sized Phocidae: new evidence from Australia. Zoological Journal of the Linnean Society. 191:1160-1180. https://doi.org/10.1093/zoolinnean/zlaa075
Rule, J.P., Adams, J.W., Marx, F.G., Evans, A.R., Tennyson, A.J., Scofield, R.P., and Fitzgerald, E.M.G. 2020c. First monk seal from the Southern Hemisphere rewrites the evolutionary history of true seals. Proceedings of the Royal Society B 287:20202318. https://doi.org/10.1098/rspb.2020.2318
Rule, J.P., Hocking, D.P., and Fitzgerald, E.M.G. 2020d. Pliocene monachine seal (Pinnipedia: Phocidae) from Australia constrains timing of pinniped turnover in the Southern Hemisphere. Journal of Vertebrate Paleontology 39: e1734015. https://doi.org/10.1080/02724634.2019.1734015
Rule, J.P., Adams, J.W., and Fitzgerald, E.M.G. 2021a. Early monk seals (Monachinae: Monachini) from the late Miocene-early Pliocene of Australia. Journal of Systematic Palaeontology, 19:441-459. https://doi.org/10.1080/14772019.2021.1920639
Rule, J.P., Kool, L., Parker, W.M.G., and Fitzgerald, E.M.G, 2021b. Turtles all the way down: Neogene pig-nosed turtle fossil from southern Australia reveals cryptic freshwater turtle invasions and extinctions. Papers in Palaeontology 8:e1414. https://doi.org/10.1002/spp2.1414
Ter, P.C. and Buckeridge, J. St. J.S. 2012. Ophiomorpha beaumarisensis isp. nov., a trave fossil from the late Neogene Beaumaris Sandstone is the burrow of a thalassinidean lobster. Proceedings of the Royal Society of Victoria, 124:223-231.
Valenzuela-Toro, A.M., Gutstein, C.S., Varas-Malca, R.M., Suarez, M.E., and Pyenson, N.D. 2013. Pinniped turnover in the South Pacific Ocean: new evidence from the Plio-Pleistocene of the Atacama Desert, Chile. Journal of Vertebrate Paleontology, 33:216-223. https://doi.org/10.1080/02724634.2012.710282
Valenzuela-Toro, A.M., Gutstein, C.S., Suárez, M.E., Otero, R., and Pyenson, N.D. 2015. Elephant seal (Mirounga sp.) from the Pleistocene of the Antofagasta Region, northern Chile. Journal of Vertebrate Paleontology, 35:e918883. https://doi.org/10.1080/02724634.2014.918883
Valenzuela-Toro, A.M., Pyenson, N.D., Gutstein, C.S., and Suarez, M.E. 2016. A new dwarf seal from the Late Neogene of South America and the evolution of pinnipeds in the Southern Hemisphere. Papers in Palaeontolgy, 2:101-115. https://doi.org/10.1002/spp2.1033
Valenzuela-Toro, A. and Pyenson, N.D. 2019. What do we know about the fossil record of pinnipeds? A historiographical investigation. Royal Society open science, 6:191394. https://doi.org/10.1098/rsos.191394
VandenBerg, A.H.M. 2016. Depositional facies and extent of the late Neogene Sandringham Sandstone in southern Victoria, Australia. Proceedings of the Royal Society of Victoria, 128:7-24.
Vickers-Rich, P. and Rich, T.H. 1993. Wildlife of Gondwana. Reed, Chatswood, 276 p.
Wallace, M.W., Dickinson, J.A., Moore, D.H., and Sandiford, M. 2005. Late Neogene strandlines of southern Victoria: a unique record of eustasy and tectonics in southeast Australia. Australian Journal of Earth Sciences, 52:277-295. https://doi.org/10.1080/08120090500139455
Walsh, S. and Naish, D. 2002. Fossil seals from late Neogene deposits in South America: a new pinniped (Carnivora, Mammalia) assemblage from Chile. Palaeontology, 45:821-842. https://doi.org/10.1111/1475-4983.00262
Warne, M.T., Archbold, N.W., Bock, P.E., Darragh, T.A., Dettmann, M.E., Douglas, J.G., Gratsianova, R.T., Grover, M., Holloway, D.J., Holmes, F.C., Irwin, R.P., Jell, P.A., Long, J.A., Mawson, R., Partridge, A.D., Pickett, J.W., Rich, T.H., Richardson, J.R., Simpson, A.J., Talent, J.A., and VandenBerg, A.H.M. 2003. Palaeontology: the biogeohistory of Victoria, p. 605-652. In Birch, W.D. (ed.), Geology of Victoria. Geological Society of Australia Special Publication 23.
Wilkinson, H.E. 1969. Description of an Upper Miocene albatross from Beaumaris, Victoria, Australia, and a review of fossil Diomedeidae. Memoirs of the National Museum of Victoria, 29:41-51. https://doi.org/10.24199/j.mmv.1969.29.04
Yonezawa, T., Kohno, N., and Hasegawa, M. 2009. The monophyletic origin of sea lions and fur seals (Carnivora; Otariidae) in the Southern Hemisphere. Gene, 441:89-99. https://doi.org/10.1016/j.gene.2009.01.022
