

 A taxonomic revision of the genus Tanystropheus (Archosauromorpha, Tanystropheidae)
A taxonomic revision of the genus Tanystropheus (Archosauromorpha, Tanystropheidae)
Article number: 22.3.80
https://doi.org/10.26879/1038
Copyright Society of Vertebrate Paleontology, December 2019
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 15 October 2019. Acceptance: 3 December 2019.
ABSTRACT
Tanystropheus represents one of the most characteristic genera of Triassic reptiles and is typified by easily recognizable, hyperelongate cervical vertebrae. First described in 1852, isolated cervical vertebrae and other remains have been referred to the genus and various species have been erected and rejected based on this material. This has resulted in a complicated and convoluted taxonomic history of the genus and confusion as to the validity of species and the referral of specimens. With the exception of the well-represented T. longobardicus, the five other species of Tanystropheus are known from isolated elements or a single, partial specimen. Here, we provide a complete overview of the taxonomic history and a revision of the genus based on first hand observations of the type material of most of the species. From this, we conclude that T. conspicuus and T. haasi should be considered nomina dubia and that T. meridensis constitutes a junior synonym to T. longobardicus. Furthermore, T. longobardicus can be subdivided into two discrete morphotypes that might represent separate species. However, a more detailed study is required to test this hypothesis. Finally, T. fossai is considered distinctly different from the other Tanystropheus taxa and is therefore referred to a separate genus, Sclerostropheus.
Stephan N.F. Spiekman. Palaeontological Institute and Museum, University of Zurich, Karl Schmid-Strasse 4, 8006, Zürich, Switzerland. stephan.spiekman@pim.uzh.ch
Torsten M. Scheyer. Palaeontological Institute and Museum, University of Zurich, Karl Schmid-Strasse 4, 8006, Zürich, Switzerland. scheyer@pim.uzh
Keywords: Tanystropheus; Archosauromorpha; new genus; Triassic; taxonomy; biogeography
Spiekman, Stephan N.F. and Scheyer, Torsten M. 2019. A taxonomic revision of the genus Tanystropheus (Archosauromorpha, Tanystropheidae). Palaeontologia Electronica 22.3.80 1–46. https://doi.org/10.26879/1038
palaeo-electronica.org/content/2019/2870-revision-of-tanystropheus
Copyright: December 2019 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0/creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/C7552279-E6B3-4363-911E-BE9D44566A3D
INTRODUCTION
Tanystropheus represents one of the most enigmatic tetrapod taxa of the Triassic due to its unique morphology and palaeobiology. Its most striking aspect is its extremely long neck, which consists of a relatively small number (13, see Rieppel et al., 2010) of bizarrely elongated cervical vertebrae with reduced neural spines. This type of cervical vertebrae is unique among tetrapods and easily recognizable.
For almost a century the genus Tanystropheus von Meyer, 1852, was only known from a number of these elongate vertebrae from the Upper and Lower Muschelkalk of Central Europe (Meyer, 1855; Huene, 1907-1908). Their position along the vertebral column was unclear, and they were tentatively interpreted as caudal vertebrae. They were only recognized as cervical vertebrae following the discovery of largely complete and articulated skeletons from the Besano Formation of Monte San Giorgio on the border between Switzerland and Italy (former Grenzbitumenzone, Anisian-Ladinian boundary, Middle Triassic; Stockar, 2010) (Peyer, 1931).
Based on these more complete specimens, it was originally proposed that the genus Tanystropheus was closely related to Sauropterygia (Peyer, 1931), but subsequent authors placed it in either “Prolacertiformes” or “Protorosauria”; two clades roughly encompassing the same taxa (e.g., Prolacerta broomi, Protorosaurus speneri, and Macrocnemus spp.) characterized by the presence of elongated cervical vertebrae (e.g., Camp, 1945a; Camp, 1945b; Wild, 1980a; Benton, 1985; Chatterjee, 1986; Evans, 1988; Benton and Allen, 1997; Jalil, 1997; Dilkes, 1998). Although originally considered to belong to Lepidosauromorpha (Wild, 1973), “Prolacertiformes/Protorosauria” were grouped within Archosauromorpha by Gow (1975), a placement which was later supported by Benton (1985) and Evans (1988) and is now firmly established. However, recent analyses strongly indicate “Prolacertiformes/Protorosauria” to be polyphyletic within Archosauromorpha, generally distinguishing a monophyletic Tanystropheidae from other “protorosaurs” like Prolacerta broomi and Protorosaurus speneri (Dilkes, 1998; Rieppel et al., 2003; Pritchard et al., 2015; Ezcurra, 2016; Spiekman, 2018).
The genus Tanystropheus is currently represented by six species. Out of these species, only T. longobardicus from the Besano Formation of Monte San Giorgio is represented by various articulated and largely complete specimens, including skull material (Wild, 1973; Nosotti, 2007). Recently, a nearly complete skeleton from the latest Ladinian or earliest Carnian of southwestern China could not be distinguished from the T. longobardicus specimens from the European Besano Formation, and was identified as T. cf. longobardicus, implying the possibility of a Tethys-wide distribution for this species (Rieppel et al., 2010). Other species are comprised of either a number of isolated remains (T. conspicuus, T. antiquus, and T. haasi) or a single incomplete specimen (T. meridensis, T. fossai) (Meyer, 1855; Huene, 1905, 1907-1908; Wild, 1973, 1980a; Rieppel, 2001; Fraser and Rieppel, 2006; Sennikov, 2011; Skawiński et al., 2017). Additionally, isolated remains from Europe, the Middle East, Asia, and North America have been attributed to the genus but have not been determined to the species level (Vickers-Rich et al., 1999; Dalla Vecchia, 2000; Rieppel, 2001; Dalla Vecchia and Avanzini, 2002; Dalla Vecchia, 2005; Li, 2007; Sues and Olsen, 2015).
Here we provide a review of the long and convoluted taxonomic history of the genus Tanystropheus and revise its taxonomy based on firsthand observations, including observations of the type material of T. conspicuus, T. longobardicus, T. meridensis, and T. fossai, and a cast of the holotype of T. haasi.
Taxonomic History of Tanystropheus
The genus Tanystropheus was erected on the basis of eight isolated, elongate bones from the Upper Muschelkalk of Bindlach near Bayreuth, Germany, and an incomplete specimen from the lowermost Keuper of Upper Silesia (Laryszów, Poland), which were identified as reptilian vertebrae and assigned to Tanystropheus conspicuus by Meyer (1855). He pointed out that these bones had previously been described by Count Georg zu Münster, who had interpreted them as limb bones of a saurian reptile, which he had named “Macroscelosaurus”. However, since this work has been lost and this genus name has fallen into disuse (nomen oblitum), the genus name Tanystropheus has received precedence (Wild, 1973, p. 148; Melville, 1981).
A number of isolated remains from the Chinle Formation (Late Triassic) of northwestern New Mexico, USA, were assigned to three new species: Tanystrophaeus bauri, Tanystrophaeus willistoni, and Tanystrophaeus longicollis (Cope, 1887). However, additional findings showed distinct differences with T. conspicuus from the Upper Muschelkalk, and this material was soon after re-assigned to a new genus of dinosaur, Coelophysis (Cope, 1889).
Later, additional elongate vertebrae from various localities were described (reported on in Huene (1905) and described in more detail in Huene (1907-1908); Supplementary Table 1). A number of these specimens from the Upper Muschelkalk were assigned to Tanystropheus conspicuus, whilst two additional species of Tanystropheus were erected. A number of elongate vertebrae from the Gogolin beds (Lower Muschelkalk) near Gogolin and Krappitz in Silesia, Poland, were identified as being similar to T. conspicuus and assigned to T. antiquus and a vertebra from the Norian of Stuttgart-Heslach, Germany, was assigned to T. posthumus. However, subsequent findings of Tanystropheus later indicated that this latter species did not belong to this genus (Peyer, 1931). Currently, this specimen is considered an otherwise indeterminable caudal vertebra of a theropod dinosaur, and the taxon “Tanystropheus posthumus” is considered a nomen dubium (Rauhut and Hungerbühler, 2000). The T. antiquus material from the Gogolin beds was recently preliminarily revised, and these specimens are considered to be of early Anisian and possibly latest Olenekian age (Skawiński et al., 2017).
More than 75 years after the initial description of the isolated remains of Tanystropheus conspicuus, excavations at the Besano Formation at Monte San Giorgio revealed largely complete and articulated specimens that could be referred to Tanystropheus (Peyer, 1930, 1931). Previously, the identity of the elongate vertebrae of Tanystropheus had been unclear, and they had been suggested to be caudal vertebrae (Meyer, 1855; Huene, 1907-1908). The articulated nature of the specimens described by Peyer (1931) clarified that they undeniably represented hyper-elongated cervical vertebrae. Furthermore, cranial remains within this material showed distinct tricuspid marginal dentition. The presence of these tricuspid teeth, as well as the elongate cervical vertebrae, was identical to that present in a now lost specimen from the same locality that had previously been described and interpreted as a pterosaur, Tribelesodon longobardicus, in which the cervical vertebrae had been misidentified as elongated phalanges due to a lack of comparative material (Bassani, 1886; Nopsca, 1923). Peyer (1931) assigned this specimen and the newly discovered material to Tanystropheus longobardicus.
Following the description of the articulated Tanystropheus longobardicus specimens, isolated remains from other localities, mainly part of the Upper and Lower Muschelkalk of Europe, were re-evaluated and elements other than elongate cervicals were identified to likely belong to the genus Tanystropheus (Huene, 1931). Among this material was a vertebra from the Erfurt Formation (Lettenkeuper, middle Ladinian) of Gaildorf, Germany, that was originally described by Plieninger (1846) and, together with other fragmented remains, including a partial upper jaw including teeth, was assigned to “Zanclodon laevis”, thus predating T. conspicuus by Meyer (1855). However all the material assigned to “Z. laevis” apart from the upper jaw fragment has been lost, and this jaw fragment has thus been assigned as the lectotype of the taxon, which differs distinctly from that of Tanystropheus spp. (Wild, 1973). “Z. laevis” was also briefly discussed, and its taxonomic history summarized by Schoch (2011), who also figured the type specimen and identified it as an archosauriform. Additionally, among the material discussed, specimens previously assigned to “Thecodontosaurus latespinatus”, “Thecodontosaurus primus”, and “Procerosaurus cruralis” were also considered to very likely belong to the genus Tanystropheus (Huene, 1931; see also the synonymy lists for T. conspicuus and T. antiquus by Wild, 1973, p. 148-149, 151). However, the assignment of “Thecodontosaurus primus” to the genus was recently questioned, and a detailed revision of this material is required to establish its affinities (Skawiński et al., 2017).
Tanystropheid remains from the Upper Buntsandstein (Röt Formation, early Anisian; Menning and Hendrich, 2016) of the Black Forest, Germany, which slightly predates the Lower Muschelkalk, were described and assigned to Tanystropheus longobardicus and Macrocnemus bassanii (Ortlam, 1966). This material was initially re-assigned to T. antiquus (Wild, 1980a). However, Wild subsequently considered this material to differ from Tanystropheus spp. to such a degree that it belonged to a different genus, but did not formally re-assign this material (Wild, 1987). Later, the Buntsandstein specimens were assigned to a new genus and species, Amotosaurus rotfeldensis, whilst T. antiquus was tentatively maintained as a valid taxon representing material from the Lower Muschelkalk (Fraser and Rieppel, 2006).
In the initial description of Tanystropheus longobardicus no detailed comparison with T. conspicuus and T. antiquus from the Germanic Basin was provided in expectation of further preparation and additional finds from excavations at Monte San Giorgio (Peyer, 1931). This comparison was eventually provided in an extensive monograph on T. longobardicus following the availability of more specimens (Wild, 1973). Therein, T. conspicuus was distinguished from T. longobardicus based on comparatively wider rib attachment sites and a concavity on the anterior end of the neural spine of the cervical vertebrae. Although these minor differences were considered not to be sufficient to define a species, the distinction between the two taxa was maintained in expectation of additional specimens from the Upper Muschelkalk that would allow for a more complete comparison. Although never providing a formal revision, Wild later considered T. conspicuus to very likely be indistinguishable from T. longobardicus (Wild, 1980a, 1980b, 1987). Tanystropheus antiquus showed more disparity from T. longobardicus in having distinctly shorter cervical vertebrae with more pronounced neural spines and zygapophyses (Wild, 1973), as was also pointed out previously in comparison to T. conspicuus (Huene, 1907-1908). Additionally, the monograph provided a systematic palaeontology section, including a synonymy list and overview of the occurrence for each of the three species in detail (Wild, 1973). From this, it followed that T. conspicuus occurred in the Upper Muschelkalk (late Anisian-early Ladinian; Menning and Hendrich, 2016) of Germany (area surrounding Bayreuth, Bindlach, Crailsheim, Schloss Stetten, Erfurt, and Göttingen) and France (Lunéville; although Peyer, 1931, did not consider this specimen, a vertebra, to belong to Tanystropheus), and in the Lettenkeuper (middle Ladinian; Menning and Hendrich, 2016) of Germany (Gaildorf, Crailsheim, and Helmstedt). The single specimen from the Upper Muschelkalk of Laryszów assigned to T. conspicuus by Meyer (1855) was not included in this list. The material assigned to T. conspicuus comprises isolated cervical, dorsal, sacral, and caudal vertebrae, as well as an isolated humerus and femora (Wild, 1973). Since the original description by Meyer (1855) provided a syntype of nine cervicals, one of these, U-MO BT 740 (Meyer, 1855, plate 30, figure 2), was assigned as the lectotype of the species. The material assigned to T. antiquus comprised a number of isolated cervical and dorsal vertebrae and a femur, which originated from the Lower Muschelkalk of Krappitz and Gogolin in Silesia, Poland, and near Jena and Rüderdorf near Berlin, Germany (Wild, 1973). Later an isolated cervical vertebra from the Lower Muschelkalk of Winterswijk (Vossenveld Formation), the Netherlands, was also assigned to the species (Wild and Oosterink, 1984) and recently more tanystropheid material from this locality was described and discussed (Spiekman et al., 2019). The assignment of specimens from Schattenmühle near Bonndorf in the Black Forest and Diedesheim near Mosbach (both Germany) to T. antiquus was considered uncertain (Wild, 1973). SMNS 16687 (no. 7 in Huene 1907-1908, p. 225, plate 93, figure 1) was established as the lectotype, since Wild (1973), as well as Fraser and Rieppel (2006), thought that the majority of the other specimens belonging to the syntype were very likely destroyed during the Second World War. However, it was recently reported that these specimens still exist (Skawiński et al., 2017). The specimens assigned to T. longobardicus were restricted to the Besano Formation of Monte San Giorgio and the Buntsandstein material assigned to the species by Ortlam (1966; Wild, 1973), which as mentioned before was soon after reassigned to T. antiquus and later to Amotosaurus rotfeldensis (Fraser and Rieppel, 2006). Because the holotype, a specimen originally assigned to Tribelesodon longobardicus (Bassani, 1886), was unfortunately destroyed in the Second World War (specimen figured in Arthaber, 1922, figure 3a), PIMUZ T 2791, the main specimen (“Hauptfund”) of the description by Peyer (1931), was established as the neotype of T. longobardicus (Wild, 1973).
Additionally, the presence of another species of Tanystropheus from Makhtesh Ramon (Anisian-Ladinian) of Israel was also briefly noted on, based on material which was first reported on by Peyer (1955). However, this new species was not erected therein, since the material was projected to be worked on by Georg Haas and identified preliminarily as Tanystropheus sp. (Wild, 1973). This material was eventually described and assigned to the species Tanystropheus haasi (Rieppel, 2001). It comprises posterior ends of two mid-cervical vertebrae, as well as a number of highly fragmentary specimens assigned to the same species based on their similar size. In the diagnosis, T. haasi was distinguished from other Tanystropheus species based on the presence of a distinct horizontal groove that separated the vertebral centrum from the neural arch, the presence of thickened margins of the articulation facets of the postzygapophyses, the presence of a straight posterior margin of the postzygapophyseal trough, which is located at the level of the articulation facet of the centrum, and the presence of a long posterior process of the neural spine that overlies the postzygapophyseal trough. Additional specimens from the same locality were referred to two different morphotypes, with one being distinctly larger and the other being distinctly smaller than T. haasi. The larger morphotype is represented by the posterior end of a posterior cervical vertebra (cervical vertebra 11 or 12) and various highly fragmented specimens, referred to Tanystropheus sp. The best-preserved specimen was estimated to be approximately 40% larger than the known 11th cervical of T. conspicuus (based on Wild, 1973). It was described as being different from T. haasi and any other species of Tanystropheus in possessing a distinct recess in the posterior margin of the postzygapophyseal trough and a horizontally oriented postzygapophysis in which the articulation facet faces ventrally rather than ventrolaterally. It additionally differed from T. haasi in possessing an oval posterior opening of the postzygapophyseal canal, which is circular in T. haasi. The smaller morphotype was considered to be very similar to T. conspicuus and is represented by a number of fragmentary cervical vertebrae, with two specimens that consist of the anterior end of the vertebral centrum being the most diagnostic. The material assigned to this morphotype does not show overlapping diagnostic morphology with the T. haasi material nor with that of the larger morphotype from Makhtesh Ramon. Furthermore, a handful of vertebrae remains referred to the genus Tanystropheus have also been described from the Jilh Formation of Saudi Arabia (dated to the Middle Triassic), which has been considered closely related to that of Makhtesh Ramon (Vickers-Rich et al., 1999). However, this Tanystropheus material was not identified to the species level.
A new species, Tanystropheus biharicus, was erected based on a single isolated cervical from the Anisian of Romania (Jurcsák, 1975), and later additional fragmentary remains were attributed to this species (Jurcsák, 1976, 1978, 1982). However, Wild (1980a) found the holotype to be indistinguishable from both T. longobardicus and T. conspicuus, and it was reassigned to T. cf. longobardicus.
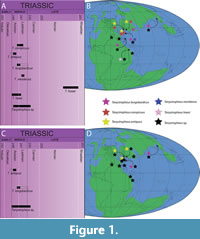 In the same study, additional previously undescribed Tanystropheus specimens were presented, and two more species were erected, Tanystropheus meridensis and Tanystropheus fossai (Wild, 1980a). Tanystropheus meridensis is known from a single specimen, PIMUZ T 3901, that originates from the Cassina beds of the Meride Limestone at Monte San Giorgio (early middle Ladinian), which are slightly younger than the Besano Formation from which the T. longobardicus specimens originate (Stockar, 2010; Figure 1). The species consists of a small specimen preserving a complete yet heavily crushed skull and the first seven cervical vertebrae. It was described as differing from T. longobardicus in the absence of palatal teeth, the quadrate being comparatively taller and more curved, the presence of paired parietals, and having slightly more elongate cervical vertebrae. Additionally, a single, incomplete tricuspid tooth from the lower Lettenkeuper near Hall in Baden-Württemberg was identified as T. cf. meridensis (Wild, 1980a). Later an additional specimen, MCSN 5562, was described from the Cascina or Cava Inferiore beds of the Meride Limestone of Monte San Giorgio, which are very slightly older than the Cassina beds in which PIMUZ T 3901 was found, yet younger than the Besano Formation (Stockar, 2010). Because the skull and anterior part of the cervical column is missing in this specimen, a direct comparison with PIMUZ T 3901 is impossible. However, this specimen showed no distinct differences from the T. longobardicus specimens and was identified either as T. cf. longobardicus (Renesto, 2005) or T. longobardicus (Nosotti, 2007). Although T. meridensis has not formally been revised, the species has been considered synonymous and indistinguishable from the small specimens of T. longobardicus (Fraser et al., 2004; Nosotti, 2007). However, it was considered possible that two taxa are represented among the Tanystropheus specimens of Monte San Giorgio, with the small specimens forming a distinct separate morphotype from the large sized specimens, meaning that these differences are attributable to specific variation rather than ontogenetic variation (Fraser et al., 2004).
In the same study, additional previously undescribed Tanystropheus specimens were presented, and two more species were erected, Tanystropheus meridensis and Tanystropheus fossai (Wild, 1980a). Tanystropheus meridensis is known from a single specimen, PIMUZ T 3901, that originates from the Cassina beds of the Meride Limestone at Monte San Giorgio (early middle Ladinian), which are slightly younger than the Besano Formation from which the T. longobardicus specimens originate (Stockar, 2010; Figure 1). The species consists of a small specimen preserving a complete yet heavily crushed skull and the first seven cervical vertebrae. It was described as differing from T. longobardicus in the absence of palatal teeth, the quadrate being comparatively taller and more curved, the presence of paired parietals, and having slightly more elongate cervical vertebrae. Additionally, a single, incomplete tricuspid tooth from the lower Lettenkeuper near Hall in Baden-Württemberg was identified as T. cf. meridensis (Wild, 1980a). Later an additional specimen, MCSN 5562, was described from the Cascina or Cava Inferiore beds of the Meride Limestone of Monte San Giorgio, which are very slightly older than the Cassina beds in which PIMUZ T 3901 was found, yet younger than the Besano Formation (Stockar, 2010). Because the skull and anterior part of the cervical column is missing in this specimen, a direct comparison with PIMUZ T 3901 is impossible. However, this specimen showed no distinct differences from the T. longobardicus specimens and was identified either as T. cf. longobardicus (Renesto, 2005) or T. longobardicus (Nosotti, 2007). Although T. meridensis has not formally been revised, the species has been considered synonymous and indistinguishable from the small specimens of T. longobardicus (Fraser et al., 2004; Nosotti, 2007). However, it was considered possible that two taxa are represented among the Tanystropheus specimens of Monte San Giorgio, with the small specimens forming a distinct separate morphotype from the large sized specimens, meaning that these differences are attributable to specific variation rather than ontogenetic variation (Fraser et al., 2004).
Tanystropheus fossai has also been described on the basis of a single specimen (MCSNB 4035), which constitutes four largely articulated cervical vertebrae and associated cervical ribs (Wild, 1980a). This specimen originates from the late Norian of Lombardy, Italy, and thus represents the latest occurrence of the genus (Wild, 1980a; Renesto and Dalla Vecchia, 2018; Figure 1). This specimen was distinguished from other species of Tanystropheus based on distinct laterally projecting crests on the lateral surface of the cervical centra and the presence of bifurcated cervical ribs. The assignment of T. fossai to Tanystropheus was corroborated (Dalla Vecchia, 2000), although similarities between these vertebrae and the caudal vertebrae of certain Triassic pterosaurs was also pointed out. However, the presence of thin and hyperelongate ribs occurring parallel to the vertebral column precludes their identification as caudal vertebrae. Another study found the vertebrae of T. fossai to lack any unequivocally diagnostic characters for the assignment to the genus and stated that they are markedly shorter proportionally than those of other Tanystropheus species (Renesto, 2005).
Two further specimens were described, a partial right coracoid from the Upper Muschelkalk of Siles in Andulusia, Spain, and a poorly preserved, fragmentary dorsal vertebra from the late Ladinian of Seiser Alm in South Tyrol, Italy. They were identified as Tanystropheus sp. indet. and ?Tanystropheus sp. indet., respectively, expanding the geographic distribution of the genus known at that time (Wild, 1980a).
A complete isolated femur from the S-charl Formation (late Anisian to early Ladinian) of Piz Ravigliel near Davos, canton Graubünden, Switzerland was briefly described and assigned to Tanystropheus sp. by Eichenberger (1986).
A number of isolated Tanystropheus remains from northern Italy have been described and assigned to Tanystropheus sp. and Tanystropheus cf. longobardicus (Dalla Vecchia, 2000; Dalla Vecchia and Avanzini, 2002; Dalla Vecchia, 2005). An isolated proximal caudal (MFSN 25761) and the proximal half of a thoracic rib, missing the articular head, were found in close association in a single block. The vertebra shares many similarities and has been assigned to the genus Tanystropheus, but, although it was considered different from the caudal vertebrae of T. conspicuus and T. longobardicus, it was not assigned on the species level because the material was considered too incomplete (Dalla Vecchia, 2000). Although the rib is undiagnostic, it is indistinguishable from the ribs of Tanystropheus, and because of its close association with the caudal vertebra, it has been suggested that it could belong to the same individual. The block contains four other unidentifiable bones. It was recovered as an isolated block and thus cannot be assigned to a specific locality, and it could have an earliest Triassic to Norian age (Dalla Vecchia, 2000). An additional specimen from layer E of the Fusea site in Friuli, Italy, which is most likely of early Carnian age, constitutes the posterior half of a cervical vertebra. Although it was described as being most similar to the middle to posterior cervical vertebrae of T. longobardicus, it was not formally assigned to the species and identified as Tanystropheus sp. (Dalla Vecchia, 2000). An additional specimen, the posterior part of a small cervical vertebra, was described from the Ladinian of the Mendel Pass in South Tyrol, Italy (Dalla Vecchia and Avanzini, 2002). It was not identified on the species level, and attributed to Tanystropheus sp. Another 30 Tanystropheus specimens were described from the late Anisian deposits of the Aupa Valley, Friuli, Italy (Dalla Vecchia, 2005). Among these remains are virtually complete cervical, dorsal, sacral, and caudal vertebrae, as well as several isolated teeth, a complete left clavicle, and a right ilium. This material might differ from other Tanystropheus species on the basis of the absence of the neural spine on the posterior end of the neural arch in the cervical vertebrae and the wide transverse process of the dorsal vertebrae, but was otherwise considered very similar to T. longobardicus, and the material was therefore referred to T. cf. T. longobardicus (Dalla Vecchia, 2005).
Since the 2000s, many new discoveries from the Middle to Late Triassic of southwestern China (Guizhou and Yunnan Provinces) have revealed a rich fauna of marine vertebrates, including several tanystropheid taxa; e.g., Dinocephalosaurus orientalis, Macrocnemus fuyuanensis, Tanystropheus sp., and T. cf. T. longobardicus (Li et al., 2004; Li, 2007; Li et al., 2007; Rieppel et al, 2008; Rieppel et al., 2010; Jiang et al., 2011), as well as potentially Fuyuansaurus acutirostris and Pectodens zhenyuensis (Fraser et al., 2013; Li et al., 2017). These findings greatly enlarged the known geographic range of the clade to the eastern part of the Triassic Tethys Ocean, including that of Tanystropheus spp., which were previously only known from its western margin (Li et al., 2007; Rieppel et al., 2010; Jaquier et al., 2017). Two Tanystropheus specimens from China are currently described, both comprising postcranial skeletons lacking the skulls and originating from the Zhuganpo Member of the Falang Formation (latest Ladinian to earliest Carnian) near Xingyi, Guizhou Province, China. The smaller specimen, IVPP V 14472, was interpreted as a juvenile specimen and comprises the posterior part of the cervical column, the trunk, the pectoral and pelvic girdles, the forelimbs, and a femur. It was considered to be very similar to T. longobardicus and identified as Tanystropheus sp. (Li, 2007). The larger specimen, GMPKU-P-1527, possesses a largely complete postcranial skeleton, including most of the cervical series, a complete trunk series and most of the tail, parts of both sides of the pectoral girdle, a complete right forelimb, parts of both pelvic girdles, and most of the right hind limb with the exception of the foot. It was found to be very similar to the large specimens of T. longobardicus known from the Besano Formation, with the only possible minor differences being the somewhat larger size of the chevron bones and the lack of slight swellings on the cervical ribs (Rieppel et al., 2010), which were observed in PIMUZ T 2819 (PIMUZ T 2189 therein). Although no distinct differences with T. longobardicus were found, the specimen was referred to T. cf. T. longobardicus because the authors considered the absence of a skull to preclude a sufficiently detailed comparison to the European large-sized T. longobardicus specimens. Nevertheless, these findings indicate a very close association between the faunas from both ends of the Tethys Ocean, as has also been established for other marine reptile clades; e.g., Sauropterygia (Li, 2006; Jiang et al., 2008; Wang et al., 2019) and Ichthyopterygia (Jiang et al., 2006). Furthermore, even for the genus Macrocnemus, a taxon generally considered to be terrestrial, a very close association has been established for specimens occurring at both sides of the Tethys (Jaquier et al., 2017).
Tanystropheid remains from the Donskaya Luka locality (Lipovskaya Formation, late Olenekian) of the Ilovlinsky District, southwestern Russia, were described and considered most closely related to Tanystropheus antiquus and Amotosaurus rotfeldensis (Sennikov, 2011). These remains were assigned to a new genus and species, Augustaburiania vatagini. This taxon is comprised of isolated specimens, and the material attributed to it consists of a number of isolated cervical and anterior dorsal vertebrae, partial sacral vertebra, caudal vertebrae, humeral fragments, femora, and a proximal fragment of a tibia. It might represent the earliest occurrence of Tanystropheidae, although recently described isolated archosauromorph remains from the Sanga do Cabral Formation of Brazil (Induan-early Olenekian) predate Au. vatagini and likely also belong to the clade (De Oliveira et al., 2018). Additionally, Sennikov (2011) re-evaluated T. antiquus and it was considered to differ sufficiently from the other Tanystropheus species to merit assignment to a separate genus, and it was re-assigned to Protanystropheus antiquus. However, most of the type material of this species was not included in this diagnosis, nor in that of Wild (1973) or Fraser and Rieppel (2006), as it was still considered lost at the time. Although a full revision of the taxon was refrained from, initial observation of the original material by Huene (1905, 1907-1908) led to the consideration that T. antiquus is a valid taxon, giving it preference over P. antiquus (Skawiński et al., 2017). However, this preliminary interpretation only covered the specimens from the Gogolin Formation, and the taxonomic status of the T. antiquus specimens from Germany and the Netherlands was considered to be uncertain until a detailed revision of original material from the Gogolin Formation is completed. Additionally, the existence of new material assigned to T. conspicuus, which originated from the lowermost Keuper (Ladinian) of Laryszów in Upper Silesia, Poland, was also briefly noted on. This is the same locality from which a specimen included in the original description of T. conspicuus by Meyer (1855) also derived (Skawiński et al., 2017).
A virtually complete and strongly elongated cervical vertebra was described from the Economy Member of the Wolfville Formation of Carrs Brook in Nova Scotia, south-eastern Canada, which is of Anisian to Carnian age (Sues and Olsen, 2015). Although only represented by a single isolated element, its size and shape does correspond to that of Tanystropheus spp. and is more similar to that of T. longobardicus, T. conspicuus, and T. haasi, than that of T. antiquus or T. fossai. It was identified as cf. Tanystropheus sp. It represents the first occurrence of Tanystropheus from North America.
In summary, prior to this study, a total of six species of Tanystropheus were acknowledged, namely T. conspicuus, T. antiquus, T. longobardicus, T. meridensis, T. fossai, and T. haasi. However, of these taxa, the validity of T. meridensis has been strongly disputed, whereas T. conspicuus has also been considered indistinguishable from T. longobardicus. Furthermore, the assignment of T. antiquus and T. fossai to the genus Tanystropheus has also been questioned, although T. antiquus is currently tentatively accepted as belonging to this genus. Additional specimens attributable to the genus Tanystropheus have been described from the Jilh Formation of Saudi Arabia (Middle Triassic), Makhtesh Ramon in Israel (Anisian-Ladinian), the Economy Member of the Wolfville Formation of south-eastern Canada (Middle to earliest Late Triassic), the Zhuganpo Member of the Falang Formation (latest Ladinian to earliest Carnian) of Guizhou Province, southern China, the earliest to Late Triassic of northern Italy, with the majority of specimens coming from the late Anisian, and the Upper Muschelkalk of Siles in Andulusia, Spain (Anisian-Ladinian) (Supplementary Table 1; Figure 1).
MATERIAL AND METHODS
This study includes all figured and referenced material of Tanystropheus, with critical taxa (T. longobardicus, T fossai, T. meridensis, T. conspicuus) having been studied personally and some more fragmentary additional material taken from the literature.
Institutional Abbreviations
BSPG, Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany; GMPKU, Geological Museum of Peking University, Beijing, China; HUJ-Pal, Paleontological Collections of the Department of Evolution, Systematics and Ecology of the Hebrew University of Jerusalem, Jerusalem, Israel; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China; MFSN, Museo Friulano di Scienze Naturali, Udine, Italy; MCSN, Museo Cantonale di Scienze Naturali di Lugano, Lugano, Switzerland; MCSNB, Museo Civico di Scienze Naturali "E. Caffi" Bergamo, Bergamo, Italy; MGUWr, Geological Museum, Institute of Geological Sciences, University of Wrocław, Wrocław, Poland; MSNM, Museo di Storia Naturale, Milan, Italy; PIMUZ, Paläontologisches Institut und Museum der Universität Zürich, Zurich, Switzerland; PIN, Paleontological Institute of the Russian Academy of Sciences, Moscow, Russia; SMNS, Staatliches Museum für Naturkunde, Stuttgart, Germany; U-MO, Urwelt-Museum Oberfranken, Bayreuth, Germany; YPM, Yale Peabody Museum, New Haven, Connecticut, USA.
RESULTS
Revision of the Various Tanystropheus Taxa and Specimens
Tanystropheus antiquus. The recent rediscovery of specimens used in the description in Huene (1907-1908) for Tanystropheus antiquus invalidates the diagnoses for this species in Wild (1973) and Fraser and Rieppel (2006), as well as that of Sennikov (2011) for Protanystropheus antiquus encompassing the same material, since all of these considered most of the original material to be lost and therefore did not include it (Skawiński et al., 2017). Based on observations of this original material, T. antiquus was considered to be a valid species and P. antiquus was rejected. However, in anticipation of a more detailed revision of these specimens and other tanystropheid material from Poland, an emended diagnosis for the species was not provided (Skawiński et al., 2017; Szczygielski, personal commun., 2019). Therefore, we currently consider only the cervical vertebrae from the Gogolin Formation that have previously been identified as T. antiquus in Huene (1907-1908) to belong to this species. Until the revision of this material has been completed, it can preliminarily be distinguished from other Tanystropheus species based on the neck vertebrae having a centrum length of less than three times their minimum height (i.e., having comparably shorter cervical vertebrae than other Tanystropheus species; sensu Fraser and Rieppel, 2006). We agree with Fraser and Rieppel (2006) that the other cervical vertebrae found in the Lower Muschelkalk of Europe (i.e., Vossenveld Formation of Winterswijk, the Netherlands, Schaumkalk Formation of Rüdersdorf and Isserstedt) likely can also be attributed to this species, as their morphology corresponds to the preliminary diagnosis, and identify this material here as T. cf. T. antiquus (following Spiekman et al., 2019). Material with tanystropheid affinities from these localities other than cervical vertebrae, including the femur (MGUWr 3894s) from the Gogolin Formation, can currently not be assigned to this species, as its preliminary diagnosis only applies to cervical vertebrae, and are identified here as Tanystropheus sp. These remains could be considered, however, if the revision of the Polish tanystropheid material also includes non-cervical material. The specimens from the Lower Muschelkalk of Bonndorf and Diedesheim, Germany, and the vertebra from Podloer Bruch near Gogolin are here considered indeterminable due to lack of diagnostic features and assigned to ?Archosauromorpha indet.
 Tanystropheus meridensis. Tanystropheus meridensis was described based on a single specimen, PIMUZ T 3901 (Figure 2A-C). This specimen from the Meride Limestone is of Ladinian age in contrast to the Anisian-Ladinian boundary age of the specimens of the Besano Formation previously assigned to T. longobardicus, and the former was considered to represent a slightly more derived form than the latter (Wild, 1980a). In this regard, it is relevant that a small-sized skeleton missing the skull and anterior part of the cervical column subsequently found in slightly older deposits of the Meride Limestone could not be distinguished from specimens referred to T. longobardicus and was identified as T. cf. longobardicus (Renesto, 2005). This was later corroborated, and the specimen was even considered to be identifiable to T. longobardicus (Nosotti, 2007). Although never formally reassigned, T. meridensis was considered to be indistinguishable from T. longobardicus in Fraser et al. (2004) and Nosotti (2007), with the holotype having been studied in detail in the latter. A new interpretation of some of the skull bones was provided therein, and it was compared to the small-sized specimens assigned to T. longobardicus housed in the collections of PIMUZ and MSNM, most notably MSNM BES SC 1018.
Tanystropheus meridensis. Tanystropheus meridensis was described based on a single specimen, PIMUZ T 3901 (Figure 2A-C). This specimen from the Meride Limestone is of Ladinian age in contrast to the Anisian-Ladinian boundary age of the specimens of the Besano Formation previously assigned to T. longobardicus, and the former was considered to represent a slightly more derived form than the latter (Wild, 1980a). In this regard, it is relevant that a small-sized skeleton missing the skull and anterior part of the cervical column subsequently found in slightly older deposits of the Meride Limestone could not be distinguished from specimens referred to T. longobardicus and was identified as T. cf. longobardicus (Renesto, 2005). This was later corroborated, and the specimen was even considered to be identifiable to T. longobardicus (Nosotti, 2007). Although never formally reassigned, T. meridensis was considered to be indistinguishable from T. longobardicus in Fraser et al. (2004) and Nosotti (2007), with the holotype having been studied in detail in the latter. A new interpretation of some of the skull bones was provided therein, and it was compared to the small-sized specimens assigned to T. longobardicus housed in the collections of PIMUZ and MSNM, most notably MSNM BES SC 1018.
Tanystropheus meridensis was distinguished based on the following combination of characters in the diagnosis of Wild (1980a): Skull morphology as in T. longobardicus with the exception of the following: presence of paired parietals, a probably edentulous palatine, and a quadrate that is more elongate and has a more concave posterior margin; a premaxilla bearing five single cusped teeth; a maxilla bearing three single cusped teeth and 12 tricuspid teeth; a lower jaw with three large single cusped and 16(?) tricuspid teeth; all teeth bear sharpened ridges, mainly on their anterior and posterior edge; cervical vertebrae similar to T. longobardicus, but slightly more elongate; fourth to sixth cervicals bearing a long horizontal lamina on the lateral margin of their centrum with a foramen positioned ventral to this lamina.
Re-analysis of PIMUZ T 3901 reveals that this diagnosis is problematic and results in the conclusion that Tanystropheus meridensis is indeed indistinguishable from the material previously assigned to T. longobardicus and should thus be considered synonymous with the latter, as will be demonstrated in the following. The parietals of PIMUZ T 3901 were considered to be unfused in Wild (1980a). However, the parietals were reinterpreted in Nosotti (2007), in which the element indicated as the right parietal in Wild (1980a) was identified as representing both parietals and the left parietal of Wild (1980a) was not identified. Personal observation (by SNFS and TMS) did not allow for a confident identification of these bones, and neither interpretation can be excluded (Figure 2B). This would imply that the fusion of the parietals cannot be assessed unambiguously, which invalidates this character as being diagnostic for T. meridensis. In any case, the fusion of the parietals can be considered a poor diagnostic character, as it is highly dependent on the ontogenetic stage of the specimen. The absence of teeth on the palatine would differentiate PIMUZ T 3901 from small specimens identified as T. longobardicus, although the palatines of larger specimens assigned to T. longobardicus are edentulous. The palatine of PIMUZ T 3901 indicated in Wild (1980a) was not identified as such in Nosotti (2007), in which that element was not identified due to the poor preservation of the skull in that region. Personal observation of this element (by SNFS) does not reveal the presence of any teeth or alveoli on this element, but also reveals no features that would identify this bone as a palatine (Figure 2B). Therefore as in Nosotti (2007), we consider the presence of the palatine and the presence of teeth on the palatine to be indeterminable in PIMUZ T 3901.
The quadrate was considered to be a diagnostic element for Tanystropheus meridensis that, apart from the characters in the diagnosis, additionally differed from that of the quadrate of T. longobardicus in having a wider pterygoid ramus and a convexity on its anterior margin (Wild, 1980a). The wider pterygoid ramus in T. meridensis was interpreted to represent a more derived state, since Macrocnemus bassanii, which was regarded to be a more basal taxon of the “prolacertiform” lineage, also was considered to bear a shorter pterygoid ramus. The pterygoid ramus of M. bassanii is indeed much shorter than that of T. longobardicus (personal observation SNFS). However, the pterygoid ramus cannot be identified in the figures presented in Wild (1980a), and personal observation (by SNFS) reveals that the quadrate is badly broken, strongly hampering any unambiguous observation of this element (Figure 2B). Nevertheless, the structure that was likely interpreted as the anterior convexity in Wild (1980a) could represent a part of the compressed pterygoid ramus, and it is not noticeably wider than that of PIMUZ T 2484. In Nosotti (2007), the shape of the left quadrate of MSNM BES SC 1018, a small-sized specimen of T. longobardicus, which was not available to Wild, was considered to be the closest in morphology to that of PIMUZ T 3901 among the PIMUZ material. This specimen is preserved under roughly the same angle as in PIMUZ T 3901. In any case, the strong compression of the specimen precludes any definitive statement on this aspect of the quadrate or its taller size and concave posterior margin mentioned in the diagnosis for T. meridensis in Wild (1980a). We therefore corroborate the assessment in Nosotti (2007) that the quadrate of PIMUZ T 3901 is indistinguishable from the quadrate of that present in the material previously assigned to T. longobardicus.
A detailed overview of the ontogenetic variation of Tanystropheus longobardicus, including the transition from a partially tricuspid marginal dentition to a dentition consisting of solely single cusped teeth, was presented by Wild (1973, p. 124-126). Regardless of whether the presented variation is related to ontogeny, this indicates that the number of tricuspid compared to single cusped teeth is variable among the small morphotype of T. longobardicus, and the dentition of PIMUZ T 3901 does not show a larger deviation than is shown within these specimens (Wild, 1973; Table 1; see also the section “The small and large morphotype of Tanystropheus from Monte San Giorgio: two taxa or ontogenetic variation?” below). The presence of cutting edges described by Wild (1980a) is corroborated. However, similar edges are also found in specimens attributed to T. longobardicus (e.g., MSNM BES SC 1018), and they likely represent a feature related to tooth wear (see Njau and Blumenschine, 2006 for similar wear of crocodylian teeth).
Wild (1980a) stated that the cervical vertebrae of PIMUZ T 3901 were slightly longer than those of specimens of similar size previously attributed to Tanystropheus longobardicus and that this provided additional support for the hypothesis that T. meridensis was more derived than T. longobardicus. The support for this claim appears to be absent, however; since the graph provided by Wild (1980a, figure 7) shows the second to sixth cervical vertebra preserved in PIMUZ T 3901 to be virtually identical in size to the corresponding vertebrae in the similarly sized specimen PIMUZ T 2795, well within the margin of deviation one can expect to be attributable to intraspecific variation. This latter specimen was previously assigned to T. longobardicus (Wild, 1973).
Finally, PIMUZ T 3901 was considered to differ from Tanystropheus longobardicus in bearing a long horizontal lamina on the lateral margin of the centrum with a foramen positioned directly below it in the fourth to sixth cervical (Wild, 1980a). The foramina cannot be observed and likely were misidentified cracks that occur throughout the specimen. The laminae are pronounced and clearly discernible, but these are similar to those visible in other small-sized species attributed to T. longobardicus (e.g., PIMUZ T 2484; Figure 2C and D).
A comparison of PIMUZ T 3901 with small-sized specimens attributed to Tanystropheus longobardicus is difficult because all specimens are heavily compressed. Nevertheless a comparison based mainly on PIMUZ T 2791, PIMUZ T 2484, and MSNM BES SC 1018 reveals no distinct differences with PIMUZ T 3901. Based on this, in addition to the refuted diagnosis of Wild (1980a) and the observations of previous studies (Fraser et al., 2004; Nosotti, 2007), T. meridensis is considered morphologically indistinguishable from small-sized specimens previously assigned to T. longobardicus. As such we propose Tanystropheus meridensis Wild 1980 to represent a junior synonym of Tanystropheus longobardicus Bassani 1886.
The small and large morphotype of Tanystropheus from Monte San Giorgio: Two taxa or ontogenetic variation? The specimens of Tanystropheus from Monte San Giorgio can be divided into two morphotypes, a small morphotype partially bearing tricuspid marginal dentition and a large morphotype bearing only single cusped dentition. A summary of the characters that distinguish the two morphotypes is presented in Table 2, and the relevant specimens of both morphotypes are listed in Table 1, Table 3, and Table 4, in which the specimens assigned to the small morphotype are indicated by an asterisk at the end of their respective specimen number. The distinction between the two morphotypes was already noted in the original description of the material (Peyer, 1931). However, at the time it was not determined whether these morphotypes represented distinct species or different ontogenetic stages of the same species in expectation of additional Tanystropheus findings from Monte San Giorgio. The variation observed in the Monte San Giorgio material was extensively described in Wild (1973, p. 124-140). The main differences that were recognized therein between the two morphotypes concerned the absence or presence of tricuspid marginal dentition, the number of teeth in the premaxilla and maxilla, the relative size of the orbits, the relative length of the maxillae, the shape of the premaxillae, frontals, parietals, and quadrates, and the shape of the bones of the palatal region and their dentition. It was concluded that there was a gradual transition in these characters from the small morphotype to the large morphotype, and therefore they were considered to represent a juvenile and adult form of the same species, respectively.
However, it has subsequently been suggested that the lack of a long postnarial process and distinct tricuspid dentition in the large morphotype, which are present in the small morphotype, does indeed indicate that the two morphotypes represent two different species (Fraser et al., 2004; but see also Renesto, 2005). We identified the differences between the two morphotypes and document herein in detail how the variation is distributed between the specimens from Monte San Giorgio to establish whether there is a gradual transition from the smaller morphotype to the larger morphotype as the specimens increase in size. If this is the case, this would be a strong indication that the observed differences represent ontogenetic variation. If no gradual transition between the two morphotypes can be observed, this would be an indication that they represent different species. In order to trace these characters through the growth series we ordered the specimens based on their relative size. Because the considered characters consist largely of cranial characters and most relevant specimens do not preserve the limbs, and because most of the considered skulls are disarticulated, relative size was based on the mean of the length of the tooth-bearing margin of the premaxilla, the maxilla, and the dentary (Table 3).
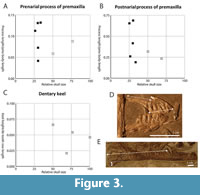 Most of these characters can only be assessed qualitatively, but the relative size of the prenarial and postnarial processes of the premaxilla and the relative size of the dentary keel were calculated for each specimen (Figure 3 and Table 4). These results indicate that the size of the prenarial and postnarial processes of the premaxilla vary strongly in size in the smaller morphotype. In the larger morphotype, these processes are either absent or very small (PIMUZ T 2790 for the prenarial process and PIMUZ T 2787 and PIMUZ T 2792 for the postnarial process). When looking at the size of the dentary keel it becomes evident that this structure is absent in all specimens of the small morphotype, whereas the relative size of the keel in the large morphotype does not appear to increase with overall size.
Most of these characters can only be assessed qualitatively, but the relative size of the prenarial and postnarial processes of the premaxilla and the relative size of the dentary keel were calculated for each specimen (Figure 3 and Table 4). These results indicate that the size of the prenarial and postnarial processes of the premaxilla vary strongly in size in the smaller morphotype. In the larger morphotype, these processes are either absent or very small (PIMUZ T 2790 for the prenarial process and PIMUZ T 2787 and PIMUZ T 2792 for the postnarial process). When looking at the size of the dentary keel it becomes evident that this structure is absent in all specimens of the small morphotype, whereas the relative size of the keel in the large morphotype does not appear to increase with overall size.
The amount of marginal teeth and the distribution of tricuspid dentition in relation to ontogeny was discussed in detail in p. 124-126 of Wild (1973) in which it was found that a gradual transition occurs from the smallest to the largest specimens of Tanystropheus from Monte San Giorgio. In the smallest specimens, all the maxillary teeth and the dentary teeth with which they articulate are tricuspid, whereas the premaxillary teeth and the dentary teeth articulating with these are pointed single-cusped teeth. As the specimens become larger, the tricuspid dentition was described as being progressively replaced by single-cusped teeth from anterior to posterior until the entire marginal dentition is made up of single-cusped teeth in the largest specimens, which were considered to be sexually mature, thus indicating that the difference in dentition can be attributed to ontogenetic variation rather than a taxonomic distinction (Wild, 1973, figure 80). Our findings, which include the specimens considered in Wild (1973), as well as PIMUZ T 1277, PIMUZ T 3901 (previously the holotype of T. meridensis), MSNM BES SC 265, and MSNM BES SC 1018, are presented in Table 1. We found that all the specimens in which the tooth count could be established with certainty bore six premaxillary teeth and 15 maxillary teeth, in contrast to Wild (1973), in which the tooth count of these elements was considered to increase with the size of the specimens. We found the position of the anteriormost tricuspid tooth to vary in the specimens bearing tricuspid dentition, ranging from the first or second tooth position on the maxilla (MSNM BES SC 1018) to the seventh (PIMUZ T 2482) counted from anterior in specimens that are roughly subequal in size (Table 3). The presence of tricuspid teeth on the posterior end of the dentary was documented in PIMUZ T 2792 (Wild, 1973). This observation could not be corroborated, and we were also not able to observe any tricuspid tooth in any of the other specimens attributed to the large morphotype.
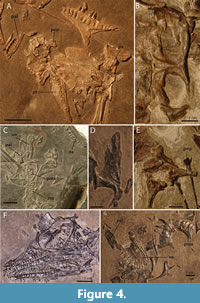 The shape of the parietal varies strongly among the observed specimens. The most consistent difference between the small and large morphotype is that the parietal is very wide and flat and does not bear a distinct supratemporal fossa in the small morphotype (Figure 4A). The parietal of the large morphotype has a much narrower intertemporal region of the parietal because of a strongly ventrally slanting supratemporal fossa. Furthermore, a distinct sagittal crest runs along the mid-line of the parietal in the large morphotype that diverges laterally on each side of the parietal lateral to the pineal foramen (Figure 4B). Additionally the parietal of PIMUZ T 2819 bears conspicuous anterolateral processes, which are absent in the smaller specimens. These processes are also absent in PIMUZ T 2787, another specimen belonging to the large morphotype. However, because the articulating parietal and frontals are preserved in ventral view, it is possible that the anterolateral processes are covered by the frontals ventrally. Although it is preserved in dorsal view, it is even possible that the parietal of PIMUZ T 2484 bears anterolateral processes, but that these were covered by the postfrontals, which cannot be identified with certainty in PIMUZ T 2819 and might have been lost or displaced. The presence of the anterolateral processes of the parietal therefore remains unclear in Tanystropheus from Monte San Giorgio with the exception of PIMUZ T 2819. The difference of the shape of the parietal between the small and the large morphotype could be attributed to ontogenetic differences, as it is strongly variable through ontogeny in extant non-avian sauropsids. In Varanus panoptes for instance, the parietal of the juvenile is also much wider and lacking the ventrally slanting supratemporal fossae seen in the adult (Werneburg et al., 2015, supplementary material).
The shape of the parietal varies strongly among the observed specimens. The most consistent difference between the small and large morphotype is that the parietal is very wide and flat and does not bear a distinct supratemporal fossa in the small morphotype (Figure 4A). The parietal of the large morphotype has a much narrower intertemporal region of the parietal because of a strongly ventrally slanting supratemporal fossa. Furthermore, a distinct sagittal crest runs along the mid-line of the parietal in the large morphotype that diverges laterally on each side of the parietal lateral to the pineal foramen (Figure 4B). Additionally the parietal of PIMUZ T 2819 bears conspicuous anterolateral processes, which are absent in the smaller specimens. These processes are also absent in PIMUZ T 2787, another specimen belonging to the large morphotype. However, because the articulating parietal and frontals are preserved in ventral view, it is possible that the anterolateral processes are covered by the frontals ventrally. Although it is preserved in dorsal view, it is even possible that the parietal of PIMUZ T 2484 bears anterolateral processes, but that these were covered by the postfrontals, which cannot be identified with certainty in PIMUZ T 2819 and might have been lost or displaced. The presence of the anterolateral processes of the parietal therefore remains unclear in Tanystropheus from Monte San Giorgio with the exception of PIMUZ T 2819. The difference of the shape of the parietal between the small and the large morphotype could be attributed to ontogenetic differences, as it is strongly variable through ontogeny in extant non-avian sauropsids. In Varanus panoptes for instance, the parietal of the juvenile is also much wider and lacking the ventrally slanting supratemporal fossae seen in the adult (Werneburg et al., 2015, supplementary material).
The most striking differences between the two morphotypes are seen in the palatal region, namely in the shape of the vomer, palatine, pterygoid, and their dentition. They differ in the presence of teeth on the pterygoid and palatine in the small morphotype, which are absent in the large morphotype, and on the vomer the dentition is larger and more recurved in the large morphotype than in the small morphotype (Figure 4, Table 1). The number of teeth in the vomer also differs in specimens belonging to the large morphotype, namely between PIMUZ T 2790 and PIMUZ T 2787 (Table 1). The tooth rows of both specimens are fully preserved, and their tooth count could therefore be established with certainty. Similarly, the tooth count of the vomer of the small specimen PIMUZ T 2779 could also unambiguously be determined. In specimens PIMUZ T 2792 and PIMUZ T 2795 observation was somewhat hampered by poor superficial preservation of the vomers, and their tooth count could be up to three teeth more than documented in Table 1. Nevertheless, it appears that the tooth count of the vomer varied in both morphotypes, and it is likely neither a taxonomically nor ontogenetically diagnostic character, since palatal tooth count has been shown to be intraspecifically variable in a number of extant palatal teeth-bearing squamates (Mahler and Kearney, 2006). The tooth count of the palatine could only be established with certainty in PIMUZ T 2484 and PIMUZ T 2482 of the small morphotype (Figure 4A and C). As stated above, in the large morphotype palatine teeth are absent (PIMUZ T 2787; Figure 4G). In PIMUZ T 2484, the tooth count is six, whereas it is five in that of the somewhat larger specimen PIMUZ T 2482. The pterygoid teeth are absent in the large morphotype (PIMUZ T 2787; Figure 4D). For the small morphotype the amount of pterygoid teeth can be counted in PIMUZ T 2795 (around 12), PIMUZ T 2484 (around 13; Figure 4A), and MSNM BES SC 1018 (around 12; Figure 4F). However, a definitive tooth count is hard to establish because the alveoli are very small, but the amount of pterygoid teeth is likely subject to intraspecific variation. Furthermore, these three specimens of the small morphotype are very similar in relative size and therefore do not allow for an assessment of the tooth count in disparate ontogenetic stages within the small morphotype (Table 3).
The vomer of the large morphotype is very wide anteriorly and thus forms a wide contact with the premaxilla and has a similar curvature as this bone (Figure 4G). It only allowed for the presence of a narrow choana laterally. The tooth row extends along the outer margin of the bone, and the vomerine teeth are quite large and recurved. The vomer of the small morphotype on the other hand is an anteroposteriorly straight bone (Figure 4C), similar to, but considerably shorter relatively, than the vomer of Macrocnemus spp. (Jaquier et al., 2017). The vomerine tooth row is straight and bears small teeth. The right vomer is fully preserved in ventral view in the smallest specimen belonging to the large morphotype (PIMUZ T 2792) and both vomers are preserved in articulation and in ventral view in the second to largest specimen of the small morphotype (PIMUZ T 2795). The difference in calculated relative estimated skull size between these two specimens is 20.4% (Table 3). Nevertheless, the vomer of PIMUZ T 2792 is indistinguishable from the vomers of larger specimens PIMUZ T 2787 and PIMUZ T 2790, and the vomers of PIMUZ T 2795 are indistinguishable from those of the smaller specimen PIMUZ T 2779. It seems unlikely that a gradual transition occurred from the morphology seen in the small morphotype to that seen in the large morphotype in the growth trajectory in between PIMUZ T 2795 and PIMUZ T 2792, and, therefore, this character appears to support a taxonomic distinction between the two morphotypes.
The palatine is well-preserved in PIMUZ T 2787 among the specimens of the large morphotype and in PIMUZ T 2484, PIMUZ T 2482, and MSNM BES SC 1018, with a likely partial palatine present in PIMUZ T 2795, among the specimens of the small morphotype (Figure 4A and G). Apart from being edentulous, the palatine of the large morphotype is different in being wider and more posteriorly extended with a smaller lateral extension articulating with the maxilla. Among the specimens of the small morphotype, there is a distinct difference in the size of the alveoli, which are comparatively very large in PIMUZ T 2484. In MSNM BES SC 1018, they are still larger than the alveoli on the pterygoid but comparatively smaller than those of PIMUZ T 2484. They are smaller still in PIMUZ T 2482 and PIMUZ T 2795, if the element in the latter specimen is indeed a palatine and not a fragment of the pterygoid. According to the relative size estimates, these four specimens are very similar in overall size (Table 3). Therefore, it appears that the observed variation is related to intraspecific variation independent of ontogeny. The shape of the palatine of PIMUZ T 2484 and PIMUZ T 2482, the two specimens of the small morphotype preserving a complete palatine, is indistinguishable. The overall shape of the palatines of the small morphotype is strongly distinct from that of the large morphotype. This and the presence of teeth on the palatine, which are absent in the large morphotype, form clear distinctions between the two morphotypes that appear unlikely to be solely attributable to ontogenetic or any other form of intraspecific variation.
The pterygoid of the large morphotype is characterized by a wide anterior portion or palatal ramus that is somewhat rounded anteriorly. This element is best preserved in PIMUZ T 2787 (Figure 4D). The pterygoid of the small morphotype has a much narrower palatal ramus that tapers to an end anteriorly, best preserved in PIMUZ T 2484 (Figure 4A). PIMUZ T 2792, the smallest specimen attributable to the large morphotype, preserves a single pterygoid, either the left pterygoid in dorsal view or the right in ventral view. It is incomplete, missing most of the quadrate ramus and part of the transverse flange. Furthermore, part of the palatal ramus is broken as indicated by its irregular medial margin, and therefore its morphology cannot unambiguously be observed. No teeth or alveoli can be observed on this element but it cannot be established whether it is preserved in dorsal or ventral view.
The ectopterygoid has been tentatively identified in PIMUZ T 2795 and PIMUZ T 2787 (Wild, 1973). Its morphology is poorly known, and the elements identified as such are similar in the two specimens.
The preservation of the opisthotic including the paroccipital process can only be observed in PIMUZ T 2484 among the specimens of the small morphotype and in PIMUZ T 2819 among the large morphotype specimens (Figure 4A and E). The difference between the two morphotypes is striking. However, none of the larger specimens belonging to the small morphotype or smaller specimens of the large morphotype preserve this element, and therefore there is no possibility of observing the morphology in the more intermediately-sized specimens. The shape of the braincase, including the opisthotic and its paroccipital process are known to change drastically in shape in extant diapsids. In Alligator mississippiensis and Varanus panoptes for instance, the paroccipital process becomes distinctly longer and narrower with age (Dufeau and Witmer, 2015, supplementary material; Werneburg et al., 2015, supplementary material), thus showing a similar disparity in shape from a juvenile to adult as seen in the small and large morphotype of Tanystropheus from Monte San Giorgio.
In the axial skeleton minor variation was mentioned (Wild, 1973). The cervical vertebrae were described as transitioning from a crescent-shaped cross section in the smaller specimens to a more triangular cross section in the larger specimens. Additionally the neural spines of the cervical vertebrae were considered to increase somewhat in relative size through ontogeny. No particular intraspecific variation was noted on in the dorsal column. The amount of caudal vertebrae bearing pleurapophyses was considered to increase with size, ranging from eight or nine in PIMUZ T 2791 to around 14 in PIMUZ T 2818. Due to the crushing of the specimens we were not able to observe the cross sectional shape of the cervical vertebrae in the material from Monte San Giorgio in detail. The isolated three-dimensional elements from the Upper Muschelkalk attributed to Tanystropheus conspicuus all represent specimens of the size range of the large morphotype, and therefore the presence of ontogenetic variation within that material cannot be confidently established.
Although not specifically discussed in Wild (1973), variation can be observed in the dorsal vertebrae of the Tanystropheus material from Monte San Giorgio in the relative height of the neural spine and length of the centrum, as well as the occurrence of laminae. Only PIMUZ T 2817 and PIMUZ T 2818 among the specimens of the large morphotype preserve an articulated dorsal column. In the former, apart from the anteriormost dorsal vertebrae, all dorsal vertebrae are partially broken, whereas in the latter, the specimen is heavily crushed and many characters of the dorsal vertebrae are obscured. Isolated dorsal vertebrae are also preserved in PIMUZ T 2787 and PIMUZ T 2792. Due to the isolated nature of these vertebrae, however, it is not possible to establish the exact position of each element in the vertebral column. Additionally the dorsal vertebral column is also complete in the Chinese specimen GMPKU-P-1527, which does not possess a skull but its size fits well within the size range of the large morphotype. However, its dorsal vertebrae are either covered by matrix or overlying ribs, or are only visible in ventral view. The dorsal part of the vertebral column is similarly disarticulated or heavily crushed in specimens of the small morphotype. The dorsal vertebrae can at least be partially counted and observed in MSNM BES SC 265 and PIMUZ T 1277. Although aspects of the dorsal column have been described for Tanystropheus from Monte San Giorgio (Wild, 1973; Renesto, 2005; Nosotti, 2007), no distinct differences between the large and small morphotype can be observed, and a comparison throughout the growth series is not possible.
The number of caudals bearing pleurapophyses in the specimens listed in Wild (1973, p. 136) could not be established with certainty due to the elements being poorly visible or too disarticulated. However, the number of pleurapophyses bearing caudals could be established in MSNM BES SC 265, a specimen of the small morphotype not available to Wild (Nosotti, 2007). The first eight caudals unambiguously bear pleurapophyses. Furthermore, in another specimen of the small morphotype, PIMUZ T 1277, at least six caudals bear pleurapophyses (Wild, 1980a). None of the specimens of the large morphotype from Monte San Giorgio allow for an exact count of caudal vertebrae bearing pleurapophyses. However, in the large Chinese specimen, GMPKU-P-1527, pleurapophyses can also be found in the first eight caudals (Rieppel et al., 2010). Although cranial material is absent in this specimen, its postcranial skeleton is morphologically indistinguishable from the large morphotype from Monte San Giorgio. Based on a comparison between these available specimens, we do not observe any distinct difference in this character between the two morphotypes as was suggested in Wild (1973).
Differences in the appendicular skeleton were documented in the shape of the pectoral and pelvic girdle elements (Wild, 1973). These differences all relate to proportional shape changes that do not appear to bear any taxonomic signal (see Wild, 1973, figures 89-93; personal observation SNFS and TMS). Similarly, variation among the limb elements is also minor and relates to the curvature of these elements (see Wild, 1973, figures 94-96; personal observation SNFS and TMS). We therefore consider these differences to be minor and easily attributable to ontogenetic variation, and they are, therefore, not considered to distinguish the two morphotypes.
In summary, the presence of prenarial and postnarial processes of the premaxillae is largely restricted to specimens of the small morphotype, and they are absent or very small in the large morphotype and the dentary keel present in the large morphotype is completely absent in the small morphotype (Table 2; Figure 3). A comparison of the relative size of the processes of the premaxilla in the small morphotype, and the dentary keel in the large morphotype reveals that much variation occurs in these characters that is apparently independent of size, indicating that it is likely not linked to ontogeny. We did not find any variation in the number of premaxillary and maxillary teeth within and between the two morphotypes in contrast to Wild (1973; see Table 1). We were also not able to identify the presence of triscupid teeth in PIMUZ T 2792 or any of the other specimens attributed to the large morphotype. We did find the position of the anteriormost tricuspid tooth on the maxilla to vary in specimens of the small morphotype, but apparently independent of size. Distinct differences are observed between the two morphotypes in the shape of the parietal and paroccipital process of the opisthotic (Table 2; Figure 4A-B and E). However, these differences could be the result of ontogenetic variation as observed in extant sauropsids. The palatal regions differ between the two morphotypes with the large morphotype having an anteriorly rounded vomer with large teeth and a narrow straight vomer with relatively much smaller teeth being present in the small morphotype (Figure 4C and G). The relatively small size difference between PIMUZ T 2792 and PIMUZ T 2795 but their very differently shaped vomers indicates that this difference is unlikely to be attributable to ontogeny. The palatine of the large morphotype differs from that of the small morphotype in being edentulous, wider, and more posteriorly extended with a relatively smaller lateral extension articulating with the maxilla (Figure 4A, C, and G). The size difference between the single specimen of the large morphotype preserving the palatine and the specimens of the small morphotype preserving the palatine is large. Nevertheless, the difference between the two morphotypes is disparate to such an extent that it seems unlikely that it is attributable to ontogeny. The pterygoid of the large morphotype differs from the small morphotype in being edentulous and with a wide and rounded palatal process (Figure 4A and D). Here, too, the size difference between the specimens of both morphotypes is large, but their morphology is similarly disparate that it seems unlikely that it is the result of ontogenetic variation. Tooth count is likely variable on all tooth-bearing palatal elements in both morphotypes independent of ontogeny (Table 1). However, the absence or presence of dentition on certain palatal elements is specific to each morphotype. In the postcranial skeleton, no consistent differences between the two morphotypes could be found.
The consistently observed differences in the shape of the premaxilla, dentary, and palatal elements are distinct and would represent a highly unusual ontogenetic transformation in Tanystropheus among sauropsids. Furthermore this transformation would have to occur in a relatively short part of the growth trajectory (between 33.7% and 50.24% of the maximum skull size estimated for Tanystropheus from Monte San Giorgio based on PIMUZ T 2819; Table 3). However, to unambiguously establish whether the two morphotypes represent different species based on a comparison of external morphology would require an overlapping size range between the two, which is not the case with the specimens currently known. Therefore, to establish whether the two morphotypes represent two distinct species requires additional study beyond a purely external morphological comparison, and we plan to use histological data and microtomographic scans to address this outstanding issue. Therefore, in the current study we are able to clearly separate two morphotypes in the Tanystropheus material from Monte San Giorgio. However, we find that it is currently not possible to exclude the possibility that the two morphotypes represent an ontogenetic series of the same taxon, and we therefore maintain the identification of the two morphotypes to T. longobardicus.
Tanystropheus conspicuus and Tanystropheus haasi. With the exception of a single undescribed dentary attributed to Tanystropheus conspicuus (SMNS 56289), both T. conspicuus and T. haasi are known from limited isolated postcranial material and are distinguished from other Tanystropheus species based on the morphology of their cervical vertebrae. These are similar in overall shape and size to those of the large morphotype of T. longobardicus, but can be easily distinguished from T. antiquus based on their relative length and are considerably larger than the cervical vertebrae known for the small morphotype of T. longobardicus and T. fossai. In order to assess the taxonomic validity of these taxa, the morphology of the cervical vertebrae of T. conspicuus and T. haasi is discussed here in detail and compared to each other and those of the large morphotype of T. longobardicus.
The diagnostic features of Tanystropheus conspicuus are currently unclear. T. conspicuus was distinguished from T. longobardicus based on the somewhat wider rib attachment sites and the bifurcating anterior end of the neural spine of the cervical vertebrae (Wild, 1973). This diagnosis was considered insufficient but maintained in expectation of the discovery of additional T. conspicuus material. The taxonomic status of T. conspicuus was later commented on in Wild (1980b), p. 204: “[...] and the probable identity of T. longobardicus and T. conspicuus from the Upper Muschelkalk (which at [the] moment cannot be proved, because of the lack of skull finds of the latter species). So in the Upper Muschelkalk only skeletal elements of adult specimens are found, never of juvenile ones. The preservation of juvenile skeletons of T. longobardicus in the Grenzbitumenzone is caused by the vicinity of the land during development of this basin [...].” The latter remark refers to the hypothesis that the small morphotype of Tanystropheus from Monte San Giorgio (T. longobardicus) was terrestrial (Wild, 1973), a view which has been disputed by various studies (e.g., Cox, 1975; Tschanz, 1986; Nosotti, 2007). Regardless, the material ascribed to T. conspicuus, although being represented only by isolated elements, largely cervical vertebrae, was considered to be indistinguishable from the large morphotype of Tanystropheus from Monte San Giorgio. The study was concluded with the following: “If it can be shown that the Upper Muschelkalk species T. conspicuus is identical with T. longobardicus, then this species would be a typical and widespread faunal element in the Tethyan and epicontinental Middle Triassic. New finds, which are under study, might confirm this assumption.” (Wild, 1980a, p. 205). However, no new material of T. conspicuus has been published on since, with the exception of the briefly mentioned fragmentary remains from Poland (Skawiński et al., 2017). In addition, GMPKU-P-1527 from the latest Ladinian or earliest Carnian of southwestern China could not be distinguished from the large morphotype of T. longobardicus (personal observation SNFS and TMS), and was identified as T. cf. T. longobardicus by Rieppel et al. (2010), indicating that at least one species of Tanystropheus occurred widespread throughout the Tethys. Regardless it is possible that multiple species of Tanystropheus co-occurred in the same habitat, as has been suggested for the fauna of Makhtesh Ramon (Rieppel, 2001) and would be the case if the small and large morphotype of T. longobardicus from Monte San Giorgio represent two separate species. Furthermore and crucially, the difference between or synonymy of T. conspicuus and the large morphotype of T. longobardicus cannot and should not be focused on temporal or spatial occurrence, but mainly on morphology (see also Simpson, 1951).
Tanystropheus haasi was distinguished within the genus Tanystropheus on the basis of 1) the presence of a deep horizontal groove directly dorsal to the centrum at the posterior end of the middle cervical vertebrae; 2) the presence of thickened margins of the articulation facets of the postzygapophyses; 3) a straight posterior margin of the postzygapophyseal trough; 4) which is furthermore located far posteriorly, namely dorsal to the posterior articulation surface of the centrum; 5) the presence of a long pointed posterior process of the neural spine that overlies the postzygapophyseal trough (Rieppel, 2001).
 We document the morphological variation between and within the cervical vertebrae of Tanystropheus conspicuus and T. haasi consisting of the diagnostic characters of T. haasi as well as additionally observed variation among and within these morphotypes, an overview of which is provided in Supplementary Table 2. The main findings are discussed below and subsequently compared to the cervical vertebrae of the large morphotype of T. longobardicus. Four cervical vertebrae that were included in the original description of T. conspicuus by Meyer (1855, plate 30) were studied firsthand, including U-MO BT 740 (Figure 5), the specimen identified as the lectotype of the species in Wild (1973), in addition to four other cervical vertebrae assigned to T. conspicuus housed in the U-MO (Figure 6). Tanystropheus haasi was studied by firsthand observation of a cast of the holotype of T. haasi (PIMUZ A/III 726, cast of HUJ-Pal. TR 1; Figure 7), and the additional material attributed to the species based on Rieppel (2001). When discussing these characters, it is important to consider that apart from the relative length, the morphology of the cervical vertebrae varies distinctly depending on the position of each vertebra within the cervical column, particularly at the anterior and posterior ends of each vertebra. This variation was described in detail for the three-dimensionally preserved cervical vertebrae of T. conspicuus, and based on this variation, the position in the cervical column of the various cervical vertebrae was assigned (Wild, 1973). However, this identification is inconclusive since all T. conspicuus vertebrae are preserved in isolation, thus not allowing the establishment of the exact position of each vertebra in the cervical column.
We document the morphological variation between and within the cervical vertebrae of Tanystropheus conspicuus and T. haasi consisting of the diagnostic characters of T. haasi as well as additionally observed variation among and within these morphotypes, an overview of which is provided in Supplementary Table 2. The main findings are discussed below and subsequently compared to the cervical vertebrae of the large morphotype of T. longobardicus. Four cervical vertebrae that were included in the original description of T. conspicuus by Meyer (1855, plate 30) were studied firsthand, including U-MO BT 740 (Figure 5), the specimen identified as the lectotype of the species in Wild (1973), in addition to four other cervical vertebrae assigned to T. conspicuus housed in the U-MO (Figure 6). Tanystropheus haasi was studied by firsthand observation of a cast of the holotype of T. haasi (PIMUZ A/III 726, cast of HUJ-Pal. TR 1; Figure 7), and the additional material attributed to the species based on Rieppel (2001). When discussing these characters, it is important to consider that apart from the relative length, the morphology of the cervical vertebrae varies distinctly depending on the position of each vertebra within the cervical column, particularly at the anterior and posterior ends of each vertebra. This variation was described in detail for the three-dimensionally preserved cervical vertebrae of T. conspicuus, and based on this variation, the position in the cervical column of the various cervical vertebrae was assigned (Wild, 1973). However, this identification is inconclusive since all T. conspicuus vertebrae are preserved in isolation, thus not allowing the establishment of the exact position of each vertebra in the cervical column.
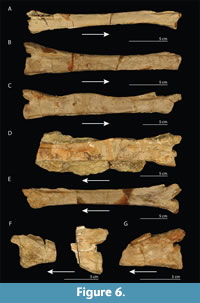 1) A horizontal groove is absent in the majority of the cervical vertebrae from the Upper Muschelkalk of Bindlacher Berg. However, specimens U-MO BT 732, U-MO BT 738, and U-MO BT 736 exhibit a larger excavation directly dorsal to the centrum than the other studied specimens, similar but somewhat less pronounced to that observed in PIMUZ A/III 726 and described for Tanystropheus haasi (Rieppel, 2001). 2) Distinct and wide, flat articulation surfaces directed lateroventrally can be observed in the majority of the studied specimens. But no distinct thickening of the articulation facets of the postzygapophyses could be observed. Similarly we consider the only preserved articulation facet of the left postzygapophysis of PIMUZ A/III 726 to not be distinctly thickened, but to be comparable in thickness and width to that of the cervical vertebrae from the Upper Muschelkalk of Bindlacher Berg. 3) In the studied specimens, a notch on the posterior margin of the postzygapophyseal trough could be observed in U-MO BT 734, U-MO BT 740, U-MO BT 736, and U-MO BT 738, whereas this margin was straight in U-MO BT 733, U-MO BT 739, and U-MO BT 732. In PIMUZ A/III 726 the posterior margin of the postzygapophyseal trough could not be observed since it is broken off posteriorly. 4) The posterior margin of the postzygapophyseal trough was located directly dorsal to the posteriormost end of the centrum in U-MO BT 733 and U-MO BT 734, whereas it is anterior to the posteriormost end of the centrum in the other studied specimens. In PIMUZ A/III 726 the posterior extent of the postzygapophyseal trough is located somewhat posterior to the posterior end of the centrum on its right side where it is complete. 5) A large posterior process of the neural spine as shown in Figure 4A of Rieppel (2001) cannot be observed in any of the studied T. conspicuus specimens. However, it is important to note that both the anterior and posterior ends of the neural spines are broken in virtually all of the specimens. In U-MO BT 736 the neural spine is not very distinct posteriorly, but it slightly protrudes over the postzygapophyseal trough. In PIMUZ A/III 726 the neural spine similarly protrudes very slightly over the postzygapophyseal trough, but it is much less expanded than n HUJ-Pal. TR 1447.
1) A horizontal groove is absent in the majority of the cervical vertebrae from the Upper Muschelkalk of Bindlacher Berg. However, specimens U-MO BT 732, U-MO BT 738, and U-MO BT 736 exhibit a larger excavation directly dorsal to the centrum than the other studied specimens, similar but somewhat less pronounced to that observed in PIMUZ A/III 726 and described for Tanystropheus haasi (Rieppel, 2001). 2) Distinct and wide, flat articulation surfaces directed lateroventrally can be observed in the majority of the studied specimens. But no distinct thickening of the articulation facets of the postzygapophyses could be observed. Similarly we consider the only preserved articulation facet of the left postzygapophysis of PIMUZ A/III 726 to not be distinctly thickened, but to be comparable in thickness and width to that of the cervical vertebrae from the Upper Muschelkalk of Bindlacher Berg. 3) In the studied specimens, a notch on the posterior margin of the postzygapophyseal trough could be observed in U-MO BT 734, U-MO BT 740, U-MO BT 736, and U-MO BT 738, whereas this margin was straight in U-MO BT 733, U-MO BT 739, and U-MO BT 732. In PIMUZ A/III 726 the posterior margin of the postzygapophyseal trough could not be observed since it is broken off posteriorly. 4) The posterior margin of the postzygapophyseal trough was located directly dorsal to the posteriormost end of the centrum in U-MO BT 733 and U-MO BT 734, whereas it is anterior to the posteriormost end of the centrum in the other studied specimens. In PIMUZ A/III 726 the posterior extent of the postzygapophyseal trough is located somewhat posterior to the posterior end of the centrum on its right side where it is complete. 5) A large posterior process of the neural spine as shown in Figure 4A of Rieppel (2001) cannot be observed in any of the studied T. conspicuus specimens. However, it is important to note that both the anterior and posterior ends of the neural spines are broken in virtually all of the specimens. In U-MO BT 736 the neural spine is not very distinct posteriorly, but it slightly protrudes over the postzygapophyseal trough. In PIMUZ A/III 726 the neural spine similarly protrudes very slightly over the postzygapophyseal trough, but it is much less expanded than n HUJ-Pal. TR 1447.
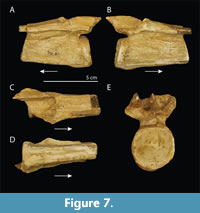 Additional variation in the cervical vertebrae could be found in the presence of the ventral foramen (foramina venae vertebralis sensu Wild, 1973, 1987; Figure 5D), the anteroposterior length of the postzygapophyseal canal, the height of the posterior opening of the neural canal, the depth of the excavation of the articular ends of the cervical centra, the shape of the articular ends of the cervical centra, how distinct the ventral keel is pronounced, and the dorsal extension of the neural spine. In U-MO BT 736 and U-MO BT 737 a distinct dorsal convexity of the neural spine can be observed slightly posterior to the mid-length of both vertebrae (Figure 5C-D). In other observed specimens, the neural spine is not markedly expanded here and straight. This character cannot be observed in PIMUZ A/III 726 since the majority of the vertebra is not preserved.
Additional variation in the cervical vertebrae could be found in the presence of the ventral foramen (foramina venae vertebralis sensu Wild, 1973, 1987; Figure 5D), the anteroposterior length of the postzygapophyseal canal, the height of the posterior opening of the neural canal, the depth of the excavation of the articular ends of the cervical centra, the shape of the articular ends of the cervical centra, how distinct the ventral keel is pronounced, and the dorsal extension of the neural spine. In U-MO BT 736 and U-MO BT 737 a distinct dorsal convexity of the neural spine can be observed slightly posterior to the mid-length of both vertebrae (Figure 5C-D). In other observed specimens, the neural spine is not markedly expanded here and straight. This character cannot be observed in PIMUZ A/III 726 since the majority of the vertebra is not preserved.
Overall, the observed variation in the Tanystropheus conspicuus material indicates that most of the features indicated as diagnostic for T. haasi fall within the range of intraspecific variation for T. conspicuus. However the horizontal groove dorsal to the posterior end of the centrum is less distinct in the specimens of T. conspicuus in which it is present than observed in T. haasi. Furthermore, the distinctly expanded posterior end of the neural spine overhanging the postzygapophyseal trough as described for HUJ-Pal. TR 1447 could not be observed for T. conspicuus, or in the cast PIMUZ A/III 726 of T. haasi. Our findings indicate that much variation occurs in the morphology of the cervical vertebrae of T. conspicuus, and that this variation overlaps with much of the morphology considered diagnostic for T. haasi. This comparison highlights the difficulty of differentiating species of tanystropheids on the basis of isolated cervical vertebrae alone, as much variation occurs along the cervical column and intraspecifically.
The majority of the characters discussed above cannot be observed in Tanystropheus longobardicus because the specimens attributed to this species of both the large and small morphotype were strongly flattened during fossilisation or because the cervical vertebrae are still in articulation with each other and therefore the pre- and postzygapophyseal regions are largely obscured. This therefore hampers a detailed comparison. The size and shape of the vertebral centra, neural spines, and zygapophyses that can be observed for the large morphotype of T. longobardicus are indistinguishable from T. conspicuus and T. haasi. Additionally, other isolated material assigned to T. conspicuus, comprising dorsal, sacral, and caudal vertebrae, as well as a humerus and femora, also cannot be distinguished from this taxon (Wild, 1973; personal observation SNFS). However, because only a very limited comparison is possible, and skull material, which is likely more diagnostic, is completely lacking for both taxa, an unambiguous taxonomic identification of T. conspicuus and T. haasi is not possible. Therefore we propose to consider both taxa as nomina dubia.
 Tanystropheus fossai. The only known specimen of Tanystropheus fossai, MCSNB 4035, consists of four articulated cervical vertebrae and associated cervical ribs and originates from the late Norian of northern Italy (Figure 8). It was interpreted to be a juvenile individual and distinguished from other species of Tanystropheus based on the absence of a neural spine and the presence of wing-like laterally projected crests on the posterior end of the centrum of the cervical vertebrae (Wild, 1980a). It cannot be determined whether the specimen represents a juvenile based solely on cervical vertebrae and ribs. Nevertheless we concur that the absence of the neural spine and laterally projected crests on the posterior ends of the centra of the cervical vertebrae represent diagnostic features not present in other tanystropheids. Additionally MCSNB 4035 also differs from other Tanystropheus taxa in having an elongated anterior free-ending process of the cervical ribs. Furthermore, dorsal to the vertebrae, a number of elongate and thin elements are preserved, which are clearly thinner than the cervical ribs preserved ventral to the vertebrae. Although figured, these elements were not commented on in Wild (1980a). They likely represent ossified tendons, similar to those present in many dinosaurian taxa (Organ and Adams 2005), but previously unknown for tanystropheids. Finally, as was also described in Wild (1980a), some of the cervical ribs appear to bifurcate (Figure 8B). It was unclear to Wild (1980a) whether these bifurcations represented a taxonomic feature or pathology. However, since no similar cases of bifurcating cervical ribs are known among amniotes, they most likely represent a pathological anomaly. Another possibility is that the apparent bifurcation of the ribs is formed by ossified tendons running parallel and attaching to the cervical ribs. The seemingly abundant presence of these tendons dorsal to the vertebrae possibly support this hypothesis. Regardless, we think it unlikely for these bifurcations to represent a taxonomic feature and therefore do not consider it a diagnostic character for the taxon. The relative length of the centra of the mid-cervical vertebrae is in correspondence with that found within the genus Tanystropheus, with the centrum being approximately 11 times longer than its minimum height in the second preserved vertebra from anterior (length 40.4 mm; height: 3.6 mm). This is in contrast to the observation presented in Renesto (2005), in which the cervical vertebrae of MCSNB 4035 were described as being considerably shorter than those of other Tanystropheus species. Although not identified as such, the morphology of the vertebrae of MCSNB 4035 was also likened to the caudal vertebrae of certain Triassic pterosaurs (Dalla Vecchia, 2000). However, it can be excluded that they indeed represent caudal vertebrae based on the associated cervical ribs. The articulation surfaces of the vertebrae appear to be amphicoelous.
Tanystropheus fossai. The only known specimen of Tanystropheus fossai, MCSNB 4035, consists of four articulated cervical vertebrae and associated cervical ribs and originates from the late Norian of northern Italy (Figure 8). It was interpreted to be a juvenile individual and distinguished from other species of Tanystropheus based on the absence of a neural spine and the presence of wing-like laterally projected crests on the posterior end of the centrum of the cervical vertebrae (Wild, 1980a). It cannot be determined whether the specimen represents a juvenile based solely on cervical vertebrae and ribs. Nevertheless we concur that the absence of the neural spine and laterally projected crests on the posterior ends of the centra of the cervical vertebrae represent diagnostic features not present in other tanystropheids. Additionally MCSNB 4035 also differs from other Tanystropheus taxa in having an elongated anterior free-ending process of the cervical ribs. Furthermore, dorsal to the vertebrae, a number of elongate and thin elements are preserved, which are clearly thinner than the cervical ribs preserved ventral to the vertebrae. Although figured, these elements were not commented on in Wild (1980a). They likely represent ossified tendons, similar to those present in many dinosaurian taxa (Organ and Adams 2005), but previously unknown for tanystropheids. Finally, as was also described in Wild (1980a), some of the cervical ribs appear to bifurcate (Figure 8B). It was unclear to Wild (1980a) whether these bifurcations represented a taxonomic feature or pathology. However, since no similar cases of bifurcating cervical ribs are known among amniotes, they most likely represent a pathological anomaly. Another possibility is that the apparent bifurcation of the ribs is formed by ossified tendons running parallel and attaching to the cervical ribs. The seemingly abundant presence of these tendons dorsal to the vertebrae possibly support this hypothesis. Regardless, we think it unlikely for these bifurcations to represent a taxonomic feature and therefore do not consider it a diagnostic character for the taxon. The relative length of the centra of the mid-cervical vertebrae is in correspondence with that found within the genus Tanystropheus, with the centrum being approximately 11 times longer than its minimum height in the second preserved vertebra from anterior (length 40.4 mm; height: 3.6 mm). This is in contrast to the observation presented in Renesto (2005), in which the cervical vertebrae of MCSNB 4035 were described as being considerably shorter than those of other Tanystropheus species. Although not identified as such, the morphology of the vertebrae of MCSNB 4035 was also likened to the caudal vertebrae of certain Triassic pterosaurs (Dalla Vecchia, 2000). However, it can be excluded that they indeed represent caudal vertebrae based on the associated cervical ribs. The articulation surfaces of the vertebrae appear to be amphicoelous.
Since the first identification of Tanystropheus fossai, many additional tanystropheid taxa with elongated cervical vertebrae have been described; e.g., Langobardisaurus pandolfii, Dinocephalosaurus orientalis, and Fuyuansaurus acutirostris (Renesto, 1994; Renesto and Dalla Vecchia, 2000; Li, 2003; Rieppel et al., 2008; Fraser et al., 2013). Most of the diagnostic features of T. fossai cannot be observed in these taxa. However, the extended anterior free-ending process of the cervical ribs present in T. fossai can also be observed in D. orientalis, as well as in Pectodens zhenyuensis, a long-necked diapsid of possibly tanystropheid affinity (Rieppel et al., 2008; Li et al., 2017). Although only known from a single, fragmentary specimen, we consider MCSNB 4035 to show distinct morphological disparity from known Tanystropheus taxa as to merit its assignment to a separate genus. This interpretation is additionally supported by the large temporal gap between MCSNB 4035 and the known Tanystropheus taxa (Figure 1) and the taxonomic diversity of long-necked tanystropheid taxa. We therefore reassign MCSNB 4035 to Sclerostropheus fossai (gen. nov.).
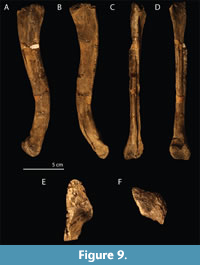 Specimens not identified on the species level. The isolated element PIMUZ A/III 771 from Piz Ravigliel represents a complete, mediolaterally crushed, right femur with a total length of 190 mm (Figure 9). It is strongly sigmoidal and bears a distinct crest-like medial trochanter that is confluent with the femoral shaft distally. This trochanter is displaced further posteriorly than in other Tanystropheus femora due to compression. The proximal head is somewhat broken, whereas the distal head is complete. It is indistinguishable from the femora of the large morphotype of T. longobardicus (e.g., PIMUZ T 2817) and those from the Upper Muschelkalk (SMNS 54623) and therefore we corroborate its identification as Tanystropheus sp. (Eichenberger, 1986).
Specimens not identified on the species level. The isolated element PIMUZ A/III 771 from Piz Ravigliel represents a complete, mediolaterally crushed, right femur with a total length of 190 mm (Figure 9). It is strongly sigmoidal and bears a distinct crest-like medial trochanter that is confluent with the femoral shaft distally. This trochanter is displaced further posteriorly than in other Tanystropheus femora due to compression. The proximal head is somewhat broken, whereas the distal head is complete. It is indistinguishable from the femora of the large morphotype of T. longobardicus (e.g., PIMUZ T 2817) and those from the Upper Muschelkalk (SMNS 54623) and therefore we corroborate its identification as Tanystropheus sp. (Eichenberger, 1986).
The isolated Tanystropheus remains from the late Anisian deposits of the Aupa Valley, Friuli, Italy, represent comprehensive material consisting of a single caudal vertebra (MFSN 25761) and possible dorsal rib (MFSN 25762) (Dalla Vecchia, 2000), and 30 specimens comprising cervical, dorsal, sacral, and caudal vertebrae, as well as teeth, a clavicle, and ilium described in Dalla Vecchia (2005). This material was identified as T. cf. T. longobardicus but was considered as possibly representing a different species based on the reduction of the neural spine on the posterior end of the cervical vertebrae and the long transverse processes of the dorsal vertebrae (Dalla Vecchia, 2005). Although representing extensive material, the absence of articulated specimens and cranial material precludes its precise taxonomic affinity, and therefore we consider it as Tanystropheus sp.
MTSN 3652, the partial isolated cervical vertebra from the late Ladinian of the Mendola Pass, Trentino, Italy, allows only for observation in lateral and ventral view and thus a very limited comparison is possible with the cervical vertebrae from the Upper Muschelkalk of Bindlacher Berg and Makhtesh Ramon, since no observations of important features such as those on the postzygapophyseal trough or the neural canal can be made. Therefore, we concur with the identification of this specimen to Tanystropheus sp. (Dalla Vecchia and Avanzini, 2002). Similarly in MFSN 25760, the posterior half of a cervical vertebra from the Fusea site of Friuli, Italy (late Ladinian to early Carnian), the same limitations in observation apply and this specimen is therefore also referred to Tanystropheus sp. (Dalla Vecchia, 2000).
YPM VPPU 022000, the virtually complete cervical vertebra from the Early to Middle Triassic Nova Scotia was identified as cf. Tanystropheus sp. by Sues and Olsen (2015). It is distinctly larger than Sclerostropheus fossai and apparently lacks wing-like laminae on its centrum and possesses a low neural spine. The relative length of the vertebra in comparison to the minimum height of its centrum (approximately 13) exceeds that known for T. antiquus (<3 sensu Fraser and Rieppel, 2006). Its size and overall morphology is in agreement with that observed for T. conspicuus, T. haasi, and the large morphotype of T. longobardicus, and therefore the specimen is identified here as Tanystropheus sp.
The articulated small-sized specimen preserving the posterior five cervical vertebrae, the trunk, forelimbs and girdles, and part of the hind limbs and girdles from the Falang Formation of Chajiang, Guizhou Province, China, IVPP V 14472, was considered morphologically indistinguishable from specimens referred to Tanystropheus longobardicus, but not identified on the species level (Li, 2007). Although its overall size is in correspondence with that of the small morphotype of T. longobardicus, no skull material is preserved that allows for the identification of characters associated with the small morphotype of T. longobardicus, such as the presence of tricuspid marginal teeth. Therefore, we corroborate its identification as Tanystropheus sp.
Apart from the material assigned to Tanystropheus haasi, two additional morphotypes of Tanystropheus from Makhtesh Ramon in Israel were recognized in (Rieppel, 2001), and it was suggested that multiple species of Tanystropheus might have coexisted there through habitat partitioning as hypothesized for the genera Nothosaurus and Saurichthys (Rieppel et al., 1997). Specimens referred to these two morphotypes were identified as Tanystropheus sp. They were recognized based on overall size and small morphological differences such as the postzygapophyses being oriented more ventrally rather than ventrolaterally, and the depth of the excavation of the horizontal groove also described for T. haasi. As shown in Supplementary Table 2 such variation also occurs in the cervical vertebrae from the Upper Muschelkalk of Bindlach, Germany. We consider this variation to be attributable to variation that occurs throughout the cervical column depending on the position of each cervical, as well as taphonomic alteration and intraspecific variation. However, the very large size of one of the two morphotypes, which is apparently distinctly larger than that of any other known Tanystropheus material, might be an indication that the two represent separate species. However, since this morphotype is represented by very limited material, we maintain its identification, as well as that of the smaller morphotype, as Tanystropheus sp. as proposed by Rieppel (2001). The Tanystropheus specimens described from the Jilh Formation of Saudia Arabia were not figured, and their identification therefore could not be assessed and we therefore tentatively maintain its identification as Tanystropheus sp. (Vickers-Rich et al., 1999).
The isolated cervical vertebrae from the Anisian of Romania were initially referred to Tanystropheus biharicus (Jurcsák, 1975, 1976, 1978, 1982) and subsequently reassigned to T. cf. longobardicus (Wild, 1980a). We find that the morphology, length to height ratio, and overall size of the vertebrae fit that seen for the large morphotype of T. longobardicu s, T. conspicuus, and T. haasi, and refer this material to Tanystropheus sp.
SYSTEMATIC PALAEONTOLOGY
DIAPSIDA Osborn 1903
ARCHOSAUROMORPHA von Huene, 1946
TANYSTROPHEIDAE Gervais, 1859
Genus TANYSTROPHEUS von Meyer, 1852
Diagnosis. Long-necked tanystropheid archosauromorphs distinguished from other genera by the following combination of characters (autapomorphy among non-archosauriform archosauromorphs marked with an asterisk): 1) the absence of shagreen-like teeth on the vomers and palatines; 2) mid-cervical vertebrae with a vertebral centrum length more than 2.5 times their minimum height; 3) a low neural spine of the cervical vertebrae; 4) very long and extremely thin cervical ribs, being approximately three times the length of their corresponding vertebra in the mid-cervical region, bearing short anterior free ending processes; 5) ribs of second sacral vertebrae not bifurcated; 6) curved anterior margin of scapular blade, which is consequently directed posteriorly; 7) the absence of hyperphalangy; 8) only two ossified distal carpals*; and 9) a thyroid fenestra present between the pubis and ischium.
Type species. Tanystropheus conspicuus (nomen dubium)
Included species. Tanystropheus longobardicus, Tanystropheus haasi (nomen dubium), and Tanystropheus antiquus.
Remarks. Tanystropheus conspicuus is considered to be represented by insufficient material to distinguish it convincingly from other Tanystropheus taxa, and we therefore consider it a nomen dubium. Nevertheless it remains the type species as the first named species of the genus, and it is diagnostic on the genus level based on the characteristic cervical vertebrae of the type material that are unique to the genus Tanystropheus.
Tanystropheus antiquus von Huene, 1905, currently only encompasses the material described in Huene (1907-1908) since the analysis of more recent material assigned to the species (e.g., Wild, 1973; Fraser and Rieppel, 2006; Sennikov, 2011) did not take most of this material into consideration as it was presumed to have been lost, which is not the case (Skawiński et al., 2017). This original material is currently under study, and, therefore, we refrain from assessing this taxon in detail and consider all material later assigned to this taxon preliminarily to Tanystropheus cf. T. antiquus.
Like Tanystropheus conspicuus, T. haasi is considered a nomen dubium as the limited material assigned to the species is insufficient to confidently distinguish the species from other species within the genus. Sclerostropheus fossai was previously attributed to Tanystropheus but is herein considered sufficiently distinct from other known Tanystropheus species to merit assignment to a separate genus.
Tanystropheus conspicuus (von Meyer, 1855) Figure 5-Figure 6
1846 Smilodon laevis (partim) Plieninger, p. 153, 247; plate 3, figure 4
1855 Tanystropheus conspicuus von Meyer, plate 30, figure 1-6, plate 46, figures 1-4
1890 Tanystropheus conspicuus Zittel, p. 567, 733; figure 514
1896 Tanystropheus conspicuus Fraas, p. 14-15
1902 Chelyzoon blezingeri von Huene, p. 51-52; plate 7, figure 1 (also mentioned in Kuhn, 1971, but appears very different from Tanystropheus)
1902 Chelyzoon latum von Huene, p. 50-51; plate 7, figure 2 (also mentioned in Kuhn, 1971, but appears very different from Tanystropheus)
1902 Procerosaurus cruralis von Huene, p. 64-65; plate 9, figure 1
1902 Pectenosaurus strunzi von Huene, p. 65; plate 6, figure 5 (synonymous with Tanystropheus sensu Kuhn, 1971, p, 11)
1905 Thecodontosaurus latespinatus von Huene, p. 349
1905 Tanystrophaeus conspicuus von Huene, p. 349
1907-1908 Thecodontosaurus latespinatus von Huene, p. 218-223, 263, 265, 270, 307, 310-312; text-figures 237-243; plate 91, figures 1-8, plate 92, figures 1-7
1907-1908 Tanystrophaeus conspicuus von Huene, p. 4, 223, 226-230, 231, 237-239, 264-265, 270, 305, 307, 312, 319, 321; text-figures 250-251, 351; plate 95, figures 1-4; plate 96, figures 1-10
1915 Tanystropheus conspicuus Broili, text; plate 2-3
1924 Tanystrophaeus conspicuus Edinger, figures 1-4
1928 Thecodontosaurus latespinatus Corroy, p. 126; text-figure 12; plate 4, figure 11
1928 Tanystrophaeus conspicuus Schmidt, p. 424-425; figure 1188
1928 Thecodontosaurus latespinatus Schmidt, p. 434; figure 1213
1931 Tanystropheus conspicuus Peyer, p. 11; text-figures 1-2; p. 21, 75-76, 92, 95-98; text-figures 24-25
1931 Tanystropheus conspicuus von Huene, 1931, throughout text and figures 1-6 (Thecodontosaurus latespinatus and Zanclodon laevis considered to be likely attributable to this species herein)
1932 Tanystropheus conspicuus von Huene p. 6-8 (Thecodontosaurus latespinatus and Zanclodon laevis considered to be likely attributable to this species herein)
1935 Macroscelosaurus conspicuus Kuhn, p. 118-119
1938 Tanystropheus conspicuus Schmidt, p. 75-76, figure 1188a
?1952 Tanystrophaeus conspicuus Rühle von Lilienstern, p. 27
1953 Tanystropheus conspicuus Adam, throughout text; figure 1
1955 Tanystropheus conspicuus Peyer, p. 489
1955 Tanystropheus conspicuus Peyer and Kuhn-Schnyder, p. 578; figure 1
1970 Thecodontosaurus latespinatus Colbert, p. 31-33
1971 Tanystropheus conspicuus Kuhn, p. 11; figures 21, 21a, 41b
1971 Thecodontosaurus latespinatus Kuhn, p. 11; figure 21
1973 Tanystropheus conspicuus Wild, p. 52-60, 67-94; figures 39-62; p. 105-106; figure 67; p. 114-115; figures 73, 97; p. 148-150
1975 Tanystropheus conspicuus Wild, p. 153; figure 1
1976 Tanystropheus conspicuus Wild, p. 20; figure 3
1980a Tanystropheus conspicuus Wild, p. 7, 10, 12-13, 25
1980b Tanystropheus conspicuus Wild, p. 201, 204-205
1986 Tanystropheus conspicuus Benton, p. 296-297
1986 Tanystropheus conspicuus Tschanz, p. 6, 59, 66-68, 84
1987 Tanystropheus conspicuus Wild, throughout text
1988 Tanystropheus conspicuus Tschanz, p. 997-998, 1002
1988 Tanystropheus conspicuus (synonymized with T. longobardicus) Evans, p. 227
1997 Tanystropheus conspicuus Benton and Allen, p. 945, 947
2000 Tanystropheus conspicuus Dalla Vecchia, p. 136, 138-139
2001 Tanystropheus conspicuus Rieppel, throughout text
2005 Tanystropheus conspicuus Dalla Vecchia, p. 40-42
2005 Tanystropheus conspicuus Renesto, p. 386
2007 Tanystropheus conspicuus Nosotti, throughout text
2012 Tanystrophaeus conspicuus Diedrich, p. 23; figure 14
2015 Tanystropheus conspicuus Pritchard, Turner, Nesbitt, Irmas, and Smith, p. 4, 9, 15
2015 Tanystropheus conspicuus Nesbitt, Flynn, Pritchard, Parrish, Ranivharimanana, Wyss, p. 23, 26-27, 62, 64
2016 Tanystropheus longobardicus Ezcurra, throughout text; figures 14a-b, 31b, 33b, 35, 43a-b
2017 Tanystropheus conspicuus Skawiński, Ziegler, Czepiński, Szermański, Tałanda, Surmik, and Niedźwiedzki, p. 4, 14-15; figure 5o-p
2018 Tanystropheus conspicuus Renesto and Saller, p. 23
Referred specimens. See Supplementary Table 1
Localities and occurrence. The Upper Muschelkalk (late Anisian to early Ladinian) of Central Europe; exact localities are listed in Supplementary Table 1. The type locality is the Upper Muschelkalk of Bindlacher Berg, near Bayreuth, Germany.
Previous diagnosis. The diagnosis for Tanystropheus conspicuus was provided in Wild (1973, p. 149; translated from German): “Up to six(?) meters long species, that can be distinguished from T. longobardicus by the wider attachment sites of the cervical ribs and the split anterior end of the neural spine for the attachment of the dorsal ligament. Although these minor differences in the construction of the cervical vertebrae of T. conspicuus are insufficient for a species definition, it is maintained as a separate species until later finds can offer clarification.”
Remarks. Tanystropheus conspicuus, represented by isolated vertebrae, femora, and a humerus, can currently not be distinguished from the large morphotype of T. longobardicus. However, the material of T. conspicuus is currently too limited to allow for a detailed comparison, and therefore the two species cannot be synonymized and we consider T. conspicuus to be a nomen dubium.
Tanystropheus antiquus (von Huene 1907-1908)
1905 Tanystrophaeus antiquus (nomen nudum) von Huene, p. 349
?1905 Thecodontosaurus primus (nomen nudum) von Huene, p. 349
1907-1908 Tanystrophaeus antiquus von Huene, p. 223-226; text-figures 246-249; p. 264-265, 305, 311; plate 93, figures 1-6, plate 94, figures 2-5
1907-1908 Tanystrophaeus primus von Huene, p. 221
?1907-1908 Thecodontosaurus primus von Huene, p. 217-218, 263, 265, 307-308, 311; plate 92, figures 8-9
1915 Tanystropheus antiquus Broili, p. 52-53, 58, 61
?1928 Tanystrophaeus cf. antiquus Corroy, p. 126-127; text-figure 13 (not Tanystropheus sensu (Peyer 1931) p. 13)
1928 Tanystrophaeus antiquus Schmidt, p. 424; figure 1187
?1928 Thecodontosaurus primus Schmidt, p. 434; figure 1212
1931 Tanystropheus antiquus Peyer, p. 93-94, 104-106; text-figure 28
1931 Tanystropheus antiquus von Huene, throughout text; figures 7-17
1935 Macroscelosaurus antiquus Kuhn, p. 119
1938 Tanystropheus antiquus Schmidt, p. 74-75, figure 1187
1944 Tanystropheus latespinatus von Huene, p. 127
1971 Tanystropheus antiquus Kuhn, p. 11; figures 21, 21a, 41a
1971 “Thecodontosaurus primus” Kuhn, p. 11, 17; figure 21
1973 Tanystropheus antiquus Wild, p. 142-143, 151-152
1975 Tanystropheus antiquus Wild, p. 153
1980a Tanystropheus antiquus (partim) Wild, p. 5, 17-18, 25
1980b Tanystropheus antiquus (partim) Wild, throughout text; plate 2
1986 Tanystropheus antiquus (partim) Tschanz, p. 84-85, 97
1986 Tanystropheus antiquus Benton, p. 296-297
1987 “Tanystropheus” antiquus (partim) Wild, throughout text
1988 Tanystropheus antiquus (partim) Tschanz, p. 1001-1002, 1008, 1010
1988 Tanystropheus antiquus Evans, p. 227
1994 “Tanystropheus” antiquus (partim) Renesto, p. 298
1997 Tanystropheus antiquus (partim) Benton and Allen, p. 945, 947
2001 Tanystropheus antiquus (partim) Rieppel, p. 273, 276
2005 Tanystropheus antiquus (partim) Dalla Vecchia, p. 41, 43
2005 Tanystropheus antiquus (partim) Renesto, p. 386
2005 “Tanystropheus” antiquus Sennikov, p. 201, 208
2006 Tanystropheus antiquus Fraser and Rieppel, p. 866, 869-870
2007 Tanystropheus antiquus Nosotti, p. 5
2011 Protanystropheus antiquus Sennikov, p.98-99, figures 6-7
2015 Protanystropheus antiquus Pritchard, Turner, Nesbitt, Irmas, and Smith, p. 12, 15
2016 Protanystropheus antiquus and Protanystropheus sp. Surmik, Boczarowski, Balin, Dulski, Szade, Kremer, and Pawlicki, p. 3-6, figure 2
2016 Protanystropheus antiquus Ezcurra, p. 24
2017 Protanystropheus antiquus Jaquier, Fraser, Furrer, and Scheyer, p. 26
2017 Tanystropheus antiquus Skawiński, Ziegler, Czepiński, Szermański, Tałanda, Surmik, and Niedźwiedzki, p. 2, 4-5, 9-13; figure 5h-m
?2017 ‘Thecodontosaurus’ primus Skawiński, Ziegler, Czepiński, Szermański, Tałanda, Surmik, and Niedźwiedzki, p. 2, 4-6, 9-12; figure 5a-c
2019 Protanystropheus Ullmann, Pandya, and Nellermoe, p. 2
2019 Tanystropheus antiquus Spiekman, Winkelhorst, Bleeker, Dorst, De Haan, and Voeten, throughout text
Syntype. SMNS 16687, SMNS 10110, MGUWr 3872s, MGUWr 3888s, MGUWr 3895s, MGUWr 3902s and some uncatalogued MGUWr specimens, all consisting of isolated cervical vertebrae. These specimens were all part of the original material used in the description of the species in Huene (1907-1908) (see also Skawiński et al., 2017). Out of these specimens no lectotype is currently assigned as the material is under revision (Szczygielski, personal commun., 2019).
Localities and occurrence. Lower part of the Gogolin Formation (lowermost Muschelkalk, lower Anisian or possibly uppermost Olenekian) of Gogolin and Krapkowice, Upper Silesia, Poland (Skawiński et al., 2017).
Previous diagnoses. The most recent diagnosis was provided in Sennikov (2011, p. 98) for Protanystropheus antiquus: “Large massive tanystropheid. Neck consisting of nine vertebrae. Cervical vertebrae elongated, their centra 2493 mm long. Length to anterior height ratio of cervical vertebral centra 3.7-6.5. Axis of cervical vertebral centra positioned at 2°-5° to horizontal. Articular surfaces of cervical vertebral centra relatively high, slightly higher than wide. Neural spines of cervical vertebrae low, elongated, in shape of flat crest, most projecting at midlength. Postzygapophyses elongated, terminating in pointed projections projecting posterior to their articular surfaces. Femur sigmoidal.”
The most recent diagnosis referring to Tanystropheus antiquus was provided in Fraser and Rieppel (2006, p. 869): “Protorosaur from the Lower Muschelkalk that has neck vertebrae with central length less than three times the minimum height [...]”.
Tanystropheus antiquus was diagnosed in Wild (1973, p. 152) as follows (translated from German): “Cervical vertebrae distinctly shortened and bearing a less reduced neural spine in comparison to T. conspicuus and T. longobardicus; the zygapophyses always overlapping. The postzygapophyses comparatively more elongated than in T. conspicuus.”
Remarks. Previous systematic palaeontology sections did not consider all the material used in the original description by Huene (1907-1908) (the species name was coined in Huene, 1905), since they thought this material was lost (Wild, 1973; Fraser and Rieppel, 2006; with SMNS 10110 being assigned as the lectotype, and Sennikov, 2011; with SMNS 16687 being assigned as the lectotype for “Protanystropheus” antiquus). However, these specimens that were presumed lost are still in existence, and thus any systematic diagnosis of the taxon should include these (Skawiński et al., 2017). Therefore, assignment of other specimens from the Lower Muschelkalk of Europe similar in morphology to that described by Huene (1907-1908) can only confidently be assigned to the species following a redescription of the original material. Because of this, this section only refers to the original material described by Huene (1907-1908). Since no emended diagnosis based on observations of the original material was provided in Skawiński et al., (2017) and revision of this material is currently ongoing, we refrain from providing a diagnosis for Tanystropheus antiquus. Nevertheless, T. antiquus can likely be preliminarily distinguished from other Tanystropheus species based on the cervical vertebrae having a central length less than three times their minimum height (sensu Fraser and Rieppel, 2006). Material that can likely be assigned to T. antiquus originates from the Lower Muschelkalk of Rüdersdorf near Berlin, Jena, and possibly Bonnhof (all Germany) (Wild, 1980a, 1980b), and the Lower Muschelkalk of Winterswijk, the Netherlands (Wild and Oosterink, 1984; Spiekman et al., 2019), as well as a single cervical vertebra from Dietersweiler near Freudenstadt, Germany (SMNS 56836; Supplementary Table 1). The assignment of the specimens previously identified as “Thecodontosaurus primus” to T. antiquus is uncertain and this material is in need of revision (Skawiński et al., 2017).
Tanystropheus cf. T. antiquus
Referred specimens. See Supplementary Table 1
Localities and occurrence. See Supplementary Table 1
Remarks. The specimens previously identified as Tanystropheus antiquus in Fraser and Rieppel (2006) or Protanystropheus antiquus (Sennikov, 2011) that do not originate from the Gogolin Formation are preliminarily referred to T. cf. T. antiquus here in expectation of a revision of the type material of T. antiquus (see also Skawiński et al., 2017; Spiekman et al., 2019), since these specimens have not been compared to the majority of the material used in the original description in Huene (1905, 1907-1908).
Tanystropheus longobardicus (Bassani 1886) Figure 2, Figure 4
Taxa first described before 1973 that are referred to Tanystropheus longobardicus were also listed in the synonymy list of T. longobardicus of Wild (1973, p. 150-151). Here, an updated synonomy list is provided for both morphotypes of T. longobardicus separately:
Small morphotype
1886 Tribelesodon longobardicus Bassani, p. 25-29
1890 Tribelesodon Zittel, p. 799
1922 Tribelesodon longobardicus von Arthaber, p. 6-7; figure 3
1923 Tribelesodon longobardicus Nopsca, throughout text; text-figures 1-4, 6
1925 Tribelesodon longobardicus Wiman, p. 3
1926 Tribelesodon longobardicus Wiman, throughout text
1930 Tanystropheus longobardicus (partim) Peyer, throughout text
1931 Tanystropheus longobardicus (partim) Peyer, throughout text; text-figures 3-23, 26; plate 1-4, 6-10, 11, figures 1-2, plate 13-14
1934 Tanystropheus longobardicus (partim) Peyer, p. 259-260
1935 Macroscelosaurus longobardicus (partim) Kuhn, p. 119-120
1937 Tanystropheus longobardicus (partim) Peyer, p. 97-105
1939 Tanystropheus longobardicus (partim) Peyer, throughout text
1955 Tanystropheus longobardicus (partim) Peyer and Kuhn-Schnyder, p. 591-604; figures 17, 19, 24-25; plate 2-3
1956 Tanystropheus longobardicus (partim) von Huene, p. 648-650; figure 657
1959 Tanystropheus longobardicus (partim) Kuhn-Schnyder, throughout text; figures 6-9, 14-15
1967 Tanystropheus longobardicus (partim) Kuhn-Schnyder, plate 1
1969 Tribelesodon longobardicus Kuhn, p. 71; figures 21-22
1971 Tanystropheus longobardicus (partim) Kuhn, p. 11; figure 21
1973 Tanystropheus longobardicus (partim) throughout text; text-figures 2-4, 7a, 8, 10a, 11c, 15-16, 20, 24, 26-27, 30, 71a, 81; plate 3, 5-6, 8, 17-18
1975 Tanystropheus longobardicus (partim) Wild, throughout text; figures 2-3, 6a
1975 Tanystropheus longobardicus (partim) Cox, p. 655
1976 Tanystropheus longobardicus (partim) Wild, throughout text; figures 1-2, 6a
1980a Tanystropheus meridensis Wild, p. 5-12; text-figures 1-4, 5a; plate 1-3
1980a Tanystropheus longobardicus (partim) Wild, throughout text; figure 5b; plate 4-5, 6b
1980b Tanystropheus longobardicus (partim) Wild, throughout text
1984 Tanystropheus longobardicus (partim) Wild and Oosterink, p. 146
1985 Tanystropheus longobardicus (partim) Benton, p. 119, 122
1986 Tanystropheus meridensis Tschanz, p. 97
1986 Tanystropheus longobardicus (partim) Tschanz, throughout text; plate 1
1987 Tanystropheus longobardicus (partim) Wild, throughout text
1987 Tanystropheus meridensis Wild, throughout text
1988 Tanystropheus longobardicus (partim) Tschanz, throughout text
1988 Tanystropheus longobardicus (partim; synonymized with T. conspicuus) Evans, p. 227
1989 Tanystropheus longobardicus (partim) Taylor, throughout text
1994 Tanystropheus longobardicus (partim) Renesto, p. 296, 298, 300
1997 Tanystropheus longobardicus (partim) Benton and Allen, p. 945, 947, 950-951
1997 Tanystropheus meridensis Benton and Allen, p. 945, 950
2000 Tanystropheus longobardicus (partim) Dalla Vecchia, p. 137
2001 Tanystropheus longobardicus (partim) Rieppel, throughout text
2001 Tanystropheus meridensis Rieppel, p. 276
2002 Tanystropheus meridensis Dalla Vecchia, p. 228
1003 Tanystropheus longobardicus (partim) Rieppel, Fraser, and Nosotti, p. 272, 276, 278, 2852
2003 Tanystropheus meridensis Rieppel, Fraser, and Nosotti, throughout text
2004 Tanystropheus longobardicus (partim) Fraser, Nosotti, and Rieppel, throughout text
2004 Tanystropheus meridensis Fraser, Nosotti, and Rieppel, throughout text
2005 Tanystropheus longobardicus (partim) Dalla Vecchia, p. 31, 40-43
2005 Tanystropheus meridensis Dalla Vecchia, p. 41-43
2005 Tanystropheus longobardicus (partim) Renesto, p. 386
2005 Tanystropheus meridensis Renesto, p. 386-387
2006 Tanystropheus longobardicus (partim) Fraser and Rieppel, p. 866, 870
2006 Tanystropheus meridensis Fraser and Rieppel, p. 866
2007 Tanystropheus longobardicus (partim) Nosotti, throughout text; figures 1-29, 36-38, 40-41, 43-45, 48-53, 56, 58-60, 62, 64-65; plate 1-4
2007 Tanystropheus meridensis Nosotti, throughout text; figure 39
2008 Tanystropheus longobardicus (partim) Rieppel, Li, and Fraser, p. 95, 99
2010 Tanystropheus longobardicus (partim) Rieppel, Jiang, Fraser, Hao, Motani, Y-L Sun, and Z-Y Sun, throughout text
2010 Tanystropheus meridensis Rieppel, Jiang, Fraser, Hao, Motani, Y-L Sun, and Z-Y Sun, p. 1088
2010 Tanystropheus longobardicus (partim) Stockar, p. 104
2010 Tanystropheus meridensis Stockar, p. 104
2015 Tanystropheus longobardicus (partim) Pritchard, Turner, Nesbitt, Irmas, and Smith, throughout text; figures 14f, 15a
2015 Tanystropheus longobardicus (partim) Nesbitt, Flynn, Pritchard, Parrish, Ranivharimanana, Wyss, throughout text; figure 49d
2016 Tanystropheus longobardicus (partim) Ezcurra, throughout text
2017 Tanystropheus longobardicus (partim) Beardmore and Furrer, throughout text; figure 2
2017 Tanystropheus longobardicus (partim) Jaquier, Fraser, Furrer, and Scheyer, p. 26
2018 Tanystropheus longobardicus (partim) Renesto and Dalla Vecchia, p. 296-297
2018 Tanystropheus longobardicus (partim) Renesto and Saller, throughout text
2018 Tanystropheus meridensis Renesto and Saller, p. 23
Large morphotype
1930 Tanystropheus longobardicus (partim) Peyer, throughout text
1931 Tanystropheus longobardicus (partim) Peyer, p. 65-71, plate 11 figure 3, plate 12
1934 Tanystropheus longobardicus (partim) Peyer, p. 259-260
1935 Macroscelosaurus longobardicus (partim) Kuhn, p. 119-120
1937 Tanystropheus longobardicus (partim) Peyer, p. 97-105
1939 Tanystropheus longobardicus (partim) Peyer, throughout text
1955 Tanystropheus longobardicus (partim) Peyer and Kuhn-Schnyder, p. 591-604; figures 18, 22-23
1956 Tanystropheus longobardicus (partim) von Huene, p. 648-650; figure 657
1959 Tanystropheus longobardicus (partim) Kuhn-Schnyder, throughout text
1967 Tanystropheus longobardicus (partim) Kuhn-Schnyder, text-figure 3
1971 Tanystropheus longobardicus (partim) Kuhn, p. 11; figure 21
1973 Tanystropheus longobardicus (partim) Wild, plates, throughout text; text-figures 1, 5-6, 7b, 9, 10b, 14, 17-19, 21, 23f, 31, 35, 64, 66c, 68, 74, 101; plate 4, 9, 13-16
1975 Tanystropheus longobardicus (partim) Wild, throughout text; figure 6b
1975 Tanystropheus longobardicus (partim) Cox, p. 655
1976 Tanystropheus longobardicus (partim) Wild, throughout text; figures 4, 6b
1980a Tanystropheus longobardicus (partim) Wild, throughout text
1980b Tanystropheus longobardicus (partim) Wild, throughout text
1984 Tanystropheus longobardicus (partim) Wild and Oosterink, p. 146
1985 Tanystropheus longobardicus (partim) Benton, p. 119, 122
1986 Tanystropheus longobardicus (partim) Tschanz, throughout text, plate 2
1987 Tanystropheus longobardicus (partim) Wild, throughout text
1988 Tanystropheus longobardicus (partim) Tschanz, throughout text
1988 Tanystropheus longobardicus (partim; synonymized with T. conspicuus) Evans, p. 227
1989 Tanystropheus longobardicus (partim) Taylor, throughout text
1994 Tanystropheus longobardicus (partim) Renesto, p. 298
1997 Tanystropheus longobardicus (partim) Benton and Allen, p. 945, 947, 950-951
2000 Tanystropheus longobardicus (partim) Dalla Vecchia, p. 137-138
2002 Tanystropheus longobardicus Dalla Vecchia, p. 228
2003 Tanystropheus longobardicus (partim) Rieppel, Fraser, and Nosotti, throughout text
2004 Tanystropheus longobardicus (partim) Fraser, Nosotti, and Rieppel, throughout text
2005 Tanystropheus longobardicus (partim) Dalla Vecchia, p. 31, 40-43
2005 Tanystropheus longobardicus (partim) Renesto, p. 386
2006 Tanystropheus longobardicus (partim) Fraser and Rieppel, p. 866, 870
2007 Tanystropheus longobardicus (partim) Nosotti, throughout text; figures 42, 47, 55
2008 Tanystropheus longobardicus (partim) Rieppel, Li, and Fraser, p. 95, 99
2010 Tanystropheus longobardicus (partim) Rieppel, Jiang, Fraser, Hao, Motani, Y-L Sun, and Z-Y Sun, throughout text; figure 2
2010 Tanystropheus longobardicus (partim) Stockar, p. 104
2015 Tanystropheus longobardicus (partim) Pritchard, Turner, Nesbitt, Irmas, and Smith, throughout text
2015 Tanystropheus longobardicus (partim) Nesbitt, Flynn, Pritchard, Parrish, Ranivharimanana, Wyss, throughout text
2016 Tanystropheus longobardicus (partim) Ezcurra, throughout text; figures 15c, 30
2017 Tanystropheus longobardicus (partim) Beardmore and Furrer, throughout text; figure 1
2017 Tanystropheus longobardicus (partim) Jaquier, Fraser, Furrer, and Scheyer, p. 26
2018 Tanystropheus longobardicus (partim) Renesto and Dalla Vecchia, p. 296-297
2018 Tanystropheus longobardicus (partim) Renesto and Saller, throughout text
Neotype. PIMUZ T 2791 (small morphotype), a strongly compressed and almost completely articulated specimen missing the posterior part of the tail.
Referred specimens. Small morphotype: PIMUZ T 2779, PIMUZ T 2781, PIMUZ T 2795, PIMUZ T 2485, PIMUZ T 2482, PIMUZ T 2484, PIMUZ T 3901, PIMUZ T 1277, MSNM BES SC 265, MSNM BES SC 1018.
Large morphotype: PIMUZ T 2787, PIMUZ T 2793, PIMUZ T 2790, PIMUZ T 2818, PIMUZ T 2819, PIMUZ T 183, SNSB-BSPG 1953 XV 2, MSNM V 3663.
Localities and occurrence. The Besano Formation (Anisian-Ladinian boundary, Middle Triassic) and Meride Limestone (Cassina beds, Ladinian, Middle Triassic) of Monte San Giorgio, the border of Switzerland (canton Ticino) and Italy (Lombardy) (Stockar, 2010).
Previous diagnosis. The most recent diagnosis of Tanystropheus longobardicus was provided in Ezcurra (2016, p. 25). Based on the set of characters treated therein, T. longobardicus was distinguished based on the following features: “frontals flared laterally as wing-like structures above the orbits; large pineal foramen enclosed between frontals and parietals; ventrally flexed anterior end of dentary; strongly posteriorly developed retroarticular process of the lower jaw; conical and straight marginal too[t]h crowns with longitudinal ridges; 13 cervical vertebrae; length of the centra of the fourth and fifth cervical vertebrae at least 14 times their heights; distal end of second sacral rib not bifurcated; two ossified distal carpals; and manual digit IV composed of four phalanges.”
Another important diagnosis was provided in Wild (1973, p. 148), in which Tanystropheus longobardicus was diagnosed based on the following combination of characters (translated from German): “Up to five or possibly six metres long lacertilian with a relatively small and lightly built skull with two temporal openings. Elongate nares positioned close to the midline of the skull. Relatively large orbits. Lacertilian-like palate with infraorbital fenestrae and interpterygoid, posteromedial, and posterolateral openings. Presence of a posttemporal fenestra. A streptostylic quadrate, movable(?) basipterygoid joint, and metakinetic skull. Triradiate jugal. Squamosal similarly shaped as in modern lacertilians. Quadratojugal and most likely postparietal absent. Small rod-shaped supratemporal present. Maxilla with 12 to 14 teeth, which in juveniles are tricuspid in the posterior part of the jaw. Premaxilla with five or six monocuspid teeth. Vomer with 12 monocuspid teeth, palatine and pterygoid only in the juvenile stage with shagreen-like teeth, of which four to six are present on the former. Lower jaw with a ventral flange anteriorly and 17 to 20(?) teeth. Twelve cervical vertebrae, of which nine are extremely elongated. The ninth cervical is generally the longest. Atlas with paired pro-atlases and a separate atlas intercentrum. Eleven dorsal, two lumbar, two sacral, and approximately 46 caudal vertebrae. In the caudal vertebrae, the 13th(?) or 14th provides the possibility of autotomy. Cervical ribs, already occurring at the atlas and axis, are more than double the length of their associated vertebrae, particularly those of the mid-cervical region, and are pseudo-holocephalous. Anterior dorsal ribs dichocephalous, middle and posterior dorsal ribs holocephalous. The capitulum is reduced in the posterior dorsal ribs. Gastral ribs each consisting of four elements, of which the two medial elements are angled at their tip. Forked chevrons positioned intervertebrally and occurring from the first(?) until approximately the 25th caudal vertebra. Pectoral girdle with a broad scapula and a broad coracoid pierced by a coracoid foramen, rhombic and long-stalked interclavicle, and a clasp-shaped clavicle. Short and strong humerus with an entepicondylar groove. Radius stronger and more curved than ulna. Four carpal elements present. Manual phalangeal formula of 2-3-4-4-3. Terminal phalanges formed by claws. Pelvic girdle consisting of a small ilium and wide ischium and pubis. Pubis bearing an obturator foramen. Femur typical lacertilian, sigmoidal. Tibia stronger and more curved than fibula. Astragalus weakly L-shaped and calcaneum disc-shaped with a foramen in between the two elements. Two distal tarsals present. Hooked fifth metatarsal. Foot adapted to swimming with age. Pedal phalangeal formula: 2-3-4-5-4. First phalanx of digit five elongated. Terminal phalanges forming powerful claws.”
Emended diagnosis. Large-sized tanystropheid distinguished by the following combination of characters (autapomorphies among non-archosauriform archosauromorphs marked with an asterisk): 1) presence of tricuspid teeth on the maxilla and dentary (small specimens only); 2) fang-like anterior marginal dentition; 3) mid cervical vertebrae with a maximum length to height ratio larger than 15*; 4) presence of 13 cervical vertebrae*; 5) presence of heterotopic bones in approximately 50% of the known specimens preserving an articulated pelvic region.
Remarks. Based on the findings in this study, the two morphotypes of Tanystropheus longobardicus that have previously been acknowledged (e.g., Wild, 1973; Fraser et al., 2004; Nosotti, 2007), can be clearly distinguished from each other based on traits summarized in Table 2. Although it is unclear whether the two represent different species or distinct ontogenetic stages of the same species, a synonymy list and specimen list is provided for both morphotypes. We are providing these separate lists as we find it important to distinguish between the two morphotypes in future comparative studies, for instance for use in phylogenetic analyses, due to the cranial discrepancy between the two.
Tanystropheus cf. T. longobardicus
Referred specimens. Small morphotype: MCSN 4451, PIMUZ T 2780, PIMUZ T 2789, PIMUZ T 2481, MSNM V 3730.
Large morphotype: PIMUZ T 2817, PIMUZ T 2483, PIMUZ T 2480, PIMUZ T 1270, PIMUZ T 1307, PIMUZ T 2794, PIMUZ T 2788, GMPKU-P-1527, MSNM BES 351.
Localities and occurrence. Small morphotype: The Besano Formation and the Cava Inferiore beds of the Meride Limestone of Monte San Giorgio on the border of Switzerland and Italy (Wild, 1973; Renesto, 2005; Stockar, 2010).
Large morphotype: The Besano Formation of Monte San Giorgio, the border of Switzerland and Italy (Wild 1973; Stockar 2010), and the upper part of the Zhuganpo Member of Falang Formation of Heshangsi of Nimaigu Village, Wusha District, Xingyi City, Guizhou Province, southwestern China.
Remarks. Various articulated specimens of Tanystropheus have been found that are indistinguishable from T. longobardicus but that do not preserve cranial material. Thus, these specimens cannot be assigned to either morphotype. However, for this list we distinguish between these specimens based on their size. If both morphotypes represent two different species, it can of course not be excluded that smaller specimens represent juvenile forms of the large morphotype.
Tanystropheus haasi (Rieppel 2001) Figure 7
1955 Tanystropheus sp. Peyer, p. 488-490; figure 3
1956 Tanystropheus / Tanystrophaeus sp. Brotzen, p. 199, 208
1973 Tanystropheus sp. Wild, p. 152
2001 Tanystropheus haasi Rieppel, throughout text; figures 1-2, 4a
2005 Tanystropheus haasi Dalla Vecchia, p. 41-43
2018 Tanystropheus haasi Renesto and Saller, p. 23
Referred specimens. See Supplementary Table 1
Localities and occurrence. Makhtesh Ramon (Anisian to Ladinian), Negev Desert, Israel
Previous diagnosis. The diagnosis for Tanystropheus haasi was provided in Rieppel (2001): “A species of the genus Tanystropheus which is diagnosed by a deep groove separating the centrum from the neural arch at the posterior end of the vertebrae of the middle cervical region (cv 6 through 8); margins of the postzygapophyseal articular facets distinctly thickened; posterior margin of the postzygapophyseal trough forming a straight transverse line in its middle portion, located above the dorsal margin of the posterior articular surface; neural spine forming a long (one third of the length of the postzgapophyseal process), pointed posterior process which projects into the postzygapophyseal trough.”
Remarks. Tanystropheus haasi is represented by fragmentary isolated cervical vertebrae. Detailed comparison with other isolated cervical vertebrae from Bindlacher Berg assigned to T. conspicuus reveals that most of the characters considered diagnostic for T. haasi (Rieppel, 2001) fall within the range of variation seen within the T. conspicuus material. This highlights that the morphology of the cervical vertebrae is highly variable depending on their relative position in the vertebral column and intraspecific variation and isolated cervical vertebrae are therefore problematic for species assignment within the genus Tanystropheus. Although the referred material of T. haasi possibly differs from the Bindlacher Berg specimens assigned to T. conspicuus in bearing a particularly deepened horizontal groove dorsal to the posterior end of the centrum and the presence of a distinctly expanded posterior end of the neural spine, we do not deem these minor differences from other Tanystropheus taxa sufficient for an assignment to a separate species. Therefore we consider T. haasi a nomen dubium since the referred material is not diagnostic.
Tanystropheus sp.
Referred specimens. See Supplementary Table 1
Localities and occurrence. See Supplementary Table 1
Remarks. These specimens identified as Tanystropheus sp. include fragmented and isolated specimens that do not possess diagnostic features for assignment to any of the known species of the genus.
Sclerostropheus gen. nov. Figure 8
zoobank.org/55B551F2-87D1-4A8C-92E8-5D751A7DF804
1980 Tanystropheus fossai Wild, p. 14-15, figure 11, plate 6e
1987 Tanystropheus fossai Wild, p. 39, 42
1994 Tanystropheus fossai Renesto, p. 296
1997 Tanystropheus fossai Benton and Allen, p. 945, 947
2000 Tanystropheus fossai Dalla Vecchia, p. 139-140
2001 Tanystropheus fossai Rieppel, p. 273, 276
2002 Tanystropheus fossai Dalla Vecchia, p. 228
2005 Tanystropheus fossai Dalla Vecchia, p. 41-43
2005 Tanystropheus fossai Renesto, p. 386
2007 Tanystropheus fossai Nosotti, p. 5
2015 Tanystropheus fossai Pritchard, Turner, Nesbitt, Irmis, and Smith, p. 15-16
2018 Tanystropheus fossai Renesto and Dalla Vecchia, p. 297
2018 Tanystropheus fossai Renesto and Saller, p. 23
Etymology. Sclerostropheus is a combination of the Ancient Greek words skleros, meaning ‘hard’ or ‘hardened’, and stropheus meaning ‘joint’. The name refers to the ossified tendons present in the cervical column of Sclerostropheus and the stiffened cervical column that is typical of tanystropheids due to their strongly elongated vertebrae and ribs.
Diagnosis. Long-necked tanystropheid differentiated from other tanystropheids by the following combination of characters (autapomorphies among non-archosauriform archosauromorphs marked with an asterisk): 1) complete absence of a neural spine in mid-cervical vertebrae*; 2) the presence of two distinct lamina running along each lateral side of the mid-cervical vertebral centra that are most pronounced at the ends of the centrum*; and 3) the presence of an elongate anterior free-ending process of the anterior to mid-cervical ribs, protruding far anteriorly beyond the anterior extent of its corresponding vertebra.
Remarks. In addition to the traits indicated in the diagnosis, MCSNB 4035 also bears bifurcating cervical ribs. However, this likely represents a pathological malformation and it is therefore not included in the diagnosis. Sclerostropheus fossai is currently poorly known from a single specimen constituting a few mid-cervical vertebrae and associated ribs. Nevertheless, considering the known taxonomic diversity of Tanystropheidae, we deem the taxon to be morphologically distinct from other taxa attributed to the genus Tanystropheus and therefore reassign it to a new, separate genus. Together with Langobardisaurus pandolfii and Tanytrachelos ahynis, S. fossai currently represents the third known Late Triassic tanystropheid taxon (Olsen, 1979; Renesto, 1994; Renesto and Dalla Vecchia, 2000, 2007).
Type species. Sclerostropheus fossai (Wild 1980) gen. nov.
Holotype. MCSNB 4035, four articulated cervical vertebrae and associated cervical ribs.
Locality and occurrence. N-slope of Canto Alto near the village of Poscante in Val Brembana, Bergamo, part of the Argillite di Riva di Solto (late Norian, Late Triassic; Rigo et al., 2009; Tackett and Tintori, 2019).
Diagnosis. Same as for the genus.
Tanystropheidae indet.
Referred specimens. GIM L53 Vert. II and BSPG As I 779
Localities and occurrence. See Supplementary Table 1
Remarks. These specimens are considered insufficient for assignment on the genus level but bear characteristics that link them to the tanystropheid clade.
DISCUSSION
Relation to Other Tanystropheid Taxa
In recent years the known diversity of tanystropheids has increased substantially with the discovery of new species assigned to new genera; e.g., Amotosaurus rotfeldensis, Langobardisaurus pandolfii, and Augustaburiania vatagini, Dinocephalosaurus orientalis, and Fuyuansaurus acutirostris (Renesto, 1994; Li, 2003; Fraser and Rieppel, 2006; Sennikov, 2011; Fraser et al., 2013); and new species of previously established genera; e.g., Macrocnemus obristi and Macrocnemus fuyuanensis (Li et al., 2007; Jiang et al., 2011; Fraser and Furrer, 2013; Jaquier et al., 2017); as well as isolated remains that can very likely be attributed to the clade; e.g., the Hayden Quarry tanystropheid material (Pritchard et al., 2015) and the isolated cervical vertebrae from the Sanga do Cabral Formation (De Oliveira et al., 2018), indicating that strongly elongated cervical vertebrae occurred in a more diverse set of early archosauromorphs than previously appreciated. Therefore, the assignment of Tanystropheus antiquus to the genus Tanystropheus should be reconsidered taking specifically into consideration the cervical morphology of other tanystropheid taxa not assigned to the genus Tanystropheus. For instance, the relative elongation of cervical vertebrae attributed to Augustaburiania vatagini is much larger (ratio centrum length/minimum height: 8.79, based on PIN, no. 1043/1392 from Sennikov (2011, figure 3h) than that of T. antiquus (less than three sensu Fraser and Rieppel, 2006). Au. vatagini is also similar to Tanystropheus spp. in the shape and size of the postzygapophyses and the presence of a low neural spine, but differs in having a distinctly curved ventral margin of the centrum. However, our results indicate that morphological characters of isolated cervical vertebrae should be considered carefully before being applied to distinguish taxa, as their morphology varies strongly within a single taxon due to variation throughout the cervical column and intraspecific variation. Much variation occurs in the degree of curvature of the ventral margin of the centrum in the cervical vertebrae attributed to Au. vatagini, which should therefore be evaluated critically in order to verify the assignment of the material to a taxon separate from the known Tanystropheus species (Sennikov, 2011). Am. rotfoldensis, which has cervical vertebrae with a relative elongation close to that seen in T. antiquus, is known from more comprehensive and articulated material and can be distinguished from Tanystropheus species based on various characters, including the total number of cervical vertebrae (which, due to the isolated nature of the material, cannot be confidently established for T. antiquus nor Au. vatagini), the tarsal morphology, and the palatal dentition (Fraser and Rieppel, 2006). Therefore, its assignment to a separate genus is justified. Unfortunately, detailed comparison of the cervical vertebrae of Am. rotfeldensis to those assigned to T. antiquus is problematic, since the known Am. rotfeldensis vertebrae are all strongly flattened, and their poor preservation severely hampers the observation of many characters.
Biogeographical Implications
Our findings show that currently two species of Tanystropheus can be distinguished: T. longobardicus and T. antiquus, with the former being divided into two distinct morphotypes and with “T. conspicuus” and “T. haasi” considered as nomina dubia. Furthermore T. antiquus and its taxonomic affinities are currently under revision (Skawiński et al., 2017). Therefore, the taxonomic diversity of the genus is reduced compared to the six different Tanystropheus species previously acknowledged. However, the two distinct morphotypes of T. longobardicus and the presence of different morphotypes of Tanystropheus at Makhtesh Ramon in Israel provide a good indication that the genus was more speciose, but they can currently not be assigned to a valid species.
The genus Tanystropheus is largely known from the Triassic Tethys coastal regions, with numerous occurrences in the western Tethys region (much of Europe, Saudi Arabia, and Israel), as well as the Germanic basin connecting to the Tethys on its northwestern margin (e.g., Hagdorn and Rieppel, 1999; Feist-Burkhardt et al., 2008), and two known occurrences in the eastern Tethys (southern China) (Figure 1). The possible exception is the only known North American specimen attributed to the genus, YPM VPPU 022000 (Sues and Olsen, 2015). The specimen appears morphologically very similar to the large morphotype of T. longobardicus, as well as to the material attributed to “T. conspicuus” and “T. haasi”, and originates from the Wolfville Formation of Carrs Brook, Nova Scotia, Canada, which is part of the Fundy basin (Sues and Olsen, 2015). This basin was, at least during certain periods in the Triassic, connected to the Germanic basin and Tethys Ocean (Leleu and Hartley, 2010). Therefore, it is reasonable to assume that Tanystropheus spp. had a Tethys-wide occurrence, including extensions into connected shallow marine basins in the northern hemisphere. YPM VPPU 022000 is also of interest as it represents the only known occurrence of the genus in non-marine sediments, since the Economy Member of the Wolfville Formation comprises fluvial sandstones with occasional aeolian dune deposits (Leleu and Hartley, 2010; Sues and Olsen, 2015). This indicates that, at least in the Fundy Basin, the genus also occurred in fresh water lake or river environments. In this regard it is important to consider that, in contrast to the viviparous Dinocephalosaurus orientalis (Liu et al., 2017), it is possible that Tanystropheus spp. had to venture on land for oviposition.
The reassignment of Sclerostropheus fossai from the late Norian of northern Italy to a new genus drastically reduces the temporal distribution of the genus Tanystropheus, which is now mainly known from the Anisian and Ladinian (Middle Triassic), with some specimens also occurring in the earliest Carnian (T. cf. T. longobardicus and Tanystropheus sp. from the Falang Formation, and Tanystropheus sp. from layer E of the Fusea site of northern Italy and possibly the Wolfville Formation), and possibly in the latest Olenekian (T. antiquus).
CONCLUSIONS
We reviewed and revised in detail the material attributed to the genus Tanystropheus, with a focus on the material from Monte San Giorgio and the specimens previously referred to T. conspicuus, T. haasi, and T. fossai. Based on a large disparity in cranial morphology, we find that T. longobardicus can be distinguished into a small and a large morphotype. These morphotypes could represent two different species rather than different ontogenetic stages of the same species as was previously suggested (e.g., Wild, 1973). However, the lack of an overlapping size range between the two morphotypes precludes an unambiguous conclusion based on our morphological comparison and further research including histological sectioning and microtomographic scans are required to establish with certainty whether the two morphotypes represent separate species. PIMUZ T 3901, previously the holotype and only known specimen of T. meridensis, is indistinguishable from the small morphotype of T. longobardicus, and, therefore, T. meridensis is established as a junior synonym of T. longobardicus. Furthermore, we found that the morphology of cervical vertebrae of Tanystropheus spp. exhibits much intraspecific variation, which is in part related to the position of the vertebra in the cervical column. A detailed comparison of cervical vertebrae assigned to T. conspicuus, T. haasi, and the large morphotype of T. longobardicus reveals that insufficient characters separate these specimens morphologically to merit assignment to a separate species. Because the material assigned to T. conspicuus and T. haasi is (largely) represented by (partial) cervical vertebrae, we consider these two species to represent nomina dubia. MCSNB 4035, the only known specimen previously assigned to T. fossai, is considered to differ distinctly from other Tanystropheus species and is therefore reassigned to a new genus: Sclerostropheus.
The known diversity of the genus Tanystropheus is reduced from six to two species. Nevertheless, the genus was likely more speciose based on fragmentary material that cannot be diagnosed confidently currently, as well as the presence of two morphotypes of T. longobardicus that might represent separate species rather than an ontogenetic series of a single species.
These findings result in new biogeographical considerations, with the genus largely being restricted temporally between the latest Olenekian or earliest Anisian and the earliest Carnian (latest Early Triassic to earliest Late Triassic), and extending spatially between the eastern and western Tethys, as well as the Germanic and Fundy basins (Figure 1). Furthermore, our study highlights the difficulty of differentiating closely related tetrapod species based on isolated material. This is particularly the case for vertebrae, as these elements are shown to exhibit much intraspecific variation, in part related to the relative position of each element in the vertebral column.
ACKNOWLEDGEMENTS
The authors would like to thank the following curators and collection managers for access to specimens: C. Klug and B. Scheffold (PIMUZ), J. Rabold and S. Eggmaier (U-MO), D-Y. Jiang (GMPKU), R. Schoch and E. Maxwell (SMNS), A. Aiello, M. Valle, and S. Perini (MCSNB), C. Dal Sasso and S. Nosotti (MSNM), and R. Stockar (MCSN). Furthermore we would like to thank D. Schwarz (MB.R/MfN), H. Hagdorn (MHI), R. Brocke and G. Riedel (SMF), I. Werneburg (GPIT), and U. Göhlich (NHMW) for information regarding Tanystropheus specimens in the collections under their care (abbreviations of certain institutions can be found in Supplementary Table 1). We would like to thank N. Fraser, O. Rieppel, S. Nosotti, F. Dalla Vecchia, and T. Szczygielski for discussions. S. Schenk (PIMUZ) is thanked for help in obtaining literature. Two reviewers and the handling editor are thanked for their constructive recommendations to the initial version of the manuscript. M. Woodert (University of Zurich) performed a pilot study on measuring the variation in the Tanystropheus specimens from Monte San Giorgio. The study was funded by SNSF (grant no. 2’5321-162775 to TMS).
REFERENCES
Adam, K.D. 1953. Ein Tanystropheus-Fund aus dem Hauptmuschelkalk bei Schloss Steffen (Kreis Künzelsau). Neues Jahrbuch für Geologie und Paläontologie Monatshefte, 1:40-43.
Arthaber, G.v. 1922. Über Entwicklung, Ausbildung und Absterben der Flugsaurier. Paläontologische Zeitschrift, 4(1):1-47.
Bassani, F. 1886. Sui fossili e sull’eta degli schisti bituminosi triasici di Besano in Lombardia. Atti Società Italiana di scienze naturali, 29:15-72.
Benton, M.J. 1985. Classification and phylogeny of the diapsid reptiles. Zoological Journal of the Linnean Society, 84:97-164.
Benton, M.J. 1986. The Late Triassic reptile Teratosaurus-a rauisuchian, not a dinosaur. Palaeontology, 29(2):293-301.
Benton, M.J. and Allen, J.L. 1997. Boreopricea from the Lower Triassic of Russia, and the relationships of the prolacertiform reptiles. Palaeontology, 40(4):931-953.
Broili, F. 1915. Beobachtungen an Tanystropheus conspicuus H. v. Meyer. Neues Jahrbuch für Mineralogie-Abhandlungen, 2:51-62.
Brotzen, F. 1956. Stratigraphical Studies on the Triassic Vertebrate Fossils from Wadi Raman, Israel. Arkiv för mineralogi och geologi; Bd. 2, nr. 9. Almqvist & Wiksell, Stockholm.
Camp, C. L. 1945a. Prolacerta and the protorosaurian reptiles. Part I. American Journal of Science 243(1):17-32. https://doi.org/10.2475/ajs.243.1.17
Camp, C. L. 1945b. Prolacerta and the protorosaurian reptiles. Part II. American Journal of Science 243(2):84-101.
Chatterjee, S. 1986. Malerisaurus langstoni, a new diapsid reptile from the Triassic of Texas. Journal of Vertebrate Paleontology, 6(4):297-312.
Colbert, E.H. 1970. A saurischian dinosaur from the Triassic of Brazil. American Museum Novitates, 2405:1-39.
Cope, E.D. 1887. A contribution to the history of the Vertebrata of the Trias of North America. Proceedings of the American Philosophical Society, 24(126):209-228.
Cope, E.D. 1889. On a new genus of Triassic Dinosauria. American Naturalist, 23(271):626.
Corroy, G. 1928. Les Vertébrés du Trias de Lorraine et le Trias Lorrain. Annales de Paléontologie, 17:81-136.
Cox, B. 1975. The longest-necked lizard? Nature, 254(5502):654.
Dalla Vecchia, F.M. 2000. Tanystropheus (Archosauromorpha, Prolacertiformes) remains from the Triassic of the Northern Friuli (NE Italy). Rivista Italiana di Paleontologia e Stratigrafia, 106(2):135-140.
Dalla Vecchia, F.M. 2005. Resti di Tanystropheus, Saurotterigi e "Rauisuchi" (Reptilia) nel Triassico medio della Val Aupa (Moggio Udinese, Udine). Gortania-Atti del Museo Friulano di Storia Naturale, 27:25-48.
Dalla Vecchia, F.M. and Avanzini, M. 2002. New findings of isolated remains of Triassic reptiles from Northeastern Italy. Bolletino della Società Paleontologica Italiana, 41(2/3):215-235.
De Oliveira, T.M., Oliveira, D., Schultz, C.L., Kerber, L., and Pinheiro, F.L. 2018. Tanystropheid archosauromorphs in the Lower Triassic of Gondwana. Acta Palaeontologica Polonica, 63(4):713-723. https://doi.org/10.4202/app.00489.2018
Diedrich, C. 2012. The Middle Triassic marine reptile biodiversity in the Germanic Basin, in the centre of the Pangaean world. Open Geosciences, 4(1):9-46. https://doi.org/10.2478/s13533-011-0060-0
Dilkes, D.W. 1998. The Early Triassic rhynchosaur Mesosuchus browni and the interrelationships of basal archosauromorph reptiles. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 353:501-541. https://doi.org/10.1098/rstb.1998.0225
Dufeau, D.L. and Witmer, L.M. 2015. Ontogeny of the middle-ear air-sinus system in Alligator mississippiensis (Archosauria: Crocodylia). PLoS One, 10(9):e0137060. https://doi.org/10.1371/journal.pone.0137060
Edinger, T. 1924. Rückenmark im Wirbelkörper! Anatomischer Anzeiger, 57:515-519.
Eichenberger, U. 1986. Die Mitteltrias der Silvretta-Decke (Ducankette und Landwassertal, Ostalpin). Mitteilungen aus dem Geologischen Institut der Eidg. Technischen Hochschule und der Universität Zürich-Neue Folge, 252.
Evans, S.E. 1988. The early history and relationships of the Diapsida, p. 221-260. In Benton, M.J. (ed.), The Phylogeny and Classification of the Tetrapods, Volume 1: Amphibians, Reptiles, Birds. Clarendon Press, Oxford.
Ezcurra, M.D. 2016. The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms. PeerJ, 4:e1778. https://doi.org/10.7717/peerj.1778
Feist-Burkhardt, S., Götz, A.E., Szulc, J., Borkhataria, R., Geluk, M., Haas, J., Hornung, J., Jordan, P., Kempf, O., and Michalik, J. 2008. Triassic, p. 749-821. In McCan, T. (ed.), The Geology of Central Europe. Geological Society of London, London.
Fraas, E. 1896. Die Schwäbischen Trias-Saurier nach dem Material der Königlichen Naturalien-Sammlung in Stuttgart zusammengestellt. Schweizerbart (E. Koch), Stuttgart
Fraser, N.C. and Furrer, H. 2013. A new species of Macrocnemus from the Middle Triassic of the eastern Swiss Alps. Swiss Journal of Geosciences, 106:199-206. https://doi.org/10.1007/s00015-013-0137-5
Fraser, N.C., Nosotti, S., and Rieppel, O. 2004. A re-evaluation of two species of Tanystropheus (Diapsida, Protorosauria), from Monte San Giorgio, southern Alps. Journal of Vertebrate Paleontology, 24(3):60A.
Fraser, N.C. and Rieppel, O. 2006. A new protorosaur (Diapsida) from the Upper Buntsandstein of the Black Forest, Germany. Journal of Vertebrate Paleontology, 26(4):866-871. https://doi.org/10.1671/0272-4634(2006)26[866:anpdft]2.0.co;2
Fraser, N.C., Rieppel, O., and Li, C. 2013. A long-snouted Protorosaur from the Middle Triassic of southern China. Journal of Vertebrate Paleontology, 33(5):1120-1126. https://doi.org/10.1080/02724634.2013.764310
Gervais, P. 1859. Zoologie et paléontologie françaises: nouvelles recherches sur les animaux vertébrés dont on trouve les ossements enfouis dans le sol de la France et sur leur comparaison avec les espèces propres aux autres régions du globe. A. Bertrand.
Gow, C.E. 1975. The morphology and relationships of Youngina capensis Broom and Prolacerta broomi Parrington. Palaeontologica Africana, 18:89-131.
Hagdorn, H. and Rieppel, O. 1999. Stratigraphy of marine reptiles in the Triassic of Central Europe. Zentralblatt für Geologie und Paläontologie, Teil I, 1998:651-678.
Huene, F.R.v. 1902. Übersicht über die Reptilien der Trias. Geologische und Paläontologische Abhandlungen (Neue Serie), 6:1-84.
Huene, F.R.v. 1905. Über die Trias-Dinosaurier Europas. Zeitschrift der Deutschen Geologischen Gesellschaft, 57:345-349.
Huene, F.R.v. 1907-1908. Die Dinosaurier der europäischen Triasformation. Geologische und Palaeontologische Abhandlungen, Supplement 1:1-419.
Huene, F.R.v. 1931. Über Tanystropheus und verwandte Formen. Neues Jahrbuch für Geologie und Paläontologie, BB, 67:65-86.
Huene, F.R.v. 1932. Die Fossile Reptil-Ordnung Saurischia: Ihre Entwicklung und Geschichte Teil I und II. Monographien zur Geologie und Palaeontologie, Serie 1 Heft 4. Gebrüder Borntraeger, Leipzig.
Huene, F.R.v. 1944. Beiträge zur Kenntnis der Protorosaurier. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Monatshefte, Abteilung B, 5:120-131.
Huene, F.R.v. 1946. Die grossen Stämme der Tetrapoden in den geologischen Zeiten. Biologisches Zentralblatt, 65:268-275.
Huene, F.R.v. 1956. Paläontologie und Phylogenie der niederen Tetrapoden. Fischer, Jena.
Jalil, N.-E. 1997. A new prolacertiform diapsid from the Triassic of North Africa and the interrelationships of the Prolacertiformes. Journal of Vertebrate Paleontology, 17(3):506-525. https://doi.org/10.1080/02724634.1997.10010998
Jaquier, V.P., Fraser, N.C., Furrer, H., and Scheyer, T.M. 2017. Osteology of a new specimen of Macrocnemus aff. M. fuyuanensis (Archosauromorpha, Protorosauria) from the Middle Triassic of Europe: potential implications for species recognition and paleogeography of tanystropheid protorosaurs. Frontiers in Earth Science, 5(91). https://doi.org/10.3389/feart.2017.00091
Jiang, D.-Y., Motani, R., Hao, W.-C., Rieppel, O., Sun, Y.-L., Schmitz, L., and Sun, Z.-Y. 2008. First record of Placodontoidea (Reptilia, Sauropterygia, Placodontia) from the eastern Tethys. Journal of Vertebrate Paleontology, 28(3):904-908. https://doi.org/10.1671/0272-4634(2008)28[904:froprs]2.0.co;2
Jiang, D.-Y., Rieppel, O., Fraser, N.C., Motani, R., Hao, W.-C., Tintori, A., Sun, Y.-L., and Sun, Z.-Y. 2011. New information on the protorosaurian reptile Macrocnemus fuyuanensis Li et al., 2007, from the Middle/Upper Triassic of Yunnan, China. Journal of Vertebrate Paleontology, 31(6):1230-1237. https://doi.org/10.1080/02724634.2011.610853
Jiang, D.-Y., Schmitz, L., Hao, W.-C., and Sun, Y.-L. 2006. A new mixosaurid ichthyosaur from the Middle Triassic of China. Journal of Vertebrate Paleontology, 26(1):60-69. https://doi.org/10.1671/0272-4634(2006)26[60:anmift]2.0.co;2
Jurcsák, T. 1975. Tanystropheus biharicus n. sp. (Reptilia, Squamata) o nouă specie pentru fauna Triasică a României. Nymphaea, 3:45-52.
Jurcsák, T. 1976. Noi descoperini de reptile fossile in Triasicul de la Aleșd. Nymphaea, 4:67-105.
Jurcsák, T. 1978. Rezultate noi in studiul saurienilor fosili de la Aleșd. Nymphaea, 6:15-60.
Jurcsák, T. 1982. Occurrences nouvelles des sauriens mésozoïques de Roumanie. Vertebrata Hungarica, 21:175-184.
Kuhn-Schnyder, E. 1959. Hand und Fuss von Tanystropheus longobardicus (Bassani). Eclogae Geologicae Helvetiae, 52(2):921-941.
Kuhn-Schnyder, E. 1967. Das Problem der Euryapsida. Éditions du Centre national de la recherche scientifique, 163:335-348.
Kuhn, O. 1935. Sauropterygia. Serie: Animalia, pars 69. Wilhelm Junk, 's-Gravenhage (The Hague).
Kuhn, O. 1969. Handbuch der Paläoherpetologie, 9. Fischer, Stuttgart.
Kuhn, O. 1971. Die Saurier der Deutschen Trias. Geiselberger, Altötting.
Leleu, S. and Hartley, A.J. 2010. Controls on the stratigraphic development of the Triassic Fundy Basin, Nova Scotia: implications for the tectonostratigraphic evolution of Triassic Atlantic rift basins. Journal of the Geological Society, 167(3):437-454. https://doi.org/10.1144/0016-76492009-092
Li, C. 2003. First record of Protorosaurid reptile (Order Protorosauria) from the Middle Triassic of China. Acta Geologica Sinica, 77(4):419-423. https://doi.org/10.1111/j.1755-6724.2003.tb00122.x
Li, C. 2007. A juvenile Tanystropheus sp. (Protorosauria, Tanystropheidae) from the Middle Triassic of Guizhou, China. Vertebrata PalAsiatica, 45(1):41.
Li, C., Fraser, N.C., Rieppel, O., Zhao, L.-J., and Wang, L.-T. 2017. A new diapsid from the Middle Triassic of southern China. Journal of Paleontology, 91:1-7. https://doi.org/10.1017/jpa.2017.12
Li, C., Rieppel, O., and LaBarbera, M.C. 2004. A Triassic aquatic protorosaur with an extremely long neck. Science, 305:1931. https://doi.org/10.1126/science.1100498
Li, C., Zhao, L.-J., and Wang, L.-T. 2007. A new species of Macrocnemus (Reptilia: Protorosauria) from the Middle Triassic of southwestern China and its palaeogeographical implication. Science in China Series D: Earth Sciences, 50(11):1601-1605. https://doi.org/10.1007/s11430-007-0118-5
Li, J. 2006. A brief summary of the Triassic marine reptiles of China. Vertebrata PalAsiatica, 44(1):99-108.
Liu, J., Organ, C.L., Benton, M.J., Brandley, M.C., and Aitchison, J.C. 2017. Live birth in an archosauromorph reptile. Nature Communications, 8:14445. https://doi.org/10.1038/ncomms14445
Mahler, D.L. and Kearney, M. 2006. The palatal dentition in squamate reptiles: morphology, development, attachment, and replacement. Fieldiana Zoology, 108:1-61. https://doi.org/10.5962/bhl.title.3855
Melville, R.V. 1981. Opinion 1186 Tanystropheus H. von Meyer, [1852] (Reptilia) conserved. Bulletin of Zoological Nomenclature, 38:188-190.
Menning , M. and Hendrich, A. 2016. Stratigraphische Tabelle von Deutschland 2016, downloaded 16 January 2019. Deutsche Stratigraphische Kommission, GeoForschungsZentrum, Potsdam. http://www.stratigraphie.de/std/Bilder/5_2.pdf
Meyer, H.v. 1855. Die Saurier des Muschelkalkes mit Rücksicht auf die Saurier aus buntem Sandstein und Keuper, p. 1-167. In Keller, H. (ed.), Zur Fauna der Vorwelt, zweite Abtheilung. Heinrich Keller, Frankfurt a.m.
Njau, J.K. and Blumenschine, R.J. 2006. A diagnosis of crocodile feeding traces on larger mammal bone, with fossil examples from the Plio-Pleistocene Olduvai Basin, Tanzania. Journal of Human Evolution, 50(2):142-162. https://doi.org/10.1016/j.jhevol.2005.08.008
Nopsca, F.B. 1923. Neubeschreibung des Trias-Pterosauriers Tribelesodon. Paläontologische Zeitschrift, 5:161-181.
Nosotti, S. 2007. Tanystropheus longobardicus (Reptilia, Protorosauria): re-interpretations of the anatomy based on new specimens from the Middle Triassic of Besano (Lombardy, northern Italy). Memorie della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano, 35:1-88.
Olsen, P.E. 1979. A new aquatic eosuchian from the Newark Supergroup (Late Triassic-Early Jurassic) of North Carolina and Virginia. Postilla, 176:1-14.
Organ, C.L. and Adams, J. 2005. The histology of ossified tendon in dinosaurs. Journal of Vertebrate Paleontology, 25(3):602-613. https://doi.org/10.1671/0272-4634(2005)025[0602:thooti]2.0.co;2
Ortlam, D. 1966. Fossile Böden als Leithorizonte für die Gliederung des Höheren Buntsandsteins im nördlichen Schwarzwald und südlichen Odenwald. Geologisches Jahrbuch, 84:485-590.
Osborn, H.F. 1903. The reptilian subclasses Diapsida and Synapsida and the early history of the Diaptosauria. Memoirs of the American Museum of Natural History, 1(8):449-519.
Peyer, B. 1930. Tanystropheus longobardicus Bass. sp. (Vorläufige Mitteilung). Cbl. Miner. usw:336-337.
Peyer, B. 1931. Die Triasfauna der Tessiner Kalkalpen II. Tanystropheus longobardicus Bass. sp. Abhandlungen der Schweizerischen Paläontologischen Gesellschaft, 50:9-110.
Peyer, B. 1934. Zehn Jahre Tessiner Fossiliengrabung des Zoologischen Museums der Universität Zürich. Verhandlungen der Schweizerischen Naturforschenden Gesellschaft:257-261.
Peyer, B. 1937. Die Triasfauna der Tessiner Kalkalpen XII. Macrocnemus bassanii Nopsca. Abhandlungen der Schweizerischen Paläontologischen Gesellschaft, 54:1-87.
Peyer, B. 1939. Über die Rekonstruktion des Skelettes von Tanystropheus. Eclogae Geologicae Helvetiae, 32(2):203-209.
Peyer, B. 1955. Demonstration von Trias-Vertebraten aus Palaestina. Eclogae Geologicae Helvetiae, 48:486-490.
Peyer, B. and Kuhn-Schnyder, E. 1955. Squamates du Trias, p. 578-605. In Piveteau, J. (ed.), Traité de Paléontologie. Masson, Paris.
Plieninger, T. 1846. Über ein neues Sauriergenus und die Einreihung der Saurier mit flachen, schneidenden Zähnen in eine Familie. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg, 2(1):148-154.
Pritchard, A.C., Turner, A.H., Nesbitt, S.J., Irmis, R.B., and Smith, N.D. 2015. Late Triassic tanystropheids (Reptilia, Archosauromorpha) from northern New Mexico (Petrified Forest Member, Chinle Formation) and the biogeography, functional morphology, and evolution of Tanystropheidae. Journal of Vertebrate Paleontology, 35:e911186. https://doi.org/10.1080/02724634.2014.911186
Rauhut, O.W. and Hungerbühler, A. 2000. A review of European Triassic theropods. Gaia, 15:75-88.
Renesto, S. 1994. A new prolacertiform reptile from the Late Triassic of Northern Italy. Rivista Italiana di Paleontologia e Stratigrafia, 100(2):285-306.
Renesto, S. 2005. A new specimen of Tanystropheus (Reptilia Protorosauria) from the Middle Triassic of Switzerland and the ecology of the genus. Rivista Italiana di Paleontologia e Stratigrafia, 111(3):377-394.
Renesto, S. and Dalla Vecchia, F.M. 2000. The unusual dentition and feeding habits of the prolacertiform reptile Langobardisaurus (Late Triassic, Northern Italy). Journal of Vertebrate Paleontology, 20(3):622-627. https://doi.org/10.1671/0272-4634(2000)020[0622:tudafh]2.0.co;2
Renesto, S. and Dalla Vecchia, F.M. 2007. A revision of Langobardisaurus rossii Bizzarini and Muscio, 1995 from the Late Triassic of Friuli (Italy). Rivista Italiana di Paleontologia e Stratigrafia, 113(2):191-201.
Renesto, S. and Dalla Vecchia, F.M. 2018. Late Triassic marine reptiles, p. 263-313. In L. Tanner (eds.), The Late Triassic World. Springer. https://doi.org/10.1007/978-3-319-68009-5_8
Renesto, S. and Saller, F. 2018. Evidences for a semi aquatic life style in the Triassic diapsid reptile Tanystropheus. Rivista Italiana di Paleontologia e Stratigrafia, 124(1):23-34.
Rieppel, O. 2001. A new species of Tanystropheus (Reptilia: Protorosauria) from the Middle Triassic of Makhtesh Ramon, Israel. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 221(2):271-287.
Rieppel, O., Fraser, N.C., and Nosotti, S. 2003. The monophyly of Protorosauria (Reptilia, Archosauromorpha): a preliminary analysis. Atti della Società italiana di scienze naturali e del Museo civico di storia naturale di Milano, 144(II):359-382.
Rieppel, O., Jiang, D.-Y., Fraser, N.C., Hao, W.-C., Motani, R., Sun, Y.-L., and Sun, Z.-Y. 2010. Tanystropheus cf. T. longobardicus from the early Late Triassic of Guizhou Province, southwestern China. Journal of Vertebrate Paleontology, 30(4):1082-1089. https://doi.org/10.1080/02724634.2010.483548
Rieppel, O., Li, C., and Fraser, N.C. 2008. The skeletal anatomy of the Triassic protorosaur Dinocephalosaurus orientalis, from the Middle Triassic of Guizhou Province, southern China. Journal of Vertebrate Paleontology, 28(1):95-110. https://doi.org/10.1671/0272-4634(2008)28[95:tsaott]2.0.co;2
Rieppel, O., Mazin, J.-M., and Tchernov, E. 1997. Speciation along rifting continental margins: a new nothosaur from the Negev (Israel). Comptes Rendus de l'Académie des Sciences-Series IIA-Earth and Planetary Science, 325(12):991-997. https://doi.org/10.1016/s1251-8050(97)82380-4
Rigo, M., Galli, M., and Jadoul, F. 2009. Late Triassic biostratigraphic constraints in the Imagna Valley (western Bergamasc Alps, Italy). Albertiana, 37:39-42.
Rühle von Lilienstern, H. 1952. Die Saurier Thüringens. Fischer, Jena.
Schmidt, M. 1928. Die Lebewelt unserer Trias. Hohenlohe'sche Buchhandlung, Öhringen.
Schmidt, M. 1938. Die Lebewelt unserer Trias (Nachtrag). Hohenlohe'sche Buchhandlung, Öhringen.
Schoch, R.R. 2011. New archosauriform remains from the German Lower Keuper. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 260(1):87-100. https://doi.org/10.1127/0077-7749/2011/0133
Sennikov, A.G. 2005. A new specialized prolacertilian (Reptilia: Archosauromorpha) from the Lower Triassic of the Orenburg Region. Paleontological Journal, 39(2):199-209.
Sennikov, A.G. 2011. New tanystropheids (Reptilia: Archosauromorpha) from the Triassic of Europe. Paleontological Journal, 45(1):90-104. https://doi.org/10.1134/s0031030111010151
Simpson, G.G. 1951. The species concept. Evolution, 5(4):285-298.
Skawiński, T., Ziegler, M., Czepiński, Ł., Szermański, M., Tałanda, M., Surmik, D., and Niedźwiedzki, G. 2017. A re-evaluation of the historical ‘dinosaur’ remains from the Middle-Upper Triassic of Poland. Historical Biology, 29(4):442-472. https://doi.org/10.1080/08912963.2016.1188385
Smith, A.G., Smith, D.G., and Funnell, B.M. 1994. Atlas of Mesozoic and Cenozoic coastlines. Cambridge University Press, New York.
Spiekman, S.N.F. 2018. A new specimen of Prolacerta broomi from the lower Fremouw Formation (Early Triassic) of Antarctica, its biogeographical implications and a taxonomic revision. Scientific reports, 8(1):17996. https://doi.org/10.1038/s41598-018-36499-6
Spiekman, S.N.F., Winkelhorst, H., Bleeker, R., Dorst, M., de Haan, R., and Voeten, D.F. 2019. Tanystropheids from the Winterswijkse Steengroeve – rare but recurring elements in the Vossenveld paleoassemblage. Grondboor & Hamer (Staringia 16), 73(5/6): 208-215.
Stockar, R. 2010. Facies, depositional environment, and palaeoecology of the Middle Triassic Cassina beds (Meride Limestone, Monte San Giorgio, Switzerland). Swiss Journal of Geosciences, 103:101-119. https://doi.org/10.1007/s00015-010-0008-2
Sues, H.-D. and Olsen, P.E. 2015. Stratigraphic and temporal context and faunal diversity of Permian-Jurassic continental tetrapod assemblages from the Fundy rift basin, eastern Canada. Atlantic Geology, 51(1):139-205. https://doi.org/10.4138/atlgeol.2015.006
Surmik, D., Boczarowski, A., Balin, K., Dulski, M., Szade, J., Kremer, B., and Pawlicki, R. 2016. Spectroscopic studies on organic matter from Triassic reptile bones, Upper Silesia, Poland. PLoS One, 11(3). https://doi.org/10.1371/journal.pone.0151143
Tackett, L.S. and Tintori, A. 2019. Low drilling frequency in Norian benthic assemblages from the southern Italian Alps and the role of specialized durophages during the Late Triassic. Palaeogeography, Palaeoclimatology, Palaeoecology, 513:25-34. https://doi.org/10.1016/j.palaeo.2018.06.034
Taylor, M.A. 1989. Neck and neck. Nature, 341:668-689. https://doi.org/10.1038/341688a0
Tschanz, K. 1986. Funktionelle Anatomie der Halswirbelsäule von Tanystropheus longobardicus (Bassani) aus der Trias (Anis/Ladin) des Monte San Giorgio (Tessin) auf der Basis vergleichend morphologischer Untersuchungen an der Halsmuskulatur rezenter Echsen. PhD thesis, University of Zurich, Zurich, Switzerland.
Tschanz, K. 1988. Allometry and Heterochrony in the growth of the neck of Triassic Prolacertiform reptiles. Palaeontology, 31(4):997-1011.
Ullmann, P.V., Pandya, S.H., and Nellermoe, R. 2019. Patterns of soft tissue and cellular preservation in relation to fossil bone tissue structure and overburden depth at the Standing Rock Hadrosaur Site, Maastrichtian Hell Creek Formation, South Dakota, USA. Cretaceous Research, 99:1-13. https://doi.org/10.1016/j.cretres.2019.02.012
Vickers-Rich, P., Rich, T., Rieppel, O., Thulborn, R.A., and McClure, H.A. 1999. A Middle Triassic vertebrate fauna from the Jilh Formation, Saudi Arabia. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 213(2):201-232. https://doi.org/10.1127/njgpa/213/1999/201
Wang, W., Li, C., Scheyer, T.M., and Zhao, L. 2019. A new species of Cyamodus (Placodontia, Sauropterygia) from the early Late Triassic of south-west China. Journal of Systematic Palaeontology, 17:1457-1476. https://doi.org/10.1080/14772019.2018.1535455
Werneburg, I., Polachowski, K.M., and Hutchinson, M.N. 2015. Bony skull development in the Argus monitor (Squamata, Varanidae, Varanus panoptes) with comments on developmental timing and adult anatomy. Zoology, 118(4):255-280. https://doi.org/10.1016/j.zool.2015.02.004
Wild, R. 1973. Die Triasfauna der Tessiner Kalkalpen XXII. Tanystropheus longobardicus (Bassani) (Neue Ergebnisse). Schweizerische Paläontologische Abhandlungen, 95.
Wild, R. 1975. Der Giraffenhals-Saurier. Die Naturwissenschaften, 62(4):149-153.
Wild, R. 1976. Neues über den Giraffenhalssaurier Tanystropheus. Natur und Museum, 106(1):13-22.
Wild, R. 1980a. Neue Funde von Tanystropheus (Reptilia, Squamata). Schweizerische Palaontologische Abhandlungen, 102.
Wild, R. 1980b. Tanystropheus (Reptilia: Squamata) and its importance for stratigraphy. Mémoires de la Société Géologique de France, n. s., 139:201-206.
Wild, R. 1987. An example of biological reasons for extinction: Tanystropheus (Reptilia, Squamata). Mémoires de la Société Géologique de France, n. s., 150:37-44.
Wild, R. and Oosterink, H. 1984. Tanystropheus (Reptilia: Squamata) aus dem Unteren Muschelkalk von Winterswijk, Holland. Grondboor en Hamer, 5:142-148.
Wiman, C. 1925. Über Dorygnathus und andere Flugsaurier. Bulletin of the Geological Institute of Uppsala, 19:23-55.
Wiman, C. 1926. Über einige Flugsaurier. Paläontologische Zeitschrift, 7(1):15-22.
Zittel, K.A. 1890. Handbuch der Palaeontologie. I. Abteilung Palaeozoologie. III Band Vertebrata. R. Oldenbourg, Munich and Leipzig.
