

 The biogeography and ecology of the Cretaceous non-avian dinosaurs of Appalachia
The biogeography and ecology of the Cretaceous non-avian dinosaurs of Appalachia
Article number: 21.1.5A
https://doi.org/10.26879/801
Copyright Society for Vertebrate Paleontology, February 2018
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 5 July 2017. Acceptance: 17 January 2018
{flike id=2123}
ABSTRACT
The Cenomanian to Maastrichtian of the Late Cretaceous saw the flooding of the interior of North America by the Western Interior Seaway, which created the eastern landmass of Appalachia and the western landmass of Laramidia. Though Appalachian dinosaur faunas are poorly known, they are nevertheless important for understanding Cretaceous dinosaur paleobiogeography and ecology. In order to better track the vicariance of eastern and western North American dinosaur faunas over the duration of the Cretaceous, the former were compared with the latter from the Aptian to Maastrichtian Stages of the Late Cretaceous using several similarity indices. The data gathered from biogeographic similarity indices suggest that an almost completely homogenous North American dinosaur fauna found in the Early Cretaceous experienced significant vicariance, splitting into a Laramidian fauna differentiated by the presence of ceratopsids, pachycephalosaurids, saurolophids, lambeosaurines, ankylosaurids, therizinosaurids, and troodontids and an Appalachian fauna characterized by the lack of the aforementioned groups and the presence of non-hadrosaurid hadrosauroids, massive hadrosauroids, basal hadrosaurids, leptoceratopsians, “intermediate”-grade tyrannosauroids, and nodosaurids between the Cenomanian and Campanian, with these two faunas later experiencing limited dispersal after the disappearance of the Western Interior Seaway from the American Interior during the Maastrichtian. Dinosaur provincialism and ecology on Appalachia are also investigated and discussed. Though the fossil record of dinosaurs for parts of the Cretaceous is poor throughout North America and in the eastern portion of the continent especially, the analyses herein nevertheless allow for a greater glimpse at dinosaur biogeography and ecology in Appalachia and in North America generally during the time.
Chase D. Brownstein. Research Associate, Stamford Museum & Nature Center, Stamford, CT. USA. chasethedinosaur@gmail.com
Keywords: paleobiogeography; paleoecology; Appalachia; Cretaceous; Dinosauria
Final citation: Brownstein, Chase D. 2018. The biogeography and ecology of the Cretaceous non-avian dinosaurs of Appalachia. Palaeontologia Electronica 21.1.5A 1-56. https://doi.org/10.26879/801
palaeo-electronica.org/content/2018/2123-appalachia-biogeography
Copyright: February 2018 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0/
INTRODUCTION
During the Albian to Cenomanian stages of the Late Cretaceous, the interior of North America was flooded by a shallow sea called the Western Interior Seaway (e.g., Russell, 1995; Roberts and Kirschbaum, 1995). The creation of the seaway caused the formation of a long, slender landmass known as Laramidia to the west and the wider, more rectangular Appalachia to the east (e.g., Schwimmer, 2002; Sampson et al., 2010), having appreciable consequences in the evolution of North American dinosaurs. The former of these two landmasses apparently experienced a heightened level of non-avian dinosaur diversification during the Campanian (Sampson et al., 2010; Loewen et al., 2013), when a high level of local dinosaur endemism partnered with regional differences in non-avian dinosaur faunas may have occurred there (Lehman, 1997; Sampson et al., 2010; Loewen et al., 2013). The cause for this rapid diversification of non-avian dinosaurs and other vertebrates on Laramidia has been hypothesized as a consequence of climate variability leading to floral variability, of orogenesis or orogenic activity, and of transgressions and regressions of the Western Interior Seaway (Horner et al., 1992; Lehman, 1997; Sampson et al., 2010; Loewen et al., 2013). There has been some disagreement regarding the existence of faunal provinces on this continent (e.g., Larson and Vavrek, 2010; Lucas et al., 2016).
Unfortunately, the terrestrial fauna of Appalachia is not well-sampled, and only a limited number of dinosaur taxa are known due to lack of Cretaceous-age terrestrial sediments in eastern North America (Baird and Horner, 1977; Baird and Galton, 1981; Baird, 1986; Schwimmer, 1986; King et al., 1988; Gates et al., 2012). Preservation bias thus exists against any articulated specimens of terrestrial animals, including non-avian dinosaurs (e.g., Schwimmer et al., 1993; Schwimmer, 1997). Schwimmer (1997) suggested that, antithetic to the hypotheses of Russell (1995), the known dinosaurs from Appalachia may represent a fair selection of non-avian dinosaur groups endemic to the continent. Schwimmer (2002) also suggested that a lack of theropod diversity or abundance on Appalachia may have also been caused by competition from the massive crocodylian Deinosuchus rugosus, a species abundant in eastern North America with bite marks on both theropod and ornithopod bones attributed to it (Schwimmer, 1997; Gallagher, 1993, 1995; Schwimmer 2002).
The recent discoveries of a leptoceratopsian from the Campanian Tar Heel Formation of North Carolina (Longrich, 2016), the description of the dinosaurs Eotrachodon orientalis from the Santonian-Campanian Mooreville Chalk Formation (Prieto-Marquez et al., 2016a) and Appalachiosaurus montgomeriensis from the Campanian Demopolis Chalk Formation (Carr et al., 2005), and the description of new dinosaur remains from microfossil sites like Ellisdale and Stokes Quarry (e.g., Gallagher, 1993; Denton et al., 2011; Schwimmer, 2015) have largely increased the non-avian dinosaur diversity of Appalachia. The dinosaur fauna of Appalachia itself was apparently dominated by relict forms isolated by the Western Interior Seaway (Schwimmer et al., 1993). However, the development of Appalachian faunas and their eventual mixing with Laramidian ones during the Maastrichtian (Schwimmer et al., 1993; Carr et al., 2005) is poorly understood.
Here, I review and statistically compare faunas known from the Aptian through Maastrichtian of eastern North America with those known from the west to better illustrate the vicariance of Appalachian and Laramidian non-avian dinosaur faunas after the creation of the Western Interior Seaway. This analysis provides a temporal framework for the evolution of non-avian dinosaur faunas on Appalachia. Additionally, non-avian dinosaur provincialism during the Coniacian, Santonian, and Campanian Stages of the Cretaceous was tested using several biogeographic similarity indices and discussed, as were ecological implications regarding competition between predatory dinosaurs and crocodyliforms in Appalachian ecosystems. Because the vast majority of Appalachian dinosaur remains occur in marine deposits and Laramidian dinosaurs are comparatively well-known from terrestrial ones, depositional bias was surely a factor that may have skewed the results of the statistical analyses conducted (e.g., Schwimmer, 1997) and is discussed below. This analysis is important for being the first major one to compare the non-avian dinosaur faunas of Appalachia with Laramidian ones statistically. Specific depositional comparisons between Appalachian and Laramidian faunas analyzed are noted in the results section.
MATERIALS AND METHODS
Permits
No permits were required for the described study, which complied with all relevant regulations. Photographs of specimens from the Arundel Clay referred to herein were supplied by Thomas Jorstad of the National Museum of Natural History. Photographs of the specimens figured herein from the Yale Peabody Museum were provided by Jamie Henderson.
Institutional Abbreviations
The following abbreviations for museum collections are used in the manuscript: USNM V/PAL: United States National Museum (Smithsonian), Washington, DC, USA; UAM(1): University of Arkansas at Fayettville, Fayettville, AK, USA; YPM VPPU: vertebrate paleontology collections, Yale Peabody Museum, New Haven, CT, USA; RMM/MCWSC: McWane Science Center, Birmingham, Alabama, USA; AMNH FARB: American Museum of Natural History (fossil amphibian, bird, and reptile collections), New York, NY, USA; ANSP: Academy of Natural Sciences at Drexel University, Philadelphia, PA, USA; AUMP: Auburn University Museum of Paleontology, Auburn, AL, USA; CCK: Columbus State University (Cretaceous research collections), Columbus, GA, USA; FMNH: Field Museum of Natural History, Chicago, IL, USA; MMNS: Mississippi Museum of Natural Science, Jackson, MS, USA; MOG: Mississippi Office of Geology, Jackson, MS, USA; PPM: Memphis Pink Palace Museum, Memphis, TN, USA; UAM: University of Alabama Museum, Tuscaloosa, AL, USA.
Methods
Faunal lists. Occurrences documenting a total of 54 major clades of non-avian dinosaurs were catalogued from the western and eastern portions of North America. Faunal lists were created for all eastern North American dinosaur-bearing units that corresponded to the Aptian, Albian, Cenomanian, Turonian, Coniacian, Santonian, Campanian, and Maastrichtian stages of the Cretaceous. The biogeographic occurrences that fed the compilation of such faunal lists were taken from an extensive review of previously published works, access to unpublished information, and personal observation, representing to the author’s knowledge the most rigorous review of Appalachian dinosaur faunas to date. Detailed review and references for the faunal lists included in Appendix 1 may be found below. Aptian Appalachian faunas were compared with those of the western North American (pre-Laramidian) Ruby Ranch Member of the Cedar Mountain Formation and units V through VII of the Cloverly Formation, whereas Albian Appalachian faunas were compared with those of the Blackleaf Formation, Wayan Formation, and Willow Tank Formation. Cenomanian Appalachian faunas were compared with the upper Cedar Mountain Formation (Mussentuchit Member), Chandler Formation, and Dunvegan Formation. Turonian Appalachian faunas and the non-avian dinosaur faunas of the Moreno Hill, Frontier, and Matanuska formations were compared. Santonian Appalachian faunas were compared with those of the Milk River Formation. Campanian Appalachian faunas were compared to those of the Wahweap, Kaiparowits, and Kirtland formations representing southern Campanian Laramidian faunas and with the Oldman, Dinosaur Park, and Judith River formations representing northern Campanian Laramidian faunas, whereas the Maastrichtian Appalachian faunas were compared with those of the Javelina, Hell Creek, Lance, and Horseshoe Canyon formations. The Laramidian (western North American) faunas used for comparisons are among the most well known (e.g., Kirkland et al., 1998; Weishampel et al., 2004, Weishampel, 2006; Gates et al., 2010; Zanno and Makovicky, 2013; Sampson et al., 2013; Farke et al., 2014) and represent a sampling of northern, middle, and southern faunas, both factors being considered to minimize statistical bias and error on the part of such faunas. As such, formations like the Foremost Formation, from which only a few taxa and indeterminate elements are known (e.g., Weishampel et al., 2004), were not included.
Statistical comparisons. For comparisons between Appalachian and Laramidian faunas, Simpson’s similarity index, the Jaccard coefficient, and Jaccard distance were employed to quantify the differentiation of the faunas of the aforementioned landmasses over time (Jaccard, 1902; Jaccard, 1912; Simpson, 1943). The former two indices show the statistical similarity of two faunas, with Simpson’s similarity index emphasizing similarity and the Jaccard coefficient emphasizing differences, whereas the Jaccard distance measures the dissimilarity between two faunas and is equal to one minus the Jaccard similarity value (e.g., Jaccard, 1902; Jaccard, 1912; Simpson, 1943). These indices were calculated by comparing dinosaurian faunas at the genus and “family” (= the next highest identifiable clade above genus) levels for the purpose of shedding light on what taxa were differentiating in the context of their parent clades as well as to better factor in the many Appalachian occurrences unable to be identified to the genus or species level. Indeterminate specimens assignable to the “family level”, even those which were the only representation of the presence of a particular clade in a faunas, were only included in “family level” calculations. This protocol was used in order to make the analyses herein more comparable to those of Gates et al. (2010), who coded their datasets at the “family”, genus, and genus-species levels. The latter level was not used in analyses herein, as so few Appalachian dinosaur fossils are identifiable past the genus level (see Appendix 1 and the Review section).
As some Appalachian dinosaur faunas with biogeographically significant records (e.g., the Owl Creek Formation) have yielded less than four distinct species, they were not included in calculations of the Jaccard coefficient, Jaccard distance or of the Simpson similarity index, and instead were compared analytically with western faunas. This group of geological units includes 14 of the Appalachian faunas examined, three of which have yielded the holotype specimens of Appalachian dinosaur taxa. The results of the calculation of these indices can be found in Appendix 1 (the Jaccard distance and Jaccard coefficient are listed in the same tables, with parentheses around values of the former).
Analysis of dinosaur faunal provincialism on Appalachia. Because it has been noted that Laramidia may have had multiple different dinosaur faunal provinces during the Campanian (e.g., Lehman, 1997; Sampson et al., 2010a; Loewen et al., 2013), dinosaur provincialism on Appalachia was also investigated for during the Coniacian, Santonian, and Campanian Stages of the Late Cretaceous. This was done by first rarefying the assemblages, which represent the most well-known (in terms of number of specimens) faunas of units from the aforementioned Stages of the Late Cretaceous and then by employing the Simpson similarity index and Jaccard coefficient to compare faunal similarities at the genus and family levels for each. Rarefaction, which calculates the expected number of taxa in a given sample A if that sample were reduced to the size of a smaller sample B (e.g., Sanders, 1968), was used to examine sampling differences between Appalachian dinosaur-bearing strata and assess for sampling bias between the corresponding faunas. The relative ages of each of the stratigraphic units was also taken into account during this process, and the ages of the formations included in this analysis of provincialism are given in the results section. The fauna of the coeval Campanian Mooreville Chalk, Blufftown, and Coffee Sand formations and an unnamed clay from Missouri were also rarefied for comparison with the Coniacian/Santonian Eutaw Formation in order to track the development of dinosaur faunas in the southeastern United States during Late Cretaceous. In comparing faunas, the presence of larger phylogenetic groups was considered. In the case of Aptian, Cenomanian, and Maastrichtian Appalachian faunas, the existence of only one comparatively well-sampled fauna in each case did not allow for considerations of provincialism. The Albian Appalachian Formations were also too poorly sampled to allow for rarefaction.
Biases. As previous studies have noted, biases in the collection of fossils, use of previously published literature, taphonomic biases among and within sedimentary units, and temporal differences among sedimentary units studied and within their faunas all play a role in skewing paleobiogeographic analyses (e.g., Nicholls and Russell, 1990; Lehman, 1997; Gates et al., 2010). Because of these factors, the specific paleoenvironments, taphonomy, and age of sediments compared herein were extensively reviewed and compared in the Results section.
Biases in the collection of fossils have been noted as an appreciable hindrance to paleobiological analyses previously (e.g., Alroy et al., 2001; Smith, 2001, 2007; Lloyd et al., 2011). Indeed, such biases among the Campanian Kaiparowits, Judith River, Dinosaur Park, and Kirtland formations were reviewed by Gates et al. (2010), who noted that significant biases exist even between these and the other Campanian faunas they compared. However, because few extensive dinosaur faunas are known from the Aptian to Santonian of North America (e.g., Carpenter et al., 1995; Kirkland et al., 1998; Main, 2013), the ability to perform statistical analysis on known faunas would be severely limited if an attempt was made to rule out certain collection biases for such comparisons. Regarding to the analysis of Appalachian dinosaur provincialism during the Campanian, collection biases are easier to take into account and are discussed below.
As noted, biases in this study reflecting taxonomically or occurrence-wise inaccurate faunal lists relied on herein may also be present (e.g., Lehman, 1997; Lloyd et al., 2011). However, given the extensive literature review undertaken for the review of Appalachian faunas and for their comparison with Laramidian faunas and the large amount of data compiled, any such discrepancies seem not to be a large bias in the analyses undertaken.
REVIEW
Aptian
Arundel Clay. The most well-represented dinosaur fauna from the Early Cretaceous of eastern North America comes from the Arundel Clay of Maryland. The classification of these elongate, discontinuing clays as a formation has been debated, and the deposits have been interpreted by at least one author as those of oxbow swamps (Kranz, 1998). Further support against the existence of the Arundel as a formation comes from the fact that the palynomorphs of the Patuxent and Arundel sediments cannot be distinguished (Brenner, 1963; Doyle and Hickey, 1973; Doyle and Robbins, 1977; Robbins, 1991; Kranz, 1998). Lipka et al. (2006) considered the Arundel Clay to be a facies contained within the Potomac Formation of late-early Aptian age. The Arundel is made up of black lignite and massive dark-grey mudstones containing limonite and siderite and is the second oldest and most fossiliferous of the three Potomac facies (Lipka et al., 2006). Here, the classification of the Arundel Clay by Lipka et al. (2006) is followed.
 The non-avian dinosaur fauna present within the Arundel Clay facies consists of a variety of theropods, at least one species of sauropod, and specimens representing at least three ornithischian clades. The theropods of this unit include the dubious “Allosaurus” medius described on the basis of a single tooth (Figure 1.1) (Marsh, 1888) and other elements that were later assigned to “Dryosaurus” grandis (Lull, 1911), Coelurus gracilis described on the basis of a pedal claw (Figure 2.5) (Marsh, 1888; Lipka, 1998), and Creosaurus potens based on a caudal vertebra (Lull, 1911). “Allosaurus” medius was regarded by Lipka (1998) as a carnosaur and Coelurus gracilis a coelurosaur. Lipka (1998) noted that Ostrom (1970) had considered the holotype pedal ungual of the latter as similar to Deinonychus and suggested a possible relation. Holtz et al. (2004) listed all three of these theropods as indeterminate.
The non-avian dinosaur fauna present within the Arundel Clay facies consists of a variety of theropods, at least one species of sauropod, and specimens representing at least three ornithischian clades. The theropods of this unit include the dubious “Allosaurus” medius described on the basis of a single tooth (Figure 1.1) (Marsh, 1888) and other elements that were later assigned to “Dryosaurus” grandis (Lull, 1911), Coelurus gracilis described on the basis of a pedal claw (Figure 2.5) (Marsh, 1888; Lipka, 1998), and Creosaurus potens based on a caudal vertebra (Lull, 1911). “Allosaurus” medius was regarded by Lipka (1998) as a carnosaur and Coelurus gracilis a coelurosaur. Lipka (1998) noted that Ostrom (1970) had considered the holotype pedal ungual of the latter as similar to Deinonychus and suggested a possible relation. Holtz et al. (2004) listed all three of these theropods as indeterminate.
Several large teeth (Figure 1.2-4) described by Lipka (1998) and reviewed and figured in Weishampel (2006) have been assigned to a large allosauroid similar to or synonymous with Acrocanthosaurus. Lipka (1998) was able to assign these large, serrated teeth to Acrocanthosaurus based on several features diagnostic to that taxon found on its teeth, discussing the controversy over assigning teeth to a specific taxon of dinosaur. This carcharodontosaur has been regarded as the apex predator of the Arundel ecosystem (Weishampel, 2006). Importantly, two distinct possibilities regarding the presence of two carnosaurs in the Arundel facies have been stated (Lipka, 1998). The first, regarding the Arundel “Creosaurus” potens and Acrocanthosaurus material, suggested they represented distinct taxa, whereas the second held that the “C.” potens material belonged to the taxon Acrocanthosaurus (Lipka, 1998). This latter hypothesis is more congruent with data on carcharodontosaur ecology from geologically younger Appalachian sites and from other Early Cretaceous localities (e.g., Weishampel et al., 2004; Appendix 1). Though the fairly diverse ornithischian and small theropod fauna and the presence of at least one species of sauropod in the Arundel may have allowed for the coexistence of two large carnosaur species, more complete carcharodontosaurid specimens must be recovered to accurately test this hypothesis (Lipka, 1998). In addition to the teeth, a massive manual ungual from the Arundel (Figure 1.5) may also be assignable to Acrocanthosaurus.
 Another clade of carnivorous theropod dinosaurs represented in the Arundel are the dromaeosaurids. Lipka (1998) assigned strongly recurved, laterally compressed teeth retrieved from the Arundel facies to Deinonychus, and several are figured herein (Figure 2.1-4). Lipka (1998) noted that these teeth and those of Acrocanthosaurus from the Arundel extended the range of both taxa during the Aptian across North America.
Another clade of carnivorous theropod dinosaurs represented in the Arundel are the dromaeosaurids. Lipka (1998) assigned strongly recurved, laterally compressed teeth retrieved from the Arundel facies to Deinonychus, and several are figured herein (Figure 2.1-4). Lipka (1998) noted that these teeth and those of Acrocanthosaurus from the Arundel extended the range of both taxa during the Aptian across North America.
One of the better records of theropod dinosaurs from the Arundel is of at least two indeterminate taxa of ornithomimosaurs. The bones originally described as “Dryosaurus” grandis by Lull (1911) and later as a species of Ornithomimus (O. affinis) (Gilmore, 1920) are currently in the collections of the USNM (Figure 3.1-6). More recently, the specimens were assigned to Archaeornithomimus (Russell, 1972), to indeterminate theropods (Smith and Galton, 1990), and to Ornithomimosauria indet. (Makovicky et al., 2004; Weishampel, 2006; Brownstein, 2017a). All but one of the original specimens of Arundel ornithomimosaurs were found at the same site near Muirkirk, Maryland (Gilmore, 1920). These original specimens include a dorsal vertebral centrum, two elongated caudal vertebrae, the distal ends of metatarsals II and III, two phalanx II-1s, a partial astragalus from the left hindlimb, a pedal phalanx III-2, a partial phalanx identified as from pedal digit IV, and a single pedal ungual (Figure 3.1-6). Additionally, a partial anteroposteriorly short pedal phalanx IV-? was assigned to “Ornithomimus affinis” (Gilmore, 1920). Gilmore (1920) noted that more material from Arundel ornithomimosaurs, including a partial tibia, phalanx and pedal ungual (USNM PAL 466054) 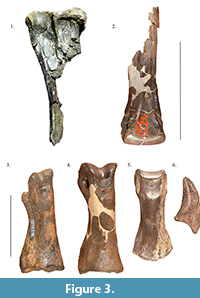 had been recovered (Figure 3.1) (Weishampel and Young, 1996), and there are also many other specimens which have not yet been described formally in the literature (pers. obs.). The original fossils found nearby Muirkirk likely come from the Dinosaur Park site, which has yielded the tibia and other pedal elements as well as a variety of other elements including many pedal unguals (Brownstein, 2017a). Notably, two morphotypes of ornithomimosaur pedal unguals (diagnosed as such based on a single flexor fossa on the ventral surfaces of the elements) are found at this site: the shorter and smaller recurved unguals like that described by Gilmore (1920), and elongated, larger unguals with more flattened ventral surfaces in lateral view and less expansive dorsal and ventral faces over their proximal articular facets (Brownstein, 2017a). This latter morphology is more akin to the pedal unguals of more derived ornithomimosaurs, suggesting the possibility of two distinct ornithomimosaurs coexisting within the Arundel facies. Two ornithomimosaur taxa are known from the Yixian Formation of China, which is of similar age to the Arundel.
had been recovered (Figure 3.1) (Weishampel and Young, 1996), and there are also many other specimens which have not yet been described formally in the literature (pers. obs.). The original fossils found nearby Muirkirk likely come from the Dinosaur Park site, which has yielded the tibia and other pedal elements as well as a variety of other elements including many pedal unguals (Brownstein, 2017a). Notably, two morphotypes of ornithomimosaur pedal unguals (diagnosed as such based on a single flexor fossa on the ventral surfaces of the elements) are found at this site: the shorter and smaller recurved unguals like that described by Gilmore (1920), and elongated, larger unguals with more flattened ventral surfaces in lateral view and less expansive dorsal and ventral faces over their proximal articular facets (Brownstein, 2017a). This latter morphology is more akin to the pedal unguals of more derived ornithomimosaurs, suggesting the possibility of two distinct ornithomimosaurs coexisting within the Arundel facies. Two ornithomimosaur taxa are known from the Yixian Formation of China, which is of similar age to the Arundel.
The sauropod material from the Arundel facies has been the subject of some taxonomic confusion (Figure 4.1-4). A sauropod tooth was named Astrodon johnstoni (Leidy, 1865). Marsh (1888) named two species of his new genus Pleurocoelus from sauropod bones discovered near Muirkirk, Maryland. Pleurocoelus nanus was named from cranial elements and multitude of other fragmentary and isolated remains of several individuals, whereas P. altus was named on the basis of a partial hindlimb (Weishampel, 2006). 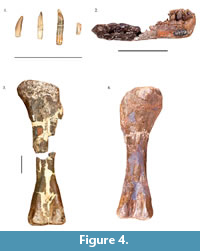 Later studies have synonymized the three taxa (Hatcher, 1903; Gilmore, 1921; Carpenter and Tidwell, 2005), though some have doubted this taxonomic classification and instead regard all three Arundel sauropod species as dubious (Rose, 2007; D’Emic, 2013). If the Arundel material does indeed belong to one valid species, the correct name would be Astrodon johnstoni, which Carpenter and Tidwell (2005) classified as a basal titanosauriform. This placement is consistent with the data on sauropod clades in North America during the Early Cretaceous (e.g., Weishampel et al., 2004; Appendix 1). Carpenter and Tidwell (2005) and Weishampel (2006) suggested that the three Arundel sauropod taxa represented different growth stages of the same taxon; Weishampel (2006) estimated juveniles at 5 m and 500 kg in size with adults approaching 20 m and 18000 kg. Juveniles of the Arundel titanosauriform may have been prey items of the carcharodontosaurids, which were present in the region.
Later studies have synonymized the three taxa (Hatcher, 1903; Gilmore, 1921; Carpenter and Tidwell, 2005), though some have doubted this taxonomic classification and instead regard all three Arundel sauropod species as dubious (Rose, 2007; D’Emic, 2013). If the Arundel material does indeed belong to one valid species, the correct name would be Astrodon johnstoni, which Carpenter and Tidwell (2005) classified as a basal titanosauriform. This placement is consistent with the data on sauropod clades in North America during the Early Cretaceous (e.g., Weishampel et al., 2004; Appendix 1). Carpenter and Tidwell (2005) and Weishampel (2006) suggested that the three Arundel sauropod taxa represented different growth stages of the same taxon; Weishampel (2006) estimated juveniles at 5 m and 500 kg in size with adults approaching 20 m and 18000 kg. Juveniles of the Arundel titanosauriform may have been prey items of the carcharodontosaurids, which were present in the region.
A single tooth was referred to the medium-sized ornithopod dinosaur Tenontosaurus sp. by Galton and Jenson (1979). This assignment was followed by Weishampel and Young (1996). Norman (2004) later assigned this specimen to Iguanodontia indet. Nevertheless, the tooth provides evidence for a large iguanodont in the Arundel ecosystem, adding to the similarities between the eastern Arundel fauna and the Aptian faunas of western North America (e.g., Ostrom, 1970; Forster, 1984; Forster, 1990; Winkler et al., 1997; Weishampel et al., 2004).
Armored dinosaurs left one of the better records of Early Cretaceous eastern North American ornithischians, and are represented by the genus Priconodon crassus in the Arundel Clay (Figure 5.1-5). This animal was first described on the basis of a single tooth (e.g., Carpenter and Kirkland, 1998), and additional teeth, an osteoderm (Weishampel, 2006), and a tibia (Carpenter, 2001; Vickaryous et al., 2004) have since been recovered. A multitude of teeth and the tibia referred to this taxon are in the collections of the United States National Museum. These teeth are all similar in being triangular, short, and having large denticles, and are assigned to nodosaurs based on the presence of a cingulum on the tooth between the base and the entirety of the crown and their narrow morphology (Weishampel, 2006). Notably, Priconodon crassus has been regarded as an unusually large nodosaurid based on the huge size of the teeth assigned to it (Carpenter, 2001). Though some have regarded this fragmentary taxon dubious (Vickaryous et al., 2004), the validity of this species has been supported by multiple studies comparing the morphology of the teeth of Priconodon with other nodosaurids (Carpenter, 2001; West and Tibert, 2004).
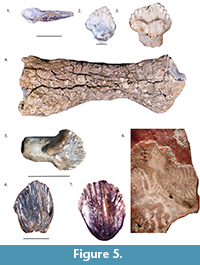 Lastly, the Arundel has surprisingly yielded teeth (Figure 5.6-7) described in detail and found to be most similar to those of neoceratopsians by Chinnery et al. (1998). The Arundel neoceratopsian teeth are especially important as they predate the age of the holotype skeleton of Aquilops americanus, which was discovered in Albian deposits (Farke et al., 2014), teeth from the Cenomanian Mussentuchit Member of the Cedar Mountain Formation (Chinnery et al., 1998), and possible ceratopsian remains from the late Albian of Idaho (Weishampel et al., 2002). This makes the Arundel teeth the oldest occurrence of neoceratopsians in North America, suggesting that the clade arrived in the continent during the middle Early Cretaceous and thus supporting the hypothesis that neoceratopsians had dispersed into North America during the Aptian (Farke et al., 2014). Farke et al. (2014) suggested that at least three interchanges of neoceratopsians, the first being of Aquilops-like taxa, occurred between North America and Asia. Therefore, the Aptian age of the Arundel teeth suggests that the Arundel neoceratopsian may have been part of this dispersal and therefore would have been similar to the small Aquilops in form.
Lastly, the Arundel has surprisingly yielded teeth (Figure 5.6-7) described in detail and found to be most similar to those of neoceratopsians by Chinnery et al. (1998). The Arundel neoceratopsian teeth are especially important as they predate the age of the holotype skeleton of Aquilops americanus, which was discovered in Albian deposits (Farke et al., 2014), teeth from the Cenomanian Mussentuchit Member of the Cedar Mountain Formation (Chinnery et al., 1998), and possible ceratopsian remains from the late Albian of Idaho (Weishampel et al., 2002). This makes the Arundel teeth the oldest occurrence of neoceratopsians in North America, suggesting that the clade arrived in the continent during the middle Early Cretaceous and thus supporting the hypothesis that neoceratopsians had dispersed into North America during the Aptian (Farke et al., 2014). Farke et al. (2014) suggested that at least three interchanges of neoceratopsians, the first being of Aquilops-like taxa, occurred between North America and Asia. Therefore, the Aptian age of the Arundel teeth suggests that the Arundel neoceratopsian may have been part of this dispersal and therefore would have been similar to the small Aquilops in form.
The Arundel Clay dinosaur fauna therefore consists of Acrocanthosaurus sp. and perhaps another large carnosaur, indeterminate coelurosaurs, the dromaeosaurid Deinonychus (including “Coelurus” gracilis), two unnamed possible species of ornithomimosaurs, the titanosauriform sauropod Astrodon johnstoni, iguanodontian dinosaurs similar to or possibly synonymous with Tenontosaurus, the large nodosaurid dinosaur Priconodon crassus, and a neoceratopsian.
Patuxent facies. Another one of the facies assigned to the Aptian by Lipka et al. (2006) is the Patuxent, the oldest unit exposed in the coastal plain of Maryland and Virginia (Stanford et al., 2004). This facies consists of sandstones mixed with light grey mudstones (Lipka et al., 2006). Though this formation has not preserved an extensive faunal list like the western Cloverly or Cedar Mountain Formations (Weems and Bachman, 1997), a somewhat extensive ichnological record of dinosaurs has been preserved in Patuxent facies. These include the tracks of theropods (Megalosauropus sp.), euornithopods (Amblydactylus sp.), and an ichnotaxon based on the tracks of a small ornithopod with possible affinities to the western form Zephyrosaurus (Hypsiloichnus marylandicus) (Weems and Bachman, 1997; Stanford, 1998; Stanford and Stanford, 1998; Stanford et al., 2004; Weishampel et al., 2004). One study (Lockley and Stanford, 2004) reported from the siliciclastic Patuxent facies the tracks of 14 different morphotypes of ornithopod, theropod, sauropod, and ankylosaur tracks. Lockley and Stanford (2004) also reported the presence of small tracks interpreted as those of hatchling dinosaurs alongside those of juveniles and adults, which they regarded as indicating the presence of nests nearby. Weems and Bachman (2015) reviewed and added to the known dinosaur ichnotaxa from the Patuxent facies, which they found to include the theropod ichnotaxon Megalosauropus sp., the ornithomimosaur ichnotaxon Ornithomimipus angustus, the sauropod ichnotaxon Brontopodus birdi (suggested to be the track of a titanosauriform), the ichnontaxon Tetrapodosaurus borealis (interpreted as a nodosaurid and compared with Propanoplosaurus), the small ornithopod taxon Hypsiloichnus marylandicus (suggested to be tracks of a dinosaur of similar grade to Zephyrosaurus schaffi), and the medium-sized to large euornithopod tracks Caririchnium leonardii (suggested to be tracks of Tenontosaurus), Gypsichnites pacensis (suggested to be tracks of an iguanodontid of similar size to Hippodraco scutodens), and Amblydactylus gethingi (suggested to be the tracks of a hadrosauroid similar to Eolambia) (Weems and Bachman, 2015).
The nodosaur genus Propanoplosaurus marylandicus (Figure 5.8) is known from a specimen constituting of both molds and casts of the skeleton of a neonate individual which was recovered alongside the ichnofossils of dinosaurs (Stanford et al., 2011). The specimen included the posterior cranium, the ribcage vertebrae, the right femur and portions of the pes, and the partial right forelimb. The holotype of Propanoplosaurus is important for being the first nodosaur skeleton from the eastern seaboard (Stanford et al., 2011) and along with Priconodon is the only valid nodosaur taxon named from the Aptian of the east coast of North America.
Arkansas Trinity Group. The Trinity Group in Arkansas consists of varying layers of fine quartz sand, clay, barite, celestite, gravel, and fossiliferous gypsum and limestone (Dane, 1929; Hunt-Foster, 2003). These sediments, which are deposited in an unconformity with eroded Paleozoic rock, originated in the Ouachita Mountains (Hunt-Foster, 2003). The record of dinosaurs from the Aptian of Arkansas is limited. However, the partial right pes of an ornithomimosaurian dinosaur (Quinn, 1973; Hunt-Foster and Kirkland, 2017) was collected from what is now termed the “Friday Site”. This specimen, UAM(1) 74, consists of metatarsals II, III, and IV and four phalanges and portions of the pedal unguals from pedal digits II, III, and IV reported from what is now termed the “Friday site” (due to it being on the property of Joe B. Friday, discoverer of the specimen) in Lockesburg Arkansas (Quinn, 1973; Hunt-Foster, 2003). Both the holotype and casts of the specimens are in the collections of the University of Arkansas.
Albian
Dakota Formation. The Albian Dakota Formation of Kansas and Nebraska has yielded the remains of a variety of dinosaurs whose relatives are also observed from the facies of the Potomac Formation. This formation consists of coastal and marine fluvial deposits in Kansas (e.g., Liggett, 2005) and as fluvial to estuarine deposits characterized by major facies changes and having common hydromorphic paleosols and disticontinous, long lignites and carbonaceous mudstones in Nebraska (e.g., Brenner et al., 2000; Joeckal et al., 2004). The most complete dinosaur skeleton ever found on the eastern margin of the Dakota Formation is the Kanas taxon Silvisaurus condrayi. The holotype specimen of this dinosaur was retrieved from a site pertaining to the Terra Cotta Clay member of the Dakota Formation in Ottawa County, Kansas, a hard, limonite-containing sandstone that was cross-bedded (Eaton, 1960). The environment in which Silvisaurus condrayi would have lived is regarded as a warm-temperate forest based on fossil leaves found nearby the Silvisaurus site (Eaton, 1960). In addition to a partial skeleton including cervical and dorsal vertebra, a sacrum, and armor including a spike possibly from the shoulder, the holotype of Silvisaurus condrayi also includes a skull and a left mandible, all of it corresponding to an individual approximately 3 m long (Eaton, 1960). During the Skull Creek highstand, the site where Silvisaurus condrayi was found would have been on the coast of the newly-formed continent of Appalachia (e.g., Eaton, 1960). The natural mold of the possible sacrum of another Silvisaurus has been retrieved from Russell County Dakota Formation exposures (Liggett, 2005).
In addition to Silvisaurus, the Dakota Formation of Kansas has yielded the tracks of ornithomimosaurs (Magnoavipes sp.), of possible ankylosaurids, and of indeterminate dinosaurs (Liggett, 2005). The probable sacrum of an ankylosaur has also been found in Cloud County, Kansas (Liggett, 2005). In Nebraska, both ornithopod footprints and the proximal end of an ornithopod femur have been recovered from Dakota Formation sediments (Joeckel et al., 2004).
Paw Paw Formation. The Paw Paw Formation of Texas is middle Albian in age and has produced the remains of two to three species of nodosaurid dinosaurs along with indeterminate nodosaur remains (e.g., Coombs, 1995; Lee, 1996; Weishampel et al., 2004). The formation is made up of ferruginous clay and sand (Hill, 1894) and was deposited in a nearshore marine setting (Scott et al., 1978). Nodosaurid dinosaurs have left a number of specimens in this formation, including a juvenile nodosaur (Jacobs et al., 1994) and two named taxa.
Pawpawsaurus campbelli is known from a complete skull lacking mandibles from Tarrant County, Texas that is morphologically similar in some ways to Silvisaurus condrayi (Lee, 1996 Paulina-Carabajal et al., 2016). Texasetes pleurohalio is known from a partial skeleton including a skull fragment, a tooth, elements from the limbs, portions of the pelvis and scapulocoracoid, and vertebrae (Coombs, 1995) and may be a synonym of Pawpawsaurus (Lee, 1996). However, in the phylogenetic analysis of Ankylosauria, the two Paw Paw nodosaur taxa and the Paw Paw nodosaur juvenile were found to be in notably different phylogenetic positions, with Pawpawsaurus the sister taxon to Europelta, Texasetes a sister taxon to Edmontonia, Denversaurus, and an unnamed nodosaur from Argentina, and the Paw Paw juvenile a sister taxon to Niobrarasaurus (Arbour et al., 2016).
Paluxy Formation. The Paluxy Formation of Texas, which is also middle Albian in age (e.g., Jacobs and Winkler, 1998; Weishampel et al., 2004; D’Emic, 2013), is a thin unit composed of shale and sandstone (Caughey, 1977). This unit has preserved arguably the most diverse of dinosaur faunas corresponding to Appalachia during the Albian. In addition to the remains of indeterminate theropods, the fossils of indeterminate dromaeosaurids have been recovered (e.g., Langston, 1974; Weishampel et al., 2004). Importantly, remains assigned to Ornithomimus sp. have also been recovered from the formation (e.g., Langston, 1974; Weishampel et al., 2004). As noted, both ornithomimosaurs and dromaeosaurs are known from Aptian to Albian deposits in the American west (Ostrom, 1969; Ostrom, 1970; Ostrom 1976; Cifelli, 1997; Cifelli and Gardner, 1997, Weishampel et al., 2004; Signac and Mackovicky, 2010; Oreska and Carrano, 2013; Brownstein, 2017a). Notably, “Laelaps” (= Dryptosaurus sp.) has also been reported from the Paluxy Formation (Langston, 1974), but these remains are likely those of indeterminate coelurosaurs or simply indeterminate theropods. Remains assigned to titanosauriforms (=“Pleurocoelus”) have also been reported from the formation (Langston, 1974; Weishampel et al., 2004). These specimens have more recently been assigned to the taxa Astrophocaudia and Cedarosaurus by D’Emic (2013). Indeterminate sauropod fossils have also been reported from the Cloverly Formation (e.g., Ostrom, 1970; Oreska and Carrano, 2013), while in Texas the ichnotaxon Brontopodus birdi has been reported from multiple localities (e.g., Langston, 1974; Pittman, 1989). Additionally, specimens assigned to nodosaurids and Tenontosaurus have been reported from the Paluxy Formation (Langston, 1974; Weishampel et al., 2004).
Glen Rose Formation. The Glen Rose Formation, which consists of alternating hard limestone and marl or marly limestone (Sellards et al., 1932), has preserved both dinosaur body specimens and ichnofossils. Trackways include the ichnotaxon Eubrontes glenrosensis (=?Acrocanthosaurus), which is found alongside the tracks of sauropods (Brontopodus) in the Paluxy River Valley (Farlow et al., 2010). There are also additional sites within this valley which have produced tridactyl tracks (Farlow et al., 2010). The Glen Rose has produced the trackways of both theropods and ornithopods (e.g., Wrather, 1922; Gould, 1929; Houston, 1933; Bird, 1939; Bird, 1944; Langston, 1974; Kuban, 1986; Pittman, 1989; Hawthorne and Bonem, 2002; Vance, 2002; Rogers, 2003). The ornithopod Tenontosaurus has also been reported from the formation (Weishampel et al., 2013). A partial juvenile titanosauriform skeleton is known from the Glen Rose as well (e.g., Langston, 1974; D’Emic, 2013), though it is not diagnostic to the genus level. In its entirety, the documented Glen Rose fauna consists of large theropods represented by tracks, sauropods, and ornithopods. This fauna is also somewhat similar to the theropod and ornithopod fauna of a track locality from the Chuta Formation of Mexico (Ferrusquía-Villafranca and Applegate, 1978).
Cenomanian
Woodbine Formation. The Woodbine Formation of Texas has preserved the singularly most complete dinosaur fauna of eastern North America during the early Late Cretaceous, representing fluvial, shelf, and deltaic deposits (Oliver, 1971; Trudel, 1994; Main, 2005; Main, 2013), and consisting primarily of sandstones and shales (Johnson, 1974).
The most diverse non-avian theropod dinosaur assemblage from the Woodbine Formation comes from a locality known as the Arlington Archosaur Site, which would have been on the Rudradia Peninsula of Appalachia during the Cenomanian (Main, 2013). This locality has produced the teeth of both dromaeosaurids and adult and juvenile allosauroids as well as the remains of other coelurosaurs (Main, 2013). The proximal portion of a large manual ungual was also recovered and may belong to an animal similar to Allosaurus, Acrocanthosaurus, or Suchomimus (Main, 2013). The latter possibility is unlikely, as no spinosaurid remains have currently been reported from North America. Main (2013) also discussed the possibility that some of the material recovered at the site could belong to tyrannosauroids. Additionally, teeth assigned to the taxon Richardoestesia have also been retrieved from the Arlington Archosaur Site (Main, 2013). Additions to the theropod fauna known from the Woodbine Formation include cf. Richardoestesia teeth and the tracks of theropods, including ornithomimosaurs (Magnoavipes) (Lee, 1997a, 1997b; Lockley et al., 2001; Lockley et al., 2011). The Arlington Archosaur Site also has preserved the remains of the hadrosauroid dinosaur Protohadros byrdi and P. sp. (Main, 2013). This genus of hadrosauroid dinosaur was originally named on the basis of a partial skull and fragmentary skeleton retrieved from Flower Mound in Denton County, Texas, and has been estimated at 7 to 8 m in length (Head, 1996; Head, 1998). This species possessed a robust set of ventrally oriented mandibles suggested as an adaptation for the consumption of low-lying plant matter (Head, 1996; Head, 1998). A set of ornithopod tracks from the Woodbine Formation have been assigned the name Caririchnium protohadrosaurichnos based on the hypothesis that they may represent the tracks of this taxon (Lee, 1997b).
Ornithischian remains assigned to indeterminate hadrosaurs and a basal nodosaurid have reported from the Woodbine Formation (Lee, 1997a). The Woodbine nodosaur, which is known from teeth, limb elements, and an osteoderm, may be a distinct species (Lee, 1997a).
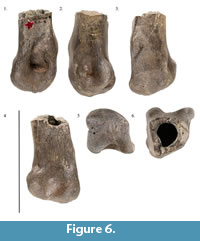 Raritan Facies. The Cenomanian Raritan facies of the upper Potomac Formation (e.g., Dalton et al., 1999; Miller et al., 2004; Lipka et al., 2006) preserves a scant but biogeographically significant non-avian dinosaur fossil record. This unit is made up of alternating clay and sand beds (Kimyai, 1966; Dalton et al., 1999; Miller et al., 2004; Lipka et al., 2006) and has produced both the only record of dinosaur tracks from the Cretaceous east of the Mississippi river and several other specimens, including an isolated distal metatarsal II of a tyrannosaur (YPM VPPU 016760) most similar to Appalachiosaurus montgomeriensis (Figure 6.1-6) (Baird, 1988, 1989; Gallagher, 1997; pers. obs.).
Raritan Facies. The Cenomanian Raritan facies of the upper Potomac Formation (e.g., Dalton et al., 1999; Miller et al., 2004; Lipka et al., 2006) preserves a scant but biogeographically significant non-avian dinosaur fossil record. This unit is made up of alternating clay and sand beds (Kimyai, 1966; Dalton et al., 1999; Miller et al., 2004; Lipka et al., 2006) and has produced both the only record of dinosaur tracks from the Cretaceous east of the Mississippi river and several other specimens, including an isolated distal metatarsal II of a tyrannosaur (YPM VPPU 016760) most similar to Appalachiosaurus montgomeriensis (Figure 6.1-6) (Baird, 1988, 1989; Gallagher, 1997; pers. obs.).
The trackway described in detail by Baird (1989) consists of the pes prints of a large theropod dinosaur. These tracks have been regarded to have affinities with those of a “megalosaurian type” (Baird, 1989). More recent studies have cast doubt on assigning any Cretaceous tracks to megalosaurs (Lockley et al., 1998). All of these tracks are now lost except for one track on display at the Rutgers University Geology Museum (Baird, 1989; Gallagher, 1997). A trace of the track at Rutgers was figured in Baird (1989) and is nearly identical in form to the Saurexallopus tracks from western North America figured by Gierlinski and Lockley (2013) in the slenderness and lengths of digits II through IV, the morphology for the impression of digit I, the relative size and shape of the digits to each other, and the presence of a noticeable hallux track alongside a somewhat centrally located noticeable metatarsophalangeal pad suggesting the presence of an arctometatarsus in the track maker. The swelled pad behind pedal digit III and the reversed hallux were noted by Gierlinski and Lockley (2013) to be features of avian pedal morphology. Additional tracks figured in Baird (1989) in photographs taken before the tracks were lost are identical to some western Saurexallopus tracks listed in Gierlinski and Lockley (2013) (Gierlinski and Lockley, 2013, figures 23.1A, C, 23.3B) and also seem to show the presence of a clearly centralized triangular metatarsophalangeal pad extremely indicative of an arctometatarsalian condition and a hallux toe indicative of an avian pes as noted by Gierlinski and Lockley (2013). Gierlinski and Lockley (2013) suggested that the trackmakers of western Saurexallopus were oviraptorosaurians, especially noting Hagryphus and Chirostenotes as plausible candidates based on the presence of a well-developed hallux toe among members of the Oviraptorosauria. This feature is shared by the larger Woodbridge tracks. Thus, it may be that the Woodbridge tracks represent some large oviraptorosaur. However, here the relationships of the tracks are regarded as equivocal among Theropoda.
In addition to theropod material, euornithopod tracks have also been recovered from the Raritan facies of the Potomac Formation (e.g., Weishampel et al., 2004). These represent an important occurrence as they evince the presence of herbivorous dinosaur taxa in the upper Potomac fauna. The possibility also remains that these tracks belong to hadrosauroids, though any assignment beyond Euornithopoda would be tentative.
The Potomac Formation clearly represents an enticing look into the non-avian dinosaur fauna of the Cenomanian of northern Appalachia. Currently, the sparse record of non-avian dinosaurs has produced at least three different types of dinosaur. Importantly, the Raritan Facies of the Potomac Formation is a terrestrial deposit, suggesting a likelihood that future dinosaur specimens may be discovered from the formation (e.g., Gallagher, 1997).
Coniacian and Santonian
Niobrara Formation (Smoky Hill Chalk). The Niobrara Formation non-avian dinosaur fauna is important for being the only known non-avian dinosaur fauna from the Coniacian of Appalachia as well as for being one of the only records of Coniacian North American dinosaurs. The two most completely known taxa from this formation are the nodosaurid Niobrarasaurus coleii and the derived non-hadrosaurid hadrosauroid Claosaurus agilis, both from Coniacian-Santonian sediments (Carpenter et al., 1995; Weishampel et al., 2004; Prieto-Márquez et al., 2016b). Additionally, the nodosaurid Heirosaurus sternbergi is known from more fragmentary remains (Carpenter et al., 1995), though it may be a synonym of Niobrarasaurus (Carpenter et al., 1995; Everhart, 2005).
Indeterminate nodosaurid and hadrosauroid specimens have also been recovered (Carpenter et al., 1995; Everhart, 2005; Everhart and Hamm, 2005; Everhart and Ewell, 2006; Everhart, 2014). One set of hadrosaurid caudal vertebrae from an animal somewhat larger than the holotype of Claosaurus agilis has been recovered with evidence of consumption by a large shark (Everhart and Ewell, 2006). The preservation of partial specimens in the marine Smoky Hill Chalk is typical of the bloat-and-float model of preservation, which often characterizes Appalachian dinosaur specimens (Schwimmer, 1997). The author follows Carpenter et al. (1995) and Prieto-Márquez et al. (2016b) in considering this fauna to be Appalachian, a hypothesis which is supported by the close position of Claosaurus to Lophorhothon as a non-hadrosaurid hadrosauroid and of Niobrarasaurus as outside the group containing the derived western nodosaurids of the Late Cretaceous (e.g., Arbour et al., 2016; Prieto-Márquez et al., 2016b), being congruent with the current understanding of Appalachian as a refugium (e.g., Schwimmer, 1997). Further discussion on the assignment of the Niobrara fauna to an Appalachian origin may be found in the discussion section.
Claosaurus agilis was a small hadrosauroid and is only known presently from a fragmentary specimen that has been described in detail (Carpenter et al., 1995) and compared with other Appalachian hadrosauroids (Prieto-Márquez et al., 2016b). Claosaurus has most recently been resolved as a derived hadrosauroid just outside to Hadrosauridae (Prieto-Márquez et al., 2016a).
Niobrarasaurus coleii is the most completely known of the three ornithischians from the Smoky Hill Chalk, and the holotype of that taxon consists of a partial specimen including portions of the skull, vertebrae, partial limbs, and a variety of osteoderms (Carpenter et al., 1995) originally described as a species of Hierosaurus (Mehl, 1936). Hierosaurus sternbergi, the original “Niobrara nodosaur”, is based on a multitude of osteoderms first described in 1905 by George Wieland (Wieland, 1905). More recently, Heirosaurus has been regarded as a nomen dubium (e.g., Carpenter et al., 1995).
McShan Formation. The McShan Formation of the southeastern United States also provides an important glimpse into Coniacian Appalachian faunas, arguably making the Appalachian dinosaur record from the Coniacian more complete than that from Laramidia. Several tyrannosaur fossils have been reported from this unit (Ebersole and King, 2011).
Eutaw Formation. The Santonian strata of southeastern North America produce the first complete non-avian dinosaur faunas from the Late Cretaceous that are distinctly ‘Appalachian’ in composition. One of the units has preserved a non-avian dinosaur fauna dating from the late Coniacian to the Campanian is the Eutaw Formation of Alabama and Mississippi, which has preserved a diverse fauna of dinosaurs including indeterminate theropods, hadrosaurids, and possibly indeterminate ankylosaurids (Kaye and Russell, 1973; Lamb, 1996; Weishampel et al., 2004). A complete review of Alabama dinosaur material (excluding Eotrachodon orientalis) found the Tombigbee Sand Member of the Eutaw Formation to have an even more diverse fauna consisting of dromaeosaurs, indeterminate tyrannosaurs (including possibly Appalachiosaurus montgomeriensis), ornithomimosaurs, nodosaurids, Lophorhothon atopus, and a multitude of hadrosaur specimens (including one notable partial specimen in the collections of the Mississippi Museum of Natural Science) (Ebersole and King, 2011).
Campanian
Demopolis Chalk Formation. The Demopolis Chalk Formation is a marine layer deposited around 78 million years ago that has yielded the most complete skeleton of an Appalachian tyrannosaur currently known: the holotype of Appalachiosaurus montgomeriensis (Carr et al., 2005). The holotype of Appalachiosaurus montgomeriensis was found at the Turnipseed Dinosaur Site and consists of the mostly complete skull and partial skeleton of a subadult tyrannosauroid (Carr et al., 2005). Multiple phylogenetic analyses using different character lists and different taxa have consistently recovered Appalachiosaurus montgomeriensis as a close outgroup to Tyrannosauridae and slightly more derived than the other currently named Appalachian tyrannosaur Dryptosaurus aquilunguis (e.g., Carr et al., 2005; Brusatte et al., 2011; Loewen et al., 2013; Fiorillo and Tykoski, 2014; Brusatte and Carr, 2016; Brusatte et. al., 2016). Two caudal vertebrae of the holotype of A. mongomeriensis are fused, possibly due to an injury the young dinosaur sustained to the tail (Carr et al., 2005). The holotype subadult specimen is estimated to have been around 6-7 meters long (Carr et al., 2005).
Additional remains of non-avian dinosaurs from the Demopolis Chalk Formation include vertebrae, hindlimb elements, and a single tooth assignable to indeterminate hadrosaurs (Ebersole and King, 2011). These large herbivores likely constituted as a prey source for Appalachiosaurus montgomeriensis.
Mooreville Chalk, Blufftown, Coffee Sand Formations, and unnamed Missouri clay unit
The Mooreville Chalk has yielded a diverse dinosaur fauna from the Santonian and Campanian. This dinosaur fauna included at least two species of hadrosauroid dinosaur (Lophorhothon atopus and Eotrachodon orientalis), the ornithomimosaur “Ornithomimus” antiquus, the dromaeosaurid Saurornitholestes, indeterminate hadrosauroids, nodosaurids, indeterminate theropods, and notably two different avian dinosaur taxa (Lull and Wright, 1942; Langston, 1960; Olson, 1975; Dobie, 1978; Lamb et al., 1993; Lamb, 1996, 1997, 1998, 2001; Chiappe et al., 2002; Weishampel et al., 2004; Kiernan and Schwimmer, 2004; Prieto-Márquez et al., 2016a; Prieto-Márquez et al., 2016b). Its equivalent to the east, the Blufftown Formation (e.g., Schwimmer, 1993; Kiernan and Schwimmer, 2004; Ebersole and King, 2011), has preserved additional non-avian dinosaur fossils, including those assigned to indeterminate hadrosaurids, ?Albertosaurus sp. and indeterminate ornithomimids (e.g., Schwimmer et al., 1993; Ebersole and King, 2011). Schwimmer et al. (1993) noted that one element assigned to Albertosaurus was virtually indistinguishable from that of Appalachiosaurus montgomeriensis, and more recent studies have considered the Blufftown “Albertosaurus” material to be of A. montgomeriensis (Ebersole and King, 2011). The holotype of Appalachiosaurus montgomeriensis itself was also previously referred to Albertosaurus prior to its recognition as a new taxon (Carr et al., 2005).
In addition to the large tyrannosauroid Appalachiosaurus montgomeriensis, smaller theropod remains have also been recovered from these Campanian strata. The dromaeosaurid dinosaur Saurornitholestes sp. was reported from the Mooreville Chalk Formation on the basis of a tooth (Kiernan and Schwimmer, 2004), and indeterminate dromaeosaurid remains are also known from the Mooreville Chalk and Blufftown Formations (e.g., Ebersole and King, 2011). Ornithomimosaurs are represented by a single element assigned to “Ornithomimus” antiquus from the Blufftown Formation (Schwimmer et al., 1993; Ebersole and King, 2011).
Lophorhothon atopus is known from a partial skull and skeleton collected from lower Campanian Mooreville Chalk sediments outcropping in Dallas County, Alabama (Langston, 1960; Prieto-Márquez et al., 2016b). This hadrosauroid dinosaur was around 7.5 m in length (Schwimmer, 2002) and has been resolved as a derived hadrosauroid close to Hadrosauridae in a recent phylogenetic analysis (Prieto-Márquez et al., 2016a). Eotrachodon orientalis, the most recently described hadrosauroid from the Mooreville Chalk Formation, was described on the basis of a juvenile specimen estimated between 4 and 5.1 m in length (Prieto-Márquez et al., 2016a; Prieto-Márquez et al., 2016b). This specimen represents the most completely known hadrosauroid dinosaur from the landmass of Appalachia and along with other Appalachian hadrosaurid taxa suggests that the landmass was where the hadrosaurid dinosaurs may have first evolved (Prieto-Márquez et al., 2016a; Prieto-Márquez et al., 2016b). Eotrachodon orientalis may have gotten somewhat larger in size than the holotype juvenile specimen, which is thought to have been several years old at the time of death (Prieto-Márquez et al., 2016b). In addition to the two named taxa from the Mooreville Chalk, indeterminate hadrosaurids (including hadrosaurines) are known from both the Blufftown and Mooreville Chalk Formations, and indeterminate ornithischian remains from the Selma group may also be from these geological units (Langston, 1960; Schwimmer et al., 1993; Ebersole and King, 2011).
In addition to the hadrosauroids of the Mooreville and Blufftown Formations, the armored nodosaurids were also present. These are represented by indeterminate remains and the notable partial specimen of a juvenile nodosaur labeled RMM 1224 from the Mooreville Chalk (Langston, 1960; Lamb, 1996; Ebersole and King, 2011; Ebersole, personal commun., 2016). This specimen is the most complete nodosaur known from east of the Mississippi river (e.g., Ebersole and King, 2011) and probably represents a distinct taxon based on its stratigraphic location in comparison to other nodosaurs.
The Chronister Site of southeastern Missouri (e.g., Fix and Darrough, 2004) represents a unique chance to view the dinosaur fauna found in western Appalachia, bearing a notable assemblage corresponding to an unnamed smectite clay unit of Santonian to Campanian age overlapping in age with the Coffee Sand Formation (Ebersole, 2009) and thus perhaps with the Mooreville Chalk and Blufftown formations (Ebersole and King, 2011). The non-avian dinosaur fauna known from the Chronister site is somewhat diverse, containing both indeterminate dromaeosaurid and tyrannosauroid material, as well as a species of hadrosauroid (Fix and Darrough, 2004). This latter species, originally named Neosaurus missouriensis and incorrectly identified as a sauropod (Gilmore and Stewart, 1945), was later assigned a new genus Parrosaurus due to the name Neosaurus being occupied (Gilmore, 1945). Later, Baird and Horner (1979) assigned P. missouriensis to Hypsibema based on the similarity of their caudal vertebrae. More recent discoveries show the site is rich in dinosaur fossils warranting study (Darrough, personal commun., 2016) and in some places the site may even constitute as a bone bed (Fix and Darrough, 2004). Additional remains of Hypsibema missouriensis, including skull elements, have also been recovered (Darrough et al., 2005; Darrough, personal commun., 2016). This dinosaur is likely assignable to its own genus, Parrosaurus, as it is not only separated from the localities where the material assigned to H. crassicauda was found but also is now known from more material, which may allow for detailed description (e.g., Darrough et al., 2005). Therefore, the name Parrosaurus missouriensis is preferred herein. This hadrosauroid was of huge size, as its distal caudal vertebrae are similar in dimensions to those of Hypsibema crassicauda (Baird and Horner, 1979). As Baird and Horner (1979) suggested, the similarity of the vertebrae of these two taxa indeed suggest a relationship between the two species. The Coffee Sand has been considered equivalent in age to the Mooreville Chalk and Blufftown Formations (Ebersole and King, 2011). Additional remains from the Coffee Sand include those of an indeterminate hadrosauroids and the limb element of a possible Eotrachodon orientalis adult (Ebersole and King, 2011; Prieto-Márquez et al., 2016b).
Tar Heel and Coachman formations. The Tar Heel and Coachman formations, which are equivalent in age (Schwimmer et al., 2015), have produced among the most extensive Appalachian dinosaur faunas. This fauna is middle Campanian in age (e.g., Baird and Horner, 1979; Self-Trail et al., 2004; Weishampel et al., 2004; Schwimmer et al., 2015), and is most extensively known from two sites. These are the Phoebus Landing site on the Cape Fear river of southern North Carolina (Bladen County) and the Stokes Quarry Site in northern South Carolina (Darlington County) (e.g., Miller, 1967; Baird and Horner, 1979; Schwimmer et al., 2015). The former site has produced four or more different species of dinosaur (e.g., Miller, 1967; Baird and Horner, 1979; Weishampel and Young, 1996; Weishampel, 2004), whereas the latter has preserved an extensive theropod dinosaur fauna and indeterminate material from hadrosaurids (Weishampel and Young, 1996; Schwimmer et al., 2015).
The Phoebus Landing fauna includes a multitude of hadrosauroid taxa. The taxon Lophorhothon has been reported from the site (Miller, 1967; Baird and Horner, 1979; Weishampel and Young, 1996; Weishampel et al., 2004) alongside material assigned to Hadrosaurus sp. and a small unnamed taxon or juvenile hadrosaur (= “Hadrosaurus” minor) (Baird and Horner, 1979). Additional remains assignable to indeterminate hadrosaurids have also been recovered from the Phoebus Landing Site and Stokes Quarry (Miller, 1967; Baird and Horner, 1979; Schwimmer et al., 2015). A right metatarsal III recovered from the Phoebus Landing site is comparable to that of Hadrosaurus foulkii and provides evidence for the presence of an ~8 m hadrosaur in the Phoebus Landing fauna (Baird and Horner, 1979). Additionally, a partial tooth from Stokes Quarry was noted by Schwimmer et al. (2015) to compare favorably with “hadrosaurines” (Hadrosauroidea indet.). Weishampel and Young (1996) documented the discovery of many hadrosaur teeth, vertebrae, and limb material, including the partial femur of a hadrosauroid.
 The behemoth hadrosauroid Hypsibema crassicauda is also known from caudal vertebrae from Phoebus Landing (Figure 7.1) and from other sites pertaining to the Tar Heel Formation (e.g., Cope, 1871; Miller, 1967; Baird and Horner, 1979; Weishampel and Young, 1996). A very large partial hadrosaur humerus from the Tar Heel Formation was described by Baird and Horner (1979) and is tentatively referred to as Hypsibema crassicauda based on its estimated complete size of 830 mm, massive for a hadrosaur (Baird and Horner, 1979). This dinosaur has been estimated at 12 m or more in length (Baird and Horner, 1979; Weishampel and Young, 1996), and based on comparisons with the vertebrae of the hadrosaurs Hadrosaurus foulkii (Cope, 1871) and Eotrachodon orientalis (Prieto-Márquez et al., 2016b) in comparison to the estimated size of each of these taxa (Weishampel and Young, 1996; Prieto-Márquez et al., 2016b), the author herein makes a tentative estimate of the size of this hadrosaur as being around 12-17 m in length. The implications of the large size attained by H. crassicauda and other Appalachian hadrosauroids are considered in the Discussion section.
The behemoth hadrosauroid Hypsibema crassicauda is also known from caudal vertebrae from Phoebus Landing (Figure 7.1) and from other sites pertaining to the Tar Heel Formation (e.g., Cope, 1871; Miller, 1967; Baird and Horner, 1979; Weishampel and Young, 1996). A very large partial hadrosaur humerus from the Tar Heel Formation was described by Baird and Horner (1979) and is tentatively referred to as Hypsibema crassicauda based on its estimated complete size of 830 mm, massive for a hadrosaur (Baird and Horner, 1979). This dinosaur has been estimated at 12 m or more in length (Baird and Horner, 1979; Weishampel and Young, 1996), and based on comparisons with the vertebrae of the hadrosaurs Hadrosaurus foulkii (Cope, 1871) and Eotrachodon orientalis (Prieto-Márquez et al., 2016b) in comparison to the estimated size of each of these taxa (Weishampel and Young, 1996; Prieto-Márquez et al., 2016b), the author herein makes a tentative estimate of the size of this hadrosaur as being around 12-17 m in length. The implications of the large size attained by H. crassicauda and other Appalachian hadrosauroids are considered in the Discussion section.
Hypsibema crassicauda has been placed as a hadrosaurid and as dubious taxon within Hadrosauroidea (e.g., Weishampel and Young, 1996; Horner et al., 2004; Weishampel, 2006; Prieto-Márquez et al., 2016b). The author concurs with the sentiment of Baird and Horner (1979) that H. crassicauda represents a valid species among Hadrosauroidea based on its caudal vertebrae being laterally uncompressed. In any case, the vertebrae assigned to H. crassicauda represent a distinct morphotype of hadrosauroid in the Phoebus Landing fauna. Further discussion of Hypsibema as a distinct hadrosauroid may be found in the discussion.
The ceratopsian dinosaurs, though extremely rare on Appalachia, left a single indication of their presence within the Tar Heel non-avian dinosaur fauna. This record includes a derived leptoceratopsid left maxilla from sediments of the Tar Heel Formation of equivalent age to the Phoebus Landing Site (Longrich, 2016). Longrich (2016) noted that Appalachia belonging to a distinct palynofloral province may have implications for the adaptations found on the Tar Heel leptoceratopsian for the consumption of less-resistant plant matter.
The theropod dinosaurs also left behind an extensive record at Phoebus Landing. Hindlimb material comparable to Dryptosaurus aquilunguis was noted by Baird and Horner (1979). The femoral material compared to D. aquilunguis and figured by Baird and Horner (1979) may show an autapomorphic feature of this taxon. This is the presence of an ovoid fossa on the medial surface of the femur just above the distal condyles (Brusatte et al., 2011). More recently, Weishampel and Young (1996) also regarded Dryptosaurus aquilunguis as present at Phoebus Landing. This large tyrannosauroid theropod is known from a holotype specimen from the New Egypt Formation of New Jersey (e.g., Brusatte et al., 2011).
In addition to Dryptosaurus aquilunguis, material assigned to the tyrannosauroid dinosaur Appalachiosaurus montgomeriensis was reported from Stokes Quarry by Schwimmer et al. (2015). Some of these elements may be from juvenile individuals (Schwimmer et al., 2015). Therefore, two large tyrannosauroid dinosaurs were present in the Tar Heel-Coachman non-avian dinosaur fauna. Adults of both of these species probably measured from 6-9 m in length, comparable in size to the crocodylian Deinosuchus rugosus (Schwimmer, 1997; Schwimmer, 2002). An address on competition between large tyrannosauroids and D. rugosus is made in the Discussion section.
Small carnivorous theropod dinosaurs are represented by at least two taxa in the Upper Tar Heel-Coachman non-avian dinosaur fauna. Schwimmer et al. (2015) identified the dromaeosaurid Saurornitholestes langstoni from diagnostic teeth and a pedal ungual. These 1.8 m long dromaeosaurids (Currie and Koppelhus, 2005) were small predators in their environment. Additionally, two teeth were noted by Schwimmer et al. (2015) to be distinct from the Coachman Saurornitholestes material and similar to those of the western species Troodon and Dromaeosaurus, though they noted that the teeth are not sufficient to affirm the presence of either of the two taxa in the Coachman Formation. Additional teeth from small theropods have been uncovered in North Carolina (Weishampel and Young, 1996).
Remains assigned to ?Ornithomimus sp. and indeterminate ornithomimosaurs have been recovered from Phoebus Landing and Stokes Quarry, respectively (Miller, 1967; Baird and Horner, 1979; Schwimmer et al., 2015). These medium-sized feathered dinosaurs represent an important addition to the theropod diversity of the Tar Heel-Coachman non-avian dinosaur fauna.
Additionally, Schwimmer et al. (2015) mentioned “bird-like” limb elements which they assigned to Maniraptora indet. These remains are indicative of the presence of small maniraptorans within the Tar Heel-Coachman non-avian dinosaur fauna. A small, pathological metatarsal was also assigned to Theropoda indet. by Schwimmer et al. (2015) from Stokes Quarry. Thus, a population of small, maniraptoran theropods likely existed among the Upper Tar Heel-Coachman Formation ecosystem.
Bladen Formation. The Bladen Formation also preserves a record of dinosaurs from the Carolinas, but from the late Campanian. This record includes the remains of indeterminate dromaeosaurids, indeterminate tyrannosaurs, Ornithomimus sp. and indeterminate hadrosauroids (Crane, 2011). This fauna, along with that of the Donoho Creek Formation below, indicates that a fairly homogeneous dinosaur fauna persisted in the Carolinas through the Campanian.
Donoho Creek Formation. The Donoho Creek Formation of the Carolinas is slightly younger in age than the Coachman Formation, dating to the late Campanian (e.g., Schwimmer et al., 2015). This formation has not preserved as diverse of a dinosaur fauna as earlier Late Cretaceous sediments in the Carolinas. However, Schwimmer et al. (2015) reported a diverse theropod assemblage from the Donoho Creek Formation including the large tyrannosauroid Appalachiosaurus montgomeriensis, the dromaeosaurid Saurornitholestes langstoni, and indeterminate ornithomimosaurians.
Indeterminate hadrosaurid material has been reported from the Donoho Creek non-avian dinosaur fauna (Weishampel and Young, 1996; Weishampel et al., 2004; Schwimmer et al., 2015), including from skull elements (Schwimmer et al., 2015). The presence of the same two theropod taxa (A. montgomeriensis and S. langstoni) in the Coachman and Donoho Creek formations suggests that theropod faunas remained somewhat homogeneous throughout the Campanian in the Carolinas.
Merchantville Formation. The Merchantville Formation of the Atlantic Coastal Plain has preserved at least three different species of dinosaurs representing three distinct clades. Hadrosaurus foulkii is present in this formation (Gallagher, 1993; Weishampel and Young, 1996; Weishampel et al., 2004). One specimen from this unit, YPM VPPU.021795, represents an indeterminate tyrannosauroid distinct from either Appalachiosaurus or Dryptosaurus (Brownstein, 2017b). This unit is early to middle Campanian in age (e.g., Miller et al., 2004).
Woodbury Formation. Like the Merchantville Formation, the Woodbury Formation is middle Campanian in age and has preserved the remains of only Hadrosaurus foulkii (e.g., Prieto-Márquez et al., 2006; Prieto-Márquez, 2011). This medium-sized basal hadrosaurid dinosaur represents, along with the Alabama taxon Eotrachodon orientalis, an important discovery from Appalachia in regards to the evolution of hadrosaurs due to its placement as a basal hadrosaurid outside Saurolophidae (Prieto-Márquez et al., 2016a; Prieto-Márquez et al., 2016b). In addition to the holotype of Hadrosaurus foulkii, a historically important specimen described by paleontologist Joseph Leidy (Leidy, 1858) and the first somewhat complete skeleton of a dinosaur known from the Americas (Prieto-Márquez et al., 2006), hindlimb material originally assigned as the holotype of the taxon “Ornithotarsus immanis” by Cope (1869) has been more recently assigned to Hadrosaurus foulkii (Baird and Horner, 1977; Weishampel and Young, 1996). Gallagher (1997) remarked on the huge size of these remains, noting that a truly large hadrosaurid was present in the Upper Cretaceous of New Jersey. Weishampel and Young (1996) gave a length estimate of Hadrosaurus foulkii at 10 m, though the individual to which the holotype of “Ornithotarsus” immanis pertains may have approached 12 or more meters in length. Additionally, a large partial femur discussed by Gallagher (1997) from the Matawan Group (which includes the Woodbury Formation) (e.g., Gallagher, 1997) may also belong to a large individual of H. foulkii, though the extremely poor condition of this bone (Gallagher, 1997) means that any assignment to a specific taxon is nearly or completely impossible. Another occurrence of large hadrosaur possibly from the Woodbury Formation is based on the single pedal phalanx of a hadrosaur assigned to “Ornithotarsus” (=Hadrosaurus foulkii) by Edward Drinker Cope, which is a third larger than the corresponding element in the holotype of H. foulkii (Weishampel and Young, 1996).
Marshalltown Formation. The most extensive non-avian dinosaur fauna from the Campanian of New Jersey and also Delaware comes from the early late Campanian Marshalltown Formation (Sugarman et al., 1995; Miller et al., 2004) consisting of fine, quartz-rich glauconite clays (Olsson, 1988; Weishampel and Young, 1996). This dinosaur fauna consists of at least two different hadrosaur taxa, a large tyrannosauroid dinosaur, ornithomimosaurs, dromaeosaurids and nodosaurids (Lauginiger, 1984; Grandstaff et al., 1992; Gallagher, 1993; Weishampel and Young, 1996; Weishampel et al., 2004; Denton et al., 2011; pers. obs.). The Ellisdale fossil site of New Jersey in particular has produced an array of fossils assignable to at least three different species of non-avian dinosaur (e.g., Grandstaff et al., 1992; Gallagher, 1993; Weishampel and Young, 1996).
Hadrosauroid dinosaurs are represented in the Marshalltown Formation by remains assigned to the gigantic hadrosauroid Hypsibema crassicauda and the hadrosaurid Hadrosaurus sp. from the Ellisdale site (Grandstaff et al., 1992; Weishampel and Young, 1996), Hadrosaurus (including H. foukii) remains from Marshalltown exposures elsewhere in New Jersey (Gallagher, 1993; Weishampel and Young, 1996, Weishampel, 2006), and indeterminate remains from both New Jersey and Delaware (Gallagher, 1993; Weishampel and Young, 1996; Weishampel et al., 2004). Additionally, indeterminate ornithopods have been reported from the Ellisdale site (Grandstaff et al., 1992).
Nodosaurids are also present in the Marshalltown Formation (Gallagher, 1993; Weishampel and Young, 1996; Weishampel et al., 2004). Though they are known only by indeterminate remains, they represent the earliest record of nodosaurids from New Jersey and the possibility remains that they correspond to an unnamed taxon or taxa.
Theropods left a diverse fauna within the Marshalltown Formation. Dryptosaurus (sometimes assigned to D. aquilunguis) was reported from the Marshalltown by Grandstaff et al. (1992), Gallagher (1993), Weishampel and Young (1996), and Weishampel et al. (2004). Ornithomimosaur specimens, including material assigned to Ornithomimus (=“Coelosaurus”) have also been collected from the Marshalltown Formation (Gallagher, 1993; Weishampel and Young, 1996; Weishampel et al., 2004; pers. obs.). Teeth referable to indeterminate dromaeosaurids are also known from the Marshalltown Formation (Kiernan and Schwimmer, 2004; Denton et al., 2011; pers. obs.), increasing the carnivorous theropod diversity of this late Campanian unit. In addition, Grandstaff et al. (1992) reported the remains of indeterminate theropods from the Ellisdale site.
Mount Laurel/Wenonah Formations. The Mount Laurel/Wenonah Formations are latest Campanian-early Maastrichtian units from New Jersey (e.g., Gallagher, 1993) that preserve a variety of dinosaur taxa. Dryptosaurus sp. has been reported from the formations, as have indeterminate ornithomimosaurs (Weishampel et al., 2004). Additionally, material assigned to Hadrosaurus sp. has been collected from the formations along with bones from indeterminate hadrosaurids (Gallagher et al., 1993). Overall, this fauna is similar in composition to that of the Marshalltown Formation.
Coon Creek and Ripley Formations. The Coon Creek and Ripley Formations represent late Campanian deposits that are equivalent in age (e.g., Ebersole and King, 2011; Ebersole, personal commun., 2016) and have yielded a non-avian dinosaur fauna consistent with faunas from earlier in the Campanian. Ebersole and King (2011) listed hadrosaurid remains as coming from the Ripley and Coon Creek Formations. Of note is a hadrosaurid skull which was recovered from the Coon Creek Formation in Tennessee (Ebersole and King, 2011). Additionally, the partial skeletons of multiple ornithopods were noted to be recovered from the Ripley Formation (Ebersole and King, 2011).
Kanguk Formation. This Campanian/Maastrichtian age deposit lies to the north of Nunavut, Canada, and has preserved a fairly diverse assemblage of dinosaurs (e.g. Gangloff, 2012; Vavrek et al., 2014). This assemblage includes a lambeosaurine and possibly another type of hadrosaurid as well as a tyrannosauroid dinosaur (Gangloff, 2012). The lambeosaurines of the Kanguk are possibly the earliest known from the continent of Appalachia, and this fauna is the only one known from the far north of the landmass.
Maastrichtian
Navesink Formation. The Maastrichtian Navesink Formation is mainly composed of glauconitic clays (e.g., Sugarman et al., 1995; Kennedy et al., 2000; Miller et al., 2004) and has preserved the best known non-avian dinosaur fauna from the Maastrichtian of Appalachia. This extensive fauna includes a diverse assemblage of theropods, including an ornithomimosaur and two possibly distinct species of tyrannosauroid, a nodosaurid, a basal hadrosaurid, and lambeosaurines.
The hadrosaurid species found within the Navesink Formation include the medium-sized Hadrosaurus “cavatus” (=foulkii), an unnamed genus of very small hadrosaur (“Hadrosaurus” minor) currently considered a hadrosaurid of undetermined affinities, remains originally designated as the holotype of Hadrosaurus minor with the possibility that they represent juvenile remains, and an indeterminate species of lambeosaurine dinosaur (Colbert, 1948; Gallagher, 1993; Gallagher, 1997; Prieto-Márquez et al., 2006). The original specimen of Hadrosaurus minor consisted of dorsal vertebrae, and has been rendered a nomen dubium (Horner et al., 2004). Thus, the Navesink specimen described by Colbert (1948) has been referred to as “Hadrosaurus” minor (e.g., Weishampel et al., 2004; Weishampel, 2006). While Baird and Horner (1977) suggested the Navesink “Hadrosaurus” minor specimen was closely related to the saurolophine Edmontosaurus from the American west, more recent studies have suggested the taxon to be an unresolved taxon within “Hadrosaurinae” (=Saurolophinae) and as a hadrosaurid of uncertain affinities (Horner et al., 2004; Prieto-Márquez et al., 2006). Prieto-Márquez et al. (2006) noted that referral of “H.” minor to Edmontosaurus would be equivocal. Furthermore, the presence of multiple species of basal hadrosaurids and non-hadrosauroids on Appalachia suggests a higher likelihood of “Hadrosaurus” minor being of a basal phylogenetic position among the hadrosaurs.
In addition to the hadrosaurids, nodosaurid remains have also been recovered from the Navesink Formation. This clade of armored dinosaurs is represented by a single vertebra (e.g., Gallagher, 1993; Weishampel et al., 2004; Weishampel, 2006). As nodosaurid remains have been recovered from the somewhat older Campanian Marshalltown Formation in New Jersey, the possibility that the Navesink vertebrae represents a distinct taxon from the Marshalltown animal is certainly possible. Regardless, this vertebrae certainly shows that another distinct group of herbivorous dinosaurs was present alongside the hadrosaurids in the Navesink ecosystem.
Theropods also left a somewhat diverse fauna behind in the Navesink Formation. The tyrannosauroids are represented by remains attributable to Dryptosaurus aquilunguis (Weishampel and Young, 1996; Weishampel, 2006), a tibia, AMNH 2550, of an unnamed tyrannosauroid (=“Laelaps” macropus) (e.g., Holtz, 2004; Weishampel et al., 2004; pers. obs.), and indeterminate material, including a taxon named Diplotomodon horrificus known from a single tooth (e.g., Weishampel and Young, 1996; Holtz, 2004; Weishampel et al., 2004; Weishampel, 2006). Additionally, the holotype of “Ornithomimus” antiquus, which has most recently been regarded as a distinct taxon of ornithomimid dinosaur, is known from the Navesink (e.g., Leidy, 1865; Weishampel and Young, 1996; Makovicky et al., 2004; Weishampel et al., 2004; Weishampel, 2006; Brusatte et al., 2012).
The Navesink Formation was the only one of the few Maastrichtian Appalachian dinosaur-bearing formations included in statistical analyses herein, being compared to the Hell Creek, Lance, Horseshoe Canyon, and Javelina Formations. The Navesink was found by Miller et al. (2004) to be approximately 69-67 million years old (early to middle Maastrichtian), whereas the dinosaur fauna of the Horseshoe Canyon Formation is early and those of the Hell Creek, Lance, and Javelina Formations are late Maastrichtian in age (e.g., Weishampel et al., 2004). The results of the statistical analyses performed on these faunas may be found in Tables 27-31 of Appendix 1.
Severn Formation. The Maastrichtian (~70.7 Ma) (Hazel et al., 1984; Baird, 1986) Severn Formation of Maryland has also produced an important record of dinosaurs. This record includes the partial femur of an ornithomimosaur assigned by Baird (1986) to “Ornithomimus” antiquus, an ornithomimosaur pedal phalanx (Hartstein et al., 1986), and indeterminate hadrosauroid limb portions and partial vertebrae (e.g., Baird, 1986; Hartstein et al., 1986).
New Egypt Formation. The New Egypt Formation preserves the most complete late Maastrichtian dinosaur fauna from the eastern United States. The dinosaur remains retrieved from this formation include the holotype of the large tyrannosauroid dinosaur Dryptosaurus aquilunguis (e.g., Brusatte et al., 2011) and are housed in the AMNH FARB and ANSP collections. This partial skeleton is one of the most complete theropod dinosaur skeletons known from the Cretaceous of eastern North America (e.g., Schwimmer, 1997; Brusatte et al., 2011). One of the most intriguing features of D. aquilunguis are its large hands and massive manual ungual. The hadrosaurid dinosaurs from the New Egypt Formation include lambeosaurines, the dubious taxon Hadrosaurus minor, and indeterminate forms (e.g., Colbert, 1948; Weishampel and Young, 1996; Weishampel et al., 2004; Prieto-Márquez et al., 2006; Weishampel, 2006).
Kingstree Formation Equivalent. This unit is late Maastrichtian in age and has preserved the vertebra of an indeterminate theropod dinosaur (Schwimmer et al., 2015).
Prairie Bluff Formation. The late Maastrichtian Prairie Bluff Formation has preserved the vertebra of an indeterminate hadrosaur (Ebersole and King, 2011; George Phillips, personal commun., 2015).
Owl Creek Formation. The Owl Creek Formation of late Maastrichtian age has preserved an extremely important record of a single type of non-avian dinosaur. This is the single tooth of a possibly chasmosaurine ceratopsid (Farke and Phillips, 2017).
RESULTS
Aptian Dinosaur Faunas. Because the Arundel Clay has been dated to the upper Aptian to Albian (e.g., Kranz, 1998; Lipka et al., 2006), it was compared with two well-known upper Aptian to Albian units of the western United States: the Ruby Ranch Member of the Cedar Mountain Formation (e.g., Kirkland et al., 1999; Mori, 2009) and the Cloverly Formation (e.g., Chen and Lubin, 1997). The age of the Cloverly Formation is not well-constrained, but several studies regarding Ostrom’s (1970) units V-VII have found an Aptian-Albian age for those sediments with dates that range from about 113-108 Ma (Burton et al., 2006; Zaleha, 2006; Farke et al., 2014). However, D’Emic and Britt (2012) found a much younger age of about 103 Ma for sediments corresponding to unit VI or VII of Ostrom (1970) (Farke et al., 2014). The age of the Ruby Ranch member is also relatively unconstrained, with some estimates placing the unit from 120-108 Ma and others at around 104 Ma, overall corresponding to the Aptian and Albian (Mori, 2009; Chure et al., 2010). Units V-VII of the Cloverly Formation represent fluvial to overbank and lacustrine environments (e.g., Ostrom, 1970; May, 1992), whereas the environment represented by the Ruby Ranch Member was a semi-arid one punctuated by low-sinuosity rivers and ephemeral ponds (e.g., Harris, 1980; Kirkland et al., 1999; Kirkland and Madsen, 2006). Though depositional differences between these formations certainly hinder precise statistical analysis by causing possible biases, the author emphasizes that Aptian dinosaur faunas containing several taxa are lacking in North America save for very few units (e.g., Weishampel et al., 2004).
Analytic comparisons of the Arundel Clay fauna and those of these two units (Table 1 of Appendix 1) show the Arundel fauna is somewhat similar in composition to that of the Ruby Ranch in containing Deinonychus, Tenontosaurus, Acrocanthosaurus, or a similar taxon, a large nodosaurid, and a large titanosauriform (e.g., Leidy, 1865; Marsh, 1888; Kranz, 1996; Weishampel, 2006; Ostrom, 1970; Weishampel et al., 2004; Mori, 2009; Woodruff, 2012; D’Emic and Foreman, 2012; Mannion et al., 2013; Oreska et al., 2013). Notably, ornithomimosaurs and neoceratopsians are absent from the Ruby Ranch Member, though they are present in both the Arundel and the Cloverly Formation along with a large nodosaurid (Sauropelta), Deinonychus, Acrocanthosaurus, Tenontosaurus, ornithomimosaurs, and titanosauriforms (Ostrom, 1969, 1970, 1976; Kranz, 1996; Chinnery et al., 1998; Makovicky and Sues, 1998; Weishampel et al., 2004; Gignac and Makovicky, 2010;Woodruff, 2012; d’Emic et al., 2012; Oreska et al., 2013; Farke et al., 2014; Brownstein, 2017a). Nevertheless, the Arundel ceratopsian material remains problematic (e.g., Farke et al., 2014), and so future work on the Arundel fauna will be needed to better comparisons between these formations. Statistical comparisons of these faunas show medium to medium-high similarity (50-70% Simpson Similarity index values, 0.5-0.6) on the family level between the Arundel and these two units, although the same comparisons yield low genus level similarity when measured with the Simpson similarity index (30-50%) and Jaccard coefficient (0.2-0.3) (Tables 2-5 of Appendix 1). The low genus-level similarity between the faunas is likely augmented because of the lack of specimens from the Arundel assignable to specific genera, as half the genera known from the Arundel (Deinonychus, Acrocanthosaurus, Tenontosaurus) are known from the Cloverly and Ruby Ranch faunas (Table 1 of Appendix 1). Acrocanthosaurus and Deinonychus specimens have also been collected from the Twin Mountain and Antlers formations of Texas and Oklahoma (Stovall and Langston, 1950; Cifelli, 1997; Harris, 1998; Currie and Carpenter, 2000; D’Emic et al., 2012), and in addition to the common occurrence of Tenontosaurus remains (e.g., Ostrom, 1970; Forster, 1984; Forster, 1990; Winkler et al., 1997; Weishampel et al., 2004), the ankylopollexian iguanodontians Hippodraco and Theiophytalia are present during the Aptian in the American west (Brill and Carpenter, 2006; McDonald et al., 2010). The lack of reported ornithomimosaur material from the Ruby Ranch member of the Cedar Mountain Formation is not regarded as significant, as the possible ornithomimosaur Nedcolbertia justinhofmanni is known from the slightly older Yellow Cat Member of the Cedar Mountain Formation (Brownstein, 2017a; Kirkland and Hunt-Foster, 2017).
Nodosaurids and titanosauriforms were also spread across North America during this time, with the latter also known from the Early Cretaceous of Texas (Sauroposeidon proteles, Astrophocaudia slaughteri, Cedarosaurus sp., and Titanosauriformes indet.) (e.g., Larkin, 1910; Langston, 1974; Cifelli, 1997; Wedel et al., 2000; Weishampel et al., 2004; D’Emic, 2013), from Utah (Brontomerus mcintoshi; Cedarosaurus weiskopfae) (Taylor et al., 2011), and from the Cloverly Formation of Montana and Wyoming (including Sauroposeidon) (Tidwell et al., 1999; Ostrom, 1970; Weishampel et al., 2004; Woodruff, 2012; D’Emic and Foreman, 2012; Mannion et al., 2013; Oreska et al., 2013). The nodosaurid taxon Sauropelta was present in the Aptian of the Little Sheep Mudstone of the Cloverly Formation in the western United States (e.g., Ostrom, 1970; Kirkland et al., 1997; Weishampel et al., 2004; Oreska et al., 2013). Additional fossils assigned to Sauropelta have been recovered from middle Cedar Mountain Formation (Jensen, 1984; Weishampel et al., 2004). Fossils of the nodosaurid taxa Hoplitosaurus sp. and Tatankacephalus cooneyorum have also been recovered from the middle Cedar Mountain Formation and the Cloverly Formation, respectively (Weishampel et al., 2004; Parsons and Parsons, 2009). Finally, the polacanthine Gastonia lorriemcwhinneyae was described in 2016 (Kinneer et al., 2016).
Though the lack of body fossils from the Patuxent facies and the Trinity Group in Arkansas warranted against statistical comparisons between the faunas of these and western units, several analytical similarities can be observed between them. Indeed, the Patuxent facies shares with the Cloverly Formation and other western units nodosaurids, small (= Zephyrosaurus-like) and large euornithopods, ornithomimosaurs (as does the Arkansas Trinity Group) (e.g., Quinn, 1973; Kirkland and Hunt-Foster, 2017), large theropods, and titanosauriforms (e.g., Ostrom, 1970; Weishampel et al., 2004; Kirkland et al., 1999; Mori et al., 2009; D’Emic, 2013) and thus supports the hypothesis that a relatively homogenous dinosaur fauna existed throughout land now within the United States during the Early Cretaceous.
Albian Dinosaur Faunas. The Dakota Formation is late Albian to early Cenomanian in age (e.g., Joeckel et al., 2004; Koch, 2007) and was compared with the similarly-aged Wayan, Blackleaf, and Willow Tank formations of the western United States (Varricchio et al., 2007; Bonde et al., 2012; Ullman et al., 2012; Krumenacker et al., 2017). The Wayan Formation of Idaho was found to be deposited between 101.8 and 95.5 Ma by Krumenacker (2010), who reviewed the formation’s stratigraphy. The Willow Tank Formation has been found to be between 98.5 and 98.1 million years old (e.g., Fleck, 1970; Troyer et al., 2006; Bonde et al., 2012), and the Blackleaf Formation has been dated to the late Albian and early Cenomanian (e.g., Dorr, 1985; Varricchio et al., 2007; Ullman et al., 2012). All of these western formations have been considered as overlapping in age (e.g., Bonde et al., 2012; figure 2 in Krumenacker et al., 2017). The Dakota Formation in southeastern Nebraska has been interpreted as representing fluvial to estuarine environments (e.g., Joeckal et al., 2004), whereas the Dakota Formation in Kansas has been interpreted as a coastal plain (e.g., Eaton, 1960), and the Blackleaf, Wayan, and Willow Tank formations all seemingly represent fluvial-deltaic environments (e.g., Kirkland et al., 1999; Bonde et al., 2008; Ullman et al., 2012; Krumenacker et al., 2017). Thus, these formations were compared due to their similar paleoenvironments and ages (Table 6 of Appendix 1). Although these formations are not as well-known as those from the Cenomanian (e.g., Krumenacker et al., 2017), the majority share the presence of iguanodonts and ankylosaurs. Genus level comparisons of these formations’ faunas yielded low values for both the Simpson similarity index and Jaccard coefficient, yet family level comparisons yielded a medium-high (50-70%; 0.5-0.7) Simpson similarity index value between the faunas of the Dakota and Wayan formations (Tables 7-10 of Appendix 1) and lower Simpson similarity index values between the Dakota and the other western faunas. The Simpson similarity index and Jaccard coefficient values found for comparisons of the Dakota with the Blackleaf and Willow Tank formations’ faunas were also low (Tables 7-10 of Appendix 1). Family-level comparisons between the Dakota and Paluxy formations yielded a 100% Simpson similarity index and a 0.6 Jaccard similarity index value.
Although the Paluxy is middle Albian in age (112.2-106 Ma) (Jacobs and Winkler, 1998; D’Emic, 2013) and the Dakota, Wayan, Blackleaf and Willow Tank formations were deposited in the late Albian to early Cenomanian (e.g., Liggett, 2005; Varricchio et al., 2007; Bonde et al., 2012; Krumenacker et al., 2017), the scarcity of dinosaur-bearing deposits from this time in North America and the relatively well-documented state of the Paluxy Formation fauna warranted statistical comparison between it and the faunas of the other aforementioned sedimentary units. Genus level similarity for both indices used was 0.0 between the Paluxy and all other formations, whereas family-level comparisons resulted in low Simpson similarity index and Jaccard coefficient values between the Paluxy fauna and those of all other formations save for the Dakota (Tables 7-10 of Appendix 1).
Because of the limited number and diversity of dinosaur taxa from the Paw Paw and Glen Rose Formations, statistical comparisons between these and other formations were not conducted. The Paw Paw formation was deposited during the Late Albian (e.g., Lee, 1996), thus being similar in age to the Dakota, Wayan, Blackleaf, and Willow Tank Formations (e.g., Liggett, 2005; Varricchio et al., 2007; Bonde et al., 2012; Ullman et al., 2012; Krumenacker et al., 2017). Like these other formations, the Paw Paw evinces that nodosaurids continued being widespread across North America during the Albian. The Glen Rose fauna further shows that large theropods, titanosauriforms, and Tenontosaurus were included in the Aptian-Albian fauna of North America.
Cenomanian Dinosaur Faunas. The non-avian dinosaur fauna of the deltaic plains (e.g., Main, 2005; Main, 2010; Main, 2013) represented by the Woodbine Formation included large carcharodontosaurs, dromaeosaurs, ornithomimosaurs, indeterminate theropods (including possible tyrannosauroids), hadrosauroids, and nodosaurids (e.g., Main, 2005, 2013). Possible allosauroid and carcharodontosaurid dinosaurs are known from the Mussentuchit Member of the Cedar Mountain Formation (Siats meekorum) (Zanno and Makovicky, 2013) and from the Turney Ranch Formation (Thayer and Ratkevich, 1995). Dromaeosaurids are also known from the Mussentuchit Member of the Cedar Mountain Formation (Garrison and Brinkman, 2007) and from the Blackleaf Formation (Ullman et al., 2012). The nodosaurid Animantarx caroljonesa is also known from the Mussentuchit Member of the Cedar Mountain Formation (Carpenter et al., 1999). Adding to similarities between the Woodbine Formation and the Mussentuchit Member of the Cedar Mountain Formation is the presence of the hadrosauroid dinosaur Eolambia in the latter unit (Kirkland et al., 1998). This taxon has been found as an outgroup to hadrosauridae behind Protohadros or as a sister taxon to P. byrdi in phylogenetic analyses (Prieto-Marquez and Norell, 2010; Wenhao and Godefroit, 2012), although another study found it to be a sister taxon of Probactrosaurus and Protohadros to be a more derived hadrosauroid (McDonald et al., 2012).
The Woodbine Formation was statistically compared to the Mussentuchit Member of the Cedar Mountain Formation of Utah and the Dunvegan Formation of western Canada (Tables 11-15). The Mussentuchit Member of the Cedar Mountain Formation is latest Albian to Cenomanian in age (~104-98 Ma) (e.g., Cifelli et al., 1997; Chure et al., 2010) and represents fluvial to deltaic environments (e.g., Kirkland et al., 1998; Carpenter et al., 1999; Kirkland et al., 1999; Garrison and Brinkman, 2007; McDonald et al., 2012; Ullman et al., 2012; Main, 2013; Zanno and Makovicky, 2013; Krumenacker et al., 2017), whereas the Dunvegan Formation represents a middle Cenomanian-age delta complex (e.g., Burns and Vavrek, 2014). The Lewisville Member of the Woodbine Formation, to which the Arlington Archosaur Site corresponds, is middle Cenomanian in age (~96-95 million years old) and preserves a coastal deltaic environment (e.g., Main, 2005; Main, 2013), making it closely comparable to the Dunvegan Formation and somewhat so to the Mussentuchit. The faunas of these units and the results of statistical analysis of the similarity of those faunas are listed in Tables 11-15 of Appendix 1.
Because the record of dinosaurs from the Raritan facies is so scant, no statistical comparisons were made between it and other Cenomanian units’ faunas. It is notable that the Raritan shares with the Mussentuchit, Wayan, and Blackleaf formations tyrannosauroid dinosaurs (e.g., Baird, 1989; Kirkland et al., 1998; Varricchio et al., 2007; Ullman et al., 2012; Zanno and Mackovicky, 2013). The size of the Raritan facies tyrannosauroid is relatively large compared to Cenomanian North American tyrannosauroids (pers. obs.). The dinosaur trackways of the Raritan are additionally important for being the only Late Cretaceous dinosaur tracks known east of the Mississippi (e.g., Baird, 1989).
Coniacian/Santonian Dinosaur Faunas. The Niobrara Chalk is late Coniacian to Santonian in age in the general area surrounding Hackleberry Creek (where Niobrarasaurus, “Heirosaurus”, indeterminate nodosaurids, and the caudal vertebrae of an indeterminate hadrosaurid have been found) and late Santonian in Logan County (where the holotype of Claosaurus was uncovered) (e.g., Carpenter et al., 1995; Weishampel et al., 2004; Everhart and Ewell, 2006; Prieto-Márquez et al., 2016a; Prieto-Márquez et al., 2016b). Thus, the Niobrara Formation is comparable in age to the Eutaw Formation, which has been dated to the Santonian (87-83 Ma) (e.g., Weishampel et al., 2004; Ebersole and King, 2011; Prieto-Márquez et al., 2016a; Prieto-Márquez et al., 2016b). Additionally, the Eutaw represents marginal marine to marine deposits and was formed during a sea transgression (Liu, 2009), somewhat comparable to the marine setting of the Niobrara Formation (e.g., Carpenter et al., 1995; Liggett, 2005; Ebersole and Ewell, 2006). Both of these formations share hadrosaurids, nodosaurids, and non-hadrosaurid hadrosauroids, whereas only hadrosaurids are observed in the latest Santonian (~84.5-83.5 Ma) Milk River Formation (thus overlapping with the Eutaw Formation entirely) (e.g., Payenberg et al., 2002; Weishampel et al., 2004; Ebersole and King, 2011; Prieto-Márquez et al., 2016a; Prieto-Márquez et al., 2016b). Additionally, the fauna of the Milk River Formation differs from those of the Eutaw and Niobrara, other Coniacian-Santonian, and Campanian Appalachian formations (e.g., Tables 17, 22 of Appendix 1; Carpenter et al., 1995; Ebersole and Ewell, 2006; Gallagher, 1993; Weishampel et al., 2004; Denton et al., 2011; Ebersole and King, 2011; Schwimmer et al., 2015) in containing pachycephalosaurids, centrosaurines, troodontids, and tyrannosaurids. Thus, it seems that major differences in the composition of dinosaur faunas emerged between Laramidia and Appalachia sometime during the “mid”-Cretaceous. Statistically, this hypothesis is supported by the low Simpson similarity index value at the family level (40%) found between the Eutaw and Milk River formations, the low Jaccard coefficient (0.2) value found from comparisons of Coniacian-Santonian Appalachian faunas (Eutaw Niobrara faunas) with that of the Milk River Formation, and the 100% Simpson similarity index value found between the Niobrara and Eutaw faunas at the family level (Tables 17-21 of Appendix 1). Unfortunately, the poor record of dinosaurs during the Turonian-Santonian of North America (e.g., Carpenter et al., 1995) hinders more precise estimates of when such faunas experienced vicariance. Indeed, all comparisons Coniacian-Santonian formations at the genus level yielded values of 0.0 for both metrics.
Though the Santonian McShan Formation preserves a heavily incomplete dinosaur fauna (e.g., Ebersole and King, 2011) and thus was not compared statistically to other Appalachian and Laramidian faunas, the presence of tyrannosauroids and hadrosauroids in its fauna supports the notion that Appalachian and Laramidian faunas had already differentiated appreciably during the Santonian.
Campanian Dinosaur Faunas. Among the several dinosaur-bearing units of the Campanian of Appalachia, the Mooreville Chalk, Blufftown and Coffee Sand formations and unnamed Missouri clay, the Tar Heel and Coachman formations, and the Marshalltown Formation were used in statistical comparisons with several western Campanian-age strata as well as with each other for the investigation herein of dinosaur provincialism on Appalachia. The Mooreville Chalk, Blufftown, and Coffee Sand formations and the unnamed Missouri clay all latest Santonian to middle Campanian in age (e.g., Ebersole, 2009; Ebersole and King, 2011; Prieto-Márquez et al., 2016a; Prieto-Márquez et al., 2016b), with the marine Mooreville Chalk being approximately 80-83 million years old (e.g., Prieto-Márquez et al., 2016b) and equivalent with the Blufftown and Coffee Sand formations (e.g., Schwimmer et al., 1993; Ebersole and King, 2011), and the apparently terrestrial (representative of an oxbow lake) (Fix and Darrough, 2004) Chronister site also apparently Campanian in age (e.g., Fix and Darrough, 2004; Ebersole, 2009). The fauna of these equivalent formations thus correspond in age to the 81-76 million year old Wahweap (Sampson et al., 2013a) and the Oldman (coeval with the Wahweap; e.g., Roberts et al., 2005; Gates et al., 2010) formations.
The dinosaur-bearing sites of the Tar Heel and Coachman formations, as noted previously, are middle Campanian in age, with the upper portion of the Tar Heel (Tar Heel II sequence) dated to be between 78.7-74.5 million years ago. This range overlaps somewhat with that of the Marshalltown Formation, which may be slightly younger at 75.7-71.2 million years of age (e.g., Miller et al., 2004; Self-Trail et al., 2004; Harris and Self-Trail, 2006; Schwimmer et al., 2015). Thus, the Marshalltown and Tar Heel-Coachman faunas overlap by at least 1.2 million years, the former overlapping with the main fossiliferous zones of the Dinosaur Park, Kaiparowits, upper Judith River, and lower Kirtland formations in age by approximately 1.5 million years and the latter by at least 2 million years (the main fossiliferous zones of these latter four units being between ~76-74 Ma; e.g., Eberth and Hamblin, 1993; Rogers et al., 1993; Horner et al., 2001; Eberth and Deino, 2005; Roberts et al., 2005; Hanson et al., 2006; Lucas et al., 2006; Gates et al., 2010). The Kaiparowits Formation has been dated to 76.6-74.4 Ma (Sampson et al., 2013a), the Dinosaur Park Formation to 76.9 and 75.8 Ma (e.g., Gates et al., 2010; Gates et al., 2012; Sampson et al., 2013a), the Hunter Wash Member (lower Kirtland Formation) to between 74.5-74.1 Ma (e.g., Gates et al., 2010), and the the Judith River Formation to between 80-75 Ma (e.g., Sullivan and Lucas, 2006). The Dinosaur Park, Judith River, Oldman, and Kirtland formations are apparently coastal plain deposits, whereas the Kaiparowits and Wahweap are apparently representative of alluvial plains (e.g., Roberts et al., 2005; Gates et al., 2010). Thus, even though the Kaiparowits Formation and the formation coeval to it correspond well to the Tar Heel, Coachman, and Marshalltown formations and the Wahweap and Oldman correspond partially to the Mooreville Chalk, Blufftown, Coffee Sand, and unnamed Missouri clay unit temporally, depositional bias may indeed be somewhat detrimental to statistical analysis. Nevertheless, the author notes that at least some eastern sites included in the formations analyzed represent coastal or inland environments (e.g., the marginal-marine Phoebus Landing site of the Tar Heel Formation, the storm-surge deposit of an inland biota at the Ellisdale site of the Marshalltown Formation, or the Chronister site of the unnamed Missouri clay unit) (e.g., Fix and Darrough, 2004; Self-Trail et al., 2004; Denton et al., 2011). Key sites such as the three just mentioned are critical for minimizing depositional bias in statistical analyses of biogeography such as those presented herein. Genus-level comparisons yielded low Jaccard coefficient and Simpson similarity index values for all faunas except for comparisons for calculation of the latter among those of the Oldman, Judith River, and Dinosaur Park formations and between the Marshalltown and Tar Heel and Coachman faunas (50-60% values for the former comparisons; 70% value for the latter) (Tables 23, 25 of Appendix 1). On the family level, high Simpson similarity index values were found from comparisons among Appalachian faunas (80-90%) and from comparisons among Laramidian faunas (80-100%) (Table 24 of Appendix 1). However, comparisons at the family level between Appalachian and Laramidian faunas yielded only low to moderately high (20-70%) values (Table 24 of Appendix 1). Family-level comparisons between these Campanian faunas for calculation of the Jaccard coefficient yielded a stronger version of this pattern of high similarity between faunas of the same landmass and low similarity between faunas of different landmasses. Comparisons between the Marshalltown, Tar Heel and Coachman, and Mooreville Chalk, Blufftown, and Coffee Sand formations and unnamed Missouri unit faunas yielded Jaccard coefficient values of 0.6-0.7, whereas comparisons between Laramidian faunas at the family level yielded Jaccard coefficient values of 0.53-0.86. Comparisons between western and eastern faunas at the family level, however, yielded Jaccard coefficient values of 0.3 or less. Even though the Oldman, Wahweap, and Mooreville Chalk, Blufftown, and Coffee Sand formations and unnamed Missouri clay unit faunas are approximately 7 million years older or less than those of the Tar Heel and Coachman formations, Marshalltown, lower Kirtland, upper Judith River, Kaiparowits, and Dinosaur Park formations, comparisons between them and these younger faunas generally yielded similar Simpson similarity index and Jaccard coefficient values to those found between formations of the same age (Tables 23-26 of Appendix 1). Indeed, both the aforementioned Appalachian faunas used in statistical comparisons and those of the Merchantville, Woodbury, Donoho Creek, Bladen, and Mt. Laurel/Wenonah formations show that Campanian Appalachian dinosaur faunas differed from Laramidian ones in containing hadrosauroids, basal hadrosaurids, and “indetermediate”-grade (Brusatte et al., 2011) tyrannosauroids as well as lacking ceratopsids, tyrannosaurids, saurolophines, lambeosaurines, pachycephalosaurids, therizinosaurs, troodontids, and ankylosaurids (e.g., Baird and Horner, 1979; Gallagher, 1993; Weishampel and Young, 1996; Gallagher, 1997; Schwimmer, 1997, 2002; Fix and Darrough, 2004; Carr et al., 2005; Brusatte et al., 2011; Denton et al., 2011; Ebersole and King, 2011; Schwimmer et al., 2015; Prieto-Márquez et al., 2016a, 2016b).
Maastrichtian Dinosaur Faunas. The Navesink Formation fauna was the only Maastrichtian Appalachian one statistically compared to western faunas of the time. The Maastrichtian-age portion of the Navesink Formation has been dated to between 69 and 67 Ma (e.g., Miller et al. 2004), whereas the Javelina Formation has been dated to 70 to 66.5 Ma (Woodward and Lehman, 2005) and the Lance and Hell Creek formations to the middle-late Maastrichtian (~69-66 Ma) generally (e.g., Johnson et al., 2002; Weishampel et al., 2004). Genus-level comparisons between the Navesink fauna and those of western formations all yielded a value of 0.0 for the Jaccard coefficient and Simpson similarity index, whereas family-level comparisons yielded medium Simpson similarity index values (40-60%) and low Jaccard coefficient values (0.2) for the same comparisons. Analytically, the faunas of both the Navesink and New Egypt formations show that Appalachia still harbored relict forms that included non-tyrannosaurid tyrannosauroids (e.g., Dryptosaurus) (e.g., Brusatte et al., 2011). However, the presence of lambeosaurines in the late Maastrichtian New Egypt Formation (which overlies the Navesink Formation) (e.g., Gallagher, 1993; Miller et al., 2004; Brusatte et al., 2011) and ceratopsids in the late Maastrichtian Owl Creek Formation (Farke and Phillips, 2017) show that several clades of dinosaur found only in the western United States during the Campanian and earlier (e.g., Schwimmer, 1997) had dispersed into the eastern United States.
DISCUSSION
Issues Regarding the Biogeographic Analysis of Cretaceous Appalachian Dinosaurs
Though the fossil record from Cretaceous Appalachian non-avian dinosaurs is important, as demonstrated in this paper, several statistical issues are encountered when trying to measure faunal similarity between Appalachian dinosaur faunas and others by means of the Jaccard coefficient and Simpson similarity index. At the genus level, calculations may not truly account for the diversity of dinosaur genera in Appalachian faunas that would be found with categorial examination. For example, the genus level calculation of an Appalachian fauna containing “Dromaeosauridae indet.” and no specific dromaeosaurid taxa for both the Simpson similarity index and Jaccard coefficient would not take into account the presence of a genus or of multiple genera of dromaeosaur. Likewise, the understudied nature of Appalachian dinosaur faunas also may bias calculations on the family level, as certain obscure genera may compose unknown families or subfamilies within a level. One example would be the gigantic hadrosauroids with massively constructed tails of Appalachia (Hypsibema crassicauda, Parrosaurus missouriensis). In future years, these animals may turn out to form a distinct family within or outside Hadrosauridae, but are simply referred to herein as hadrosauroids of uncertain phylogenetic placement.
Analytic comparison of Appalachian non-avian dinosaur faunas to the western faunas in Appendix 1 and to other Cretaceous faunas listed in Weishampel et al. (2004) also shows that even the most speciose Appalachian non-avian dinosaur faunas from the Coniacian to Maastrichtian are somewhat lacking in the number of represented dinosaur groups and known species compared to faunas of similar environments and times (i.e., when compared to the Nemegt Formation of early Maastrichtian Mongolia) (e.g., Weishampel et al., 2004; Appendix 1). However, there may be both taphonomic and ecological reasons for this disparity, as discussed above and below.
The Turonian of Appalachia is clearly poorly known. During the Aptian to Cenomanian, however, eastern North American faunas have a relatively similar number and composition of represented groups to western ones, and so the results from the aforementioned interval may be considered as more accurate.
One sampling bias among Appalachian faunas themselves also occurs temporally, as certain stages of the Cretaceous are currently known to be recorded by more units in eastern North America than other stages. This is true especially in the cases of the Coniacian to Campanian stages of the Late Cretaceous, when a variety of relatively species-rich faunas are known from all across the landmass of Appalachia (e.g., Baird and Horner, 1979; Gallagher, 1993; Carpenter et al., 1995; Denton et al., 2011; Ebersole and King, 2011; Schwimmer et al., 2015; Schwimmer, 2016). Contrast such a record with that from the Cenomanian, when the Woodbine Formation gives the only record of dinosaurs from the aforementioned eastern landmass (e.g., Main, 2013). As such, it is important to note that the Cenomanian, Turonian, and Maastrichtian faunas catalogued in this paper are lacking in the number and species-richness of stratigraphic units compared to the Coniacian-Campanian ones. In the case of the Cenomanian dinosaur faunas known from Appalachia, the small number of species may be contributed to the lack of vertebrate fossils known from the Potomac Formation (e.g., Baird, 1989; Gallagher, 1997; Main, 2013) and the lack of dinosaur-bearing Turonian strata in the eastern portion of North America. Finally, though the Maastrichtian faunas hail from across the eastern United States, only the Navesink Formation has produced more than three clades of dinosaur (e.g., Gallagher, 1993). Future work will hopefully reveal the dinosaur faunas of other Maastrichtian localities pertaining to Appalachia.
Importantly, sampling biases clearly exist between the much better sampled western North American (e.g., Kirkland et al., 1998; Roberts et al., 2005; Jinnah et al., 2009; Gates et al., 2010) and poorly sampled Appalachian ones that are mostly representative of marine deposits (e.g., Schwimmer, 1997). Future work will be needed to reduce this bias. Additionally, the comparatively slow rate at which new dinosaur taxa are named from Appalachia means that Appalachian faunas are only somewhat represented in the known fossil record. However, because the author extensively reviewed the fauna of Appalachian dinosaur faunas statistically compared herein through review of the literature and personal observation, the comparisons herein represent currently the most complete glimpse at Appalachian dinosaur faunas. Specific discussion regarding the presence of dinosaur genera in certain provinces may be found below.
As Late Cretaceous dinosaurs have been excavated from sediments in New Jersey and Delaware since the 19th century and dinosaur specimens have only been collected, identified, and described in detail from southeastern North America rather recently (Langston, 1960; Schwimmer, 1993; Weishampel and Young, 1996; Carr et al., 2005; Ebersole and King, 2011), sampling biases certainly exist among Campanian-Maastrichtian Appalachian dinosaur faunas of these areas and needed to be taken into account for analysis of dinosaur provincialism on the landmass. Though it has been critiqued as a method of comparing species richness (e.g., Alroy, 2000, 2010a, 2010b, 2010c; Alroy et al., 2008), the method of rarefaction (Sanders, 1968) was used to assess the comparability of Appalachian Campanian faunas analyzed herein. The results of rarefaction on the most well-known (in terms of numbers of specimens) Appalachian faunas also allows for insight into the different sampling between each formation’s fauna. The Conacian-Santonian faunas that were used in rarefaction were those of the Eutaw Formation and Niobrara Formation. The results of rarefaction of these assemblages yielded an estimated number of clades of five for the Eutaw Formation when rarefied using the total number of specimens from the Niobrara. This result shows the unequal nature of sampling between the two formations, a discrepancy already present when comparing the number of specimens from each. This is relatively unsurprising, as the dinosaur fauna from the Niobrara Formation consists entirely of animals which were washed out into 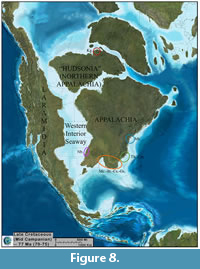 the Western Interior Seaway (e.g., Carpenter et al., 1995). Nevertheless, it may be assumed that the dinosaurs of the Niobrara correspond to the coastal areas of western Appalachia, as the portion of the landmass west of the Appalachians was essentially flat (e.g., Sampson et al., 2010a; Figure 8.1), and thus the rivers and creeks of the area probably could not generate the force needed to wash the remains of the dinosaurs found in the Niobrara Formation out to sea from far inland. Additionally, previous studies have found Claosaurus to be closer to Appalachian hadrosauroids and basal hadrosaurids (i.e., Lophorhothon, Hadrosaurus, Eotrachodon) than Laramidian forms of similar age (i.e., Jeyawati rugoculus) (Prieto-Márquez et al., 2016a). Finally, the localities where the type specimens of “Heirosaurus” and Niobrarasaurus were collected are in the middle of Kansas (e.g., Carpenter et al., 1995), which would have been much closer to the western shore of Appalachia than the eastern shore of Laramidia during the Santonian (e.g., figure 6 in Schröder-Adams, 2014). As such, the author follows previous studies (e.g., Prieto-Márquez et al., 2016b) in regarding the Niobrara Formation dinosaur fauna as Appalachian in origin.
the Western Interior Seaway (e.g., Carpenter et al., 1995). Nevertheless, it may be assumed that the dinosaurs of the Niobrara correspond to the coastal areas of western Appalachia, as the portion of the landmass west of the Appalachians was essentially flat (e.g., Sampson et al., 2010a; Figure 8.1), and thus the rivers and creeks of the area probably could not generate the force needed to wash the remains of the dinosaurs found in the Niobrara Formation out to sea from far inland. Additionally, previous studies have found Claosaurus to be closer to Appalachian hadrosauroids and basal hadrosaurids (i.e., Lophorhothon, Hadrosaurus, Eotrachodon) than Laramidian forms of similar age (i.e., Jeyawati rugoculus) (Prieto-Márquez et al., 2016a). Finally, the localities where the type specimens of “Heirosaurus” and Niobrarasaurus were collected are in the middle of Kansas (e.g., Carpenter et al., 1995), which would have been much closer to the western shore of Appalachia than the eastern shore of Laramidia during the Santonian (e.g., figure 6 in Schröder-Adams, 2014). As such, the author follows previous studies (e.g., Prieto-Márquez et al., 2016b) in regarding the Niobrara Formation dinosaur fauna as Appalachian in origin.
The results of rarefaction on the most well-known (in terms of specimens) faunas of the Campanian of Appalachia showed that there was little difference between different assemblages in terms of sampling. The number of specimens from the Marshalltown Formation was the smallest out of the number of specimens from each of the Campanian Formations whose faunas were rarefied (these were the fauna of the Mooreville Chalk, Blufftown, and Coffee Sand formations and unnamed Missouri clay unit, that of the Tar Heel and Coachman formations, and the Marshalltown Formation), and thus it was used as the subsample value (n). The resulting estimated number of clades for the Mooreville Chalk, Blufftown, and Coffee Sand formations and unnamed Missouri clay unit was seven, one less than that of the faunal list from the equivalent units. The rarefaction of the Tar Heel and Coachman formations using this value produced an estimated number of nine clades of dinosaur, the same number represented from the formation in the faunal list. Thus, these three groups of units may be considered significantly comparable.
The Appalachian Dinosaur Fauna
The landmass of Appalachia harbored a diverse dinosaur fauna from the Aptian to the Maastrichtian stages of the Cretaceous. The data compiled in Tables 1-5 of Appendix 1 suggest that North American faunas of the Aptian were differentiated at the genus level to an extent, but a distinct east-west division was not present. A summary of the stage-by-stage results of the biogeographic analyses performed can be found in Table 1. At the family level, the biogeographic data compiled from Aptian North American dinosaur faunas suggest that even though genera were different across regions, the same clades were present across the continent. This Aptian North American fauna would therefore have included basal ornithomimosaurs, the carcharodontosaurid Acrocanthosaurus akotensis, dromaeosaurids (especially the taxon Deinonychus), nodosaurids, iguanodontians, basal neoceratopsian dinosaurs, possibly the orodromine dinosaur Zephyrosaurus, and titanosauriform sauropods. Albian-Cenomanian North American faunas include nodosaurids, dromaeosaurids, macronarian sauropods, carcharodontosaurids, and basal hadrosauroids. The absence of therizinosaurs and oviraptorosaurs from eastern North America during this time is considered ambiguous, as a large portion of the Arundel facies theropod material remains to be formally studied. During the Albian, orodromine dinosaurs are no longer found in Appalachian dinosaur faunas (Table 6 of Appendix 1), though orodromines and other small ornithopod dinosaurs are known from all three western North American dinosaur faunas from the same time (Table 6 of Appendix 1). Nevertheless, western and eastern faunas still share nodosaurids, iguanodontians, and titanosauriformes (Table 6 of Appendix 1). The lack of faunal data from the Dakota Formation and Paluxy Formation may have caused the calculation of inflated values for their Simpson similarity index and Jaccard coefficient. Among the better-known western faunas, these values are all fairly high (Tables 7-10 of Appendix 1), suggesting relative faunal homogeneity in the western portion of North America.
The Cenomanian Woodbine Formation of Appalachia, when compared to the Mussentuchit Member of the Cedar Mountain Formation, is clearly differentiated at the genus level but more similar at the family level to the latter fauna in both the calculated values for the Simpson similarity index and for the Jaccard coefficient (Tables 11-15 of Appendix 1). Both of these two aforementioned faunas are dissimilar to the more incomplete Chandler and Dunvegan formations from the north of the continent.
During the Turonian, though faunal comparisons could not be made, several significant fossils paint a picture of Appalachian non-avian dinosaur ecology. The Appalachiosaurus-like tyrannosauroid material from the Potomac Formation (Baird, 1989) suggests that tyrannosaurs were gaining in size on Appalachia during the time and may also suggest that Appalachiosaurus and other Appalachian tyrannosaurs were endemic to the landmass. The possible oviraptorosaur tracks from the Raritan Formation would also be the first record of large oviraptorosaurs, let alone oviraptorosaurs, from the eastern portion of North America. However, the lack of information and surviving tracks from this trackway makes any assignment at this time of the footprints to a specific theropod group dubious until more study of the trackway is done. In the west, several groups which prove to be absent from Appalachia for the rest of the Cretaceous appear, including the ceratopsoids (represented by Zuniceratops) and the therizinosaurids (represented by Nothronychus) (Table 16 of Appendix 1). Thus, it may be that the Turonian marks the beginning of the major differentiation of eastern and western Appalachian faunas.
By the Coniacian and Santonian, the difference between eastern and western North American faunas becomes clear. Notably, the Appalachian faunas of the time are arguably better known than those from Laramidia. In Appalachia, the non-hadrosaurid hadrosauroids and non-tyrannosaurid tyrannosauroids are still present, whereas in the western Milk River fauna, they are not. A distinctly Appalachian fauna of the combination of hadrosauroids, hadrosaurids, nodosaurids, tyrannosauroids, ornithomimosaurs, and dromaeosaurids appears, notably devoid of two major groups of herbivorous dinosaurs found in the west: the pachycephalosaurids and ceratopsoids. The Simpson similarity index and Jaccard coefficient values between the Appalachian faunas are very high (Table 1), whereas those between the Appalachian faunas and Milk River Formation fauna are low. Importantly, the first occurrences of hadrosaurids on Appalachia, their probable place of origin, are from the Coniacian and Santonian (Prieto-Márquez et al., 2016a, Prieto-Márquez et al., 2016b), suggesting that the group first evolved during this time. The hadrosaurids from the Niobrara Formation would have coexisted with the relatively small hadrosauroid Claosaurus agilis (Table 17 of Appendix 1).
The Campanian non-avian dinosaur faunas of Appalachia show best the distinct non-avian dinosaur fauna which appeared on the continent. Ornithopods of uncertain affinities, hadrosauroids, basal hadrosaurids, nodosaurids, and leptoceratopsians constitute the ornithischian fauna, while at least two or more taxa of derived non-tyrannosaurid tyrannosauroids (Dryptosaurus aquilunguis, ?Dryptosaurus sp., Appalachiosaurus montgomeriensis) and indeterminate tyrannosauroid remains, multiple species of dromaeosaurids (Saurornitholestes langstoni, Saurornitholestes sp., ?Dromaeosaurus sp.) as well as indeterminate specimens, ornithomimosaurs (“Ornithomimus” antiquus, Ornithomimus sp., indeterminate specimens), indeterminate maniraptorans, indeterminate coelurosaurs, and indeterminate theropods represent the saurischian fauna. At least three different ecomorphs of hadrosauroid dinosaur existed during this time. These were smaller non-hadrosaurid hadrosauroids (represented in the Campanian by Lophorhothon atopus and indeterminate remains), medium-sized (~6-10 m long) hadrosaurids (Eotrachodon orientalis, Hadrosaurus sp., a possibly unnamed taxon from the Coachman Formation, indeterminate remains from the Donoho Creek, Bladen, Coon Creek/Ripey, and Demopolis Chalk Formations) and large to supermassive (~10-17 m long) hadrosauroid taxa (Hadrosaurus foulkii based on the large Woodbury Formation remains, Hypsibema crassicauda, and Parrosaurus missouriensis) (Table 30 of Appendix 1). The hypothesis that large hadrosaurid taxa represent adults of Hadrosaurus is rejected based on the large size of the holotype of the latter taxon and referred specimens of similar size (comparable to adult specimens of western taxa) (e.g., Gallagher, 1993; Prieto-Márquez et al., 2006; pers. obs.). Though the holotype of Eotrachodon is a juvenile of only several years of age (e.g., Prieto-Márquez et al., 2016a; Prieto-Márquez et al., 2016b), Prieto-Márquez et al. (2016b) mentioned that several Appalachian specimens referable to hadrosaurids that are 40%+ larger than the same elements in the Eotrachodon holotype could be referable to that taxon. However, based on the estimated 4-5.1 m length for the holotype of Eotrachodon (e.g., Prieto-Márquez et al., 2016b), such adult animals would be around 5.6-7.4 m in length, much smaller than the estimated length for taxa like Hypsibema and Parrosaurus (e.g., Weishampel and Young, 1996; this paper). It should also be noted that, in the case of hadrosaurs for which individuals of varying sizes are known, very large adult individuals comparable to the size of Hypsibema are rare (e.g., for Edmontosaurus) (e.g., Horner et al., 2011), whereas the Phoebus Landing site has produced many specimens assignable to Hypsibema (e.g., Baird and Horner, 1979). Additionally, the author regards the differences between the vertebrae of Hypsibema crassicauda and Hadrosaurus foulkii as further evidence these taxa are not synonymous.
The possible lack of nodosaurid dinosaurs within the Upper Tar Heel-Coachman fauna is likely due to taphonomic bias, as their remains have occurred both north and south of the Carolinas during the same interval of the Campanian (e.g., Ebersole and King, 2011; Schwimmer et al., 2015). The Coniacian-Santonian taxon Claosaurus agilis and the indeterminate hadrosaurids of the same time from Appalachia may be grouped into the first and second of these ecomorphs, respectively.
The presence of these three distinct ecomorphs of hadrosauroids as well and several nodosaurid taxa from the Campanian of Appalachia may be due to the lack or rareness of three herbivorous dinosaur groups on the eastern landmass present on Laramidia. The ceratopsids, lambeosaurines, and ankylosaurids are almost certainly absent from Appalachia proper until the Maastrichtian (Appendix 1). Both Appalachia and Laramidia share a lack of sauropod dinosaurs from the Cenomanian to Maastrichtian, a phenomenon termed the sauropod hiatus on the latter Landmass (e.g., Lucas and Hunt, 1989; Lehman, 2001; D’Emic et al., 2010; D’Emic et al., 2011; Mannion and Upchurch, 2011; D’Emic et al., 2012). Mallon and Anderson (2013) suggested the presence of a niche partitioning system was present among the herbivorous dinosaurs of the Dinosaur Park Formation and also hypothesized particular niches for each of the large herbivorous clades they examined. Ceratopsids and hadrosaurids were regarded by Mallon and Anderson (2013) as consumers of similar foods, though they also suggested that the hadrosaurs were higher browsers than their ceratopsid contemporaries. Nodosaurids and ankylosaurids were recovered as low browsers, though the former group was suggested to have persisted on tougher food than the latter. The lack of ceratopsids may have also been a factor in the evolution of the Black Creek leptoceratopsian to consume less resistant plant material (Longrich, 2016).
Hadrosaurids were likely the most common herbivorous dinosaurs on Appalachia, and make up more than half of the known dinosaur fauna in southern Appalachia (e.g., Ebersole and King, 2011). Two to five or more taxa of hadrosauroids are present in almost all Appalachian ecosystems sampled from the Coniacian to late Campanian (Tables 23, 31 of Appendix 1), while a maximum of only five taxa is reached in southern Laramidian faunas (with most western hadrosauroid faunas containing on average one lambeosaurine and one to two saurolophines per geological unit) (Table 31 of Appendix 1).
Though ankylosaurs only make up 5% of dinosaur faunas in assemblages from southeastern Appalachia and are the rarest of dinosaurs in the Atlantic Coastal Plain during the Late Cretaceous (Weishampel and Young, 1996; Gallagher, 1997; Ebersole and King, 2011), probably around three taxa (e.g., Niobrarasaurus coeii, the unnamed Mooreville taxon, and possibly Hierosaurus sternbergii) as well as multiple indeterminate remains of nodosaurid dinosaurs, are known from the Coniacian through Campanian of Gulf Coastal and Atlantic Coastal plains. This number is indeed comparable to the three named taxa of nodosaurids known from Laramidia during the same stages of the Late Cretaceous (e.g., Weishampel et al., 2004; Appendix 1), though notably the Appalachian forms only represent those which existed near the coast of about half of Appalachia. Though Butler and Barrett (2008) found a positive correlation between ankylosaurs and marine environments, Arbour et al. (2016) only found significant evidence for association of nodosaurids with the paleoenvironment they died within for those from the Paw Paw Formation. In the case of other nodosaurid-bearing strata from the Cretaceous, there is little statistical evidence for such associations (Arbour et al., 2016). However, the geography of Appalachia, which as noted was almost entirely flat west of the Appalachians, likely could not have produced waterways with forces great enough to carry dinosaur corpses over long distances. Thus, it may be more likely that, in the case of Appalachian nodosaurids, the carrying of specimens out to sea indicate such animals were living near the shore.
Theropod faunas from southern Appalachia are mostly consistent with those from southern Laramidia. Both contain ornithomimosaurs and usually one to two dromaeosaurid taxa. Notably, though large tyrannosauroids are known from Laramidia and Appalachia, the Appalachian forms are derived non-tyrannosaurid tyrannosauroids while all Laramidian forms are nested within Tyrannosauridae (e.g., Weishampel et al., 2004; Carr et al., 2005; Brusatte et al., 2011; Loewen et al., 2013; Brusatte and Carr, 2016; Brusatte et al., 2016). The only taxon of dromaeosaurid reported with certainty from Appalachia is Saurornitholestes, found to be basal eudromaeosaur outside Dromaeosaurinae in at least one analysis (DePalma et al., 2015). Notably, three clades of dinosaurs present on Laramidia (the troodontids, therizinosaurids and oviraptorosaurs) are absent from Appalachia from the Coniacian through the Maastrichtian, though given the small sample size of Appalachian dinosaurs, it is possible that future work will discover the presence of these groups on the eastern landmass. This is especially possible considering the scarcity of these groups on Laramidia (e.g., Weishampel et al., 2004).
Alongside non-tyrannosaurid tyrannosauroids, the presence of at least one taxon of basal eudromaeosaur (Saurornitholestes) in addition to hadrosauroids and nodosaurids with a lack of ceratopsids and lambeosaurines until the Maastrichtian is consistent with Schwimmer (1997) in that non-avian dinosaurs from Appalachia represent relict faunas. At least one group of dinosaurs markedly successful in the Late Cretaceous (the hadrosaurids) seems to have evolved on the continent and remains only in the form of basal taxa. Additionally, one clade of dinosaurs (leptoceratopsians) do not occur on Appalachia until the middle Campanian (Table 31 of Appendix 1), suggesting that they had migrated to the landmass during the middle Late Cretaceous. Therefore, it is probably better to refer to Appalachian non-avian dinosaur faunas as consisting of both relict and endemic clades as well as of those which had migrated to the landmass after it became isolated. Lambeosaurines and ceratopsids likely represent nonnative species which only occurred in Appalachia proper during the Maastrichtian (e.g., Farke and Phillips, 2017). This is supported by the fact that the former clade only occurs on Appalachia during the late Campanian/Maastrichtian, whilst the latter does so during the Maastrichtian (e.g., Gangloff, 2012; Farke and Phillips, 2017). Arbour et al. (2016) additionally speculated that the ankylosaurids, which also are not found on Appalachia, were also isolated from the continent by the Western Interior Seaway.
It is notable that some dinosaur taxa are found across a wide geographic range on Appalachia and in formations of slightly different ages. Of the Coniacian-Campanian Appalachian hadrosauroid taxa (Claosaurus agilis, Lophorhothon atopus, Hadrosaurus foulkii, Hadrosaurus sp., Hypsibema crassicauda, Parrosaurus missouriensis, Eotrachodon orientalis, unnamed Kanguk lambeosaurine), four are known from multiple formations comprising multiple different provinces (Lophorhothon atopus; Hadrosaurus foulkii, Hadrosaurus sp., Hypsibema crassicauda) (see below) and from multiple different intervals of time (Tables 23, 31, 38 of Appendix 1). This is the same for the tyrannosauroid taxa Appalachiosaurus montgomeriensis and Dryptosaurus aquilunguis, the dromaeosaurid taxon Saurornitholestes langstoni, and the ornithomimosaur “Ornithomimus” antiquus, and may be due to the geography of Appalachia allowing dinosaurs to migrate across the landscape or alternatively be a signal that taxa reported from many wide-ranging and temporally disparate faunas are actually divisible into multiple species. Notably, reports of the three hadrosauroids listed above that have been reported from faunas other than from whence their holotype was recovered are mostly based on one or a few elements (Baird and Horner, 1979; Grandstaff et al., 1992; Prieto-Márquez et al., 2016b). This suggests that an even wider variety of hadrosauroid species may have existed on Appalachia then stated herein. For example, if the multiple occurrences of these three aforementioned taxa are regarded as distinct species, a total of about 10 or more, rather than seven to eight, hadrosauroid taxa are known from the surviving deposits of the landmass of Appalachia. Accounting for the badly-sampled interior, west, and north of Appalachia as well as the lack of localities corresponding to the landmass from the Coniacian and Santonian, the actual number of distinct hadrosauroid species from the Coniacian-Campanian of the landmass may have been twice or thrice as many as currently known. This may also apply to other ornithischian and theropod dinosaurs, though the latter group seems to have been constrained in diversity in some areas for several reasons ecologically (see below).
Dinosaur Provincialism on Appalachia
Very little work has been done on the possibility of non-avian dinosaur provincialism on Appalachia. Schwimmer (2016) suggested the presence of distinct northern and southern non-avian dinosaur faunas on Appalachia, though did not mention the presence of ornithomimosaurians in the Atlantic Coastal Plain and also included Maastrichtian lambeosaurines within the northern dinosaur province, using the presence of these crested hadrosaurs to help differentiate the different provinces. The results herein, however, nevertheless support the hypothesis of Schwimmer (2016) that dinosaur provincialism did occur on Appalachia. On Laramidia, the presence of non-avian dinosaur provincialism is notably debated (e.g., Lehman, 1997, Sampson et al., 2010; Sampson et al., 2013a; Lucas et al., 2016).
The rarefaction of Coniacian/Santonian dinosaur faunas of Appalachia, as noted previously, showed the discrepancy between the sample bins of the Eutaw and Niobrara Formations. However, a few conclusions may still be reached about the biogeographic relations of the Eutaw and Niobrara Formations. Firstly, their dinosaur faunas may be distinguished from each other by the different genus of hadrosauroid found in each. Claosaurus agilis is only known from the Santonian of the Niobrara (e.g., Carpenter et al., 2011), whereas the Eutaw Formation preserves bones assignable to Lophorhothon (e.g., Ebersole and King, 2011). Secondly, both share nodosaurids, hadrosauroids, and hadrosaurids, three key components of the general Appalachian fauna discussed later in this manuscript. The calculated Simpson similarity index and Jaccard coefficient values do show support for the differentiation of these two faunas, as though they have values of 0.0 on the genus level, on the family level the Simpson similarity index has a value of 100% and the Jaccard coefficient has a value of 0.4 (as noted, calculation of the Jaccard coefficient emphasizes differences). Thus, it may be that a distinct dinosaur fauna from that of the southeastern portion of of Appalachia existed in the western portion of the landmass during the Santonian, but further discoveries will be needed to better validate this hypothesis.
Because the rarefaction of Campanian Appalachian dinosaur faunas showed relatively small differences between the represented number of clades from each of the three formations analyzed, the hypothesis of dinosaur provincialism can be more confidently tested for this stage of the Late Cretaceous. The Simpson similarity index and Jaccard index values for comparisons between the three formations analyzed are low and the Jaccard distance values are high between the southern Appalachian Mooreville Chalk, Blufftown, and Coffee Sand formations and the unnamed Missouri deposit and the Marshalltown Formation of the Atlantic Coastal Plain (Tables 32-37 of Appendix 1), whereas the former two indices are moderate to high and the latter is low in value between either of the former two formations and the upper Tar Heel and Coachman Formations (Tables 32-37 of Appendix 1). Thus, the presence of distinct northern and southern Appalachian non-avian dinosaur provinces is supported by the calculated values of Simpson similarity index and Jaccard coefficient. The Atlantic Coastal Plain province is the name suggested herein for the northern fauna, and the Gulf Coastal Plain province is the name suggested herein for the southern.
The Campanian Atlantic Coastal Plain province can be characterized by the presence of one or more species of tyrannosauroid (Dryptosaurus aquilunguis, ?Dryptosaurus sp.), a scarcity of dromaeosaurids compared to their presence in southern deposits (Tables 31, 46 of Appendix 1), and the presence of the hadrosaurid Hadrosaurus foulkii and remains assignable to Hadrosaurus sp. A Campanian southeastern Appalachian province can be defined by the inclusion of only one taxon of tyrannosauroid that is unknown in the Atlantic Coastal Plain (Appalachiosaurus montgomeriensis), the presence of the dromaeosaurid Saurornitholestes and numerous remains from indeterminate dromaeosaurids (Table 46 of Appendix 1; Ebersole and King, 2011), and the presence of the hadrosaurids Eotrachodon orientalis, Parrosaurus missouriensis, and hadrosauroid Lophorhothon atopus (Table 31 of Appendix 1).
The upper Tar Heel and Coachman Formations, as indicated by the Simpson similarity index, Jaccard coefficient, and Jaccard distance values as well as analytical observations, thus represents an intersection between these two faunas. It also includes several taxa unique to itself among the three Campanian Appalachian faunas analyzed, including a leptoceratopsian and a second morphotype of dromaeosaurid besides Saurornitholestes (i.e., ?Dromaeosaurus sp.) (Schwimmer et al., 2015). That these taxa are not present in the other Campanian Appalachian formations (Table 31 of Appendix 1) may simply be a product of their rarity. However, the distinct combination of dinosaurs in the Atlantic (Dryptosaurus, Hadrosaurus) and Gulf Coastal Plains (Lophorhothon, Appalachiosaurus) found in the Tar Heel and Coachman Formations suggest the Atlantic and Gulf Coastal Plain provinces may have graded into each other on a longitudinal axis, similar to the hypothesis Gates et al. (2010) suggested regarding Laramidian provinces. This mixed fauna is referred to herein as the Carolinian province.
Though several of the formations comprising the Gulf Coastal Plain province are of older age than the Tar Heel, Coachman, and Marshalltown formations (e.g., Schwimmer et al., 1993; Miller et al., 2004; Self Trail et al., 2004; Harris and Self-Trail, 2006; Ebersole and King, 2011; Schwimmer et al., 2015), it should be noted that Atlantic Coastal Plain dinosaur faunas from early Campanian formations of comparable age to southern formations (i.e., the Merchantville Formation) (Miller et al., 2004) contain elements distinct from southern Appalachian faunas, including Hadrosaurus, an indeterminate but distinct taxon of tyrannosauroid, and several specimens of ornithomimosaur (only one specimen of ornithomimosaur is documented from the Campanian of the Gulf Coastal Plain) (e.g., Schwimmer et al., 1993; Ebersole and King, 2011). Among the Gulf Coastal Plain units, the Demopolis Chalk, which is of middle-late Campanian age (e.g., Carr et al., 2005; Ebersole and King, 2011) bears the ~78 million year old holotype specimen of Appalachiosaurus, which is apparently not found in the Atlantic Coastal Plain (e.g., Schwimmer, 2016; pers. obs.). Further examination of the specimens of these early Campanian formations (the hadrosauroid/hadrosaurid and non-tyrannosauroid coelurosaurian taxa especially) will be needed to corroborate the results of biogeographic comparisons between the earlier Gulf Coastal Plain and later Atlantic Coastal Plain and Carolinian faunas, though the current weight of evidence does seem to suggest that Appalachian dinosaur provincialism occurred during the Campanian Stage.
Additionally, one other stratigraphic unit may suggest the presence of another dinosaur province. Though the small sample size of the Campanian/Maastrichtian Kanguk Formation warrants only analytical comparison with the faunas, the presence of lambeosaurines in this unit (Gangloff, 2012) may suggest the presence of another non-avian dinosaur province in northern Appalachia. However, the hypothesis of the existence of a northern Appalachian province requires further data from the northern reaches of North America to be thoroughly tested.
Loewen et al. (2013) suggested the presence of dinosaur provincialism on Laramidia was due to transgressions and regressions of the Western Interior Seaway isolating local dinosaur faunas from each other along the Sevier orogeny. Indeed, the dinosaur faunas from the eastern seaboard of North America were proximal to the Appalachian mountains (e.g., Schwimmer, 2002), and so transgressions and regressions of the Atlantic Ocean may have resulted in a similar result, creating the northern and southern dinosaur faunas. The rarefaction of the Mooreville Chalk, Blufftown, and Coffee Sand formations and unnamed Missouri clay fauna with the subsample of the Eutaw Formation (=45) gives an estimated number of seven clades, showing that a small discrepancy exists between bins. However, these two units, when compared, have Simpson similarity index values of 100% on both the genus and family levels, as well as high Jaccard coefficient values and thus low Jaccard distance values (Table 49 of Appendix 1). Thus, they may be grouped together in the Gulf Coastal Plain province. This grouping provides further evidence that the Niobrara Formation represents another fauna, which is referred to herein as the Western Appalachian fauna (Figure 8.1).
Thus, three to five non-avian dinosaur provinces on Appalachia may be apparent from current data. Four of these (the Atlantic Coastal Plain, Gulf Coastal Plain, Carolinian, and Western Appalachian provinces) are quantifiable based on data from the calculation of Jaccard coefficient and Simpson similarity index values. A possible fifth, from northern Appalachia and represented in the Kanguk Formation, may be differentiated by the presence of lambeosaurines, but further data from northern North America is required to support its existence.
Possible Ecological Constraints on Appalachian Dinosaur Diversity and Commonality
Several patterns in Appalachian dinosaur biogeography may relate to Appalachian ecology. One such case involves the tyrannosauroids of Appalachia. Though several tyrannosaurid tyrannosauroids are present in far southern Laramidia during the Campanian, only the species Appalachiosaurus mongomeriensis is present in the Gulf Coastal Plain province of Appalachia during the Campanian (e.g., Tables 23, 31 of Appendix 1). Additionally, most remains of this sole taxon are probably juveniles (e.g., Schwimmer, 2002; Carr, Williamson and Schwimmer, 2005; Schwimmer et al., 2015). Schwimmer (1997), Schwimmer (2002), and Schwimmer et al. (2015) suggested that rather than large theropod dinosaurs, the massive crocodylian Deinosuchus rugosus constituted the apex predator of southeastern Appalachian ecosystems. Multiple observations from this study and others both indirectly and directly support this hypothesis, and as such previous work on the abundance of Deinosuchus is reviewed herein. Firstly, this large, predatory crocodylian was extremely abundant in the Gulf Coastal Plain (Schwimmer and Williams, 1993; Schwimmer and Williams, 1997; Schwimmer, 1997; Schwimmer, 2002) and reached sizes of up to 10 meters in length (Schwimmer, 1997). Secondly, possible evidence of predation on medium-sized theropods (~400 kg) (including tyrannosauroids) by Deinosuchus rugosus is known from bones recovered from the Ellisdale site in New Jersey and from Georgia (Schwimmer, 2002). Deinosuchus rugosus remains are also extremely common in the Gulf Coastal plain (e.g., Schwimmer and Williams, 1993; Schwimmer, 1997; Schwimmer and Williams, 1997; Schwimmer, 2002). However, Deinosuchus rugosus remains have been noted as less common in sediments from the Coachman and Tar Heel Formations of the Carolinas (Baird and Horner, 1979; Schwimmer et al., 2015). Finally, only one significant Deinosuchus rugosus bearing locality is known from the Atlantic Coastal Plain (Schwimmer, 2002). Notably, only one scute possibly assignable to Deinosuchus has been recovered from the Missouri Chronister Site (Darrough, personal commun., 2016), which may be due to the separation of the Chronister site from other Gulf Coastal Plain sites by the Mississippi Water Gap causing a change in fauna. However, the western portion of Appalachia is poorly sampled and so any attempt to estimate western Appalachian Deinosuchus populations should be considered tentative. Although, as noted herein, there is a temporal difference of several million years between the most dinosaur-speciose southern Appalachian faunas and those from the Carolinas and Atlantic Coastal Plain, it does seem that across this ~10 million year timescale (e.g., Miller et al., 2004; Harris and Self-Trail, 2006; Ebersole and King, 2011) the abundance of Deinosuchus in certain formations and rareness in others noted by Schwimmer (2002) correlates with the number of species and abundance of tyrannosauroids as well as with the abundance of dromaeosaurid material as reviewed and compared in this study. This correlation was also noted by Schwimmer (2002). However, the extensive review of Appalachian dinosaur faunas and specimens (for rarefaction) herein importantly adds additional support to this hypothesis. At least two to three different taxa of tyrannosauroid existed in the Atlantic Coastal Plain (Dryptosaurus, the Merchantville taxon, and possibly others represented by fragmentary or isolated specimens) (e.g., Gallagher, 1993; Schwimmer, 2016; Table 22 of Appendix 1; pers. obs.), while the south has only Appalachiosaurus montgomeriensis during the Campanian (e.g., Carr et al., 2005; Ebersole and King, 2011; this paper). Sampling and depositional biases as causes of this difference in the diversity of tyrannosauroids between the Atlantic and Gulf Coastal plains is unlikely, as the Merchantville, Marshalltown, Mooreville Chalk, Blufftown, etc., all represent marginal marine to marine deposits (e.g., Gallagher, 1993; Miller et al., 2004; Carr et al., 2005; Ebersole and King, 2011) and the results of rarefaction indicate minimal differences between examined formations (this paper). This latter statement is especially true when the thousands of dinosaur specimens collected from the Ellisdale site but not yet closely evaluated (e.g., Denton et al., 2011; Denton, personal commun., 2017) and thus excluded here is taken into account. However, further collection and description of Merchantville Formation dinosaur specimens will be needed for the use of that unit in rarefaction and statistical analysis.
This apparent north-south (though note Appalachian coastal plains were rotated relatively southward compared to the condition today) (e.g., Schwimmer, 1997; Schwimmer, 2002) correlation between the abundance of Deinosuchus and the diversity and numerousness of large and small theropods supports the hypothesis of Schwimmer (2002) that the large crocodylian may have been outcompeting large, predatory theropod dinosaurs in the Gulf Coastal plain and became rarer towards the middle of Appalachia. However, as the vast majority of Appalachian dinosaur occurrences from the Gulf Coastal Plain, Carolinian, and Atlantic Coastal Plain provinces are from marine sediments, it may be that the preservation of Deinosuchus and other crocodylians in these sediments was taphonomically favored. Thus, better evidence for large crocodyliforms being the apex predators of their respective ecosystems in Appalachia comes from the Arlington Archosaur Site of the Woodbine Formation, which preserves a deltaic ecosystem. The presence of a large, generalist crocodyliform within southern Appalachian ecosystems is notably not restricted to Campanian faunas. The large crocodyliform Deltasuchus motherali is known from the Cenomanian Woodbine Formation where it was apparently very common (e.g., Main et al., 2012; Noto et al., 2012; Main, 2013; Adams et al., 2017). Evidence of feeding on dinosaurs by this large crocodyliform has been documented (Noto et al., 2012) and the possibility that for ~20 million years (Cenomanian to Campanian) crocodyliforms were the apex predators in at least Texas ecosystems has been discussed (Main et al., 2012). Importantly, theropod remains are rare at the Arlington Archosaur Site, the locality of the discovery of Deltasuchus motherali (e.g., Main et al., 2012; Main, 2013; Adams et al., 2017). The commonality of this unnamed crocodyliform in an inland ecosystem combined with direct evidence of feeding on dinosaurs further suggests that crocodyliforms were taking on the role of apex predators in Appalachian ecosystems. However, as with many other hypotheses regarding the ecology of the inhabitants of Appalachia, more specimens will be needed to support the hypothesis of crocodyliforms being apex predators in Appalachia.
The possibility of ecological competition helping to create distinct dinosaur provinces may also be applicable to other Mesozoic and especially Campanian southern Laramidian ecosystems, where Deinosuchus were also somewhat abundant and where the most diverse Laramidian crocodylomorph assemblage (comparable to that of the Upper Tar Heel and Coachman formations per Baird and Horner, 1979; Schwimmer et al., 2015) occurred (Schwimmer, 2002; Sampson et al., 2013b).
Comparisons of Appalachian and European Coniacian-Campanian Faunas
Though the data from this paper corresponds to Laramidian and Appalachian dinosaurs, a comparison with those from the Cretaceous of Europe is warranted. Both faunas are similar in sharing leptoceratopsians, derived non-hadrosaurid hadrosauroids, nodosaurids, and dromaeosaurids (e.g., Weishampel, 2004; Dalla Vecchia, 2009; Kirkland et al., 2013). Notably, the European hadrosauroid taxon Tethyshadros insularis was recovered by Prieto-Márquez et al. (2016a) as the sister taxon of Hadrosauridae and therefore as one closer node to that clade than the Appalachian taxon Claosaurus agilis and as two closer nodes than Lophorhothon atopus. This phylogenetic data is suggestive that interchanges between European and Appalachian non-avian dinosaur faunas occurred during the middle Late Cretaceous. This evidence for interchange between Europe and Appalachia supports the hypothesis that the immigration of leptoceratopsians to Appalachia was from Europe rather than from Laramidia (e.g., Lindgren et al., 2007; Longrich, 2016).
Appalachian and European faunas can be distinguished from each other in several ways. First, the dominant clade of large (~5+ meter) predatory theropods on Appalachia was the tyrannosauroidea, while in Europe the ceratosaurian abelisaurids seemingly take this role (e.g., Weishampel et al., 2004; Tortosa et al., 2013). Secondly, ornithomimosaurs are fairly common and widespread on Appalachia during the Late Cretaceous (e.g., Weishampel et al., 2004; Erickson and Ebersole, 2011; Schwimmer et al., 2015), whereas during the same time only one probable occurrence is known from Europe (Mateus et al., 2009). European nodosaurids seem to form a distinct clade, the Struthiosaurinae (Kirkland et al., 2013), while the majority of Appalachian taxa (Texasetes pleurohalio, the unnamed Paw Paw juvenile, Silvisaurus condrayi, Niobrarasaurus coleii) have consistently not been found as close relatives of any ‘struthiosaurines’ (e.g., Carpenter, 2001; Vickaryous et al., 2004; Arbour et al., 2016). These major differences affirm that Appalachian faunas were distinct from European ones, and suggest along with the data recounted herein on Laramidian and Appalachian non-avian dinosaurs that Appalachia itself was a distinct non-avian dinosaur faunal zone during the majority of the Late Cretaceous.
CONCLUSIONS
Appalachian non-avian dinosaurs represent a distinct fauna, which included both relict (hadrosauroids, Saurornitholestes, comparatively abundant nodosaurids, non-tyrannosaurid tyrannosauroids), endemic (non-saurolophid hadrosaurids), and immigrant (leptoceratopsians) forms. This fauna lasted from the Coniacian to the Late Campanian with its possible origins in the Turonian, and changed by the immigration of nonnative clades from the American west by the Maastrichtian. Jaccard coefficient and Simpson similarity index comparisons when combined with the results of rarefaction suggest the presence of three to five different non-avian dinosaur provinces on Appalachia from the Coniacian to Campanian Stages of the Late Cretaceous. Four are named the Gulf Coastal Plain, Atlantic Coastal Plain, Carolinian, and Western Appalachian faunas herein, while a possible fifth is represented by the Kanguk Formation in Nunavut, Canada. Hadrosauroids occurred in at least three different ecomorphs (small hadrosauroids, medium-sized hadrosaurids and hadrosauroids, and supermassive hadrosaurids), nodosaurids were relatively abundant, and tyrannosauroids constituted as the apex predators towards the middle of Appalachia. In the southern portion of the landmass, Deltasuchus and Deinosuchus rugosus crocodylians may have outcompeted large theropod dinosaurs as apex predators, though more specimens will be needed to quantify this hypothesis. The Appalachian non-avian dinosaur record and the possibility of ecological interactions shaping Appalachian dinosaur provinces have implications for the creation of southern Laramidian dinosaur provinces and for dinosaur provinciality generally.
ACKNOWLEDGEMENTS
The author would like to thank T. Jorstad of the Smithsonian Institution and J. Henderson of the Yale Peabody Museum for providing photos of specimens figured herein, and D. Schwimmer of Columbus State University, J. Ebersole of the McWane Science Center, G. Darrough of the Bollinger County Museum of Natural History and Lost World Studios, M. Fix of the University of Missouri-St. Louis, R. Denton of Geoconcepts Engineering Inc., M. Suter of the University of Arkansas at Fayetteville, R. Johnson of the Monmouth Amateur Paleontologists’ Society, G. Phillips of the Mississippi Museum of Natural Science, and members of the North Carolina Fossil Club for our discussions regarding Appalachian dinosaurs and for providing me with papers, information on certain specimens and sites, and other particulars that were very helpful in writing this manuscript. The author would also like to thank D. Brinkman of the Yale Peabody Museum and C. Mehling of the American Museum of Natural History for allowing me access to the Yale Peabody Museum and American Museum of Natural History collections, respectively, and for our discussions regarding the specimens I studied in those collections and on Appalachian dinosaurs generally. Finally, I would like to thank L. Wilson and three anonymous reviewers for their review of this manuscript.
REFERENCES
Adams, T.L., Noto, C.R., and Drumheller, S. 2015. The crocodyliform diversity of the Woodbine Formation (Cenomanian) of Texas and the transition from Early to Mid Cretaceous ecosystems. Society of Vertebrate Paleontology Abstracts with Programs, 75:77.
Alroy, J., Marshall, C.R., Bambach, R.K., Bezusko, K., Foote, M., Fursich, F.T., Hansen, T.A., Holland, S.M., Ivany, L.C., Jablonski, D., Jacobs, D.K., Jones, D.C., Kosnik, M.A., Lidgard, S., Low, S., Miller, A.I., Novack-Gottshall, P.M., Olszewski, T.D., Patzkowsky, M.E., Raup, D.M., Roy, K., Sepkoski, J.J., Sommers, M.G., Wagner, P.J., and Webber, A. 2001. Effects of sampling standardization on estimates of Phanerozoic marine diversification. Proceedings of the National Academy of Sciences of the United States of America, 98:6261-6266. https://doi.org/10.1073/pnas.111144698
Arbour, V.M. and Currie, P.J. 2013. Euoplocephalus tutus and the diversity of ankylosaurid dinosaurs in the Late Cretaceous of Alberta, Canada, and Montana, USA. PLoS ONE, 8(5): e62421. https://doi.org/10.1371/journal.pone.0062421
Arbour, V.M., Burns, M.E., Sullivan, R.M., Lucas, S.G., Cantrell, A.K., Fry, J., and Suazo, T.L. 2014. A new ankylosaurid dinosaur from the Upper Cretaceous (Kirtlandian) of New Mexico with implications for ankylosaurid diversity in the Upper Cretaceous of Western North America. PLOS ONE, 9(9):e108804. https://doi.org/10.1371/journal.pone.0108804
Arbour, V.M., Zanno, L.E., and Gates, T. 2016. Ankylosaurian dinosaur palaeoenvironmental associations were influenced by extirpation, sea-level fluctuation, and geodispersal. Palaeogeography, Palaeoclimatology, Palaeoecology, 449:289-299. https://doi.org/10.1016/j.palaeo.2016.02.033
Baird, D. 1986. Upper Cretaceous reptiles from the Severn Formation of Maryland. The Mosasaur, 3:63-85.
Baird, D. 1988. Medial Cretaceous carnivorous dinosaur bone and footprints from New Jersey. Journal of Vertebrate Paleontology, 8(suppl. 3):8A.
Baird, D. 1989. Medial Cretaceous carnivorous dinosaur and footprints from New Jersey. The Mosasaur, 4:53-64.
Baird, D. and Galton, P.M. 1981. Pterosaur bones from the Upper Cretaceous of Delaware. Journal of Vertebrate Paleontology, 1:67-71. https://doi.org/10.1080/02724634.1981.10011880
Baird, D. and Horner, J.R. 1977. A fresh look at the dinosaurs of New Jersey and Delaware. Bulletin of the New Jersey Academy of Sciences, 22:50.
Baird, D. and Horner, J.R. 1979. Cretaceous dinosaurs of North Carolina. Brimleyana, 2:1-28.
Bird, R.T. 1939. Thunder in his footsteps. Natural History, 43:254-261.
Bird, R.T. 1944. Did Brontosaurus ever walk on land? Natural History, 53:60-67.
Bonde, J., Varricchio, D.J., Jackson, F.D., Loope, D.B., and Shirk, A.M. 2008. Dinosaurs and dunes! Sedimentology and paleontology of the Mesozoic in the Valley of Fire State Park. Geological Society of America Field Guide 11:249-262.
Bonde, J.W., Varricchio, D.J., Bryant, G., and Jackson, F.D. 2012. Mesozoic vertebrate paleontology of Valley of Fire State Park, southern Nevada, p. 108-126. In Hattori, E.M. (ed.), Field Trip Guide Book for the 71st Annual Meeting of The Society Of Vertebrate Paleontology, Paris Los Vegas, Las Vegas, Nevada, USA.
Boyd, C.A., Drumheller, S.K., and Gates, T.A. 2013. Crocodyliform feeding traces on juvenile ornithischian dinosaurs from the Upper Cretaceous (Campanian) Kaiparowits Formation, Utah. PLoS ONE, 8(2):e57605. https://doi.org/10.1371/journal.pone.0057605
Brenner, J.G. 1963. The Spores and Pollen of the Potomac Group of Maryland, p. 15. State of Maryland, Board of Natural Resources, Dept. of Geology, Mines, and Water Resources, Baltimore, Maryland, USA.
Brenner, R.L., Ludvigson, G.A., Witzke, B.J., Zawistoski, A.N., Kvale, E.P., Ravn, R.L., and Joeckel, R.M. 2000. Late Albian Kiowa-Skull Creek marine transgression, lower Dakota Formation, eastern margin of Western Interior Seaway, USA, Journal of Sedimentary Research, 70:868-878. https://doi.org/10.1306/2DC4093E-0E47-11D7-8643000102C1865D
Brill, K. and Carpenter, K. 2006. A Description of a New Ornithopod from the Lytle Member of the Purgatoire Formation (Lower Cretaceous) and a Reassessment of the Skull of Camptosaurus, p. 49-67. In Carpenter, K. (ed.), Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Indiana University Press, Bloomington, Indiana, USA.
Brown, C.M. and Druckenmiller, P. 2011. Basal ornithopod (Dinosauria: Ornithischia) teeth from the Prince Creek Formation (early Maastrichtian) of Alaska. Canadian Journal of Earth Sciences, 48:1342-1354. https://doi.org/10.1139/e11-017
Brown, C.M., Evans, D.C., Ryan, M.J., and Russell, A.P. 2013. New data on the diversity and abundance of small-bodied ornithopods (Dinosauria, Ornithischia) from the Belly River Group (Campanian) of Alberta. Journal of Vertebrate Paleontology, 33(3):495. https://doi.org/10.1080/02724634.2013.746229
Brownstein, C.D. 2017a. Description of Arundel Clay ornithomimosaur material and a reinterpretation of Nedcolbertia justinhofmanni as an "Ostrich Dinosaur": biogeographic implications. PeerJ, 5:e3110. https://doi.org/10.7717/peerj.3110.
Brownstein, C.D. 2017b. A tyrannosauroid metatarsus from the Merchantville Formation of Delaware increases the diversity of non-tyrannosaurid tyrannosauroids on Appalachia. PeerJ,5:e4123. https://doi.org/10.7717/peerj.4123.
Brusatte, S.L. and Carr, T.D. 2016. The phylogeny and evolutionary history of tyrannosauroid dinosaurs. Scientific Reports, 6:e20252. https://doi.org/10.1038/srep20252
Brusatte, S.L., Averianov, A., Sues, H.D., Muir, A., and Butler, I.B. 2016. New tyrannosaur from the mid-Cretaceous of Uzbekistan clarifies evolution of giant body sizes and advanced senses in tyrant dinosaurs. Proceedings of the National Academy of Sciences of the United States of America, 113(13):3447-3452. https://doi.org/10.1073/pnas.1600140113
Brusatte, S.L., Benson, R.B., and Norell, M.A. 2011. The anatomy of Dryptosaurus aquilunguis (Dinosauria: Theropoda) and a review of its tyrannosauroid affinities. American Museum Novitates, 3717:1-53.
Brusatte, S.L., Choiniere, J.N., Benson, R.B., Carr, T.D., and Norell, M.A. 2012. Theropoda dinosaurs from the Late Cretaceous of Eastern North America: anatomy, systematics, biogeography and new information from historic specimens. Journal of Vertebrate Paleontology, 32(supp. 5):70.
Burns, M.E. and Vavrek, M.J. 2014. Probable ankylosaur ossicles from the Middle Cenomanian Dunvegan Formation of northwestern Alberta, Canada. PLoS ONE, 9(5):e96075. https://doi.org/10.1371/journal.pone.0096075
Burton, D., Greenhalgh, B.W., Britt, B.B., Kowallis, B.J., Elliott, W.S., and Barrick, R. 2006. New radiometric ages from the Cedar Mountain Formation, Utah and the Cloverly Formation, Wyoming: implications for contained dinosaur faunas. Geological Society of America Abstracts with Programs, 38:52.
Carpenter, K. 1982. Baby dinosaurs from the Late Cretaceous Lance and Hell Creek formations and a description of a new species of theropod. Contributions to Geology, University of Wyoming, 20(2):123-134.
Carpenter, K. 2001. Phylogenetic analysis of the Ankylosauria, p. 455-483. In Carpenter, K. (ed.), The Armored Dinosaurs. Indiana University Press, Bloomington, Indiana, USA.
Carpenter, K. and Cifelli, R.L. 2016. A possible juvenile ceratopsoid ilium from the Cenomanian of central Utah, USA. Cretaceous Research, 60:167-175. https://doi.org/10.1016/j.cretres.2015.11.016
Carpenter, K. and Kirkland, J.I. 1998. Review of Lower and middle Cretaceous ankylosaurs from North America. In Lucas, S.G., Kirkland, J.I., and Estep, J.W. (eds.), Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin, 14:249-270.
Carpenter, K., Bartlett, J., Bird, J., and Barrick, R. 2008. Ankylosaurs from the Price River Quarries, Cedar Mountain Formation (Lower Cretaceous), east-central Utah. Journal of Vertebrate Paleontology, 28(4):1089-1101. https://doi.org/10.1671/0272-4634-28.4.1089
Carpenter, K., Dilkes, D., Weishampel, D.B. 1995. The dinosaurs of the Niobrara Chalk Formation (Upper Cretaceous, Kansas). Journal of Vertebrate Paleontology, 15(2):275-297. https://doi.org/10.1080/02724634.1995.10011230
Carpenter, K., Kirkland, J.I., Burge, D.L., and Bird, J. 1999. Ankylosaurs (Dinosauria: Ornithischia) of the Cedar Mountain Formation, Utah, and their stratigraphic distribution. In Gillette, D. (ed.), Vertebrate Paleontology in Utah. Utah Geological Survey Miscellaneous Publication, 99(1):243-251.
Carpenter, K., Kirkland, J.I., Birge, D., and Bird, J. 2001. Disarticulated skull of a new primitive ankylosaurid from the Lower Cretaceous of Utah. In Carpenter, K. (ed.), 2001. The Armored Dinosaurs. Indiana University Press, Bloomington, Indiana, USA.
Carr, T.D. and Williamson, T.E. 2010. Bistahieversor sealeyi, gen. et sp. nov., a new tyrannosauroid from New Mexico and the origin of deep snouts in Tyrannosauroidea. Journal of Vertebrate Paleontology, 30(1):1-16. https://doi.org/10.1080/02724630903413032
Carr, T.D., Williamson, T.E., and Schwimmer, D.R. 2005. A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama. Journal of Vertebrate Paleontology, 25(1):119-143.https://doi.org/10.1671/0272-4634(2005)025%5B0119:ANGASO%5D2.0.CO;2
Caughey, C.A. 1977. Depositional systems in the Paluxy Formation (Lower Cretaceous), Northeast Texas oil, gas, and groundwater resources. Geological Circular, Bureau of Economic Geology, The University of Texas at Austin, 77-8:1-59.
Chen, Z.Q. and Lubin, S. 1997. A fission track study of the terrigenous sedimentary sequences of the Morrison and Cloverly Formations in northeastern Bighorn Basin, Wyoming. The Mountain Geologist, 34:51-62.
Chiappe, L.M., Lamb, J.P., and Ericson, P.G.P. 2002. New Enantiornithine bird from the marine Upper Cretaceous of Alabama. Journal of Vertebrate Paleontology, 22(1):170-174.
Chinnery, B.J. 2004. Description of Prenoceratops pieganensis gen et sp. nov. (Dinosauria: Neoceratopsia) from the Two Medicine Formation of Montana. Journal of Vertebrate Paleontology, 24(3):572-590. https://doi.org/10.1671/0272-4634(2004)024%5B0572:DOPPGE%5D2.0.CO;2
Chinnery, B.J. and Horner, J.R. 2007. A new neoceratopsian dinosaur linking North American and Asian taxa. Journal of Vertebrate Paleontology, 27(3):625-641. https://doi.org/10.1671/0272-4634(2007)27%5B625:ANNDLN%5D2.0.CO;2
Chinnery, B.J., Lipka, T.R., Kirkland, J.I., Parrish, J.M., and Brett-Surman, M.K. 1998. Neoceratopsian teeth from the Lower and middle Cretaceous of North America. In Lucas, S.G., Kirkland, J.I., and Estep, J.W. (eds.), Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin, 14:297-302.
Chure, D., Britt, B., Whitlock, J.A., and Wilson, J.A. 2010. First complete sauropod dinosaur skull from the Cretaceous of the Americas and the evolution of sauropod dentition. Naturwissenschaften, 97(4):379-391. https://doi.org/10.1007/s00114-010-0650-6
Cifelli, R.L. 1997. First notice on Mesozoic mammals from Oklahoma. Oklahoma Geology Notes, 57:4-17.
Cifelli, R.L. and Gardner, J.D. 1997. Additions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous), southeastern Oklahoma. Oklahoma Geology Notes, 57(4):124-131.
Cifelli, R.L., Lipka T.R., Schaff C.R., and Rowe, T.B. 1999. First Early Cretaceous mammal from the eastern seaboard of the United States. Journal of Vertebrate Paleontology, 19(2):199-203.
Colbert, E.H. 1948. A hadrosaurian dinosaur from New Jersey. Proceedings of the Academy of Natural Sciences of Philadelphia 100:23-37.
Coombs, W.P. 1995. A nodosaurid ankylosaur (Dinosauria: Ornithischia) from the Lower Cretaceous of Texas. Journal of Vertebrate Paleontology, 15(2):298-312. https://doi.org/10.1080/02724634.1995.10011231
Cope, E.D. 1871. Observations on the distribution of certain extinct vertebrata in North Carolina. Proceedings of the American Philosophical Society, 12(86):210-216.
Crane, C.D. 2011. Vertebrate Paleontology and Taphonomy of the Late Cretaceous (Campanian) Bladen Formation, Bladen County, North Carolina. M.Sc., East Carolina University, Greenville North Carolina, USA.
Currie, P.J. and Carpenter, K. 2000. A new specimen of Acrocanthosaurus atokensis (Theropoda, Dinosauria) from the Lower Cretaceous Antlers Formation (Lower Cretaceous, Aptian) of Oklahoma, USA. Geodiversitas, 22:207-246.
D’Emic, M.D. 2013. Revision of the sauropod dinosaurs of the Lower Cretaceous Trinity Group, southern USA, with the description of a new genus. Journal of Systematic Palaeontology. 11(6): 707-726. https://doi.org/10.1080/14772019.2012.667446
D’Emic, M.D., and Britt, B. 2008. Reevaluation of the phylogenetic affinities and age of a basal titanosauriform (Sauropoda: Dinosauria) from the Early Cretaceous Cloverly Formation of North America. Journal of Vertebrate Paleontology, 28(suppl. 3):68A.
D’Emic, M.D. and Foreman, B.Z. 2012. The beginning of the sauropod dinosaur hiatus in North America: insights from the Lower Cretaceous Cloverly Formation of Wyoming. Journal of Vertebrate Paleontology, 32(4): 883-902. https://doi.org/10.1080/02724634.2012.671204
D’Emic, M.D., Melstrom, K.M., and Eddy, D.R. 2012. Paleobiology and geographic range of the large-bodied Cretaceous theropod dinosaur Acrocanthosaurus atokensis. Palaeogeography, Palaeoclimatology, Palaeoecology, 333-334:13-23.https://doi.org/10.1016/j.palaeo.2012.03.003
D’Emic, M.D., Wilson, J.A., and Thompson, W. 2010. The end of the sauropod hiatus in North America. Palaeogeography, Palaeoclimatology, Palaeoecology, 297:486-490. https://doi.org/10.1016/j.palaeo.2010.08.032
D’Emic, M.D., Wilson, J.A., Williamson, T.E. 2011. A sauropod dinosaur pes from the latest Cretaceous of North America and the validity of Alamosaurus sanjuanensis (Sauropoda, Titanosauria). Journal of Vertebrate Paleontology, 31:1072-1079. https://doi.org/10.1080/02724634.2011.595856
Dalla Vecchia, F.M. 2009. Tethyshadros insularis, a new hadrosauroid dinosaur (Ornithischia) from the Upper Cretaceous of Italy. Journal of Vertebrate Paleontology, 2(4):1100-1116. https://doi.org/10.1671/039.029.0428
Dane, C.H. 1929. Upper Cretaceous formations of southwestern Arkansas. Arkansas Geological Survey Bulletin, 1:215.
Darrough, G.E., Fix, M.F., Parris, D.C., and Grandstaff, B.S. 2005.Chronister Dinosaur Site investigations: New information on the Cretaceous of Missouri. Journal of Vertebrate Paleontology, 25(3):49A.
Denton, R.K., O’Neill, R.C., Grandstaff, B.S., and Parris, D.C. 2011. The Ellisdale Site (Late Cretaceous, Campanian) - is there a rationale for an “Ellisdalean” land faunal age?. Geological Society of America Abstracts with Programs, 43(2):85.
DePalma, R.A., Burnham, D.A, Martin, L.A., Larson, P.L., and Bakker, R.T. 2015. The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation. Paleontological Contributions, 14:1-16.
Dobie, J.L. 1978. A fossil amniote egg from an Upper Cretaceous deposit (Mooreville Chalk of the Selma Group) in Alabama. Copeia, 3:460-464.
Dorr, J.A. 1985. Newfound Early Cretaceous dinosaurs and other fossils in southeastern Idaho and westernmost Wyoming. Contributions from the Museum of Paleontology: University of Michigan, 27(3):73-85.
Doyle, J.A. and Hickey, L.J. 1976. Pollen and leaves from the Mid-Cretaceous Potomac Group and their bearing on early angiosperm evolution. Origin and Early Evolution of Angiosperms :136-206.
Doyle, J.A. and Robbins, E.T. 1977. Angiosperm pollen donation of the continental Cretaceous of the Atlantic Coastal Plain and its application to deep wells in the Salisbury Embayment. Palynology, 1:43-78.
Eaton, T.H. 1960. A new armored dinosaur from the Cretaceous of Kansas. The University of Kansas Paleontological Contributions: Vertebrata, 8:1-24.
Ebersole, S.M. 2009. Biostratigraphy, Paleogeography, and Paleoenvironments of the Upper Cretaceous (Campanian) Northern Mississippi Embayment. PhD Dissertation, University of Alabama, Tuscaloosa, Alabama, USA.
Ebersole, S.M. and King, J.L. 2011. A review of non-avian dinosaurs from the Late Cretaceous of Alabama, Mississippi, Georgia, and Tennessee. Bulletin of the Alabama Museum of Natural History, 28:81-93.
Eberth, D.A. and Deino, A.L. 2005. New 40Ar/39Ar ages from three bentonites in the Bearpaw, Horseshoe Canyon, and Scollard formations (Upper Cretaceous-Paleocene) of southern Alberta, Canada. In Braman, D.R., Therrien, F., Koppelhus, E.B., and Taylor, W. (eds.), Dinosaur Park Symposium: Short Papers, Abstracts, and Program, p. 23-24. Royal Tyrrell Museum of Paleontology, Drumheller, Alberta, Canada.
Eberth, D.A. and Hamblin, A.H. 1993. Tectonic, stratigraphic, and sedimentological significance of a regional disconformity in the upper Judith River Formation (Belly River Wedge) of southern Alberta, Saskatchewan, and northern Montana. Canadian Journal of Earth Science, 30:174-200. https://doi.org/10.1139/e93-016
Everhart, M.J. 2005. Oceans of Kansas: A Natural History of the Western Interior Sea. Indiana University Press, Bloomington, Indiana, USA.
Everhart, M.J. and Ewell, K. 2006. Shark-bitten dinosaur (Hadrosauridae) vertebrae from the Niobrara Chalk (Upper Coniacian) of western Kansas. Transactions of the Kansas Academy of Sciences, 109(1-2):27-35. https://doi.org/10.1660/0022-8443(2006)109%5B27:SDHCVF%5D2.0.CO;2
Everhart, M.J. and Hamm, S.A. 2005. A new nodosaur specimen (Dinosauria: Nodosauridae) from the Smoky Hill Chalk (Upper Cretaceous) of western Kansas. Transactions of the Kansas Academy of Science, 108(1/2):15-21.
Fallow, J.O., O’Brien, M., Kuban, G.J., Dattilo, B.F., Bates, K.T., Falkingham, P.L., Piñuela, L., Rose, A., Freels, A., Kumagai, C., Libben, C., Smith, J., and Whitcraft, J. 2010. Dinosaur tracksites of the Paluxy River (Glen Rose Formation, Lower Cretaceous), Dinosaur Valley State Park, Somervell County, Texas, USA. Actas de V Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno, p. 15-16. Salas de los Infantes, Burgos, Spain.
Farke, A.A. and Phillips, G.E. 2017. The first reported ceratopsid dinosaur from eastern North America (Owl Creek Formation, Upper Cretaceous, Mississippi, USA). PeerJ, 5:e3342. https://doi.org/10.7717/peerj.3342
Farke, A.A., Ryan, M.J., Barrett, P.M., Tanke, D.H., Braman, D.R., Loewen, M.A., and Graham, M.R. 2011. A new centrosaurine from the Late Cretaceous of Alberta, Canada, and the evolution of parietal ornamentation in horned dinosaurs. Acta Palaeontologica Polonica, 56(4):691-702.https://doi.org/10.4202/app.2010.0121
Ferrusquía-Villafranca, I. and Applegate, S.P. 1978. Mesozoic volcanosedimentary rocks and dinosaur tracks in the southwest Pacific region of Mexico. Revista - Instituto de Geología, Universidad Nacional Autónoma de México, 2(2):150-162.
Fiorillo, A.R. and Tykoski, R.S.T. 2012. A new species of the centrosaurine ceratopsid Pachyrhinosaurus from the North Slope (Prince Creek Formation: Maastrichtian) of Alaska. Acta Palaeontologica Polonica, 57:561-573. https://doi.org/10.4202/app.2011.0033
Fiorillo, A.R. and Tykoski, R.S.T. 2014. A diminutive new tyrannosaur from the top of the world. PLoS ONE, 9(3):e91287. https://doi.org/10.1371/journal.pone.0091287
Fix, M.F. and Darrough, G.E. 2004. Dinosauria and associated vertebrate fauna of the Late Cretaceous Chronister Site of southeast Missouri. Geological Society of America Abstracts with Programs, 36(3):14.
Forster, C.A. 1984. The paleoecology of the ornithopod dinosaur Tenontosaurus tilletti from the Cloverly Formation, Big Horn Basin of Wyoming and Montana. The Mosasaur, 2:151-163.
Forster, C.A. 1990. The postcranial skeleton of the ornithopod dinosaur Tenontosaurus tilletti. Journal of Vertebrate Paleontology, 10: 273-294.
Freedman Fowler, E.A. and Horner, J.R. 2015. A new brachylophosaurine hadrosaur (Dinosauria: Ornithischia) with an intermediate nasal crest from the Campanian Judith River Formation of northcentral Montana. PLoS ONE, 10(11):e0141304. https://doi.org/10.1371/journal.pone.0141304
Gallagher, W.B. 1993. The Cretaceous-Tertiary mass extinction event in North Atlantic Coastal Plain. The Mosasaur, 5:75-154.
Gallagher, W.B. 1995. Evidence of juvenile dinosaurs and dinosaurian growth stages in the Late Cretaceous deposits of the Atlantic Coastal Plain. Bulletin of the New Jersey Academy of Science, 40:5-8.
Gallagher, W.B. 1997. When Dinosaurs Roamed New Jersey. Rutgers University Press, New Brunswick, New Jersey.
Galton, P.M. and Jensen, J.A. 1979. Remains of ornithopod dinosaurs from the Lower Cretaceous of North America. Brigham Young University Geological Studies, 25:1-10.
Gangloff, R.A. Dinosaurs under the Aurora. Bloomington, Indiana University Press, 2012.
Garrison, J.R. and Brinkman, D.B. 2007. A multidisciplinary study of the Lower Cretaceous Cedar Mountain Formation, Mussentuchit Wash, Utah: a determination of the paleoenvironment and paleoecology of the Eolambia caroljonesa dinosaur quarry. Cretaceous Research, 28:461-494. https://doi.org/10.1016/j.cretres.2006.07.007
Gates, T.A., Horner, J.R., Hanna, R.R., and Nelson, C.R. 2011. New unadorned hadrosaurine hadrosaurid (Dinosauria, Ornithopoda) from the Campanian of North America. Journal of Vertebrate Paleontology, 31(4):798-811. https://doi.org/10.1080/02724634.2011.577854
Gates, T.A., Jinnah, Z., Levitt, C., and Getty, M.A. 2014. New hadrosaurid specimens from the lower-middle Campanian Wahweap Formation of Utah, p. 156-173. In Eberth, D.A. and Evans, D.C. (eds.), Hadrosaurs. Indiana University Press, Bloomington, Indiana, USA.
Gates, T.A., Prieto-Márquez, A., and Zanno, L.E. 2012. Mountain building triggered Late Cretaceous North American megaherbivore dinosaur radiation. PLoS ONE, 7(8):e42135. https://doi.org/10.1371/journal.pone.0042135
Gates, T.A., Sampson, S.D., Zanno, L.E., Roberts, E.M., Eaton, J.G., Nydam, R.L., Hutchison, J.H., Smith, J.A., Loewen, M.A., and Getty, M.A. 2010. Biogeography of terrestrial and freshwater vertebrates from the Late Cretaceous (Campanian) Western Interior of North America. Palaeogeography, Palaeoclimatology, Palaeoecology, 291: 371-387. https://doi.org/10.1016/j.palaeo.2010.03.008
Gierlinski G. and Lockley, M.G. 2013. A trackmaker for Saurexallopus: Ichnological evidence for oviraptorosaurian tracks from the Upper Cretaceous of western North America, p. 526-529. In Titus, A.L. and Loewen, M.A. (eds.), At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah. Indiana University Press, Bloomington, Indiana, USA.
Gignac, P.M. and Makovicky, P.J. 2010. A description of Deinonychus antirrhopus bite marks and estimates of bite force using tooth indentation simulations. Journal of Vertebrate Paleontology, 30(4):1169-1177.
Gilmore, C.W. 1920. Osteology of the carnivorous Dinosauria in the United States National Museum, with special reference to the genera Antrodemus (Allosaurus) and Ceratosaurus. Bulletin of the United States National Museum, 60:1-154.
Gilmore, C.W. 1945. Parrosaurus, n. name, replacing Neosaurus Gilmore, 1945. Journal of Paleontology 19(5):540.
Gilmore, C.W. and Stewart, D.R. 1945. A new sauropod dinosaur from the Upper Cretaceous of Missouri. Journal of Paleontology, 19(1):23-29.
Gould, C.N. 1929. Comanchean reptiles from Kansas, Oklahoma, and Texas. Bulletin of the Geological Society of America, 40:457-462.
Grandstaff, B.S., Parris, D.C., Denton, R.K., and Gallagher, W.B. 1992. Alphadon (Marsupialia) and Multituberculata (Allotheria) in the Cretaceous of eastern North America. Journal of Vertebrate Paleontology, 12(2):217-222. https://doi.org/10.1080/02724634.1992.10011450
Hanson, R.E., Befus, K.S., Breyer, J.A., Busbey, A.B., Giffin, W.R., Hargrove, U.S., and Lehman, T.M. 2006. Volcanology, geochronology, and stratigraphic framework of Upper Cretaceous basaltic maar deposits in the Big Bend Area of West Texas. Geological Society of America Abstracts with Programs, 38(7):564.
Harris, B. and Self-Trail, J. 2006. Late Cretaceous base level lowering in Campanian and Maastrichtian depositional sequences, Kure Beach, North Carolina. Stratigraphy, 3(3):195-216.
Harris, J.D. 1998. A reanalysis of Acrocanthosaurus atokensis, its phylogenetic status, and paleobiogeographic implications, based on a new specimen from Texas. New Mexico Museum of Natural History and Science Bulletin, 13:1-75.
Hartstein, E.F. and Decina, L.E. 1986. A new Severn Formation (Early Middle Maastrichtian, Late Cretaceous) locality in Prince Georges County, Maryland. The Mosasaur, 3:87-95.
Hawthorne, J.M. and Bonem, R.M. 2002. Ichnology, stratigraphy and paleoenvironment of the Boerne Lake spillway dinosaur tracksite, south-central Texas. Texas Journal of Science, 54(4):309-324.
Hazel, J.E., Edwards, L.E., and Bybell, L.M. 1984. Significant unconformities and the hiatuses represented by them in the Paleogene of the Atlantic and Gulf Coastal Provinces. In Schlee, J. (ed.), Interregional Unconformities. American Association of Petroleum Geologists Memoir, 36:59-66.
Hill, R.T. 1894. Geology of parts of Texas: Indian Territory and Arkansas adjacent to the Red River region. Bulletin of the Geological Society of America, 5:297-380.
Holtz, T.R., Molnar, R.E., and Currie, P.J. 2004. Basal Tetanurae, p 71-110. In Weishampel, D.B., Dodson, P., and Osmólska, H. (eds.), The Dinosauria, 2nd Edition. University of California Press, Berkeley, California, USA.
Horner, J.R., Goodwin, M.B., and Myhrvold, N. 2011. Dinosaur census reveals abundant Tyrannosaurus and rare ontogenetic stages in the Upper Cretaceous Hell Creek Formation (Maastrichtian), Montana, USA. PLoS ONE, 6(2):e16574. https://doi.org/10.1371/journal.pone.0016574
Horner, J.R., Schmitt, J.G., Jackson, F., and Hanna, R. 2001. Bones and rocks of the Upper Cretaceous Two Medicine-Judith River Clastic Wedge Complex, Montana. In Hill, C.L. (ed.), Guidebook for the Field Trips, Society of Vertebrate Paleontology 61st Annual Meeting: Mesozoic and Cenozoic Paleontology in the Western Plains and Rocky Mountains, Museum of the Rockies Occasional Paper, pp. 3-14. Museum of the Rockies, Bozeman, Montana, USA.
Horner, J.R., Varricchio, D.J., and Goodwin, M.B. 1992. Marine transgressions and the evolution of Cretaceous dinosaurs. Nature, 358:59-61. https://doi.org/10.1038/358059a0
Horner, J.R., Weishampel, D.B., and Forster, C.A. 2004. Hadrosauridae, p 438-463. In Weishampel, D.B., Dodson, P., and Osmólska, H. (eds.), The Dinosauria, 2nd Edition. University of California Press, Berkeley, California, USA.
Houston, S.H. 1933. Fossil footprints in Comanchean limestone beds, Bandera County, Texas. Journal of Geology, 41(6):650-653.
Hunt-Foster, R.K. 2003. An Early Cretaceous theropod foot from southwestern Arkansas: Proceedings Journal of the Arkansas Undergraduate Research Conference, 10:87-103.
Hunt-Foster, R.K. and Kirkand, J.I. Evaluating the enigmatic Early Cretaceous ornithomimosaur record in North America. Society of Vertebrate Paleontology Annual Meeting Abstracts :133.
Hunt, A.P. and Lucas, S.G. 2003. A new hadrosaur track from the Upper Cretaceous Fruitland Formation of northwestern New Mexico. New Mexico Geological Society Guidebook, 54th Field Conference, Geology of the Zuni Plateau, 379:379-381.
Jaccard, P. 1902. Lois de distribution florale. Bulletin de la Socíeté Vaudoise des Sciences Naturelles, 38:67-130. (in French).
Jaccard, P. 1912. The distribution of the flora in the alpine zone. New Phytologist, 11(2):37-50.
Jacobs, L.L., Winkler, D.W., Murry, P.A., and Maurice, J.M. 1994. A nodosaurid scuteling from the Texas shore of the Western Interior Seaway, p. 337-346. In Carpenter, K., Hirsch, K.F., and Horner, J. (eds.), Dinosaur Eggs and Babies. Cambridge University Press, Cambridge, Massachusetts, USA.
Jasinski, S.E. and Sullivan, R.M. 2011. Re-evaluation of pachycephalosaurids from the Fruitland-Kirtland transition (Kirtlandian, late Campanian), San Juan Basin, New Mexico, with a description of a new species of Stegoceras and a reassessment of Texacephale langstoni. New Mexico Museum of Natural History and Science Bulletin, 53:202-215.
Jensen, J.A. 1984. Continuing study of new Jurassic/Cretaceous vertebrate faunas from Colorado and Utah. National Geographic Society Research Reports, 16:373-381.
Jinnah, Z.A., Roberts, E.M., Deino, A.L., Larson, J.S., Link, P.K., and Fanning, C.M. 2009. New 40Ar-39Ar and detrital zircon U-Pb ages for the Upper Cretaceous Wahweap and Kaiparowits formations on the Kaiparowits Plateau, Utah: implications for regional correlation, provenance, and biostratigraphy. Cretaceous Research, 30:287-299. https://doi.org/10.1016/j.cretres.2008.07.012
Joeckel, R.M., Cunningham, J.M., Corner, R.G., Brown, G.W., Phillips, P.L., and Ludvigson, G. A. 2004. Late Albian dinosaur tracks from the cratonic (eastern) margin of the Western Interior seaway, Nebraska, USA. Ichnos, 11:275-284. https://doi.org/10.1080/10420940490442377
Johnson, R.O.1974. Lithofacies and Depositional Environments of the Rush Creek Member of the Woodbine Formation (Gulfian) of North Central Texas. M.Sc. Thesis, The University of Texas at Arlington, Arlington, Texas, USA.
Johnson, K.R., Nichols, D.J., and Hartman, J.H. 2002. Hell Creek Formation: A 2001 synthesis. Geological Society of America Special Paper, 361:503-510.
Kaye, J.M. and Russell, D.A. 1973. The oldest record of hadrosaurian dinosaurs in North America. Journal of Paleontology, 47:91-93.
Kennedy, W.J., Landman, N.H., Cobban, W.A., and Johnson, R.O. 2000. Additions to the ammonite fauna of the Upper Cretaceous Navesink Formation of New Jersey. American Museum Novitates, 3306:1-30.
Kiernan, K. and Schwimmer, D.R. 2004. First record of a velociraptorine theropod (Tetanurae, Dromaeosauridae) from the eastern Gulf Coastal United States. The Mosasaur, 7:89-93.
Kimyai, A. 1966. New plant microfossils from the Raritan Formation (Cretaceous) in New Jersey. Micropaleontology, 12(4):461-476.
King, D.T., Abbott-King, J.P., Bell, G.L., Lamb, J.P., Dobie, J.L., and Womochel, D.R. 1988. Stratigraphy and depositional environments of the Turnipseed dinosaur site in the Upper Cretaceous Demopolis Chalk of Montgomery County, Alabama. Journal of the Alabama Academy of Sciences, 59:34-48.
Kinneer, B., Carpenter, K., and Shaw, A. 2016. Redescription of Gastonia burgei (Dinosauria: Ankylosauria, Polacanthidae), and description of a new species. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, 282 (1):37-80.
Kirkland, J.I. 1998. A new hadrosaurid from the upper Cedar Mountain Formation (Albian-Cenomanian: Cretaceous) of eastern Utah - the oldest known hadrosaurid (lambeosaurine?). In Lucas, S.G., Kirkland, J.I., and Estep, J.W. (eds.), Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin, 14:283-295.
Kirkland, J.I., Alcalá, L., Loewen, M.A., Espílez, E., Mampel, L., and Wiersma, J.P. 2013. The basal nodosaurid ankylosaur Europelta carbonensis n. gen., n. sp. from the Lower Cretaceous (Lower Albian) Escucha Formation of northeastern Spain. PLoS ONE, 8(12):e80405. https://doi.org/10.1371/journal.pone.0080405
Kirkland, J.I., Britt, B., Burge, D.L., Carpenter, K., Cifelli, R., DeCourten, F., Eaton, J., Hasiotis, S., and Lawton, T. 1997. Lower to Middle Cretaceous dinosaur faunas of the central Colorado Plateau: a key to understanding 35 million years of tectonics, sedimentology, evolution, and biogeography. Brigham Young University Geology Studies, 42(2):69-103.
Kirkland, J.I., Cifelli, R.L., Britt, B.B., Burge, D.L., DeCourten F.L., Eaton, J.G., and Parrish, J.M. 1999. Distribution of vertebrate faunas in the Cedar Mountain Formation, east-central Utah, p. 210-218. In Gillette, D.D. (ed.), Vertebrate Paleontology in Utah. Utah Geological Survey Miscellaneous Publication, 99-1.
Kirkland, J.I. and Madsen, S.K., 2007. The Lower Cretaceous Cedar Mountain Formation of eastern Utah: the view up an always interesting learning curve. In Lund, W.R. (ed.), Field Guide to Geological Excursions in southern Utah. Utah Geological Association Publication, 35:1-108.
Kirkland, J.I. and Wolfe, D.G. 2001. First definitive therizinosaurid (Dinosauria; Theropoda) from North America. Journal of Vertebrate Paleontology, 21(3):410-414.
Kranz, P.M. 1996. Notes on the sedimentary iron ores of Maryland and their dinosaurian fauna. Maryland Geological Survey Special Paper, 3:87-115.
Kranz, P.M. 1998. Mostly dinosaurs: a review of the vertebrates of the Potomac Group (Aptian Arundel Formation), USA. In Lucas, S.G., Kirkland, J.I., and Estep, J.W. (eds.), Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin, 14:235-238.
Krumenacker, L. 2010. Chronostratigraphy and Paleontology of the Mid-Cretaceous Wayan Formation of Eastern Idaho, with a Description of the First Oryctodromeus Specimens from Idaho. M.Sc. Thesis, Brigham Young University, Provo, Utah, USA.
Krumenacker, L.J., Simon, D.J., Scofield, G., and Varricchio, D.J. 2017. Theropod dinosaurs from the Albian-Cenomanian Wayan Formation of eastern Idaho. Historical Biology, 29(2): 170-186. https://doi.org/10.1080/08912963.2015.1137913
Kuban, G.J. 1989. Color distinctions and other curious features of dinosaur tracks near Glen Rose, Texas, p. 428-440. In Gillette, D.D. and Lockley, M.G., (eds.), Dinosaur Tracks and Traces. Cambridge University Press, Cambridge, Massachusetts, USA.
Lamb, J.P. 1996. Ankylosauria from the Upper Cretaceous of Alabama. Journal of Vertebrate Paleontology, 16(suppl.):58A.
Lamb, J.P. 1997. Marsh was right: Ichthyornis had a beak! Journal of Vertebrate Paleontology, 17(3):59A.
Lamb, J.P. 1998. Lophorhothon, an iguanodontian, not a hadrosaur. Journal of Vertebrate Paleontology, 18(3):58A.
Lamb, J.P. 2001. Dinosaur egg with embryo from the Cretaceous (Campanian) Mooreville Chalk Formation, Alabama. Journal of Vertebrate Paleontology, 21(3):70-71A.
Lamb, J.P., Chiappe, L.M., and Ericson, P.G.P. 1993. A marine Enantionithine from the Cretaceous of Alabama. Journal of Vertebrate Paleontology, 13(3):45A.
Langston, W. 1960. The vertebrate fauna of the Selma Formation of Alabama, part VI: the dinosaurs. Fieldiana: Geological Memoirs, 3(5):315-359.
Langston, W. 1974. Nonmammalian Comanchean tetrapods. Geoscience and Man, 8:77-102.
Larkin, P. 1910. The occurrence of a sauropod dinosaur in the Trinity Cretaceous of Oklahoma. Journal of Geology, 17:93-98.
Lee, Y.N. 1996. A new nodosaurid ankylosaur (Dinosauria: Ornithischia) from the Paw Paw Formation (Late Albian) of Texas. Journal of Vertebrate Paleontology, 16:232-345. https://doi.org/10.1080/02724634.1996.10011311
Lee, Y.N. 1997a. The Archosauria from the Woodbine Formation (Cenomanian) in Texas. Journal of Paleontology, 71(6):1147-1156.
Lee, Y.N. 1997b. Bird and dinosaur footprints in the Woodbine Formation (Cenomanian), Texas. Cretaceous Research, 18:849-864. https://doi.org/10.1006/cres.1997.0091
Lehman, T.M. 1997. Late Campanian dinosaur biogeography in the western interior of North America, p. 223-240. In Wolberg, D. and Stump, E. (eds.), Dinofest International Proceedings. Philadelphia Academy of Natural Sciences, Philadelphia, Pennsylvania, USA.
Lehman, T.M., 2001. Late Cretaceous dinosaur provinciality, p. 310-328. In Tanke, D. and Carpenter, K. (eds.), Mesozoic Vertebrate Life. Indiana University Press, Bloomington, Indiana, USA.
Leidy J. 1858. Hadrosaurus foulkii, a new saurian from the Cretaceous of New Jersey, related to Iguanodon. Proceedings of the Academy of Natural Sciences, 10:213-218.
Liggett, G.A. 2005. A review of the dinosaurs from Kansas. Transactions of the Kansas Academy of Science, 108(1/2):1-14.
Lindgren, J., Currie, P.J., Siverson, M., Rees, J., Cederström, P., and Lindgren, F. 2007. The first neoceratopsian dinosaur remains from Europe. Palaeontology, 50(4):929-937. https://doi.org/10.1111/j.1475-4983.2007.00690.x.
Lipka, T.R. 1998. The affinities of the enigmatic theropods of the Arundel Clay facies (Aptian), Potomac Formation, Atlantic Coastal Plain of Maryland. In Lucas, S.G., Kirkland, J.I., and Estep J.W., (eds.), Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin, 14:229-234.
Liu, K. 2005. Facies changes of the Eutaw Formation (Coniacian-Santonian), onshore to offshore, Northeastern Gulf of Mexico area. Gulf Coast Association of Geological Societies Transactions, 55:431-441.
Lloyd, G.T., Young, J.R., and Smith, A.B. 2012. Taxonomic structure of the fossil record is shaped by sampling bias. Systematic Biology, 61(1):80-89. https://doi.org/10.1093/sysbio/syr076
Lockley, M.G., Cart, K., Martin, J., and Milner, A. 2011. New theropod tracksites from the Upper Cretaceous “Mesaverde” Group, western Colorado: implications for ornithomimosaur track morphology. New Mexico Museum of Natural History and Science Bulletin, 53:321-329.
Lockley, M.G., Meyer, C.A., and Santon, V.F. 1998. Megalosauripus and the problematic concept of megalosaur footprints. Gaia, 15:313-337.
Lockley, M.G. and Stanford, R. 2004. Maryland’s Early Cretaceous dinosaur diversity as revealed by a track bonanza. Geological Society of America Abstracts with Programs, 36(2):116.
Lockley, M.G., Wright, J.L., and Matsukawa, M. 2001. A new look at Magnoavipes and so-called “Big Bird” tracks from Dinosaur Ridge (Cretaceous, Colorado). Mountain Geologist, 38: 137-146.
Loewen, M.A., Irmis, R.B., Sertich, J.J.W., Currie, P.J., and Sampson, S.D. 2013. Tyrant dinosaur evolution tracks the rise and fall of Late Cretaceous oceans. PLoS ONE, 8(11):e79420. https://doi.org/10.1371/journal.pone.0079420
Longrich, N.R. 2011. Titanoceratops ouranos, a giant horned dinosaur from the Late Campanian of New Mexico. Cretaceous Research, 32(3): 264-276. https://doi.org/10.1016/j.cretres.2010.12.007
Longrich, N.R. 2013. Judiceratops tigris, a new horned dinosaur from the Middle Campanian Judith River Formation of Montana. Bulletin of the Peabody Museum of Natural History, 54:51-65.
Longrich, N.R. 2014. The horned dinosaurs Pentaceratops and Kosmoceratops from the upper Campanian of Alberta and implications for dinosaur biogeography. Cretaceous Research, 51: 292-308. https://doi.org/10.1016/j.cretres.2014.06.011
Longrich, N.R. 2016. A ceratopsian dinosaur from the Late Cretaceous of eastern North America, and implications for dinosaur biogeography. Cretaceous Research, 57:199-207. https://doi.org/10.1016/j.cretres.2015.08.004
Longrich, N.R., Barnes, K., Clark, S., and Millar, L. 2013. Caenagnathidae from the Upper Campanian Aguja Formation of West Texas, and a revision of the Caenagnathinae. Bulletin of the Peabody Museum of Natural History, 54:23-49. https://doi.org/10.3374/014.054.0102
Lucas, S.G. and Hunt, A.P. 1989. Alamosaurus and the sauropod hiatus in the Cretaceous of the North American Western Interior. Geological Society of America Special Paper, 238:75-85.
Lucas, S.G., Hunt, A.P., and Sullivan, R.M. 2006. Stratigraphy and age of the Upper Cretaceous Fruitland Formation, west-central San Juan Basin, New Mexico. In Lucas, S.G. and Sullivan, R.M. (eds.), Late Cretaceous Vertebrates from the Western Interior, p. 1-6. New Mexico Museum of Natural History and Science, Albuquerque, New Mexico, USA.
Lucas, S.G., Sullivan, R.M., Lichtig, A.J., Dalman S.G., and Jasinski S.E. 2016. Late Cretaceous dinosaur biogeography and endemism in the Western Interior basin, North America: A critical re-evaluation. New Mexico Museum of Natural History and Science Bulletin, 71:195-213.
Lull, R.S. 1911. Systematic paleontology of the Lower Cretaceous deposits of Maryland: Vertebrata. Maryland Geological Survey, Lower Cretaceous Volume :183-211.
Lund, E.K., O’Connor, P.M., Loewen, M.A., and Jinnah, Z.A. 2016. A new centrosaurine ceratopsid, Machairoceratops cronusi gen et sp. nov., from the Upper Sand Member of the Wahweap Formation (Middle Campanian), southern Utah. PLoS ONE, 11(5):e0154403. https://doi.org/10.1371/journal.pone.0154403
Main, D.J. 2005. Paleoenvironments and aleoecology of the Cenomanian Woodbine Formation of Texas: Paleobiogeography of the Hadrosaurs (Dinosauria: Ornithischia). M.Sc. Thesis, The University of Texas at Arlington, Arlington, Texas, USA.
Main, D.J. 2013. Appalachian Delta Plain Paleoecology of the Cretaceous Woodbine Formation at the Arlington Archosaur Site in North Texas. PhD Dissertation, University of Texas at Arlington, Arlington, Texas, USA.
Main, D.J., Noto, C.R., and Drumheller, S.K. 2012. Crocodiles of the Texas Cretaceous: The Campanian of Big Bend to the Cenomanian of North Texas, a comparison of great size, feeding behavior and paleoecology. Geological Society of America Abstracts with Programs, 44(1):3.
Makovicky, P.J., Kobayashi, Y., and Currie, P.J. 2004. Ornithomimosauria, p. 137-150. In Weishampel, D.B., Dodson, P., and Osmólska, H. (eds.), The Dinosauria, 2nd Edition. University of California Press, Berkeley, California, USA.
Makovicky, P.J. and Sues, H.D. 1998. Anatomy and phylogenetic relationships of the theropod dinosaur Microvenator celer from the Lower Cretaceous of Montana. American Museum Novitates 3240:1-27.
Mallon, J.C. and Anderson, J.S. 2013. Skull ecomorphology of megaherbivorous dinosaurs from the Dinosaur Park Formation (Upper Campanian) of Alberta, Canada. PLoS ONE, 8(7):e67182. https://doi.org/10.1371/journal.pone.0067182
Mallon, J.C., Ott, C.J., Larson, P.L., Iuliano, E.M., and Evans, D.C. 2016. Spiclypeus shipporum gen. et sp. nov., a boldly audacious new chasmosaurine ceratopsid (Dinosauria: Ornithischia) from the Judith River Formation (Upper Cretaceous: Campanian) of Montana, USA. PLoS ONE, 11(5):e0154218. https://doi.org/10.1371/journal.pone.0154218
Mannion, P. D. and Upchurch, P. 2011. A re-evaluation of the ‘mid-Cretaceous sauropod hiatus’ and the impact of uneven sampling of the fossil record on patterns of regional dinosaur extinction. Palaeogeography, Palaeoclimatology, Palaeoecology, 299:529-540. https://doi.org/10.1016/j.palaeo.2010.12.003
Mannion, P.D., Upchurch, P., Barnes, R.N., and Mateus, O. 2013. Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms. Zoological Journal of the Linnean Society, 168:98-206.
Marsh, O.C. 1888. Notice of a new genus of Sauropoda and other new dinosaurs from the Potomac Formation. American Journal of Science, (Series 3) 35:89-94.
Marsh, O.C. 1889. Notice of gigantic horned Dinosauria from the Cretaceous. American Journal of Sciences, 38:173-175.
McDonald, A.T., Bird, J., Kirkland, J.I., and Dodson, P. 2012. Osteology of the basal hadrosauroid Eolambia caroljonesa (Dinosauria: Ornithopoda) from the Cedar Mountain Formation of Utah. PLoS ONE, 7(10):e45712. https://doi.org/10.1371/journal.pone.0045712
McDonald, A.T., Kirkland, J.I., DeBlieux, D.D., Madsen, S.K., Cavin, J., Milner, A.R.C., and Panzarin, L. 2010. New basal iguanodonts from the Cedar Mountain Formation of Utah and the evolution of thumb-spiked dinosaurs. PLoS ONE, 5(11):e14075. https://doi.org/10.1371/journal.pone.0014075
McDonald, A.T., Wolfe, D.G., and Kirkland, J.I. 2010. A new basal hadrosauroid (Dinosauria: Ornithopoda) from the Turonian of New Mexico. Journal of Vertebrate Paleontology, 30(3): 799-812. https://doi.org/10.1080/02724631003763516
Mehl, M.G. 1936. Hierosaurus coleii: a new aquatic dinosaur from the Niobrara Cretaceous of Kansas. Denison University Bulletin, 31(3):1-20.
Miller, H.W. 1967. Cretaceous vertebrates from Phoebus Landing, North Carolina. Proceedings of the Academy of Natural Sciences, 119:219-239.
Miller, K.G., Sugarman, P.J., Browning, J.V., Kominz, M.A., Olsson, R.K., Feigenson, M.D., and Hernandez, J.C. 2004. Upper Cretaceous sequences and sea-level history, New Jersey Coastal Plain. Bulletin of the Geological Society of America, 116(3):368-393.
Mori, H. 2009. Dinosaurian Faunas of the Cedar Mountain Formation and LA-ICP-MS Detrital Zircon Ages for Three Stratigraphic Sections. MSc, Brigham Young University. All Theses and Dissertations. http://scholarsarchive.byu.edu/etd/2000.
Mori, H., Druckenmiller, P.S., and Erickson, G.M. 2015. A new Arctic hadrosaurid from the Prince Creek Formation (lower Maastrichtian) of northern Alaska. Acta Palaeontologica Polonica, 61(1):15-32. https://doi.org/10.4202/app.00152.2015
Nicholls, E.L. and Russell, A.P. 1990. Paleobiogeography of the Cretaceous Western Interior Seaway of North America; the vertebrate evidence. Palaeogeography, Palaeoclimatology, Palaeoecology, 79:(1-2):149-169. https://doi.org/10.1016/0031-0182(90)90110-S
Norman, D.B. 2004. Basal Iguanodontia, p. 413-437. In Weishampel, D.B., Dodson, P., and Osmólska, H. (eds.), The Dinosauria, 2nd Edition. University of California Press, Berkeley, California, USA.
Noto, C.R., Main, D.J., and Drumheller, S.K. 2012. Feeding traces and paleobiology of a Cretaceous (Cenomanian) crocodyliform: Example from the Woodbine Formation of Texas. Palaios, 27(2):1-11.
Oliver, W.B. 1971. Depositional systems within the Woodbine Formation (Upper Cretaceous), northeast Texas. Reports of Investigation, Bureau of Economic Geology, The University of Texas at Austin, 73:1-28.
Olson, S.L. 1975. Ichthyornis in the Cretaceous of Alabama. Wilson Bulletin, 87: 103-105.
Oreska, M.P.J., Carrano, M.T., and Dzikiewicz, K.M. 2013. Vertebrate paleontology of the Cloverly Formation (Lower Cretaceous), I: faunal composition, biogeographic relationships, and sampling. Journal of Vertebrate Paleontology, 33(2):264-292. https://doi.org/10.1080/02724634.2012.717567
Ostrom, J.H. 1969. A new theropod dinosaur from the Lower Cretaceous of Montana. Postilla, 128:1-17.
Ostrom, J.H. 1970. Stratigraphy and paleontology of the Cloverly Formation (Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana. Bulletin of the Peabody Museum of Natural History, 35:1-234.
Ostrom, J.H. 1976. On a new specimen of the Lower Cretaceous theropod dinosaur Deinonychus antirrhopus. Breviora, 439:1-21.
Parsons, W.L. and Parsons, K.M. 2009. A new ankylosaur (Dinosauria: Ankylosauria) from the Lower Cretaceous Cloverly Formation of central Montana. Canadian Journal of Earth Sciences, 46(10):721-738. https://doi.org/10.1139/E09-045
Pasch, A.D. and May, K.C. 1998. First occurrence of a hadrosaur (Dinosauria) from the Matanuska Formation (Turonian) in the Talkeetna Mountains of south-central Alaska. In Clough, J.G. and Larson, F. (eds.), Short notes on Alaska geology 1997. Alaska Division of Geological and Geophysical Surveys Professional Report, 118H:99-109.
Paulina-Carabajal, A., Lee, Y.N., and Jacobs, L.L. 2016. Endocranial morphology of the primitive nodosaurid dinosaur Pawpawsaurus campbelli from the Early Cretaceous of North America. PLoS ONE, 11(3):e0150845. https://doi.org/10.1371/journal.pone.0150845
Payenberg, T.H.D., Braman, D.R., Davis, D.W., and Miall, A.D. 2002. Litho- and chronostratigraphic relationships of the Santonian-Campanian Milk River Formation in southern Alberta and Eagle Formation in Montana utilising stratigraphy, U-Pb geochronology, and palynology. Canadian Journal of Earth Sciences, 39:1553-1577. https://doi.org/10.1139/e02-050
Penkalski, P. and Blows, W.T. 2013. Scolosaurus cutleri (Ornithischia: Ankylosauria) from the Upper Cretaceous Dinosaur Park Formation of Alberta, Canada. Canadian Journal of Earth Sciences, 50(2):171-182. https://doi.org/10.1139/cjes-2012-0098
Pittman, J.G. 1989. Stratigraphy, lithology, depositional environment, and track type of dinosaur track-bearing beds of the Gulf Coastal Plain, p. 135-153. In Gillette, D.D. and Lockley, M.G. (eds.), Dinosaur Tracks and Traces. Cambridge University Press, Cambridge, Massachusetts, USA.
Prieto-Márquez A. 2011. Revised diagnoses of Hadrosaurus foulkii Leidy, 1858 (the type genus and species of Hadrosauridae Cope, 1869) and Claosaurus agilis Marsh, 1872 (Dinosauria: Ornithopoda) from the Late Cretaceous of North America. Zootaxa, 2765:61-68.
Prieto-Marquez, A. and Norell, M.A. 2010. Anatomy and relationships of Gilmoreosaurus mongoliensis (Dinosauria: Hadrosauroidea) from the Late Cretaceous of Central Asia. American Museum Novitates, 3694:1-52.
Prieto-Márquez, A., Erickson, G.M., and Ebersole, J.A. 2016. A primitive hadrosaurid from southeastern North America and the origin and early evolution of 'duck-billed' dinosaurs. Journal of Vertebrate Paleontology, 36(2):e1054495.
Prieto-Márquez, A., Erickson, G.M., and Ebersole, J.A. 2016. Anatomy and osteohistology of the basal hadrosaurid dinosaur Eotrachodon from the uppermost Santonian (Cretaceous) of southern Appalachia. PeerJ, 4:e1872.
Prieto-Márquez, A., Weishampel, D.B., and Horner, J.R. 2006. The dinosaur Hadrosaurus foulkii, from the Campanian of the East Coast of North America, with a reevaluation of the genus. Acta Palaeontologica Polonica, 51(1):77-98.
Quinn, J.H. 1973. Arkansas Dinosaur. Geological Society of America Abstracts with Programs, 5(3):276-277.
Ratkevich, R. 1998. New Cretaceous brachiosaurid dinosaur, Sonorasaurus thompsoni gen et sp. nov, from Arizona. Arizona-Nevada Academy of Science Bulletin, 31:71-82.
Robbins, E.I. 1991. Age of Early Cretaceous palynomorphs in the Muirkirk clay pit fossil locality (Prince George’s County, MD). US Geological Survey Open-File Report, 91-613:1-7.
Roberts, E.M., Deino, A.L., and Chan, M.A., 2005. 40Ar/39Ar age of the Kaiparowits Formation, southern Utah, and correlation of contemporaneous Campanian strata and vertebrate faunas along the margin of the Western Interior Basin. Cretaceous Research, 26:307-318.
Roberts, L.N.R. and Kirschbaum, M.A. 1995. Paleogeography of the Late Cretaceous of the Western Interior of middle North America-Coal distribution and sediment accumulation. U.S. Geological Survey Professional Paper, 1561:1-115.
Rogers, J.V. 2003. Pachycheilosuchus trinquei, a new procoelous crocodyliform from the Lower Cretaceous (Albian) Glen Rose Formation of Texas. Journal of Vertebrate Paleontology, 23(1):128-145.
Rogers, R.R., Swisher, C.C., and Horner, J.R. 1993. 40Ar/39Ar age and correlation of the nonmarine Two Medicine Formation (Upper Cretaceous), northwestern Montana. Canadian Journal of Earth Science, 30:1066-1075.
Russell, D.A. 1972. Ostrich dinosaurs from the Late Cretaceous of western Canada. Canadian Journal of Earth Sciences, 9:375-402.
Russell, D.A. 1995. China and the lost worlds of the dinosaurian era. Historical Biology, 10(1):3-12.
Ryan, M.J. 2007. A new basal centrosaurine ceratopsid from the Oldman Formation, southeastern Alberta. Journal of Paleontology, 81(2):376-396.
Ryan, M.J., Evans, D.C., Currie, P.J., Brown, C.M., and Brinkman, D. 2012. New leptoceratopsids from the Upper Cretaceous of Alberta, Canada. Cretaceous Research, 35:69-80.
Ryan, M.J., Evans, D.C., Currie, P.J., and Loewen, M.A. 2014. A New chasmosaurine from northern Laramidia expands frill disparity in ceratopsid dinosaurs. Naturwissenschaften, 101: 505.
Ryan, M.J., Russell, A.P., and Hartman, S. 2010. A new chasmosaurine ceratopsid from the Judith River Formation, Montana, p. 181-188. In Ryan, M.J., Chinnery-Allgeier, N.J., and Eberth, D.A. (eds.), New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium. Indiana University Press, Bloomington, Indiana, USA.
Sampson, S.D., Loewen, M.A., Farke, A.A., Roberts, E.M., Forster, C.A., Smith, J.A., and Titus, A.L. 2010. New horned dinosaurs from Utah provide evidence for intracontinental dinosaur endemism. PLoS ONE, 5(9):e12292.
Sampson, S.D., Loewen, M.A., Roberts, E.M., and Getty, M.A. 2013. A new macrovertebrate assemblage from the Late Cretaceous (Campanian) of southern Utah, p. 599-617. In Titus, A.L. and Loewen, M.A. (eds.), At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah. Indiana University Press, Bloomington, Indiana, USA.
Sampson, S.D., Lund, E.K., Loewen, M.A., Farke, A.A., and Clayton, K.E. 2013. A remarkable short-snouted horned dinosaur from the Late Cretaceous (late Campanian) of southern Laramidia. Proceedings of the Royal Society B: Biological Sciences, 280(1766):20131186.
Sanders, H.L. 1968. Marine benthic diversity: A comparative study. The American Naturalist, 102(925):243.
Schröder-Adams, C. 2014. The Cretaceous Polar and Western Interior seas: paleoenvironmental history and paleoceanographic linkages. Sedimentary Geology, 301:26-40. https://doi.org/10.1016/j.sedgeo.2013.12.003
Schwimmer, D.R. 1986. Late Cretaceous fossils from the Blufftown Formation (Campanian) in Western Georgia. The Mosasaur, 3:109-123.
Schwimmer, D.R. 1997. Late Cretaceous dinosaurs in Eastern USA: A taphonomic and biogeographic model of occurrences, p. 203-211. In Wolberg, E. and Stump, E. (eds.), Dinofest International Proceedings. Philadelphia Academy of Natural Sciences, Philadelphia, Pennsylvania, USA.
Schwimmer, D.R. 2002. King of the Crocodylians: The Paleobiology of Deinosuchus. Indiana University Press, Bloomington, Indiana, USA.
Schwimmer, D.R. 2016. Was there a southeastern dinosaur province in the Late Cretaceous?. Geological Society of America Abstracts with Programs, 48(3):22-23.
Schwimmer, D.R., Sanders, A.E., Erickson, B.R., and Weems, R.E. 2015. A Late Cretaceous dinosaur and reptile assemblage from South Carolina, USA. Transactions of the American Philosophical Society, 105(2):1-157.
Schwimmer, D.R. and Williams, G.D. 1993. A giant crocodile from Alabama and observations on the paleobiology of southeastern crocodilians. Journal of Vertebrate Paleontology, 13(suppl. 3):56A.
Schwimmer, D.R. and Williams, G.D. 1997. New Specimens of Deinosuchus rugosus, and further insights into chelonivory by Late Cretaceous eusuchian crocodiles. Journal of Vertebrate Paleontology, 16(suppl. 3):64A.
Schwimmer, D.R., Williams, G.D., Dobie, J.L., and Siesser, W.G. 1993. Late Cretaceous dinosaurs from the Blufftown Formation in western Georgia and eastern Alabama. Journal of Vertebrate Paleontology, 67(2):288-296.
Scott, R.W., Fee, D., Magee, R., and Laalli, H. 1978. Epeiric depositional models for the Lower Cretaceous Washita Group. Reports of Investigation, Bureau of Economic Geology, The University of Texas at Austin, 94:1-23.
Self-Trail, J.M., Christopher, R.A., Prowell, D., and Weems, R.E. 2004. The age of dinosaur-bearing strata at Phoebus Landing, Cape Fear River, North Carolina. Geological Society of America Abstracts with Programs, 36(2):117.
Sellards, E.W., Adkins, W.A., and Plummer, F.B. 1932. The Geology of Texas. Bulletin of the University of Texas, 3232:315-316.
Simpson, G.G. 1943. Mammals and the nature of continents. American Journal of Science, 241:1-31.
Smith, A.B. 2001. Large-scale heterogeneity of the fossil record: implications for Phanerozoic biodiversity studies. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 356:351-367. https://doi.org/10.1098%2Frstb.2000.0768
Smith, A.B. 2007. Marine diversity through the Phanerozoic: problems and prospects. Journal of the Geological Society of London, 164:731-745. https://doi.org/10.1144/0016/76492006-184
Smith, D. and Galton, P.M. 1990. Osteology of Archaeornithomimus asiaticus (Upper Cretaceous, Iren Dabasu Formation, People’s Republic of China). Journal of Vertebrate Paleontology, 10:255-265. https://doi.org/10.1080/02724634.1990.10011811
Stanford, R. 1998. Feathered theropods in the Smithsonian’s “back yard”? p. 56. In Wolberg, D.L., Gittis, K., Miller, S., Carey, L., and Raynor, A. (eds.), The Dinofest Symposium. Academy of Natural Sciences, Philadelphia, Pennsylvania, USA.
Stanford, R. and Stanford, S.M. 1998.... and the sauropod wore sneakers!, p. 57. In Wolberg, D.L., Gittis, K., Miller, S., Carey, L., and Raynor, A. (eds.), The Dinofest Symposium. Academy of Natural Sciences, Philadelphia, Pennsylvania, USA.
Stanford, R., Weems, R.E., and Lockley, M.G. 2004. A new dinosaur ichnotaxon from the Lower Cretaceous Patuxent Formation of Maryland and Virginia. Ichnos, 11:251-259. https://doi.org/10.1080/10420940490428797
Stanford, R., Weishampel, D.B., and DeLeon, V.B. 2011. The first hatchling dinosaur reported from the Eastern United States: Propanoplosaurus marylandicus (Dinosauria: Ankylosauria) from the Early Cretaceous of Maryland, USA. Journal of Vertebrate Paleontology, 85(5):916-924. https://doi.org/10.1666/10-113.1
Storer, J.E. 1975. Dinosaur tracks, Columbosauripus ungulatus (Saurischia: Coelurosauria), from the Dunvegan Formation (Cenomanian) of northeastern British Columbia. Canadian Journal of Earth Sciences, 12:1805-1807. https://doi.org/10.1139/e75-160
Stovall, J.W. and Langston, W. 1950. Acrocanthosaurus atokensis, a new genus and species of Lower Cretaceous Theropoda from Oklahoma. American Midland Naturalist, 43:686-728.
Sugarman, P.J., Miller, K.G., Burky, D., and Feigenson, M.D. 1995. Uppermost Campanian-Maastrichtian strontium isotopic; biostratigraphic and sequence stratigraphic framework of the New Jersey Coastal Plain. Geological Society of America Bulletin, 107:19-37. https://doi.org/10.1130/0016-7606(1995)107%3C0019:UCMSIB%3E2.3.CO;2
Sullivan, R.M. 2003. Revision of the dinosaur Stegoceras Lambe (Ornithischia, Pachycephalosauridae). Journal of Vertebrate Paleontology, 23(1):181-207. https://doi.org/10.1671/0272-4634(2003)23[181:ROTDSL]2.0.CO;2
Sullivan, R.M. 2006. A taxonomic review of the Pachycephalosauridae (Dinosauria: Ornithischia). New Mexico Museum of Natural History and Science Bulletin, 35:347-365.
Sullivan, R.M. and Lucas, S.G. 2006. The Kirtlandian land-vertebrate “age”-faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of western North America. In Lucas, S.G. and Sullivan, R.M. (eds.), Late Cretaceous vertebrates from the Western Interior. New Mexico Museum of Natural History and Science Bulletin 35:7-29.
Taylor, M.P., Wedel, M.J., and Cifelli, R.L. 2011. Brontomerus mcintoshi, a new sauropod dinosaur from the Lower Cretaceous Cedar Mountain Formation, Utah, USA. Acta Palaeontologica Polonica, 56(1):75-98. https://doi.org/10.4202/app.2010.0073
Thayer, D.W. and Ratkevich, R.P. 1995. In-progress dinosaur excavation in the mid-Cretaceous Turney Ranch Formation, southeastern Arizona. Mesa Southwest Museum and Southwest Paleontological Society. Proceedings of the Fossils of Arizona Symposium III, Mesa, Arizona, USA.
Troyer, R., Barth, A.P., Wooden, J.L., and Jacobson, C. 2006. Provenance and timing of Sevier Foreland Basin sediments in the Valley of Fire, southern Nevada, from U-Pb geochronology. Geological Society of America Abstracts with Programs, 38(7):369.
Trudel, P. 1994. Stratigraphic Sequences and Facies Architecture of the Woodbine - Eagle Ford Interval, Upper Cretaceous, North Central Texas. Masters Thesis, Tarleton State University.
Ullman, P.V., Varricchio, D., and Knell, M.J. 2012. Taphonomy and taxonomy of a vertebrate microsite in the Mid-Cretaceous (Albian-Cenomanian) Blackleaf Formation, southwest Montana. Historical Biology, 24:311-328. https://doi.org/10.1080/08912963.2011.602405
Vance, T. 2002. Annotated checklist and bibliography of the prehistoric reptile and dinosaur tracks and trackways of Texas. Occasional Papers of the Dallas Paleontological Society, 5:17-79.
Varricchio, D.J., Martin A.J., and Katsura, Y. 2007. First trace and body fossil evidence of a burrowing, denning dinosaur. Proceedings of the Royal Society B: Biological Sciences 274(1616):1361-1368. https://doi.org/10.1098/rspb.2006.0443
Vavrek, M.J., Hills, L.V., and Currie, P.J. 2014. A hadrosaurid (Dinosauria: Ornithischia) from the Late Cretaceous (Campanian) Kanguk Formation of Axel Heiberg Island, Nunavut, Canada, and its ecological and geographical implications. Arctic, 67(1):1-9. https://doi.org/10.14430/arctic4362
Vavrek, M.J., and Larsson, H.C.E., 2010. Low beta diversity of Maastrichtian dinosaurs of North America. Proceedings of the National Academy of Science, 107(18):8265-8268. https://doi.org/10.1073/pnas.0913645107
Vickaryous, M.K., Maryańska, T., and Weishampel, D.B. 2004. Ankylosauria, p. 363-392. In Weishampel, D.B., Dodson, P., and Osmólska, H. (eds.), The Dinosauria, 2nd Edition. University of California Press, Berkeley, California, USA.
Watanabe, A., Erickson, G.M., and Druckenmiller, P.S. 2013. An ornithomimosaurian from the upper Cretaceous Prince Creek Formation of Alaska. Journal of Vertebrate Paleontology, 33: 1169-1175. https://doi.org/10.1080/02724634.2013.770750
Wedel, M.J., Cifelli, R.L., and Sanders, R.K. 2000. Sauroposeidon proteles, a new sauropod from the Early Cretaceous of Oklahoma. Journal of Vertebrate Paleontology, 20(1):109-114. https://doi.org/10.1671/0272-4634(2000)020%5B0109:SPANSF%5D2.0.CO;2
Weems, R.E. and Bachman, J.M. 1997. Cretaceous anuran and dinosaur footprints from the Patuxent Formation of Virginia. Proceedings of the Biological Society of Washington, 110(1):1-17.
Weems, R.E. and Bachman, J.M. 2015. The Lower Cretaceous Patuxent Formation Ichnofauna of Virginia. Ichnos, 22:208-219. https://doi.org/10.1080/10420940.2015.1063493
Weishampel, D.B. 2006. Another look at the dinosaurs of the East Coast of North America, p. 129-168. In ‘Coletivo Arqueológico-Paleontológico Salense, (eds.), Actas III Jornadas Dinosaurios Entorno. Salas de los Infantes, Burgos, Spain.
Weishampel, D.B. and Young, L. 1996. Dinosaurs of the East Coast. Johns Hopkins University Press, Baltimore, Maryland, USA.
Weishampel, D.B., Barrett, P.M., Coria, R.A., Loeuff, J.L., Xing, X., Xijin, Z., Sahni, A., Gomani, E.M.P., and Noto, C.R. 2004. Dinosaur Distribution, p. 517-617. In Weishampel, D.B., Dodson, P., and Osmólska, H. (eds.), The Dinosauria, 2nd Edition. University of California Press, Berkeley, California, USA.
Weishampel, D.B., Meers, M.B., Akerston, W.A., and McCrady, A.D. 2002. New Early Cretaceous dinosaur remains, including possible ceratopsians, from the Wayan Formation of eastern Idaho. Idaho Museum of Natural History Occasional Paper, 37:5-17.
Wenhao, W. and Godefroit, P. 2012. Anatomy and relationships of Bolong yixianensis, an Early Cretaceous Iguanodontoid Dinosaur from Western Liaoning, China, p. 293-333. In Godefroit, P. (ed.), Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems. Indiana University Press, Bloomington, Indiana, USA.
West, A. and Tibert, N. 2004. Quantitative analysis for the type material of Priconodon crassus: a distinct taxon from the Arundel Formation in souther Maryland. Geological Society of America Abstracts with Programs, 36(5):423.
Wieland, G.R. 1909. A new armored saurian from the Niobrara. American Journal of Sciences 27(4):250-252.
Williston, S.W. 1905. A new armored dinosaur from the Upper Cretaceous of Wyoming. Science, 22(564):503-504.
Winkler, D.A., Murry, P.A., and Jacobs, L.L. 1997. A new species of Tenontosaurus (Dinosauria: Ornithopoda) from the Early Cretaceous of Texas. Journal of Vertebrate Paleontology, 17: 330-348. https://doi.org/10.1080/02724634.1997.10010978
Woodward, H.N. and Lehman, T.M. 2008. Bone histology and microanatomy of Alamosaurus sanjuanensis (Sauropoda: Titanosauria) from the Maastrichtian of Big Bend National Park, Texas. Journal of Vertebrate Paleontology, 29(3):807-821. https://doi.org/10.1671/039.029.0310
Wolfe, D.G. and Kirkland, J.I. 1998. Zuniceratops christopheri n. gen. and n. sp., a ceratopsian dinosaur from the Moreno Hill Formation (Cretaceous, Turonian) of west-central New Mexico. In Lucas, S.G., Kirkland, J.I., and Estep, J.W. (eds.), Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin, 14:307-317.
Woodruff, D.C. 2012. A new titanosauriform from the Early Cretaceous Cloverly Formation of Montana. Cretaceous Research, 36:58-66. https://doi.org/10.1016/j.cretres.2012.02.003
Wrather, W.E. 1922. Dinosaur tracks in Hamilton County, Texas. Journal of Geology, 30(5):354-360.
Zaleha, M.J. 2006. Sevier orogenesis and nonmarine basin filling: Implications of new stratigraphic correlations of Lower Cretaceous strata throughout Wyoming, USA. Geological Society of America Bulletin, 118:886-896. https://doi.org/10.1130/B25715.1
Zanno, L.E. and Makovicky, P.J. 2013. Neovenatorid theropods are apex predators in the Late Cretaceous of North America. Nature Communications, 4:1-9. https://doi.org/10.1038/ncomms3827
Zanno, L.E., Gillette, D.D., Albright, L.B., and Titus, A.L. 2009. A new North American therizinosaurid and the role of herbivory in 'predatory' dinosaur evolution. Proceedings of the Royal Society B: Biological Sciences, 276:3505-3511. https://doi.org/10.1098/rspb.2009.1029
