

 Postcranial anatomy of the Spanish Upper Triassic sauropterygian Paludidraco multidentatus (Simosauridae)
Postcranial anatomy of the Spanish Upper Triassic sauropterygian Paludidraco multidentatus (Simosauridae)
Article number: 27.2.a40
https://doi.org/10.26879/1405
Copyright Palaeontological Association, August 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Appendices
Submission: 14 May 2024. Acceptance: 31 July 2024.
ABSTRACT
Simosauridae is an eosauropterygian clade known by two representatives: Simosaurus gaillardoti, from the Ladinian (Middle Triassic) of France and Germany, and Paludidraco multidentatus, from the Carnian (Upper Triassic) of Spain. However, the fossil record of this clade is relatively abundant, being mostly represented by isolated cranial and postcranial material, which ranges from the uppermost Anisian or the lowermost Ladinian of the Middle East to the Carnian of Europe. Therefore, the clade is considered as exclusive of the western region of the Paleotethys. Information about the postcranial anatomy of the group is mostly restricted to that corresponding to S. gaillardoti, even though the holotype of P. multidentatus preserves a large part of its postcranial skeleton, very well preserved. Thus, a detailed description of this bizarre taxon has not been performed so far. We provide a comprehensive description of the postcranium of the holotype of P. multidentatus, which has been carefully prepared for this study. The detailed comparative study with the postcranial skeleton of S. gaillardoti allows us to improve knowledge about it, as well as about Simosauridae in relation to other sauropterygian clades.
Alberto Cabezuelo-Hernández. Grupo de Biología Evolutiva, Departamento de Física Matemática y de Fluidos. Facultad de Ciencias, Universidad Nacional de Educación a Distancia (UNED), Avda. Esparta s/n, 28232. Las Rozas. Madrid, Spain. (Corresponding author) paleo.alb@gmail.com
ORCID iD: 0000-0002-2990-8384
Carlos De Miguel Chaves. Grupo de Biología Evolutiva, Departamento de Física Matemática y de Fluidos. Facultad de Ciencias, Universidad Nacional de Educación a Distancia (UNED), Avda. Esparta s/n, 28232. Las Rozas. Madrid, Spain. carlosmiguelchaves@gmail.com
ORCID iD: 0000-0002-4002-9540
Francisco Ortega. Grupo de Biología Evolutiva, Departamento de Física Matemática y de Fluidos. Facultad de Ciencias, Universidad Nacional de Educación a Distancia (UNED), Avda. Esparta s/n, 28232. Las Rozas. Madrid, Spain. fortega@ccia.uned.es
ORCID iD: 0000-0002-7431-354X
Adán Pérez-García Grupo de Biología Evolutiva, Departamento de Física Matemática y de Fluidos. Facultad de Ciencias, Universidad Nacional de Educación a Distancia (UNED), Avda. Esparta s/n, 28232. Las Rozas. Madrid, Spain. a.perez.garcia@ccia.uned.es
ORCID iD: 0000-0002-9074-6855
Keywords: El Atance fossil site; Eosauropterygia; Iberian Peninsula; Late Triassic; postcranium; simosaurid
Final citation: Cabezuelo-Hernández, Alberto, De Miguel Chaves, Carlos, Ortega, Francisco, and Pérez-García, Adán. 2024. Postcranial anatomy of the Spanish Upper Triassic sauropterygian Paludidraco multidentatus (Simosauridae). Palaeontologia Electronica, 27 (2):a40.
https://doi.org/10.26879/1405
palaeo-electronica.org/content/2024/5299-postcranium-of-paludidraco
Copyright: August 2024 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Sauropterygia is a group of Mesozoic marine reptiles with a fossil record ranging from the Lower Triassic (Olenekian) (Rieppel, 1994, 1999, 2000; Sennikov, 2001; Bardet et al., 2014) to the uppermost Cretaceous (Maastrichtian) (Rieppel, 2000; Bardet et al., 2014). Simosauridae is a poorly known eosauropterygian clade erected by Huene in 1948, after the description of its type species, Simosaurus gaillardoti Meyer, 1842, from the Ladinian (Middle Triassic) of France and Germany (Rieppel, 1994). This lineage is currently recognized as represented by two valid species: S. gaillardoti and the recently described Paludidraco multidentatus de Miguel Chaves et al., 2018a, from the Carnian (Late Triassic) of Spain (see García-Ávila et al., 2021 and references therein). Material from this lineage of indeterminate specific and even generic attribution has been identified in several Middle East and European Middle to Late Triassic sites. In this sense, the oldest records of Simosauridae correspond to the uppermost Anisian or the lowermost Ladinian of Israel (Haas, 1981; Rieppel et al., 1999) and Saudi Arabia (Vickers-Rich et al., 1999; Kear et al., 2010), whereas the youngest ones comes from the Carnian of Italy (Dalla Vecchia, 2006, 2008, 2017), Germany (Rieppel, 1996, 2000), and Spain (de Miguel Chaves et al., 2015, 2018a, 2020a, b; García-Ávila et al., 2021). Simosauridae is, therefore, an eosauropterygian clade exclusively known in the western region of the Paleotethys (de Miguel Chaves et al., 2020a, b).
Postcranial remains of Simosauridae are relatively abundant, but they mostly correspond to isolated elements. By contrast, scarce articulated skeletons or associated elements of a single individual are known for this clade. Both the cranial and the postcranial anatomy of S. gaillardoti is relatively well known, having been described and figured in previous studies, for more than a century (Jaekel, 1905; Huene, 1921, 1948, 1952, 1959; Kuhn-Schnyder, 1961; Rieppel, 1994, 2000; de Miguel Chaves et al., 2020a). On the contrary, the anatomy of the recently defined P. multidentatus was only briefly described in de Miguel Chaves et al., (2018a), based on a relatively complete skeleton (the holotype) and an isolated skull (the paratype). This Spanish taxon was interpreted as a filter feeder with slow-moving habits (de Miguel Chaves et al., 2018a), notably differing from the active predatory lifestyle recognized for S. gaillardoti. In this context, the markedly different cranial and postcranial anatomy of these two simosaurid representatives suggests a high ecological disparity in the clade (de Miguel Chaves et al., 2018a). Due to the scarce information so far available about this strange taxon, and the good preservation of the skeleton corresponding to its holotype, a detailed preparation has been performed. This allows us to present here an exhaustive description of the postcranium of P. multidentatus, based on the reference specimen (i.e., its holotype). For its characterization, detailed anatomical comparisons with the postcranium of its sister taxon S. gaillardoti, as well as with that of other sauropterygians, are performed. Thus, knowledge about the postcranial anatomy of one of the main sauropterygian lineages known from the Triassic record of both Europe and the Middle East (i.e., Simosauridae) is notably increased. The information obtained in this work, along with that of future studies involving other P. multidentatus specimens (see García-Ávila et al., 2021) currently under preparation and study, will allow a complete inclusion of this taxon in a phylogenetical analysis and provide an updated phylogeny for Simosauridae.
Institutional abbreviations. GBA, Geologische Bundesanstalt, Vienna, Austria; GPIT-PV, Paleontological Collection of the Geologisch-Paläontologisches Institut, Tübingen Universität, Tübingen, Germany; HUJ-PAL, Paleontological collection of the Hebrew University of Jerusalem, Jerusalem, Israel; MFSN, Museo Friulano di Storia Naturale, Udine, Italy; MNHN, Muséum National d’Histoire Naturelle, Paris, France; MUPA-ATZ, El Atance collection, Museo de Paleontología de Castilla-La Mancha, Cuenca, Spain; SMNS, Staatliches Museum für Naturkunde Stuttgart, Stuttgart, Germany.
Anatomical abbreviations. ac, acetabulum; ap, anterior process; c, centrum; cf, coracoid facet; clc, clavicular corner; clf, clavicular facet; cll, clavicular lamina; clp, clavicular process; clr, clavicular ramus; cof, coracoid foramen; cr, cervical rib; dc, deltopectoral crest; diap; diapophysis; dps, deltopectoral scar; dr, dorsal rib; f, foramen; fe, femur; gl, glenoid; icl, interclavicle; iclr, interclavicular ramus; ipr, infraprezygapophysis; iprf, infraprezygapophyseal facet; ipt, infrapost-zygapophysis; iptf, infrapostzygapophyseal facet; isf, iliac facet for the ischium; itr, inner trochanter; j, jaw; lcl, left clavicle; lco, left coracoid; lg, longitudinal grooves; lil, left ilium; lis, left ischium; lpu, left pubis; na, neural arch; nc, neural canal; nk, neural keel; ns, neural spine; par, parapophysis; pr, prezygapohysis; prf, prezygapophyseal facet; prl, prespinal lamina; pt, postzygapophysis; ptf, postzygapophyseal facet; ptl, postspinal lamina; puf, iliac facet for the pubis; rcl, right clavicle; rco, right coracoid; rfe, right femur; ril, right ilium; rsc, right scapula; sab, supracetabular buttress; scg, scapular groove; sy, symphysis; tp, transverse process; vk, ventral keel; zs, zygosphene; zt, zygantrum; zw, zygantral wedge.
MATERIALS AND METHODS
 The holotype of Paludidraco multidentatus, MUPA-ATZ0101, comes from the Keuper Facies of El Atance fossil site (Guadalajara, Central Spain), corresponding to the base of the Julian substage, i.e., the basal term of the Carnian (García-Ávila et al., 2021). It was discovered and collected during a field campaign, in 2015, and was briefly described in a short note, in which the taxon P. multidentatus was defined (see de Miguel Chaves et al., 2018a). Most postcranial remains of MUPA-ATZ0101 were collected in two blocks (Figure 1A-B’; Figure 2A-B’). Additionally, in the case of having suffered disarticulation in the biostratinomic phase, several elements from this individual were either removed from the skeleton during its extraction in the field or were removed during the preparation (see Appendix 1, Figure S1). The larger block has been prepared in both dorsal and ventral views (Figure 1A-B’); this region of the skeleton being currently exposed in this last view. Much of this preparation has been carried out after the original description of the taxon (de Miguel Chaves et al., 2018a), to facilitate the detailed study carried out here. Most elements of that block remain articulated as originally found in the field, except those extracted with the aim of providing a complete and detailed characterization, or some that were not preserved in articulation (see Appendix 1, Figure S1). The block preserves elements from the cervical region to the posterior dorsal area (Figure 1A-B’). The proximal region of the block displays posterior cervical vertebrae, for which the most anterior ones are poorly preserved. The elements corresponding to the cervicals are identified, in Figure 1, Figure 3-Figure 4, and in Appendix 1, Figures S1-2, as ‘C’ (cervical vertebra) and ‘cc’ (cervical centrum), followed by a ‘#’ sign plus a numeral as their exact position within the cervical series is unknown (from #1 to #10). The cervical elements from #1 to #5 were collected in the field from a more anterior position of this individual. They preserved a continuous, although inexact, anatomical position. Hence, they are named with a ‘#’ sign followed by a correlative number (Figure 3; see Appendix 1, Figure S1). Most dorsal vertebrae are preserved in articulation. Following the previous nomenclature, they are identified, in Figure 1, Figure 4 and in Figures S1-3 in the Appendix 1, as ‘D’ (dorsal vertebra), ‘dc’ (dorsal centrum) and ‘da’ (dorsal neural arch), followed by a numeral when their exact position is known, and by a ‘#’ sign plus a numeral when their position, although relatively continuous in the dorsal series, cannot be precisely determined (from #24 to #41) (Figure 5, Figure 6, Figure 7; see Appendix 1, Figure S1). All the disarticulated dorsal centra are identified as ‘centrum’ followed by a ‘#’ sign plus a numeral (from #1 to #9) (Figure 8; see Appendix 1, Figure S1), since their exact correspondence to their neural arches is unknown. The disarticulated ribs figured in this study follow the same nomenclature as the disarticulated centra (see Appendix 1, Figure S1).
The holotype of Paludidraco multidentatus, MUPA-ATZ0101, comes from the Keuper Facies of El Atance fossil site (Guadalajara, Central Spain), corresponding to the base of the Julian substage, i.e., the basal term of the Carnian (García-Ávila et al., 2021). It was discovered and collected during a field campaign, in 2015, and was briefly described in a short note, in which the taxon P. multidentatus was defined (see de Miguel Chaves et al., 2018a). Most postcranial remains of MUPA-ATZ0101 were collected in two blocks (Figure 1A-B’; Figure 2A-B’). Additionally, in the case of having suffered disarticulation in the biostratinomic phase, several elements from this individual were either removed from the skeleton during its extraction in the field or were removed during the preparation (see Appendix 1, Figure S1). The larger block has been prepared in both dorsal and ventral views (Figure 1A-B’); this region of the skeleton being currently exposed in this last view. Much of this preparation has been carried out after the original description of the taxon (de Miguel Chaves et al., 2018a), to facilitate the detailed study carried out here. Most elements of that block remain articulated as originally found in the field, except those extracted with the aim of providing a complete and detailed characterization, or some that were not preserved in articulation (see Appendix 1, Figure S1). The block preserves elements from the cervical region to the posterior dorsal area (Figure 1A-B’). The proximal region of the block displays posterior cervical vertebrae, for which the most anterior ones are poorly preserved. The elements corresponding to the cervicals are identified, in Figure 1, Figure 3-Figure 4, and in Appendix 1, Figures S1-2, as ‘C’ (cervical vertebra) and ‘cc’ (cervical centrum), followed by a ‘#’ sign plus a numeral as their exact position within the cervical series is unknown (from #1 to #10). The cervical elements from #1 to #5 were collected in the field from a more anterior position of this individual. They preserved a continuous, although inexact, anatomical position. Hence, they are named with a ‘#’ sign followed by a correlative number (Figure 3; see Appendix 1, Figure S1). Most dorsal vertebrae are preserved in articulation. Following the previous nomenclature, they are identified, in Figure 1, Figure 4 and in Figures S1-3 in the Appendix 1, as ‘D’ (dorsal vertebra), ‘dc’ (dorsal centrum) and ‘da’ (dorsal neural arch), followed by a numeral when their exact position is known, and by a ‘#’ sign plus a numeral when their position, although relatively continuous in the dorsal series, cannot be precisely determined (from #24 to #41) (Figure 5, Figure 6, Figure 7; see Appendix 1, Figure S1). All the disarticulated dorsal centra are identified as ‘centrum’ followed by a ‘#’ sign plus a numeral (from #1 to #9) (Figure 8; see Appendix 1, Figure S1), since their exact correspondence to their neural arches is unknown. The disarticulated ribs figured in this study follow the same nomenclature as the disarticulated centra (see Appendix 1, Figure S1).
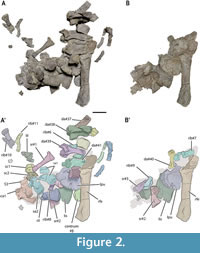 The smaller block (Figure 2A-B’) was completely removed in the laboratory, after the description of the taxon (see de Miguel Chaves et al., 2018a), to allow a detailed characterization of all elements included in it (see Appendix 1, Figure S1). It preserved distal dorsal, sacral, and proximal caudal vertebrae (some of them partially articulated); ribs; isolated gastral fragments; girdle elements; and a femur. Some of the elements from this block suffered from partial distortion and displacement. However, the vertebral elements preserve their original position within the vertebral series (Figure 2A-B’; Appendix 1, Figure S1). The vertebral elements are numbered following the above indicated nomenclature (i.e., using numerals for the elements of known position, and a ‘#’ sign followed by a numeral for the elements with unknown exact position). Thus, the sacral and caudal vertebrae are identified, in Figure 2, Figure 9-Figure 10 and in Figure S1 in the Appendix 1, as ‘S’ (sacral vertebra), ‘sa’ (sacral neural arch), ‘sc’ (sacral centrum), ‘ca’ (caudal neural arch) and ‘cac’ (caudal centrum).
The smaller block (Figure 2A-B’) was completely removed in the laboratory, after the description of the taxon (see de Miguel Chaves et al., 2018a), to allow a detailed characterization of all elements included in it (see Appendix 1, Figure S1). It preserved distal dorsal, sacral, and proximal caudal vertebrae (some of them partially articulated); ribs; isolated gastral fragments; girdle elements; and a femur. Some of the elements from this block suffered from partial distortion and displacement. However, the vertebral elements preserve their original position within the vertebral series (Figure 2A-B’; Appendix 1, Figure S1). The vertebral elements are numbered following the above indicated nomenclature (i.e., using numerals for the elements of known position, and a ‘#’ sign followed by a numeral for the elements with unknown exact position). Thus, the sacral and caudal vertebrae are identified, in Figure 2, Figure 9-Figure 10 and in Figure S1 in the Appendix 1, as ‘S’ (sacral vertebra), ‘sa’ (sacral neural arch), ‘sc’ (sacral centrum), ‘ca’ (caudal neural arch) and ‘cac’ (caudal centrum).
The material was photographed with a Nikon D3500 camera and subsequently processed for the illustrations using both Adobe Photoshop (v. 22.0.1) and Adobe Illustrator. Specimens corresponding to representatives of the main lineages of Sauropterygia (i.e., Placodontia, Pachypleurosauria, Nothosauria, and Pistosauroidea), housed in the collections of the GBA, GPIT-PV, HUJ-PAL, MNHN, and SMNS, were first-hand studied for the exhaustive comparative analyses and the detailed characterization of P. multidentatus performed here, in addition to the information available in the literature. Several measurements were taken for MUPA-ATZ0101, and for several specimens of Simosaurus gaillardoti housed at the SMNS, relevant for the comparative study between both taxa, using a 200 x 0.02 mm caliper (see Appendix 2, Tables S1-S7, Figures S1-S4 for these measurements).
SYSTEMATIC PALAEONTOLOGY
Superorder SAUROPTERYGIA Owen, 1860
Order EOSAUROPTERYGIA Rieppel, 1994
Superfamily NOTHOSAUROIDEA Baur, 1889
Family SIMOSAURIDAE Huene, 1948
Type species. Simosaurus gaillardoti Meyer, 1842
Included species. Simosaurus gaillardoti, Paludidraco multidentatus.
 Distribution. Uppermost Anisian or lowermost Ladinian to Carnian (Middle to Upper Triassic) of Europe and the Middle East (Rieppel, 1994, 1996, 2000; Dalla Vecchia, 2008; Kear et al., 2010; de Miguel Chaves et al., 2015, 2018a, 2020a, b; Segesdi and Ösi, 2021).
Distribution. Uppermost Anisian or lowermost Ladinian to Carnian (Middle to Upper Triassic) of Europe and the Middle East (Rieppel, 1994, 1996, 2000; Dalla Vecchia, 2008; Kear et al., 2010; de Miguel Chaves et al., 2015, 2018a, 2020a, b; Segesdi and Ösi, 2021).
Emended diagnosis (modified from de Miguel Chaves et al., 2020a). Nothosauroidea with the following synapomorphies: relatively short and blunt snout, without rostral constriction; unreduced prefrontals; jugals extending anteriorly along the lateral margin of the orbit; undepressed temporal region; absence of premaxillary and dentary fangs; absence of enlarged teeth on the maxilla; mandibular articulation displaced to a level distinctly behind the occipital condyle; at least 32 dorsal vertebrae; socketed vertebral centra; zygosphene without articular facets; ovoid to oblong transverse processes of the dorsal vertebrae; infraprezygapophyses and infrapostzygapophyses present, at least in the dorsal, the sacral, and the most anterior caudal vertebrae; flattened middle-to-distal shaft of the dorsal ribs; presence of an anteromedial process in the clavicle; presence of a postero-medially expanded lamina in the clavicle; markedly set off iliac blade; and humeral mid-shaft distinctly narrower than the proximal and distal regions. It is also characterized by the following exclusive combination of characters: distinctly constricted vertebral centra (in ventral and lateral views); at least twice high than wide transverse processes of the dorsal vertebrae (not laterally prominent); relatively high and latero-medially constrained neural spines of the dorsal vertebrae; conspicuously ornamented neural spines (with lateral longitudinal grooves in the upper third and apical ornamentation); presence of pre- and postspinal laminae from the middle dorsal to the sacral vertebrae; presence of a zygantral wedge, at least from the dorsal to the sacral vertebrae; medio-laterally expanded distal region of the dorsal ribs; three or four sacral vertebrae; distinct expansion of distal region of sacral ribs (in dorso-ventral direction); absence of entepicondylar foramen; and humerus longer than the femur.
SIMOSAURUS Meyer, 1842
Simosaurus gaillardoti Meyer, 1842
Neotype. MNHN AC.9028, an almost complete skull (figure 50 in Rieppel, 2000; figures 1b, 2b in de Miguel Chaves et al., 2018b).
Locality and horizon. Réhainvillers, Lunéville, France; Upper Muschelkalk, Ladinian (Middle Triassic) (Hagdorn and Rieppel, 1999).
Distribution. Ladinian (Middle Triassic) levels outcropping in an area including northeastern France and southwestern Germany (Rieppel, 1994; de Miguel Chaves et al., 2018b).
Emended diagnosis (modified from de Miguel Chaves et al., 2020a). Member of Simosauridae differing from Paludidraco multidentatus by the following characters: relatively short snout; anteriorly positioned external nares; wide and oval upper temporal fenestra; oval pineal foramen, slightly displaced posteriorly; supraoccipital located below the occipital-parietal exposure; supraoccipital exposed more or less vertically on the occiput; relatively robust jaw; splenial located in the lingual surface of the jaw; teeth located dorsally in the jaw; big, conical, and blunt teeth, anchored in deep sockets (thecodont implantation); five premaxillary teeth; less than 35 dorsal vertebrae; platycoelous centra (except the most anterior cervicals); as high as long cervical to sacral centra; markedly deep neural canal of the vertebral centra (except in the anterior cervicals, unknown for the sacral and caudal vertebrae); strongly constricted dorsal centra (in ventral view); not pachyostotic sensu stricto dorsal vertebrae; transverse processes of the dorsal vertebrae with a ventral notch, and not contacting the articular surface of the centra; short zygantral wedge (from the dorsal to the sacral vertebrae, but absent in the cervicals); vertebral arches without neural keel; laterally expanded postzygapophyses of dorsal vertebrae; absence of infrapre- and infrapostzygapophyes in the cervical vertebrae; bifurcated zygosphene, with a marked ventral indentation; not pachyostotic sensu stricto dorsal ribs; absence of a marked distal expansion of the dorsal ribs; straight or slightly convex distal margin of the dorsal ribs; expanded distal region of the sacral ribs (in cranio-caudal direction); absence of clavicular foramina; T-shaped interclavicle; well-developed coracoid foramen; straight posteromedial margin of the coracoid; postacetabular process not exceeding the posterior margin of the ilium; closed obturator foramen of the pubis; notched pubis; pubis with a prepubic process; slim humerus relative to the femur; deltopectoral scar less than one fifth of the humerus total length; presence of ectepicondylar groove; not reduced spinal width of sacral vertebrae.
PALUDIDRACO de Miguel Chaves, Ortega and Pérez-García, 2018a
Paludidraco multidentatus de Miguel Chaves, Ortega and Pérez-García, 2018a
(Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12, Figure 13, Figure 14, Figure 15)
Type material. Holotype, MUPA-ATZ0101, a relatively complete and partially articulated skeleton, including the skull, both incomplete mandibular rami, most of the dorsal vertebrae and ribs, the sacral and some caudal vertebrae, and elements of the girdles and the appendicular regions (Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12, Figure 13, Figure 14, Figure 15 in this study; figures 1a, 2a-e in de Miguel Chaves et al., 2018a). Paratype, MUPA-ATZ0102, a skull and most of the left lower jaw (figure 2f-I in de Miguel Chaves et al., 2018a).
Type locality and horizon. El Atance fossil site (Sigüenza, Guadalajara, Central Spain). Keuper Facies. Basal Julian, Carnian (Late Triassic) (García-Ávila et al., 2021).
Distribution. Exclusively known in its type locality (de Miguel Chaves et al., 2018a).
Emended diagnosis (modified from de Miguel Chaves et al., 2020a). Member of Simosauridae differing from Simosaurus gaillardoti by the following characters: relatively elongated snout; retracted external nares; narrow upper temporal fenestra; elongated pineal foramen, close to the middle area of the skull table; horizontal exposition of the supraoccipital at the posterior end of the parietal table; supraoccipital horizontally sutured with the parietal; very slender and fragile jaw; splenial located dorsally to the dentary; teeth inserted in pleurodont implantation; numerous small, recurved, and mesiodistally compressed teeth; more than 15 premaxillary teeth; teeth situated laterally in the jaw; at least 40 dorsal vertebrae; amphicoelous centra; higher than long cervical to sacral centra; moderately deep neural canal of the vertebral centra; not strongly constricted dorsal centra (in ventral view); highly pachyostotic sensu stricto dorsal vertebrae; high transverse processes of the dorsal vertebrae, without ventral notch, contacting the articular surface of the centra; elongated zygantral wedge (at least in the posterior dorsal and the sacral vertebrae, unknown for the cervical, the anterior and the middle dorsal, and the caudal vertebrae); vertebral arches with neural keel; not laterally expanded postzygapophyses of dorsal vertebrae; presence of infrapre- and infrapostzygapophyes in the cervical vertebrae; not bifurcated zygosphene, with a slight ventral indentation; pachyostotic dorsal ribs sensu stricto; strong distal expansion of the dorsal ribs; unevenly convex distal margin of the dorsal ribs; not expanded distal region of the sacral ribs (in cranio-caudal direction); presence of two symmetrical clavicular foramina; semi-lunate interclavicle; reduced or absent coracoid foramen; angled posteromedial margin of the coracoid; postacetabular process exceeding the posterior margin of the ilium; open obturator foramen of the pubis; not notched pubis; pubis without prepubic process; massive humerus relative to the femur; deltopectoral scar one third of the humerus total length; absence of ectepicondylar groove; spinal width of sacral vertebrae notably reduced.
DESCRIPTION OF THE POSTCRANIAL SKELETON OF PALUDIDRACO MULTIDENTATUS
Postcranial Axial Skeleton
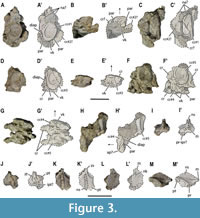
 Cervical vertebrae. At least seven disarticulated centra corresponding to the anterior cervical region (Figure 3A-H’; Appendix 1, Figure S1), and four or five articulated vertebrae corresponding to the most posterior cervical region are identified (Figure 1A, B; Figure 4A-E’). Hence, a minimum of 11 or 12 cervicals is recognized for Paludidraco multidentatus based on the study of its holotype. All identified cervical centra are amphicoelous, with relatively deep concave articular surfaces; and higher than wider, and wider than longer. The neural canal in none of the anterior cervical centra is observable. The less deformed of these centra (i.e., cc#1 in Figure 3A-C’) has a height/length ratio of 2 (see Appendix 2, Table S1). The cervical centra from cc#1 to cc#6 in Figure 3-Figure 4, bear a distinct ventral keel, which separates two ventral fossae, lacking nutritive foramina (Figure 3B-B’, E-E’, G-G’; Figure 4B-B’). A ventral keel is not developed in the posterior cervical centra (this character is unknown in the cervicals cc#7 and cc#8, being confirmed as absent from cc#9; Figure 4B-B’). Two facets for the rib articulation can be distinguished on both the right and the left laterals of the anterior cervical centra: a vertical and dorsally located diapophysis; and a horizontal and more ventrally located parapohysis (Figure 3A-A’, C-C’). These facets are closely situated to each other, being separated by a small circular channel (Figure 3F-F’).
Cervical vertebrae. At least seven disarticulated centra corresponding to the anterior cervical region (Figure 3A-H’; Appendix 1, Figure S1), and four or five articulated vertebrae corresponding to the most posterior cervical region are identified (Figure 1A, B; Figure 4A-E’). Hence, a minimum of 11 or 12 cervicals is recognized for Paludidraco multidentatus based on the study of its holotype. All identified cervical centra are amphicoelous, with relatively deep concave articular surfaces; and higher than wider, and wider than longer. The neural canal in none of the anterior cervical centra is observable. The less deformed of these centra (i.e., cc#1 in Figure 3A-C’) has a height/length ratio of 2 (see Appendix 2, Table S1). The cervical centra from cc#1 to cc#6 in Figure 3-Figure 4, bear a distinct ventral keel, which separates two ventral fossae, lacking nutritive foramina (Figure 3B-B’, E-E’, G-G’; Figure 4B-B’). A ventral keel is not developed in the posterior cervical centra (this character is unknown in the cervicals cc#7 and cc#8, being confirmed as absent from cc#9; Figure 4B-B’). Two facets for the rib articulation can be distinguished on both the right and the left laterals of the anterior cervical centra: a vertical and dorsally located diapophysis; and a horizontal and more ventrally located parapohysis (Figure 3A-A’, C-C’). These facets are closely situated to each other, being separated by a small circular channel (Figure 3F-F’).
Most neural arches of the proximal cervical region are strongly crushed and badly preserved; thus, a detailed description cannot be provided except for one of them (Figure 3I-M’). Small structures that project anteriorly and posteriorly from the main neural arch body correspond to the broken right pre- and postzygapohysis, respectively (Figure 3I-K’, M-M’). The postzygapophysis is placed notably higher than the prezygapophysis (Figure 3M-M’), as in the posterior cervicals (Figure 4D-D’). The presence of infrapre- and infrapostzygapophysis cannot be confirmed in this element (Figure 3I-J’). This neural arch also preserves a zygosphene, but it is badly preserved for a detailed characterization (Figure 3I-I’, K-K’). An incipient and poorly developed ventral keel (termed here as the neural keel) is present in this fragmentary cervical neural arch, at the roof of the neural canal (Figure 3L-L’).
The posterior cervical neural arches are relatively well preserved, and some of the centra corresponding to these vertebrae are still in articulation with them. Their neural spines slant slightly in the posterior direction. They are not latero-medially constrained or widened towards the tip, and are dorso-ventrally short, being almost as high as long (see Figure 4A-B’, D-D’; and Appendix 2, Table S1). The upper third of the spines are conspicuously ornamented, small tubercles and pits being dorsally present, and with the lateral development of longitudinal grooves (Figure 4B-B’, D-E’). The presence of pre- and postspinal laminae in the neural arches cannot be evaluated due to the tight articulation of these elements. The diapophyses of the posterior region of the cervical series (i.e., from vertebra C#8 to C#10 in Figure 4D-D’) gradually shift dorsally to lie on the neural arches, where they develop as incipient transverse processes in the last vertebrae. The last cervical is difficult to identify due to the preservation of this articulated vertebral series. However, the vertebra C#9 in Figure 4B-B’, D-E’ clearly corresponds to the cervical region, as its centrum displays an individualized parapophysis (i.e., not yet fused with the diapophysis) (Figure 4D-D’). The following vertebra (i.e., C#10 in Figure 4B-B’, D-E’) is interpreted here as the last cervical, as its corresponding centrum (i.e., cc#10 in Figure 4B-B’, E-E’) seems to display a poorly developed, small parapophysis in its ventro-lateral side (Figure 4B-B’). This centrum is notably constricted both in lateral and ventral views. The neural canal of cc#10 is slightly hourglass-shaped and moderately deep, two deep depressions developing laterally to it (Figure 4B-B’). Despite some zygosphenes are preserved in this posterior cervical series (Figure 4D-D’), their morphology cannot be described due to their bad preservation. The distance between the prezygapophyses (i.e., the prezygapophyseal distance) of each of these cervicals is clearly greater than the width of the corresponding centrum, but the articulation of these vertebrae hinders the access to this measure. The prezygapophyseal distance is interpreted as probably equivalent to the distance between the postzygapophyses (i.e., the postzygapophyseal distance) in the posterior cervicals, as their postzygapophyses are more laterally projected than those of the dorsal vertebrae (see vertebra C#6 vs vertebra D2; Figure 4A-A’, C-C’). Small infrapre- and infrapostzygapophyses can be identified in the posterior region of the cervical series (e.g., vertebrae C#8, C#10; Figure 4B-B’, D-D’).
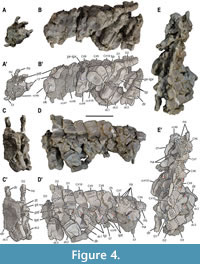
 Dorsal vertebrae. A minimum of 40 or 41 dorsal vertebrae were present in the individual MUPA-ATZ0101 based on the neural arches (even though only 38 dorsal centra are preserved). The vertebral elements in Figure 5, Figure 6, Figure 7, Figure 8 correspond to disarticulated elements of the posterior dorsal region of MUPA-ATZ0101 (see Appendix 1, Figure S1). Most dorsal vertebrae are preserved as articulated elements in the main block (Figure 1A-B’). The first dorsal vertebrae (from D1 to D5) display single rib facets (i.e., transverse processes) shared between the centrum and the neural arch (Figure 1A-A’; Figure 4D-D’; Appendix 1, Figure S2A, B), corresponding to the pectoral vertebrae sensu Seeley (1874).
Dorsal vertebrae. A minimum of 40 or 41 dorsal vertebrae were present in the individual MUPA-ATZ0101 based on the neural arches (even though only 38 dorsal centra are preserved). The vertebral elements in Figure 5, Figure 6, Figure 7, Figure 8 correspond to disarticulated elements of the posterior dorsal region of MUPA-ATZ0101 (see Appendix 1, Figure S1). Most dorsal vertebrae are preserved as articulated elements in the main block (Figure 1A-B’). The first dorsal vertebrae (from D1 to D5) display single rib facets (i.e., transverse processes) shared between the centrum and the neural arch (Figure 1A-A’; Figure 4D-D’; Appendix 1, Figure S2A, B), corresponding to the pectoral vertebrae sensu Seeley (1874).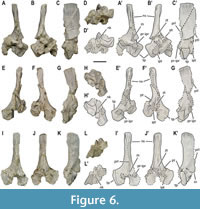 From the vertebra D6 in Figure 1A-A’ and in Figure S2B in the Appendix 1 onwards, the transverse processes are exclusively developed on the neural arch (Figure 1A-A’; Appendix 1, Figure S2B-E). The zygapophyseal articulations form the here termed ‘X-shaped’ wedge-and-socket system: the wedge being formed by the pre-infraprezygapophyseal complex, that fits into the socket-like post-infrapostzygapophyseal complex of the preceding vertebra, which forms a distinct ‘X’ shape (Figure 5, Figure 6, Figure 7; Appendix 1, Figure S2D). The middle dorsal vertebrae show highly pachyostotic sensu stricto (see Houssaye et al., 2008 for terminology) centra, zygapophyses, transverse processes, and neural spines. The pachyostosis sensu stricto is gradually lost towards the posterior dorsal region (see Appendix 1, Figure S2C-I). The neurocentral suture is open throughout the dorsal region, as indicated by the separation of the neural arches from their respective centra.
From the vertebra D6 in Figure 1A-A’ and in Figure S2B in the Appendix 1 onwards, the transverse processes are exclusively developed on the neural arch (Figure 1A-A’; Appendix 1, Figure S2B-E). The zygapophyseal articulations form the here termed ‘X-shaped’ wedge-and-socket system: the wedge being formed by the pre-infraprezygapophyseal complex, that fits into the socket-like post-infrapostzygapophyseal complex of the preceding vertebra, which forms a distinct ‘X’ shape (Figure 5, Figure 6, Figure 7; Appendix 1, Figure S2D). The middle dorsal vertebrae show highly pachyostotic sensu stricto (see Houssaye et al., 2008 for terminology) centra, zygapophyses, transverse processes, and neural spines. The pachyostosis sensu stricto is gradually lost towards the posterior dorsal region (see Appendix 1, Figure S2C-I). The neurocentral suture is open throughout the dorsal region, as indicated by the separation of the neural arches from their respective centra.
 The dorsal centra are amphicoelous and non-notochordal. The articular surfaces of the anterior and middle dorsal centra are slightly oval, higher than wider, and wider than long (Figure 8A-AB). The dorsal centra have a height/length average ratio of 1.5 (see Appendix 2, Table S1). However, the posterior centra are as high as wide, being cranio-caudally shorter than those of the preceding dorsals (Figure 8AC-AJ). They show a height/length average ratio of 1.4 (some are highly deformed to know the value of this ratio). All dorsal centra with an available dorsal view show the eosauropterygian cruciform articulation platform for the neural arch (Rieppel 1994). The dorsal centra from the anterior to the middle region display a relatively deep and wide neural canal with parallel walls (Figure 4D-D’; Figure 8C, G; Appendix 1, Figure S2G, H). The neural canal becomes hourglass shaped and shallower in most posterior dorsal centra (Figure 8AI). As in the cervical centrum cc#10, the dorsal centra develop two depressions laterally to the middle region of the neural canal. In the best-preserved elements, these depressions delimit four triangular areas
The dorsal centra are amphicoelous and non-notochordal. The articular surfaces of the anterior and middle dorsal centra are slightly oval, higher than wider, and wider than long (Figure 8A-AB). The dorsal centra have a height/length average ratio of 1.5 (see Appendix 2, Table S1). However, the posterior centra are as high as wide, being cranio-caudally shorter than those of the preceding dorsals (Figure 8AC-AJ). They show a height/length average ratio of 1.4 (some are highly deformed to know the value of this ratio). All dorsal centra with an available dorsal view show the eosauropterygian cruciform articulation platform for the neural arch (Rieppel 1994). The dorsal centra from the anterior to the middle region display a relatively deep and wide neural canal with parallel walls (Figure 4D-D’; Figure 8C, G; Appendix 1, Figure S2G, H). The neural canal becomes hourglass shaped and shallower in most posterior dorsal centra (Figure 8AI). As in the cervical centrum cc#10, the dorsal centra develop two depressions laterally to the middle region of the neural canal. In the best-preserved elements, these depressions delimit four triangular areas (Figure 8C, G, W; Appendix 1, Figure S2H) (i.e., the ‘socketed’ configuration sensu Sanz 1984). The dorsal centra are from moderately constricted in the anterior dorsal region (in lateral view, the ventral view being unavailable; Figure 1A-A’; Appendix 1, Figure S3A, B) to not or very slightly constricted (in ventral and lateral views) in the middle dorsal region (Figure 8B, D; Appendix 1, Figure S3C), becoming notably constricted in the most posterior dorsal region (e.g., Figure 8J, V, X), where the last centra are distinctly constricted (in ventral and lateral views) (e.g., Figure 8AH, AJ; Appendix 1, Figure S3A, B). The articular facets for the neural arches of the dorsal centra are laterally projected, being anteriorly placed in the mid-dorsal vertebrae and reaching a more central position in the posterior ones. Additionally, some dorsal centra bear small and unpaired foramina on their ventro-lateral surfaces (see Figure 1G).
(Figure 8C, G, W; Appendix 1, Figure S2H) (i.e., the ‘socketed’ configuration sensu Sanz 1984). The dorsal centra are from moderately constricted in the anterior dorsal region (in lateral view, the ventral view being unavailable; Figure 1A-A’; Appendix 1, Figure S3A, B) to not or very slightly constricted (in ventral and lateral views) in the middle dorsal region (Figure 8B, D; Appendix 1, Figure S3C), becoming notably constricted in the most posterior dorsal region (e.g., Figure 8J, V, X), where the last centra are distinctly constricted (in ventral and lateral views) (e.g., Figure 8AH, AJ; Appendix 1, Figure S3A, B). The articular facets for the neural arches of the dorsal centra are laterally projected, being anteriorly placed in the mid-dorsal vertebrae and reaching a more central position in the posterior ones. Additionally, some dorsal centra bear small and unpaired foramina on their ventro-lateral surfaces (see Figure 1G).
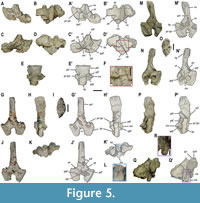
The dorsal neural arches show latero-medially constrained neural spines that gradually widen apically (Figure 1B-B’; Figure 5M-N’; Appendix 1, Figure S3D). The spines are cranio-caudally shorter than higher (height/length average ratio of approximately 2; see Appendix 2, Table S1) (Figure 5, Figure 6, Figure 7), except those of the first dorsals (see vertebrae D1 and D2 in Figure 4D-D’). The neural spines gradually shorten cranio-caudally towards the posterior dorsal region. The height of the neural spines is similar to the width of the neural arch (see Appendix 2, Table S1). The upper third of the spines is conspicuously ornamented with the lateral development of distinct longitudinal grooves (Figure 4D-D’; Figure 5H-H’, P-P’; Figure 6C-C’, G-G’, K-K’). Dorsally, the neural spines of anterior-to-middle dorsal vertebrae display small tubercles and pits, as in the cervicals (Appendix 1, Figure S2E, F), whereas those of posterior dorsal vertebrae display transversal ridges (Figure 5I). The inclination of the neural spines decreases from the anterior to the posterior region of the dorsal series, becoming vertical in the posterior dorsal vertebrae (Figure 1A, B; Figure 5H-H’, P-P’; Figure 6C-C’, K-K’). The neural spines of the pachyostotic sensu stricto vertebrae are swollen, its cross section being rectangular in shape (see vertebra D17 in Figure S2F in the Appendix 1). This cross-section gradually shifts to sub-ellipsoidal as the neural spines become less pachyostotic sensu stricto (Figure 5I, O). The apical margin of the neural spines ranges from slightly convex to flat in lateral view, being always horizontal relative to the centrum ventral rim. The transverse processes are at least twice as high as wide (see Appendix 2, Table S1), not laterally prominent, and showing a morphology from ovoid (in the anterior to middle dorsal vertebrae; Appendix 1, Figure S2B-E) to oblong (in the most posterior dorsal vertebrae) (Figure 5H-H’, P-P’). They completely reach the articular surface of the centrum (Figure 5A-A’, G-G’, M-M’; Appendix 1, Figure S3D). The neural canal is sub-rectangular and high relative to the neural arch total height. The detailed characterization of the zygosphene of the dorsal vertebrae is hampered by both the tight articulation of the dorsal vertebrae in the block and the poor preservation in the disarticulated neural arches. Nevertheless, the dorsal neural arches da#22 and da#31 in Figure 1B-B’ (corresponding to those in Figure 1F, H, I, respectively) preserve a complete zygosphene. This structure dorsally merges with a thin vertical and anteriorly well-projected lamina (i.e., the prespinal lamina) (Figure 1F). Ventrally, the zygosphene is widened in its base, cranially tapering to a pointed tip, and bears a slight ventral indentation, but cannot be considered as bifurcated (Figure 1I). The zygantrum of the dorsal vertebrae is formed by two fossae separated by a vertical septum, which is dorsally continued by a thin lamina (i.e., the postspinal lamina), which is not as prominently projected as the prespinal lamina (Figure 1F; Figure 5, Figure 6, Figure 7). This lamina is not present in the first dorsals, in which the vertical septum displays a ‘rudder’-like morphology (see vertebra D2 in Figure 4C-C’). The zygantrum elongates posteriorly in a wedge-like fashion (named here as the zygantral wedge), notably exceeding beyond the postzygapophyses (Figure 1H; Figure 5E-E’, P-P’, Q-Q’). The zygantral wedge articulates with the zygosphene of the following vertebra. The neural keel is strongly developed in the dorsal neural arches, becoming gradually thinner towards the posterior dorsal series (Figure 1H; Figure 5F, L, R; Figure 6D-D’, H-H’, L-L’; Appendix 1, Figure S3D, E). The prezygapophyseal distance is likely greater than the centrum width based on the disarticulated elements of a similar anatomical position. This ratio cannot be provided for any of the articulated dorsal vertebra, as they are all preserved in lateral view. The prezygapophyseal distance is slightly greater than that between the postzygapophyses (see Appendix 2, Table S1). The pre-infraprezygapohyseal complex is individualized from the main body of the neural arch by a conspicuous constriction. The prezygapophyseal articular facets are oval. They are dorso-medially inclined, displaying and angle of about 45º relative to the horizontal plane in the disarticulated posterior dorsal vertebrae (this angle is unknown in the articulated anterior and middle dorsals). The prezygapophyseal articular facets are larger than those of the infraprezygapophyses. The postzygapophyses of the first dorsals are almost horizontal (see vertebra D2 in Figure 4C-C’). However, the postzygapophyses of the posterior dorsal vertebrae are latero-ventrally inclined and not laterally expanded, and the infrapostzygapophyses are dorsally directed, being more or less inclined (this configuration is unknown in the middle dorsal vertebrae due to their tight articulation) (Figure 5C-C’, J-J’, N-N’; Figure 6B-B’, J-J; Figure 7F-F’).
 Sacral vertebrae. Three or four sacral vertebrae are recognized (see Discussion). Three unequivocal centra of MUPA-ATZ0101 are sacral elements. One of them is preserved in articulation with its neural arch (S3 in Figure 2A-A’). The other two are disarticulated but they were found close to their probably corresponding neural arches (see sc1 and sc2 in Figure 2A-A’). The centra sc1 and sc2 in Figure 2A-A’ (corresponding to those in Figure 9F-M) are highly deformed. The centrum sc1 (Figure 9F-I) is as high as wide, but notably short, as those of the most posterior dorsal region. By contrast, the centra sc2 (Figure 9J-M) and that of the sacral vertebra S3 in Figure 2A-A’ (corresponding to that in Figure 10A-E’) are longer than sc1 (Figure 9K-M, Figure 10C-D’). The centrum of the sacral vertebra S3 lacks a ventral rim. Therefore, the height/length ratio cannot be provided for this element. The overall morphology of the sacral centra is similar to that of most posterior dorsals, being markedly amphicoelous and ventrally constricted, and with an hourglass-shaped neural canal, which is shallower than that of the middle dorsal centra. The sacral centra contribute to the rib articulation, so that the neurocentral suture reaches the lower part of the rib articulation (see Figure 10C-D’).
Sacral vertebrae. Three or four sacral vertebrae are recognized (see Discussion). Three unequivocal centra of MUPA-ATZ0101 are sacral elements. One of them is preserved in articulation with its neural arch (S3 in Figure 2A-A’). The other two are disarticulated but they were found close to their probably corresponding neural arches (see sc1 and sc2 in Figure 2A-A’). The centra sc1 and sc2 in Figure 2A-A’ (corresponding to those in Figure 9F-M) are highly deformed. The centrum sc1 (Figure 9F-I) is as high as wide, but notably short, as those of the most posterior dorsal region. By contrast, the centra sc2 (Figure 9J-M) and that of the sacral vertebra S3 in Figure 2A-A’ (corresponding to that in Figure 10A-E’) are longer than sc1 (Figure 9K-M, Figure 10C-D’). The centrum of the sacral vertebra S3 lacks a ventral rim. Therefore, the height/length ratio cannot be provided for this element. The overall morphology of the sacral centra is similar to that of most posterior dorsals, being markedly amphicoelous and ventrally constricted, and with an hourglass-shaped neural canal, which is shallower than that of the middle dorsal centra. The sacral centra contribute to the rib articulation, so that the neurocentral suture reaches the lower part of the rib articulation (see Figure 10C-D’).
 The neural spines of the sacral vertebrae are notably thin (see Appendix 2, Table S1) (Figure 9A-C’; Figure 10A-B’, E-E’). They show a sub-rounded lateral profile (Figure 9E-E’; Figure 10C-C’). A thin neural keel is present at the ventral region of this element (Figure 9D-D’). The transverse processes barely project laterally, and they are slightly concave and high. The height of the neural spines is also similar to the width of the neural arches (see Appendix 2, Table S1). The prezygapophyseal articular facets are inclined about 50º relative to the horizontal plane. The prezygapophyseal and postzygapophyseal distances are reduced in comparison to those of the dorsal vertebrae. The prezygapophyseal distance is now subequal to the centrum width and roughly greater than that between the postzygapophyses (see Appendix 2, Table S1). The sacral vertebra S3 completely preserves the zygosphene. It is ventrally widened and does not bear a ventral indentation (see Figure 10A-A’). The zygosphene acquires an “L” shape in lateral view, forming an angle of 90º relative to the sagittal plane (Figure 10C-C’).
The neural spines of the sacral vertebrae are notably thin (see Appendix 2, Table S1) (Figure 9A-C’; Figure 10A-B’, E-E’). They show a sub-rounded lateral profile (Figure 9E-E’; Figure 10C-C’). A thin neural keel is present at the ventral region of this element (Figure 9D-D’). The transverse processes barely project laterally, and they are slightly concave and high. The height of the neural spines is also similar to the width of the neural arches (see Appendix 2, Table S1). The prezygapophyseal articular facets are inclined about 50º relative to the horizontal plane. The prezygapophyseal and postzygapophyseal distances are reduced in comparison to those of the dorsal vertebrae. The prezygapophyseal distance is now subequal to the centrum width and roughly greater than that between the postzygapophyses (see Appendix 2, Table S1). The sacral vertebra S3 completely preserves the zygosphene. It is ventrally widened and does not bear a ventral indentation (see Figure 10A-A’). The zygosphene acquires an “L” shape in lateral view, forming an angle of 90º relative to the sagittal plane (Figure 10C-C’).
Caudal vertebrae. The centrum cac#1 in Figure S1 in the Appendix 1 (corresponding to that of Figure 10F-I) is recognized as corresponding to a vertebra of the proximal caudal region based on the small size, the smaller height/length ratio relative to the dorsals (1.18), the markedly constricted shape in ventral view, and the absence of facets for the chevrons. The neural canal is similar to that of the middle dorsals, with parallel borders, but shallower (Figure 10H). The neural arch ca1 in Figure 2A-A’ (corresponding to that in Figure 10A-E’) is identified here as that corresponding to the first caudal vertebra (see Discussion). It has a remarkably thin neural spine (see Appendix 2, Table S1) (Figure 10B-B’), which is sub-rectangular in lateral view (Figure 10D-D’), its height being slightly bigger than the neural arch width (see Appendix 2, Table S1). Both the prezygapophyseal and the postzygapophyseal distances are reduced compared to those of the dorsal vertebrae. The prezygapophyseal distance is equal to that between the postzygapophyses (see Appendix 2, Table S1). The transverse processes are barely prominent laterally, and they are slightly more than twice higher than long (see Appendix 2, Table S1). The inclination angle of the prezygapophyseal facets cannot be provided, since the tight articulation with the preceding vertebra does not allow observing that region.
 Ribs and gastralia. Some cervical ribs, which only preserve their most proximal part, are still articulated with their respective centra (Figure 3F-H’; Figure 4D-D’). The anterior cervical ribs are double-headed, enclosing a circular channel together with the diapophysis and parapophysis. The presence of a free anterior process cannot be ascertained due to the poor preservation of these elements. The rib #1 in Figure S1B in the Appendix 1 (corresponding to that in Figure 11A, B), removed from the block between the vertebral elements C#10 and cc#10, probably represents a posterior cervical rib, based on its triangular shape but seemingly confluent articular facets (Rieppel 1994).
Ribs and gastralia. Some cervical ribs, which only preserve their most proximal part, are still articulated with their respective centra (Figure 3F-H’; Figure 4D-D’). The anterior cervical ribs are double-headed, enclosing a circular channel together with the diapophysis and parapophysis. The presence of a free anterior process cannot be ascertained due to the poor preservation of these elements. The rib #1 in Figure S1B in the Appendix 1 (corresponding to that in Figure 11A, B), removed from the block between the vertebral elements C#10 and cc#10, probably represents a posterior cervical rib, based on its triangular shape but seemingly confluent articular facets (Rieppel 1994).
The dorsal ribs of the right side are mostly disarticulated, whereas those of the left side preserve their original position. They are single-headed, those of the middle region being highly pachyostotic sensu stricto throughout the entire element, from the articular region to the distal one (Figure 1A-B’, Figure 11E; Appendix 1, Figure S3F). They are all conspicuously ornamented with numerous longitudinal striae along the shaft, and numerous tiny pits in their distal region (Figure 1E; Figure 11E, F). The anterior dorsal ribs are notably more curved than those of the middle and posterior dorsal regions (Figure 11D), those of the first dorsals being slim and barely curved (Figure 11C). Both the anterior and the posterior dorsal ribs are shorter than those of the middle dorsal region (Figure 1A-B’; Appendix 2, Table S2). The ribs display a funnel-like (sensu de Miguel Chaves et al., 2020a), flat to slightly concave articular region, being individualized from the rib shaft by a slight constriction (Figure 11C-G; Appendix 1, Figure S3F, G). The rib shaft gradually widens and flattens from its middle region onwards (i.e., elliptical in section) developing a strong distal expansion medio-laterally, and a heavily thickened distal region due to the pachyostosis sensu stricto (Figure 1A-B’, E; Figure 11E). The distal margin of anterior and middle dorsal ribs is unevenly convex, so that its middle region outstands the lateral sides (Figure 11E). The posterior dorsal ribs are not pachyostotic sensu stricto nor distally expanded (Figure 1A-B’; Figure 11F). Four ribs are tentatively identified as the posteriormost dorsal ribs (Figure 11G-J).
Three ribs are unequivocally recognized as sacrals (Figure 2A, B; Figure 11K-M). These elements have a distinct morphology: the rib sr#1 in Figure 2A-A’ (corresponding to that in Figure 11K) has a markedly expanded proximal region, whereas the distal one is slightly expanded, and the shaft is straight and slender; the ribs sr#2 and sr#3 in Figure 2B-B’ (corresponding to those in Figure 11L, M, respectively) have both the proximal and distal regions distinctly expanded, with curved shafts, acquiring a characteristic hourglass shape (Figure 11L, M). The rib #10 in Figure 2A-A’ (corresponding to that in Figure 11N) could potentially represent a sacral one based on its expanded proximal and distal regions, but it is heavily crushed to confirm this anatomical attribution. The rib #11 in Figure 2A-A’ (corresponding to that in Figure 11O) is unequivocally recognized as a caudal element based on its slender and curved morphology, and its cylindrical shaft that tapers into a non-expanded and pointed distal end.
MUPA-ATZ0101 preserves numerous gastral remains (Figure 1B-B’). However, they are very fragmentary and/or disarticulated to provide either the detailed description of the gastral elements or the number of gastral units per element. As in the ribs, the gastralia are highly ornamented with numerous and thin striae (Figure 1C, D).
Girdle Bones
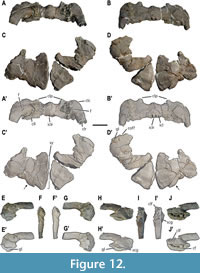 Pectoral girdle. The relatively complete dermal pectoral girdle (i.e., clavicles plus interclavicle) of MUPA-ATZ0101 is preserved (Figure 12A-B’). Both clavicles are relatively complete, only lacking the distal part of the right and left clavicular rami, and the distal part of the clavicular laminae. The dermal pectoral girdle is a wide element (see Appendix 2, Table S3 for measurements) with strongly angulated clavicles (i.e., forming an angle of almost 90o) that articulate with each other immediately anterior to the interclavicle (Figure 12A-B’). The main body of each clavicle is continued by the clavicular rami, which are postero-dorsally projected, and by a postero-medially projected lamina (here termed the clavicular lamina). The clavicular corner (sensu Storrs, 1991; i.e., the region where the clavicle bends) acquires an expanded aspect. Distinct anteriorly projected processes develop at about the mid-half of each clavicle (Figure 12A-B’). The latter are broken in their distal tip; nonetheless, they are interpreted as probably short relative to the clavicle total length based on the preserved borders. A large foramen is present in the dorsal surface of each clavicle, that of the right one being partially eroded, but being symmetrical to the other (Figure 12A-A’). The interclavicle is a small (in relation to the total width of the dermal pectoral girdle; see Appendix 2, Table S3 for measurements), and semi-lunate element that articulates between both clavicles, in a posterior position. Posteriorly, the interclavicle bears a slim medial ramus that is distally broken, its total length being unknown.
Pectoral girdle. The relatively complete dermal pectoral girdle (i.e., clavicles plus interclavicle) of MUPA-ATZ0101 is preserved (Figure 12A-B’). Both clavicles are relatively complete, only lacking the distal part of the right and left clavicular rami, and the distal part of the clavicular laminae. The dermal pectoral girdle is a wide element (see Appendix 2, Table S3 for measurements) with strongly angulated clavicles (i.e., forming an angle of almost 90o) that articulate with each other immediately anterior to the interclavicle (Figure 12A-B’). The main body of each clavicle is continued by the clavicular rami, which are postero-dorsally projected, and by a postero-medially projected lamina (here termed the clavicular lamina). The clavicular corner (sensu Storrs, 1991; i.e., the region where the clavicle bends) acquires an expanded aspect. Distinct anteriorly projected processes develop at about the mid-half of each clavicle (Figure 12A-B’). The latter are broken in their distal tip; nonetheless, they are interpreted as probably short relative to the clavicle total length based on the preserved borders. A large foramen is present in the dorsal surface of each clavicle, that of the right one being partially eroded, but being symmetrical to the other (Figure 12A-A’). The interclavicle is a small (in relation to the total width of the dermal pectoral girdle; see Appendix 2, Table S3 for measurements), and semi-lunate element that articulates between both clavicles, in a posterior position. Posteriorly, the interclavicle bears a slim medial ramus that is distally broken, its total length being unknown.
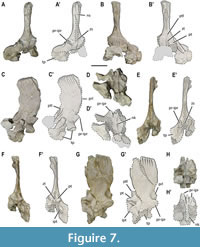
Both coracoids of MUPA-ATZ0101 are incomplete (Figure 1B-B’, Figure 12C-D’) (see Appendix 2, Table S4 for measurements) and show a relatively remarkable taphonomic distortion, especially in their middle and proximal regions. Their anterior and posterior margins are concave, displaying a waisted configuration (sensu Rieppel, 2000). By contrast, the lateral and medial margins are convex, the latter being notably larger. A poorly developed concavity exclusively preserved in the lateral margin of the right coracoid could potentially represent a reduced coracoid foramen (see Figure 12D-D’). The dorsal surface of the left coracoid is concave, but that of the right one is flat, not being possible to recognize which of them correspond to the original morphology. The ventral is convex in both bones. The posterior margin in these elements is thick, the thickness gradually decreases towards the anterior margin, where it becomes extremely thin, especially near the medial margin. The medial region of the left coracoid displays an angled posterior margin of about 150o (Figure 12C-D’), this part being barely preserved in the other. The coracoid symphysis is large, occupying most of the medial region.
Both scapulae of MUPA-ATZ0101 are relatively complete (Figure 12E-J’) (see Appendix 2, Table S5 for measurements), the left scapula undergoing relatively extensive latero-medial compression in its glenoidal portion (Figure 12F-F’), whereas the right one seems to be less distorted in this area (Figure 12I-I’). The scapulae bear a glenoidal portion well differentiated from the scapular blade by a constriction. They bear a large glenoid facet for the humerus articulation, which is larger than that for the coracoid. Above the glenoid of the right clavicle, a well-defined groove is developed (Figure 12H-H’), this character not being observable in the left scapula due to its distortion. The medial surfaces of the scapulae are slightly concave. No coracoid foramen is visible in either of the scapulae of MUPA-ATZ0101 (Figure 12G-G’, J-J’). The right scapula preserves a well-defined clavicular facet, developed in its anteromedial region, which is medially recurved to receive the posterior ramus of the corresponding clavicle (Figure 12I-I’). The scapular blades are robust and postero-dorsally projected, and they are slightly curved dorso-ventrally. They display essentially parallel lateral margins along all their complete length (Figure 12F-F’, I-I’), distally tapering to a blunt end with an oval section (see Figure 1A; Appendix 1, Figure S1A).
 Pelvic girdle. Remains of both ilia are preserved for MUPA-ATZ0101 (Figure 2A-A'; Figure 13A-G’), the right one being complete (see Appendix 2, Table S6 for measurements), whereas the left one only preserves the partial proximal portion (Figure 13F-G’). The right ilium suffers from a severe crush deformity in its anterior part because of taphonomic distortion in cranio-caudal direction. As a result, both that iliac blade and the ventral portion of this bone are bent (Figure 13C-E’). This ilium shows a well-developed iliac blade with a discrete spina preacetabuli, and a postacetabular process that, despite being bent due to deformation, exceeds the posterior margin of the ventral portion (Figure 13A-B’). The iliac blade is separated from the ventral portion of the bone by a constriction (Figure 13A-B’). The ventral portion participates in the formation of the acetabulum, where a sinuous concavity appears in the ventral margin of the lateral side, anteriorly forming the supracetabular buttress. A large facet conforms the acetabulum, which is located low on the ventral portion, right below the supracetabular buttress (Figure 13A-A’). Two more or less oval and similarly sized articular facets are developed ventrally to the acetabulum: an anterior facet for the pubis, and a posterior one for the ischium (Figure 13E-E’). Both ilia show strongly ornamented medial sides and iliac blades with conspicuous rugosities and striae (Figure 13C-C’, D-D’, F-F’), whereas the lateral sides of these bones are smooth (Figure 13A-A’, G-G’).
Pelvic girdle. Remains of both ilia are preserved for MUPA-ATZ0101 (Figure 2A-A'; Figure 13A-G’), the right one being complete (see Appendix 2, Table S6 for measurements), whereas the left one only preserves the partial proximal portion (Figure 13F-G’). The right ilium suffers from a severe crush deformity in its anterior part because of taphonomic distortion in cranio-caudal direction. As a result, both that iliac blade and the ventral portion of this bone are bent (Figure 13C-E’). This ilium shows a well-developed iliac blade with a discrete spina preacetabuli, and a postacetabular process that, despite being bent due to deformation, exceeds the posterior margin of the ventral portion (Figure 13A-B’). The iliac blade is separated from the ventral portion of the bone by a constriction (Figure 13A-B’). The ventral portion participates in the formation of the acetabulum, where a sinuous concavity appears in the ventral margin of the lateral side, anteriorly forming the supracetabular buttress. A large facet conforms the acetabulum, which is located low on the ventral portion, right below the supracetabular buttress (Figure 13A-A’). Two more or less oval and similarly sized articular facets are developed ventrally to the acetabulum: an anterior facet for the pubis, and a posterior one for the ischium (Figure 13E-E’). Both ilia show strongly ornamented medial sides and iliac blades with conspicuous rugosities and striae (Figure 13C-C’, D-D’, F-F’), whereas the lateral sides of these bones are smooth (Figure 13A-A’, G-G’).
MUPA-ATZ0101 only preserves the left pubis (Figure 2, Figure 13H-J). This element is almost complete (see Appendix 2, Table S4 for measurements). It is relatively well preserved, although being very compressed medio-laterally due to taphonomic distortion. The pubis bears concave anterior and posterior margins and expanded convex lateral and medial ones (Figure 13H-I’). The lateral margin participates in the formation of the acetabulum, but most of this region is eroded (Figure 13H-H’). The obturator foramen is open and oval, being located ventro-proximally in the lateral margin (Figure 13H-H’). The medial margin is distorted; however, an evenly convex and non-notched contour is interpreted for it based on the preserved region (Figure 13H-I’). The pubic symphysis occupies half of the medial margin of this bone. The medial surface of the lateral margin (i.e., the anatomically dorsal side) is distinctly ornamented, whereas the lateral surface (i.e., the anatomically ventral side) is smoother (Figure 13J).
The left ischium is the only one preserved for MUPA-ATZ0101 (Figure 2, Figure 13K-M). This element is almost complete (see Appendix 2, Table S4 for measurements), only lacking part of its articular region (Figure 13K-L’). As in the pubis, it is medio-laterally compressed due to taphonomic distortion. The ischium bears strongly concave anterior and posterior margins, the anterior concavity seemingly being more pronounced, which would conform an asymmetrical (i.e., axe-like rather than fan-like) shape for this element. The lateral and especially the medial margins are expanded (Figure 13K-L’). The lateral margin of the ischium is incomplete, but the preserved area allows to recognize that it participates in the formation of the acetabulum (Figure 13L-L’). The ischium bears a short and wide shaft. The symphysis occupies less than half of its medial margin (Figure 13L-L’). As in the pubis, the medial (i.e., the anatomically dorsal side) surface of the proximal region is highly ornamented, whereas its lateral (i.e., the anatomically ventral side) surface is smoother (Figure 13M). The posterior margin of the pubis and the anterior margin of the ischium conform a large thyroid fenestra.
Limb Bones
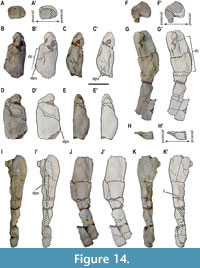 Forelimb elements. Both partial humeri of MUPA-ATZ0101 are preserved (Figure 14; Appendix 1, Figure S1). The left humerus only preserves the proximal half (Figure 14A-E’), but with a more complete proximal region than that of the right one. The right humerus is almost complete, only lacking the most proximal area of the articular region, and a small portion of the distal epiphysis (Figure 14F-K’). The estimated total length for a complete humerus, considering the preserved regions in both humeri of MUPA-ATZ0101, is about 290 mm (see Appendix 2, Table S7 for measurements). The most complete humerus shows that was strongly arched with a relatively well-developed deltopectoral crest (Figure 14G-G’, J-J’), its postaxial margin being concave (Figure 14G-G’, J-J’). The proximal humeral regions are massive, displaying a subrounded contour (Figure 14A-A’, F-F’). The proximal region of the left humerus is interpreted as preserving the original volume, being larger than the medially crushed proximal area of the right one. The right humerus shows that the distal region was more expanded than the proximal one. The humerus is notably constricted in its mid-diaphyseal region (Figure 14G-G’, J-J’). The distal region of this element is preaxially voluminous, whereas it flattens towards the postaxial margin (Figure 14H-H’). A distinct scar, termed here as the deltopectoral scar, is developed in the deltopectoral crest of both humeri, that of the right humerus being completely preserved. The deltopectoral scar in the latter element extends over the preaxial flexion, its length being approximately one third of the humerus total length (Figure 14B-D’, G-I’). A small foramen is observed in the postaxial margin of the right humerus (Figure 14K-K’), its presence cannot be confirmed in the left humerus due to preservation reasons. The humerus of this taxon lacks an entepicondylar foramen, an ectepicondylar groove, and a separation for the radial and ulnar condyles (Figure 14G-H’, J-J’). No zeugopodial or autopodial forelimb elements are preserved in MUPA-ATZ0101.
Forelimb elements. Both partial humeri of MUPA-ATZ0101 are preserved (Figure 14; Appendix 1, Figure S1). The left humerus only preserves the proximal half (Figure 14A-E’), but with a more complete proximal region than that of the right one. The right humerus is almost complete, only lacking the most proximal area of the articular region, and a small portion of the distal epiphysis (Figure 14F-K’). The estimated total length for a complete humerus, considering the preserved regions in both humeri of MUPA-ATZ0101, is about 290 mm (see Appendix 2, Table S7 for measurements). The most complete humerus shows that was strongly arched with a relatively well-developed deltopectoral crest (Figure 14G-G’, J-J’), its postaxial margin being concave (Figure 14G-G’, J-J’). The proximal humeral regions are massive, displaying a subrounded contour (Figure 14A-A’, F-F’). The proximal region of the left humerus is interpreted as preserving the original volume, being larger than the medially crushed proximal area of the right one. The right humerus shows that the distal region was more expanded than the proximal one. The humerus is notably constricted in its mid-diaphyseal region (Figure 14G-G’, J-J’). The distal region of this element is preaxially voluminous, whereas it flattens towards the postaxial margin (Figure 14H-H’). A distinct scar, termed here as the deltopectoral scar, is developed in the deltopectoral crest of both humeri, that of the right humerus being completely preserved. The deltopectoral scar in the latter element extends over the preaxial flexion, its length being approximately one third of the humerus total length (Figure 14B-D’, G-I’). A small foramen is observed in the postaxial margin of the right humerus (Figure 14K-K’), its presence cannot be confirmed in the left humerus due to preservation reasons. The humerus of this taxon lacks an entepicondylar foramen, an ectepicondylar groove, and a separation for the radial and ulnar condyles (Figure 14G-H’, J-J’). No zeugopodial or autopodial forelimb elements are preserved in MUPA-ATZ0101.
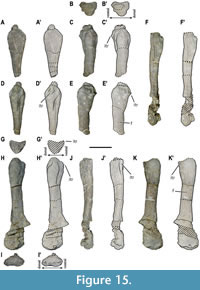 Hindlimb elements. MUPA-ATZ0101 preserves both partial femora (Figure 15; Appendix 1, Figure S1). The left femur only preserves the almost complete proximal half (Figure 15A-E’). The right one is almost complete, only lacking the proximalmost part of the articular region (Figure 15F-K’). The estimated total length for a complete femur of the taxon studied here, considering the preserved regions in both femora of MUPA-ATZ0101, is estimated to be about 235 mm (see Appendix 2, Table S7 for measurements). The femur is less massive and robust (based on the proximal diameter) than the humerus. It shows a small foramen in the proximal half of the shaft in postaxial view, being symmetrical in both femora (Figure 15E-E’, K-K’). The right femur shows that this element has a slightly sigmoidal shape (Figure 15H-K’). The proximal femoral region shows a triangular morphology in cross-section due to the weakly developed inner trochanter (Figure 15B-B’, G-G’). The femur of this taxon shows a weakly developed intertrochanteric fossa (Figure 15B-E’, G-K’). Thus, the inner trochanter is not set off from the femoral shaft. The femur is 1.23 times shorter than the humerus, as well as less expanded both proximally and distally. The expansion of the distal region is only slightly wider than that of the proximal one. A weak intercondylar fossa is developed in the postero-ventral part of the femoral distal region (Figure 15 I-I’, K-K’), separating the articular facets for the tibia and the fibula. No zeugopodial or autopodial hindlimb elements are preserved in MUPA-ATZ0101.
Hindlimb elements. MUPA-ATZ0101 preserves both partial femora (Figure 15; Appendix 1, Figure S1). The left femur only preserves the almost complete proximal half (Figure 15A-E’). The right one is almost complete, only lacking the proximalmost part of the articular region (Figure 15F-K’). The estimated total length for a complete femur of the taxon studied here, considering the preserved regions in both femora of MUPA-ATZ0101, is estimated to be about 235 mm (see Appendix 2, Table S7 for measurements). The femur is less massive and robust (based on the proximal diameter) than the humerus. It shows a small foramen in the proximal half of the shaft in postaxial view, being symmetrical in both femora (Figure 15E-E’, K-K’). The right femur shows that this element has a slightly sigmoidal shape (Figure 15H-K’). The proximal femoral region shows a triangular morphology in cross-section due to the weakly developed inner trochanter (Figure 15B-B’, G-G’). The femur of this taxon shows a weakly developed intertrochanteric fossa (Figure 15B-E’, G-K’). Thus, the inner trochanter is not set off from the femoral shaft. The femur is 1.23 times shorter than the humerus, as well as less expanded both proximally and distally. The expansion of the distal region is only slightly wider than that of the proximal one. A weak intercondylar fossa is developed in the postero-ventral part of the femoral distal region (Figure 15 I-I’, K-K’), separating the articular facets for the tibia and the fibula. No zeugopodial or autopodial hindlimb elements are preserved in MUPA-ATZ0101.
DISCUSSION
Comparison of the Vertebral Elements and Implications
General vertebral characters. The holotype of Paludidraco multidentatus (MUPA-ATZ0101) shows distinctly constricted centra in ventral view (except those pachyostotic sensu stricto) (Figure 8X, AB, AF, AJ). This condition was considered a synapomorphy of Simosauridae within Nothosauroidea (de Miguel Chaves et al., 2018a; 2020a). However, several centra attributed to Nothosaurus sp. (e.g., SMNS 80266, SMNS 83945) and Nothosaurus giganteus Münster, 1834 (e.g., SMNS 84057) are distinctly constricted in ventral view, differing from those typically displaying parallel lateral margins and not constricted in ventral view of most nothosaurians (see Appendix 1, Figure S4A-B vs L-Q, Y-Z). Therefore, this character state is not synapomorphic of Simosauridae within Nothosauroidea, contrary to the previous considerations (Rieppel, 1999; de Miguel Chaves et al., 2018a, 2020a). The centra of Simosauridae usually display a more pronounced constriction (in lateral view) than those constricted of some nothosaurs (compare the elements in Figure S4D, H with those in Figure S4N, Q in the Appendix 1; and see Dalla Vecchia, 2008, figure 3; de Miguel Chaves et al., 2015, figure 2; de Miguel Chaves et al., 2020a, figure 3). However, this character can be subject to positional variability (see Figure 8 in this study, and de Miguel Chaves et al., 2020a, figure 1), some of the centra being as slightly constricted (Figure 8B, F) as those of some nothosaurs (see Appendix 1, Figure S4N, Q). For this reason, this character cannot be considered diagnostic of Simosauridae and, thus, it is considered here in the exclusive character combination of this clade. The centra of P. multidentatus exclusively share the socketed configuration with those of Simosaurus gaillardoti (e.g., Figure S5H, M, R; S6O, AA in the Appendix 1) and the vertebrae of indeterminate simosaurids within Sauropterygia (see Rieppel, 1994; Dalla Vecchia, 2008; de Miguel Chaves et al., 2015). Therefore, this condition is considered here as a synapormorphy of the clade. Small and unpaired foramina are sporadically observed here in the dorsal centra of MUPA-ATZ0101 (Figure 1G) and S. gaillardoti (ACH, pers. obs.), a condition also recognized in some Nothosaurus vertebrae (e.g., Sanz, 1983; Rieppel et al., 1999; Liu et al., 2014; Klein et al., 2022). They differ from the large, paired and/or symmetrical subcentral foramina of Pistosauroidea (Sander et al., 1997; Rieppel et al., 1999; Liu et al., 2014), considered synapomorphic of Pistosauria (O’Keefe, 2001; Dalla Vecchia, 2006; Sato et al., 2010; 2014a) or Plesiosauria within Sauropterygia (Brown, 1981; Storrs, 1991; Sander et al., 1997; Druckenmiller and Russell, 2008; Brum et al., 2022).
MUPA-ATZ0101 shows that the neural arches of P. multidentatus display infrapre- and infrapostzygapophyses in all preserved vertebrae (i.e., a character confirmed from the posterior cervicals to the proximal caudals). The presence of infrapre- and infrapostzygapophyses was recognized as a synapomorphy of the clade Simosauridae within Nothosauroidea (de Miguel Chaves et al., 2018a; 2020a). Similar zygapophyseal articulations were exclusively identified within Sauropterygia in the pistosauroid Bobosaurus forojuliensis Dalla Vecchia, 2006, from the lower Carnian of Italy, but with a reversed polarity (i.e., the socket is formed by the pre-infraprezygapophyses, and the wedge is formed by the post-infrapostzygapophyses) and lacking the ‘X’ configuration of the socket (see Dalla-Vecchia, 2006, 2017). Therefore, the presence of an ‘X-shaped’ wedge-and-socket system is here identified as exclusive of Simosauridae within Sauropterygia. On the one hand, the neural arches of P. multidentatus exclusively share with those of S. gaillardoti and those of indeterminate simosaurids within Sauropterygia the presence of ovoid to oblong and not laterally prominent transverse processes (e.g., Vickers-Rich et al., 1999; Dalla Vecchia, 2008, 2017; de Miguel Chaves et al., 2015; Segesdi and Ősi, 2021). This condition is considered here a synapomorphy of the clade. On the other hand, the not laterally prominent transverse processes of Nothosauria (i.e., the sister group of Simosauridae) are slenderer, less dorsoventrally developed, and ‘8-shaped’ (sensu Klein et al., 2022) (Storrs, 1991; Rieppel, 1998a; Diedrich, 2013a; and see Sanz, 1984; Rieppel and Wild, 1996; Diedrich, 2014). Paludidraco multidentatus shares with S. gaillardoti (and the so far documented vertebrae attributable to indeterminate simosaurids) the presence of conspicuously ornamented neural spines of the dorsal vertebrae, with lateral longitudinal grooves in the upper dorsal third, being relatively high and latero-medially constrained (see Skuphos, 1893; Rieppel, 1994, 1996; Dalla Vecchia, 2008; de Miguel Chaves et al., 2015, 2020a; Segesdi and Ösi, 2021). Neural spines with longitudinal grooves or distinct apical ornamentation are also present in some basal pistosauroids (e.g., Augustasaurus hagdorni Sander et al., 1997 or Pistosaurus longaevus Meyer, 1839; see Diedrich, 2013a; Sander et al., 2013), some pachypleurosaurs (e.g., Serpianosaurus mirigiolensis Rieppel, 1989; Serpianosaurus germanicus Diedrich, 2013b), and some nothosaurs (e.g., Nothosaurus tchernovi Haas, 1980; Nothosaurus jagisteus Rieppel, 2001). Nevertheless, the spines of the latter taxa differ from those of Simosauridae in being considerably lower (i.e., A. hagdorni, S. mirigiolensis, S. germanicus), showing a sub-rounded profile (i.e., A. hagdorni), or displaying a rounded section in the dorsal vertebrae (i.e., P. longaevus). The moderately high dorsal neural spines in Simosauridae are proportionally lower than those of most plesiosaurs (see Druckenmiller and Russell, 2008); and the nothosaurs Nothosaurus mirabilis Münster, 1834, from the Ladinian of Germany; and Nothosaurus haasi Rieppel et al., 1997 and N. tchernovi, both from the upper Anisian or lower Ladinian of Israel (Rieppel, 2000; Dalla-Vecchia, 2006; Klein et al., 2022). Some other Triassic eosauropterygians also present moderately high neural spines (e.g., Nothosaurus juvenilis Edinger, 1921, from the Anisian of Germany [see Diedrich, 2013c]; Wumengosaurus delicatomandibularis Jiang et al., 2008, from the Anisian of China [see Wu et al., 2011]; or P. longaevus, from the Anisian and Ladinian of Germany [see Diedrich, 2013a]). The presence of pre- and postspinal laminae and a zygantral wedge in P. multidentatus, at least from the dorsal to the sacral vertebrae, is shared with S. gaillardoti (Appendix 1, Figures S6A-D, F, H; S7B, C, K, L, N) and with the isolated vertebrae of indeterminate simosaurids (e.g., Skuphos, 1893; Dalla Vecchia, 2008). Prespinal laminae have also been reported in the nothosaurian N. mirabilis (Klein et al., 2022), and the pistosauroid B. forojuliensis (Dalla-Vecchia, 2017), being larger in these taxa; and a zygantral wedge is also present in the dorsal vertebrae of some vertebrae attributable to indeterminate species of Nothosaurus (e.g., SMNS 80266) and in N. giganteus (e.g., SMNS 84061, SMNS 84453) (ACH, pers. obs.), but it is absent in all other sauropterygian taxa analyzed in this study (i.e., Placodontia, Pachypleurosauria, most nothosaurians, and Pistosaurus). The zygosphene of Simosauridae is poorly known, but based on the available material, it seems that it never develops articular facets (Dalla Vecchia, 2008; de Miguel Chaves et al., 2015). By contrast, the zygosphene in Nothosauria displays well-developed and individualized articular facets (Sanz, 1984; Rieppel et al., 1997, 1999; de Miguel Chaves et al., 2016). As in all other basal eosauropterygians, the prezygapophyseal distance of the presacral vertebrae in Simosauridae is always larger than the centrum width (see description section; Rieppel, 1994, 2000; Dalla Vecchia, 2008; de Miguel Chaves et al., 2015, 2020a).
Simosauridae presents a greater number of dorsal vertebrae (32 elements or more) (S. gaillardoti: 32–34 [based on GPIT-PV-31566; see Huene, 1959; Rieppel, 1994; ACH, pers. obs.]; and P. multidentatus: 40–41 [based on MUPA-ATZ0101; see Appendix 1, Figure S1]) than other sauropterygians (see Rieppel, 2000), only comparable to the eosauropterygian W. delicatomandibularis, from the Anisian of China, with a maximum of 30–31 dorsal vertebrae (Jiang et al., 2008). The cervical and caudal regions are poorly known in Simosauridae, these regions only being partially preserved in both the most complete specimens of S. gaillardoti (i.e., GPIT-PV-31566, SMNS 14733) and the holotype of P. multidentatus. In this sense, the holotype of the simosaurid ‘Partanosaurus zitteli’ Skuphos, 1893 (considered as a nomen dubium; see de Miguel Chaves et al., 2020a), GBA 1893/010/0001, also corresponds to a partial skeleton, but it lacks the cervical and caudal regions. Considering the above discussion, the presence of at least 32 dorsal vertebrae is here considered as a synapomorphy of Simosauridae, with S. gaillardoti having fewer than 35 dorsal vertebrae, and P. multidentatus having at least 40 of these elements.
Within Simosauridae, several vertebral differences were previously reported by de Miguel Chaves et al., (2020a) between P. multidentatus (i.e., pachyostotic zygapophyses; amphicoelous centra; higher than long dorsal centra; and high transverse processes of the dorsal vertebrae, without ventral notch, contacting the articular surface of the centra) and S. gaillardoti (i.e., absence of zygapophyseal pachyostosis; platycoelous centra; longer than high or at least as long as high dorsal centra; and transverse processes of the dorsal vertebrae with a ventral notch, and not contacting the articular surface of the centra). The detailed comparison of P. multidentatus with S. gaillardoti allows to improve the previously established vertebral characterization for both taxa. In this sense, in addition to the always higher than long cervical centra, P. multidentatus displays pachyostotic sensu stricto dorsal vertebrae, not only affecting the zygapophyses (i.e., with bloated neural spines, transverse processes, zygapophyses, and centra). Therefore, they differ from the vertebrae of S. gaillardoti (i.e., non-pachyostotic sensu stricto, and always with as high as long centra; see Appendix 1, Figures S5, S6 and Appendix 2, Table 1). Moreover, new vertebral characters, not present in S. gaillardoti, are recognized here for P. multidentatus. Thus, P. multidentatus displays a moderately deep neural canal of the dorsal vertebral centra, unlike that markedly deep condition in the dorsal centra of S. gaillardoti (Appendix 1, Figure S6O, AE). The neural canal of the cervical centra in both taxa is moderately deep compared to that of the dorsals of S. gaillardoti (Appendix 1, Figure S5H, M, R), but this character state is unknown for the sacrals and caudals of the latter taxon. An elongated zygantral wedge, notably exceeding the postzygapophyses, is present at least in the posterior dorsal and sacral vertebrae of P. multidentatus, this character state being unknown for the cervicals, anterior and middle dorsals due to the tight articulation; and for the caudals due to preservation issues. On the contrary, a short zygantral wedge is always present from the dorsal to the sacral vertebrae of S. gaillardoti (see Appendix 1, Figures S6H; S7K, N), being absent in the cervicals. The neural arches of P. multidentatus display a distinct neural keel, unlike those of S. gaillardoti, which lack this character (see Appendix 1, Figure S6C, H). The postzygapophyses of the dorsal vertebrae in P. multidentatus never expand laterally, whereas those of S. gaillardoti are notably expanded laterally (see Appendix 1, Figure S6D, I). The cervical vertebrae of P. multidentatus present infrapre- and infrapostzygapophyes, unlike those of S. gaillardoti (see Rieppel, 1994, Figure 13). The latter character is absent in any other sauropterygian and, thus, considered here as an autapomorphy of P. multidentatus. In addition, the articulation between most preserved vertebrae in MUPA-ATZ0101 hampers the detail characterization of the zygosphene in P. multidentatus, being probably not bifurcated unlike the condition for S. gaillardoti (see Appendix 1, Figure S6E).
Some vertebral features of P. multidentatus are shared with other sauropterygians, but not with S. gaillardoti. Hence, amphicoelous vertebrae are also present in Pachypleurosauria, some Nothosauria and some Pistosauroidea (Rieppel, 2000), but they do not display socketed centra. Additionally, the vertebrae of these taxa differ from those of P. multidentatus in being: smaller, and proportionally longer in pachypleurosaurs and Lariosaurus spp. (Rieppel, 2000; Diedrich, 2013b, c, 2014); longer than high in the basal pistosauroids Corosaurus alcovensis Case 1936, Yunguisaurus liae Cheng et al., 2006 (Storrs, 1991; Rieppel, 1998a; Sato et al., 2010, 2014a) and the specimen attributed to cf. Cymatosaurus by Sander et al., (2013); and typically wider than high in Plesiosauria (Brown, 1981; Vincent and Storrs, 2019; Persons et al., 2022). Pachyostotic sensu stricto vertebrae are commonly found in Pachypleurosauria and some Nothosauria (Rieppel, 2000). A neural keel apparently has never been mentioned in the literature for any sauropterygian. However, some dorsal vertebrae attributed to Nothosaurus sp. (e.g., SMNS 84707, SMNS 80266, or SMNS 90098) and to N. giganteus (e.g., SMNS 80204, SMNS 84061, or SMNS 84453) present this character (ACH, pers. obs.), but it is absent in all other sauropterygian taxa analyzed in this study (i.e., Placodontia, Pachypleurosauria, most nothosaurians, and Pistosaurus).
The sacral elements in Simosauridae. The number of sacral vertebrae is usually challenging to determine in Sauropterygia if they are not fully articulated with their sacral ribs, or if the latter elements are not connected to the ilium (Rieppel, 1994; Dalla Vecchia, 2017). This issue arises because there are usually no major differences with the immediately anterior or posterior vertebrae, and because the most posterior dorsal and the most anterior caudal ribs may converge on the ilium in addition to the true sacral ribs (Rieppel, 1994). This challenge is further complicated by the fact that some eosauropterygians show ‘sacralization’ (sensu Rieppel, 1994) of the last dorsal and first caudal vertebrae, so they are morphologically similar to the true sacral ones (Rieppel, 1994, 2000; Klein et al., 2022). In MUPA-ATZ0101, five neural arches (i.e., da#41, sa1, S2, S3, and ca1; Figure 7E-H’; Figure 9A-E’; Figure 10A-E’) of the most posterior region display a different morphology and proportions compared to the preceding dorsal ones (see “sacral vertebrae” in the description section and Appendix 2, Table S1). The neural spines in sa1, S2 and S3 are sub-rounded, and cranio-caudally and dorso-ventrally shorter than those of da#41 and ca1 (Figure 7E-H’; Figure 10A-E’). These three elements (i.e., sa1, S2, and S3) show a morphology highly similar to those considered sacrals for S. gaillardoti (Rieppel, 1994) (see Appendix 1, Figure S7A-C, K-O). Therefore, they unequivocally represent sacral vertebrae, with a potential fourth additional one (i.e., da#41), which would represent the first sacral. The element ca1 is considered here as the first caudal vertebra based on its more sub-rectangular and slightly cranio-caudally longer neural spine. Considering the above discussion, the sacral number recognized for MUPA-ATZ0101 is three or four. A minimum of three sacral vertebrae were recognized by Rieppel (1994) in S. gaillardoti (based on SMNS 14733), although a fourth element (i.e., the vertebra #113; see Appendix 1, Figure S7L) was likely to represent the fourth sacral vertebra (Rieppel, 1994). The vertebra #113 is anatomically and proportionally identical to the preceding sacral vertebrae of SMNS 14733 (see Appendix 1, Figure S7K-O and Appendix 2, Table S1). The relatively complete and articulated specimen of S. gaillardoti GPIT-PV-31566 displays three unequivocal sacral ribs directly contacting the ilium in the left anatomical side of the body (Appendix 1, Figure S8A-C), as reported by Rieppel (1994) and contrary to the five erroneously reported by Huene (1959). However, an additional rib may represent the fourth sacral rib based on its length and morphology (ACH, pers. obs.). Hence, S. gaillardoti probably has four sacral vertebrae and, consequently, a sacral count of three or four vertebrae is considered here for Simosauridae.
Comparison of the Rib Elements and Implications
The holotype of Paludidraco multidentatus has double-headed cervical ribs as is commonly present in basal eosauropterygians (Rieppel, 1994, 2000). Nevertheless, the presence of an anterior free process in the cervical ribs cannot be confirmed in the holotype of P. multidentatus. The dorsal ribs of P. multidentatus, Simosaurus gaillardoti, ‘Partanosaurus zitteli’, and other indeterminate simosaurids share a middle-to-distal flattened rib shaft, and a medio-laterally expanded distal region (Rieppel, 1994, 1996; Dalla Vecchia, 2008; de Miguel Chaves et al., 2020a; Reolid and Reolid, 2020). By contrast, the dorsal ribs of Nothosauria usually display a rounded shaft, with a distal region that is either non-expanded or slightly expanded (Reolid and Reolid, 2020; Klein et al., 2022), and an ‘8-shaped’ articular region (sensu Klein et al., 2022). Therefore, the middle-to-distal flattened shaft of the ribs in Simosauridae is considered here as synapomorphic within Sauropterygia. The strong distal expansion and the highly pachyostotic sensu stricto condition of the dorsal ribs in P. multidentatus was recognized by de Miguel Chaves et al. (2020a) as a difference relative to the condition in S. gaillardoti and ‘Pa. zitteli ’ (i.e., displaying not pachyostotic sensu stricto, and moderately expanded dorsal ribs). The detailed comparison between S. gaillardoti and P. multidentatus allows us to identify here that the unevenly convex distal margin of the dorsal ribs in the Spanish taxon is not present in S. gaillardoti, where it is straight or slightly convex (see de Miguel Chaves et al., 2020a, figure 4). Several other sauropterygians display pachyostotic sensu stricto dorsal ribs (Rieppel and Lin, 1995; Rieppel, 1998b; Lin and Rieppel, 1998; Jiang et al., 2008, 2014; Liu et al., 2011; Cheng et al., 2016; Lin et al., 2017), but this condition is generally restricted to the proximal half of the bones, lacking a distal expansion. The sacral ribs of MUPA-ATZ0101, as those of S. gaillardoti and ‘Pa. zitteli’, are distinctly expanded distally in dorso-ventral direction (Appendix 1, Figure S4E, I, K), a character previously recognized as a synapomorphy of Simosauridae within Sauropterygia (de Miguel Chaves et al., 2018a, 2020a). However, this dorso-ventral expansion of the sacral ribs is also identified here in some specimens attributed to Nothosaurus sp. (e.g., SMNS 80266, SMNS 84898) or Nothosaurus giganteus (e.g., SMNS 87690) (see Appendix 1, Figure S4R, T, W) and, thus, is no longer considered here as exclusive of Simosauridae. Additionally, the sacral ribs of S. gaillardoti, especially those more robust, display a cranio-caudally directed distal expansion (Appendix 1, Figure S4F), which is exclusive within Nothosauroidea.
Comparison of the Girdle Bone Elements and Implications
Pectoral girdle. The presence of an ‘anterolateral’ process in the clavicle (sensu Rieppel, 1994, and here re-named as anteromedial considering its precise anatomical position) was recognized as a synapomorphy of the clade Simosauridae within Nothosauroidea (de Miguel Chaves et al., 2018a, 2020a). Clavicular processes are also present in the Triassic eosauropterygians Brevicaudosaurus jiyangshanensis Shang et al., 2020, from the Ladinian of China; and Diandongosaurus acutidentatus Shang et al., 2011 (Sato et al., 2014b; Xue-Qing et al., 2015) and Dianmeisaurus gracilis Shang and Li, 2015 (Shang et al., 2017), both from the Anisian of China. However, the processes in these pachypleurosaur taxa are in an anterolateral position. The presence of strongly angulated clavicles, a postero-medially expanded clavicular lamina, and a scapular blade with parallel margins and a blunt end with oval section is shared between Paludidraco multidentatus (based on its holotype, MUPA-ATZ0101) and Simosaurus gaillardoti (see Appendix 1, Figure S9). A different configuration is present in some indeterminate members of Simosauridae not attributable to S. gaillardoti or to P. multidentatus for the clavicles (i.e., curved clavicles in the specimen MFSN 34904 from the Carnian of Italy; see Dalla Vecchia 2008) and for the scapulae (i.e., scapular blade with a widened distal end in the holotype of ‘Partanosaurus zitteli’; see de Miguel Chaves et al., 2020a and references therein). An expanded clavicular lamina is present in all the clavicles attributed to Simosauridae, but it is absent in Nothosauria, (ACH, pers. obs.) (see Appendix 1, Figure S9A-B, G-J; Dalla Vecchia, 2008, figure 6; Rieppel, 1994, figure 25; Rieppel et al., 1999, figure 11) and, thus, it is considered here as synapomorphic of Simosauridae within Nothosauroidea. A similar lamina, but less expanded, is developed in the clavicles of the placodont Placodus gigas Agassiz, 1833 (see Rieppel, 1994, figure 54D, E). Additionally, some differences are recognized here between the pectoral girdle elements of P. multidentatus (based on its holotype) in relation to those of S. gaillardoti: presence of two symmetrical clavicular foramina (unlike the clavicles of S. gaillardoti, which lack these foramina [see Appendix 1, Figure S9A, G, I]); a semi-lunate interclavicle (unlike that T-shaped of S. gaillardoti [see Appendix 1, Figure S9B, I-J]); reduced or absent coracoid foramen (unlike that well-developed of S. gaillardoti [see Appendix 1, Figure S9E-J]); and an angled posteromedial margin of the coracoid (unlike that straight of S. gaillardoti [see Appendix 1, Figure S9C-D, K-N]). Both an angled posteromedial margin of the coracoid and the presence of a clavicular foramina have not been described for any sauropterygian in the literature. However, some small and medially located, rather than dorsally, foramina, have been identified in clavicles attributed to Nothosaurus sp. (ACH, pers. obs., based on the specimens SMNS 84073, SMNS 59822). Therefore, the presence of both two symmetrical foramina located dorsally in the clavicles, and an angled posteromedial margin of the coracoid, are here considered exclusive characters of P. multidentatus.
Pelvic girdle. An iliac blade set off from the acetabular region by a distinct constriction, the presence of a spina preacetabuli and a postacetabular process in the ilium, and waisted ischia and pubes, are characters present in both Paludidraco multidentatus (based on its holotype, MUPA-ATZ0101) and Simosaurus gaillardoti. The general morphology of the pelvic girdle elements in Simosauridae is shared with that of the other basal eosauropterygians (i.e., a waisted pubis and ischium) (Rieppel, 2000). The set off iliac blade of P. multidentatus and S. gaillardoti is also shared with the only other known ilium attributed to Simosauridae (i.e., the specimen MFSN 34914; see Dalla Vecchia, 2008) and other non-nothosauroid sauropterygians (e.g., Placodus gigas or Wumengosaurus delicatomandibularis), but not with those of Nothosauria (Rieppel, 2000). Hence, the presence of a set off iliac blade is considered here a synapomorphy of Simosauridae within Nothosauroidea. A small spina preacetabuli is also present in the isolated ilium SMNS 84493 (ACH, pers. obs.) attributed to Nothosaurus sp., and in the specimen attributed to Nothosaurus cf. juvenilis by Renesto (2010). The seemingly shared asymmetrical morphology (i.e., axe-like) of the ischia of P. multidentatus (based on its holotype, MUPA-ATZ0101) and S. gaillardoti would differ from the typically symmetrical (i.e., fan-like) morphology in the nothosaurians (see Shang, 2006; Klein et al., 2022). However, as it is mentioned above (see “pelvic girdle” in the description section), the state of this character is dubious in the holotype of P. multidentatus and, thus, it is not considered here in the diagnosis of Simosauridae. In addition, some differences are identified here between the pelvic girdle elements of the holotype of P. multidentatus and those of S. gaillardoti. In this sense, P. multidentatus presents a long postacetabular process that extends beyond the posterior margin of the ilium. In contrast, the postacetabular process of S. gaillardoti does not extend beyond the posterior margin of the ilium (see Appendix 1, Figure S7D-E, P-Q). A postacetabular process exceeding the posterior margin of the ilium is shared between P. multidentatus and other sauropterygians (e.g., W. delicatomandibularis [see Wu et al., 2011, figure 4E, F]; P. gigas [see Rieppel, 2000, figure 10A, B]). The Spanish taxon also displays a pubis with an open obturator foramen and a medial margin not notched, whereas S. gaillardoti displays an enclosed obturator foramen and notched pubis (see Appendix 1, Figure S7I, U). Pubes with an open obturator foramen are also present in some species of Nothosaurus and Lariosaurus (e.g., Nothosaurus mirabilis, Lariosaurus hongguoensis Jiang et al., 2006); however, the foramen is usually slit-like (Rieppel, 2000) rather than rounded or oval as in S. gaillardoti and P. multidentatus. Moreover, the pubic and ischial shafts are wide in P. multidentatus, unlike those strongly constricted in S. gaillardoti (see Appendix 1, Figure S7I, J, U, V).
Comparison of the Limb Bone Elements and Implications
Forelimb elements. The humerus of Paludidraco multidentatus (based on its holotype, MUPA-ATZ0101) and that of Simosaurus gaillardoti share a strongly arched preaxial margin. They also display a distinct diaphyseal constriction, resulting in a mid-shaft narrower than the proximal and the distal regions. Consequently, the humerus of Simosauridae differs from that of its sister group Nothosauria, where it is not strongly arched, and the mid-shaft is not distinctly constricted, being subequal ( Nothosaurus spp.) or distinctly wider ( Lariosaurus spp.) than the proximal region (Jiang et al., 2006; Ji et al., 2014; Li and Liu, 2020; Klein et al., 2022). A distinctly constricted humeral mid-shaft is shared by Simosauridae with other non-nothosaurian eosauropterygians, such as Cymatosaurus spp. (see Sander et al., 2013 and references therein; Klein et al., 2015) or Serpianosaurus germanicus (Diedrich, 2013b). In addition, the humerus of Simosauridae displays the following characters: it lacks an entepicondylar foramen, unlike that commonly present in nothosaurians (Rieppel, 1998b, 2000; Bickelmann and Sander, 2008); it is longer than the femur, the opposite condition being usually present in Nothosauria (Rieppel, 1994, 1998b; Rieppel and Wild, 1996; Shang, 2006; Ji et al., 2014; Shang et al., 2022); and it presents a deltopectoral scar in the preaxial margin, which is absent in any other sauropterygian (ACH, pers. obs.) and, thus, it is considered here as a synapomorphy of Simosauridae.
The detailed comparison of the humerus of P. multidentatus with that of S. gaillardoti allows to recognize several differences. In this sense, the holotype of P. multidentatus (MUPA-ATZ0101) shows a more massive humerus in relation to the femur, and with a less pronounced diaphyseal constriction than that of S. gaillardoti (see Appendix 1, Figure S10). In addition, S. gaillardoti seems to have a higher humerus/femur length ratio (1.29 based on SMNS 14733 and GPIT-PV-31566) than MUPA-ATZ0101 (1.23) (see Rieppel, 1994; Huene, 1959; and Appendix 2, Table S7). Nevertheless, this measurement is based on incomplete elements for P. multidentatus, making this ratio dubious for this taxon and, hence, it is not considered here in the diagnosis; and a shallow ectepicondylar groove, which is absent in the almost complete right humerus of MUPA-ATZ0101 (see Appendix 1, Figure S10B, C, N, O). Moreover, the deltopectoral scar of S. gaillardoti ranges from small and rounded (e.g., SMNS 18686, SMNS 18698) to probably absent (e.g., SMNS 14733, SMNS 17590), being elongated (less than one fifth of the humerus total length) and relatively large exclusively in the humerus SMNS 52095. Therefore, the deltopectoral scar in S. gaillardoti never reaches one third of the humerus total length as in MUPA-ATZ0101 (Appendix 1, Figure S11A-E, P, Q, T, U). Several humeri of S. gaillardoti display a small and elongated foramen on their postaxial margin (e.g., SMNS 14733, SMNS 18686, SMNS 52095) (Appendix 1, Figure S11L-O, R, S, V, W), whereas in others the presence of this character cannot be confirmed due to preservation reasons (e.g., SMNS 17590; SMNS 18287) or it is absent (e.g., SMNS 18698, SMNS 7956). A foramen in the postaxial margin is also present in the right humerus of MUPA-ATZ0101 (Appendix 1, Figure S11F, G).
Hindlimb elements. The femur in both Paludidraco multidentatus and Simosaurus gaillardoti is notably less developed (i.e., slenderer, less massive) than the humerus, as in all other basal eosauropterygians (Rieppel, 1998b, 2000). The femur in Simosauridae shares the general morphology of the basal eosauropterygians, being weakly-curved, slim and with a weak development of the inner trochanter (i.e., not set off from the femur proximal region) (Rieppel, 2000). Hence, the femur of Simosauridae does not display any diagnostic character compared to the other basal eosauropterygians, differing only from that of Placodontia (i.e., with a well-defined and set off inner trochanter; Rieppel, 2000) and Plesiosauria (i.e., paddle-like; O’Keefe, 2001). In addition to the differences indicated above relative to the humerus/femur length ratio, the femur of P. multidentatus differs from that of S. gaillardoti in the presence of a femoral foramen (Appendix 1, Figure S11F-I) and in the higher robustness compared to the similarly sized femur #81 of SMNS 14733 (see Appendix 1, Figure S10G-L, R-X).
CONCLUSIONS
We provide a detailed description of the postcranial anatomy of the holotype of the Spanish simosaurid Paludidraco multidentatus, from the Carnian (Upper Triassic) of El Atance (Guadalajara, Central Spain). This specimen, which was only briefly described in its original publication, is exceptionally well-preserved and represents one of the most complete Iberian Triassic sauropterygians. Further preparation of this specimen has revealed previously unrecognized characters for this bizarre taxon, as well as for the relatively poorly known Simosauridae. Additionally, an updated diagnosis for Simosauridae is proposed based on a direct and detailed comparison of the holotype of P. multidentatus and all postcranial remains attributed to Simosaurus gaillardoti, the other representative of this clade identified as valid and identified in the Ladinian of France and Germany. Consequently, characters that were not previously considered are here identified as synapomorphies for Simosauridae within Sauropterygia (e.g., socketed vertebral centra, ovoid transverse processes of the dorsal vertebrae, and at least 32 dorsal vertebrae) and within Nothosauroidea (e.g., zygosphene without articular facets, and distinctly constricted humeral shaft). On the other hand, the vertebral centra distinctly constricted in ventral view, and the distinct expansion of distal area of sacral ribs, are no longer considered exclusive characters of Simosauridae, as their presence in certain Nothosaurus taxa is evidenced here. Finally, three characters not identified in any other sauropterygian are proposed as additional autapomorphies of P. multidentatus: infrapre- and infrapostzygapophyes in the cervical vertebrae, two symmetrical clavicular foramina, and an angled posteromedial margin of the coracoid. Paludidraco multidentatus can be unequivocally attributed to Simosauridae based on numerous characters shared with S. gaillardoti, as established in previous and the present discussions (e.g., relatively short and blunt snout, without rostral constriction; absence of premaxillary and dentary fangs; socketed vertebral centra; transverse processes of the dorsal vertebrae ovoid to oblong; infraprezygapophyses and infrapostzygapophyses present, at least in the dorsal, the sacral, and the most anterior caudal vertebrae; or presence of an anteromedial process in the clavicle). In addition, this Spanish taxon has several postcranial characters that notably differ from those of S. gaillardoti (e.g., highly pachyostotic sensu stricto dorsal vertebrae and ribs, semi-lunate interclavicle, not notched pubis, or a massive humerus).
ACKNOWLEDGEMENTS
This study was funded by the FPU grant (ref. FPU20/01945) (ACH) and the FPU mobility grant (ref. EST23/00375) (ACH) of the Spanish Ministerio de Universidades. The authors thank R. Schoch, E. Maxwell, E. Mujal (SMNS), and I. Werneburg (GPIT-PV) for the access to the specimens studied here, F. Marcos and her team (Universidad Complutense de Madrid, Madrid, Spain) for the preparation of the material, and the two anonymous reviewers for their kind and useful comments. This research received the support of the Viceconsejería de Cultura of the Consejería de Educación, Cultura y Deportes of Castilla-La Mancha (research project SBPLY/16/180801/000021) (APG).
REFERENCES
Agassiz, L. 1833-1843. Recherches sur les Poissons fossils. Neuchatel et Soleure 3 + 8, 390:1–32.
Bardet, N., Falconnet, J., Fischer, V., Houssaye, A., Jouve, S., Pereda Suberbiola, X., Pérez-García, A., Rage, J.C., and Vincent, P. 2014. Mesozoic marine reptile palaeobiogeography in response to drifting plates. Gondwana Research, 26:869–887.
https://doi.org/10.1016/j.gr.2014.05.005
Baur, G. Palaeohatteria Credner, and the Proganosauria. American Journal of Science 1889, 37:310–313.
Bickelmann, C. and Sander, P.M. 2008. A partial skeleton and isolated humeri of Nothosaurus (Reptilia: Eosauropterygia) from Winterswijk, The Netherlands. Journal of Vertebrate Paleontology, 28:326–338.
https://doi.org/10.1671/0272-4634 (2008)28[326:APSAIH]2.0.CO;2
Brown, D.S. 1981. The English Upper Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and classification of the Plesiosauria. Bulletin of British Museum of Natural History (Geology), 35:253–347.
Brum, A.S., Simões, T.R., Souza, G.A., Pinheiro, A.E.P., Figueiredo, R.G., Caldwell, M.W., Sayão, J.M., and Kellner, A.W.A. 2022. Ontogeny and evolution of the elasmosaurid neck highlight greater diversity of Antarctic plesiosaurians. Palaeontology, 65:1–21.
https://doi.org/10.1111/pala.12593
Case, E.C. 1936. A nothosaur from the Triassic of Wyoming. University of Michigan Contributions from the Museum of Paleontology, 5:1–36.
Cheng, Y.-N., Sato, T., Wu, X.-C., and Li, C. 2006. First complete pistosauroid from the Triassic of China. Journal of Vertebrate Paleontology, 26:501–504.
https://doi.org/10.1671/0272-4634 (2006)26[501:fcpftt]2.0.co;2
Cheng, Y., Wu, X., Sato, T., and Shang, H. 2016. Dawazisaurus brevis, a new eosauropterygian from the Middle Triassic of Yunnan, China. Acta Geologica Sinica (English Edition), 90:401–424.
https://doi.org/10.1111/1755-6724.12680
Dalla Vecchia, F.M. 2006. A new sauropterygian reptile with plesiosaurian affinity from the Late Triassic of Italy. Rivista Italiana di Paleontologia e Stratigrafia, 112:207–225.
Dalla Vecchia, F.M. 2008. First record of Simosaurus (Sauropterygia, Nothosauroidea) from the Carnian (Late Triassic) of Italy. Rivista Italiana Di Paleontologia e Stratigrafia, 114:273–285.
https://doi.org/10.13130/2039-4942/5902
Dalla Vecchia, F.M. 2017. Comments on the skeletal anatomy of the Triassic reptile Bobosaurus forojuliensis (Sauropterygia, Pistosauroidea). Gortania. Geologia, Paleontologia, Paletnologia, 38:39–75.
De Miguel Chaves, C., Pérez-García, A., Cobos, A., Royo-Torres, R., Ortega, F., and Alcalá, L. 2015. A diverse Late Triassic tetrapod fauna from Manzanera (Teruel, Spain). Geobios, 48:479–490.
https://doi.org/10.1016/j.geobios.2015.09.002
De Miguel Chaves, C., García-Gil, S., Ortega, F., Sanz, J.L., and Pérez-García, A. 2016. First Triassic tetrapod (Sauropterygia, Nothosauridae) from Castilla y León: Evidence of an unknown taxon for the Spanish record. Journal of Iberian Geology, 42:29–38.
https://doi.org/10.5209/rev_JIGE.2016.v42.n1.51210
De Miguel Chaves, C., Ortega, F., and Pérez-García, A. 2018a. New highly pachyostotic nothosauroid interpreted as a filter-feeding Triassic marine reptile. Biology Letters, 14:1–5.
https://doi.org/10.1098/rsbl.2018.0130
De Miguel Chaves, C., Ortega, F., and Pérez-García, A. 2018b. Cranial variability of the European Middle Triassic sauropterygian Simosaurus gaillardoti. Acta Palaeontologica Polonica, 63:315–326.
https://doi.org/10.4202/app.00471.2018
De Miguel Chaves, C., Ortega, F., and Pérez-García, A. 2020a. On the validity of the Middle Triassic sauropterygian ‘Partanosaurus zitteli ’ and the diversity of Simosauridae. Journal of Vertebrate Paleontology, 39:1–12.
https://doi.org/10.1080/02724634.2019.1704770
De Miguel Chaves, C., Ortega, F., and Pérez-García, A. 2020b. The Iberian Triassic fossil record of Sauropterygia: an update. Journal of Iberian Geology, 46:445–464.
https://doi.org/10.1007/s41513-020-00137-w
Diedrich, C.G. 2013a. The oldest “subaquatic flying” reptile in the world - Pistosaurus longaevus Meyer, 1839 (Sauropterygia) from the Middle Triassic of Europe. In Tanner, L.H., Spielmann, J.A., and Lucas, S.G. (eds.), 2013, The Triassic System. New Mexico Museum of Natural History and Science, Bulletin 61, pp. 169–215.
Diedrich, C.G. 2013b. The marine pachypleurosaur Serpianosaurus germanicus nov. spec.-skeleton and isolated bone remains from the pelsonian (Middle Triassic) of the European Germanic Basin carbonate intertidals and its paleobiology and taphonomy. In Tanner, L.H., Spielmann, J.A., and Lucas, S.G. (eds.), 2013, The Triassic System. New Mexico Museum of Natural History and Science, Bulletin 61, pp. 159–168.
Diedrich, C.G. 2013c. Shallow marine sauropterygian reptile biodiversity and change in the Bad Sulza formation (Illyrian, Middle Triassic) of Central Germany, and a contribution to the evolution of Nothosaurus in the Germanic Basin. In Tanner, L.H., Spielmann, J.A., and Lucas, S.G. (eds.), 2013, The Triassic System. New Mexico Museum of Natural History and Science, Bulletin 61, pp. 132–158.
Diedrich, C.G. 2014. Marine Lariosaurus (Sauropterygia) records from the Middle Triassic Germanic Basin - Stratigraphic and palaeobiogeographic importance for Tethyan and Germanic Basin correlation. Albertiana, 42:31–41.
Druckenmiller, P.S. and Russell, A.P. 2008. A phylogeny of Plesiosauria (Sauropterygia) and its bearing on the systematic status of Leptocleidus Andrews, 1922. Zootaxa, 1863:1–120.
https://doi.org/10.11646/zootaxa.1863.1.1
Edinger, T. 1921. Über Nothosaurus I. Ein Steinkern der Schädelhöhl. Senckenbergiana, 3:121–129.
García-Ávila, M., De La Horra, R., De Miguel Chaves, C., Juncal, M.A., Pérez-García, A., Ortega, F. and Diez, J.B. 2021. Palynological and sedimentological implications of the sauropterygian Upper Triassic site of El Atance (Central Iberian Peninsula). Review of Palaeobotany and Palynology, 295:1–15.
https://doi.org/10.1016/j.revpalbo.2021.104541
Haas, G. 1980. Ein Nothosaurier-Schädel aus dem Muschelkalk des Wadi Ramon (Negev. Israel). Annalen des Naturhistorischen Museums Wien, 83:19–125.
Haas, G. 1981. A fragmentary skull of Simosaurus (Reptilia: Sauropterygia) from the Middle Triassic of the Makhtesh Ramon, Israel. Israel Journal of Zoology, 30:30–34.
Hagdorn, H. and Rieppel, O. 1999. Stratigraphy of marine reptiles in the Triassic of central Europe. Zentralblatt für Geologie und Paläontologie, Teil I:651–678.
Houssaye, A., De Buffrénil, V., Rage, J.-C., and Bardet, N. 2008. An analysis of vertebral ‘pachyostosis’ in Carentonosaurus mineaui (Mosasauroidea, Squamata) from the Cenomanian (early Late Cretaceous) of France, with comments on its phylogenetic and functional significance. Journal of Vertebrate Paleontology, 28:685–691.
https://doi.org/10.1671/0272-4634 (2008)28[685:AAOVPI]2.0.CO;2
Jaekel, O. 1905. Über den Schädelbau der Nothosauriden. Sitzungsberichte Der Gesellschaft Naturforschender Freunde in Berlin, 60–84.
Ji, C., Jiang, D.Y., Rieppel, O., Motani, R., Tintori, A., and Sun, Z.Y. 2014. A new specimen of Nothosaurus youngi from the Middle Triassic of Guizhou, China. Journal of Vertebrate Paleontology, 34:465–470.
https://doi.org/10.1080/02724634.2013.808204
Jiang, D., Maisch, M.W., Sun, Z., Sun, Y., and Hao, W. 2006. A new species of Lariosaurus (Reptilia, Sauropterygia) from the Middle Anisian (Middle Triassic) of southwestern China. Neues Jahrbuch Für Geologie Und Paläontologie-Abhandlungen, 242:19–42.
Jiang, D.-Y., Rieppel, O., Motani, R., Hao, W.-C., Sun, Y.-L., Schmitz, L., and Sun, Z.-Y. 2008. A New Middle Triassic Eosauropterygian (Reptilia, Sauropterygia) from Southwestern China. Journal of Vertebrate Paleontology, 28:1055–1062.
https://doi.org/10.1671/0272-4634-28.4.1055
Jiang, D.-Y., Motani, R., Tintori, A., Rieppel, O., Chen, G.-B., Huang, J.-D., Zhang, R., Sun, Z.-Y., and Ji, C. 2014. The Early Triassic eosauropterygian Majiashanosaurus discocoracoidis, gen. et sp. nov. (Reptilia, Sauropterygia), from Chaohu, Anhui Province, People’s Republic of China. Journal of Vertebrate Paleontology, 34:1044–1052.
https://doi.org/10.1080/02724634.2014.846264
Kear, B.P., Rich T.H., Vickers-Rich, P., Ali, M.A., Al-Mufarrih, Y.A., Matari, A.H., Masary, A.M., and Halawani, M.A. 2010. A review of aquatic vertebrate remains from the Middle-Upper Triassic Jilh Formation of Saudi Arabia. Proceedings of the Royal Society of Victoria, 122:1–8.
https://doi.org/10.1071/RS10001
Klein, N., Voeten, D.F.A.E., Lankamp, J., Bleeker, R., Sichelschmidt, O.J., Liebrand, M., NIEWEG, D.C., and Martin Sander, P. 2015. Postcranial material of Nothosaurus marchicus from the Lower Muschelkalk (Anisian) of Winterswijk, The Netherlands, with remarks on swimming styles and taphonomy. Palaontologische Zeitschrift, 89:961–981.
https://doi.org/10.1007/s12542-015-0273-5
Klein, N., Eggmaier, S., and Hagdorn, H. 2022. The redescription of the holotype of Nothosaurus mirabilis (Diapsida, Eosauropterygia) - a historical skeleton from the Muschelkalk (Middle Triassic, Anisian) near Bayreuth (southern Germany). PeerJ, 10:e13818.
https://doi.org/10.7717/peerj.13818
Kuhn-Schnyder, E. 1961. Der Schädel von Simosaurus. Paläontologische Zeitschrift, 35:95–113.
Li, Q. and Liu, J. 2020. An Early Triassic sauropterygian and associated fauna from South China provide insights into Triassic ecosystem health. Communications Biology, 3:1–11.
https://doi.org/10.1038/s42003-020-0778-7
Lin, K. and Rieppel, O. 1998. Functional morphology and ontogeny of Keichousaurus hui (Reptilia, Sauropterygia). Fieldiana: Geology, 39:1–35.
https://doi.org/10.5962/bhl.title.5174
Lin, W.-B., Jiang, D.-Y., Rieppel, O., Motani, R., Ji, C., Tintori, A., Sun, Z.-Y., and Zhou, M. 2017. A new specimen of Lariosaurus xingyiensis (Reptilia, Sauropterygia) from the Ladinian (Middle Triassic) Zhuganpo Member, Falang Formation, Guizhou, China. Journal of Vertebrate Paleontology, 37:e1278703.
https://doi.org/10.1080/02724634.2017.1278703
Liu, J., Rieppel, O., Jiang, D., Aitchison, J.C., Motani, R., Zhang, Q.-Y., Zhou, C.-Y., and Sun, Y.-Y. 2011. A new pachypleurosaur (Reptilia: Sauropterygia) from the Lower Middle Triassic of Southwestern China and the phylogenetic relationships of Chinese pachypleurosaurs. Journal of Vertebrate Paleontology, 31:292–302.
https://doi.org/10.1080/02724634.2011.550363
Liu, J., Hu, S.X., Rieppel, O., Jiang, D.Y., Benton, M.J., Kelley, N.P., Aitchison, J.C., Zhou, C.Y., Wen, W., Huang, J.Y., Xie, T., and Lv, T. 2014. A gigantic nothosaur (Reptilia: Sauropterygia) from the Middle Triassic of SW China and its implication for the Triassic biotic recovery. Scientific Reports, 4:1–9.
https://doi.org/10.1038/srep07142
Münster, G. 1834. Vorläufige Nachricht über einige neue Reptilien im Muschelkalke von Baiern. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde, 1834:521–527.
O’Keefe, F.R. 2001. A cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia). Acta Zoologica Fennica, 213:1–63.
Owen, R. 1860. Palaeontology; or, a Systematic Summary of Extinct Animals and their Geologic Remains. Adam and Charles Black, Edinburgh, 420 p.
https://doi.org/10.5962/bhl.title.13917
Persons, W.S., Street, H.P., and Kelley, A. 2022. A long-snouted and long-necked polycotylid plesiosaur from the Late Cretaceous of North America. IScience, 105033.
https://doi.org/10.1016/j.isci.2022.105033
Renesto, S. 2010. A new specimen of Nothosaurus from the latest Anisian (Middle Triassic) Besano Formation (Grenzbitumenzone) of Italy. Rivista Italiana Di Paleontologia e Stratigrafia, 116:145–160.
Reolid, M. and Reolid, J. 2020. First record of Triassic marine reptiles (Nothosauria, Sauropterygia) from the Alpujarride complex (internal zones of the Betic Cordillera, Spain). Estudios Geológicos, 76:1–16.
https://doi.org/10.3989/EGEOL.43592.535
Rieppel, O. 1989. A New Pachypleurosaur (Reptilia: Sauropterygia) from the Middle Triassic of Monte San Giorgio, Switzerland. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 323:1–73.
https://doi.org/10.1098/rstb.1989.0001
Rieppel, O. 1994. Osteology of Simosaurus gaillardoti and the relationships of stem-group Sauropterygia. Fieldiana: Geology, 28:1–85.
https://doi.org/10.5962/bhl.title.3399
Rieppel, O. 1996. The status of the sauropterygian reptile Partanosaurus zitteli Skuphos from the Middle Triassic of the Austrian Alps, with comments on Microleptosaurus schlosseri Skuphos. Paläontologische Zeitschrift, 70:567–577.
Rieppel, O. 1998a. Corosaurus alcovensis Case and the phylogenetic interrelationships of Triassic stem-group Sauropterygia. Zoological Journal of the Linnean Society, 124:1–41.
https://doi.org/10.1111/j.1096-3642.1998.tb00568.x
Rieppel, O. 1998b. The status of the sauropterygian reptile genera Ceresiosaurus, Lariosaurus, and Silvestrosaurus from the Middle Triassic of Europe. Fieldiana: Geology, 38:1–46.
https://doi.org/10.5962/bhl.title.3413
Rieppel, O. 1999. The sauropterygian genera Chinchenia, Kwangsisaurus, and Sanchiaosaurus from the Lower and Middle Triassic of China. Journal of Vertebrate Paleontology, 19:321–337.
https://doi.org/10.1080/02724634.1999.10011144
Rieppel, O. 2000. Part 12A. Sauropterygia I. In Wellnhofer, P. (ed.), Handbuch der Paläoherpetologie. München: Verlag Friedrich Pfeil, pp. 1–134.
Rieppel, O. 2001. A new species of Nothosaurus (Reptilia: Sauropterygia) from the Upper Muschelkalk (Lower Ladinian) of southwestern Germany. Palaeontographica Abteilung A, 263:137–162.
https://doi.org/10.1127/pala/263/2001/137
Rieppel, O. and Lin, K. 1995. Pachypleurosaurs (Reptilia: Sauropterygia) from the Lower Muschelkalk, and a review of the Pachypleurosauroidea. Fieldiana: Geology, 32:1–44.
https://doi.org/10.5962/bhl.title.3474
Rieppel, O. and Wild, R. 1996. A Revision of the Genus Nothosaurus (Reptilia: Sauropterygia) from the Germanic Triassic, with Comments on the Status of Conchiosaurus clavatus. Fieldiana: Geology, 34:1–82.
https://doi.org/10.5962/bhl.title.2691
Rieppel, O., Mazin, J.-M., and Tchernov, E. 1997. Speciation along rifting continental margins: a new Nothosaur from the Negev (Israel). Earth & Planetary Sciences, 325:991–997.
https://doi.org/10.1016/s1251-8050 (97)82380-4
Rieppel, O., Mazin, J.-M., and Tchernov, E. 1999. Sauropterygia from the Middle Triassic of Makhtesh Ramon, Negev, Israel. Fieldiana: Geology, 40:1–85.
https://doi.org/10.5962/bhl.title.3354
Sander, P.M., Rieppel, O., and Bucher, H. 1997. A new pistosaurid (Reptilia: Sauropterygia) from the Middle Triassic of Nevada and its implications for the origin of the plesiosaurs. Journal of Vertebrate Paleontology, 17:526–533.
https://doi.org/10.1080/02724634.1997.10010999
Sander, P.M., Klein, N., Albers, P.C.H., Bickelmann, C., and Winkelhorst, H. 2013. Postcranial morphology of a basal Pistosauroidea (Sauropterygia) from the Lower Muschelkalk of Winterswijk, The Netherlands. Palaontologische Zeitschrift, 88:55–71.
https://doi.org/10.1007/s12542-013-0181-5
Sanz, J.L. 1983. Consideraciones sobre el género Pistosaurus. El Suborden Pistosauria (Reptilia, Sauropterygia). Estudios Geológicos, 39:451–458.
Sanz, J.L. 1984. Osteología comparada de las familias Nothosauridae y Simosauridae (Reptilia, Sauropterygia). Estudios Geológicos, 40:81–105.
https://doi.org/10.3989/egeol.84401-2652
Sato, T., Cheng, Y.N., Wu, X.C., and Li, C. 2010. Osteology of Yunguisaurus Cheng et al., 2006 (Reptilia; Sauropterygia), a Triassic pistosauroid from China. Paleontological Research, 14:179–195.
https://doi.org/10.2517/1342-8144-14.3.179
Sato, T., Zhao, L.J., Wu, X.C., and Li, C. 2014a. A new specimen of the Triassic pistosauroid Yunguisaurus, with implications for the origin of Plesiosauria (Reptilia, Sauropterygia). Palaeontology, 57:55–76.
https://doi.org/10.1111/pala.12048
Sato, T., Cheng, Y.N., Wu, X.C., and Shan, H.Y. 2014b. Diandongosaurus acutidentatus Shang, Wu & Li, 2011 (Diapsida: Sauropterygia) and the relationships of Chinese eosauropterygians. Geological Magazine, 151:121–133.
https://doi.org/10.1017/S0016756813000356
Seeley, H.G. 1874. On Muraenosaurus leedsi, a plesiosaurian from the Oxford Clay. Part I, Quarterly Journal of the Geological Society of London, 30:197–208.
https://doi.org/10.1144/GSL.JGS.1874.030.01-04.35
Segesdi, M. and Ősi, A. 2021. Sauropterygian remains from the Middle Triassic of Villány, Hungary–new information on the aquatic reptile fauna of Tisza Megaunit (Triassic southern Eurasian shelf region). Palaeobiodiversity and Palaeoenvironments, 101:985–1011.
https://doi.org/10.1007/s12549-020-00480-x
Sennikov, A.G. 2001. Discovery of a Primitive Sauropterygian from the Lower Triassic of the Donskaya Luka (Don Basin) and the Range of Triassic Marine Reptiles in Russia. Paleontological Journal, 35:301–309.
Shang, Q.-H. 2006. A new species of Nothosaurus from the early Middle Triassic of Guizhou, China. Vertebrata PalAsiatica, 44:237–249.
Shang, Q.-H., Wu, X.-C., and Li, C. 2011. A new Eosauropterygian from the Middle Triassic of Eastern Yunnan Province, Southwestern China. Vertebrata PalAsiatica, 4:155–171.
https://doi.org/10.1017/S0016756813000356
Shang, Q.-H. and Li, C. 2015. A new small-sized eosauropterygian (Diapsida: Sauropterygia) from the Middle Triassic of Luoping, Yunnan, southwestern China. Vertebrata PalAsiatica, 53:265–280.
Shang, Q.-H., Li, C., and Wu, X.-C. 2017. New information on Dianmeisaurus gracilis Shang & Li, 2015. Vertebrata PalAsiatica, 4:145–161.
Shang, Q.-H., Wu, X.-C., and Li, C. 2020. A New Ladinian Nothosauroid (Sauropterygia) from Fuyuan, Yunnan Province, China. Journal of Vertebrate Paleontology, 40:e1789651.
https://doi.org/10.1080/02724634.2020.1789651
Shang, Q.-H., Li, C., and Wang, W. 2022. Nothosaurus luopingensis n. sp. (Sauropterygia) from the Anisian, Middle Triassic of Luoping, Yunnan Province, China. Vertebrata PalAsiatica.
https://doi.org/10.19615/j.cnki.2096-9899.220524
Skuphos, T. 1893. Uber Partanosaurus Zitteli Skuphos und Microleptosaurus Schlosseri nov. gen., nov. spec., aus den Vorarlberger Partnachschichten. Abhandlungen der kaiserlichköniglichen geologischen Reichsanstalt, 15:1–16.
Storrs, G.W. 1991. Anatomy and relationships of Corosaurus alcovensis (Diapsida: Sauropterygia) and the Triassic Alcova Limestone of Wyoming. Bulletin of the Peabody Museum of Natural History Yale University, 44:1–151.
Vickers-Rich, P., Rich, T.H., Rieppel, O., Thulborn, R.A., and Mcclure, H.A. 1999. A Middle Triassic vertebrate fauna from the Jilh Formation, Saudi Arabia. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 213:201–232.
Vincent, P. and Storrs, G.W. 2019. Lindwurmia, a new genus of Plesiosauria (Reptilia: Sauropterygia) from the earliest Jurassic of Halberstadt, northwest Germany. Science of Nature, 106:1–2.
https://doi.org/10.1007/s00114-018-1600-y
von Huene, F. 1921. Neue beobachtungen an Simosaurus. Acta Zoologica, 2:1–2.
von Huene, F. 1948. Simosaurus and Corosaurus. American Journal of Science, 246:41–43.
https://doi.org/10.2475/ajs.246.1.41
von Huene, F. 1952. Skelett und Verwandtschaft von Simosaurus. Palaeontographica A, 102:163–182.
von Huene, F. 1959. Simosaurus guilielmi aus dem unteren Mittelkeuper von Obersontheim. Palaeontographica A, 113:180–184.
von Meyer, H. 1839. Mittheilung an Professor Bronn gerichtet. Neues Jahrbuch für Mineralogie Geogosie und Geol Petrefaktenkunde, 1839:559–560.
von Meyer, H. 1842. Simosaurus, die Stumpfschnauze, ein Saurier aus dem Muschelkalke von Luneville. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefakten-Kunde, 1842:184–197.
Wu, X.-C., Zhao, L.-J., and Sato, T. 2011. New information on Wumengosaurus delicatomandibularis Jiang et al., 2008 (Diapsida: Sauropterygia), with a revision of the osteology and phylogeny of the taxon. Journal of Vertebrate Paleontology, 31:70–83.
Xue-Qing, L., Wen-Bin, L., Rieppel, O., Zuo-Yu, S., Zhi-Guang, L., Hao, L., and Da-Yong, J. 2015. A new specimen of Diandongosaurus acutidentatus (Sauropterygia) from the Middle Triassic of Yunnan, China. Vertebrata PalAsiatica, 10:281–290.
