

 A Miocene ant species of the genus Forelius Emery, 1888 (Hymenoptera: Formicidae: Dolichoderinae) from Mexico
A Miocene ant species of the genus Forelius Emery, 1888 (Hymenoptera: Formicidae: Dolichoderinae) from Mexico
Article number: 26.3.a36
https://doi.org/10.26879/1294
Copyright Paleontological Society, September 2023
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 14 April 2023. Acceptance: 3 September 2023.
ABSTRACT
The first fossil ant species of the New World genus Forelius (Formicidae: Dolichoderinae) is described. The type material is a Miocene amber inclusion from the Montecristo mine near Simojovel, Chiapas, southwestern Mexico. The new species is named Forelius chenpauch sp. nov. It differs from its congeners by having a unique combination of characters: antennal scapes reaching the posterior margin of the head; pronotum with six erect hairs; mesonotum with four erect hairs; propodeum with four erect hairs; front and dorsum of head, dorsum of pronotum, dorsum of mesonotum and propodeum covered with a fine striation. Forelius chenpauch sp. nov. has a rounded spiracle, which could be considered a plesiomorphic character closely related to the Forelius group from the north of the Amazon basin. Accordingly, the current divergence estimates of Forelius as a single genus from the Leptomyrmex + Forelius + Dorymyrmex clade (ca. 27 Ma) and the occurrence of Forelius chenpauch sp. nov. in the fossil record (ca. 23 Ma), may suggest that the rounded spiracle is a plesiomorphic state probably widespread since the Miocene from southern Mexico to northern South America.
Fernando Varela-Hernández. Laboratorio de Sistemática Molecular, Escuela de Estudios Superiores del Jicarero, Universidad Autónoma del Estado de Morelos, C. P. 62909, Jojutla, Morelos, México. Corresponding author. fernando.varela@uaem.mx
Francisco Riquelme. Laboratorio de Sistemática Molecular, Escuela de Estudios Superiores del Jicarero, Universidad Autónoma del Estado de Morelos, C. P. 62909, Jojutla, Morelos, México. Corresponding author. francisco.riquelme@uaem.mx
Roberto J. Guerrero. Universidad del Magdalena, Santa Marta, Magdalena, Colombia, rguerrero@unimagdalena.edu.co
Keywords: ant fossil; amber- Lagerstätte; Chiapas amber; Leptomyrmecini tribe; new species
Final citation: Varela-Hernández, Fernando, Riquelme, Francisco, and Guerrero, Roberto J. 2023. A Miocene ant species of the genus Forelius Emery, 1888 (Hymenoptera: Formicidae: Dolichoderinae) from Mexico. Palaeontologia Electronica, 26(3):a36.
https://doi.org/10.26879/1294
palaeo-electronica.org/content/2023/3951-a-new-ant-fossil-species
Copyright: September 2023 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
https://zoobank.org/62019DB5-8F62-4276-994C-8867124393A6
INTRODUCTION
The ant genus Forelius Emery, 1888 (Leptomyrmecini, Dolichoderinae) is endemic to the New World. It is distributed in arid and semi-arid ecosystems from the USA to Argentina (Guerrero, 2019). At the beginning of the twentieth century, the genus was considered monotypic, with Forelius mccooki (McCook, 1880) being the only recognizable species. Since then, new species were subsequently described or recently transferred to the genus (Emery, 1905; Cuezzo, 2000; Guerrero and Fernández, 2008; Guerrero, 2021). Cuezzo (2000) has extensively reviewed the nomenclature and taxonomy of the genus Forelius, describing new species and expanding their distribution range, particularly those in the southern hemisphere. Forelius comprises 19 extant species (Bolton, 2023) with no fossil species described to date. Ten years ago, a fossil ant specimen assigned to Forelius was recorded for the first time in Miocene Mexican amber, but no formally described species (Durán-Díaz et al., 2013).
In the present paper, we describe the first species that match the genus Forelius (sensu Shattuck, 1992 and Cuezzo, 2000), based on a Miocene amber inclusion recently recovered from the Montecristo mine in Simojovel, Chiapas, Mexico. Due to the preservation quality of its fossil inclusions, the Simojovel amber mines are part of a fossil preservation site known as Amber-Lagerstätte (Riquelme et al., 2014a, b); which also includes the mines and pits from Totolapa and Estrella de Belén near Palenque in the Chiapas Highlands (Riquelme et al., 2015). The Simojovel amber deposits belong to the Miocene-assigned Balumtum and Mazantic strata (Perrilliat et al., 2010; Durán-Díaz et al., 2013; Riquelme et al., 2014a, b). Another amber outcrop near Simojovel assigned to the La Quinta strata, predominantly marine and of Late Oligocene age has also been reported (Allison, 1967; Frost and Langenheim, 1974). The sedimentary environment of the amber deposits suggests a lowland fluvial environment close to the coastal plain (Allison, 1967; Frost and Langenheim, 1974; Graham, 1999; Langenheim, 2003). An extinct leguminous tree of the genus Hymenaea Linné is the botanical source of Chiapas amber from Miocene (Langenheim, 2003; Riquelme et al., 2014a)
MATERIALS AND METHODS
The fossil specimen studied here is an amber inclusion from the Montecristo mine near Simojovel, Chiapas, Southwestern Mexico. The fossil is embedded in a piece of translucent golden amber that has been cut and polished to obtain a flat surface, which allows observation by attenuating the optical distortions caused by the amber. Morphological data were collected using an Olympus® AZ binocular dissecting microscope (Laboratorio de Sistemática Molecular, Universidad Autónoma del Estado de Morelos), a Carl Zeiss® AXIO ZOOM.V16 microscope coupled to an Axiocam MRc5 camera, and the ZEN 2012 (Blue edition) program to stack the photomicrographs (Laboratorio de Microscopía y Fotografía de la Biodiversidad II, Instituto de Biología, Universidad Nacional Autónoma de México). Schematic drawings were hand traced using a stereomicroscope and photomicrographs in an electronic tablet. The Corel Draw X7® program was used for graphic editing. Taxonomy and terminology follow Shattuck (1992) for the genus and for the species Cuezzo (2000). Measurements and indices follow Cuezzo (2000) using an ocular micrometer at 80x. All the following measurements are expressed in millimeters:
TL = Total length. In lateral view, taken from anterior clypeal margin (excluding mandibles) to apex of last segment of metasoma.
HL = Head length. Maximum length, in full face view, from apex of clypeal plate to middle of posterior vertex margin.
HW = Head width. Maximum width in full face view excluding eyes.
EL = Eye length. Maximum length of eye in full face view
EW = Eye width. Width of eye perpendicular to maximum length.
IOD = Distance between inner-most edges of eyes. Measured in full face view, distance between edges of eyes.
COD = Clypeus-ocular distance. Shortest distance between posterior margin of clypeus and anterior margin of eye.
SL = Scape length. Maximum length of first antennal segment, excluding neck and basal condyle.
PSL = Propodeal spiracle length. Maximum length of propodeal spiracle.
PSW = Maximum width of propodeal spiracle. Measured perpendicularly to maximum length.
MsL = Mesosoma length. Straight-line distance measured in lateral view from anterior margin of pronotum (excluding collar) to posterior extremity of metapleural lobe.
CI = Cephalic index: (HW/HL)*100.
OI = Ocular index: (EL/EW)*100.
SI= Scape index: (SL/HW)*100.
The fossil inclusion was assigned as a holotype CPAL.464, which belongs to the Colección de Paleontología maintained at the Universidad Autonoma del Estado de Morelos, Mexico.
SYSTEMATIC PALEONTOLOGY
Class Insecta Linnaeus, 1758
Order Hymenoptera Linnaeus, 1758
Family Formicidae Latreille, 1809
Subfamily Dolichoderinae Forel, 1878
Tribe Leptomyrmecini Emery, 1913
Genus Forelius Emery, 1888
Type species. Iridomyrmex mccooki McCook, 1880
Forelius chenpauch sp. nov. Varela-Hernández, Guerrero and Riquelme
(Figure 1-Figure 2)
zoobank.org/795FBFFB-5934-4259-899E-E60A69DA1319
Etymology. The specific epithet chenpauch means “from the amber cave” in the Tzotzil language, a name given by the local people of the Simojovel to the amber mines or pits.
 Type material. Holotype CPAL.464 Worker, amber inclusion, complete specimen. It is deposited in the Colección de Paleontología, UAEM, Morelos, Mexico (Figure 1A)
Type material. Holotype CPAL.464 Worker, amber inclusion, complete specimen. It is deposited in the Colección de Paleontología, UAEM, Morelos, Mexico (Figure 1A)
Locality. Mexico, Chiapas, Simojovel, Montecristo mine: 17° 7' 45.99" N, 92°41'43" W.
Distribution. It is only known from the Miocene strata of Simojovel, Chiapas, southern Mexico.
Horizon. The Mazantic and Balumtum strata, Early Miocene (ca. 23 Ma).
Diagnosis. Antennal scape reaching the posterior margin of the head; pronotum with six erect hairs; mesonotum with four erect hairs; propodeum with four erect thin hairs; front and dorsum of head, dorsum of pronotum, dorsum of mesonotum, and propodeum covered with fine longitudinal striae.
Worker description
Measurements (mm). TL: 2.0; HL: 0.6; HW: 0.6; EL: 0.1; EW: 0.1; IOD: 0.4; SL: 0.5; PSL: 0.05; PSW: 0.05; MsL: 0.8; CI: 100; OI: 100; SI: 83.33.
Head. As long as broad; vertex of head weakly concave; antennal scapes reaching the posterior margin of the head; mandible with a basal and masticator margin differentiated, with six observable teeth in the latter (Figure 2B); clypeus with two slender, erect setae projecting onto the mandibles at the middle part, about the level of the antennal insertions; two other shorter setae on each side of the erect setae described previously: one at the lateral edge of the mandible and another between the latter and the one at the middle of the mandible; the dorsal face of mandibles covered with decumbent pilosity; dorsum of head finely striated (Figure 1B, Figure 2A, 2B).
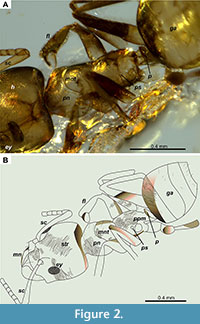 Mesosoma. Dorsal profile discontinuous. In dorsal view, well-impressed promesonotal suture; weak mesonotal groove in lateral view; pronotum with six thin, erect hairs, four located anteriorly, longer than the other two located posteriorly, two at the anteriormost humeral and the other two at the middle; dorsal surface of pronotum and mesonotum with weak transversal striae; dorsal and posterior face of propodeum rounded, with no apparent angle that differentiates the two faces; one pair of thin, erect hairs located dorsolaterally on the propodeum dorsal face and another pair posterolaterally on the propodeum posterior face; propodeal spiracle small and rounded (Figure 2A, 2B). Legs without pilosity.
Mesosoma. Dorsal profile discontinuous. In dorsal view, well-impressed promesonotal suture; weak mesonotal groove in lateral view; pronotum with six thin, erect hairs, four located anteriorly, longer than the other two located posteriorly, two at the anteriormost humeral and the other two at the middle; dorsal surface of pronotum and mesonotum with weak transversal striae; dorsal and posterior face of propodeum rounded, with no apparent angle that differentiates the two faces; one pair of thin, erect hairs located dorsolaterally on the propodeum dorsal face and another pair posterolaterally on the propodeum posterior face; propodeal spiracle small and rounded (Figure 2A, 2B). Legs without pilosity.
Petiole. Scale present, inclined anteriorly, with a pair of erect setae, barely visible.
Gaster. First and second tergite with a row of short, thin, erect setae running along the apical fringe of tergites, sternites lost; gaster concolorous yellow with mesosoma.
Remarks. The worker (Holotype CPAL.464) studied here is classified in the genus Forelius since it presents the diagnostic characters of the genus, that is, the anterior clypeal margin with two long curved setae is observed in the specimen, which, although in the specimen the mandible is open, these clypeal hairs project reaching the external margin of the mandible. The configuration of the mandible and the dentition pattern on the masticatory margin agree with that of Forelius according to Shattuck (1992), that is, mandible with a smooth basal and masticatory margin differentiated, the latter armed with 6-7 teeth. The petiolar scale is reduced and strongly inclined anteriorly as well, however, in the specimen it appears relatively straight (Figure 1-Figure 2), but it is due to the position within the amber piece. The latter also makes it difficult to observe the first gastral segment projecting anteriorly and concealing petiole in dorsal view; however, an inclined view of the piece allowed us to observe a shallow depression on the anterior face of the first gastral tergite. Forelius chenpauch sp. nov. is a species with a rounded spiracle but differs from other Forelius species with this same condition by the characters listed below (Table 1).
Forelius chenpauch sp. nov. could be confused with extant dolichoderine ants of the genus Liometopum Mayr, 1861 and the genus Tapinoma Foerster, 1850, which can currently be found in Mexico. However, Forelius chenpauch sp. nov. is distinguished by having a mandible with a basal margin distinct from the masticatory margin (Figure 1A, 1B, Figure 2B), whereas Liometopum and Tapinoma have an undifferentiated basal margin. In addition, Forelius chenpauch sp. nov. has a petiole scale anteriorly inclined (Figure 1A, Figure 2A, 2B). In contrast, Liometopum has a vertical petiolar scale, that is, never anteriorly inclined; and in the currently known Neotropical species of Tapinoma the petiolar scale is strongly reduced, in most hidden by the first gastral segment (Guerrero unpublished data).
On the other hand, the genus Forelius and the genus Azteca Forel, 1878 have been recorded in Mexican and Dominican amber (Duran-Ruiz et al., 2013; Barden, 2017). However, the fossil record of Forelius is uncommon in both deposits, whereas Azteca is more frequent. Forelius is composed of hyperactive thermophilic ground-nesting ants (Ward, 2005) that are adapted to living in arid to semi-arid areas (Shattuck, 1992; Cuezzo, 2000) whilst Azteca encompasses ants that nest mainly in trees (Longino, 2007). Notably, the species Azteca alpha Wilson, 1985 constitutes 30 to 50% of the total number of ants found in Dominican amber (Barden, 2017). The highest frequency of Azteca records is consistent with an arboreal environment. The sedimentary data in Mexican amber deposits suggests a lowland fluvial environment close to the coastal plain (Allison, 1967; Frost and Langenheim, 1974; Graham, 1999; Langenheim, 2003; Riquelme et al., 2014a, 2015). Accordingly, the leguminous trees of Hymenaea (the botanical source of Mexican amber) were part of a tropical forest resembling modern mangroves (Langenheim, 2003) where a warmer (+1.99 °C) and drier (−337.7 mm) tropical climate have been estimated than the current average records (Hernández-Hernández et al., 2020). Although forested areas may have predominated, the Miocene tropical forest in southern Mexico possibly presented minor arid to semi-arid zones that the Forelius could exploit to locate the colonies.
DISCUSSION
Taxonomy in the genus Forelius has received attention in the last decades. Initially, Shattuck (1992) offered clear morphological boundaries for the genus, allowing it to be differentiated from other relatively similar dolichoderine ants (e.g., Dorymyrmex Mayr, 1866; Linepithema Mayr, 1866 or Tapinoma). Cuezzo (2000) reviewed the Forelius species in the New World recognizing 18 species, some of which present uncertainties in their taxonomic limits and need to be revised—i.e., evidence of intergradation between F. pruinosus (Roger, 1863) and F. mccooki (McCook, 1860) (Ward, 2005). Later works have increased the number of species within the genus (Guerrero and Fernandez 2008) and resolved some nomenclatural problems (Guerrero 2021) within the genus. In this work, we describe the first fossil Forelius species preserved in amber; a species with possible evolutionary affinity with those of northern South America and Central America due to the presence of the rounded spiracle.
The shape of the spiracle in Forelius was used as a taxonomic argument by Kusnezov (1963) to propose different genus names. This author proposed that the species with an elongated propodeal spiracle (slit-shaped), exclusive to South America, should be named Neoforelius Kusnezov, 1953 (a junior synonym of Forelius Shattuck, 1992) while those with a rounded propodeal spiracle should be classified as Forelius. Recent studies suggest that species with rounded propodeal spiracle occur north of the Amazon basin (Guerrero and Fernández, 2008), except for F. antarticus and F. heyerii (Guerrero, 2021). The other species with slit-shaped propodeal spiracles occur south of the Amazon basin. This distributional pattern separates two groups with a polymorphism associated to that trait (Guerrero and Fernández, 2008).
The occurrence of rounded spiracles on the new species Forelius chenpauch sp. nov. reinforces a biogepographic pattern previously described by Guerrero and Fernández, (2008), in which the few species found north of the Amazon basin all have round spiracles, while south of the Amazon many species have elongate, slit-like spiracles, and only two species have round spiracles. Based on the distribution pattern, round spiracles could be plesiomorphic, apomorphic, or homoplasious in the genus Forelius. The fossil species Forelius chenpauch sp. nov. has a rounded propodeal spiracle, which could be considered a closely related plesiomorphic trait to those species distributed north of the Amazon basin (e.g., Forelius pruinosus). Thus, Forelius chenpauch sp. nov. is an essential clue to understanding the evolution of the Forelius genus. Whitin Leptomyrmecini, Forelius and Dorymyrmex probably diverged more than 35 Ma ago (Ward et al., 2010). If we consider the estimated upper limit for the occurrence of Forelius crown group (ca. 27 Ma), then Forelius chenpauch sp. nov., which lived in the Miocene (ca. 23 Ma) of southern Mexico, may be closely related to extant species distributed in northern of South America and Central America (i.e., north of the Amazon basin).
If the Forelius divergence times are true (Ward et al., 2010) and Forelius chenpauch sp. nov. shares common ancestry with the rest of the species distributed north of the Amazon basin, then it could be suggested that the rounded spiracle is a plesiomorphic state probably widespread since the Miocene from Southern Mexico to northern South America. Although the previously exposed hypothesis could receive support from the fossil record and estimation of divergence times, the presence of this morphological trait in F. antarcticus and F. heyeri is not supported; a possible explanation would be the independent evolution north and south of the Amazon (Guerrero and Fernández 2008). Another subsequent diversification of species with slit-shaped spiracle should occur south of the Amazon basin (Guerrero and Fernández, 2008). Further phylogenetic work on the genus is needed to differentiate among these hypotheses. Phylogenetic information integrating a broad taxon sampling within Forelius, genomic data and detailed morphological analysis of fossil species such as Forelius chenpauch sp. nov. can provide more robust hypotheses about the evolution of the genus Forelius and fill in the gaps regarding the evolution of propodeal spiracle shape.
ACKNOWLEDGMENTS
We thank S. Guzmán-Gómez at the LMF2-LANABIO (IBUNAM) and V. Córdova-Tabares at the ENCB-IPN, for photomicrography assistance. We also thank J. Núñez -Dávalos for the drawing. RJG is supported by MinCiencias through the project “Patrones de diversidad histórica y ecológica de las hormigas en el socioecosistema bosque seco tropical de Colombia y sus implicaciones para la conservación” with contract 2021-1029 signed by the Universidad del Magdalena and Icetex. We thank the Handling Editor and anonymous reviewers whose comments have improved the final version of this paper.
REFERENCES
Barden, P. and Grimaldi, D.A. 2016. Adaptive radiation in socially advanced stem-group ants from Cretaceous. Current Biology, 26(4):515–521.
https://doi.org/10.1016/j.cub.2015.12.060
Barden, P. 2017. Fossil ants (Hymenoptera: Formicidae): ancient diversity and the rise of modern lineages. Myrmecological News, 24:1–30.
Bolton, B. 2023. An online catalog of the ants of the world.
http://antcat.org/
Cuezzo, F. 2000. Revisión del género Forelius (Hymenoptera: Formicidae: Dolichoderinae). Sociobiology, 35(2A):197–275.
Cuezzo, F. 2003. Subfamilia Dolichoderinae, p. 291–297. En Fernández, F. (ed.), Introducción a las hormigas de la región Neotropical. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá.
Durán-Ruíz, C., Riquelme, F., Coutiño-José, M., Carbot-Chanona, G., Castaño-Meneses, G., and Ramos-Arias, M. 2013. Ants from Miocene Totolapa amber (Chiapas, Mexico) with the first record of the genus Forelius (Hymenoptera, Formicidae). Canadian Journal of Earth Sciences, 50:485–502.
https://doi.org/10.1139/cjes-2012-0166
Emery, C. 1888d. Über den sogenannten Kaumagen einigerAmeisen. Zeitschrift für Wissenschaftliche Zoologie, 46:378–412.
Emery, C. 1905. Studi sulle formiche della fauna neotropica. Bulletino della Società Entomologica Italiana, 37(1905):107–194.
Emery, C. 1913. Hymenoptera. Fam. Formicidae. Subfam. Dolichoderinae. Genera Insectorum 137:1–50.
Forel, A. 1878. Études myrmécologiques en 1878 (première partie) avec l'anatomie du gésier des fourmis. Bulletin de la Société Vaudoise des Sciences Naturelles, 15:337–392.
Frost, S.H. and Langenheim, R.L. 1974. Cenozoic reef biofacies, tertiary larger Foraminifera and scleractinian corals from Chiapas, Mexico. Northern Illinois University Press, DeKalb.
Graham, A. 1999. Studies in neotropical paleobotany. XIII. An Oligo-Miocene palynoflora from Simojovel (Chiapas, Mexico). American Journal of Botany, 86(1):17–31.
https://doi.org/10.2307/2656951
Guerrero, R.J. and Fernández, F. 2008. A new species of the ant genus Forelius (Formicidae: Dolichoderinae) from the dry forest of Colombia. Zootaxa, 1958:51–60.
https://doi.org/10.11646/zootaxa.1958.1.5
Guerrero, R.J. 2019. Subfamilia Dolichoderinae, p. 693–720. In Fernández, F., Guerrero, R.J., and Denise, T. (eds.), Hormigas de Colombia. Universidad Nacional de Colombia, Bogotá.
https://doi.org/10.21676/9789587837650
Guerrero, R.J. 2021. Transfer of two South American ant species from Tapinoma Foerster 1850 to Forelius Emery 1888 (Hymenoptera: Formicidae: Dolichoderinae). Zootaxa, 4920(3):428–438.
https://doi.org/10.11646/zootaxa.4920.3.8
Hernández-Hernández, M.J., Cruz, J.A., and Castañeda-Posadas, C. 2020. Paleoclimatic and vegetation reconstruction of the Miocene Southern Mexico using fossil flowers. Journal of South American Earth of Sciences, 104(2020):102827.
https://doi.org/10.1016/j.jsames.2020.102827
Kusnezov, N. 1963. Zoogeografía de las hormigas en Sudamérica. Acta Zoológica Lilloana, 19:25–186.
Langenheim, J.H. 2003. Plant resins: chemistry, evolution, ecology and ethnobotany. Timber Press, Portland, Cambridge,UK.
https://doi.org/10.2307/4129644
Latreille, P.A. 1809. Genera crustaceorum et insectorum secundum ordinem naturalem in familias disposita, iconibus exemplisque plurimus explicata. Tomus 4. A. Koenig, Parisiis et Argentorati [= Paris and Strasbourg].
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines genera, species cum characteribus, differentris, synonymis, locis. Editos decima, reformata. Laurentii Salvii, Stockholm.
https://doi.org/10.5962/bhl.title.157601
Longino, J. 2007. A taxonomic review of the genus Azteca (Hymenoptera: Formicidae) in Costa Rica and a global revision of the aurita group. Zootaxa, 1491:1–63.
https://doi.org/10.11646/zootaxa.1491.1.1
Perrilliat, M.C., Vega, F.J., and Coutiño, M.A. 2010. Miocene mollusks from the Simojovel area in Chiapas, Southwestern Mexico. Journal of South American Earth Sciences, 30(2):111–119.
https://doi.org/10.1016/j.jsames.2010.04.005
Riquelme, F., Ruvalcaba-Sil, J.L., Alvarado-Ortega, J., Estrada-Ruiz, E., Galicia-Chávez, M., Porras-Múzquiz, H., Stojanof, V., Siddons, D.P., and Miller, L. 2014a. Amber from México: Coahuilite, Simojovelite and Bacalite. Cambridge/MRS Proceedings, 1618:169–180.
https://doi.org/10.1557/opl.2014.466
Riquelme, F., Hernández-Patricio, M., Martínez-Dávalos, A., Rodríguez-Villafuerte, M., Montejo-Cruz, M., Alvarado-Ortega, J., Ruvalcaba-Sil, J.L., and Zúñiga-Mijangos, L. 2014b. Two flat-backed polydesmidan millipedes from the Miocene Chiapas-amber Lagerstätte, Mexico. PLoS ONE, 9(8):e105877.
https://doi.org/10.1371/journal.pone.0105877
Riquelme, F., Montejo-Cruz, M., Luna-Castro, B., and Zúñiga-Mijangos, L. 2015. Fossil Jumping-bristletail from the Chiapas amber: Neomachilellus (Praeneomachilellus) ezetaelenensis sp. nov. (Microcoryphia: Meinertellidae). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 275(1):93–106.
https://doi.org/ 10.1127/njgpa/2015/0453
Shattuck, S.O. 1992. Generic revision of the ant subfamily Dolichoderinae (Hymenoptera: Formicidae). Sociobiology, 21(1):1–181.
Ward, P.S. 2005. A synoptic review of the ants of California (Hymenoptera: Formicidae). Zootaxa, 936:1–68.
https://doi.org/10.11646/zootaxa.936.1.1
Ward, P.S., Brady, S.G., Fisher, B.L., and Schultz, T.R. 2010. Phylogeny and biogeography of Dolichoderine ants: effects of data partitioning and relict taxa on historical inference. Systematic Biology, 59(3):342–362.
https://doi.org/10.1093/sysbio/syq012
