

 Pleistocene cave hyenas in the Iberian Peninsula: New insights from Los Aprendices cave (Moncayo, Zaragoza)
Pleistocene cave hyenas in the Iberian Peninsula: New insights from Los Aprendices cave (Moncayo, Zaragoza)
Article number: 20.1.11A
https://doi.org/10.26879/622
Copyright Society for Vertebrate Paleontology, March 2017
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 23 December 2015. Acceptance: 13 March 2017
{flike id=1786}
ABSTRACT
A new Pleistocene paleontological site, Los Aprendices, located in the northwestern part of the Iberian Peninsula in the area of the Moncayo (Zaragoza) is presented. The layer with fossil remains has been dated by amino acid racemization to 143.8 ± 38.9 ka (earliest Late Pleistocene or latest Middle Pleistocene). Five mammal species have been identified in the assemblage: Crocuta spelaea (Goldfuss, 1823) Capra pyrenaica (Schinz, 1838), Lagomorpha indet, Arvicolidae indet and Galemys pyrenaicus (Geoffroy, 1811). The remains of C. spelaea represent a mostly complete skeleton in anatomical semi-connection. The hyena specimen represents the most complete skeleton ever recovered in Iberia and one of the most complete remains in Europe. It has been compared anatomically and biometrically with both European cave hyenas and extant spotted hyenas. In addition, a taphonomic study has been carried out in order to understand the origin and preservation of these exceptional remains. The results suggest rapid burial with few scavenging modifications putatively produced by a medium sized carnivore. A review of the Pleistocene Iberian record of Crocuta spp. has been carried out, enabling us to establish one of the earliest records of C. spelaea in the recently discovered Los Aprendices cave, and also showing that the most extensive geographical distribution of this species occurred during the Late Pleistocene (MIS4-2).
Víctor Sauqué. Grupo Aragosaurus-IUCA, Dpto. Ciencias de la Tierra, Facultad de Ciencias, Universidad de Zaragoza, Calle Pedro Cerbuna 12, 50009, Zaragoza, Spain; vsauque@gmail.com
Laboratorio de Zooarqueología y Tafonomía de Zonas Áridas (LaZTA), IDACOR, CONICET/FFyH UNC. Av. H. Yrigoyen 174, 5000, Córdoba, Argentina
Raquel Rabal-Garcés. Grupo Aragosaurus-IUCA, Dpto. Ciencias de la Tierra, Facultad de Ciencias, Universidad de Zaragoza, Calle Pedro Cerbuna 12, 50009, Zaragoza, Spain; rrabalg@gmail.com
Joan Madurell-Malaperia. Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona, Edifici ICTA-ICP, c\de les columnes sn, 08193, Cerdanyola del Vallès, Barcelona, Spain; joan.madurell@icp.cat
Mario Gisbert. Centro de espeleología de Aragón (CEA); asismario@hotmail.com
Samuel Zamora. Instituto Geológico y Minero de España, Calle Manuel Lasala, 44, 9ºB, 50006, Zaragoza, Spain; s.zamora@igme.es
Trinidad de Torres. Laboratory of Biomolecular Stratigraphy, Madrid School of Mines; trinidad.torres@upm.es
José Eugenio Ortiz. Laboratory of Biomolecular Stratigraphy, Madrid School of Mines joseeugenio.ortiz@upm.es
Gloria Cuenca-Bescós. Grupo Aragosaurus-IUCA, Dpto. Ciencias de la Tierra, Facultad de Ciencias, Universidad de Zaragoza, Calle Pedro Cerbuna 12, 50009, Zaragoza, Spain; cuencag@unizar.es
Keywords: Pleistocene; Crocuta spelaea; taphonomy; distribution; Iberian Peninsula
Final citation: Sauqué, Víctor, Rabal-Garcés, Raquel, Madurell-Malaperia, Joan, Gisbert, Mario, Zamora, Samuel, de Torres, Trinidad, Ortiz, José Eugenio, and Cuenca-Bescós, Gloria. 2017. Pleistocene cave hyenas in the Iberian Peninsula: New insights from Los Aprendices cave (Moncayo, Zaragoza). Palaeontologia Electronica 20.1.11A: 1-38. https://doi.org/10.26879/622
palaeo-electronica.org/content/2017/1786-crocuta-spelaea-in-iberias
Copyright: © March 2017 Society of Vertebrate Paleontology. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0/
INTRODUCTION
The cave hyena (Crocuta spelaea) is a well-known and widespread taxon in the Late Pleistocene of Europe. The first mention of the cave hyena in the paleontological literature dates back to illustrations by Esper (1774) portraying the teeth of animals recovered from the Zoolithen Cave in Bavaria (Germany; Diedrich, 2008, 2011a). Several decades later, Goldfuss (1823) formally erected the new species Hyaena spelaea on the basis of an almost complete cranium recovered from Zoolithen Cave. This species was later transferred to the genus Crocuta by Soergel (1937). Some scholars consider the cave hyena a subspecies of the living African spotted hyena and refer to Crocuta crocuta intermedia (Bonifay, 1971) or Crocuta crocuta spelaea (see Kurtén, 1957; Werdelin and Solounias, 1991).
During the Late Pleistocene cave hyenas were the main non-anthropic accumulators of bone remains in caves (Fosse, 1997; Diedrich, 2014). As a consequence of its activity, hundreds of accumulations have been documented in European caves (Fosse, 1997; Fosse et al., 1998; Diedrich and Žák, 2006; Turner et al., 2008; Fourvel, 2012; Diedrich, 2014; Fourvel et al., 2014). In the Late Pleistocene of the Iberian Peninsula, the cave hyena was a common element in large mammal assemblages, being recorded from numerous sites (Arribas et al., 2010; Varela, 2011). Its last Iberian occurrence was in the south of the Peninsula, in Las Ventanas Cave (Granada, Spain) at the end of the Late Pleistocene (ca. 12.780 cal ka BP, Carrión et al., 2001).
Despite their ubiquitous presence in European Late Pleistocene assemblages, complete or even partial skeletons of Crocuta spelaea are very rare in the fossil record. The most complete specimens are known from Koněprusy Cave (Czech Republic; Diedrich and Žák, 2006) and from Les Oubliettes (Gargas, France; Cardoso, 1993). The lack of more complete skeletons of this species in the fossil record may be a consequence of their scavenger dietary habits, including cannibalism (Diedrich, 2005, 2011a, 2011b, 2011c, 2011d, 2012a, 2012b, 2014).
During a paleontological survey in 2012 in the Moncayo Massif, several bones of cave hyena were discovered in a cave called Los Aprendices (Zaragoza, Spain). The aim of this paper is to present the former cave and the excavated hyena remains. The studied specimens include the best-preserved skeleton of Crocuta spelaea in the Iberian Peninsula, and one of the most complete in Europe. This skeleton allows for a taphonomic study to understand the processes undergone by the carcass and to identify which animals might have scavenged on it. Furthermore, a biometric analysis of such a well-preserved specimen provides a good comparison with other specimens from Iberia and South-Western and Central Europe.
LOS APRENDICES CAVE: THE HISTORY OF ITS DISCOVERY AND ITS GEOGRAPHICAL AND GEOLOGICAL LOCATION
History of Its Discovery
 The site of Los Aprendices was discovered in 2012 by members of the University of Zaragoza and the CEA (Centro de Espeleología de Aragón; Speleological Centre of Aragón) in the course of a prospection survey in the caves of the Moncayo Massif (“Prospection of the karstic caves in the Moncayo area located within the municipal areas of Purujosa, Trasobares and Calcena Nº Exp 087/2011”). Los Aprendices Cave was already known in the region and while mapping it in 2005 members of the CEA unblocked a passage leading to a gallery with a detritus cone that probably corresponds to the former entrance to the cave. During the work associated with the unblocking of the gallery several large carnivore metapodials in anatomical correspondence were discovered (Figure 1.1). Preliminary study of the recovered remains at the University of Zaragoza suggested that they belong to a cave hyena. Taking that information into account, a systematic field survey was begun in the Los Aprendices Cave in order to extract other bones putatively corresponding to the same specimen.
The site of Los Aprendices was discovered in 2012 by members of the University of Zaragoza and the CEA (Centro de Espeleología de Aragón; Speleological Centre of Aragón) in the course of a prospection survey in the caves of the Moncayo Massif (“Prospection of the karstic caves in the Moncayo area located within the municipal areas of Purujosa, Trasobares and Calcena Nº Exp 087/2011”). Los Aprendices Cave was already known in the region and while mapping it in 2005 members of the CEA unblocked a passage leading to a gallery with a detritus cone that probably corresponds to the former entrance to the cave. During the work associated with the unblocking of the gallery several large carnivore metapodials in anatomical correspondence were discovered (Figure 1.1). Preliminary study of the recovered remains at the University of Zaragoza suggested that they belong to a cave hyena. Taking that information into account, a systematic field survey was begun in the Los Aprendices Cave in order to extract other bones putatively corresponding to the same specimen.
Description of the Cave and the Infill
 Los Aprendices Cave is situated in the Sierra del Moncayo, which is located in the central part of the Iberian Range, northern Spain (Figure 2.1). As a result of the altitude of the Sierra and its geographical location between the Duero and Ebro river basins, the area receives major hydrological input from Atlantic fronts. The entrance of the cave is at an altitude of 910 m in the western slopes of the Peñas del Cabo, among the foothills of the Moncayo, just a few kilometres to the south of the village of Calcena (Zaragoza, Spain) (Figure 2.2-3). The cave extends into the limestone-dolomite breccias and vacuolar dolomites of the Cortes de Tajuña Formation, which is Lower Jurassic in age. It consists of a large sloping gallery comprising a big hall in its initial section, which gradually diminishes in size, with a reduction particularly in the height of the ceiling (Figure 2.4). To reach the gallery where the fossil remains were found we have to pass through a narrow aisle (some 50 cm in height) that leads to another gallery at a lower level than the main hall (Figure 3.1-2). At the end of this gallery is the sediment cone that is probably associated with a former entrance to the cave (Gisbert and Pastor, 2009). This former entrance must have been the one used by the animals to reach the cone (Figure 3.5).
Los Aprendices Cave is situated in the Sierra del Moncayo, which is located in the central part of the Iberian Range, northern Spain (Figure 2.1). As a result of the altitude of the Sierra and its geographical location between the Duero and Ebro river basins, the area receives major hydrological input from Atlantic fronts. The entrance of the cave is at an altitude of 910 m in the western slopes of the Peñas del Cabo, among the foothills of the Moncayo, just a few kilometres to the south of the village of Calcena (Zaragoza, Spain) (Figure 2.2-3). The cave extends into the limestone-dolomite breccias and vacuolar dolomites of the Cortes de Tajuña Formation, which is Lower Jurassic in age. It consists of a large sloping gallery comprising a big hall in its initial section, which gradually diminishes in size, with a reduction particularly in the height of the ceiling (Figure 2.4). To reach the gallery where the fossil remains were found we have to pass through a narrow aisle (some 50 cm in height) that leads to another gallery at a lower level than the main hall (Figure 3.1-2). At the end of this gallery is the sediment cone that is probably associated with a former entrance to the cave (Gisbert and Pastor, 2009). This former entrance must have been the one used by the animals to reach the cone (Figure 3.5).
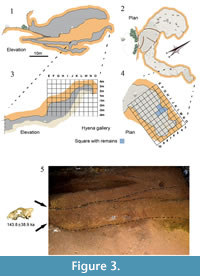 As mentioned above, the sediment cone blocks off a former entrance. It has a depth of 7 m and consists of a superposition of conglomerate levels. The level at which the cave hyena was found is composed of a clayey matrix and clasts with a maximum diameter of 5 cm and an average size of roughly 2 cm. Neither the adjacent nor the overlying levels have reported macrofaunal remains. The level with the cave hyena includes few clasts in the area where the skeleton occurred, but was a conglomerate supported by the clayey matrix (Figure 3.5).
As mentioned above, the sediment cone blocks off a former entrance. It has a depth of 7 m and consists of a superposition of conglomerate levels. The level at which the cave hyena was found is composed of a clayey matrix and clasts with a maximum diameter of 5 cm and an average size of roughly 2 cm. Neither the adjacent nor the overlying levels have reported macrofaunal remains. The level with the cave hyena includes few clasts in the area where the skeleton occurred, but was a conglomerate supported by the clayey matrix (Figure 3.5).
The large mammal association recovered from Los Aprendices Cave includes Capra pyrenaica and Crocuta spelaea. By screening and washing the sediment from the cave, the following microfaunal remains were also recovered: Lagomorpha indet., Arvicolidae indet., Galemys pyrenaicus and Serpentes indet.
To investigate the chronology of the site, two bone samples were initially sent for C14 dating at the Poznan Radiocarbon Laboratory (Poland). Unfortunately, there was no collagen in the bones, precluding a successful analysis. Subsequently, samples were sent to be analysed by amino acid racemization. For this purpose, we chose three molars of Capra pyrenaica, found in the same level as the hyena. The analysis was carried out at the Laboratory of Biomolecular Stratigraphy (Laboratorio de Estratigrafía Biomolecular; LEB) of the Mines Engineering School Polytechnic University of Madrid. The sample was prepared in accordance with the LEB protocol and analysed in a HPLC-1100 high-performance liquid chromatograph equipped with a fluorescence detector. The age obtained by this process was 143.8 ± 38.9 ka (see Appendix 1 with abundance and racemization relations of various amino acids). Given the associated fauna at the site, we regard it as more coherent for the sigma to be negative, with the site thus dating to the early stages of the Late Pleistocene.
The fossil remains studied in the present paper are housed in the Museo de Ciencias Naturales of the University of Zaragoza (with reference abbreviations from MPZ 2014/574 to MPZ 2014/718). The remains were recovered with the authorization and financial support of the Government of Aragon and the Moncayo Natural Park.
MATERIAL AND METHODS
Identifiable remains and bone fragments larger than 4 cm have been studied. For a general taxonomic identification the following references were followed: Pales and Lambert (1971) and Walker (1985). Measurements were taken with digital calipers (Mitutoyo Digimatic Caliper CD-800 CX, Japan) in accordance with the protocols of Hollister (1918), Bonifay (1971), von den Driesch (1976), Clot (1980), Cardoso (1993) and Blasco and Montes (1997). All measurements are given in millimetres. The Los Aprendices specimen has been compared to individuals of the extant species collected from different localities and stored at the Royal Museum for Central Africa, Belguim (RMCA), the Museu de Ciències Naturals de Barcelona, Spain (MZB); and Museu de Geologia de Barcelona, Spain (MGB), and to Late Pleistocene fossils from Gabasa 1 (Huesca, Northern Spain) stored at the Museo Arqueológico Provincial de Huesca, Spain. In addition, the morphometric data have been compared with collections from western Europe published by Bonifay (1971), Blasco and Montes (1997), Altuna and Mariezkurrena (2000), García (2003), Testu (2006), Fourvel (2012) and others.
In order to assess the skeletal representation in the assemblage from Los Aprendices, we referred to the number of remains (NR), the number of identified specimens (NISP), the minimum number of elements (MNE) and the minimum number of individuals (MNI), which were calculated in accordance with Lyman (1994). To calculate the MNI, the teeth were used, because these are the most common anatomical element, and the degree of eruption and dental wear were also taken into account. The age of the Crocuta spelaea specimen was determined in accordance with the revision by Fourvel (2012) of the methods of Mills (1982) and Brugal et al. (1997).
Various types of marks produced by carnivore teeth have been differentiated (pits, punctures, grooves, furrowing, crenulated edges and impact points), following the definitions by Haynes (1980, 1983), Binford (1981) and Sala (2012). The measurements were taken with electronic digital calipers. To identify the carnivore marks, these measurements were compared with data provided by Delaney-Rivera et al. (2009), Domínguez-Rodrigo and Piqueras (2003), Saladié et al. (2013) and Rabal-Garcés et al. (2012).
To ascertain whether the breakage of the bones took place in fresh bone, immediately post-mortem, or sometime after burial, as well as the possible causes of this breakage, we follow the criteria proposed by Villa and Mahieu (1991). This method takes into account the delineation (curved, intermediate or transverse), the angle (oblique, right angle or mixed) and the type of edge of the breaks shown by long bones greater than 4 cm in length (irregular or smooth). Further, the breakage index has also been taken into account. This refers to the size of the diaphysis preserved in relation to the total size of the bone in question, both in terms of its length and its circumference. The breakage indices that refer to the length of the diaphysis are L1 (preserved length < 25% of total length), L2 (preserved length between 25% and 50% of total length), L3 (preserved length between 50% and 75% of total length) and L4 (preserved length > 75% of total length). The breakage indices for the circumference are as follows: C1 (preserved circumference < 50% of total circumference), C2 (preserved circumference > 50% of total circumference) and C3 (the circumference is complete or almost complete, at least in some part).
Dentognathic and postcranial abbreviations: Bd, Breadth of the distal end; Bp, Breadth of proximal end; BPC, Proximal articular surface breadth; BM, body mass; c, lower canine; C, upper canine; d, distal; DMD, mesiodistal diameter; DPA, Olecranic maximum depth; DVL, vestibule-lingual diameter; GB, greatest breadth; GL, greatest length; H condyle md, height of mandibular condyle; Hdiast, height of ramus below the diastema; Hm1, height of ramus below m1; Hp4, height of ramus below p4; Hramus, mandibular ramus height; i2, second lower incisive; i3, third lower incisive; I1, first upper incisive; I2, second upper incisive; I3, third upper incisive; FL, femur length; HL, humerus length; l, left; L, length; Lm1, lower carnassial length; max, maximum; min, minimum; Mtc, metacarpus; Mtt, metatarsus; m1, first lower molar; n, sample size; p, proximal; p2, second lower premolar; p3, third lower premolar; p4, fourth lower premolar; P1, first upper premolar; P2, second upper premolar; P3, third upper premolar; P4, fourth upper premolar; r, right; SD, smallest breadth of the diaphysis; SKL, skull length; W, Width; W condyle md, width of the mandibular condyle; X: mean; σ, standard deviation; 1st Pha: first phalanx; 2nd Pha: second phalanx. Sites: Ap, Los Aprendices: L.K, Labeko Koba; S.I, Sima I; Bo, Bolinkoba; Ga, Gabasa; Fo, Fontainhas; Va, Valdegoba; C.B, Cueva del Búho; To, El Toll; Ol, Olopte B; C.O, Cova de l'Or; Mo, Mollet; Ata, Atapuerca; TD 8, Atapuerca Trinchera Dolina 8; LV, Lunel-Viel; Pe, Petralona; Mos, Mosbach; Ka, Kazanka; Context: A, archaeological; P, palaeontological. Chronology: P, Pleistocene; MP, Middle Pleistocene; ELP, early Late Pleistocene; LP, Late Pleistocene; EH, early Holocene; MPal, middle Palaeolithic; EUPal, early Upper Palaeolithic; UPal, Upper Palaeolithic; Ms, Mousterian; Chat, Chatelperronian; Per, Perigordian; Au, Aurignacian; Gr, Gravettian; So, Solutrean; Mg, Magdalenian; Az, Azilian.
RESULTS
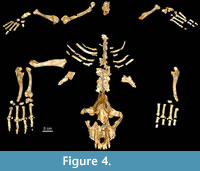
A total of 217 remains have been recovered from the site of Los Aprendices, 209 of which have been identified taxonomically. Eight fossils larger than 4 cm that have not been identified taxonomically have also been recovered. The MNE is 120. A total of 194 remains of Crocuta spelaea have been recovered, belonging to a single adult individual. The Los Aprendices individual fits into class 4 (6-10 years) according to Fourvel (2012) (Figure 4, Figure 5).The individual was found in anatomical semi-connection in the sediment cone within an area of scarcely two square metres (Figure 3.4-5, Table 1). Further, 15 remains belonging to at least two individuals of Capra pyrenaica have been recovered, one of the individuals being an adult and the other a neonate (see Table 2).
SYSTEMATIC PALAEONTOLOGY
Order CARNIVORA Bowdich, 1821
Family HYAENIDAE Gray, 1821
Genus CROCUTA Kaup, 1828
Type Species. Crocuta crocuta Erxleben, 1777
Crocuta spelaea Goldfuss, 1823
(Figure 4, Figure 5, Figure 6, Figure 7)
1823 Hyaena spelaea Goldfuss p.456-462, figs. 16, 17.
 1993 Crocuta crocuta spelaea Cardoso, p. 82-99, figs. 1, 2, 3, 4.
1993 Crocuta crocuta spelaea Cardoso, p. 82-99, figs. 1, 2, 3, 4.
v. 1997 Crocuta crocuta Blasco and Montes , p.12, 14, 17-21, pl, 2, 3, 4, 5, 6, 7.
1998 Crocuta crocuta spelaea Iñigo, Molero and Maldonado, p. 66, 68, fig. 1.
1999 Crocuta spelaea Baryshnikov, p. 167, 168, figs. 2, 3, 4.
2002 Crocuta crocuta Davis, p. 54, 55, fig. 21.
2005 Crocuta crocuta Arbizu, Álvarez-Lao, and Adan, p. 134, pl, 1..
2006 Crocuta crocuta spelaea Diedrich and Žák, p. 248, 249, figs. 5, 6.
2006 Crocuta crocuta spelaea Testu, p. 273, 274 , pl, 12, 13
2008 Crocuta crocuta spelaea Diedrich, p. 825, fig. 3.
2010 Crocuta crocuta Arsuaga, Baquedano, Pérez-González, Sala, García, Álvarez-Lao, Laplana, Huguet, Sevilla, Blain, Quam, Ruiz Zapata, Sala, Gil García, Uzquiano, and Pantoja, p. 63, fig. 9.
 2010 Crocuta spelaea Rosell, Blasco, Rivals, Cebrià, Morales, Rodríguez, Serrat, and Carbonell, fig. 4.
2010 Crocuta spelaea Rosell, Blasco, Rivals, Cebrià, Morales, Rodríguez, Serrat, and Carbonell, fig. 4.
2012a Crocuta crocuta spelaea Diedrich, p. 67-74, figs. 3, 4, 5, 6, 7, 8, 9,10.
2012b Crocuta crocuta spelaea Diedrich, p. 173-177, figs. 3, 4, 5.,6,7
v. 2012 Crocuta crocuta Martínez-Sánchez, López-García, Alcalá-Ortiz, Blain, and Rabal-Garcés, p. 551, fig. 2.
2014 Crocuta crocuta Maroto, fig. 4.
2015 Crocuta crocuta Álvarez-Lao, Ruiz-Zapata, Gil-García, Ballesteros, and Jiménez-Sánchez, fig. 5.
2015 Crocuta crocuta spelaea Fourvel, Fosse, Fernandez, and Antoine, p. 248, 249, 250, fig. 3.
Holotype.Hyaena spelaea Goldfuss, 1823. Incomplete cranium (Goldfuss-Museum Bonn No. M2609).
Type Locality. Zoolithen Cave, Geilenreuth, Wiensent Valley, Bavaria, Germany.
Localities. The species is found in hundreds of Pleistocene caves throughout Europe.
 Material. cranium (MPZ 2014/657), one mandible (MPZ 2014/593), one axis (MPZ 2014/589), five cervical vertebrae (MPZ 2014/592, MPZ 2014/605, MPZ 2014/607, MPZ 2014/668, MPZ 2014/675), seven thoracic vertebrae (MPZ 2014/641, MPZ 2014/656, MPZ 2014/661, MPZ 2014/667, MPZ 2014/670, MPZ 2014/674, MPZ 2014/690), four lumbar vertebrae (MPZ 2014/582, MPZ 2014/586, MPZ 2014/701, MPZ 2014/703), ten ribs (MPZ 2014/591, MPZ 2014/604, MPZ 2014/612, MPZ 2014/637, MPZ 2014/646, MPZ 2014/651, MPZ 2014/652, MPZ 2014/658, MPZ 2014/662, MPZ 2014/666), two scapulae (MPZ 2014/584, MPZ 2014/588), one humerus (MPZ 2014/672), two ulnae (MPZ 2014/610; MPZ 2014/676), one radius (MPZ 2014/611), one scapholunate (MPZ 2014/613), two capitate (MPZ 2014/630, MPZ 2014/659) one pisiform (MPZ 2014/614), eight metacarpi (MPZ 2014/575, MPZ 2014/581, MPZ 2014/615, MPZ 2014/616, MPZ 2014/617, MPZ 2014/618, MPZ 2014/684, MPZ 2014/685), one femur (MPZ 2014/587), two tibiae (MPZ 2014/671, MPZ 2014/681), one talus (MPZ 2014/682), two calcanei (MPZ 2014/632, MPZ 2014/677), one unciform (MPZ 2014/678), two cuboids (MPZ 2014/ 623, MPZ 2014/ 629), two scaphoids (MPZ 2014/623, MPZ 2014/629), two cuneiforms (MPZ 2014/62, MPZ 2014/628), eight metatarsi (MPZ 2014/633, MPZ 2014/634, MPZ 2014/635, MPZ 2014/636, MPZ 2014/642, MPZ 2014/643, MPZ 2014/644, MPZ 2014/645) and twenty-four phalanxes (MPZ 2014/575, MPZ 2014/576, MPZ 2014/577, MPZ 2014/578, MPZ 2014/579, MPZ 2014/580, MPZ 2014/583, MPZ 2014/590, MPZ 2014/594, MPZ 2014/599, MPZ 2014/600, MPZ 2014/601, MPZ 2014/602, MPZ 2014/603, MPZ 2014/620, MPZ 2014/621, MPZ 2014/622, MPZ 2014/626, MPZ 2014/627, MPZ 2014/631, MPZ 2014/639, MPZ 2014/654, MPZ 2014/655, MPZ 2014/680).
Material. cranium (MPZ 2014/657), one mandible (MPZ 2014/593), one axis (MPZ 2014/589), five cervical vertebrae (MPZ 2014/592, MPZ 2014/605, MPZ 2014/607, MPZ 2014/668, MPZ 2014/675), seven thoracic vertebrae (MPZ 2014/641, MPZ 2014/656, MPZ 2014/661, MPZ 2014/667, MPZ 2014/670, MPZ 2014/674, MPZ 2014/690), four lumbar vertebrae (MPZ 2014/582, MPZ 2014/586, MPZ 2014/701, MPZ 2014/703), ten ribs (MPZ 2014/591, MPZ 2014/604, MPZ 2014/612, MPZ 2014/637, MPZ 2014/646, MPZ 2014/651, MPZ 2014/652, MPZ 2014/658, MPZ 2014/662, MPZ 2014/666), two scapulae (MPZ 2014/584, MPZ 2014/588), one humerus (MPZ 2014/672), two ulnae (MPZ 2014/610; MPZ 2014/676), one radius (MPZ 2014/611), one scapholunate (MPZ 2014/613), two capitate (MPZ 2014/630, MPZ 2014/659) one pisiform (MPZ 2014/614), eight metacarpi (MPZ 2014/575, MPZ 2014/581, MPZ 2014/615, MPZ 2014/616, MPZ 2014/617, MPZ 2014/618, MPZ 2014/684, MPZ 2014/685), one femur (MPZ 2014/587), two tibiae (MPZ 2014/671, MPZ 2014/681), one talus (MPZ 2014/682), two calcanei (MPZ 2014/632, MPZ 2014/677), one unciform (MPZ 2014/678), two cuboids (MPZ 2014/ 623, MPZ 2014/ 629), two scaphoids (MPZ 2014/623, MPZ 2014/629), two cuneiforms (MPZ 2014/62, MPZ 2014/628), eight metatarsi (MPZ 2014/633, MPZ 2014/634, MPZ 2014/635, MPZ 2014/636, MPZ 2014/642, MPZ 2014/643, MPZ 2014/644, MPZ 2014/645) and twenty-four phalanxes (MPZ 2014/575, MPZ 2014/576, MPZ 2014/577, MPZ 2014/578, MPZ 2014/579, MPZ 2014/580, MPZ 2014/583, MPZ 2014/590, MPZ 2014/594, MPZ 2014/599, MPZ 2014/600, MPZ 2014/601, MPZ 2014/602, MPZ 2014/603, MPZ 2014/620, MPZ 2014/621, MPZ 2014/622, MPZ 2014/626, MPZ 2014/627, MPZ 2014/631, MPZ 2014/639, MPZ 2014/654, MPZ 2014/655, MPZ 2014/680).
The cave hyena is the predominant taxon in Los Aprendices. As the elements were found in anatomical position and, presumably because there is no duplication of elements, they have been assigned to a single individual. We here describe the cranium, mandible, teeth, because these are elements of great taxonomic importance.
Description
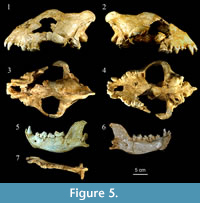
Cranium. The cranium (MPZ 2014/657) is almost complete, lacking a fragment of the right zygomatic arch and part of the nasals including a fragment of the right side. The sagittal crest is slightly deformed, and the left side of the neurocranium is flattened (Figure 5). The diastema between the canine and P1 is reduced, and P1 and P2 are in contact (Figure 5.4). In extant Crocuta crocuta, P1-P2 are separated by a distance of several mm. The infraorbital foramen is located above the point of contact between P2 and P3. The dental series displays the moderate curvature typical of C. spelaea (García, 2003). The palate is wide at the P4, and even though it is broken a pronounced concavity characteristic of the genus Crocuta can be observed (Morales et al., 1987). The cranium has a wide snout with a slight post-canine constriction. The rostral region (splanchnocranium) is short and wide, conferring upon it a “U”-shape in dorsal view, whereas in Hyaena it is “V”-shaped (García, 2003). One of the most notable characteristics of the cranium is the absence of M1. This element is generally lacking in the genus Crocuta but is present in Hyaena, in which M1 has three cuspids (Kurtén, 1957; Werdelin and Solounias, 1991; García, 2003).
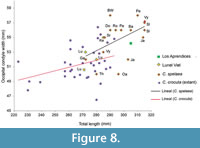
The cranium has a total length of 305 mm, which lies clearly outside the range of extant Crocuta crocuta (see Figure 5, 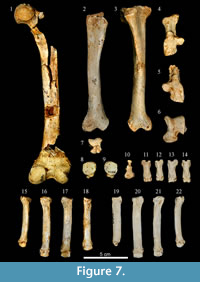 Figure 8; Appendix 2). However, it matches the size documented for cave hyenas and is larger than the C. c. intermedia (Serres et al., 1828) of Lunel Viel (Bonifay, 1971). In general terms the measurements of the skull of Los Aprendices are similar but lightly larger than those of Crocuta crocuta praespelaea from Mosbach, Kazanka and Petralona (Kurtén, 1962; Kurtén and Poulianos, 1981; Schütt, 1971; Baryshnikov, 2014). The specimen from Los Aprendices is similar in size to cave hyenas from central Europe at the sites of Badel Cave, the Perick Caves, and Rösenbecker Cave (Diedrich, 2011a), as well as at the French site of Jaurens (Ballesio, 1979). In comparison with other crania from the Iberian Peninsula, the specimen from Los Aprendices is clearly smaller as compared with the specimen from Fontainhas (L=317 mm) in Portugal (Cardoso, 1993), and slightly larger than the cranium from Gabasa 1 (L=294mm) (Blasco and Montes, 1997). Furthermore, it is larger than the latest Early Pleistocene cranium from Gran Dolina TD4W (L=265mm), which is ascribed to C. crocuta (García and Arsuaga, 1999).
Figure 8; Appendix 2). However, it matches the size documented for cave hyenas and is larger than the C. c. intermedia (Serres et al., 1828) of Lunel Viel (Bonifay, 1971). In general terms the measurements of the skull of Los Aprendices are similar but lightly larger than those of Crocuta crocuta praespelaea from Mosbach, Kazanka and Petralona (Kurtén, 1962; Kurtén and Poulianos, 1981; Schütt, 1971; Baryshnikov, 2014). The specimen from Los Aprendices is similar in size to cave hyenas from central Europe at the sites of Badel Cave, the Perick Caves, and Rösenbecker Cave (Diedrich, 2011a), as well as at the French site of Jaurens (Ballesio, 1979). In comparison with other crania from the Iberian Peninsula, the specimen from Los Aprendices is clearly smaller as compared with the specimen from Fontainhas (L=317 mm) in Portugal (Cardoso, 1993), and slightly larger than the cranium from Gabasa 1 (L=294mm) (Blasco and Montes, 1997). Furthermore, it is larger than the latest Early Pleistocene cranium from Gran Dolina TD4W (L=265mm), which is ascribed to C. crocuta (García and Arsuaga, 1999).
Upper dentition. C1. The length and width of the upper canines from Los Aprendices are within the range of variation of extant Crocuta crocuta but are clearly larger than average (Appendix 3).
P1. P1 is a small simple tooth, the crown of which forms a low cone crossed by a ridge that transversely divides it into two.
 P2. Mesial and distal accessory cusps are present and clearly developed. The outline of the tooth is rectangular, with the distal part buccolingually wider than the mesial part. The length and width of P2 from Los Aprendices fall within the range of extant Crocuta crocuta but close to the maximum values. The P2 from Los Aprendices is clearly larger than the P2 from the cranium of C. crocuta from Gran Dolina TD4W. The P2 from Los Aprendices is larger than C. crocuta spp. from Petralona (Baryshnikov and Tsoukala, 2010) and the C. c praespelaea from Kazanka and Mosbach (Baryshnikov, 2014), which according to Baryshnikov (2014) has the largest P2 of the spelaea forms. It is remarkable that the Los Aprendices P2 is the largest premolar recovered in the Iberian Peninsula of fossil record of Crocuta genus.
P2. Mesial and distal accessory cusps are present and clearly developed. The outline of the tooth is rectangular, with the distal part buccolingually wider than the mesial part. The length and width of P2 from Los Aprendices fall within the range of extant Crocuta crocuta but close to the maximum values. The P2 from Los Aprendices is clearly larger than the P2 from the cranium of C. crocuta from Gran Dolina TD4W. The P2 from Los Aprendices is larger than C. crocuta spp. from Petralona (Baryshnikov and Tsoukala, 2010) and the C. c praespelaea from Kazanka and Mosbach (Baryshnikov, 2014), which according to Baryshnikov (2014) has the largest P2 of the spelaea forms. It is remarkable that the Los Aprendices P2 is the largest premolar recovered in the Iberian Peninsula of fossil record of Crocuta genus.
P3. P3 is characterized by the great development of the protocone, which is surrounded by a well-developed cingulum and lacks accessory cusps. This tooth is an important part of the process of bone crushing (Schütt, 1971), and therefore its structure can be used to evaluate the degree of adaptation the dentition to feeding on carrion (Baryshnikov, 2014). This tooth presents an extremely high crown, which is also seen in other populations of Crocuta spelaea. It indicates a Crocuta spelaea specialization for breaking bones related to scavenging. The length and width values of P3 from Los Aprendices are within the range of variation of the Iberian Pleistocene cave hyena and exceed the measurements of extant C. crocuta (Appendix 3).
 P4. The anterior face of the protocone is approximately level with the anterior face of the parastyle, which is typical of the genus Crocuta (Wederlin and Solounias, 1991) Its greatest length and width markedly exceed those of extant C. crocuta and C. c. intermedia (Cardoso, 1993), and it is also larger than the C. crocuta from the Middle Pleistocene of Gran Dolina TD4W (García, 2003). The Los Aprendices specimen is of similar length and width as C. c. praespelaea from Kazanka (Baryshnikov, 2014) and is slighty larger than C. c. praespelaea from Mosbach (Bayshnikov, 2014) and C. crocuta spp. from Petralona (Baryshnikov and Tsoukala, 2010). Therefore, its values are similar to those of C. spelaea from the Iberian Peninsula, such as Gabasa 1 (Blasco and Montes, 1997), Fontainhas (Cardoso, 1993), Sima I, Kobaedaerra (Castaños, 1987), Labeko Koba (Altuna and Mariezkurrena, 2000), El Toll, Olopte B and Cova de l’Or (J.M.-M personal commun., 2015.) (Appendix 3). The extreme metrical variability of dental remains (Figure 9, Appendix 3) most likely coincides with ecomorphotypic variation, so it is not recommended to use cave hyenas for biochronological purposes (e.g., Kurtén, 1957; Kurtén and Poulianos 1977; Klein and Scott 1989; Baryshnikov 1999, Fourvel et al., 2015).
P4. The anterior face of the protocone is approximately level with the anterior face of the parastyle, which is typical of the genus Crocuta (Wederlin and Solounias, 1991) Its greatest length and width markedly exceed those of extant C. crocuta and C. c. intermedia (Cardoso, 1993), and it is also larger than the C. crocuta from the Middle Pleistocene of Gran Dolina TD4W (García, 2003). The Los Aprendices specimen is of similar length and width as C. c. praespelaea from Kazanka (Baryshnikov, 2014) and is slighty larger than C. c. praespelaea from Mosbach (Bayshnikov, 2014) and C. crocuta spp. from Petralona (Baryshnikov and Tsoukala, 2010). Therefore, its values are similar to those of C. spelaea from the Iberian Peninsula, such as Gabasa 1 (Blasco and Montes, 1997), Fontainhas (Cardoso, 1993), Sima I, Kobaedaerra (Castaños, 1987), Labeko Koba (Altuna and Mariezkurrena, 2000), El Toll, Olopte B and Cova de l’Or (J.M.-M personal commun., 2015.) (Appendix 3). The extreme metrical variability of dental remains (Figure 9, Appendix 3) most likely coincides with ecomorphotypic variation, so it is not recommended to use cave hyenas for biochronological purposes (e.g., Kurtén, 1957; Kurtén and Poulianos 1977; Klein and Scott 1989; Baryshnikov 1999, Fourvel et al., 2015).
Mandible. The left hemimandible from Los Aprendices is well preserved and practically complete. The corpus preserves the i1, i2, c1, p2, p3, p4 and m1, all showing wear. The ramus has a complete and well preserved coronoid process, articular and angular apophyses. The corpus presents a basal edge that in lateral view is strongly convex at the level of m1 (Figure 5.5-6). This feature is characteristic of Crocuta (Bonifay, 1971; Testu, 2006). The hemimandible is mesiolingually curved. In buccal view, the ramus in its distal part shows a deep, subtriangular masseteric fossa that extends from the protoconid of m1 to the angular process and the condyle. The single mental foramen is placed at the level of p2. The arrangement of the first two premolars (p2 and p3) is not overlapping, and p4 and m1 overlap as in Crocuta (Testu, 2006). The disposition of the premolars in the dental series is clearly curved (Figure 5.7); this curvature is caused by the increase in size and is a distinctive feature of C. spelaea (García, 2003).
Los Aprendices hemimandible is larger mesiodistally as compared with extant Crocuta crocuta (Appendix 4). It is also far larger than fossil hyenas from the Middle Pleistocene, such as Atapuerca TD 8 (García, 2003) and Lunel Viel (Bonifay, 1971). However, the distance c1-m1 is smaller than in C. c. praespelaea from Kazanka (Baryshnikov, 2014). On the other hand, the mandible fits perfectly within the range of C. spelaea and is one of the largest specimens of the Iberian Peninsula (Appendix 4). The biometric analysis of the hemimandible further reinforces the inclusion of the studied specimen in the hypodigm of C. spelaea.
Lower dentition. c1. The c1 is within the range of variation shown by the population of Pleistocene cave hyenas and larger than extant Crocuta crocuta (Appendix 5).
p2. The main cusp, the protoconid, is buccolingually wide and mesially surrounded by a well-developed cingulum. The distal accessory cusp is well-developed and separated from the protoconid by a transverse groove. The p2 of Los Aprendices is clearly larger than the average for extant C. crocuta , C. c. intermedia and C. c. praespelaea from Petralona (Appendix 5). On the other hand, its values fit perfectly with the C. spelaea from Labeko Koba (Altuna and Mariezkurrena, 2000) and are slightly smaller than the individual of C. c. praespelaea from Kazanka (Baryshnikov, 2014). It is also slightly larger than the population of C. spelaea form Gabasa (Blasco and Montes, 1997).
p3. The protoconid has a slightly distal tilt, and shows a marked cingulum on the mesial and distal edges. No accessory cusps are developed. The p3 of Los Aprendices is the largest from the Iberian fossil record and definitely larger than extant Crocuta crocuta (Appendix 5).
p4. p4 displays well-developed accessory cuspid with the distal one being mesiodistally longer than the mesial one. As in p3, the protoconid displays a slight distal tilt. The p4 fits perfectly within the range of Late Pleistocene cave hyenas, being larger than extant Crocuta crocuta and C. c. intermedia, which we classified as a C. crocuta from the Middle Pleistocene. Furthermore, the measurements from Los Aprendices are similar to the C. c. praespelaea (Appendix 5).
m1. The paraconid is clearly mesiodistally larger than the protoconid, a taxonomic character only seen in Crocuta among Middle to Late Pleistocene bone-cracking hyaenas (Werdelin and Solounias, 1991). The m1 presents a well-developed cingulum on the lingual side that extends up to the anterior edge. Distally, a mesiodistally short talonid is evident, which is a character typical of the genus Crocuta. Finally, the distolingual part of the protoconid displays a small but well-marked metaconid. Several scholars consider the presence of well-developed metaconid in the m1 as an ancestral character in the genus Crocuta, barely present in the last European spotted hyaenas. However, the extreme variability in the development and presence of m1 metaconid in extant and extinct representatives of the genus Crocuta prevent its use for taxonomic proposes (Werdelin and Solounias, 1991; García, 2003).
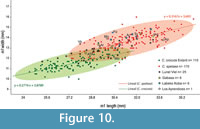 The length and width of m1 from Los Aprendices are within the range of variation of the Iberian Pleistocene cave hyena and exceed the measurements of extant Crocuta crocuta (Appendix 5). In Figure 10, it can be observed that there are two distinct populations: one for the C. spelaea and another population made up of extant C. crocuta and the specimens of C. crocuta from the Middle Pleistocene, such as C. c. intermedia (Bonifay, 1971) C. crocuta from Gran Dolina (García, 2003).
The length and width of m1 from Los Aprendices are within the range of variation of the Iberian Pleistocene cave hyena and exceed the measurements of extant Crocuta crocuta (Appendix 5). In Figure 10, it can be observed that there are two distinct populations: one for the C. spelaea and another population made up of extant C. crocuta and the specimens of C. crocuta from the Middle Pleistocene, such as C. c. intermedia (Bonifay, 1971) C. crocuta from Gran Dolina (García, 2003).
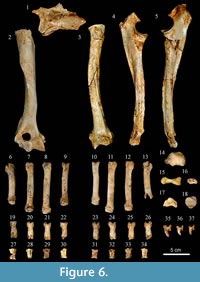
Forelimbs. The forelimb bones (Figure 6; Appendix 6) include both incomplete scapulae, one nearly complete humerus, both complete ulnas, one complete radius, some carpals such as the scapholunar, pisiform, cuneiform and unciform, and all eight metacarpals except the Mtc I.
Humerus. The proximal epiphysis of the humerus is broken and the head is missing. In cranial view, the diaphysis is straight laterally flattened, with caudal bending in the proximal third. The deltoid crest is very prominent and connected with the humeral crest. The distal trochlea is wide and the condyle is broken. Above this there is a well-developed oval supratrochlear foramen. In the distal part of the diaphysis, there is a deep and well-developed olecranon fossa. As a consequence of the broken or absent epiphysis, is not possible to metrically compare the humerus with other known samples.
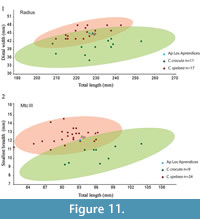 Radius. The proximal epiphysis has an oval outline. The bicipital tuberosity is prominent, oval and is placed on the proximolateral part of the diaphysis. The diaphysis has a semicircular cross section and is anteroposteriorly flattened. It shows strong torsion in the distal part of the bone. The distal epiphysis is mediolaterally wider than the diaphysis and has a very marked styloid process that is pointed and projected distally. The radius length of the Los Aprendices specimen (227 mm) is within the range of variation of Iberian Pleistocene cave hyenas. Its length also lies within the range of extant Crocuta crocuta, but is slightly below average (Appendix 6, Figure 11.1). Despite being shorter than that of extant C. crocuta, it is moderately mediolaterally wider, which gives a characteristic robust aspect to C. spelaea (Kurtén, 1957).
Radius. The proximal epiphysis has an oval outline. The bicipital tuberosity is prominent, oval and is placed on the proximolateral part of the diaphysis. The diaphysis has a semicircular cross section and is anteroposteriorly flattened. It shows strong torsion in the distal part of the bone. The distal epiphysis is mediolaterally wider than the diaphysis and has a very marked styloid process that is pointed and projected distally. The radius length of the Los Aprendices specimen (227 mm) is within the range of variation of Iberian Pleistocene cave hyenas. Its length also lies within the range of extant Crocuta crocuta, but is slightly below average (Appendix 6, Figure 11.1). Despite being shorter than that of extant C. crocuta, it is moderately mediolaterally wider, which gives a characteristic robust aspect to C. spelaea (Kurtén, 1957).
Ulna. The proximal part of the olecranon process is not preserved and the diaphysis displays more curvature and robusticity as compared with extant Crocuta crocuta. The triangular area below the lesser semilunar notch is deep and connected to the depressed area for the interosseous ligament, which can be seen on the lateral part of the diaphysis. The styloid process is robust with a rounded articular circumference that projects posteriorly.
Metacarpals. The metacarpals are clearly shorter than the average in extant Crocuta crocuta and are mediolaterally robust (Appendix 6, Figure 11.2). The length and width of MtcpIII falls outside the range of variation of extant C. crocuta, being closer to the minimum values for proximodistal length and close to the maximum values for mediolateral breadth (Appendix 6).
Hindlimbs. Of the hindlimbs (Figure 7; Appendix 6), only one broken femur and both tibiae are more or less complete. One astragalus and both calcanei complete some tarsals have also been recovered: one navicular, both cuboids, one first cuneiform and all eight metatarsals except Mtt I.
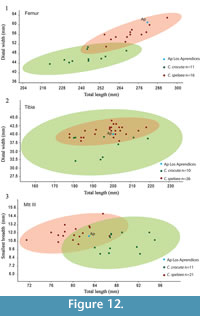 Femur. The diaphysis is straight in the proximal part and slightly anterioposteriorly curved in the distal part. It has a circular cross-section, which becomes oval in the distal part. The proximal epiphysis is broken and the great trochanter is missing. The neck is complete and connects the diaphysis with the head, which is spherical shape and presents an oval fovea capitis. The distal epiphysis of femur consisted of two similar condyles. The lateral one is partially broken. The condyles are similar, situated at the same level and separated by a intercondyloid fossa. The femur length of the Los Aprendices specimen is within the range of variation of extant Crocuta crocuta but very close to the maximum values and clearly greater than the average (Appendix 6). The distal epiphysis mediolateral width is outside the range of variation of extant C. crocuta (Appendix 6). The femur from Los Aprendices is similar in size to the femur from Labeko Koba (Altuna and Mariezkurrena, 2000) and larger than the femur from TD5, which belongs to C. crocuta (García, 2003). In Figure 12.1, it can be observed that there are two clusters that differentiate C. spelaea from extant C. crocuta.
Femur. The diaphysis is straight in the proximal part and slightly anterioposteriorly curved in the distal part. It has a circular cross-section, which becomes oval in the distal part. The proximal epiphysis is broken and the great trochanter is missing. The neck is complete and connects the diaphysis with the head, which is spherical shape and presents an oval fovea capitis. The distal epiphysis of femur consisted of two similar condyles. The lateral one is partially broken. The condyles are similar, situated at the same level and separated by a intercondyloid fossa. The femur length of the Los Aprendices specimen is within the range of variation of extant Crocuta crocuta but very close to the maximum values and clearly greater than the average (Appendix 6). The distal epiphysis mediolateral width is outside the range of variation of extant C. crocuta (Appendix 6). The femur from Los Aprendices is similar in size to the femur from Labeko Koba (Altuna and Mariezkurrena, 2000) and larger than the femur from TD5, which belongs to C. crocuta (García, 2003). In Figure 12.1, it can be observed that there are two clusters that differentiate C. spelaea from extant C. crocuta.
Tibia. The tibia is short and robust. It is slightly convex on the proximomedial side. The proximal epiphysis has a triangular cross-section, and this morphology continues throughout the diaphysis except for the distal portion, where it is oval. The tibial crest is wide and well-developed. The distal epiphysis is subrectangular, with a well differentiated, laterally projecting medial rounded malleolus. The length and width of tibia from Los Aprendices are within the range of variation of the cave hyena and extant Crocuta crocuta (Appendix 6, Figure 12.2).
Metatarsals. The metatarsals are shorter than those of extant Crocuta crocuta and more robust (Appendix 6, Figure 12.3). As occurs with Mtc III, the length and width of Mtt III is outside the range of variation of extant C. crocuta, but close to the length minimum and wider than the maximum value (Appendix 6, Figure 12.3).
TAPHONOMY
The recovered specimen of Crocuta spelaea displays several gnaw marks and fractures putatively corresponding to carnivore activity and for this reason a detailed taphonomic study was carried out.
Breakage
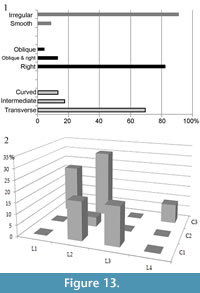 Breakage of the long bones was analysed in accordance with the protocol of Villa and Mahieu (1991). The delineation of the fractures of the bones tends to be transverse (70%). The predominant angles are right angles (80%), and the edges are mainly irregular (85%) (Figure 13.1). With regard to the length and section of the preserved diaphysis there is a predominance (30%) of bones with a length greater than a quarter but less than half of the total length of the bone (L2), and a complete diaphysis (C3). Second in frequency (21.8%) are bones with a length less than a quarter of the total and a complete diaphysis section (C3). It is noteworthy that only 34% of the bones have a diaphysis section of less than a third of the total (C1). Likewise, bones with a length of less than a quarter of the total only represent 21.8% of the total (Figure 13.2). The presence of a high number of bones with transverse delineation, right angles and irregular edges is related to fracturing when the bone is dry, and this pattern indicates that the bone has been broken due to sedimentary pressure.
Breakage of the long bones was analysed in accordance with the protocol of Villa and Mahieu (1991). The delineation of the fractures of the bones tends to be transverse (70%). The predominant angles are right angles (80%), and the edges are mainly irregular (85%) (Figure 13.1). With regard to the length and section of the preserved diaphysis there is a predominance (30%) of bones with a length greater than a quarter but less than half of the total length of the bone (L2), and a complete diaphysis (C3). Second in frequency (21.8%) are bones with a length less than a quarter of the total and a complete diaphysis section (C3). It is noteworthy that only 34% of the bones have a diaphysis section of less than a third of the total (C1). Likewise, bones with a length of less than a quarter of the total only represent 21.8% of the total (Figure 13.2). The presence of a high number of bones with transverse delineation, right angles and irregular edges is related to fracturing when the bone is dry, and this pattern indicates that the bone has been broken due to sedimentary pressure.
Gnaw Marks
The skeleton of the hyena from Los Aprendices displays modifications produced by carnivores, showing evidence of scavenge. Twenty bones with bite marks have been found, which - with a NISP of 191 - yields of percentage of 10.5%. The part of the skeleton with the greatest modification is the zeugopod-stylopod region, with 25% of the remains altered. The axial skeleton also presents a considerable degree of alteration, with 11.5% of the remains altered, while the autopods show a much lower level of alteration, amounting to just 4.2%. 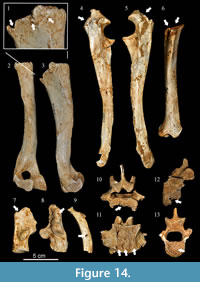 The low modification of the autopodium could be related to low bone consumption intensity, which also is supported by the fact that no ingested bone flakes have been recovered. (Table 3, Figure 14).
The low modification of the autopodium could be related to low bone consumption intensity, which also is supported by the fact that no ingested bone flakes have been recovered. (Table 3, Figure 14).
Table 4 presents the data for the different types of bite marks documented and their distribution among the different bone elements. The main conclusion from the analysis is that the most abundant type of mark on the hyena bones is “furrowing”, which appears on 15 remains, followed by “punctures” and “pits”, which in total appear on 12 remains (Figure 14). In terms of bone consumption sequence, the skeleton of Los Aprendices shows low bone consumption.
Measurements have been taken on the tooth marks produced by carnivores (“pits”, “punctures” and grooves). All the marks measured were found on spongy bone or fine cortical bone. The mean width of the depressions (“pits” and “punctures”) is 2.91 mm, and the mean length is 3.77 mm. The grooves have a mean width of 1.60 mm and a mean length of 5.70 mm (Table 5).
DISCUSSION
The Iberian Record of Crocuta
The earliest European occurrence of Crocuta crocuta is from the latest Early Pleistocene layer TD4-5 of Gran Dolina in the Sierra de Atapuerca (Spain) (ca. 0.9 Ma; García and Arsuaga, 1999). The presence of the taxon is almost continuous in the TD sequence. The sequence from TD3-4-TD8 is dated to between 473 ±130 and 900±150 ka (Moreno et al., 2015). A closer age, but just after the Brunhes-Matuyama boundary is shown by the record from Casal Selce in Italy (Sardella and Petrucci, 2012). The arrival of C. crocuta and the extinction of Pachycrocuta brevirostris, last recorded in layer EVT7 of Vallaparadís Estació (ca. 0.85 Ma; Madurell-Malapeira et al., 2010), mark the beginning of the Galerian Mammal Age in Europe. This faunal turnover has been called “the Crocuta crocuta event” (Martínez-Navarro, 2010).
During the Middle Pleistocene, Crocuta crocuta was the most common hyaenid recorded in the Iberian Peninsula. According to the paleontological literature fossils of C. c. intermedia have been recovered from a variety of sites in the Iberian Peninsula (Figure 4, Table 1), such as Atapuerca TD4 (Aguirre, 1987), the Cueva del Congosto (Carrión et al., 2006; Arribas et al., 2010), the Cuevas de Pedraza (Carrión et al., 2006; Arribas et al., 2010), Pinilla del Valle (Arribas et al., 2010), Villacastín (Carrión et al., 2006) and Cúllar (Carrión et al., 2006). In fact, the subspecies C. c. intermedia has been subject to broad criticism, as the specimens of C.c. intermedia and modern-day C. crocuta present a similar morphology and cannot be biometrically distinguished (Bonifay, 1971; García, 2003; Arribas et al., 2010). This is due to the fact that their metric parameters lie within the range of the present-day C. crocuta population (Kurtén, 1957; García, 2003). For some authors the hyaenid from Atapuerca TD4 in fact belongs to C. crocuta (García and Arsuaga, 1999; García, 2003) as does the one found at Pinilla del Valle (Arsuaga et al ., 2012).
Crocuta spelaea is first recorded in Western Europe around 300 ka (Baryshnikov and Tsoukala, 2010). It is not until the Middle-Late Pleistocene boundary that the larger cave hyena from appears (Arribas et al., 2010). C. spelaea is craniodentally larger than C. crocuta and also displays a relative shortening of the distal limb segments, especially the radius, tibia, metacarpals and metatarsals (Kurtén, 1957).
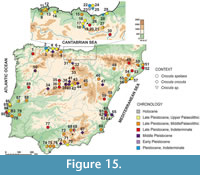 The cave hyena enjoyed notable success, becoming a ubiquitous taxon in European faunal associations of the Late Pleistocene from the Urals to the Iberian Peninsula (Figure 15, Appendix 7) (Arribas et al., 2010; Carrión et al., 2006, Stuart and Lister, 2014). Some authors hold that there are anatomical differences that are not sufficient for making a taxonomic distinction between individuals from the Eurasian Pleistocene and those from Africa (Turner et al., 2008). Results of studies of ancient DNA of these fossil remains suggest that the populations of European spotted hyena are a subset of the African population and not a distinct species (Rohland et al., 2005). Other authors suggest that there are still discrepancies between molecular and paleontological data. These authors suggest that the evolutionary timescale of spotted hyenas is not fully resolved, so further analyses are needed in order to reconcile the molecular and morphological data (Sheng et al., 2014).In the Iberian Peninsula there are numerous sites associated with the accumulating activity of cave hyenas, sites such as the Cueva del Búho (Iñigo et al., 1998), Torrejones (Arribas, 1997; Carrión et al., 2006) Gabasa 1 (Blasco and Montes, 1997), Las Ventanas (Carrión et al., 2001), Nerja (Arribas et al., 2004), Zafarraya (Geraads, 1995 Arribas et al., 2010) and Gorham (Carrión et al., 2008). The presence of Crocuta spelaea extended throughout the Late Pleistocene, lasting almost to the LGM (Last Glacial Maximum) as also the case in northern Eurasia (Stuart and Lister, 2014). However in Cueva de las Ventanas there is an indirect record (coprolites) that is dated to ca. 12 780 cal yr BP (Carrión et al., 2001), which is one of the latest records of this taxon in Europe. The reason for the disappearance of cave hyenas from Europe may be related to climatic factors or to other factors, such as reduced availability of prey or the ecological impact of hominids on the physical environment (Varela et al., 2010; Varela, 2011; Stuart and Lister, 2014).
The cave hyena enjoyed notable success, becoming a ubiquitous taxon in European faunal associations of the Late Pleistocene from the Urals to the Iberian Peninsula (Figure 15, Appendix 7) (Arribas et al., 2010; Carrión et al., 2006, Stuart and Lister, 2014). Some authors hold that there are anatomical differences that are not sufficient for making a taxonomic distinction between individuals from the Eurasian Pleistocene and those from Africa (Turner et al., 2008). Results of studies of ancient DNA of these fossil remains suggest that the populations of European spotted hyena are a subset of the African population and not a distinct species (Rohland et al., 2005). Other authors suggest that there are still discrepancies between molecular and paleontological data. These authors suggest that the evolutionary timescale of spotted hyenas is not fully resolved, so further analyses are needed in order to reconcile the molecular and morphological data (Sheng et al., 2014).In the Iberian Peninsula there are numerous sites associated with the accumulating activity of cave hyenas, sites such as the Cueva del Búho (Iñigo et al., 1998), Torrejones (Arribas, 1997; Carrión et al., 2006) Gabasa 1 (Blasco and Montes, 1997), Las Ventanas (Carrión et al., 2001), Nerja (Arribas et al., 2004), Zafarraya (Geraads, 1995 Arribas et al., 2010) and Gorham (Carrión et al., 2008). The presence of Crocuta spelaea extended throughout the Late Pleistocene, lasting almost to the LGM (Last Glacial Maximum) as also the case in northern Eurasia (Stuart and Lister, 2014). However in Cueva de las Ventanas there is an indirect record (coprolites) that is dated to ca. 12 780 cal yr BP (Carrión et al., 2001), which is one of the latest records of this taxon in Europe. The reason for the disappearance of cave hyenas from Europe may be related to climatic factors or to other factors, such as reduced availability of prey or the ecological impact of hominids on the physical environment (Varela et al., 2010; Varela, 2011; Stuart and Lister, 2014).
Although the taxon is present at many sites, this is often in the form of teeth, either from adult or juvenile individuals, and isolated bones, or indirect evidence as coprolites (Carrión et al., 2001, 2006; Álvarez-Lao, 2002; Arsuaga et al., 2012). As a result of their gregarious, scavenging and cannibalistic behaviour, remains of Crocuta spelaea tend to be highly modified by gnawing, and associated skeletal remains are extremely rare (Diedrich, 2005, 2011a, 2012b, 2014). Up to now, the best-preserved remains of C. spelaea in the Iberian Peninsula were the crania from Gabasa 1 (Blasco and Montes, 1997) and the cranium from Fontainhas (Cardoso, 1993). Within Europe the most complete fossil is the one from Les Oubliettes (Gargas, France) (Cardoso, 1993) and the skeleton from Koněprusy Cave (Czech Republic) (Diedrich, 2012a). As a result of its taphonomic history, the C. spelaea skeleton from Los Aprendices is one of the most complete and best-preserved in Europe, and certainly the best-preserved in the Iberian Peninsula (Figure 15).
Taxonomic Attribution of the Studied Material
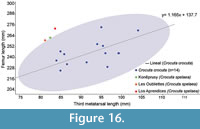 The characteristics of Crocuta spelaea have been described by other authors based on observations from isolated remains of cave hyena (Reynolds, 1902; Kurtén, 1957). The Los Aprendices skeleton provides a unique opportunity to study the anatomical proportions of cave hyenas based on a semi-complete individual. The Los Aprendices skeleton presents some characteristics of C. spelaea, such as the short diastema between C and P1, and P1 and P2 in contact, while in extant C. crocuta P1 and P2 are separated by a diastema. The dental series displays a moderate curvature in the maxillary and mandible, which is characteristic of C. spelaea (García, 2003). Furthermore, the skeleton has a huge skull, a powerful mandible and long proximal hindlimb elements (femur) compared to other cave hyenas. By contrast, medium and distal hindlimb parts are more robust and shorter than those of extant C. crocuta. In the fossil record, there are few articulated C. spelaea skeletons, in fact only three have been recovered so far. The skeleton of Koněprusy Cave (Czech Republic; Diedrich and Žák, 2006), Les Oubliettes (Gargas, France; Cardoso, 1993) and Los Aprendices. Despite the scarce material of C. spelaea, these skeletons present a similar femur length and metatarsus length ratio. This pattern is different from extant C. crocuta (Figure 16). These features agree with those previously reported for C. spelaea and support the idea that C. spelaea and C. crocuta are different species (Kurtén, 1957; Baryshnikov, 1999; Vinuesa et al., 2015) (Table 3, Figure 12). The biometric analysis of cranial elements reveals that there are two clearly distinct populations: one is formed by the extant C. crocuta, and the other by the cave hyena C. spelaea, which is the species characteristic of the Eurasian Late Pleistocene (Figure 7, Figure 8, Figure 12). On the other hand, there are no morphological or metrical criteria to distinguish C. c. intermedia from extant C. crocuta (Kurtén, 1957; García, 2003). Therefore, the cave hyena could be considered as a Late Pleistocene form that evolved from the C. crocuta, which arrived to the Iberian Peninsula in “the Crocuta crocuta event” (Martínez-Navarro, 2010). In this context, the Los Aprendices skeleton is the first skeleton in the Peninsula, which clearly belongs to C. spelaea. Furthermore, the level where it was found has been dated to 143.8 ± 38.9 ka, meaning that we are dealing with one of the oldest - if not the oldest - and most complete records of Crocutaspelaea in the Iberian Peninsula (Figure 15, Appendix 7).
The characteristics of Crocuta spelaea have been described by other authors based on observations from isolated remains of cave hyena (Reynolds, 1902; Kurtén, 1957). The Los Aprendices skeleton provides a unique opportunity to study the anatomical proportions of cave hyenas based on a semi-complete individual. The Los Aprendices skeleton presents some characteristics of C. spelaea, such as the short diastema between C and P1, and P1 and P2 in contact, while in extant C. crocuta P1 and P2 are separated by a diastema. The dental series displays a moderate curvature in the maxillary and mandible, which is characteristic of C. spelaea (García, 2003). Furthermore, the skeleton has a huge skull, a powerful mandible and long proximal hindlimb elements (femur) compared to other cave hyenas. By contrast, medium and distal hindlimb parts are more robust and shorter than those of extant C. crocuta. In the fossil record, there are few articulated C. spelaea skeletons, in fact only three have been recovered so far. The skeleton of Koněprusy Cave (Czech Republic; Diedrich and Žák, 2006), Les Oubliettes (Gargas, France; Cardoso, 1993) and Los Aprendices. Despite the scarce material of C. spelaea, these skeletons present a similar femur length and metatarsus length ratio. This pattern is different from extant C. crocuta (Figure 16). These features agree with those previously reported for C. spelaea and support the idea that C. spelaea and C. crocuta are different species (Kurtén, 1957; Baryshnikov, 1999; Vinuesa et al., 2015) (Table 3, Figure 12). The biometric analysis of cranial elements reveals that there are two clearly distinct populations: one is formed by the extant C. crocuta, and the other by the cave hyena C. spelaea, which is the species characteristic of the Eurasian Late Pleistocene (Figure 7, Figure 8, Figure 12). On the other hand, there are no morphological or metrical criteria to distinguish C. c. intermedia from extant C. crocuta (Kurtén, 1957; García, 2003). Therefore, the cave hyena could be considered as a Late Pleistocene form that evolved from the C. crocuta, which arrived to the Iberian Peninsula in “the Crocuta crocuta event” (Martínez-Navarro, 2010). In this context, the Los Aprendices skeleton is the first skeleton in the Peninsula, which clearly belongs to C. spelaea. Furthermore, the level where it was found has been dated to 143.8 ± 38.9 ka, meaning that we are dealing with one of the oldest - if not the oldest - and most complete records of Crocutaspelaea in the Iberian Peninsula (Figure 15, Appendix 7).
Paleobiological Inferences from the Studied Material
The size of the Los Aprendices skeleton, as well as the size of Crocuta spelaea, were obtained using published regression equations for body mass on postcranial measurements in modern carnivores (Anyonge, 1993; Christiansen, 1999), as well as equations that provide mass estimates from craniodental variables (Van Valkenburgh, 1990) (Table 6).
Regression equations with craniodental variables, such as skull length (SKL) and lower carnassial length (Lm1; equation recalculated with data from Van Valkenburgh, 1990), were used to estimate the body mass of Crocuta spelaea and compared it with the mass estimates for extant C. crocuta obtained by Palmqvist et al. (2011). In Table 6, it can be observed that the equations overestimate the size of extant C. crocuta, and it is likely that this is also the case in C. spelaea. On the other hand, the regression equations for body mass (BM) on limbs, including humerus length (HL) and femur length (FL), offer a better correlation between the real body mass of spotted hyenas and the results obtained by regression equations (Palmqvist et al., 2011).
The study of Palmqvist et al. (2011) shows that the most accurately estimated body mass was obtained with the mean of all estimated masses. The mean mass calculated for Crocuta spelaea is 88 kg, which is 60% more than the extant C. crocuta. This increase in size of C. spelaea from the C. crocuta from the Middle Pleistocene may have been due to climatic factors, for different sizes are detected in glacial and interglacial periods. It is not possible to distinguish these periods on the basis of the size of the hyenas, but it seems clear that temperature and mean body size are inversely related in accordance with Bergmann’s rule (Kurtén, 1957; Kurtén and Poulianos, 1977; Klein and Scott, 1989, Fourvel et al., 2015).
In the case of Los Aprendices, the estimated body mass is 103 kg, which makes it one of the largest cave hyenas ever recorded. However, the body mass could only be estimated but some of equations cited, due to the incompleteness of the studied humerus. According to the later data, the body mass could be overestimated as a consequence of the observed deviations for the regression equations based only on cranial values.
The crural index (tibia length/femur length) in the skeleton of Los Aprendices is 0.74, which is similar to the values of Crocuta spelaea (0.75) and Pachycrocuta brevirostris (0.74) and clearly lower than in extant C. crocuta (0.82) (Palmqvist et al., 2011). The shortening of the tibia of C. spelaea suggests a less cursorial lifestyle. Also, such shortening could provide great power and more stability to dismember and carry large parts of carcasses without dragging (Spoor, 1985; Turner and Antón, 1996; Palmqvist et al., 2011).
Crocuta spelaea was a heavier and more powerful animal than extant C. crocuta, so C. spelaea could use its weight and strength to reduce larger prey. Consequently, this huge carnivore was capable of accumulating bones of large prey like horses (Equus ferus), woolly rhinos (Coelodonta antiquitatis), giant deer (Megaloceros giganteus) and woolly mammoth (Mammuthus primigenius) (Diedrich, 2012b, 2014).
In the last years, actuo-taphonomic studies show that Crocuta crocuta seldom creates large accumulations of bones despite having the ability. On the contrary, it is thought that the cave hyena made large accumulations of bones during the Late Pleistocene throughout Europe (Turner et al., 2008; see references in Diedrich, 2014; Fosse, 1997; Fourvel, 2012, Fourvel et al., 2015). Therefore, apart from the morphological differences between C. spelaea and C. crocuta, ethological differences might exist as well. Another point that supports the separation of C. crocuta and C. spelaea into two different species is the morphology of their brains, which may be related to the differences in behaviour of these carnivores (Vinuesa et al., 2015). However, it should be borne in mind that behavioural differences could also be due to climatic factors, as well as availability of prey (Fourvel and Mwebi, 2011).
Taphonomy
The characteristics of the breakage in the specimen from Los Aprendices are typical of sites at which the fractures were produced after deposition and with the bone in a dry state, as at Sarrians, Besouze (Villa and Mahieu, 1991) or Coro Tracito (Rabal-Garcés, 2013). This type of breakage is produced by trampling or alternatively by the pressure of sediment or falling blocks (Villa and Mahieu, 1991). In the case of Los Aprendices, it is more likely to have been produced by the weight of the sediment of the cone than by trampling, given how inaccessible the area is. At sites where the breakage is produced on fresh bones and where the accumulation is due to anthropic activity or to carnivores such as hyenas, by contrast, the breaks are characterized by a curved delineation, gentle angles and regular edges. This pattern is observed at Fontbrégoua (Villa and Mahieu, 1991), Gran Dolina stratum Aurora TD-6 (Díez et al., 1999), Abric Romaní L.B., Vanguard Cave (Cáceres, 2002), and the Cueva del Camino (Arsuaga et al., 2012).
The cranium presents a deformed sagittal crest. This type of deformation, without breaking the cranium, suggests that the carcass underwent rapid burial while the individual still preserved some of its soft tissues and the bone had collagen that endowed it with plasticity.
The length of the bones recovered and the sections of diaphysis suggest that the accumulation does not correspond to remains accumulated by bone-breaking carnivores such as hyenas, which destroy the bones and leave abundant diaphysis fragments with an incomplete circumference (L1-L2 and C1-C2) (Villa and Mahieu, 1991; Díez et al., 1999; Cáceres, 2002). The specimen from Los Aprendices presents a predominance of complete diaphyseal sections. However, modified remains may also take the form of diaphyseal cylinders (C3) from which the two epiphyses are lacking. This is observed in various dens in central Europe such as Balve Cave (Diedrich, 2011b), Koněprusy Caves-Prošek Dome, Nad Kačákem, Turská Maštal Cave (Diedrich and Žák, 2006), Bottrop (Diedrich, 2012b) and Westeregeln (Diedrich, 2007).
With regards to the tooth marks at Los Aprendices, it is difficult to establish which animal produced them, since the modifications in question were produced on the carcass of an isolated hyena. The accumulations generated by hyenas tend to show a great number of carnivore tooth marks (Diedrich, 2011a, 2011b, 2011c, 2012b), with 60-100% of the sample altered. At fossil sites where alterations to remains have been attributed to hyenas (Blumenschine, 1986; Blumenschine and Marean, 1993; Domínguez-Rodrigo, 1994; Marean and Kim, 1998; Yravedra, 2006; Villa et al., 2010), the percentage of bones with tooth marks is greater than 40%. By contrast, accumulations produced by medium-sized carnivores result in a percentage of altered remains of no more than 25% (Brain, 1981; Sauqué et al., 2014). This criterion might suggest the activity of a medium-sized carnivore at Los Aprendices.
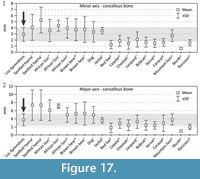 The sizes of the marks (“pits” and “punctures”) found on the hyena from Los Aprendices were compared with data obtained in experiments undertaken by Selvaggio and Wilder (2001), Domínguez-Rodrigo and Piqueras (2003), Saladié (2009) and Delaney-Rivera et al. (2009) (Figure 17). The sizes of the marks from Los Aprendices show great similarity to those of the puma, with the mean values, maxima and minima almost the same. Judging by the size of the marks, other carnivores with a size range from ca 10 kg to 50 kg could be responsible. There is no evidence of the existence of any other carnivore in Los Aprendices Cave, making it difficult to propose a reliable ascription of the bite marks in question. Nonetheless, it is most likely that these were caused by a large- to medium-sized carnivore. The sizes of the marks as well as the patterns of alteration are very similar to those found at the sites of Los Rincones (Sauqué et al., 2014).
The sizes of the marks (“pits” and “punctures”) found on the hyena from Los Aprendices were compared with data obtained in experiments undertaken by Selvaggio and Wilder (2001), Domínguez-Rodrigo and Piqueras (2003), Saladié (2009) and Delaney-Rivera et al. (2009) (Figure 17). The sizes of the marks from Los Aprendices show great similarity to those of the puma, with the mean values, maxima and minima almost the same. Judging by the size of the marks, other carnivores with a size range from ca 10 kg to 50 kg could be responsible. There is no evidence of the existence of any other carnivore in Los Aprendices Cave, making it difficult to propose a reliable ascription of the bite marks in question. Nonetheless, it is most likely that these were caused by a large- to medium-sized carnivore. The sizes of the marks as well as the patterns of alteration are very similar to those found at the sites of Los Rincones (Sauqué et al., 2014).
One factor to keep in mind is that cannibalistic behaviour is very common in Crocuta crocuta (Frank, 1994). Present-day hyenas accumulate bones from individuals of their own species, as has been observed in Africa (Cooper, 1993; East et al., 1989). This behaviour has also been recorded in Crocuta spelaea; in numerous hyena dens crania have been found with bite marks and with jugal arches torn off and mandibles broken (Diedrich, 2005, 2011a, 2011c). Examples are seen in the material from Perick Cave (Diedrich, 2005), Sloup Cave (Diedrich, 2012b), Konĕprusy and Chlum Komin Cave (Diedrich and Žák, 2006).
The frequency of individual marks on each bone also varies greatly depending to the accumulating agent in question. Leopards rarely (<5%) leave more than 10 marks on a bone, whereas in the case of hyenas it is common for bones to present numerous individual marks, as many as 42 on a single bone (Selvaggio, 1994; Domínguez-Rodrigo and Pickering, 2010). At Los Aprendices there are no elements with more than seven marks on a single bone (see humerus MPZ 2014/672).
 In summary, the low level of post-depositional breakage, the high number of elements represented, the presence of these elements in anatomical semi-connection, the fact that no element is repeated, as well as the left-right mirror symmetry of the elements and the low degree of modification caused by teeth, suggest that the hyena from Los Aprendices was in a complete state when it reached the sediment cone blocking the former entrance to the cave. Once dead, its carcass was presumably scavenged by a medium-sized carnivore. Both the modification pattern and the size of the marks are very similar to those that could have been produced by either a wolf or a European leopard. The presence of both these taxa has been established at a Late Pleistocene site in the vicinity of the site of Los Rincones (Sauqué et al., 2014, 2016a, 2016b). Moreover, taphonomic analysis of the hyena indicates that the marks were not produced by another cave hyena, suggesting that it is most likely that the animal met its death in a solitary context. In other words, if the hyena had been with a clan of cave hyenas at the time of its death, these other hyenas would have scavenged on its carcass, causing a large part of the skeleton to disappear and dispersing the bones (Figure 18). Cannibalistic and scavenging activities of this sort have been recorded in many central European sites (Diedrich, 2005, 2011a, 2011b, 2011c, 2011d, 2012a, 2012b, 2014; Villa et al., 2010).
In summary, the low level of post-depositional breakage, the high number of elements represented, the presence of these elements in anatomical semi-connection, the fact that no element is repeated, as well as the left-right mirror symmetry of the elements and the low degree of modification caused by teeth, suggest that the hyena from Los Aprendices was in a complete state when it reached the sediment cone blocking the former entrance to the cave. Once dead, its carcass was presumably scavenged by a medium-sized carnivore. Both the modification pattern and the size of the marks are very similar to those that could have been produced by either a wolf or a European leopard. The presence of both these taxa has been established at a Late Pleistocene site in the vicinity of the site of Los Rincones (Sauqué et al., 2014, 2016a, 2016b). Moreover, taphonomic analysis of the hyena indicates that the marks were not produced by another cave hyena, suggesting that it is most likely that the animal met its death in a solitary context. In other words, if the hyena had been with a clan of cave hyenas at the time of its death, these other hyenas would have scavenged on its carcass, causing a large part of the skeleton to disappear and dispersing the bones (Figure 18). Cannibalistic and scavenging activities of this sort have been recorded in many central European sites (Diedrich, 2005, 2011a, 2011b, 2011c, 2011d, 2012a, 2012b, 2014; Villa et al., 2010).
CONCLUSIONS
Los Aprendices Cave (Zaragoza Province, Spain), has yielded a partially complete skeleton of Crocuta spelaea associated with Capra pyrenaica, Lagomorpha indet., Arvicolidae indet. and Galemys pyrenaicus.
The skeleton is the best preserved Pleistocene Crocuta specimen in the Iberian Peninsula and one of the most complete specimens in the European fossil record. Cave hyena fossils are usually fragmentary, and it is difficult to establish the relationships between the different features of the cranial and postcranial elements. For this reason we consider that the skeleton form Los Aprendices may be a reference for future studies. A mass of 103 kg has been estimated for Los Aprendices skeleton, making it one of the largest C. spelaea in the European record. This skeleton presents the characteristic features of C. spelaea such as: P3 with an extremely high crown, short diastema between C and P1, P1 and P2 in contact, disposition of the premolars in the dental series clearly curved, relative shortening of the distal parts of the extremities and larger size compared with C. crocuta. These features could be related with its behaviour as the main non anthropogenic bone accumulator during Pleistocene.
The Crocuta spelaea from Los Aprendices preserves 56 % of the total skeleton, providing critical information on the final moments of its life that suggest that it was solitary at the time of death. The skeleton is partially complete and 107 anatomical elements and a total of 194 remains have been recovered. The hyena from Los Aprendices died in the sediment cone that led to the cave, and it was subsequently modified by a large-medium-sized carnivore (probably a wolf or leopard). The pattern of modification and the type of bite marks, as well as the size of these marks, suggest that it was not scavenged by another cave hyena, as habitually occurs among this species (see references in Diedrich, 2014).
The level from which the skeleton was recovered has been dated by amino acid racemization to 143.8 ± 38.9 ka, which would place it either in the Middle Pleistocene or in the early Late Pleistocene, suggesting that Los Aprendices is one of the oldest records of Crocuta spelaea in the Iberian Peninsula.
The greatest expansion of the cave hyena in the Iberian Peninsula took place during the Late Pleistocene, between MIS 4 and MIS 2. Furthermore, the species may have survived longer in this area than in other parts of Europe, with records dated close to the Holocene. In our review of cave hyena distribution in the Iberian Peninsula we reallocated some Crocuta crocuta to C. spelaea mainly based on morphometry. In this review we included 90 sites with presence of cave hyena, which makes the Iberian Peninsula one of the areas with the highest density of sites containing cave hyena remains in Europe.
ACKNOWLEDGMENTS
We should like to thank the Centro Espeleológico de Aragón (CEA), and the Instituto Pirenaico de Ecología, in particular R. García-González, who is in charge of the vertebrate collection. We also thank to J. Colmenar and D. Castanera for their help taking the photographs and S. Serrano for helping with the manuscript. The authors thank R. Glasgow for revising the English grammar.
Various investigators have provided us with information about hyena remains in the Asturias (D. Álvarez Lao), Valencia (A. Sanchis) and France (P. Fosse). The authors thank L. Montes, P. Utrilla and M.F. Blasco for granting access to unpublished data on Gabasa as well as their valuable comments. The authors thank F. Gutierrez and I. Sasowsky for their valuable comments about karst and geology. We would like to thank C. Tormo and D. Santamaría for their help with Figure 4.
This work has been funded by the Spanish Ministerio de Economía y Competitividad (CGL2011-28681, CGR2014-5473-P and CGL2012- 38434-C03-01.) and the Generalitat de Catalunya (2014 SGR 416 GRC). We sincerely thank J. Quesada, V. Vicedo, E. Gilissen and W. Wendelen for providing access to the collections under their care.
A particular thanks to palaeoartist Gianfranco Mensi for his precise reconstruction. Finally, many thanks go to all the members of the excavation team (J. Ruiz, D. Ramón del Río, D. Castanera) of the 2012, field campaigns at Los Aprendices.
The Government of Aragon and the University of Zaragoza have partially subsidised the palaeontological activities at Los Aprendices. S.Z. is funded by a Ramón y Cajal Grant (RYC-2012-10576) from the Spanish Ministry of Economy and Competitiveness.
The authors also thank the reviewers for the useful suggestions that greatly improved the manuscript. Finally, the authors thank the careful work of the editorial team of Palaeontologia Electronica.
REFERENCES
Aguirre, E. 1987. Los yacimientos mesopleistocenos en cavidades de la Sierra de Atapuerca, p. 201-204 In Aguirre, E., Carbonell, E., and Bermudez de Castro, J.M. (eds.), El hombre fósil de Ibeas y el Pleistoceno de la Sierra de Atapuerca. Junta de Castilla Y Leon Valladolid.
Altuna, J. 1971. Los mamíferos del yacimiento prehistórico de Morín (Santander), p. 367-399. In Gonzalez Echegaray, J. and Freeman, L.G (eds.). Cueva Morín. Excavaciones 1966-1968. Publicaciones del Patronato de las cuevas Prehistóricas de la Provincia de Santander, Santander.
Altuna, J. 1972. Fauna de Mamíferos de los yacimientos prehistóricos de Guipúzcoa. Munibe, 24:1-464.
Altuna, J. 1990. Caza y alimentación procedente de macromamíferos durante el Paleolítico de Amalda, p. 149-192. In Altuna, J., Baldeón, A., and Mariezkurrena, K. (eds.), La Cueva de Amalda (Zestoa, País Vasco): Ocupaciones Paleolíticas y Postpaleolíticas (volume B4). Fundación José Miguel de Barandiarán, San Sebastián.
Altuna, J. and Mariezkurrena, K. 1984. Bases de subsistencia de origen animal de los pobladores de Ekain, p. 211-280. In Altuna, J. and Merino, J.M. (eds.), El Yacimiento Prehistórico de la cueva de Ekain (Deba, Gipuzkoa). Sociedad de Estudios Vascos, San Sebastián.
Altuna, J. and Mariezkurrena, K. 2000. Macromamíferos del yacimiento de Labeko Koba (Arrasate, País Vasco). Munibe, 52:107-181.
Altuna, J., Mariezkurrena, K., Armendariz, A., Del Barrio, L., Ugalde, T., and Peñalver, J. 1982. Carta arqueológica de Guipúzcoa. Munibe, 34:1-242.
Álvarez Lao, D.J. 2002. Recuperación y estudio de la colección de Vertebrados Cuaternarios del Departamento de Geología de la Universidad de Oviedo. Revista Española de Paleontología, 17:187-198.
Álvarez-Lao, D.J. 2003. Macromamíferos fósiles del Pleistoceno de Asturias. Revisión bibliográfica y datos actuales. Naturalia Cantabricae, 2:11-23.
Álvarez-Lao, D.J. and García-García, N. 2006. A new site from the Spanish Middle Pleistocene with cold-resistant faunal elements: La Parte (Asturias, Spain). Quaternary International, 142:107-118.
Álvarez-Lao, D.J., Ruiz-Zapata, M.B, Gil-García, M.J, Ballesteros, D., and Jiménez-Sánchez, M. 2015. Palaeoenvironmental research at Rexidora Cave: New evidence of cold and dry conditions in NW Iberia during MIS 3. Quaternary International, 379:35-46.
Anyonge, W. 1993. Body mass in large extant and extinct carnivores. Journal of Zoology, 231:339-350.
Arbizu, M., Arsuaga, J.L., and Adán, G.E. 2005. La Cueva del Forno/Conde (Tuñón, Asturias): un yacimiento del tránsito del Paleolítico Medio y Superior en la Cornisa Cantábrica, p. 10-38. In Montes, R. and Lasheras, J.A. (eds.), Neanderthales cantábricos, Estado de la Cuestión. Museo Nacional y Centro de Investigación de Altamira, Santander.
Arbizu, M., Álvarez-Lao, D., and Adan, G.E. 2005. Una guarida de Crocuta crocuta en Leguín (Echauri, Navarra). Munibe, 57:131-138.
Argant, A. 1991. Carnivores Quaternaires de Bourgogne. Documents des Laboratoires de Géologie de Lyon, Lyon.
Arribas, A. 1997. Un leopardo, Panthera pardus (Linnaeus, 1758), en el Pleistoceno de la Cueva de los Torrejones (Tamajón, Guadalajara, España). Geogaceta, 22:19-22.
Arribas, A., Garrido, G., and García Solano, J.A. 2010. Los hiénidos del Plio-Pleistoceno español: géneros y especies, distribución temporal y espacial. Zona Arqueológica, 13:34-49.
Arribas Herrera, A., Aura Tortosa, J.E., Carrión, J.D., Jordá Pardo, J.F., and Pérez Ripoll, M. 2004. Presencia de hiena manchada en los depósitos basales (Pleistoceno superior final) del yacimiento arqueológico de la Cueva de Nerja (Málaga, España ), Revista Española de Paleontología, 19:109-121.
Arribas Herrera, A., Díez Fernández-Lomana, J.C., and Jordá Pardo, J.F. 1997. Primeras ocupaciones en los depósitos pleistocenos de la Cueva de los Torrejones (Sistema Central español, Tamajón, Guadalajara). Litoestratigrafía y Actividad Biológica: Cuaternario y Geomorfología, 11:55-66.
Arribas, A., Solano, J.G., Garrido, G. and Carroza, JA. 2008. El registro diferencial de carnívoros y homínidos en el karst del Pleistoceno del Sistema Central español: condicionantes geológicos y espaciales, p. 39-52. In Díez, J.C., (ed.) Zooarqueología Hoy: Encuentros Hispano-Argentinos.
Arribas, A., Garrido, G., and García Solano, J.A. 2010. Los hiénidos del Plio-Pleistoceno español: géneros y especies, distribución temporal y espacial. Zona Arqueológica, 13:34-49.
Arrizabalaga, A. 2000. El yacimiento arqueológico de Labeko Koba (Arrasate, País Vasco). Entorno. Crónica de las investigaciones. Estratigrafía y estructuras. Cronología absoluta. Munibe, 52:15-72.
Arsuaga, J.L., Baquedano, E., Pérez-González, A., Sala, M.T.N., García, N., Álvarez-Lao, D., Laplana, C., Huguet, R., Sevilla, P., Blain, H.-A., Quam, R., Ruiz Zapata, M.B, Sala, P., Gil García, M.J., Uzquiano, P. and Pantoja, A. 2010. El yacimiento kárstico del Pleistoceno Superior de la Cueva del Camino en el Calvero de la Higuera (Pinilla del Valle, Madrid). Zona Arqueológica, 13:349-368.
Arsuaga, J.L., Baquedano, E., Pérez-González, A., Sala, N., Quam, R.M., Rodríguez, L., García, R., García, N., Álvarez-Lao, D.J., Laplana, C., Huguet, R., Sevilla, P., Maldonado, E., Blain, H.A., Ruiz-Zapata, M.B., Sala, P., Gil-García, M.J., Uzquiano, P., Pantoja, A., and Márquez, B. 2012. Understanding the ancient habitats of the last-interglacial (late MIS 5) Neanderthals of central Iberia: paleoenvironmental and taphonomic evidence from the Cueva del Camino (Spain) site. Quaternary International, 275:55-75.
Aura Tortosa, J.E., Jodá Pardo, J.F., Avezuela Aristu, B., Pérez Ripoll, M., Tiffagom, M., and Morales Pérez, J.V. 2011 . La Cueva de Nerja (Málaga, España) y el Gravetiense en Andalucía. Cuaternario y Arqueología. Homenaje a Francisco Giles Pacheco, ASPHA and Diputación Provincial de Cadiz, Cadiz.
Ballesio, R. 1979. Le gisement pléistocène supérieur de la grotte de Jaurens à Nespouls, Corrèze, France. Les Carnivores (Mammalia, Carnivora). Canidae et Hyaenidae. Nouvelles Archives du Muséum d’Histoire naturelle de Lyon, Lyon.
Barandiarán, I. 1973. Excavaciones en la Cueva de los Casares (Riba de Saelices, Guadalajara). Excavaciones Arqueológicas en España, 76.
Barroso, C., Riquelme, J.A., Moigne, A.M., and Banes, L. 2006. Les faunes de grands mammifères du Pléistocène supérieur de la Grotte du Boquete de Zafarraya. Etude Paléontologique, Paléoécologique et Archéozoologique, p. 675-891. In Barroso, C. and de Lumley, H. (eds.), La Grotte du Boquete de Zafarraya, Málaga.
Baryshnikov, G. 1999. Chronological and geographical variability of Crocuta spelaea (Carnivora, Hyaenidae) from the Pleistocene of Russia. Deinsea, 6:155-174.
Baryshnikov, G. 2014. Late Pleistocene hyena Crocuta ultima ussurica (Mammalia: Carnivora: Hyaenidae) from the Paleolithic site in Geographical Society Cave in the Russian far east. Proceedings of the Zoological Institute RAS , 318(3):197-225.
Baryshnikov, G. and Tsoukala, E. 2010. New analysis of the Pleistocene carnivores from Petralona cave (Macedonia, Greece) based on the collection of the Thessaloniki Aristotle university. Geobios, 43:389-402.
Binford, L.R. 1981. Bones. Ancient Men and Modern Myths. Academic Press, New York.
Blasco , M.F. and Montes , L. 1997. Los hiénidos del yacimiento musteriense de Gabasa 1 (Huesca, España). Bolskan, 14:9-27.
Blumenschine, R. 1986. Carcass consumption sequences and the archaeological distinction of scavenging and hunting. Journal of Human Evolution, 15:639-659.
Blumenschine, R. and Marean, C.W. 1993. A carnivore’s view of archaeological bone Assemblages, p.273-300. In Hudson J. (ed.), From Bones to Behavior. Ethno Archaeological and Experimental Contributions to the Interpretation of Faunal Remains. Illinois Center of Archaeological Investigations, Southern Illinois University press, Illinois
Bonifay, M.F. 1971. Carnivores Quaternaires du Sud-Est de la France. Mémoires du Muséum d'Histoire Naturelle, Paris, Paris.
Bowdich, T.E. 1821. An Analysis of the Natural Classifications of Mammalia for the use of Students and Traveller s. J. Smith, Paris.
Brain, C.K. 1981. The Hunters or the Hunted? An Introduction to African Cave Taphonomy. University of Chicago Press, Chicago.
Brugal, J.P., Fosse, P., and Guadelli, J.-L. 1997. Comparative study of bone assemblages made by recent and pleistocene hyenids, p. 158-187. In Hannus, L.A., Rossum, L., and Peter, W. (eds.), Proceedings of the Bone Modification Conference. Hot Spring, Dakota City.
Cabrera Valdés, V. 1984. El yacimiento de la Cueva de “El Castillo”. Biblioteca Praehistorica Hispana, 22, Madrid.
Cáceres, I. 2002. Tafonomía de yacimientos antrópicos en Karst. Complejo Galería (Sierra de Atapuerca, Burgos), Vanguard Cave (Gibraltar) y Abric Romaní (Capellades, Barcelona). Unpublished PhD Thesis, Universitat Rovira i Virgili, Tarragona.
Cardoso, J.L. 1993. La hyènes des «Oubliettes» de Gargas, Crocuta crocuta spelaea (Mammalia, Carnivora). Bulletin du Muséum National d’histoire Naturelle. Section C, Sciences de la terre, Paléontologie, Géologie, Minéralogie, 15:79-104.
Cardoso, J.L. 1995. Grutas do Maciço Hespérico de Portugal com faunas de grandes mamíferos plistocénicos. Breve síntese. Cadernos Laboratorio Xeolóxico de Laxe, 20:213-229.
Cardoso, J.L. 1996. Les grands Mammifères du Pléistocène supérieur du Portugal. Essai de synthèse. Geobios, 29:235-250.
Carrión, J.S., Finlayson, C., Fernández, S., Finlayson, G., Allue, E., López-Sáez, J.A., López-García, G., Gil-Romera, G., Bailey, G., and González-Sampériz, P. 2008. A coastal reservoir of biodiversity for Upper Pleistocene human populations: palaeoecological investigations in Gorham’s Cave (Gibraltar) in the context of the Iberian Peninsula. Quaternary Science Review, 27:2118-2135.
Carrión, J.S., Riquelme, J.A., Navarro, C., and Munuera, M. 2001. Pollen in hyaena coprolites reflects late glacial landscape in southern Spain. Palaeogeography, Palaeoclimatology,Palaeoecology, 176:193-205.
Carrión, J.S., Scott, L., Arribas, A., Fuentes, N., Gil, G., and Montoya, E. 2006. Pleistocene landscapes in central Iberia inferred from pollen analysis of hyena coprolites. Journal of Quaternary Science, 21:1-12.
Carrión, J.S., Yll, E.I., Walker, M.J., Legaz, A.J., Chaín, C., and López, A. 2003. Glacial refugia of temperate, Mediterranean and Ibero‐North African flora in south‐eastern Spain: new evidence from cave pollen at two Neanderthal man sites. Global Ecology and Biogeography, 12:119-129.
Castaños, P.M. 1983. Estudio de los macromamíferos del yacimiento prehistórico de Bolinkoba (Abadiano-Vizcaya). Kobie Paleoantropología, 13:261-298.
Castaños, P.M. 2005. Revisión actualizada de las faunas de macromamíferos del Würm antiguo en la Región Cantábrica. Museo de Altamira. Monografías, 20:201-207.
Castaños, P. 1987. Los carnívoros prehistóricos de Vizcaya. Kobie Paleoantropología, 16:7-76.
Christiansen, P. 1999. What size were Arctodus simus and Ursus spelaeus (Carnivora: Ursidae)? Annales Zoologici Fennici, 36:93-102.
Clot, A. 1980. La grotte de la Carrière (Gerde, Haute-Pyrénés, France). Stratigraphie et Paléontologie des Carnivores. Unpublished PhD Thesis, Université Paul Sabatier, Toulouse.
Cooper, S.M. 1993. Denning behavior of spotted hyaenas (Crocuta crocuta) in Botswana. African Journal of Ecology, 31:178-180.
Cortés Sánchez, M., Muñoz, V.E., Sanchidrián, J.L., and Simón, M.D. 1996 . El Paleolítico en Andalucía. La dinámica de los grupos predadores en la prehistoria andaluza. Ensayo de Síntesis. Repertorio bibliográfico de 225 años de investigación (1770-1995), Córdoba.
Cuenca-Bescós, G., Ramón del Río, D., and Canudo J.I. 2010. La Cueva de los Huesos de Obón: un cubil de hiena del Pleistoceno medio de Teruel (España). Zona Arqueológica, 13:271-280.
Currant, A.P. 2000. A review of the quaternary mammals of Gibraltar, p. 201-205. In Stringer, C.B., Barton, R.N.E., and Finlayson, J.C. (eds.), Neanderthals on the Edge. Oxbow Books,Oxford
Daura, J., Sanz, M., García, N., Allué, E., Vaquero, M., Fierro, E., Carrión, J.S., López-García, J.M., Blain, H.A., Sánchez-Marco, A., Valls, C., Albert, R.M., Fornós, J.J., Julià, R., Fullola, J.M, and Zilhão, J. 2013. Terrasses de la Riera dels Canyars (Gavà, Barcelona): the landscape of Heinrich Stadial 4 north of the “Ebro frontier” and implications for modern human dispersal into Iberia. Quaternary Science Review, 60:26-48.
Daura, J., Sanz, M., Pike, A.W.G., Subirà, M.E., Fornós, J.J., Fullola, J.M., Julià, R., and Zilhão, J. 2010. Stratigraphic context and direct dating of the Neandertal mandible from Cova del Gegant (Sitges, Barcelona). Journal of Human Evolution, 59:109-122.
Davis, S.J.M. 2002. The mammals and birds from the Gruta do Caldeirão, Portugal Revista Portuguesa de Arqueologia, 5:29-98.
Davis, S.J.M., Robert, I., and Zilhão, J., 2010. La Cueva de Caldeirão, (Portugal Central) ¿El hogar de hienas, hombres, quebrantahuesos...? Zona Arqueológica, 13:283-296.
Delaney-Rivera, C., Plummer, T.W., Hodgson, J.A., Forrest, F., Hertel, F., and Oliver, J.S., 2009. Pits and pitfalls: taxonomic variability and patterning in tooth mark dimensions. Journal of Archaeological Science, 36:2597-2608.
de Serres, M., Dubreuil, J., and B. Jeanjean. 1828. Mémoires sur les diverses espèces d’Hyènes fossiles découvertes dans les cavernes de Lunel-Vieil (Hérault). Mémoires du Muséum d'Histoire Naturelle, Paris, 17:269-312.
Diedrich, C. 2005. Eine oberpleistozäne Population von Crocuta crocuta spelaea (Goldfuss 1823) aus dem eiszeitlichen Fleckenhyänenhorst Perick-Höhlen von Hemer (Sauerland, NW Deutschland) und ihr Kannibalismus. Philippia, 12:93-115.
Diedrich, C. 2007. The Upper Pleistocene Crocuta crocuta spelaea (Goldfuss 1823) population and its prey from the gypsum karst den site Westeregeln near Magdeburg (Middle Germany). Abhandlungen und Berichte des Museums fiir Natur- und Heimatskunde zu Magdeburg, 30:57-83.
Diedrich, C. 2008. The rediscovery holotypes of the Upper Pleistocene spotted hyena Crocuta crocuta spelaea (Goldfuss 1823) and the steppe lion Panthera leo spelaea (Goldfuss 1810) and taphonomic discussion to the Zoolithen Cave hyena den at Geilenreuth (Bavaria South-Germany). Zoological Journal of Linnean Society, 154:822-831.
Diedrich, C. 2011a. The late Pleistocene spotted hyena Crocuta crocuta spelaea (Goldfuss 1823) population with its type specimens from the Zoolithen Cave at Gailenreuth (Bavaria, South Germany): a hyena cub raising den of specialised cave bear scavengers in boreal forest environments of Central Europe, Historical Biology, 23:335-367
Diedrich, C. 2011b. One of Europe’s last glacial Crocuta crocuta spelaea (Goldfuss 1823) clans from the Rösenbeck Cave hyena den (Germany) and contribution to cranial shape variability. Biological Journal of the Linnean Society, 103:191-220.
Diedrich, C. 2011c. Periodical use of the Balve Cave (NW Germany) as a Late Pleistocene Crocuta crocuta spelaea (Goldfuss, 1823) den: hyena occupations and bone accumulations vs. human Middle Palaeolithic activity. Quaternary International, 233:171-184.
Diedrich, C. 2011d. The Crocuta crocuta spelaea (Goldfuss, 1823) population and its prey from the late Pleistocene Teufelskammer cave hyena den besides the famous Paleolithic Neanderthal cave (NRW, NW Germany). Historical Biology, 23:237-270.
Diedrich, C. 2011e. A clan of Late Pleistocene hyenas, Crocuta crocuta spelaea (Goldfuss 1823), from the Rösenbeck Cave (Germany) and a contribution to cranial shape variability. Biological Journal of the Linnena Society, 103:191-220.
Diedrich, C. 2012a. Europe’s first Upper Pleistocene Crocuta crocuta spelaea (Goldfuss 1823) skeleton from the Koněprusy Caves: a hyena cave prey depot site in the Bohemian Karst (Czech Republic). Historical Biology, 24:63-89.
Diedrich, C. 2012b. The Ice Age spotted Crocuta crocuta spelaea (Goldfuss, 1823) population, their excrements and prey from the Late Pleistocene hyena den Sloup Cave in the Moravian Karst; Czech Republic. Historical Biology, 24:161-185.
Diedrich, C. 2014. Palaeopopulations of Late Pleistocene Top Predators in Europe: Ice Age Spotted Hyenas and Steppe Lions in Battle and Competition about Prey. Paleontol. J. http://dx.doi.org/10.1155/2014/106203.
Diedrich, C. and Žák, K. 2006. Upper Pleistocene hyena Crocutacrocutaspelaea (Goldfuss, 1823) prey deposit and den sites in horizontal and vertical caves of the Bohemian Karst (Czech Republic). Bulletin Geosciences, 81:237-276.
Díez, C., Alonso, R., Bengoechea, A., Colina, A., Jordá, J.F., Navarro, M., Ortiz, J.E., Pérez, S., and Torres, T. 2008. El Paleolítico Medio en el Valle del Arlanza (Burgos): los sitios de La Ermita, Millán y La Mina. Cuaternario y Geomorfología: Revista de la Sociedad Española de Geomorfología y Asociación Española para el Estudio del Cuaternario, 22:35-157.
Díez, J.C., Fernández-Jalvo, Y., Rosell, J., and Cáceres, I. 1999. Zooarchaeology and taphonomy of Aurora Stratum (Gran Dolina, Sierra de Atapuerca, Spain). Journal of Human Evolution, 37:623-652.
Díez, J.C., García, M.A., Gil, E., Jordá, J.F., Ortega, A.I., Sánchez, A., and Sánchez, B. 1988-1989. La cueva de Valdegoba (Burgos). Primera campaña de excavaciones. Zephyrus, XLI-XLII:55-74.
Domingo, M.S., Alberdi, M.T., Sánchez Chillón, B., and Cerdeño, E. 2005. La fauna cuaternaria de la Cornisa Cantábrica en las colecciones del Museo Nacional de Ciencias Naturales. Munibe, 57:5-30.
Domínguez-Rodrigo, M. 1994. Dinámica trófica, estrategias de consumo y alteraciones óseas en la sabana africana: resumen de un Proyecto de Investigación Etoarqueológico (1991-1993). Trabajos de Prehistoria, 51:15-37.
Domínguez-Rodrigo, M. and Pickering, T.R. 2010. A multivariate approach for discriminating bone accumulations created by Spotted hyenas and leopards: Harnessing actualistic data from East and Southern Africa. Journal of Taphonomy, 8:155-179.
Domínguez-Rodrigo, M. and Piqueras, A. 2003. The use of tooth pits to identify carnivore taxa in tooth marked archaeofaunas and their relevance to reconstruct hominid carcass processing behaviours. Journal of Archaeological Science, 30:1385-1391.
East, M.L., Hofer, H., and Türk, A. 1989. Functions of birth dens in spotted hyaenas (Crocuta crocuta). Journal of Zoology London, 219:690-697.
Ehrenberg, K. 1938-1940. Die Fuchs-oder Teufelsleucken bei Eggenburg, Niederdonau. Abhandl. Zoologisch-Botanischen Gesellschaft in Wien, 17:1-301.
Esper, J.F. 1774. Ausführliche Nachricht von neuentdeckten Zoolithen unbekannter vierfüssiger Thiere und denen sie enthaltenden, so wie verschiedenen anderen, denkwürdigen Grüften der Oberbürgischen Lande des Marggrafthums Bayreuth. Nürnberg.
Estévez, J. 1979. La fauna del Pleistoceno catalán, vol. 2. Unpublished PhD Thesis, Universitat de Barcelona, Barcelona.
Estévez, J. 1987. La fauna de L'Arbreda (sector Alfa) en el conjunt de faunes del Pleistocè superior. Cypsela, VI:73-87.
Erxleben, J.C.P. 1777. Anfangsgründe der Naturlehre und Systema regni animalis. Dieterich, Göttingen und Gotha, Göttingen.
Falguères, C., Yokohama, Y., and Arrizabalaga, A.. 2005. La geocronología del yacimiento pleistocénico de Lezetxiki (Arrasate, País Vasco). Crítica de las dataciones existentes y algunas nuevas aportaciones. Munibe, 57:93-106.
Fernández, S., Carrión, J.S., Fuentes, N., González-Sampériz, P., Montoya, E., Gil, G., Vega-Toscano, G., and Riquelme, J.A. 2007. The Holocene and upper Pleistocene pollen sequence of Carihuela cave, southern Spain. Geobios, 40:75-90.
Fosse, P. 1997. Variabilité des assemblages osseux créés par l’hyène des cavernes. Paléo, 9:15-54.
Fosse, P., Brugal, J.P., Guadelli, J.L., Michel, P., and Tournepiche, J.F. 1998. Les repaires d’hyenes des cavernes en Europe occidentale: presentation et comparisons de quelques assemblages osseux. In Brugal, J-P., Meignen, L., and Patou- Mathis, M. (eds.), Economie Prehistorique: Les Comportements de Substance au Paleolithique. XVIII Rencontres Internationals d’Archeologie et d’Historie d’Antibes. Sophia Antipolis: Editions APDCA, 44-61.
Fourvel, J.B. 2012. Hyénidés modernes et fossiles d’Europe et d’Afrique: taphonomie comparée de leurs assemblages osseux. Unpublished PhD Thesis,Université de Toulouse Le Mirail, Toulouse.
Fourvel, J.B., Fosse, P., and Avery, G. 2014. Spotted, striped or brown? Taphonomic studies at dens of extant hyaenas in eastern and southern Africa, Quaternary International, 369:38-50.
Fourvel, J.B., Fosse, P., Fernandez, P., and Antoine, P.O. 2015. Large mammals of Fouvent-Saint-Andoche (Haute-Saône, France): a glimpse into a Late Pleistocene hyena den. Geodiversitas, 37(2):237-266.
Fourvel, J.B. and Mwebi, O. 2011. Hyenas’ level of dependence on livestock in pastoralist areas in the Republic of Djibouti and Kenya: relation between prey availability and bone consumption sequence. Prédateurs dans tous leurs états: évolution, biodiversité, interactions, mythes, symboles: actes des rencontres, Antibes. 21-23 October 2010, 157-176.
Frank, L.G. 1994. When hyenas kill their own. New Scientist , 141:38-41.
García, N. 2003. Osos y otros carnívoros de la Sierra de Atapuerca. Fundación Oso de Asturias, Oviedo.
García, N. and Arsuaga, J.L. 1999. Carnivores from the Early Pleistocene hominid-bearing Trinchera Dolina 6 (Sierra de Atapuerca, Spain). Journal of Human Evolution, 37:415-430.
Geoffroy, E. 1811. Memoire sur les espéces des genres Musaraigne et Mygale . Annales du Muséum National d'Histoire Naturelle, 17: 169-194.
Geraads, D. 1995. Carnívoros musterienses de la Cueva de Zafarraya (Málaga). Cuaternario y Geomorfología, 9(3-4):51-57.
Gisbert, M. and Pastor, M. 2009. Cuevas y Simas de la provincia de Zaragoza. Centro de Espeleología de Aragón (CEA), Zaragoza.
Goldfuss, A. 1823. Osteologische Beiträge zur Kenntnis verschiedener Säugethiere der Vorwelt. Nova Acta Physico-Medica Academiae Caesarae Leopoldino Carolinae Naturae Curiosorum, 11(2):451-490.
González Echegaray, J. 1980. El yacimiento de la cueva de “El Pendo”: excavaciones 1953-57. Consejo Superior de Investigaciones Científicas, Madrid.
Grandal D’Anglade, A., López-González, F., and Vidal Romaní, J.R. 1997. Condicionantes en la distribución de macromamíferos en Galicia (NW Península Ibérica) durante el Cuaternario superior. Cadernos Laboratorio Xeolóxico de Laxe , 22:43-66.
Gray, J.E. 1821. On the natural arrangement of vertebrose animals. London Medical Repository, 15:269-310
Gusi, F., Gibert, J., Agustí, J., and Pérez Cueva, A. 1987. Nuevos datos del yacimiento Cova del Tossal de la Font (Vilafamés, Castellón). Cuadernos de prehistoria y arqueología castellonenses, 10:7-18.
Gusi, F., Olària, C., Ollé, A., Saladié, P., Vallverdú, J., Cáceres, I., van der Made, J., Expósito, I., Burjachs, F., López-Polín, L., Lorenzo, C., Bennàsar, M., Salazar-García, D.C., and Carbonell, E. 2013. La Cova de Dalt del Tossal de la Font (Vilafamés, Castellón): conclusiones preliminares de las intervenciones arqueológicas (1982-1987/2004-2012). Cuadernos de Prehistoria y Arqueología Castellonenses, 31:17-37.
Haynes, G. 1980. Evidence of carnivore gnawing on Pleistocene and Recent mammalian bones. Paleobiology, 6:341-351.
Haynes, G. 1983. A guide for differentiating mammalian carnivore taxa responsable for gnaw damage to herbivore limb bones. Paleobiology, 9:164-172.
Hollister, N. 1918. East African Mammals in the United States National Museum (volume 99). US Government Printing Office, Washington.
Iñigo, C., Molero, G., and Maldonado, E. 1998. Los carnívoros del yacimiento pleistoceno de Cueva del Buho (Segovia, España) y sus huellas de actividad. Estudios Geológicos, 54:65-73.
Iturbe, G., Fumanal, M.P., Carrión, J.S., Cortell, E., Martínez, R., Guillem, P.M., Garralda, M.D., and Vandermeersch, B. 1993. Cova Beneito (Muro, Alicante): una perspectiva interdisciplinar. Recerques del Museu d'Alcoi, 2:23-88.
Jiménez Fuentes, E., Cardoso, J.L., and Crespo, E.G. 1998. Presencia de Agrionemys (= Testudo) hermanni (Gmelin, 1789) en el Paleolítico Medio de la Gruta Nova da Columbeira (Bombarral, provincia de Estremadura, Portugal). Stvdia Geologica Salmanticensia, 34:123-139.
Kaup, J.J. 1828. Über Hyaena, Uromastix, Basiliscus, Corythaeolus, Acontias. Isis von Oken (Leipzig), 21:1144-1150.
Klein, R.G. and Scott, K. 1989. Glacial/interglacial size variation in fossil spotted hyenas (Crocuta crocuta) from Britain. Quaternary Research, 32:88-95.
Kurtén, B. 1957. The bears and hyaenas of the interglacials. Quaternaria, 4:69-81.
Kurtén, B. 1962. The spotted hyena (Crocuta crocuta) from the middle Pleistocene of Mosbach at Wiesbaden, Germany. Commentationes Biologicae, 24:1-9.
Kurtén, K. and Poulianos, A.N. 1977. New stratigraphic data and faunal material from Petralona Cave with special reference to the Carnivora. Anthropos, 4:47-130.
Kurtén, B. and A.N. Poulianos. 1981. Fossil Carnivora of Petralona Cave. Status of 1980. Anthropos, 8:9-56.
Llana Rodríguez, C., Villar Quinteiro, R., and Martínez Cortizas, A. 1996. Secuencia paleoambiental y cultural de la Cueva de A Valiña (Castroverde, Lugo): una ocupación Chatelperroniense en Galicia, p. 97-112. In Pérez Alberti, A. (ed.), Dinámica y Evolución de Medios Cuaternarios, Xunta de Galicia, Santiago de Compostela.
Lloveras, L., Moreno-García, M., Nadal, J., Maroto, J., Soler, J., and Soler, N. 2010. The application of actualistic studies to assess the taphonomic origin of Musterian rabbit accumulations from Arbreda Cave (North-East Iberia). Archaeofauna, 19:99-119.
López-Quintana, J.C., Castaños, P., Guenaga Lizasu, A., Murelaga, X., Areso, P., and Uriz, A. 2005. La cueva de Atxagakoa (Forua, Bizkaia): Ocupación humana y guarida de carnívoros durante el Musteriense en Urdaibai. Illunzar, 5:11-24.
Lyman, R.L. 1994. Vertebrate Taphonomy. Cambridge University Press, Cambridge.
Marean, C.W. and Kim, S.Y. 1998. Mousterian largemammal remains from Kobeh Cave: Behavioral implications for Neanderthals and early modern humans. Current Anthropology, 39:79-113.
Madurell-Malapeira, J., Minwer-Barakat, R., Alba, D.M., Garcés, M., Gómez, M., Aurell-Garrido, J., Ros-Montoya, S., Moyà-Solà, S., and Berástegui, X. 2010. The Vallparadís section (Terrassa, Iberian Peninsula) and the latest Villafranchian faunas of Europe. Quaternary Science Review, 29:3972-3982.
Maíllo Fernández, J.M., Valladas, H., Cabrera Valdés, V., and Bernardo de Quirós, F. 2001. Nuevas dataciones para el Paleolítico superior de Cueva Morín (Villanueva de Villaescusa, Cantabria). Espacio, Tiempo y Forma. Serie I, Prehistoria y Arqueología, 14:145-150.
Maroto, J. 1993. La cueva de los Ermitons (Sales de Llierca, Girona): un yacimiento del Paleolítico medio final. Espacio, Tiempo y Forma. Serie I, Prehistoria y Arqueología, 6:13-30.
Maroto, J. 2014. El conjunto del Reclau Viver, p. 246-257. In Sala Ramos (ed.), Los cazadores recolectores del Pleistoceno y del Holoceno en Iberia y el Estrecho de Gibraltar: Estado actual del conocimiento del registro arqueológico. Universidad de Burgos y Fundación Atapuerca, Burgos.
Maroto, J., Soler, N., and Mir, A. 1987. La cueva de Mollet I (Serinyà, Gerona). Cypsela VI:101-110.
Maroto, J., Vaquero, M., Arrizabalaga, Á., Baena, J., Baquedano, E., Jordá, J., Juliá, R., Montes, R., Van Der Plicht, J., Rasines, P., and Wood, R. 2012. Current issues in late Middle Palaeolithic chronology: New assessments from Northern Iberia. Quaternary International, 247:15-25.
Martínez-Navarro, B. 2010 Early Pleistocene faunas of Eurasia and hominin dispersals, p. 207-224. In Out of Africa I. The First Hominin Colonization of Eurasia Fleagle, J.G., Shea, J.J., Grine, F.E., Baden, A.L., and Leakey, R.E. (eds.),Springer, New York.
Martínez-Sánchez, R., López-García, J.M., Alcalá-Ortiz, A., Blain, H.A., and Rabal-Garcés, R. 2012. Bears and Hyenas from the Latest Pleistocene of Southern Iberia: Sima de Abraham, Priego de Córdoba, Andalusia. Journal of Taphonomy, 10:545-559.
Mills, M.G.L. 1982. Notes on age determination, growth and measurements of brown hyaenas Hyaena brunnea from the Kalahari Gemsbok National Park. Koedoe, 25:55-61.
Morales, J., Soria, D., and Soto, E. 1987. Los carnívoros del Pleistoceno Medio de Atapuerca, p. 135-152 In Aguirre, E., Carbonell, E., and Bermudez de Castro, J.M. (eds.), El hombre fósil de Ibeas y el Pleistoceno de la Sierra de Atapuerca. Junta de Castilla Y Leon, Valladolid.
Moreno, D., Falguères, C., Pérez-González, A., Voinchet, P., Ghaleb, B., Despriée, J., Sala, R., Carbonell, E., Bermúdez de Castro, J.M., and Arsuaga, J.L. 2015. New radiometric dates on the lowest stratigraphical section (TD1 to TD6) of Gran Dolina site (Atapuerca, Spain). Quaternary Geochronology, 30:535-540
Pales, L. and Lambert, C. 1971. Atlas Ostéologique pour Servir à la Identification des Mammifères du Quaternaire. Centre National de la Recherche Scientifique, Bordeaux.
Palmqvist, P., Martínez-Navarro, B., Pérez-Claros, J.A., Torregrosa, V., Figueirido, B., Jiménez-Arenas, J.M., Patrocinio Espinares, M., Ros-Montonya, S., and De Renzi, M. 2011. The giant hyena Pachycrocuta brevirostris : modelling the bone-cracking behavior of an extinct carnivore. Quaternary International, 243:61-79.
Pérez Ripoll, M. 1977. Los mamíferos del yacimiento musteriense de Cova Negra (Játiva, Valencia). Serie de Trabajos Varios, 53.
Quam, R.M., Arsuaga, J.L., Bermúdez de Castro, J.M., Díez, J.C., Lorenzo, C., Carretero, J.M., García, N., and Ortega, AI. 2001. Human remains from Valdegoba cave (Huérmeces, Burgos, Spain). Journal of Human Evolution, 41:385-435.
Rabal-Garcés, R. 2013. Estudio paleontológico de Ursus spelaeus Rosenmüller, 1794 del Pleistoceno superior de Coro Tracito (Tella, Huesca, España). Unpublished PhD Thesis, Universidad de Zaragoza, Zaragoza.
Rabal-Garcés, R., Cuenca-Bescós, G., Canudo, J.I., and Torres, T. 2012. Was the European cave bear an occasional scavenger? Lethaia, 45:96-108.
Raposo, L. 1995. Ambientes, territorios y subsistencia en el Paleolítico Medio de Portugal. Complutum, 6:57-77
Raposo, L. and Cardoso, J.L. 1998. Las industrias líticas de la Gruta Nova de Columbeira (Bombarral, Portugal) en el contexto del Musteriense final de la Península Ibérica. Trabajos de Prehistoria, 55(1):39-62.
Reynolds, S.H. 1902 A monograph of the British Pleistocene Mammalia Part II. The Cave Hyena. Palaeontological Society Monographs, 2:1-25
Riquelme Cantal, J.A. and Carrión García, J.S. 2010. La Cueva de las Ventanas, Píñar, (Granada): reconstrucción arqueológica de un cubil de hiena tardiglaciar en el sur de la Península Ibérica. Zona Arqueológica, 13:345-351.
Roche, J. 1964. Le Paléolithique supérieur portugais. Bilan de nos connaissances et problèmes. Bulletin de la Société Préhistorique Française. Études et Travaux, 61:11-27.
Rodríguez Vidal, J., Cáceres, L.M., Alvarez, G., Martínez Aguirre, A., Alcaraz, J.M., Riquelme, J.A., and Recio, J.M. 2003. Secuencia cronoestratigráfica del Pleistoceno Superior en Sierra Morena.: la cueva de La Sima (Constantina, Sevilla). Geocaceta, 33:47-50.
Rodríguez Vidal, J., Giles Pacheco, F., and Riquelme Cantal, J.M. 2010. El registro fósil de hienas en las cuevas de Gorham’s y Vanguard (Gibraltar): contexto paleogeográfico. Zona Arqueológica, 13:354-364.
Rosell, J., Blasco, R., Fernández-Laso, M.C., Vaquero, M., and Carbonell, E. 2012. Connecting areas: Faunal refits as a diagnostic element to identify synchronicity in the Abric Romaní archaeological assemblages. Quaternary International, 252:56-67.
Rosell, J., Blasco, R., Morales, J.I., Rivals, F., Rodríguez-Hidalgo, A., Cebrià, A., and Camarós, E. 2012. Compartint l’espai: la interacció entre homínids i carnívors als nordest peninsular (Cova del Toll i Cova de les Teixoneres, Moià, Bages), I Actes d’Arqueologia de la Catalunya Central. Homenatge a Miquel Cura, 47-51.
Rosell, J., Blasco, R., Rivals, F., Camarós, E., Arilla, M., Fàbregas, J., Rufà, A., and Sánchez C. 2014. Amagant jes proves: superposicions de l’activitat de carnívors i Neandertals a la Cova de les Teixoneres (Moià, Bages). Actes II Jornades d’Arqueologia de la Catalunya Central, 2012:74-77.
Rosell, J., Blasco, R., Rivals, F., Cebrià, A., Morales, J.I., Rodríguez, A., Serrat, D., and Carbonell, E. 2010. Las ocupaciones en la Cova de les Teixoneres (Moià, Barcelona): relaciones espaciales y grado de competencia entre hienas, osos y neandertales durante el Pleistoceno superior. Zona Arqueológica, 13:393-402.
Rohland, N., Pollack, J.L., Nagel, D., Beavual, C., Airvaux, J., Pääbo, S., and Hofreiter, M. 2005. The population history of extant and extinct hyenas. Molecular Biology Evolution, 22:2435-2443.
Rueda i Torres, J.M. 1993. L’Acció antrópica sobre les matèries dures animals durant el Plistocè del Nord-Est de Catalunya. Unpublished PhD Thesis, Universitat de Girona, Girona.
Sala, M.T.N., Arsuaga, J.L., Laplana, C., Ruiz, B., Gil, M.J., García, N., Aranburu, A., and Algaba, M. 2011. Un paisaje de la Meseta durante el Pleistoceno Superior. Aspectos paleontológicos de la Cueva de la Zarzamora (Segovia, España). Boletín de la Real Sociedad Española de Historia Natural. Sección Geológica, 105:67-85.
Sala, N. 2012. Tafonomía de yacimientos kársticos de carnívoros en el Pleistoceno. Unpublished PhD Thesis, Universidad Complutense de Madrid, Madrid.
Saladié, P. 2009. Mossegades d’omnívors. Aproximació experimental i aplicación zooarqueològica als jaciments de la Sierra de Atapuerca. Unpublished PhD Thesis, Universitat Rovira i Virgili, Tarragona.
Saladié, P., Huguet, R., Díez, C., Rodríguez‐Hidalgo, A., and Carbonell, E. 2013. Taphonomic modifications produced by modern brown bears (Ursus arctos). International Journal of Osteoarchaeology, 23(1):13-33.
Samper Carro, S.C. and Martínez-Moreno, J. 2014. Who let the hyenas out? Taphonomic analysis of the faunal assemblage from GL-1 of Cova del Gegant (Sitges, Spain). Quaternary International, 330:19-35.
Sánchez, A., Fraile, A., van der Made, J., Morales, J., Quiralte, V., Salesa, M.J., Sánchez, I.M., Sanchiz, B., Soria, D., Jiménez, J., Barbadillo, L.J., Laplana, C., and Szyndlar, Z. 2005. Primeros datos faunísticos del Neolítico madrileño: la cueva de La Ventana (Torrelaguna, Madrid). In Arias Cabal, P., Ontañón Peredo, R., and García-Moncó Piñeiro, C. (eds.), III Congreso del Neolítico en la Península Ibérica. Monografias del Instituto Internacional de Investigaciones Prehistóricas de Cantabria 1, Universidad de Cantabria, Santander, 155-165.
Sanz, A., Sala, N., Algaba, M., and Arsuaga, J.L. 2015. El Portalón del Tejadilla, nueva ventana al Pleistoceno superior de la Meseta Castellana. Libro de resúmenes de la XXI Bienal de la Real Sociedad Española de Historia Natural, Burgos, 28-29.
Sardella, R. and Petrucci, M. 2012. The earliest Middle Pleistocene Crocuta crocuta (Erxleben, 1777) at Casal Selce (Rome, Italy). Quaternary International, 267:103-110.
Sarrión, I. 1983. La fauna würmiense de la Cueva de San Antón (Villanueva de Viver, Castellón). Spélaion, 2:23-37.
Sarrión, I. 1990. El yacimiento del Pleistoceno medio de la Cova del Corb (Ondara, Alicante). Archivo de Prehistoria Levantina, XX:43-77.
Sarrión, I. 2010. El cáprido del yacimiento de la Cova del Molí Mató, Agres, Alacant. Recerques del Museu d’Alcoi, 19:7-18.
Sauqué, V., Galán, J., Nuñez-Lahuerta, C., Sola-Almagro, C., Rabal-Garcés, R., Martínez, I., Mazo, C., and Cuenca-Bescós, G. 2014. Neandertales en el Pleistoceno Superior del valle del Ebro, paeloambiente y evidencias tafonómicas en Aguilón P-7. In Royo-Torres, R., Verdú, F.J., and Alcalá, L. (coord.), XXX Jornadas de Paleontología de la Sociedad Española de Paleontología. Fundamental, 24:223-226.
Sauqué, V., García-González, R., and Cuenca-Bescós, G. 2016a. A Late Pleistocene (MIS3) ungulate mammal assemblage (Los Rincones, Zaragoza, Spain) in the Eurosiberian-Mediterranean boundary. Historical Biology, 28(3):358-389.
Sauqué, V., Rabal-Garcés, R., and Cuenca-Bescós, G. 2016b. Carnivores from los Rincones, a leopard den in the highest mountain of the Iberian range (Moncayo, Zaragoza, Spain) Historical Biology, 28(4):479-506.
Sauqué, V., Rabal-Garcés, R., Sola-Almagro, C., and Cuenca-Bescós. 2014. Bone accumulation by leopards in the Late Pleistocene in the Moncayo massif (Zaragoza, NE Spain). PLoS One, 9(3):e92144.
Schinz, R. 1838. Descripción de Capra pyrenaica. Nouveaux Memoires de la Societe Helvetique d’Histoire Naturell, Neuchatel, Swiss.
Schütt, G. 1971. Die Hyänen der Mosbacher Sande (Altpleistozän, Wiesbaden/Hessen) mit einem Beitrag zur Stammesgeschichte der Gattung Crocuta. Mainzer Naturwissenschaftliches Archiv, 10:29-76.
Selvaggio, M.M. 1994. Identifying the timing and sequence of hominid and carnivore involvement with Plio-Pleistocene bone assemblages from carnivore tooth marks and stone-tool butchery marks on bone surfaces. PhD Thesis, Rutgers University, New Jersey.
Selvaggio, M.M. and Wilder, J. 2001. Identifying the involvement of multiple carnivore taxa with archaeological bone assemblages. Journal of Archaeological Science, 28:465-470.
Senosiáin, M.A., Arsuaga, J.L., and Álvarez, G.E.A. 2005. La cueva del Forno/Conde (Tuñón, Asturias): un yacimiento del tránsito del Paleolítico medio y superior en la Cornisa Cantábrica, p. 425-441. In Montes Barquín, R. and Lasheras Corruchaga, J.A. (eds.), Neandertales cantábricos, estado de la cuestión : Subdirección General de Publicaciones, Información y Documentación Monografías del Museo Nacional de Altamira, Santander.
Sesé, C., Alberdi, M.T., Mazo, A., and Morales, J. 2001. Mamíferos del Mioceno, Plioceno y Pleistoceno de la Cuenca de Guadix-Baza (Granada, España): revisión de las asociaciones faunísticas más características. Paleontologia i Evolució, 32-33:31-36.
Sesé, C. and Soto, E. 2009. Aportación del Museo Nacional de Ciencias Naturales al conocimiento de los mamíferos del Cuaternario de España. In Lobón-Cerviá, J. and Morales, J. (eds.), Notas para la historia reciente del Museo Nacional de Ciencias Naturales. Homenaje a María Dolores Soria Mayor, Consejo Superior de Investigaciones Científica, Madrid, 153-210.
Sheng, G.L., Soubrier, J., Liu, J.Y., Werdelin, L., Llamas, B., Thomson, V.A., Tuke, J., Wu, L.J., Hou, X.D., Chen, Q.J., Lai, X.L., and Cooper, A. 2014. Pleistocene Chinese cave hyenas and the recent Eurasian history of the spotted hyena, Crocuta crocuta. Molecular Ecology, 23:522-533.
Soergel, W.1937. Die Stellung der Hyaena spelaea GOLDF. aus der Lindentaler Hyänenhöhle bei Gera. Beiträge zur Geologie von Thüringen, 4:171-189.
Spoor, C.F. 1985. Body proportions in Hyaenidae. Anatomischer Anzeiger, 160:215-220.
Stringer, C., Finlayson, J.C., Barton, R.N.E., Fernández-Jalvo, Y., Cáceres, I., Sabin, RC., Rhodes, E.J., Currant, A.P., Rodríguez-Vidal, J., Giles-Pacheco, F., and Riquelme-Cantal, J.A. 2008. Neanderthal exploitation of marine mammals in Gibraltar. Proceedings of the National Academy of Sciences of the United States of America, 105 (38):14319-14324.
Stuart, A.J. and Lister, A.M. 2014. New radiocarbon evidence on the extirpation of the spotted hyaena (Crocuta crocuta (Erxl.) in northern Eurasia. Quaternary Science Review, 96:108-116.
Testu, A. 2006. Etude paleontologique et biostratigraphique des Felidae et Hyaenidae pleistocenes de l'Europe mediterraneenne. Unpublished PhD Thesis, Universite de Perpignan, Perpignan.
Tiffagom, M. and Sanchis Sierra, A. 2008. Excavaciones arqueológicas en la Cova de l’Assut de Bellús. Primeras valoraciones. Archivo de Prehistoria Levantina, 27:371-380.
Torrente Casado, R. 2010. Aportes al Estudio de las Primeras Ocupaciones Humanas en el Sur de la Península Ibérica. El caso de Cúllar-Baza 1. Arqueología y Territorio, 7:1-19.
Torres, T. 2013. La Historia del oso de las Cavernas: Vida y Muerte de un Animal Desaparecido. Escuela Técnica Superior de Ingenieros de Minas, Madrid.
Turner, A. 1984. The interpretation of variation in fossil specimens of spotted hyaena (Crocuta crocuta Erxleben, 1777) from Sterkfontein Valley sites (Mammalia: Carnivora). Annals of the Transvaal Museum, 33:399-418.
Turner, A. and Antón, M. 1996. The giant hyena, Pachycrocuta brevirostris (Mammalia, Carnivora, Hyaenidae). Geobios, 29:455-468.
Turner, A., Antón, M., and Werdelin, L. 2008. Taxonomy and evolutionary patterns in the fossil Hyaenidae of Europe. Geobios, 41:677-687.
Utrilla, P., Montes, L., Blasco, F., Torres, T., and Ortiz, J.E. 2010. La cueva de Gabasa revisada 15 años después: un cubil para las hienas y un cazadero para los Neandertales. Zona Arqueológica, 13:377-389.
Valente, M.J. 2004. Humans and Carnivores in the Early Upper Paleolithic in Portugal: Data from Pego do Diabo Cave . Revue de Paléobiologie, 23(2):611-626.
Van Valkenburgh, B. 1990. Skeletal and dental predictors of body mass in carnivores, p. 181-205. In Damuth, J. and MacFadden, B.J. (eds.), Body Size in Mammalian Paleobiology: Estimation and Biological Implications. Cambridge University Press, Cambridge.
Varela, S. 2011. Aplicación de los modelos de distribución de especies hacia el Pasado. Análisis de la distribución y extinción las poblaciones europeas de hiena manchada ( Crocuta crocuta (Erxleben, 1777)) durante el Pleistoceno. Unpublished PhD Thesis, Universidad Autonoma de Madrid, Madrid.
Varela, S., Lobo, J.M., Rodríguez, J., and Batra, P. 2010. Were the Late Pleistocene climatic changes responsible for the disappearance of the European spotted hyena populations? Hindcasting a species geographic distribution across time. Quaternary Science Review, 29:2027-2035.
Villa, P., Goni, M.F.S., Cuenca-Bescós, G., Grün, R., Ajas, A., Pimienta, J.C.G., and Lees, W. 2010. The archaeology and paleoenvironment of an Upper Pleistocene hyena den: an integrated approach. Journal of Archaeological Science, 37:919-935.
Villa, P. and Mahieu, E. 1991. Breakage patterns of human long bones. Journal of Human Evolution, 21:27-48.
Villalta, J.F. 1972. Presencia de la Marmota y otros elementos de la fauna esteparia en el Pleistoceno catalán. Acta Geológica Hispánica, VII(6):170-173.
Vinuesa, V., Iurino, D.A., Madurell-Malapeira, J., Liu, J., Fortuny, J., Sardella, R., and Alba, D.M. 2015. Inferences of social behavior in bone-cracking hyaenids (Carnivora, Hyaenidae) based on digital paleoneurological techniques: Implications for human-carnivoran interactions in the Pleistocene. Quaternary International. doi:10.1016/j.quaint.2015.10.037
von den Driesch, A. 1976. A Guide to the Measurement of Animal Bones from Archaeological Sites. Peabody Museum Press, Harvard.
Walker, R. 1985. A Guide to Post-Cranial Bones of East African Animals. Hylochoerus Press, Norwich.
Werdelin, L. and Solounias, N. 1991. The Hyaenidae: Taxonomy, systematic, and evolution. Fossils and Strata, 30:1-104.
Yravedra, J. 2003. Estado de la cuestión sobre la subsistencia del musteriense en el interior y la fachada de la Península Ibérica. Zephyrus, 56:61-84
Yravedra, J. 2006. Acumulaciones biológicas en yacimientos arqueológicos: Amalda VII y Esquilleu III-IV. Trabajos de Prehistoria, 62:55-78.
Yravedra, J. 2007. Aproximaciones tafonómicas a los cazadores de la segunda mitad del Pleistoceno Superior de la mitad Norte del interior de la Península Ibérica. Arqueoweb. Revista Sobre Arqueología en Internet, 9(1). http://pendientedemigracion.ucm.es/info/arqueoweb/pdf/9-2
Yravedra, J. and Gómez Castanedo, A. 2010. Estudio zooarqueológico y tafonómico del yacimiento del Otero (Secadura, Voto, Cantabria). Espacio Tiempo y Forma. Serie I, Prehistoria y Arqueología, 3:21-38.
Yravedra, J., Gómez Castanedo, A., and Muñoz Fernández, E. 2010. Estrategias de subsistencia en el Yacimiento Paleolítico del Ruso (Igollo de Camargo, Cantabria, España). Espacio, Tiempo y Forma. Serie I, Prehistoria y Arqueología , 3:39-58.
