

Article Search
Volume 27.2
May–August 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.2 May-August 2024
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
 J. Michael Hils
J. Michael Hils
Department of Geological Sciences
Ohio University
316 Clippinger Laboratories
Athens, Ohio 45701
USA
jhils2@udayton.edu
Michael Hils completed his Master's degree in geology at Ohio University in 2014 after earning his B.S. with a double major in geology and biology from the University of Dayton. He currently teaches at University of Dayton.

 Daniel I. Hembree
Daniel I. Hembree
Department of Geological Sciences
Ohio University
316 Clippinger Laboratories
Athens, Ohio 45701
USA
hembree@ohio.edu
Daniel Hembree is an Associate Professor in the Department of Geological Sciences at Ohio University. His research interests primarily lie with animal-substrate interactions in ancient and modern continental environments. These interactions are preserved in the fossil record as trace fossils. Trace fossils provide an in situ record of ancient biodiversity, ecology, and environment. The study of trace fossils, therefore, provides vital information for accurate paleoenvironmental reconstructions. This involves not only the study of paleosols and continental trace fossils throughout geologic time, but also the experimental study of burrowing behaviors of extant terrestrial annelids, arthropods, amphibians, and reptiles. Current research projects involve the study of the influence of climate changes on ancient soils and soil ecosystems including those of the Pennsylvanian and Permian of southeast Ohio, Permian of eastern Kansas, and the Eocene to Miocene of Colorado and Wyoming.
APPENDIX 1.
Video of Hogna lenta using compaction burrowing technique. Click on image to run movie.

APPENDIX 2.
Video of Hogna lenta depositing silk on compacted sediment and glass of the terrarium. Click on image to run movie.

APPENDIX 3.
Video of Hogna lenta gathering sediment with silk to form a trapdoor. Click on image to run movie.

APPENDIX 4.
Video of Gorgyrella inermis using silk to gather material into a bundle. Click on image to run movie.

APPENDIX 5.
Video of Gorgyrella inermis gathering silken bundle of sediment and removing it from the burrow. Click on image to run movie.

APPENDIX 6.
Video of Gorgyrella inermis depositing silk along the sides of its burrow. Click on image to run movie.

APPENDIX 7.
Quantitative data for the burrows produced by Aphonopelma chalcodes. All measurements in cm. SR = subvertical ramp.
APPENDIX 8.
Bray-Curtis result matrix for Aphonopelma chalcodes. This matrix displays the comparison of burrows produced by Gorgyrella inermis, Hogna lenta, and A. chalcodes . Cells are filled in black where burrows are compared to themselves. Cells with scores of high (1.0-0.8), moderate (0.7-0.6) to low (0.5-0.3) similarity are filled in blue with increasing lightness. See Table 1 and Appendix 7 for architecture key. (PDF only)
APPENDIX 9.
Quantitative data for the burrows produced by Hadrurus arizonensis. All measurements in cm. SR = subvertical ramp, HB = helical burrow, MW = mazework, UB = U-shaped burrow.
APPENDIX 10.
Bray-Curtis result matrix for Hadrurus arizonensis,Gorgyrella inermis, and Hogna lenta. Formatting is the same as Appendix 8. See Table 1 and Appendix 9 for architecture key. (PDF only)
APPENDIX 11.
Quantitative data for the burrows produced by Mastigoproctus giganteus. All measurements in cm. SR = subvertical ramp, MW = mazework, UB = U-shaped burrow, VS = vertical shaft.
APPENDIX 12.
Bray-Curtis result matrix for Mastigoproctus giganteus , Gorgyrella inermis, and Hogna lenta. Formatting is the same as Appendix 8. See Table 1 and Appendix 11 for architecture key. (PDF only)
APPENDIX 13.
Quantitative data for the burrows produced by Narceus americanus. All measurements in cm unless otherwise noted. SV = subvertical burrow, VS = vertical shaft, HB = helical burrow, OS = O-shaped burrow.
APPENDIX 14.
Bray-Curtis result matrix for Narceus americanus, Gorgyrella inermis, and Hogna lenta. Formatting is the same as Appendix 8. See Table 1 and Appendix 13 for architecture key. (PDF only)
TABLE 1. Experimental design and burrows produced. Sediment density is given in kgf/cm2. 1, Experiment 1; 2, Experiment 2; 3, Experiment 3. Sediment moisture values are in percent total volume. VS: vertical shaft, VC: vertical shaft with terminal chamber, SS: subvertical shaft, SC: subvertical shaft with terminal chamber, JS: J-shaped burrow, YS: Y-shaped burrow, IC: isolated chamber.
TABLE 2. Qualitative data for each burrow architecture. Burrows are arranged by increasing complexity. That distribution architectures produced by each species is provided, as well as the sum of all burrow architectures developed. (PDF only)
TABLE 3. Quantitative measurements of vertical shafts produced by Gorgyrella inermis and Hogna lenta. (PDF only)
TABLE 4. Quantitative measurements of subvertical shafts produced by Gorgyrella inermis and Hogna lenta.
TABLE 5. Quantitative measurements of other burrow architectures produced by Gorgyrella inermis and Hogna lenta. See Table 1 for architecture key.
TABLE 6. Mann-Whitney and Kolmogorov-Smirnov test results ( p values) from the comparison of the burrows produced by arthropods considered in this study. 1) Gorgyrella and Hogna, 2) Spiders and Aphonopelma, 3) Spiders and Hadrurus, 4) Spiders and Mastigoproctus, 5) Spiders and Narceus. Boxed values indicate statistical difference ( p <0.05). All measurements are in cm, and all angles are in degrees.
TABLE 7. Matrix of Bray-Curtis test results for the comparison of Gorgyrella inermis and Hogna lenta burrows to G. inermis burrows. Cells are filled in black where burrows are compared to themselves. Cells with scores of high to moderate similarity (1.0-0.7) are filled in blue with increasing lightness. See Table 1 for burrow architecture key. (PDF only)
TABLE 8. Matrix of Bray-Curtis test results for the comparison of Gorgyrella inermis and Hogna lenta burrow to H. lenta burrows. Cells are filled in black where burrows are compared to themselves. Cells with scores of high to moderate similarity (1.0-0.7) are filled in blue with increasing lightness. See Table 1 for burrow architecture key. (PDF only)
TABLE 9. Mann-Whitney and Kolmogorov-Smirnov test results ( p values) from the comparison of clusters in each dendrogram. 1) Gorgyrella and Hogna (Figure 18), 2) Spiders and Aphonopelma (Figure 20), 3) Spiders and Hadrurus (Figure 21), 4) Spiders and Mastigoproctus (Figure 22), 5) Spiders and Narceus (Figure 23), 6) All arthropods (Figure 24). Boxed values indicate statistical difference ( p <0.05). All measurements are in cm, and all angles are in degrees.
TABLE 10. Mann-Whitney and Kolmogorov-Smirnov test results (p values) from the comparison of the properties of Gorgyrella and Hogna burrows produced under different experimental conditions including terrarium size, sediment density, and sediment moisture. No results indicated significant differences (p <0.05).
TABLE 11. Mann-Whitney and Kolmogorov-Smirnov test results (p values) between burrows produced by Hogna lenta individuals with a brood and those without a brood. No results indicated significant differences ( p <0.05).
FIGURE 1. Basic spider morphology. Modified from Dipenaar-Schoeman (2002).
FIGURE 2. Representatives of spiders used in this study. 1, Adult Gorgyrella inermis. 2, Juvenile G. inermis. 3, Hogna lenta.

FIGURE 3. Quantitative properties of burrows. 1, Measurements of burrows included maximum depth (D), slope (OA), total length (L), branching angle (BA), and tunnel and shaft width (w), height (h), and circumference (c). Width is defined as parallel to the hinge-line of the burrow trapdoor; height is defined as perpendicular to the hinge-line of the burrow trapdoor. 2, Complexity (C) is the sum of the number of segments (s), chambers (h), and surface openings (e) of a single burrow system. 3, Tortuosity is a measure of the average sinuosity of all of the segments of a burrow system. The tortuosity of a single segment is found by dividing the total length (u) by the straight-line distance (v). Modified from Hembree et al., 2012.
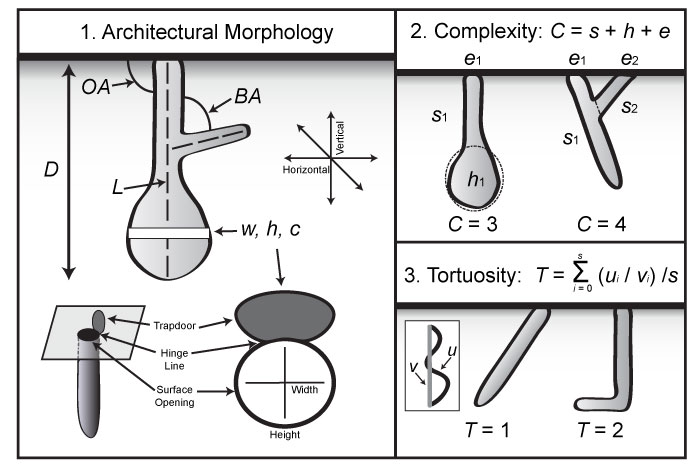
FIGURE 4. Representatives of other laboratory animals used for burrow comparisons. 1, A female Mexican blond tarantula (Aphonopelma chalcodes), 2, An Arizona hairy desert scorpion (Hadrurus arizonensis), 3, A whip scorpion (Mastigoproctus giganteus), 4, A giant American millipede (Narceus americanus).

FIGURE 5. Burrow production by Hogna lenta. 1-2, Spider pushes headfirst into the sediment until it is nearly covered. 3, Spider turns around inside the chamber to face outward. 4, Spider applies silk to the sediment and moves it around. 5, Spider covers itself in silk-lined sediment. 6, Spinnerets gathering sediment together to form a trapdoor. The sediment surrounding the spider and its burrow has been lightened.

FIGURE 6. Brooding behavior of Hogna lenta. 1, An egg sac. 2, H. lenta with offspring. 3, A vertical shaft (H5) that served as a brood burrow.

FIGURE 7. Burrow production by Gorgyrella inermis. 1, Spider excavates a small depression in the sediment. 2, Spider deposits silk in sediment at the base of the burrow, 3-4, Spider turns around inside of burrow and picks up bundles of sediment. 5, The spider throws the sediment from the burrow. 6, Spider deposits silk along the burrow wall.

FIGURE 8. Surface features produced by Gorgyrella inermis. 1, Sediment forcefully removed from the burrow by the spider throwing packets with its chelicerae. 2, Trapdoor held open at a burrow entrance. 3, A silk mat produced by a spider that did not construct a burrow. 4, A cocoon built by a spider that did not burrow. The cocoon did not extend into the sediment.

FIGURE 9. Surface features produced by Hogna lenta. 1, Open trapdoor at the burrow entrance. 2, Turret produced by building a burrow entrance against the sides of the terrarium. 3, Silk strands deposited along the sides of the terrarium.

FIGURE 10. Striations on sides of burrow casts resulting from scratch marks in the silk linings of spider burrows. 1, Burrow produced by juvenile Gorgyrella inermis. 2, Burrow produced by adult G. inermis. 3-4, Burrows produced by Hogna lenta. The surfaces of H. lenta burrow casts include sediment, resulting from the thin layer of silk that lined these burrows.
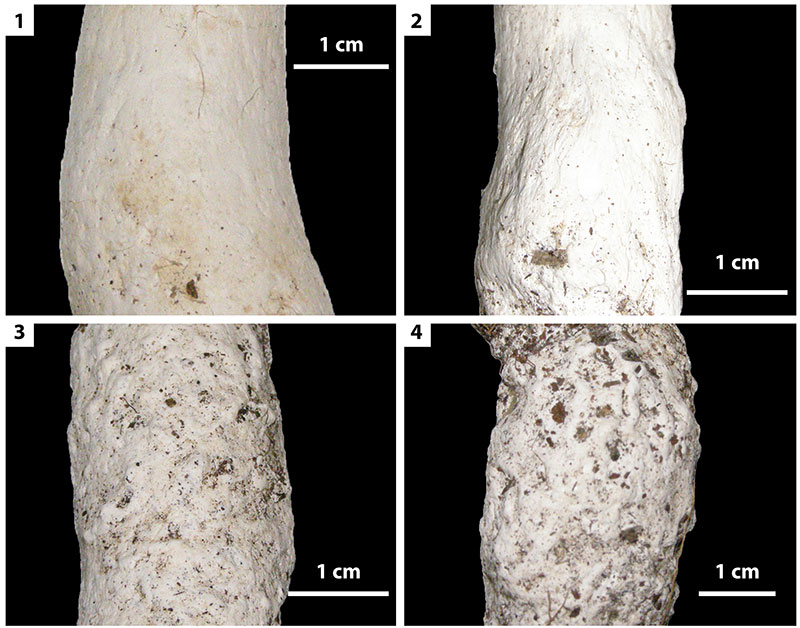
FIGURE 11. Changes in surficial features due to increasing stability of sediment. The spiders used less silk as sediment becomes more stable. This resulted in burrow casts with rougher sides than those with more silk. The first burrow was constructed in sediment composed of 50% silt-clay and had a thick silk lining. The third burrow was constructed in 100% silt-clay and had almost no silk lining.
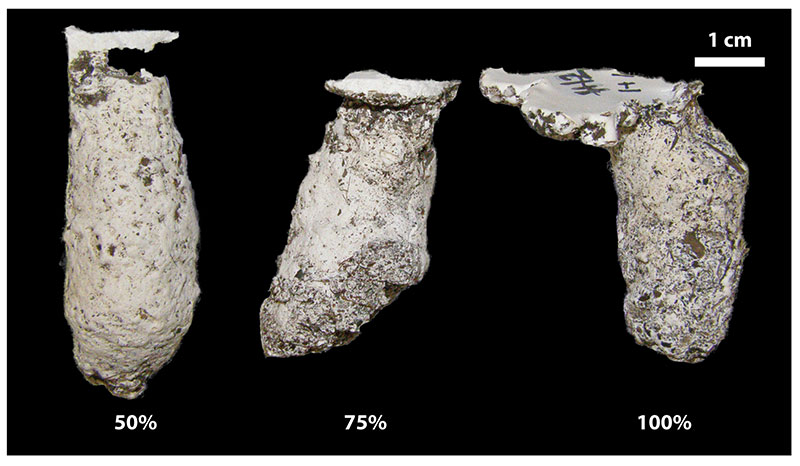
FIGURE 12. Vertical shafts. 1, Burrow produced by an adult Gorgyrella inermis (G9). 2, Burrow produced by Hogna lenta. This burrow served as a brood burrow in addition to a dwelling structure (H5). 3, Burrow produced by juvenile G. inermis (G15). 4, Burrow produced by H. lenta (H7).

FIGURE 13. Subvertical shafts. 1, Burrow produced by adult Gorgyrella inermis (G24). 2, Burrow produced by adult G. inermis (G5). 3, Burrow produced by Hogna lenta (H14). 4, Burrow produced by H. lenta (H16).

FIGURE 14. Vertical shafts with terminal chambers. 1, Burrow produced by juvenile Gorgyrella inermis (G2). 2, Burrow produced by adult G. inermis (G17). 3, Burrow produced by adult G. inermis (G16). 4, Burrow produced by Hogna lenta (H4).
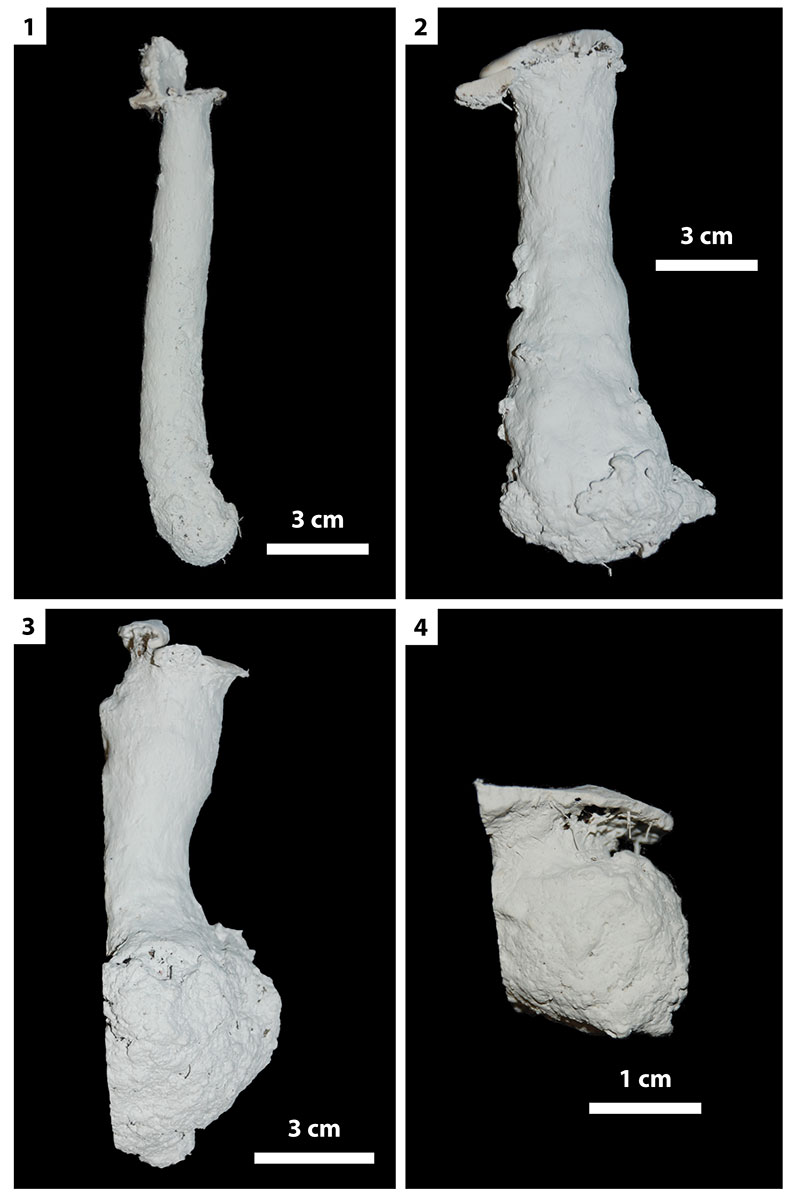
FIGURE 15. Other burrow architectures. 1, Y-shaped burrow produced by Hogna lenta (H4). 2, Isolated chamber produced by adult Gorgyrella inermis (G6). 3, Subvertical shaft with terminal chamber produced by adult G. inermis (G27). 4, J-shaped burrow produced by juvenile G. inermis (G23).
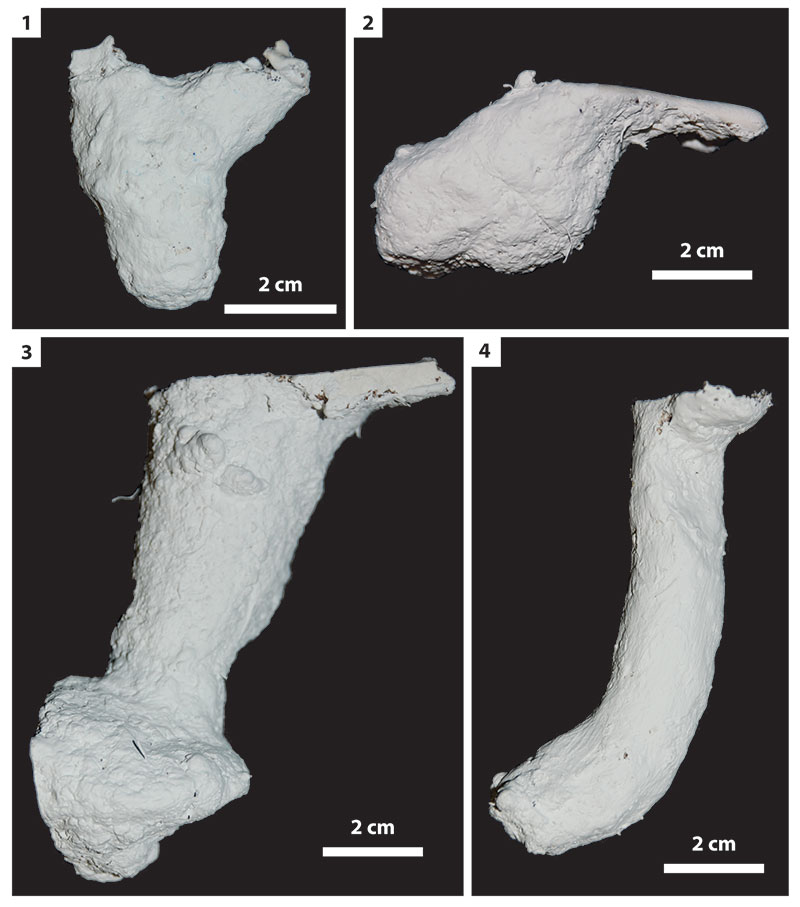
FIGURE 16. Cluster analysis of all burrows produced by Gorgyrella inermis in the study. Cluster A contains burrows with a vertical component. Cluster B contains burrows with a subvertical component and two burrows with vertical components. Values located at the nodes of the dendrogram are the mean Bray-Curtis similarity score of all burrows emanating from that node cluster. Brackets above the dendrogram group together main clusters and provide the mean Bray-Curtis similarity score of the burrows in that cluster. Clusters indicated by letters in yellow circles are clusters of spider burrows that form in all analyses. Burrow architecture is indicated in the key at the bottom of the figure; see Table 1 for key. Tracemaker is indicated by the color of the box with the associated name.
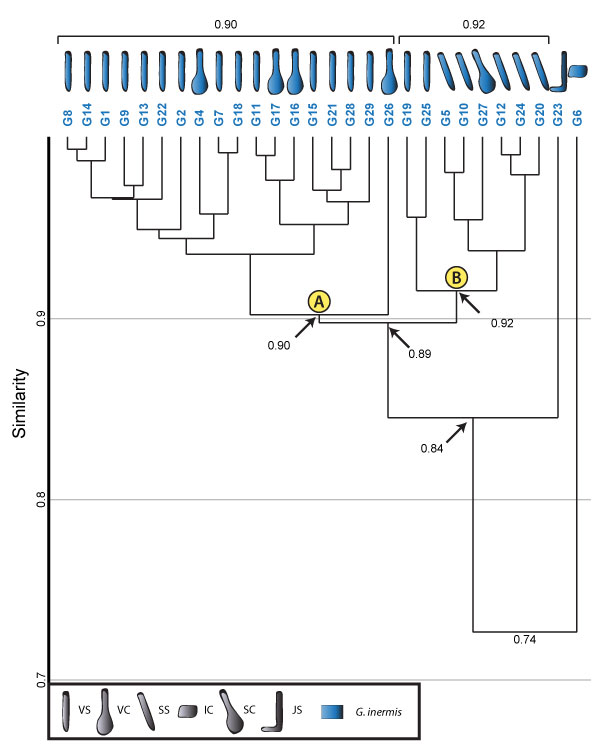
FIGURE 17. Cluster analysis of all burrows produced by Hogna lenta in the study. Cluster A consists mainly of burrows with a vertical component. Cluster B consists of burrows with a subvertical component. Formatting is the same as in Figure 16.
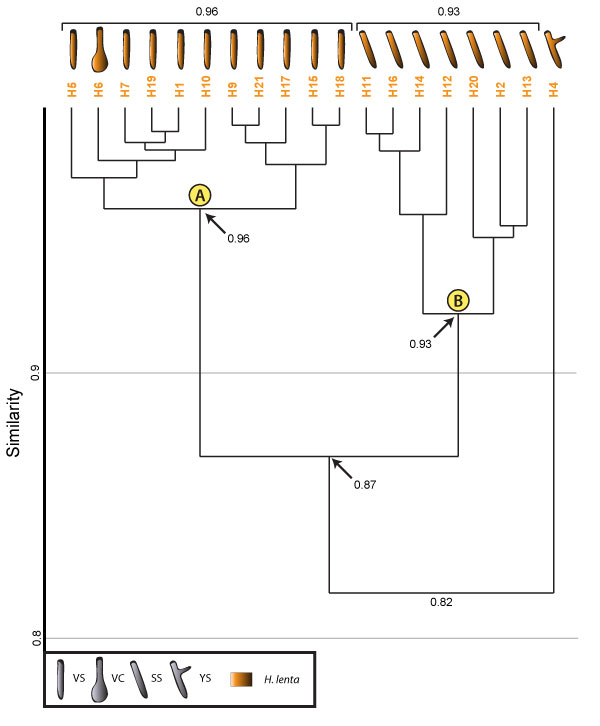
FIGURE 18. Cluster analysis of all burrows produced by Gorgyrella inermis and Hogna lenta. Clusters A (vertical) and B (subvertical) are maintained, but an additional cluster, C, forms. Cluster C contains the J- and Y-shaped burrows. The isolated chamber is not included in any cluster. Formatting is the same as in Figure 16.
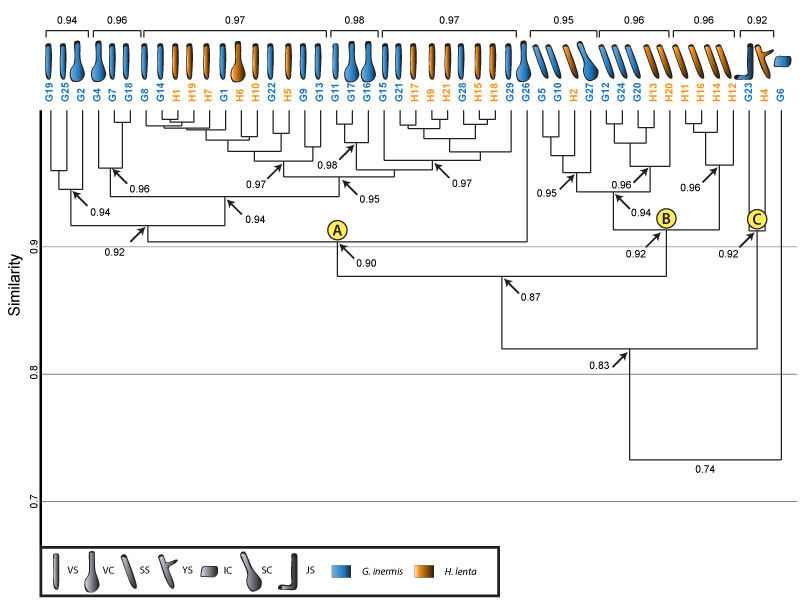
FIGURE 19. Combination of Bray-Curtis matrix and cluster analysis to explain how the cluster analysis groups burrow into the dendrogram. The burrows in the matrix were rearranged into the order (from left to right) produced by the dendrogram. Clusters, numbered 1-9, were matched with groupings in the matrix. The groupings in the matrix are highlighted with a white box. This box contains all of the similarity values contained within that cluster. All similarity values within a grouping are either 1.0 (identical) or 0.9 (highly similar). The separation between Clusters A and B is seen between grouping 5 and grouping 6. The cells above and the cells to the left of grouping 5 (similarity scores contained in Cluster A) have similarity values of 1.0 and 0.9, whereas cells below and cells to the right of grouping 5 (similarity scores contained in Cluster B) have similarity values of 0.9 and 0.8. The small range of similarity scores (1.0-0.8) when comparing both clusters explains why the node containing both clusters has a similarity score of 0.87. See Table 1 for architecture key.
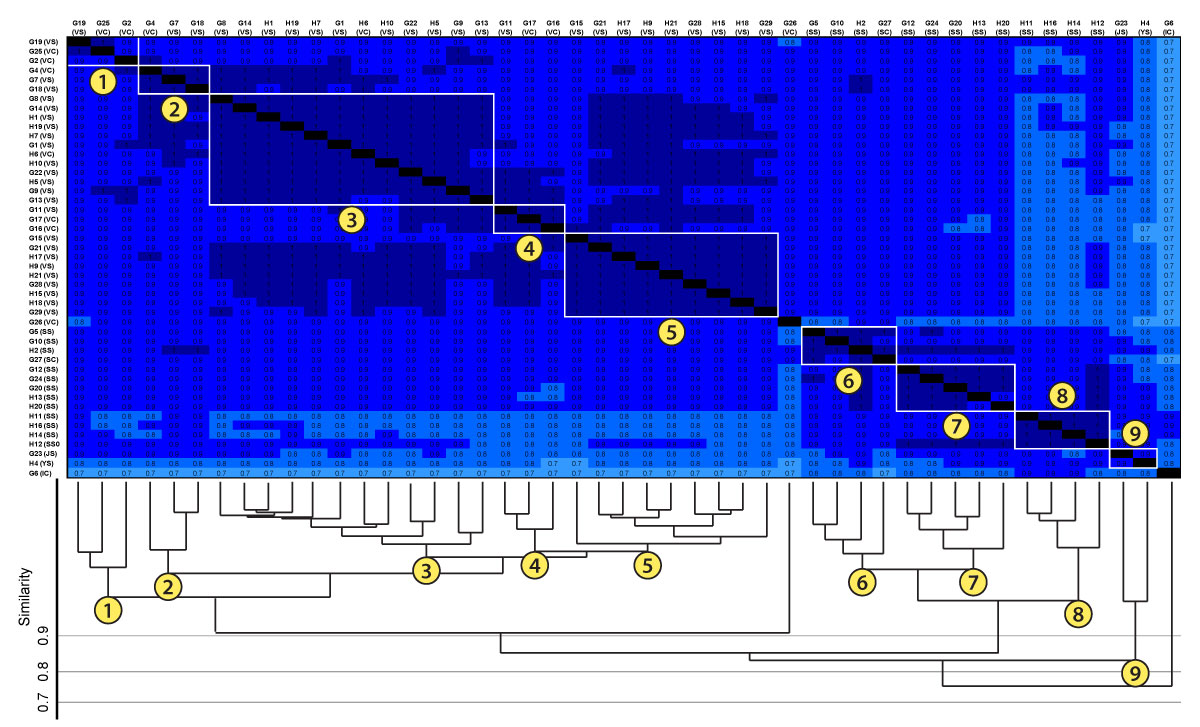
FIGURE 20. Cluster analysis of burrows produced by Aphonopelma chalcodes, Gorgyrella inermis , and Hogna lenta. Clusters A, B, and C are maintained, and the burrows G6 and A2 form a new cluster. The larger A. chalcodes burrow, A1, does not fall into a cluster. Formatting is the same as in Figure 16. Clusters indicated by letters in blue circles form only in this analysis. The subvertical ramp (SR) is an architecture unique to A. chalcodes.
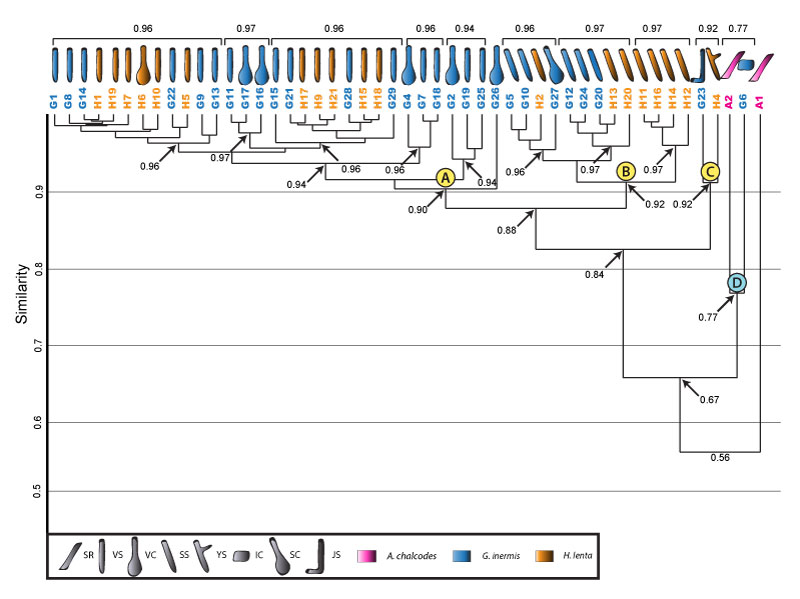
FIGURE 21. Cluster analysis of burrows produced by Hadrurus arizonensis, Gorgyrella inermis, and Hogna lenta. Spider Clusters A, B, and C are maintained, and a separate scorpion cluster is produced. Formatting and key is the same as in Figures 16 and 21. Mazeworks (MW), U-shaped burrows (US), subvertical ramps (SR), and helical burrows (HB) are produced H. arizonensis.
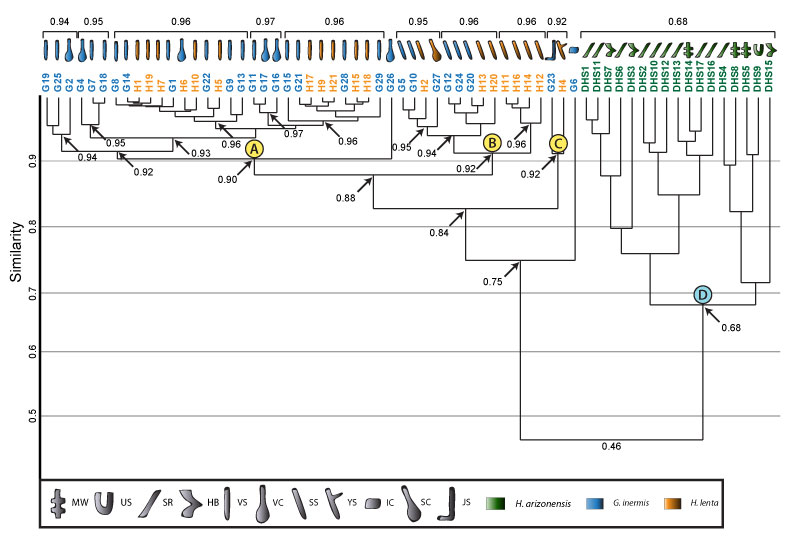
FIGURE 22. Cluster analysis of burrows produced by Mastigoproctus giganteus, Gorgyrella inermis, and Hogna lenta. Spider Clusters A, B, and C are maintained with the addition of morphologically similar whip scorpion burrows, and G6 forms a new cluster. Unique burrow architectures produced by M. giganteus form a separate cluster. Formatting and key is the same as in Figures 16 and 21. Mazeworks (MW), U-shaped burrows (US), and subvertical ramps (SR) are produced only by M. giganteus.
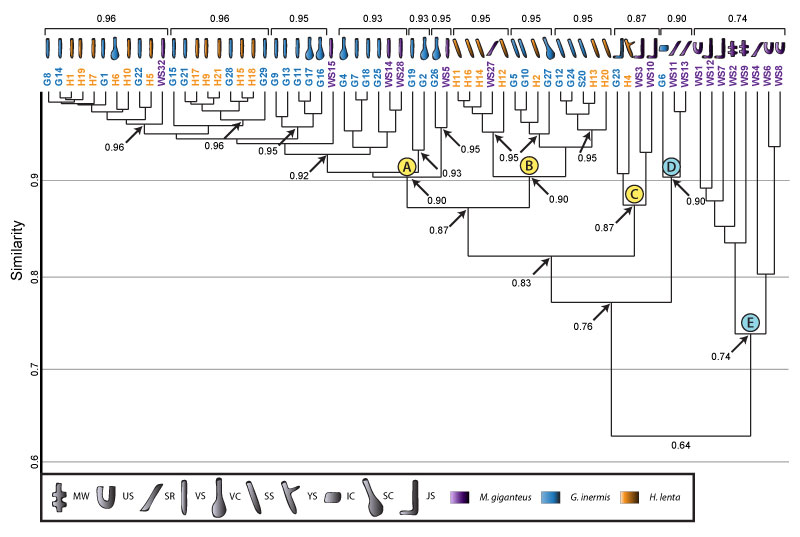
FIGURE 23. Cluster analysis of burrows produced by Narceus americanus, Gorgyrella inermis, and Hogna lenta. Spider Clusters A, B, and C are maintained and include burrows produced by all three species. G6 forms a new cluster with the most dissimilar burrows produced by N. americanus. Formatting and key is the same as in Figures 16 and 21. O-shaped burrows (OS), U-shaped burrows (US), and subvertical ramps (SR) are only produced by N. americanus.
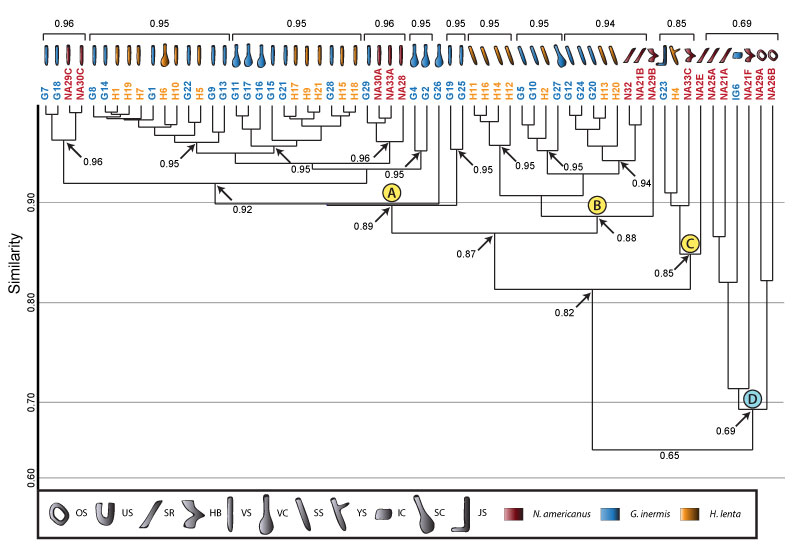
FIGURE 24. Combined cluster analysis of burrows produced by all arthropods considered in this study. Clusters A, B, C, and D consist of burrows produced by small arachnids and millipedes. Clusters E and F consist of burrows produced by large arachnids and millipedes. The burrow architectures placed into each cluster and the tracemaker are indicated by symbols above each cluster. Multiple species boxes placed below a burrow symbol mean each species produced that burrow architecture and that it is placed into that cluster. Formatting and key is the same as in Figures 16 and 21.
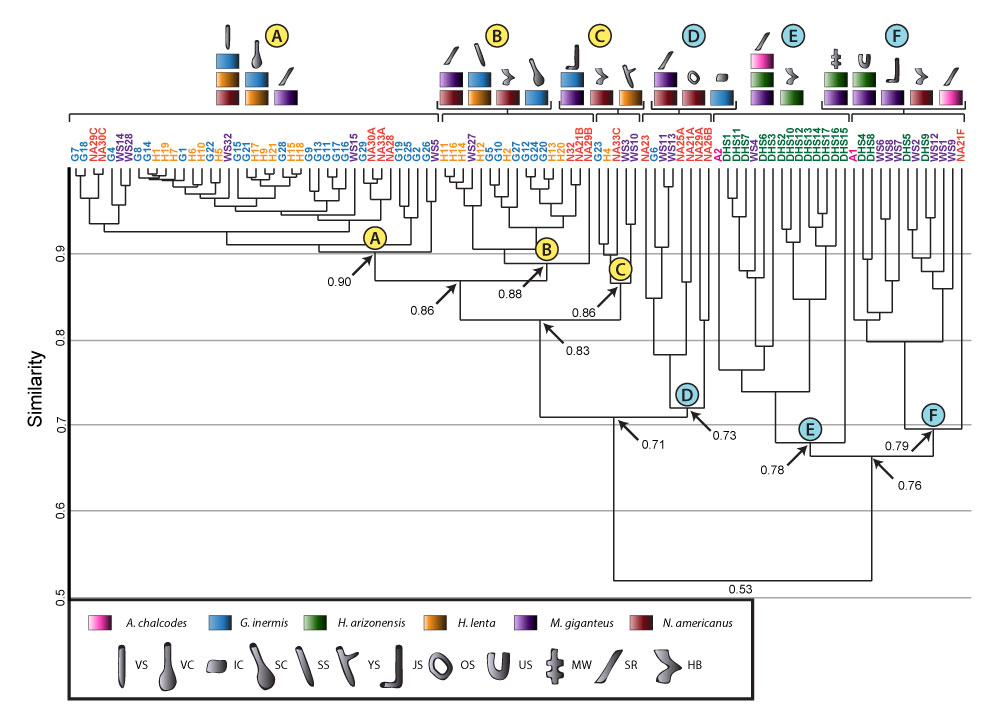
Neoichnology of the burrowing spiders Gorgyrella inermis (Mygalomorphae: Idiopidae) and Hogna lenta (Araneomorphae: Lycosidae)
Plain Language Abstract
The form of modern traces is related to the body shape of the animal that made the trace, the behaviors expressed by the animal when making or using the trace, and the environment in which the trace was made. Studying fossil traces may provide information about environmental conditions and the types of organisms present in the past. The goals of this study were: 1) to document the shapes of burrows produced by two species of burrowing spiders, 2) to determine if environmental conditions affected the shapes of the burrows produced, and 3) to determine how similar the spider burrows were to those of other arthropods. South African trapdoor spiders (Gorgyrella inermis) and field wolf spiders (Hogna lenta) were provided with aquariums filled with sediment in which to construct burrows. Gorgyrella inermis individuals produced vertical shafts, subvertical shafts, vertical shafts with terminal chambers, subvertical shafts with terminal chambers, isolated chambers, and J-shaped burrows. Hogna lenta individuals produced vertical shafts, subvertical shafts, vertical shafts with terminal chambers, and Y-shaped burrows. A very important feature to all of these burrows was the presence of a silk lining. The burrows made by G. inermis and H. lenta were found to be highly similar to one another. When sediment conditions were altered, the burrows were statistically similar, but the variety of burrow shapes decreased. The spider burrows were compared to four other burrowing animals: a tarantula (Aphonopelma chalcodes), a scorpion (Hadrurus arizonensis), a whip scorpion (Mastigoproctus giganteus), and a millipede (Narceus americanus). The result of these comparisons suggests that the burrows made by the scorpions, the tarantulas, and larger whip scorpions were quite different than those of G. inermis and H. lenta. The smaller spider produced burrows that were more similar to small whip scorpion burrows and the millipede burrows.
Resumen en Español
Neoicnología de las arañas excavadoras de madrigueras Gorgyrella inermis (Mygalomorphae: Idiopidae) y Hogna lenta (Araneomorphae: Lycosidae)
Los arácnidos excavadores de madrigueras son importantes para los actuales ecosistemas del suelo, pero el conocimiento de estos animales en los antiguos ecosistemas del suelo es limitado. En este estudio, se investigaron dos especies de arañas excavadoras de madrigueras: Gorgyrella inermis ("South African trapdoor spider") y Hogna lenta ("field wolf spider"). Se estudiaron individuos de cada especie para investigar sus técnicas de excavación y su comportamiento y, además, para categorizar las morfologías de sus madrigueras. Los experimentos se llevaron a cabo con variaciones en la densidad y humedad del sedimento para evaluar los efectos de las condiciones ambientales sobre la morfología de las madrigueras. Las arañas produjeron siete arquitecturas diferentes de madrigueras: pozos verticales, pozos verticales con cámaras terminales, pozos subverticales, pozos subverticales con cámaras terminales, madrigueras con forma de J, madrigueras con forma de Y, y cámaras aisladas. Todas estas arquitecturas de madriguera tienen características comunes que las hacen identificables como madrigueras de araña. La densidad y la humedad del sedimento tuvieron poca influencia en la morfología de las madrigueras, pero la diversidad de arquitecturas fue mayor en sedimentos de densidad y humedad moderadas. Los resultados de este estudio muestran que las arañas producen estructuras biogénicas únicas que se pueden distinguir de las madrigueras de otros organismos del suelo. Los datos recogidos en este estudio pueden ser utilizados para interpretar mejor la paleoecología y la historia evolutiva de las arañas y los artrópodos del suelo.
Palabras clave: icnofósiles; huellas fósiles; continental; artrópodos; arácnido; paleoecología
Traducción: Enrique Peñalver
Résumé en Français
Néoichnologie des araignées fouisseuses Gorgyrella inermis (Mygalomorphae: Idiopidae) et Hogna lenta (Araneomorphae: Lycosidae)
Les arachnides fouisseurs sont importants pour les écosystèmes de sols modernes, mais la connaissance de ces animaux dans les écosystèmes de sols anciens est limitée. Dans cette étude, deux espèces d'araignées fouisseuses ont été étudié: Gorgyrella inermis (mygale fouisseuse noire d'Afrique du Sud) et Hogna lenta (araignée aranéomorphe). Les individus de chaque espèce ont été étudiés pour enquêter leurs techniques et comportements de fouissages et pour catégoriser les morphologies de leurs terriers. Les expériences ont été effectuées avec des variations de la densité et de l'humidité des sédiments et évaluer les effets des conditions environnementales sur la morphologie du terrier. Sept architectures de terriers ont été produites par ces araignées: des puits verticaux, des puits verticaux avec des chambres terminales, des puits sous-verticaux, de puits sous-verticaux avec des chambres terminales, des terriers en forme de J, des terriers en forme de Y, et des chambres isolées. Toutes les architectures de terriers ont des caractéristiques communes qui les rendent identifiables comme des terriers d'araignée. La densité et l'humidité des sédiments avaient peu d'influence sur la morphologie terrier, mais la diversité de l'architecture était plus grande dans les sédiments de densité et d'humidité modérée. Les résultats de cette étude montrent que les araignées produisent des structures biogéniques uniques qui peuvent être distinguées des terriers d'autres organismes du sol. Les résultats de cette étude peuvent être utilisés pour mieux interpréter la paléoécologie et l'évolution des araignées et des arthropodes du sol.
Mots-clés: ichnofossiles; fossiles traces; continental; arthropodes; arachnide; paléoécologie
Translator: Kenny J. Travouillon
Deutsche Zusammenfassung
Neochnologie der grabenden Spinnen Gorgyrella inermis (Mygalomorphae: Idiopidae) und Hogna lenta (Araneomorphae: Lycosidae)
Grabende Arachniden sind wichtig für moderne Bodenökosysteme, jedoch ist das Wissen über prähistorische Bodenökosysteme limitiert. In dieser Studie wurden zwei Arten von grabenden Spinnen untersucht: Gorgyrella inermis (Südafrikanische Falltürspinne) und Hogna lenta (Wiesenwolfsspinne). Um die Grabtechniken und Verhaltensweisen zu ermitteln und um die Bauten-Morphologie zu kategorisieren wurden Individuen jeder Art untersucht. Zur Evaluierung der Effekte von Umweltbedingungen auf die Baubeschaffenheit wurden Experimente mit variierender Sedimentdichte und Feuchtigkeit durchgeführt. Die Spinne produzierten sieben Baustile: vertikale Schächte, vertikale Schächte mit endständigen Kammern, subvertikale Schächte, subvertikale Schächte mit endständigen Kammern, J-förmige Grabbauten, Y-förmige Grabbauten und isolierte Kammern. Jede Architektur weist gemeinsame Merkmale auf, die sie als Spinnenbauten charakterisiert. Sedimentdichte und Feuchtigkeit haben wenig Einfluss auf die Morphologie der Grabbauten, jedoch war die Diversität der Architektur bei Sedimenten mit moderater Dichte und Feuchtigkeit am höchsten. Die Ergebnisse dieser Untersuchung zeigen, dass Spinnen einzigartige biogene Strukturen erzeugen, die von den Grabbauten anderer Organismen unterschieden werden können. Das in dieser Studie gesammelte Datenmaterial kann dazu genutzt werden die Paläoökologie und Evolutionsgeschichte der Spinnen und Bodenarthropoden besser zu interpretieren.
Schlüsselwörter: Ichnofossilien; Spurenfossilien; kontinental; Arthropode; Arachnide; Paläoökologie
Translator: Eva Gebauer
Arabic
in progress
Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024