

Article Search
Volume 27.2
May–August 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.2 May-August 2024
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
APPENDIX 1.
Descriptions of new dental characters employed in the phylogenetic analyses; some of them were modified from Bryant et al. (1993), Wolsan (1993), Baskin (2004) and Valenciano et al. (2015, 2016).
Characters and character states used in the phylogenetic analysis
(1) Posterior lacerate and jugular foramina: “fused” into a single foramen (0); tendency to separate openings with the jugular foramen distolateral to the posterior lacerate foramen (1).
(2) Shape of upper incisor row: parabolic (0); straight (1).
(3) Position of the infraorbital foramen: above P3 (0); above P4 parastyle (1).
(4) Incisive foramen: located at the level of C (0); located at the level of the diastema I3-C (1).
(5) Mastoid process: reduced, located in dorsal view in line with the middle point of the orbit (0); enlarged, located laterally exceeding to the orbit (1).
(6) Relative position of the mastoid and paroccipìtal processes: relatively close (0); mastoid process located mesially to the paroccipital process (1).
(7) Paroccipital process: not reduced (0); reduced (1).
(8) Height of the mandibular corpus: low and thin mandibular corpus (0); high and thick mandibular corpus (1).
(9) Orientation of the I3 cusp: spreaded out laterally (0); in line with the cusps of I1-2 (1).
(10) Orientation of Canine: spreaded out laterally, with an arrangement of the tip non parallel (0); ventrally directed, with a parallel arrangement of the tip (1).
(11) P1. Present (0); absent (1).
(12) P2. Mesiodistal axis of P2: in line with the tooth row (0); rotated buccolingually (1).
(13) P2. Distal accessory cusp: absent (0); present (1).
(14) P2. Occlusal shape: subrectangular (0); triangular (1).
(15) P2. Buccal wall: rectilinear (0); conspicuous concavity (1).
(16) P3. Robustness ratio [(maximum width/ maximum length) x 100]: slender P3 (less than 60) (0); Robust P3 (60 or more than 60) (1).
(17) P3. Mesial accessory cusp: reduced or absent (0); present (1).
(18) P3. Distal accessory cusp: present (0); absent (1).
(19) P3. Buccal wall: rectilinear wall (0); conspicuous concavity (1).
(20) P3. Basal cingulum: weak (0); strong (1).
(21) P4/3 length ratio. Maximum length of P4 in relation to maximum length of P3 ratio [(L P4/L P3) x 100]: less than 170 (0); more than 170 (1).
(22) P4. Robustness ratio [(maximum width/ maximum length) x 100]: slender P4 (less than 60) (0); robust P4 (more than 60) (1).
(23) P4 protocone: low (0); knoblike (1).
(24) P4. Protocone cingulum: weak or absent (0); prominent and complete (1).
(25) P4. Protocone: in front or anterior to the mesiobuccal corner (0); displaced distally (1).
(26) P4. Parastyle: Absent or weak (0); Strong and low (1); Strong and high (2).
(27) P4. Buccal wall: rectilinear (0); with a pronounced concavity between paracone and metastyle (1).
(28) P4. Basal cingulum surrounding the tooth: weak (0); strong (1).
(29) Relative size between maximum length of M1 and the maximum length of P4: M1 larger than or equal in size to P4 (values >100) (0); M1 smaller than P4 (values between 70-100) (1); M1 much smaller than P4 (values <70) (2).
(30) M1. Robustness ratio [(maximum width on the buccolingual area/ maximum length) x 100]: relatively robust M1 (from 130 to 150) (0); slender M1 (more than 160) (1), very slender M1 (more than 200) (2).
(31) M1. Stylar area: small (0); enlarged (1).
(32) M1. Metacone related to paracone: normal size (0); very reduced (1).
(33) M1. Metaconule: present (0); absent (1).
(34) M1. Enlargement of the distal base of the metaconule: present (0); absent (1).
(35) M1. Protocone position: mesolingually located (0); almost centrally on the middle of the talone (1).
(36) M1. Protocone shape: ridge-shaped (0); conical cusp-like shape (1).
(37) M1. Lingual platform: not completely enclose the protocone (0); completely enclose the protocone (1).
(38) M1. Length of the lingual wall: lesser than the buccal wall (unexpanded lingual platform) (0); longer than the buccal wall (moderately lingual platform) (1); much longer than the buccal wall (very expanded lingual platform) (2).
(39) M1. Lingual platform: oval shape (0); with a concavity in the middle point (1).
(40) p1. Present (0); absent (1).
(41) p2. Position of the mesiodistal axis of p2: in line with the tooth row (0); buccolingually rotated (1).
(42) p2. Distal accessory cuspid: absent (0); present (1).
(43) p2. Robustness ratio [(maximum width/ maximum length) x 100]: slender p2 (less than 50) (0); relatively robust p2 (from 50 to 70) (1); very robust p2 (more than 70) (2).
(44) p2 length compared to p3: p2 not reduced (0); p2 reduced (1).
(45) p2-4. Basal cingula with a mesial and distal thickened cristids: cingula and cristid weaks (0); complete and strong cingula and cristids (1).
(46) p3. Mesial accessory cuspid: absent (0); present (1).
(47) p3. Distal accessory cuspid: present (0); absent (1).
(48) p3. Distal thickened of p3: absent (0); present (1).
(49) p4. Length ratio in relation to m1 [(maximum length p4/ maximum length m1) x 100]: from 50 to 60, indicating a relatively not reduced p4 (0); more than 60, indicating a p4 relatively enlarged (1).
(50) p4. Mesial accesory cuspid: absent or poorly developed (0); present, well developed (1).
(51) p4. Distal accessory cuspid: High and well developed (0); low and reduced (1); Absent (2).
(52) p4. Backward inclination of the main cuspid: practically vertical (90º-80º) (0); with backward inclination (less than 80º) (1).
(53) p4, unworn principal cusp: equals or exceeds height of m1 paraconid (0); lower than m1 paraconid (1).
(54) p4. Distal area: not buccolingually thickened, oval shaped (0); buccolingually thickened, quadrangular shaped (1).
(55) p4. Lingual expansion or basal bulge: absent (0); present (1).
(56) m1. Height of protoconid in relation to paraconid: protoconid higher than paraconid (0); protoconid similar in height than the paraconid (1).
(57) m 1. Metaconid: individualized, with a moderate size (0); reduced (1); absent (2).
(58) m1. Relative length of talonid with respect the total m1 length: the talonid 1/3 of the total length (0); equal or less than 1/4 of the total length (1).
(59) m1. Width talonid ratio [(maximum talonid width/ maximum width in the base of the protoconid-metaconid) x 100]: talonid not widened (values between 85-100) (0); reduced talonid (< 85) (1).
(60) m1. Height of hypoconid: low (0); high (1).
(61) m1. Position of hypoconid: labially located (0); centrally positioned or almost centrally positioned (1).
(62) m1. Orientation of the hypoconid: almost vertical (0); orientated towards the protoconid (1).
(63) m1. Entoconid: individualized (0); transform in an entocristid (1); absent (2).
(64) m1 talonid: closed and deep basin (0); open and shallow basin with a low entocristid and a beveled lingual wall of the talonid (1); basin lost (2).
(65) m1. Shape of the hypoconid: pyramidal (0); trending to a crest-like shape (1).
(66) m1. Hypoconid size: medium (0); enlarged (1); reduced (2).
(67) m1. Hypoconulid: very reduced to absent (0); present not reduced (1).
(68) (39) m2. Present (0); absent (1).
(69) m2 paraconid: present (0); (1) very week or absent (1).
(70) m2. Metaconid: present (0); absent (1).
APPENDIX 2.
Character-taxon matrix used for phylogenetic analyses.

APPENDIX 3.
Character-taxon matrix in nexus format. (Available online in zipped format)
 Alberto Valenciano. Departamento de Geología Sedimentaria y Cambio Medioambiental. Instituto de Geociencias (CSIC, UCM). Calle José Antonio Novais 12, 28040. Madrid, Spain and Departamento de Paleontología Universidad Complutense de Madrid (UCM), Facultad de Ciencias Geológicas UCM. Calle José Antonio Novais 12, 28040, Madrid, Spain. alb3rtovv@gmail.com
Alberto Valenciano. Departamento de Geología Sedimentaria y Cambio Medioambiental. Instituto de Geociencias (CSIC, UCM). Calle José Antonio Novais 12, 28040. Madrid, Spain and Departamento de Paleontología Universidad Complutense de Madrid (UCM), Facultad de Ciencias Geológicas UCM. Calle José Antonio Novais 12, 28040, Madrid, Spain. alb3rtovv@gmail.com
Alberto Valenciano is a PhD. granted candidate at Instituto de Geociencias and Universidad Complutense de Madrid. His research is focused on the extinct giant mustelids from the Neogene and Quaternary of Europe, Asia, Africa and North America (e.g. Ekorus, Enhydritherium, Enhydriodon, Eomellivora, Hoplictis, Megalictis, Oligobunis, Plesiogulo, Sivaonyx...). He is especially interested in comparative, biometric, morphological and three-dimensional studies of extinct large and giant musteloids (including mustelids, ailurids, procyonids and mephitids) to understand their paleobiology and their evolutive relationships. At the same time, He is studying some carnivorans (mustelids, mephitids and ursids) from the vertebrate site of Batallones (Bat-1 and Bat-3), late Miocene, Vallesian MN 10 (Torrejón de Velasco, Spain). Moreover He is also collaborating with several vertebrate paleontologist researchers studying small Neogene musteloids, bears and amphicyonids from Europe, North America, Turkey and Africa.

 Juan Abella. Universidad Estatal Península de Santa Elena. Kilómetro 1,5 de la Vía Santa Elena- La Libertad. Edificio Instituto de Investigacion Científica y Desarrollo Tecnológico INCYT), Segundo Piso. 240210 La Libertad, Ecuador. jabella@upse.edu.ec and Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona. Edifici Institut de Ciència i Tecnologia Ambientals - Institut Català de Paleontologia (ICTA-ICP), Campus de la UAB, Carrer de les Columnes s/n, 08193 Cerdanyola del Vallès, 08193, Barcelona, Spain. juan.abella@icp.cat
Juan Abella. Universidad Estatal Península de Santa Elena. Kilómetro 1,5 de la Vía Santa Elena- La Libertad. Edificio Instituto de Investigacion Científica y Desarrollo Tecnológico INCYT), Segundo Piso. 240210 La Libertad, Ecuador. jabella@upse.edu.ec and Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona. Edifici Institut de Ciència i Tecnologia Ambientals - Institut Català de Paleontologia (ICTA-ICP), Campus de la UAB, Carrer de les Columnes s/n, 08193 Cerdanyola del Vallès, 08193, Barcelona, Spain. juan.abella@icp.cat
Juan Abella is a PhD, staff member of the Universidad Estatal Península de Santa Elena, Ecuador. Although his main research in palaeontology dealt with several groups of mammalian carnivorans, mainly the Arctoidea, and his PhD thesis was study of the fossil remains of the basal bear Indarctos arctoides from the Batallones-3 site. Recently, he is also carrying out a research project about the Cenozoic evolution of the Ecuadorian faunas, with emphasis in marine vertebrates. He is author of more than 20 works published in scientific journals and books and he has participated in more than 5 research projects.
 Ursula B. Göhlich. Naturhistorisches Museum Wien, Dept. Geology and Paleontology Burgring 7, 1010 Vienna, Austria. ursula.goehlich@nhm-wien.ac.at
Ursula B. Göhlich. Naturhistorisches Museum Wien, Dept. Geology and Paleontology Burgring 7, 1010 Vienna, Austria. ursula.goehlich@nhm-wien.ac.at
Dr. Ursula Göhlich is curator of vertebrate paleontology at the Natural History Museum of Vienna since 2007. She received her PhD in 1997 from the University of Munich (Germany) followed by several Postdoc projects e.g. at the University of Lyon (France) (as an Alexander-von-Humboldt fellow) and at the Natural History Museum of Los Angeles (USA). In 2011 she obtained her habilitation from the University of Munich, where she is an associated professor since then.
She is specialized on European Neogene proboscideans and birds and addresses taxonomy, biogeography, biostratigraphy, and paleoecology, but her research also covers other vertebrate groups and topics.
 M. Ángeles Álvarez-Sierra. Departamento de Geología Sedimentaria y Cambio Medioambiental. Instituto de Geociencias (CSIC, UCM). Calle José Antonio Novais 12, 28040. Madrid, Spain and Departamento de Paleontología Universidad Complutense de Madrid (UCM), Facultad de Ciencias Geológicas UCM. Calle José Antonio Novais 12, 28040, Madrid, Spain. masierra@ucm.es
M. Ángeles Álvarez-Sierra. Departamento de Geología Sedimentaria y Cambio Medioambiental. Instituto de Geociencias (CSIC, UCM). Calle José Antonio Novais 12, 28040. Madrid, Spain and Departamento de Paleontología Universidad Complutense de Madrid (UCM), Facultad de Ciencias Geológicas UCM. Calle José Antonio Novais 12, 28040, Madrid, Spain. masierra@ucm.es
M. Ángeles Álvarez-Sierra is a professor of paleontology at the Universidad Complutense of Madrid, Spain. She is focused on European Cenozoic small mammals (mainly rodents) and concerned about paleobiology and biostratigraphy. She participates in excavations and studies of several fossil sites in Spain (Cuenca de El Ebro, Madrid -Batallones, Somosaguas-, Calatayud-Teruel, Loranca). She is author of more than 100 works published in scientific journals and books and she has participated in more than 25 research projects.
 Jorge Morales. Departamento de Paleobiología. Museo Nacional de Ciencias Naturales-CSIC, Calle José Gutiérrez Abascal, 2, 28006, Madrid, Spain. mcnm166@mncn.csic.es
Jorge Morales. Departamento de Paleobiología. Museo Nacional de Ciencias Naturales-CSIC, Calle José Gutiérrez Abascal, 2, 28006, Madrid, Spain. mcnm166@mncn.csic.es
Jorge Morales is Research Professor at the National Museum of Natural Sciences of Madrid, CSIC, Spain. He focuses his research on ruminant and carnivoran systematics and evolution from Eurasia and Africa. He has been notably involved in field excavations in the Miocene of Spain and Africa. He leads the research project of late Miocene of Batallones fossil vertebrate sites (Madrid basin, Spain). These localities are yielding some of the most interesting, richest and best-preserved Neogene (late Miocene) vertebrate assemblages of the Iberian Peninsula.
FIGURE 1. Vienna Basin with geographic and stratigraphic position of localities (asterisks) (modified after Harzhauser et al., 2004, figures 1 and 2).
FIGURE 2. Fossil remains of Eomellivora fricki (Pia, 1939) from Wien XII-Altmannsdorf and Gaiselberg. 1-4, Holotype NHMW 2016/0065/0001 from Wien XII-Altmannsdorf,left maxilla fragment, 1, lateral view, 2, occlusal view, 3, rostral view, 4, caudal view; 5-6, right maxilla fragment, 5, lateral view, 6, occlusal view; 7, fragment of the left zygomatic arch; 8, fragment of the left temporo-mandibular joint; 9-10, fragment with the right mastoid process (mp), the ventral part of the nuchal crest (nc), the external auditory meatus (eam), and part of the glenoid cavity (gc), 9, lateral view, 10, dorsal view; 11-14, NHMW 1977/1948/0130 right hemimandible from Gaiselberg. 11, lateral view, 12, medial view, 13, occlusal view, 14, ventral view. Scale bar equals 5 cm.
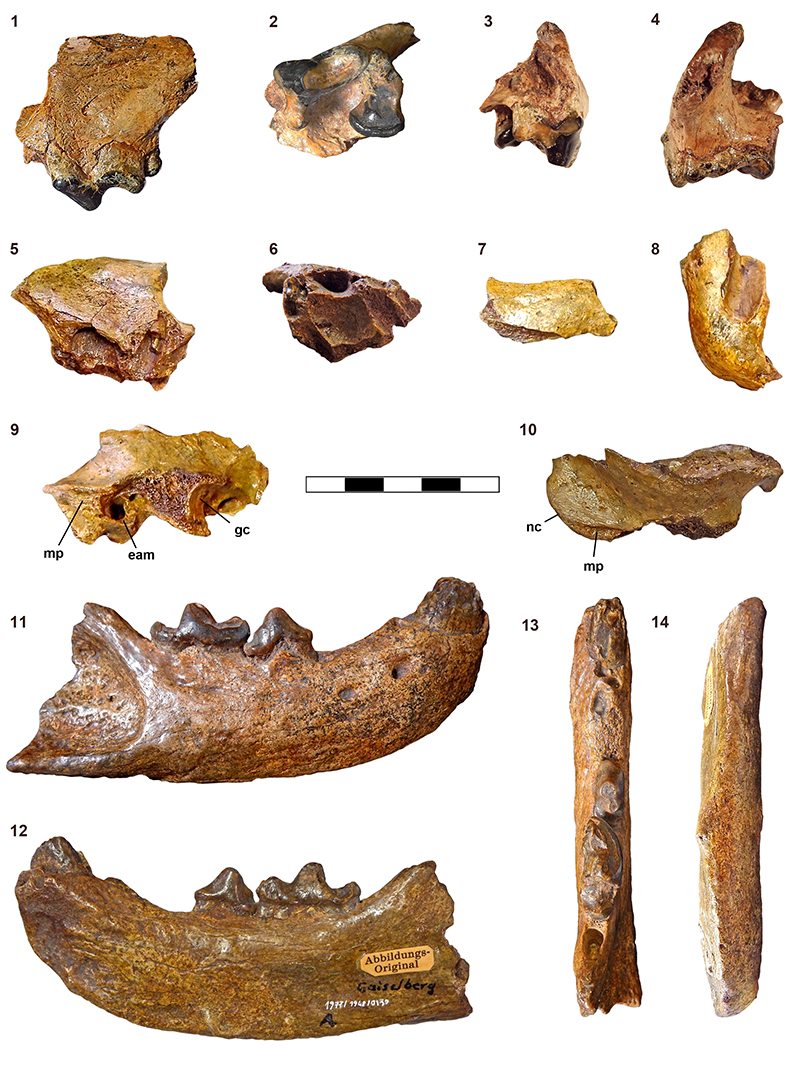
FIGURE 3. Phylogenetic relationship of Eomellivora fricki within Eomellivora, and of some extinct Neogene mustelids and living carnivorans. Canis lupus is the outgroup. Searches were performed by means of the Branch and Bound and a Bootstrap analysis through 1000 replicates. A single tree is obtained (length 172 steps, consistency index (CI)=0.4593, retention index (RI)=0.5811). The numbers below nodes are Bremer indices, and the numbers above nodes are Bootstrap support percentages (only shown ≥ 50). Letters (A-K) indicated selected nodes. The synapomorphies for each node are reported in Table 4.
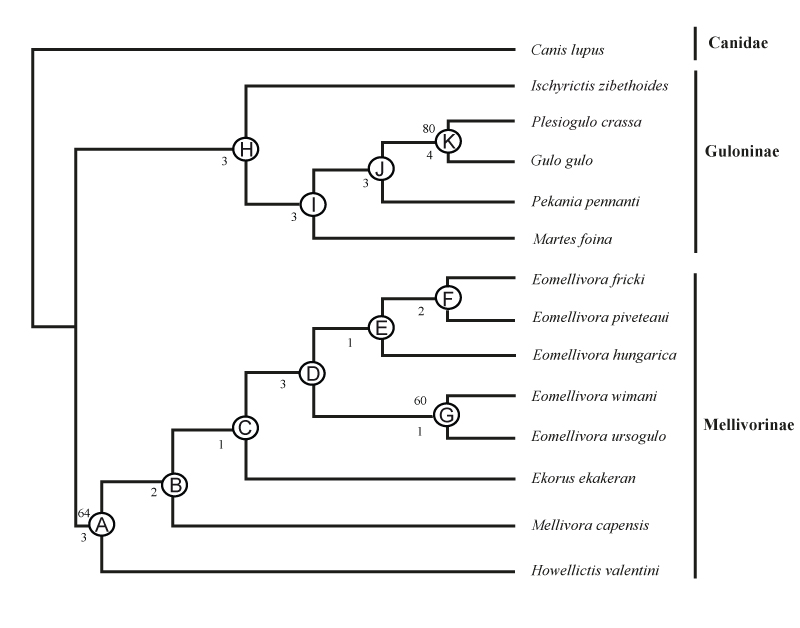
FIGURE 4. Main comparative material of the upper dentition of species of Eomellivora considered in the present manuscript. 1, Holotype of Eomellivora fricki NHMW 2016/0065/0001 from Wien XII-Altmannsdorf (Austria), MN9; 2, Eomellivora piveteaui MNHN-TRQ-1005 from type locality Yassiören (Turkey), MN9; 3, Eomellivora piveteaui Bat-3´13.185 from Batallones (Spain), MN10; 4, Holotype of Eomellivora ursogulo PIN-No.268 from Grebeniki (Ukraine), MN11; 5, Lectotype of Eomellivora wimani PMU-M3692 from Shangyingou (China), MN12-13; 6, Paratype (M1) of Eomellivora hungarica MFGI-Ob-3831 from Polgárdi 2 (Hungary), MN13; 8, cast of the holotype of Eomellivora ursogulo NHMW 2016/0085/0001 from Grebeniki (Ukraine), MN11; 9, cast of the holotype of Ekorus ekakeran KNM-LT 23125 from Lothagan (Kenya), 7 m.y.a. 1-6, occlusal view, 7-9, lateral view. Scale bar equals 5 cm.

FIGURE 5. Main comparative material of mandible and lower dentition of species of Eomellivora considered in this manuscript. 1-2, Eomellivora fricki NHMW 1977/1948/0130 from Gaiselberg (Austria), MN9. 1, lateral view, 2, occlusal view; 3-4, Eomellivora piveteaui Bat-3´13.230 from Batallones (Spain), MN10. 3, lateral view, 4, occlusal view; 5-6, Eomellivora wimani PMU-M3693 from Shangyingou (China), MN12-13 (same specimen as PMU-M3692). 5, lateral view, 6, occlusal view; 7-8, Holotype of Eomellivora ursogulo PIN-No.269a from Grebeniki (Ukraine), MN11. 7, lateral view, 8, occlusal view; 9-10, Holotype of Eomellivora hungarica MFGI-Ob-2676 from Polgárdi 2 (Hungary), MN13. 9, lateral view, 10, occlusal view. Scale bar equals 5 cm.

FIGURE 6. Scatter diagrams of measurements (mm) of the dentition of Eomellivora and Ekorus ekakeran. 1, P4; 2, M1; 3, c; 4, p4; 5, m1; 6, m2. Sources: WXII (Wien XII-Altmannsdorf) present manuscript; GAI (Gaiselberg) present manuscript; SH (Shangyingou), and LI (Liuwangou), Zdansky (1924); GRT (Gritsev), Wolsan and Semenov (1996); NO (Novaya Emetovka), Orlov (1948); GYÖ (Györszentmárton), Kretzoi (1965), KRF (Kern River Formation site 50), Stock and Hall (1933); CIM (Cimislia), Wolsan and Semenov (1996); YAS (Yassiören), Ozansoy (1965) and for P4 and M1, estimations based on pictures of MNHN-TRQ-1005, rather than the evidently confusing original data provided in Ozansoy (1965); WSS (Wissberg), Tobien (1955); RPI (Ravin de la Pluie), Koufos (2012); BAT (Batallones), Valenciano et al. (2015); LVF (Los Valles de Fuentidueña), Crusafont-Pairó and Ginsburg (1973) and for p2 Valenciano et al. (2015); KLF (Kalfa), Lungu (1978) and for M1 Valenciano et al. (2015); GRE (Grebeniki), Orlov (1948); CSA (Csákvár), Kretzoi (1942); POL (Polgárdi 2); LOT (Lothagam) Werdelin, 2003.
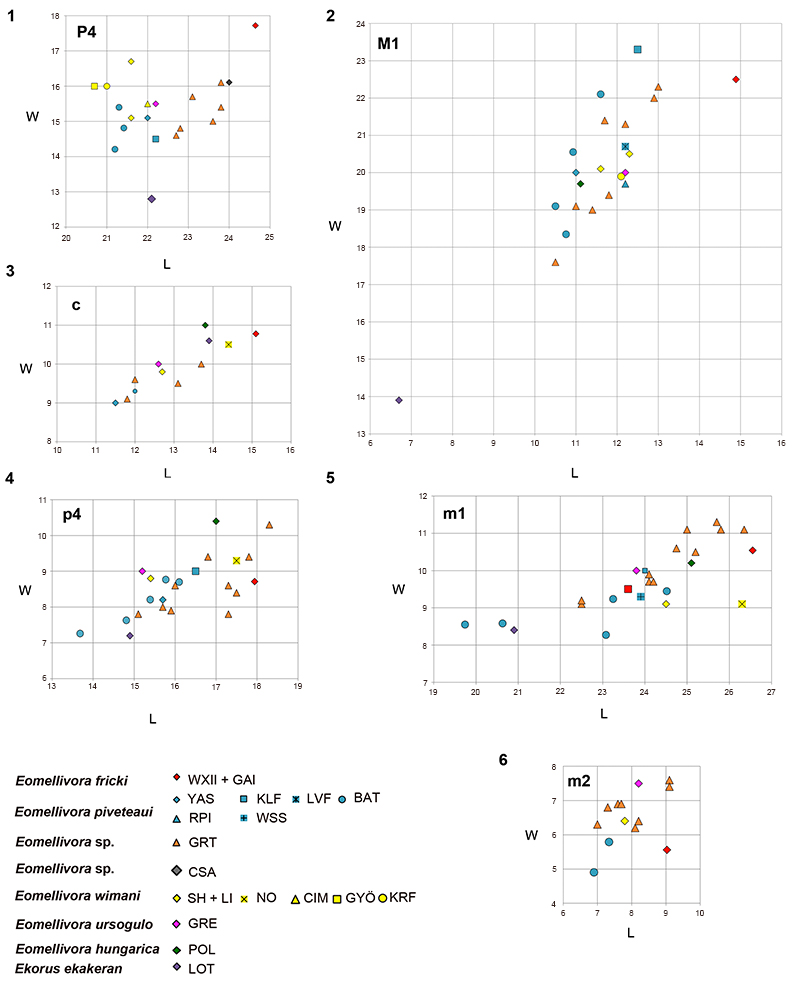
TABLE 1. Fossil vertebrate fauna Wien XII-Altmannsdorf (Griesergasse, Oswaldgasse) (updated by U.B.Göhlich, based on Pia (1939) and Thenius (1948)).
| Wien XII-Altmannsdorf |
| Carnivora |
| Eomellivora fricki (formerly Hadrictis) |
| Lycaena chaeretis |
| Perissodactyla |
| Aceratherium incisivum |
| Hippotherium sp. |
| Artiodactyla |
| Miotragocerus pannoniae |
| Propotamochoerus palaeochoerus |
| Proboscidea |
| Deinotherium giganteum |
TABLE 2. Fossil vertebrate fauna from Gaiselberg (uddated by U.B. Göhlich, based on Zapfe (1949), Thenius (1950), Pickford (2016) and J. Giaourtsakis (personal commun., Athens, 2014).
| Gaiselberg |
| Carnivora |
| Eomellivora fricki (formerly Hadrictis) |
| Amphicyon sp. |
| Felidae indet. |
| Perissodactyla |
| Aceratherium incisivum |
| Hoploaceratherium sp. |
| Hippotherium sp. |
| Anchitherium aurelianense |
| Chalicotherium goldfussi |
| Artiodactyla |
| Conohyus doati (formerly Hyotherium palaeochoerus) |
| Dorcatherium naui |
| Miotragocerus pannoniae |
| Bovidae indet (smaller than M. pannoniae) |
| Proboscidea |
| Deinotherium giganteum |
| Tetralophodon longirostris |
TABLE 3. Upper and lower teeth measurements (in mm) of Eomellivora fricki (Pia, 1939). L (length) and W (Width). The measurements for the p2 and m2 of NHMW 1977/1948/0130 are based on its alveolus.
| P4 | M1 | c | p2 | p4 | m1 | m2 | ||||||||
| L | W | L | W | L | W | L | W | L | W | L | W | L | W | |
| NHMW 2016/0065/0001 | 24.6 | 17.7 | 14.9 | 22.5 | ||||||||||
| NHMW 1977/1948/0130 | 15.1 | 10.8 | 11.5 | 5.7 | 17.9 | 8.7 | 26.5 | 10.5 | 9 | 5.6 | ||||
TABLE 4. Synapomorphies for selected nodes, with associated character numbers and states (in parenthesis). Italics denote ambiguous synapomorphies.
| Node | Character: state |
| A | 6 (1), 11 (1), 20 (1), 32 (1), 35 (1), 40 (1), 41 (1), 52 (1), 57 (2), 58 (1), 59 (1), 62 (1), 63 (2), 64 (2), 70 (1) |
| B | 12 (1), 13 (1), 16 (1), 22 (1); 26 (2), 27 (1), 28 (1), 42 (1), 46 (1); 48 (1), 50 (1), 56 (1) |
| C | 2 (1), 3 (1), 11(1), 17 (1), 19 (1), 31 (1), 61 (1) |
| D | 13 (0), 24 (1), 26 (1), 27 (0), 42 (0), 54 (1), 60 (1) |
| E | 17 (0), 30 (1), 36 (0), 69 (0) |
| F | 32 (0), 35 (0), 50 (0) |
| G | 14 (1), 15 (1), 38 (2), 43 (2) |
| H | 3 (1), 7 (1), 18 (1), 38 (2), 52 (0),55 (1), 65 (1), 66 (0), 67 (1) |
| I | 8 (0), 21 (1), 27 (1), 36 (0), 43 (0), 47 (1), 49 (0), 51 (1) |
| J | 23 (0), 24 (1), 25 (1), 26 (1), 28 (1), 32 (1) |
| K | 1 (0), 6 (1), 7 (0), 8 (1), 9 (0), 15 (1), 19 (1), 20 (1), 41 (1), 43 (2), 45 (1), 48 (1), 51 (2), 53 (1), 56 (1) |
Re-evaluation of the very large Eomellivora fricki (Pia, 1939) (Carnivora, Mustelidae, Mellivorinae) from the Late Miocene of Austria
Plain Language Abstract
The mustelids is a family of carnivorous mammals that includes weasels, ferrets, badgers, minks, otters, martens and wolverines. This manuscript provides information about a very large extinct relative of the living honey badger (Mellivora capensis), described initially as Hadrictis fricki Pia, 1939 from the Late Miocene of Austria. After making a re-evaluation of these fossils, we conclude that Hadrictis can be considered as a synonymy of Eomellivora, therefore naming it as Eomellivora fricki, being one of the earliest representatives of the genus. In addition, based on the dentition, Eomellivora fricki emerge as one of the largest mustelids ever. Also, a detailed comparison with other species of Eomellivora has revealed marked differences with all of them, demonstrating that E. fricki is a valid species clearly distinguishable from other mellivorines. Based on our phylogenetic analyses, the Late Miocene large-sized mustelids Eomellivora and Ekorus are more closely related with the honey badger than with the other living mustelids.
Resumen en Español
Reevaluación de la especie de gran tamaño Eomellivora fricki (Pia, 1939) (Carnivora, Mustelidae, Mellivorinae) del Mioceno Tardío de Austria
Reevaluamos el material austríaco de Hadrictis fricki Pia, 1939, de las localidades Wien XII-Altmannsdorf y Gaiselberg (MN9, Vallesiense, Mioceno Tardío), concluyendo que Hadrictis puede ser considerado una sinonimia de Eomellivora Zdansky, 1924; por lo tanto, a este taxón lo consideramos Eomellivora fricki. Esta especie es uno de los primeros representantes del género, junto con E. piveteaui Ozansoy, 1965. Nuestros análisis filogenéticos indican que Eomellivora forma un grupo monofilético, estableciendo el clado hermano del grande y derivado Ekorus ekakeran del Mioceno Tardío. Eomellivora fricki muestra una morfología dental primitiva y es la especie más grande del género. Esta especie muestra la complejidad del género Eomellivora, en el que coexistían tanto especies grandes como pequeñas desde el inicio del Mioceno Tardío.
Palabras clave: Eomellivora; Hadrictis; Mellivorinae; Mustelidae; Mioceno
Traducción: Enrique Peñalver (Sociedad Española de Paleontología)
Résumé en Français
Réévaluation de l'espèce de très grande taille Eomellivora fricki (Pia, 1939) (Carnivora, Mustelidae, Mellivorinae) du Miocène récent d'Autriche
Nous avons réévalué le matériel autrichien d'Hadrictis fricki Pia, 1939, provenant des localités Wien XII-Altmannsdorf et Gaiselberg (MN9, Vallésien, Miocène récent), et avons conclu qu'Hadrictis peut être considéré comme un synonyme d'Eomellivora Zdansky, 1924. Nous avons donc renommé cette espèce Eomellivora fricki. Cette espèce est une des plus anciennes du genre, avec E. piveteaui Ozansoy, 1965. Nos analyses phylogénétiques indiquent qu'Eomellivora forme un groupe monophylétique, groupe-frère du taxon dérivé et de grande taille Ekorus ekakeran du Miocène récent. Eomellivora fricki montre une morphologie dentaire primitive et est la plus grande espèce du genre. Cette espèce illustre la complexité du genre Eomellivora, dans lequel des espèces de petite et de grande taille ont coexisté depuis le début du Miocène récent.
Mots-clés : Eomellivora ; Hadrictis ; Mellivorinae ; Mustelidae ; Miocène
Translator:Antoine Souron
Deutsche Zusammenfassung
Neubeurteilung der großwüchsigen Art Eomellivora fricki (Pia, 1939) (Carnivora, Mustelidae, Mellivorinae) aus dem Obermiozän von Österreich
Die Neubeurteilung des österreichischen Materials von Hadrictis fricki Pia, 1939, aus den Lokalitäten Wien XII-Altmannsdorf und Gaiselberg (MN9, Vallesium, Obermiozän) ergibt, dass Hadrictis als Synonym von Eomellivora Zdansky, 1924 angesehen werden kann; hieraus resultiert die neue Namenskombination Eomellivora fricki. Diese Art ist zusammen mit E. piveteaui Ozansoy, 1965 einer der frühesten Vertreter dieser Gattung. Unsere phylogenetische Analyse zeigt Eomellivora als monophyletisches Taxon und als Schwestergruppe zur großen und fortschrittlichen obermiozänen Art Ekorus ekakeran. Eomellivora fricki zeichnet sich durch eine ursprüngliche Zahnmorphologie aus und stellt die größte Art innerhalb der Gattung dar. Diese Art verdeutlicht die Komplexität der Gattung Eomellivora, innerhalb derer große und kleine Arten seit Beginn des Obermiozäns koexistieren.
Schlüsselwörter: Eomellivora; Hadrictis; Mellivorinae; Mustelidae; Miozän
Translator: Authors
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024