

Article Search
Volume 27.2
May–August 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.2 May-August 2024
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
 C.B. Skovsted. Swedish Museum of Natural History, Department of Palaeobiology, SE-104 05 Stockholm, Sweden (corresponding author). christian.skovsted@nrm.se
C.B. Skovsted. Swedish Museum of Natural History, Department of Palaeobiology, SE-104 05 Stockholm, Sweden (corresponding author). christian.skovsted@nrm.se
Christian B. Skovsted is Senior Curator of invertebrate palaeontology at the Swedish Museum of Natural History, Stockholm, Sweden. From 2014 he also serves as Editor of GFF, the scientific journal of the Geological Society of Sweden. He received his PhD from Uppsala University in 2004 with a thesis concerning the Early Cambrian fauna of Small Shelly Fossils (SSF) from North East Greenland. Since then he has expanded his work to include Cambrian SSF faunas from all parts of the globe. In recent years he has mainly concentrated on the scleritome structure and taxonomy of the problematic tommotiids and their close relatives, the brachiopods.

 I. Knight. Government of Newfoundland and Labrador, Department of Natural Resources, Geological Survey, P.O. Box 8700, St. John’s, Newfoundland and Labrador, A1B 4J6. ianknight@gov.nl.ca
I. Knight. Government of Newfoundland and Labrador, Department of Natural Resources, Geological Survey, P.O. Box 8700, St. John’s, Newfoundland and Labrador, A1B 4J6. ianknight@gov.nl.ca
Ian Knight is a project geologist in the mapping section of the Geological Survey of Newfoundland and Labrador. He principally studies the stratigraphy of and maps the Lower Paleozoic shelf succession of the western Newfoundland. He also has worked on the Cambrian-Ordovician shelf succession in North-East Greenland with the Geological Survey of Greenland, the Carboniferous basins of western Newfoundland and the Proterozoic Ramah Group in northern Labrador.
 U. Balthasar. School of Geography, Earth and Environmental Sciences, Plymouth University, Drake Circus, Plymouth PL4 8AA, UK. uwe.balthasar@plymouth.ac.uk
U. Balthasar. School of Geography, Earth and Environmental Sciences, Plymouth University, Drake Circus, Plymouth PL4 8AA, UK. uwe.balthasar@plymouth.ac.uk
Uwe Balthasar did his undergraduate at the Philipps-Universität Marburg (Germany) before moving on to do a PhD at Cambridge on early Cambrian brachiopods and fossil preservation. After postdocs in Uppsala and Glasgow, he took up a lectureship in Palaeontology at Plymouth University in 2014. In addition to his interest in the early evolution of brachiopods and fossil preservation, he is interested in the nature of aragonite-calcite seas and how they impacted on the evolution of biomineralisation.
 W.D. Boyce. Government of Newfoundland and Labrador, Department of Natural Resources, Geological Survey, P.O. Box 8700, St. John’s, Newfoundland and Labrador, A1B 4J6. dougboyce@gov.nl.ca
W.D. Boyce. Government of Newfoundland and Labrador, Department of Natural Resources, Geological Survey, P.O. Box 8700, St. John’s, Newfoundland and Labrador, A1B 4J6. dougboyce@gov.nl.ca
W. Douglas Boyce specializes in Cambrian to Silurian trilobite faunas, and sometimes molluscs, fish and plants. He has been the Paleontologist for the Government of Newfoundland and Labrador, Canada since mid-1984. He also serves as Associate Curator of Paleontology, Provincial Museum Division, The Rooms Corporation of Newfoundland & Labrador. He received his MSc from Memorial University in 1984 with a thesis documenting new Early Ordovician trilobite faunas from the Boat Harbour and Catoche formations of western Newfoundland. He is a Past Chair of the Geological Association of Canada’s Paleontology Division and a Past President of the GAC Newfoundland Labrador Section.
APPENDIX 1.
Table listing all identified brachiopod specimens from all investigated samples collected by the authors from the Forteau Formation of southern Labrador and western Newfoundland (both sections on Great Northern Penninsula and Gros Morne National Park) with locality name and stratigraphical unit indicated. For each sample all brachiopod specimens were counted where the posterior margin is preserved and identification to species and valve type was possible (v = ventral valves; d = dorsal valves; a = articulated specimens. For Setatella significance Skovsted, Streng, Knight and Holmer, 2010 valve fragments were also counted. The total number of specimens for each species and valve type is indicated at the bottom of the table.
To download zipped file of Excel spread sheets click here.
APPENDIX 2.
Table listing all identified brachiopod specimens from all investigated samples from the Forteau Formation of southern Labrador and western Newfoundland derived from the Institute of Cambrian Studies (ICS; Boulder Colorado and University of Chicago, Michigan) and Professor John S. Peel (JSP; Uppsala) as well as samples collected by the authors from float and/or poorly constrained sections west of Forteau (F-1) and in the vicinity of Fox Cove (FCX) in southern Labrador with approximate locality details and stratigraphical units indicated. For each sample all brachiopod specimens were counted where the posterior margin is preserved and identification to species and valve type was possible (v = ventral valves; d = dorsal valves; a = articulated specimens. The total number of specimens for each species and valve type is indicated at the bottom of the table.
To download zipped file of Excel spread sheets click here.
FIGURE 1. Geology map of western Newfoundland and southern Labrador showing main geological terranes and outlining the location of the study areas discussed in this article and illustrated in more detailed maps in Figure 2, Figure 4, Figure 7.
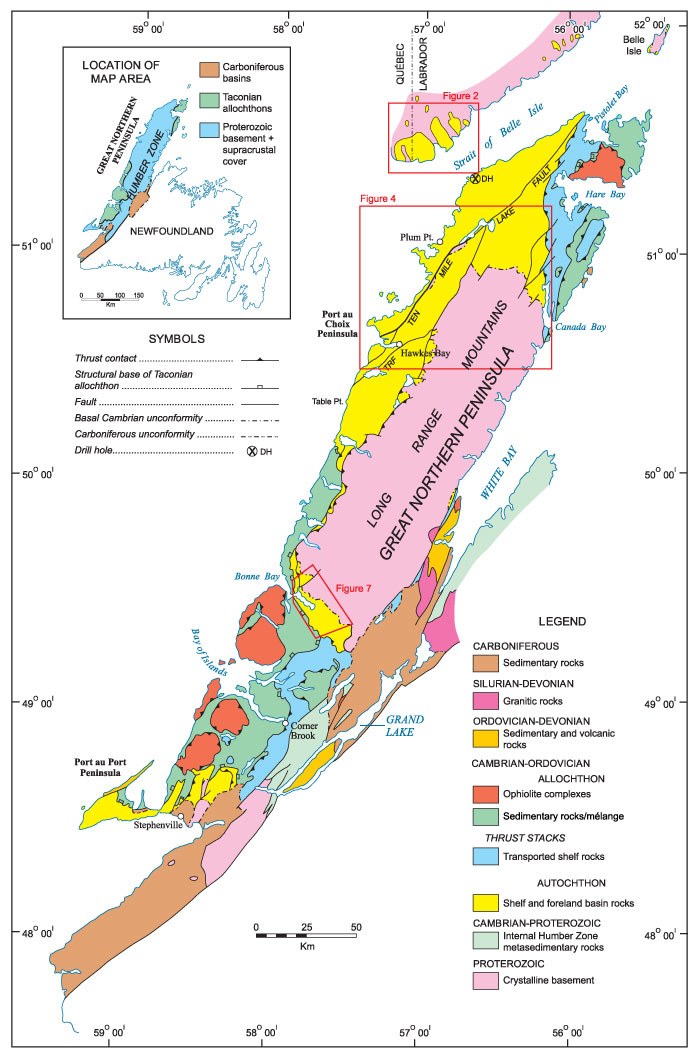
FIGURE 2. Geology map of the south coast of Labrador showing localities in the Forteau Formation, Labrador Group that were studied and yielded a rich brachiopod fauna (map based on Bostock et al., 1983 and Gower, 2010). LLQ - L’Anse-au-Loup quarry, DCQ - Diablo Cove quarry
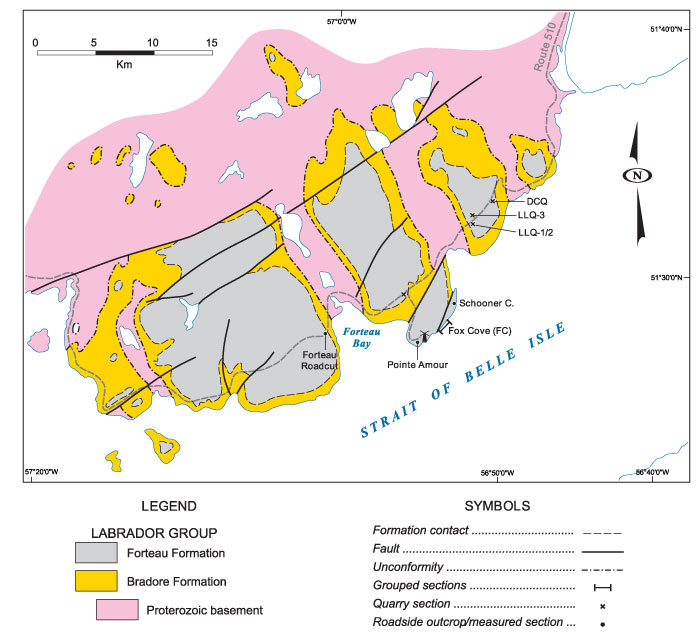
FIGURE 3. Sections measured in the Forteau Formation of south Labrador coast. FC1 (open shelf facies of James and Kobluk, 1978) and FC2a (Patch reef facies of James and Kobluk, 1978; bioherm of Debrenne and James, 1981) are adjacent to each other on the foreshore east of Fox Cove; FC2 is divided into a lower (FC2a) and upper part (FC2b) separated by a vertical fault that cuts out most of the Middle shale; FC3 is measured to the southwest of FC2 and is a composite of three short sections that are correlated over more than 50 m on the base of the Upper limestone (upper biostrome of Debrenne and James, 1981). Legend is applicable to all stratigraphic figures (Figure 5-Figure 6, Figure 8-Figure 9).

FIGURE 4. Geology map of central part of the Great Northern Peninsula showing faunally productive localities studied at Mount St. Margaret, along Route 432 north of the Long Range Proterozoic massif, and along the Big East River resource road at the south end of Highlands of St. John. MSM - Mount St. Margaret, ECP - East Castor Pond road, R432-5 etc - Route 432; BER-5 etc - Big East River resource road.
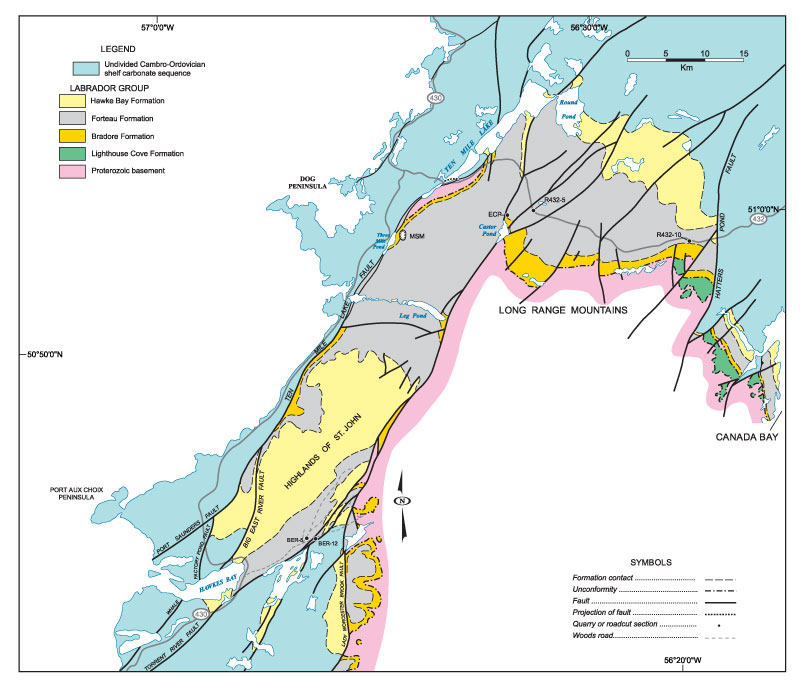
FIGURE 5. Stratigraphic section through the incomplete Forteau Formation exposed at Mount St. Margaret quarry (MSM). The archaeocyathid interval at top of section is partly based on scattered roadside outcrops above the quarry. The succession is host to brachiopod assemblage 1 and 2.
FIGURE 6. Measured sections displaying samples collected from rocks of the upper heterolithic interval of the Upper limestone, Forteau Formation on Route 432 east of Ten Mile Lake (R432-5) and Big East River resource road northeast of Hawkes Bay (BER-5) (see Figure 4). Both sections host brachiopod assemblage 2. The succession at R432-5 comprises skeletal carbonate and shale overlain by calcareous siltstone and white quartz arenite. The incomplete succession at BER-5 is lithologically similar to the lower carbonate interval of R432-5 and may tentatively suggest they are distant correlatives of the same parasequence. The brachiopod fauna hosted by lithofacies of small-scale transgression are inherited from the restricted fauna associated with the long-term regressive phase of the Forteau shelf sedimentation.

FIGURE 7. Geology map of the Gros Morne area showing location of sections in the Forteau Formation (based on Knight, 2013, figure 3). Samples GM07 -1 etc. were collected randomly in 2007 and are linked to localities GM11-1 etc. that were mapped, sectioned, and sampled in 2011 (see Figure 8-Figure 9).
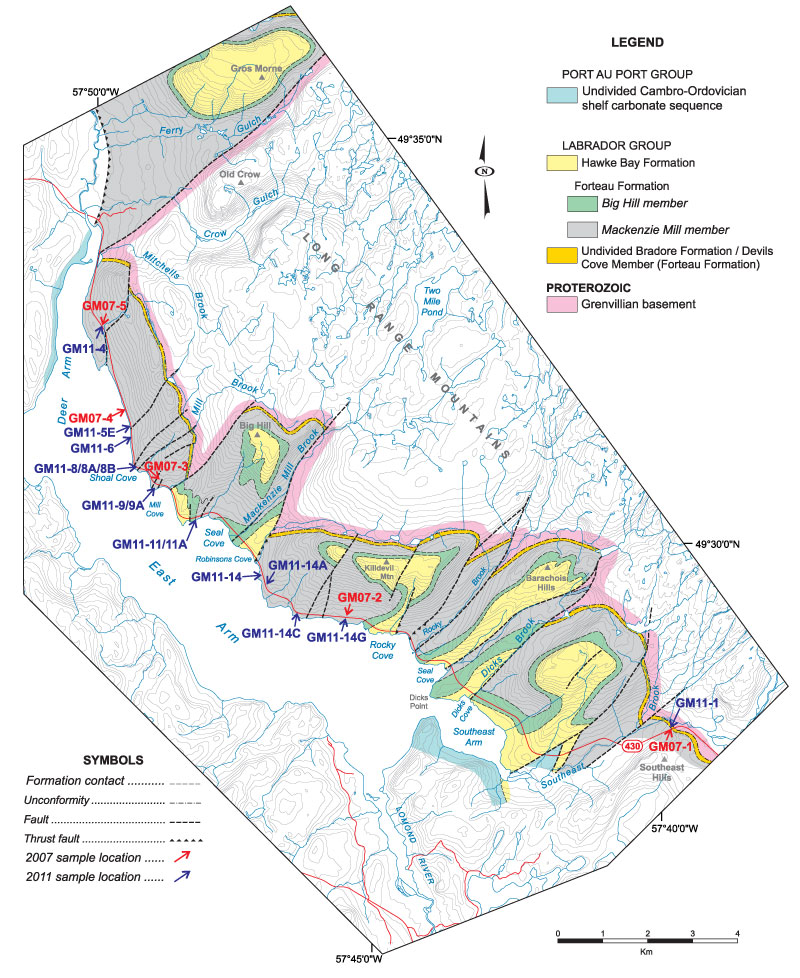
FIGURE 8. Stratigraphic sections through the Forteau Formation in Gros Morne National Park (based on Knight, 2013, figure 4) showing the correlation of limestone intervals and samples that yielded assemblage 3 brachiopod fauna. DAL - Deer Arm Limestone. DCL – Devils Cove limestone.
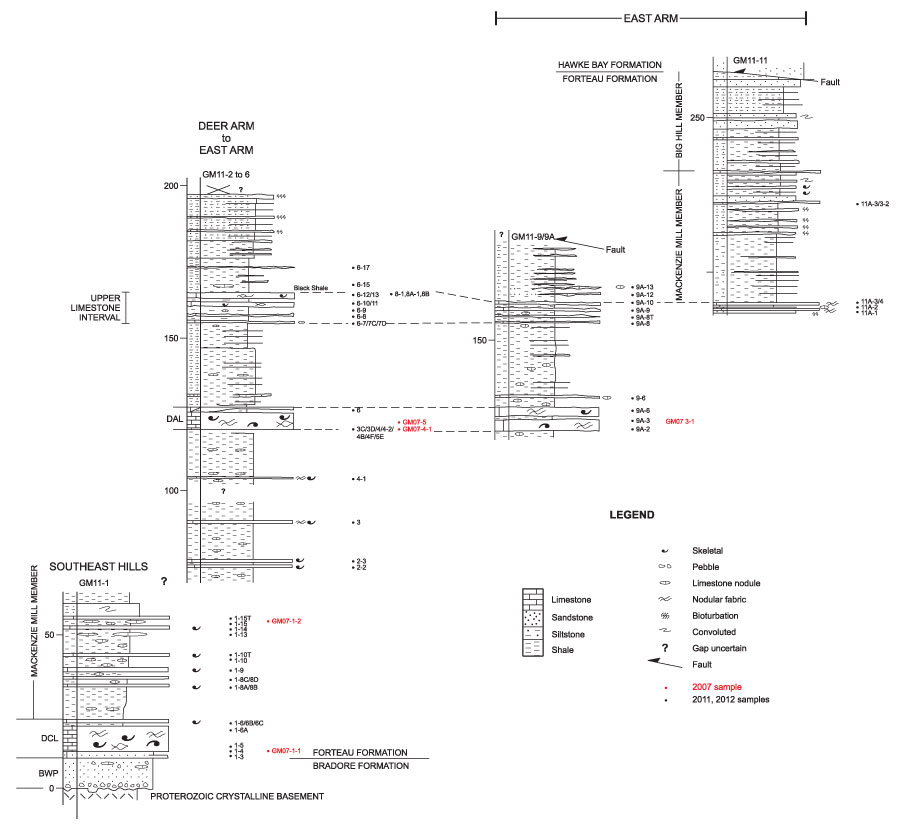
FIGURE 9. Sections for localities GM11-14, 14A and 14G at the eastern end of East Arm (see Figure 7) showing location of samples that yielded assemblage 3 brachiopod fauna. The folded and faulted succession there is difficult to correlate with any certainty to the stratigraphy of Figure 8. Nonetheless, two limestone intervals in GM11-14 are tentatively correlated with the Deer Arm (DAL) and Upper limestone interval of Figure 8. The offset of sections (GM11-14 and 14A) reflects the stratigraphic uncertainty of the structurally displaced succession although GM11-14G may correlate with the upper GM11-14A section.

FIGURE 10. Distribution of brachiopod taxa recovered from sections measured in the Forteau Formation of the south Labrador coast. Shaded area indicates known stratigraphic range of individual taxa. Correlation of adjacent sections is approximate only. For more exact correlation of sections and samples see Figure 3.
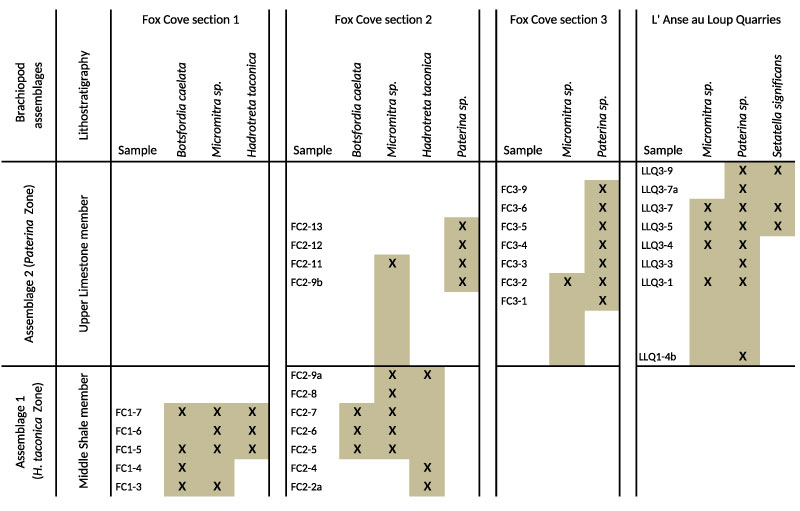
FIGURE 11. Distribution of brachiopod taxa recovered from sections measured in the Forteau Formation of the Great Northern Peninsula of western Newfoundland. Shaded area indicates known stratigraphic range of individual taxa. Correlation of adjacent sections is approximate only. For more exact correlation of sections and samples see Figure 6-Figure 7.
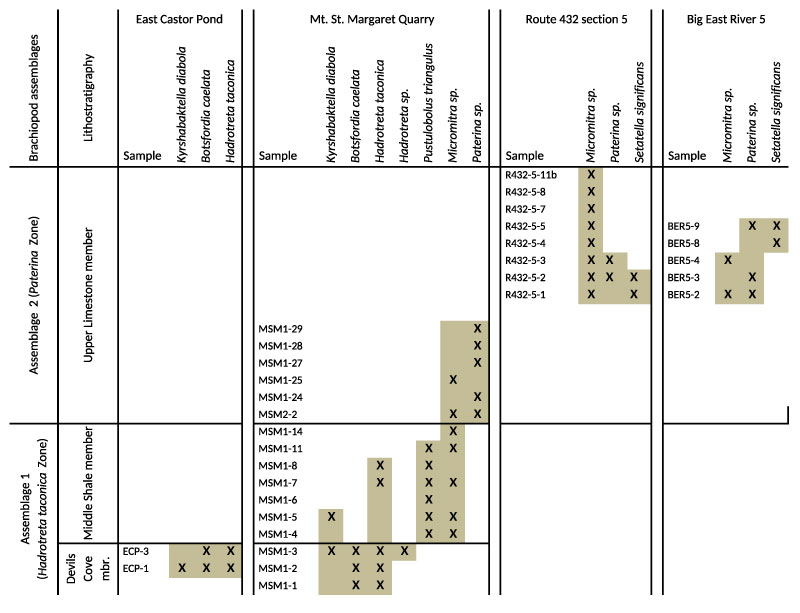
FIGURE 12. Distribution of brachiopod taxa recovered from sections measured in the Forteau Formation of the Gros Morne National Park of western Newfoundland. Shaded area indicates known stratigraphic range of individual taxa. Correlation of adjacent sections is approximate only. For more exact correlation of sections and samples see Figure 9-Figure 10.
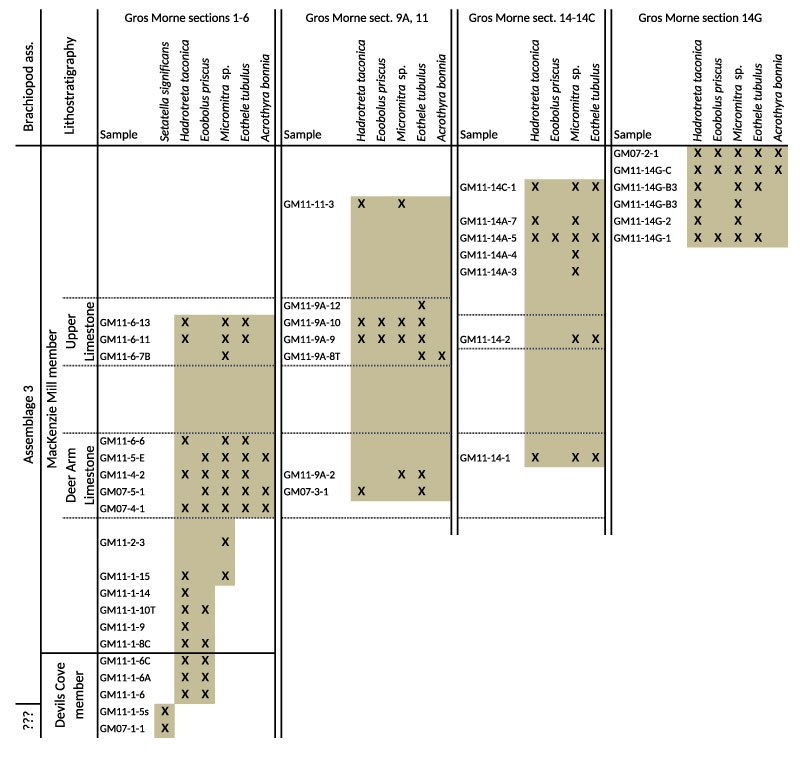
FIGURE 13. Environmental preferences of studied brachiopod taxa distributed along a simplified cross section of the eastern Laurentian shelf during the early Cambrian with the relative position of the three general study areas indicated. A. Representing the environment during the deepening phase of the transgressive systems tract (Middle shale of Labrador and the Great Northern Peninsula and Devils Cove member and lower part of the Mackenzie Mill member in Gros Morne area). B. Representing the environment during the regressive high stand system tract with the development of a prograding shelf (Upper limestone of Labrador and the Great Northern Peninsula and the upper part of the Mackenzie Mill member in Gros Morne area).
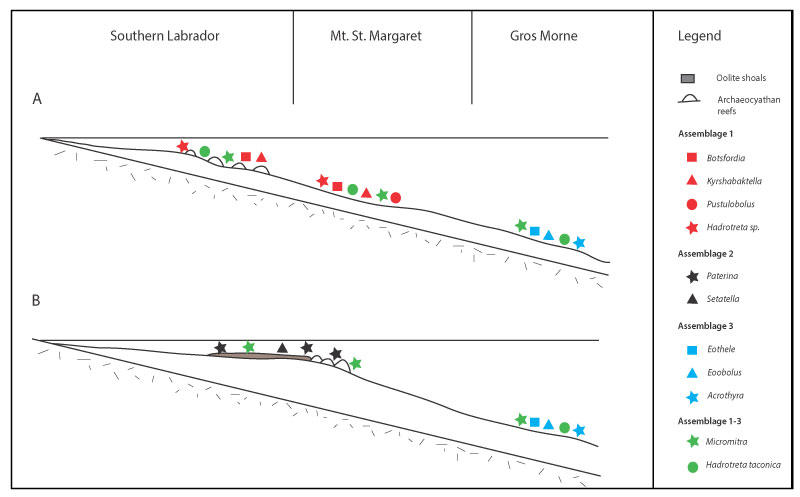
FIGURE 14. Kyrshabaktella diabola n. sp. from the Forteau Formation of southern Labrador and western Newfoundland. 1-2, Ventral valve NFM F-2516 from Devils Cove member, East Castor Pond, sample ECP-01; 1, external view; 2, detail of larval shell and resorbed pedicle groove. 3-4, Ventral valve NFM F-2517 (Holotype) from Devils Cove member, Mount St. Margaret, sample MSM-03; 3, internal view; 4, oblique posterior view showing pedicle groove. 5, Ventral valve exterior NFM F-2518 from Devils Cove member, Mount St. Margaret, sample ICS1520. 6-8, Dorsal valve NFM F-2519 from Lower Shale member, Mount St. Margaret, sample MSM-5; 6, internal view; 7, oblique internal view; 8, detail of internal surface showing vascula lateralia and median tongue with muscle scars. 9-10, Dorsal valve NFM F-2520 from the Lower shale, Fox Cove, sample ICS1575; 9, internal view; 10, detail of dorsal interior showing dorsal pseudointerarea and median tongue. All scalebars equal 500 µm except in figure 2 where the bar equals 125 µm

FIGURE 15. Eoobolus priscus (Poulsen, 1932) from the Forteau Formation of western Newfoundland. 1, Ventral valve exterior NFM 662 from Deer Arm Limestone, sample JSP 1982-01. 2-3, Ventral valve NFM F-2570 from Deer Arm Limestone, sample GM07-4-1; 2, external view; 3, detail of pustulose adult surface ornament. 4, Ventral valve interior NFM F-2571 from Deer Arm Limestone, sample GM07-4-1 showing arrangement of muscle scars. 5-6, Ventral valve NFM F-2574 from Deer Arm Limestone, sample GM07-5-1; 5, internal view; 6, oblique interal view showing imprints of pedicle nerves. 7, Ventral valve interior NFM F-2572 from Deer Arm Limestone, sample GM07-4-1. 8, Dorsal valve NFM F-2522 from Deer Arm Limestone, sample ICS 1421. 9, Ventral valve interior NFM F-2566 from Mackenzie Mills member, sample GM11-9A-10. 10-12, Dorsal valve NFM F-2567 from Mackenzie Mills member, sample GM11-9A-10; 10, internal view; 11, oblique internal view; 12, detail of columnar shell structure from naturally broken edge. Scalebars equal 250 µm in figures 1-2 and 4-11, 50 µm in figure 3 and 10 µm in figure 12.

FIGURE 16. Pustulobolus triangulus n. gen et. n. sp. from the Forteau Formation, Lower shale, Mount St. Margaret of western Newfoundland. 1-2, Ventral valve NFM F-2523, sample MSM-11; 1, oblique lateral view; 2, external view. 3-4, Ventral valve NFM F-2524, sample MSM-11; 3, detail of larval shell; 4, detail of pitted larval shell ornament. 5, Ventral valve exterior NFM F-2525, sample MSM-11. 6, Ventral valve exterior NFM F-2526, sample MSM-5. 7-8, Ventral valve NFM F-2527, sample MSM-5; 7, exterior view; 8, detail of pustulose adult surface ornament. 9-10, Dorsal Valve NFM F-2528, sample MSM-7; 9, exterior view; 10, detail of larval shell in oblique view. 11, Conjoined shell NFM F-2529, sample MSM-11, showing outline of dorsal valve and triangular ventral pseudointerarea. Scalebars equal 250 µm in figures 1-2, 5-7, 9 and 11; 50 µm in figures 3 and 10; 20 µm in figure 8 and 5 µm in figure 4.
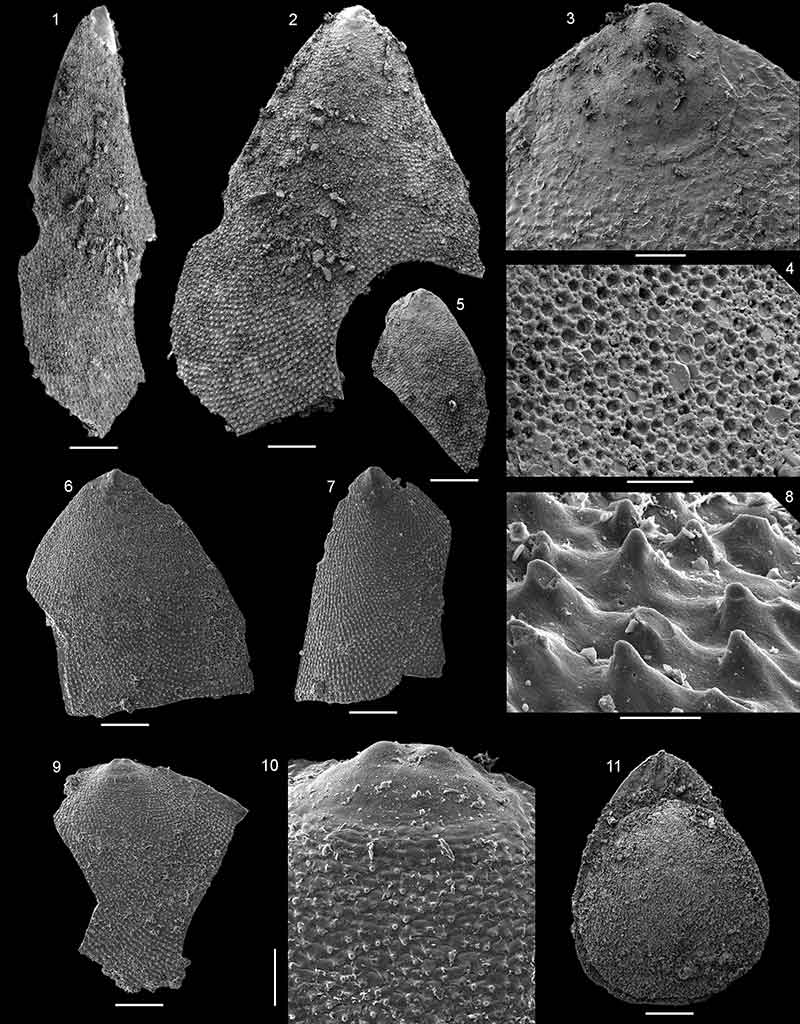
FIGURE 17. Pustulobolus triangulus n. gen et. n. sp. from the Forteau Formation, Lower shale, Mount St. Margaret of western Newfoundland, all from sample MSM-11. 1, 4, Ventral valve interior NFM F-2530, 1, interior view; 4, oblique view of interior. 2-3, Ventral valve NFM F-2531 (holotype); 2, interior view; 3, detail of ventral pseudointerarea with pustulose ornament lateral to flexure lines. 5, Ventral valve NFM F-2532, oblique view of interior showing cavities under pseudointerarea. 6, Ventral valve interior NFM F-2533. 7, Ventral valve interior NFM F-2534. 8-10, Dorsal valve NFM F-2535; 8, interior view; 9, detail of posterior margin; 10, detail of larval shell. Scalebars equal 250 µm in figures 1-2 and 4-8; 100 µm in figures 3 and 9; 20 µm in figure 10.

FIGURE 18. Botsfordia caelata (Hall, 1847) from the Forteau Formation of southern Labrador and western Newfoundland. 1, 8, Ventral valve NFM F-2536 from sample ICS1565; 1, exterior view; 8, detail of larval shell with pitted ornament. 2-4, Ventral valve NFM F-2537 from Devils Cove member, East Castor Pond, sample ECP-03; 2, external view; 3, oblique posterolateral view; 4, detail of pustulose adult shell ornament. 5-6, Ventral valve NFM F-2538 from Devils Cove member, East Castor Pond, sample ECP-03; 5, interior view; 6, oblique interior view. 7, Ventral valve exterior NFM F-2539 from Devils Cove member, East Castor Pond, sample ECP-03. 9-10, Ventral valve NFM F-2540 from Devils Cove member, East Castor Pond, sample ECP-03; 9, interior view; 10, detail interior surface in oblique view showing imprints of pedicle nerves. 11-13, Dorsal valve NFM F-2541 from Devils Cove member, East Castor Pond, sample ECP-03; 11, interior view; 12, oblique interior view; 13, detail of internal surface with dorsal pseudointerarea and posterolateral muscle scars. Scalebars equal 500 µm in figures 1-3, 5-7, 9 and 11-12; 100 µm in figures 4, 10 and 13; 10 µm in figure 8.

FIGURE 19. Eothele tubulus Ushatinskaya in Voronova et al., 1987 from the Forteau Formation of western Newfoundland. 1-3, Ventral valve NFM F-2575 from Deer Arm Limestone, sample GM07-5-1; 1, external view; 2, detail of larval shell showing pitted larval and pustulose adult shell ornament; 3, detail of posterior margin in oblique view showing oval opening of pedicle tube. 4, Ventral valve NFM F-2573 from Deer Arm Limestone, sample GM07-4-1. 5-6, Ventral valve NFM F-2568 from Deer Arm Limestone, sample GM07-3-1; 5, internal view; 6, oblique view showing median tongue and imprints of vascula lateralia. 7-8, Dorsal valve NFM F-2576 from Deer Arm Limestone, sample GM07-5-1; 7, external view; 8, oblique posterior view. 9-10, Dorsal valve NFM F-2521 from Deer Arm Limestone, sample GM07-5-1; 9, oblique lateral view; 10, oblique posterior view. 11-12, Dorsal valve NFM F-2569 from Mackenzie Mill member, sample GM07-2-1; 11, internal view showing dorsal pseudointerarea; 12, oblique lateral view. All scalebars equal 500 µm except in figures 2-3 that equals 100 µm.

FIGURE 20. Acrothyra bonnia n. sp. from the Forteau Formation of western Newfoundland, Deer Arm Limestone, sample GM07-5-1. 1-3, Ventral valve NFM F-2559; 1, external view; 2, oblique lateral view; 3, detail of larval shell and pedicle foramen. 4, Ventral vave NFM F-2560, oblique lateral view. 5-6, Dorsal valve NFM F-2561; 5, external view; 6, oblique posterior view. 7-9, Ventral valve NFM F-2562 (Holotype), 7, internal view; 8, oblique lateral view of posterior margin and apical process; 9, detail of oblique anterior view of shell interior showing internal opening of pedicle foramen and muscle scars lateral to apical process. 10-11, Ventral valve NFM F-2563, 10, internal view; 11, oblique anterior view. 12-13, Ventral valve NFM F-2564, 12, internal view; 13, detail of ventral pseudointerarea in oblique posterior view showing triangular intertrough. 14, Dorsal view NFM F-2565, oblique lateral view showing low median ridge. All scalebars equal 250 µm except in figures 3, 9 and 13 that equals 100 µm.
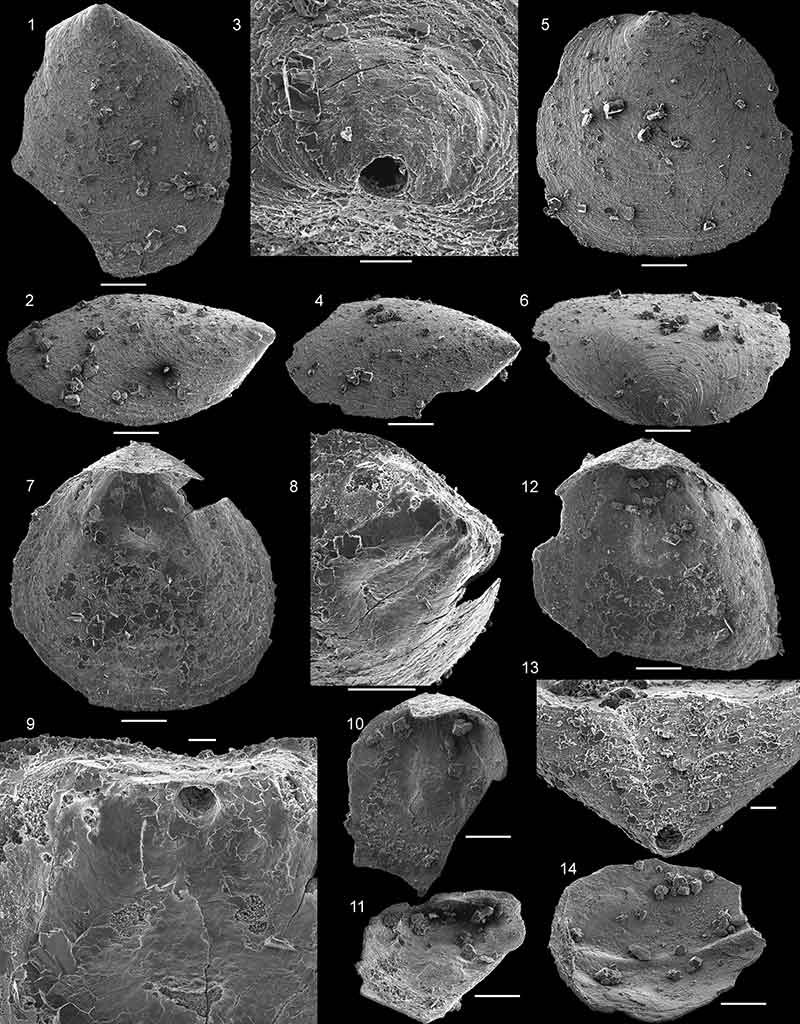
FIGURE 21. Hadrotreta taconica (Walcott, 1887) from the Forteau Formation of southern Labrador and western Newfoundland. 1-4, Ventral valve NFM F-2542 from sample ICS1577; 1, external view; 2, oblique lateral view; 3, oblique posterolateral view; 4, detail of apex showing larval shell. 5-6, Ventral valve NFM F-2543 from sample ICS1577; 5, external view; 6, detail of adult shell ornament. 7, Ventral valve NFM F-2544 from sample ICS1565 in external view. 8, Ventral valve NFM F-2545 from sample ICS1565, detail of posterior margin in oblique lateral view showing position of pedicle foramen at junction between larval and adult shell. 9, Conjoined shell NFM F-2546 ICS1575 in lateral view. 10-12, Ventral valve NFM F-2547 from sample ICS1545; 10, internal view; 11, oblique anterior view; 12, detail of apical process, pedicle foramen and apical pits. 13, Ventral valve NFM F-2548 from sample ICS1553; oblique lateral internal view. 14, Dorsal valve NFM F-2549 from sample ICS1548; external view. 15-17, Dorsal valve NFM F-2550 from sample ICS1553; 15, internal view; 16, detail of posterior margin showing dorsal pseudointerarea; 17, oblique lateral view. 18-19, Dorsal valve NFM F-2551 from sample ICS1575; 18, internal view; 19, oblique lateral view. Scalebars equal 200 µm in figures 1-3, 5, 7 and 9-19, 100 µm in figures 4 and 6 and 50 µm in figure 8.
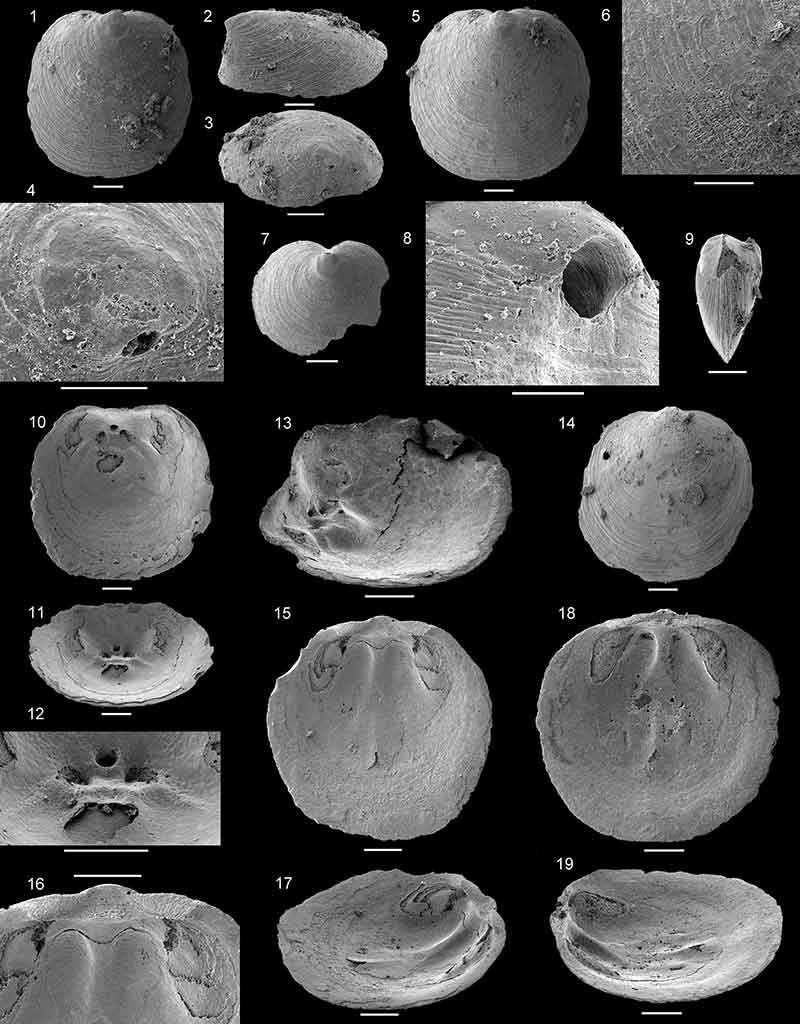
FIGURE 22. Hadrotreta sp. from the Forteau Formation of southern Labrador and western Newfoundland. 1-2, Ventral valve NFM F-2552 from sample ICS1553; 1, oblique posterior view showing procline pseudointerarea; 2, external view. 3-4, Ventral valve NFM F-2553 from sample ICS1577; 3, external view; 4, detail of larval shell with drop-shaped pedicle foramen. 5-6, Ventral valve NFM F-2554 from Lower shale, Mount St. Margaret, sample MSM-3; 5, internal view showing vascula lateralia and apical process; 6, oblique anterior view showing apical pits and internal opening of pedicle foramen. Scalebars equal 500 µm in figure 3, 200 µm in figures 1-2, 5-6 and 100 µm in figure 4.

FIGURE 23. Paterinids from the Forteau Formation. 1-5, Paterina sp. 1-3, ventral valve NFM F-2555 from sample ICS1548; 1, posterior view; 2, external view; 3, lateral view. 4-5, Dorsal valve NFM F-2556 from sample LLQ3-5; 4, posterior view; 5, external view. 6-9, Micromitra sp. 6-8, Ventral valve NFM F-2557 from sample MSM-11; 6, posterior view; 7, external view; 8, lateral view. 9, Dorsal valve NFM F-2558 from sample MSM-7; oblique posterolateral view. All scalebars equal 500 µm.

FIGURE 24. Setatella significans Skovsted, Streng, Knight and Holmer, 2010 from the Forteau Formation, The upper biostrome, L’anse au Loupe, southern Labrador. 1-2, Ventral valve NFM F-792 sample LLQ3-7; 1, internal view of posterior margin; 2, posterior view showing pseudointerarea. 3-4, Ventral valve NFM F-790, sample LLQ3-9; 3, internal view; 4, oblique lateral view. 5, Ventral valve NFM F-789, sample LLQ3-7, external view. 6, Shell fragment NFM F-793, sample LLQ3-9. All scalebar equals 250 µm.

TABLE 1. Brachiopod assemblages of the Forteau Formation of southern Labrador and western Newfoundland, Canada with the presence of each taxon and assemblage in investigated stratigraphical sections indicated.
| Sampled sections | ||||||||||||||
| Brachiopod taxa studied | Fox Cove Middle shale | MSM Devils Cove member | MSM Middle shale | Fox Cove Upper lmst. | L'Anse au Loupe 1-3 | MSM Upper lmst. | Big East River section 5 | Route 432 section 5 | *Gros Morne 1-6 | Gros Morne 9A-11 | Gros Morne 14-14C | Gros Morne 14G | Assemblages | |
| Inner shelf, Transgressive Stage | Inner shelf, Regressive Stage | Outer shelf | ||||||||||||
| Micromitra sp. | x | x | x | x | x | x | x | x | x | x | x | 1, 2, 3 | ||
| Hadrotreta taconica | x | x | x | x | x | x | x | 1, 3 | ||||||
| Eothele tubulus | x | x | x | x | 3 | |||||||||
| Eoobolus priscus | x | x | x | x | ||||||||||
| Acrothyra bonnia | x | x | x | |||||||||||
| Paterina sp. | x | x | x | x | x | 2 | ||||||||
| Setatella significans | x | x | x | |||||||||||
| Botsfordia caelata | x | x | 1 | |||||||||||
| Kyrshabaktella diabola | x | x | x | |||||||||||
| Pustulobolus triangulus | ? | x | ||||||||||||
| Hadrotreta sp. | x | x | ||||||||||||
| *) excludes the Devils Cove member | ||||||||||||||
TABLE 2. Distribution of Botsfordia caelata (Hall, 1847) and Eoobolus priscus (Poulsen, 1932) in GGU (Grønlands Geologiske Undersøgelse - now part of the Geological Survey of Denmark and Greenland) samples collected from the upper Bastion Formation at Albert Heim Bjerge and C.H. Ostenfeld Nunatak of North-East Greenland. Absolute number of specimens for each investigated GGU sample with relative proportions of the two species for each area indicated. See Skovsted and Holmer (2005) and Skovsted (2004, 2006) for more details.
| Section + sample | Taxon | Botsfordia caelata | Eoobolus priscus | |
| Albert Heim Bjerge | ||||
| GGU 314804 | 10 | |||
| GGU 314806 | 8 | |||
| GGU 314807 | 61 | |||
| GGU 314808 | 19 | |||
| GGU 314809 | 23 | 97 | ||
| GGU 314835 | 246 | |||
| GGU 314836 | 5 | |||
| GGU 314837 | 1 | 9 | ||
| Ʃ | 368 | 111 | ||
| % | 77% | 23% | ||
| CH Ostenfeld Nunatak | ||||
| GGU 314901 | 2 | 3 | ||
| GGU 314903 | 7 | 3 | ||
| GGU 314904 | 5 | 38 | ||
| GGU 314906 | 75 | |||
| GGU 314908 | 4 | 61 | ||
| GGU 314909 | 1 | 9 | ||
| GGU 314910 | 22 | 5 | ||
| GGU 314918 | 61 | |||
| GGU 314919 | 2 | |||
| GGU 314931 | 21 | |||
| GGU 314933 | 12 | 94 | ||
| GGU 314934 | 3 | |||
| GGU 314936 | 2 | |||
| GGU 314937 | 1 | |||
| Ʃ | 116 | 315 | ||
| % | 27% | 73% | ||
Depth related brachiopod faunas from the lower Cambrian Forteau Formation of southern Labrador and western Newfoundland, Canada
Plain Language Abstract
A diverse fauna of fossil brachiopods (lamp-shells) from the early part of the Cambrian Period (about 520-512 million years before present) is found in limestones of the Forteau Formation of southern Labrador and western Newfoundland (Canada). The fauna contains 11 species with phosphatic shells that were extracted from the rock using weak acids. Several species are well known from other regions of North America and the rest of the world but the fauna also contains three species and one genus that are new to science.
Rocks of the Forteau Formation were formed on the sea floor along the shelf of paleocontinent Laurentia (mainly present day North America but also including Greenland and parts of Scotland and Argentina) but the nature of the rocks reveal that the conditions on the sea floor varied very much in the sampled areas. In southern Labrador the formation is mainly composed of shallow water carbonate rocks with abundant reefs while mud rocks dominate the sequence in western Newfoundland reflecting deeper water environments.
For the first time, we show that the brachiopod faunas of the deep and shallow water environments of the Forteau Formation differ very much in composition, and that some species are restricted to shallow or deep water environments. By comparing the fauna of the Forteau Formation to other brachiopod faunas from eastern North America (from Greenland, Quebec, Vermont, New York State, and Pennsylvania) we show that the same factors seem to control the distribution of brachiopods also in these areas. This discovery indicates that fossil brachiopods can be used as a tool to evaluate at which water depth Cambrian sedimentary rocks were deposited.
Resumen en Español
Faunas de braquiópodos en relación con la profundidad de la Formación Forteau del Cámbrico inferior del sur de Labrador y el oeste de Terranova
Se describe una fauna diversa de braquiópodos organofosfáticos de finales del Cámbrico temprano (Serie 2, Etapa 3-4) de la Formación Forteau del sur de Labrador y el oeste de Terranova. La fauna total incluye 11 especies que representan una amplia selección de grupos de braquiópodos del Cámbrico. Se reconocen tres asociaciones distintas: las asociaciones 1 y 2 se encuentran en carbonatos con origen en aguas poco profundas en asociación con arqueociatos en el sur de Labrador y en el lado occidental de la Gran Península del Norte de Terranova occidental. La Asociación 3 se encuentra en un ambiente de plataforma distal del Parque Nacional Gros Morne. Las asociaciones 1 y 2 se encuentran en continuidad estratigráfica y definen dos biozonas de braquiópodos, una zona inferior de Hadrotreta taconica y una zona superior de Paterina, respectivamente. La presencia de H. taconica en la Asociación 3 de braquiópodos posiblemente indica contemporaneidad con la Asociación 1, pero en un entorno de aguas profundas. La identificación de diferentes asociaciones de braquiópodos contemporáneas (1 y 3) en ambientes poco profundos y profundos de la Formación Forteau permite por primera vez un análisis de las limitaciones ambientales que determinan la distribución de taxones individuales de braquiópodos en la sucesión del Cámbrico inferior de Laurentia oriental. La comparación con las faunas de otras áreas indica que los patrones de distribución identificados pueden extenderse a otras faunas de braquiópodos que se encuentran a lo largo de la paleocosta/margen del Cámbrico en Laurentia oriental. El estudio indica que algunos taxones determinados de braquiópodos se pueden usar como indicadores de paleoprofundidad; Botsfordia caelata en ambientes poco profundos y Eoobolus priscus y Eothele tubulus en aguas profundas. Se describen los siguientes taxones como nuevos: Kyrshabaktella diabola n. sp., Pustulobolus triangulus n. gen et n. sp. y Acrothyra bonnia n. sp.
Palabras clave: Brachiopoda; Linguliformea; Cámbrico; Paleoecología; Bioestratigrafía; Laurentia
Traducción: Enrique Peñalver (Sociedad Española de Paleontología)
Résumé en Français
Des faunes de brachiopodes variant selon la profondeur dans le Cambrien inférieur de la formation de Forteau, sud du Labrador et ouest de Terre-Neuve
Une faune diverse de brachiopodes organophosphatiques est décrite dans la fin du Cambrien ancien (Série 2, Stage 3-4) de la formation de Forteau au sud du Labrador et à l’ouest de Terre-Neuve. Au total, la faune inclut 11 espèces représentant une large sélection de groupes de brachiopodes cambriens. Trois assemblages distincts sont reconnus : les assemblages 1 et 2 sont trouvés dans des carbonates d’eau peu profonde en association avec des archéocyathes au sud du Labrador et sur la côte ouest de la Grande Péninsule du Nord à l’ouest de Terre-Neuve. L’assemblage 3 est trouvé dans un milieu de plateforme distale dans le Parc National du Gros-Morne. Les assemblages 1 et 2 sont trouvés en continuité stratigraphique et définissent deux biozones à brachiopodes, une zone inférieure à Hadroteta taconica et une zone supérieure à Paterina, respectivement. La présence d’H. taconica dans l’assemblage 3 de brachiopodes indique une possible équivalence temporelle avec l’assemblage 1 mais dans un milieu d’eau profonde. L’identification d’assemblages de brachiopodes distincts et temporellement équivalents (1 et 3) dans des environnements d’eaux profonde et peu profonde dans la formation de Forteau permet pour la première fois une analyse des contraintes environnementales déterminant la distribution des taxons individuels de brachiopodes dans la succession du Cambrien inférieur à l’est de la Laurentia. Une comparaison avec les faunes d’autres régions indique que les schémas de distribution identifiés peuvent être étendus aux autres faunes de brachiopodes trouvées le long de la paléocôte de l’est de la Laurentia au Cambrien. L’étude indique que des taxons spécifiques de brachiopodes peuvent être utilisés comme des indicateurs de paléobathymétrie : Botsfordia caelata dans des environnements d’eau peu profonde, Eoobolus priscus et Eothele tubulus dans des eaux plus profondes. Les nouveaux taxons suivants sont décrits : Kyrshabaktella diabola n. sp., Pustulobolus triangulus n. gen. et n. sp., et Acrothyra bonnia n. sp.
Mots-clés : Brachiopoda ; Linguliformea ; Cambrien ; paléoécologie ; biostratigraphie ; Laurentia ; nouveau genre ; nouvelle espèce
Translator: Antoine Souron
Deutsche Zusammenfassung
Tiefen-abhängige Brachiopoden-Faunen aus der unteren kambrischen Forteau Formation von Süd-Labrador und West-Neufundland
Es wird eine diverse Fauna organophosphatischer Brachiopoden aus der spätkambrischen (Serie 2, Stufe 3-4) Forteau Formation von Süd-Labrador und West-Neufundland beschrieben. Die gesamte Fauna beinhaltet 11 Arten, die eine breitgefächerte Auswahl an kambrischen Brachiopoden-Gruppen repräsentieren. Es werden drei unterschiedliche Assemblagen erkannt: die Assemblagen 1 und 2 werden in Flachwasserkarbonaten zusammen mit Archaeocyathiden in Süd-Labrador und auf der Westseite der Großen Nordinsel von West-Neufundland gefunden. Die Assemblage 3 ist in einem distalen Schelf-Milieu des Gros Morne National Parks zu finden. Die Assemblagen 1 und 2 sind stratigraphisch kontinuierlich und definieren zwei Brachiopoden-Biozonen: die untere Hadrotreta taconica Zone und die höhere Paterina Zone. Das Vorkommen von H. taconica in der Brachiopoden Assemblage 3 weist möglicherweise auf eine Zeitgleichheit von Assemblage 1 in einem Tiefwassermilieu hin. Die Identifizierung von erkennbar zeitgleichen Brachiopoden Assemblagen (1 und 3) in Flach-und -Tiefwasserbereichen der Forteau Formation erlaubt es zum ersten Mal die Umweltbedingungen zu analysieren, um die individuellen Brachiopoden-Taxa während der spätkambrischen Abfolge von Ost-Laurentia bestimmen. Vergleiche mit Faunen aus anderen Gebieten weisen darauf hin, dass die identifizierten Verteilungsmuster auf andere Brachiopoden-Faunen entlang der kambrischen Paläoküste bzw. des Laurentia-Randes erweitert werden können. Diese Studie zeigt, dass bestimmte Brachiopoden-Taxa als Indikatoren für Paläotiefe genutzt werden können; Botsfordia caelata in flacher Umgebung und Eoobolus priscus und Eothele tubulus in tieferen Gewässern. Die folgenden neuen Taxa werden beschrieben: Kyrshabaktella diabola n. sp., Pustulobolus triangulus n. gen et n. sp., Acrothyra bonnia n. sp.
Schlüsselwörter: Brachiopoda; Linguliformea; Kambrium; Paläoökologie; Biostratigraphie; Laurentia
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024